
Long-term fructose-enriched diet introduced immediately after weaning does not induce oxidative...
7
Long-term fructose-enriched diet introduced immediately after weaning does not induce oxidative stress in the rat liver Jelena Nestorov a, ⁎ , Alhadi M. Glban a , Ana Mijušković b , Aleksandra Nikolić-Kokić b , Ivana Elaković a , Nataša Veličković a , Gordana Matić a a Department of Biochemistry Institute for Biological Research “Siniša Stanković,” University of Belgrade, 11060 Belgrade, Serbia b Department of Physiology, Institute for Biological Research “Siniša Stanković,” University of Belgrade, 11060 Belgrade, Serbia ARTICLE INFO ABSTRACT Article history: Received 19 December 2013 Revised 30 May 2014 Accepted 9 June 2014 Increased fructose consumption is correlated with the rising prevalence of obesity, metabolic syndrome, and type 2 diabetes. It is believed that reactive oxygen species contribute to the development and progression of metabolic disturbances, especially those associated with insulin resistance. Dietary fructose produces both pro-oxidative and antioxidative effects, depending upon the experimental conditions, dosage, duration of treatment, and pathophysiological milieu. The effects of fructose overconsumption on young populations, which have an increased risk of developing metabolic disorders in adulthood, have not been fully elucidated. We have previously shown that rats subjected to a long-term fructose-enriched diet immediately after weaning display impaired hepatic insulin sensitivity. In this study, we tested the hypothesis that long-term fructose consumption induces alterations in the redox setting of the liver. Starting from the 21st day after birth, male Wistar rats were maintained for 9 weeks on a standard diet (control) or a fructose-enriched diet that consisted of standard food and 10% fructose solution instead of drinking water. The expression and activity of antioxidant enzymes as well as lipid peroxidation and protein damage markers were measured. The results showed that a fructose-enriched diet led to an increased expression of mitochondrial manganese superoxide dismutase but did not affect antioxidant enzymes activity, lipid peroxidation, thiol content, and the level of protein oxidation. Therefore, our results suggest that the decrease in hepatic insulin sensitivity that was previously observed in rats that were kept on the same diet regime might be attributed to molecular mechanisms other than redox disbalance. A possible fructose-related micronutrient deficiency should be examined. © 2014 Elsevier Inc. All rights reserved. Keywords: Antioxidant enzymes Fructose-fed rat Liver Oxidative stress Young rats 1. Introduction High-calorie diets and sedentary lifestyles have been correlated with the rising prevalence of obesity, metabolic syndrome, type 2 diabetes, and coronary diseases [1,2]. Widespread overcon- sumption of solid fats and added sugars, especially fructose- sweetened beverages, is the hallmark of modern lifestyles and particularly prominent in young populations [3]. Although young organisms and adults differ largely by their metabolic and physiological profiles, most of the previous studies NUTRITION RESEARCH 34 (2014) 646 – 652 Abbreviations: CAT, catalase; iNOS, inducible nitric oxide synthase; GPx, glutathione peroxidase; GR, glutathione reductase; Hsp, heat shock protein; SOD1, copper-zinc superoxide dismutase 1; SOD2, mitochondrial manganese superoxide dismutase 2; TBARS, thiobarbituric acid reactive substances. ⁎ Corresponding author. E-mail address: [email protected] (J. Nestorov). Available online at www.sciencedirect.com ScienceDirect www.nrjournal.com http://dx.doi.org/10.1016/j.nutres.2014.06.006 0271-5317/© 2014 Elsevier Inc. All rights reserved.
Transcript of Long-term fructose-enriched diet introduced immediately after weaning does not induce oxidative...

N U T R I T I O N R E S E A R C H 3 4 ( 2 0 1 4 ) 6 4 6 – 6 5 2
Ava i l ab l e on l i ne a t www.sc i enced i r ec t . com
ScienceDirectwww.n r j ou rna l . com
Long-term fructose-enriched diet introduced
immediately after weaning does not induceoxidative stress in the rat liverJelena Nestorova,⁎, Alhadi M. Glbana, Ana Mijuškovićb, Aleksandra Nikolić-Kokić b,Ivana Elakovića, Nataša Veličkovića, Gordana Matića
a Department of Biochemistry Institute for Biological Research “Siniša Stanković,” University of Belgrade, 11060 Belgrade, Serbiab Department of Physiology, Institute for Biological Research “Siniša Stanković,” University of Belgrade, 11060 Belgrade, Serbia
A R T I C L E I N F O
Abbreviations: CAT, catalase; iNOS, inducibshock protein; SOD1, copper-zinc superoxthiobarbituric acid reactive substances.⁎ Corresponding author.
E-mail address: [email protected] (J.
http://dx.doi.org/10.1016/j.nutres.2014.06.0060271-5317/© 2014 Elsevier Inc. All rights rese
A B S T R A C T
Article history:Received 19 December 2013Revised 30 May 2014Accepted 9 June 2014
Increased fructose consumption is correlated with the rising prevalence of obesity, metabolicsyndrome, and type 2 diabetes. It is believed that reactive oxygen species contribute to thedevelopment and progression of metabolic disturbances, especially those associated withinsulin resistance. Dietary fructose produces both pro-oxidative and antioxidative effects,depending upon the experimental conditions, dosage, duration of treatment, andpathophysiological milieu. The effects of fructose overconsumption on young populations,which have an increased risk of developing metabolic disorders in adulthood, have not beenfully elucidated.Wehavepreviously shownthat rats subjected toa long-termfructose-enricheddiet immediately after weaning display impaired hepatic insulin sensitivity. In this study, wetested the hypothesis that long-term fructose consumption induces alterations in the redoxsettingof the liver. Starting from the21stdayafter birth,maleWistar ratsweremaintained for 9weeks on a standard diet (control) or a fructose-enriched diet that consisted of standard foodand 10% fructose solution instead of drinkingwater. The expression and activity of antioxidantenzymes aswell as lipid peroxidation andprotein damagemarkersweremeasured. The resultsshowed that a fructose-enriched diet led to an increased expression of mitochondrialmanganese superoxide dismutase but did not affect antioxidant enzymes activity, lipidperoxidation, thiol content, and the level of protein oxidation. Therefore, our results suggestthat the decrease in hepatic insulin sensitivity that was previously observed in rats that werekept on the same diet regime might be attributed to molecular mechanisms other than redoxdisbalance. A possible fructose-related micronutrient deficiency should be examined.
© 2014 Elsevier Inc. All rights reserved.
Keywords:Antioxidant enzymesFructose-fed ratLiverOxidative stressYoung rats
1. Introduction
High-calorie diets and sedentary lifestyles have been correlatedwith the rising prevalence of obesity,metabolic syndrome, type2 diabetes, and coronary diseases [1,2]. Widespread overcon-
le nitric oxide synthase; Gide dismutase 1; SOD2
Nestorov).
rved.
sumption of solid fats and added sugars, especially fructose-sweetened beverages, is the hallmark of modern lifestyles andparticularly prominent in young populations [3]. Althoughyoung organisms and adults differ largely by their metabolicand physiological profiles, most of the previous studies
Px, glutathione peroxidase; GR, glutathione reductase; Hsp, hea, mitochondrial manganese superoxide dismutase 2; TBARS
t,

Table 1 – Diet composition
Ingredient Standard diet Fructose-enricheddiet
Fructose solution (g/L) – 100Protein 20% 20%Lysine 0.90% 0.90%Methionine + cystine 0.75% 0.75%Cellulose 8% 8%Phosphorus 0.50% 0.50%Sodium 0.15%-0.25% 0.15%-0.25%Calcium 1% 1%Manganese (mg/kg) 30 30Copper (mg/kg) 20 20Zinc (mg/kg) 100 100
647N U T R I T I O N R E S E A R C H 3 4 ( 2 0 1 4 ) 6 4 6 – 6 5 2
investigated fructose-inducedmetabolic disturbances inadults.Therefore, the link between increased fructose consumption inchildhood and the development of metabolic disorders inadulthood is still not clear.
The fructose-fed rat represents a commonly used animalmodel for studying diet-induced metabolic disturbances [4].Previous animal studies have shown that fructose-rich diets caninduce most of the features of metabolic syndrome, includinghypertension, insulin resistance, abdominal obesity, hepaticsteatosis, endothelial dysfunction, and inflammation [5,6]. It hasbeen suggested that oxidative stress participates in the develop-ment and progression of these metabolic disturbances [7,8].
Oxidative stress is a condition that occurs when the pro-oxidants (radicals) overwhelms the antioxidant defense. Theantioxidant defense system includes antioxidant enzymes, suchas superoxide dismutase (SOD); catalase (CAT); glutathioneperoxidase (GPx); glutathione reductase (GR); and nonenzymaticlow-molecular weight compounds, including reduced gluta-thione; vitamins A, E, and C; beta-carotene; and bilirubin. Inaddition, numerous proteins, including heat shock proteins(Hsp): Hsp70 and Hsp90, assist in the repair of oxidativelydamaged cellular biomolecules.
The induction of some metabolic syndrome features bychronic fructose feeding or by a single dose of fructose [9] wasassociated with oxidative stress and disruption of antioxidantmechanisms [10].However, a largediscrepancy in thecourseandthe intensity of fructose-induced alterations in the functioningof antioxidant enzymes has been observed. Protective effects offructose and its phosphorylated forms after short-term applica-tion were also demonstrated in oxidative stress–related condi-tions, and several studies have reported their antioxidative andcytoprotective effects [11-14]. In general, it appears that negativeeffects of fructose mostly emerge after long-term exposure,whereas its acute application seems to protect the cells and canbe beneficial under some pathophysiological conditions [11].
Our previous studies show that a 9-week fructose-enricheddiet, applied immediately after weaning, led to a decrease ininsulin sensitivity, alterations of lipidmetabolism, and low-gradeinflammation in rat livers [15,16]. The question now though iswhether the same diet regime promotes oxidative stress, whichmay be responsible for previously observed metabolic distur-bances, such as decreased hepatic insulin sensitivity. In thisstudy, we tested the hypothesis that long-term fructose con-sumption in the period from weaning to adulthood inducesalterations in the redox setting of the liver, as themain fructose-metabolizing tissue. To test this hypothesis, we measured theexpression and the activity of antioxidant enzymes aswell as thelevel of markers of lipid and protein damage in the liver of ratssubjected to a fructose-rich diet immediately after weaning.
Iron (mg/kg) 100 100Iodine (mg/kg) 0.5 0.5Selenium (mg/kg) 0.1 0.1Vitamin A (IU/kg) 10000 10000Vitamin D3 (IU/kg) 1600 1600Vitamin E (mg/kg) 25 25Vitamin B12 (mg/kg) 0.02 0.02
Control animals were fed with commercial standard chow anddrinking water, and rats on fructose-enriched diet were fed with thesame chow and 10% (wt/vol) fructose solution instead of drinkingwater. Animals were exposed to these diets during 9 weeks.
2. Methods and materials
2.1. Animals and diets
Male Wistar rats (21 days old) were randomly divided into 2experimental groups with nine rats each. The control group (C)was provided with standard chow and drinking water, and thefructose group (F) was provided with the same chow but10% (wt/vol) fructose solution instead of the drinking water.
Both experimental groups had access to food and drinking fluidfor 9 weeks ad libitum. The detailed composition of the diets ispresented in Table 1. The choice of fructose concentration wasbased on data stating that 10% fructose solution closelyresembles the intake of sweet beverages that are characteristicof the Western diet [17]. Rats were kept under standardconditions, at 22°C with a 12-hour light/dark cycle. During the9 weeks of treatment, food and liquid intake was measureddaily. Caloric intake was calculated as the sum of caloriesconsumed as both food and liquid. Body mass was measuredweekly, whereas the livers were weighed immediately aftersacrifice. The procedures complied with the EEC Directive (86/609/EEC) on theprotectionof animalsused for experimentalandother scientific purposes, and theywere approved by theEthicalCommittee for theUseof LaboratoryAnimals of the Institute forBiological Research “Siniša Stanković,” University of Belgrade.
2.2. Tissue collection and determination of glucoseconcentration in the blood
After overnight fasting, rats were euthanized by rapiddecapitation. Trunk blood was quickly collected into EDTA-containing tubes and agitated slowly. Glucose concentrationin the blood was immediately measured with MultiCare strips(Biochemical Systems International, Arezzo, Italy). After bloodcollection, the livers were perfused with cold 0.9% NaCl,excised, and stored in liquid nitrogen.
2.3. Tissue preparation and determination of antioxidantenzymes activity
For preparation of whole cell extracts, the livers were homoge-nized in 10 vol (wt/vol) of Tris buffer (50mMTris, 0.25M sucrose,1 mM EDTA, pH 7.4) and sonicated (3 × 10 s at 10 MHz on ice),

Table 2 – Food, liquid, and caloric intakes; body and livermass; and fasting glucose level
Control Fructose
Food intake (g/day/animal) 20.57 ± 2.63 14.20 ± 2.01*Liquid intake (mL/day/animal) 49.33 ± 5.39 74.10 ± 11.36*Total caloric intake (kJ/day/animal) 226.23 ± 28.90 283.65 ± 24.51*Caloric intake from fructoserelative to total caloric intake (%)
0 45
Body mass (g) 367.78 ± 48.42 358.33 ± 43.24Weight gain (g) 328.11 ± 45.68 319.75 ± 39.35Mass of liver (g) 12.48 ± 1.82 10.77 ± 1.46Liver-to-body ratio (×100) 3.67 ± 0.25 3.48 ± 0.41Blood glucose (mmol/L) 5.20 ± 0.42 4.84 ± 0.62
Male 21-day-old rats were maintained for 9 weeks on the standarddiet (control) or on the fructose-enriched diet that was comprised ofstandard food but 10% fructose drinking solution. Food and liquidintakes were measured daily. Total caloric intake was calculated assumof calories ingested as foodand liquid. Bodymasswasmeasuredweekly, whereas the livers were weighed immediately after the ratswere euthanized and presented as absolute units and in relation tobody mass at the end of experiment. Blood glucose level wasdetermined at the end of experiment, after overnight fasting ofanimals. Values are expressed as the means ± SD (n = 9 animals pergroup). P < .05 was considered statistically significant. ⁎P < .05.
648 N U T R I T I O N R E S E A R C H 3 4 ( 2 0 1 4 ) 6 4 6 – 6 5 2
before60minutesof centrifugationat105000g. Thesupernatantswere used as whole cell extracts. Total SOD activity wasdetermined by the adrenaline method [18]. One SOD unit wasdefined as the amount of the enzyme necessary to decrease therate of adrenalin auto-oxidation by 50% at pH 10.2. Fordetermination of manganese superoxide dismutase 2 (SOD2)activity, the assaywas performed after preincubationwith 8mMpotassium cyanide. The copper-zink superoxide dismutase 1(SOD1) activitywas calculatedas thedifferencebetween the totalSOD and SOD2 activities. Catalase activity was determinedaccording to Claiborne [19]. One unit of CAT activity was definedas the amount of the enzyme that decomposes 1mmol H2O2 perminute at 25°CandpH7.0. Theactivity ofGPxwasdeterminedbythe glutathione reduction of t-butyl hydroperoxide, using amodification of the assay described by Paglia and Valenine [20].One unit of GPx activity was defined as the amount of theenzyme needed to oxidize 1 mmol NADPH per minute at 25°CandpH7.0. Glutathione reductase activitywas determinedusingthe method of Glatzle et al [21]. One unit of GR activity wasdefined as the amount of the enzyme needed to oxidize 1 nmolNADPH per minute at 25°C and pH 7.4. All enzyme activities areexpressed as units (U) per mg of protein.
2.4. Assessment of total thiols, thiobarbituric acid reactivesubstances, and protein carbonyl groups
Thiol content was determined after incubation (20 minutes,37°C) of the whole cell extracts with 5 mM 5,5-dithiobis-2-nitrobenzoic acid (DTNB) (ε412 = 13600 M−1 ∙ cm−1).
Formeasurement of lipid peroxidation products and proteincarbonyl groups, livers were homogenized in 10 vol (wt/vol) ofTrisbufferpH7.4without sucrose, and then theywere sonicatedand centrifuged at 6000g. The degree of lipid peroxidation wasassessed by estimating the thiobarbituric acid reactive sub-stances (TBARS) [22]. The absorbance wasmeasured at 530 nm.TBARS concentrationwas calculated using themolar extinctioncoefficient of malondialdehyde (ε530 = 15600 M−1 ∙ cm−1), andexpressed in nmol per mg of protein.
Colorimetric analysis of protein carbonylswas carried out byamodifiedmethodof Levine [23]. Liverhomogenate (100μL)wasmixedwith 400 μL of 10mM2,4-dinitrophenylhydrazine (DNPH)in 2 M HCl. Samples were kept in the dark for 45 minutes, withoccasional mixing. After the addition of 500 μL of ice-cold20% trichloroacetic acid, the samples were centrifuged for10 minutes at 10000 rpm at 4°C. The supernatants were thenaspirated and discarded. The pellets were washed 3 times with1 mL of 1:1 (vol/vol) ethanol:ethyl acetate and followed bycentrifugation for 10minutesat 10000 rpm.The final precipitatewas resolubilized in 100 μL of 8 M urea. For each sample, theprocedure was carried out with a blank containing liverhomogenate and 2 M HCl without DNPH. The absorbance wasmeasured at 375 nm. Protein carbonyl contentwas expressed innanomoles per milligram of protein, using molar absorptioncoefficient ε375 = 22000 M−1 ∙ cm−1.
2.5. SDS-Polyacrylamide gel electrophoresisand immunoblotting
Proteins were resolved in 12% SDS-polyacrylamide gels andtransferred to polyvinylidene difluoride (PVDF) membrane.
SOD1, SOD2, GR, and GPx were detected by Abcam antibodies(ab13489, ab13533, ab16801, and ab22604, respectively). Induciblenitric oxide synthase (iNOS) was detected by 610330 antibody(BD Transduction Laboratories, San Jose, CA, USA); CAT, byAbgent antibody AP8623-c; and β-actin, by AC-15 antibody(Sigma-Aldrich, St. Louis, MO, USA), whereas Hsp70 andHsp90 were detected by StressGen antibodies SPA-820 andSPA-830, respectively. After incubation with secondary anti-bodies, the immunoreactive proteins were visualized by theenhanced chemifluorescence method (Amersham Biosci-ences Limited,Buckinghamshire, UK). Quantitative analysisof immunoreactive bands was performed using ImageQuantsoftware. β-Actin was used as equal load control.
2.6. Statistical analyses
Eachassaywasperformed in triplicate for each sample.Dataarepresented as means ± SEM, except for food, liquid, and caloricintake; liver and body masses; and fasting glucose level, whichare expressed as means ± SD. Between-group differences wereassessed using the Student t test. A probability level of P < .05was considered statistically significant.
3. Results and discussion
During thenineweeks of treatment, food and liquid intakesweremeasured daily. Food intake was significantly reduced, whereasthe liquid and caloric intakes were significantly higher infructose-fed rats, as compared with controls (Table 2). Despitethe increased energy intake observed in fructose-fed rats, nosignificant differences in body mass were detected between theexperimental groups (Fig. 1). There were also no significantdifferences perceived between the groups in liver mass, liver-to-body ratio, or blood glucose concentration (Table 2). Because rats

0
50
100
150
200
250
300
350
400
450
0 1 2 3 4 5 6 7 8 9
Bod
y m
ass
(g)
Week
C F
Fig. 1 – Time course of the average body mass of controland fructose-fed rats. Body mass was measured weeklyover a 9-week experimental period. Groups: C, control (■);F, fructose-fed rats (▲). Values are means ± SD (n = 9).
649N U T R I T I O N R E S E A R C H 3 4 ( 2 0 1 4 ) 6 4 6 – 6 5 2
were not fed a protein and micronutrient-fortified diet, reducedchow intake in the fructose-fed group could have led to adecrease in the total mineral and vitamin density per unit ofenergy consumed. Therefore, the possibility that fructose-enriched diet induced, at least marginally, micronutrient(vitamins andminerals) deficiency cannot be ruledout. Excessivefructose intake may impair mineral status and, thus, contributeto the development of diet-induced metabolic disorders [24-27].
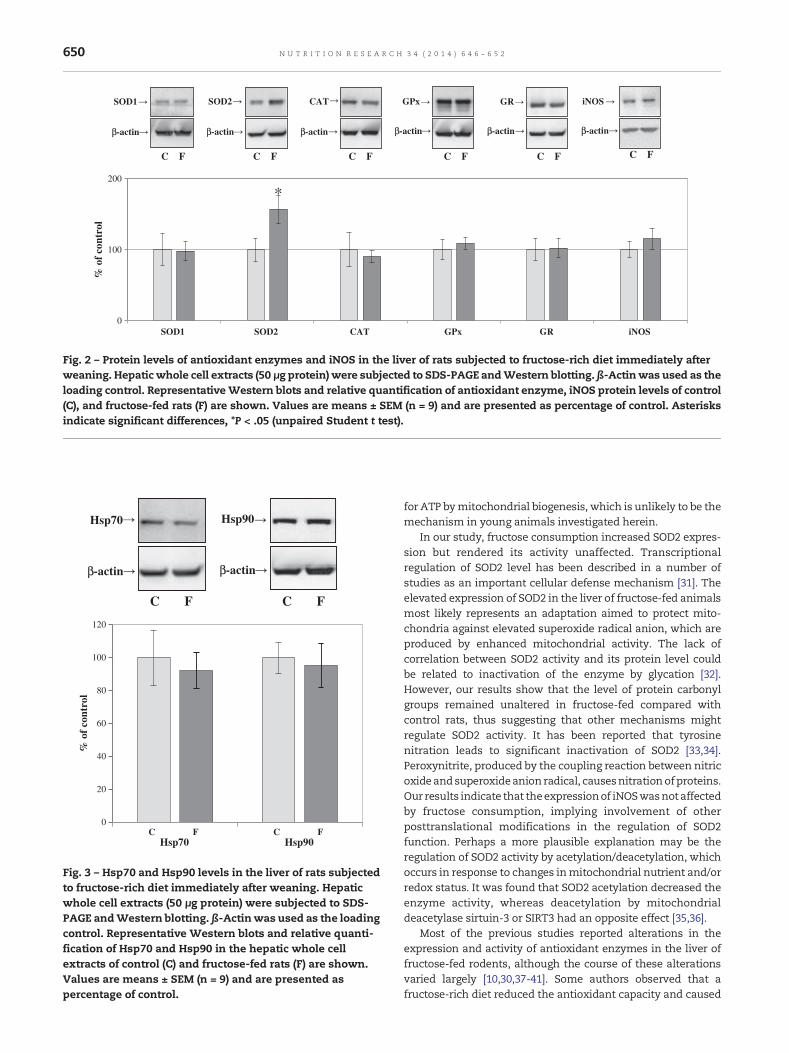
In previous research, we presented histological analyses ofrat liver, after the same diet regime, which showed no signs ofhepatic steatosis, inflammatory cell infiltration, or hepatocytesnecrosis [15]. In this study, we demonstrated that fructoseconsumption did not affect the activity of antioxidant enzymesand did not provoke oxidative damage of proteins and lipids inthe liver of rats subjected to fructose-enricheddiet immediatelyafter weaning. No significant differences in the activities ofSOD1, SOD2,CAT, GPx, andGRbetween the fructose-fedand thecontrol groups were detected (Table 3). The expression ofmitochondrial SOD2 was elevated by 50% (P < .05) in fructose-fed rats, and this was the only antioxidant enzyme whose
Table 3 – Antioxidant enzyme activities, total thiol (SH),TBARS, and protein carbonyl level in the liver of ratssubjected to fructose-rich diet immediately after weaning
Control Fructose
SOD1 (U/mg protein) 32.20 ± 0.94 27.70 ± 4.83SOD2 (U/mg protein) 4.64 ± 0.75 3.52 ± 0.76CAT (U/mg protein) 309.89 ± 12.66 297.98 ± 30.68GPx (U/mg protein) 631.00 ± 40.00 618.00 ± 67.00GR (U/mg protein) 39.72 ± 2.52 43.78 ± 3.18SH groups (mM/mg protein) 0.052 ± 0.012 0.050 ± 0.017TBARS (nmol/mg protein) 0.432 ± 0.018 0.465 ± 0.006Protein carbonyl (nmol/mg protein) 5.81 ± 0.77 5.06 ± 0.47
Antioxidant enzyme activities in hepatic whole cell extracts ofcontrol and fructose-fed rats are expressed as units per milligramof protein. The level of total thiols is expressed as millimoles perliter of −SH groups per milligram of protein. The level of TBARS andcarbonyl groups is expressed as nanomoles per milligram ofprotein. Values are expressed as the means ± SEM (n = 9).
expressionwas altered (Fig. 2). Keeping inmind that nitric oxideand superoxide anion radicals react to form the peroxynitrite(which is amore potent oxidant),we examined the protein levelof iNOS, a nitric oxide generating enzyme, the expression ofwhich is up-regulated under pro-oxidative conditions. The levelof iNOS remained unaltered in fructose-fed rats when com-pared to controls (Fig. 2). In addition, no significant differencesin the total thiol content, the level of TBARS, and the level ofprotein carbonyls between fructose-fed and control rats wereobserved (Table 3). The expression of molecular chaperonesHsp70 and Hsp90, which can serve as markers of proteindamage, also remained unchanged by the fructose-enricheddiet (Fig. 3).
Liver is themain fructose-metabolizing tissue, which, due tothe high activity of fructokinase, rapidly absorbs fructose fromthe blood and converts it into fructose-1-phosphate. A conse-quent rapid depletionofATP and free phosphate inhepatocyteshas to be compensated by the enhancedmitochondrial activity.In line with this, we previously observed increased mitochon-drial β-oxidation of fatty acids in the liver of rats subjected to afructose-enricheddiet immediately afterweaning (unpublisheddata). However, the facilitated activity of electron transportchain inevitably leads to an enhancement of electron leakageand superoxide radical anion production [28]. MitochondrialSOD2 rapidly removes superoxide anion radicals that areproduced during oxidative phosphorylation. Therefore, theincreased expression of SOD2 in the liver of fructose-fedanimals most likely represents an adaptation to altered redoxconditions. It is important to point out that, in our study,fructose consumption led to an increased expression of SOD2but rendered the enzyme activity unaffected. This impliesthat animals subjected to a long-term fructose-enriched dietimmediately after weaning have a capacity to adapt their redoxsystem to such a diet. In support of this assumption, total thiolcontent and the level of TBARS (as markers of general redoxconditions and lipid peroxidation, respectively) remainedunaltered in the fructose-fed group as compared with thecontrol group (Table 3). Apart from oxidative stress, long-termfructose consumption was also associated with carbonyl stresscytotoxicity [11]. Previous studies have shown that fructose-induced glycation promotes the generation of reactive oxygenspecies and that reactive oxygen species accelerate the glyca-tion processes, thus suggesting a strong relationship betweencarbonyl and oxidative stress [11]. In this study, we demon-strated that the level ofprotein carbonyls remainedunaltered inthe fructose-fed rats compared to control rats, thereby provid-ing additional support for our finding that fructose overcon-sumption did not induce oxidative stress. Moreover, the level ofmolecular chaperones Hsp70 and Hsp90, the expression ofwhich is up-regulated in response to stress [29], remainedunchained in the fructose-fed group. This suggests that therewas no accumulation of damaged proteins in the liver.
In a similar study thatwas conducted on 90-day-old animalsfed with fructose-enriched chow, it was reported that dietaryfructose provoked lipid peroxidation and decreased SOD2activity in the liver. At the same time, the amount of differentmitochondrial proteins and mitochondrial mass significantlyincreased in the fructose-fed group as compared with thecontrol group [30]. These results imply that adult animalssubjected to a fructose-rich diet meet the increased demands
0
100
200
SOD1 SOD2 CAT GPx GR iNOS
% o
f co
ntro
l
*
β-actin
SOD1
β-actin
SOD2
β-actin
CAT
β-actin
GPx
β-actin
GR
C F C FC FC FC F
iNOS
β-actin
C F
Fig. 2 – Protein levels of antioxidant enzymes and iNOS in the liver of rats subjected to fructose-rich diet immediately afterweaning. Hepatic whole cell extracts (50 μg protein) were subjected to SDS-PAGE andWestern blotting. β-Actinwas used as theloading control. RepresentativeWestern blots and relative quantification of antioxidant enzyme, iNOS protein levels of control(C), and fructose-fed rats (F) are shown. Values are means ± SEM (n = 9) and are presented as percentage of control. Asterisksindicate significant differences, *P < .05 (unpaired Student t test).
β-actin
Hsp70
C F
Hsp90
β-actin
C F
0
20
40
60
80
100
120
Hsp70 Hsp90
% o
f co
ntro
l
C F C F
Fig. 3 – Hsp70 and Hsp90 levels in the liver of rats subjectedto fructose-rich diet immediately after weaning. Hepaticwhole cell extracts (50 μg protein) were subjected to SDS-PAGE andWestern blotting. β-Actinwas used as the loadingcontrol. Representative Western blots and relative quanti-fication of Hsp70 and Hsp90 in the hepatic whole cellextracts of control (C) and fructose-fed rats (F) are shown.Values are means ± SEM (n = 9) and are presented aspercentage of control.
650 N U T R I T I O N R E S E A R C H 3 4 ( 2 0 1 4 ) 6 4 6 – 6 5 2
for ATP bymitochondrial biogenesis, which is unlikely to be themechanism in young animals investigated herein.
In our study, fructose consumption increased SOD2 expres-sion but rendered its activity unaffected. Transcriptionalregulation of SOD2 level has been described in a number ofstudies as an important cellular defense mechanism [31]. Theelevated expression of SOD2 in the liver of fructose-fed animalsmost likely represents an adaptation aimed to protect mito-chondria against elevated superoxide radical anion, which areproduced by enhanced mitochondrial activity. The lack ofcorrelation between SOD2 activity and its protein level couldbe related to inactivation of the enzyme by glycation [32].However, our results show that the level of protein carbonylgroups remained unaltered in fructose-fed compared withcontrol rats, thus suggesting that other mechanisms mightregulate SOD2 activity. It has been reported that tyrosinenitration leads to significant inactivation of SOD2 [33,34].Peroxynitrite, produced by the coupling reaction between nitricoxideandsuperoxideanion radical, causesnitrationofproteins.Our results indicate that theexpressionof iNOSwasnot affectedby fructose consumption, implying involvement of otherposttranslational modifications in the regulation of SOD2function. Perhaps a more plausible explanation may be theregulation of SOD2 activity by acetylation/deacetylation, whichoccurs in response to changes inmitochondrial nutrient and/orredox status. It was found that SOD2 acetylation decreased theenzyme activity, whereas deacetylation by mitochondrialdeacetylase sirtuin-3 or SIRT3 had an opposite effect [35,36].
Most of the previous studies reported alterations in theexpression and activity of antioxidant enzymes in the liver offructose-fed rodents, although the course of these alterationsvaried largely [10,30,37-41]. Some authors observed that afructose-rich diet reduced the antioxidant capacity and caused

651N U T R I T I O N R E S E A R C H 3 4 ( 2 0 1 4 ) 6 4 6 – 6 5 2
oxidative damages in the liver [30,41,42], whereas othersreported the absence of oxidative stress [40] or adaptation tomodest oxidative stress [10] in fructose-fed rats. Alternatively,Girard et al [38] noticed that a fructose-rich diet enhanced thetotal antioxidant capacity of the liver. Our results show that theabsence of oxidative stress increased SOD2 expression withoutchanges in the enzyme activity but no alterations in the activityand expression of other antioxidant enzymes in the liver offructose-fed rats. There are studies reporting antioxidative andcytoprotective effects of fructose [11-14], but it seems thatfructose exerts protective effects only after short-term applica-tion. Its long-term consumption exerts mostly negative effects[11]. Accordingly, antioxidative activity of fructose couldprovoke down-regulation of enzymatic antioxidative systembecause cells need balanced redox milieu to maintain redoxsignaling cascades. Still, such modulations would ultimatelylead to oxidative stress because, in the long run, fructoseantioxidative properties may not match the efficiency ofantioxidant enzymes. In addition, the ability of antioxidantdefense systems to react efficiently under conditions ofdisturbed homeostasis is largely dependent upon age. Most ofthe previous studies reporting pro-oxidative effects of fructosewere performed on adult male individuals, whereas in thisstudy, the animals were 21-day-old juvenile rats at thebeginning of the treatment. In addition, the discrepanciesbetween the results might stem from the differences in thetreatmentduration (ranging from3weeks to17months)and thefructose concentration (ranging from 10% to 60%).
Previously, we stated that a fructose-rich diet appliedimmediately afterweaningaffects insulin sensitivity and induceshypertension, hyperlipidemia, and low-grade inflammation[15,16,43]. This is in line with similar studies showing thatlong-term fructose consumption induces profound metabolicdisturbances [5]. However, in the current study, our hypothesisstating that this diet induces alterations in the hepatic redoxsetting, which may contribute to development and progressionof insulin resistance, was not confirmed. Thus, we can assumethat fructose-provoked impairment of insulin signaling isnot induced by oxidative stress, at least not at a young age. Theunderpinningmolecularmechanismsmight includeapreviouslyobserved increase in adipose tissue lipolysis, followed by anincreased plasma concentration of nonesterified fatty acids[44] and a hepatic low-grade inflammation [16]. Yet, we cannotrule out the possibility that prolonged fructose overconsump-tion might eventually induce oxidative damages and, conse-quently, contribute to the progression of hepatic insulinresistance in later adulthood.
As limitations of our study, we should mention a relativelysmall sample size and a cross-sectional study design. Alongitudinal study might provide better insight into cause-and-effect relationships between fructose overconsumption andredox-related changes in the liver. Furthermore, the fructose-fedanimals inour studywerenot fedaproteinandmicronutrientfortified diet, and they exhibited a reduced food intake. Althoughthere were no visible signs of malnutrition, a possibility offructose-inducedmarginalmicronutrientdeficiency,whichcouldaffect hepatic metabolism, cannot be ruled out.
In conclusion, our results show that a fructose-enricheddiet, applied immediately after weaning, modulates onlySOD2 expression in the rat liver, but it has no effect on the
activity of antioxidant enzymes (including SOD2), lipidperoxidation, total thiol content, and oxidative damage ofproteins. Although previous studies provided evidence of adirect link between oxidative stress and the induction ofinsulin resistance, our results suggest that the decrease inhepatic insulin sensitivity, which was previously observed inthe rats kept on the same diet regime, occurred in the absenceof oxidative stress. Therefore, this might be attributed tomolecular mechanisms other than redox imbalance.
Acknowledgment
Thisworkwas supported by theMinistry of Education, Scienceand Technological Development of the Republic of Serbia,Grant III41009. There are no conflicts of interest regarding thepublication of this article.
R E F E R E N C E S
[1] Thorp AA, Owen N, Neuhaus M, Dunstan DW. Sedentarybehaviors and subsequent health outcomes in adults asystematic review of longitudinal studies, 1996-2011. Am JPrev Med 2011;41:207–15.
[2] Malik VS, Popkin BM, Bray GA, Despres JP, Hu FB. Sugar-sweetened beverages, obesity, type 2 diabetes mellitus, andcardiovascular disease risk. Circulation 2010;121:1356–64.
[3] Ford CN, Slining MM, Popkin BM. Trends in dietary intakeamong US 2- to 6-year-old children, 1989-2008. J Acad NutrDiet 2013;113:35–42.
[4] Tran LT, Yuen VG, McNeill JH. The fructose-fed rat: a reviewon the mechanisms of fructose-induced insulin resistanceand hypertension. Mol Cell Biochem 2009;332:145–59.
[5] Dekker MJ, Su Q, Baker C, Rutledge AC, Adeli K. Fructose: ahighly lipogenic nutrient implicated in insulin resistance,hepatic steatosis, and the metabolic syndrome. Am J PhysiolEndocrinol Metab 2010;299:E685–94.
[6] Tappy L, Le KA, Tran C, Paquot N. Fructose and metabolicdiseases:newfindings,newquestions.Nutrition2010;26:1044–9.
[7] Grattagliano I, Palmieri VO, Portincasa P, Moschetta A,Palasciano G. Oxidative stress-induced risk factors associatedwith the metabolic syndrome: a unifying hypothesis. J NutrBiochem 2008;19:491–504.
[8] Rains JL, Jain SK. Oxidative stress, insulin signaling, anddiabetes. Free Radic Biol Med 2011;50:567–75.
[9] Moreno JA, Hong E. A single oral dose of fructose inducessome features ofmetabolic syndrome in rats: role of oxidativestress. Nutr Metab Cardiovasc Dis 2013;23:536–42.
[10] Francini F, Castro MC, Schinella G, Garcia ME, Maiztegui B,Raschia MA, et al. Changes induced by a fructose-rich diet onhepatic metabolism and the antioxidant system. Life Sci2010;86:965–71.
[11] SemchyshynHM. Fructation in vivo: detrimental and protectiveeffects of fructose. Biomed Res Int 2013;2013:343914.
[12] Frenzel J, Richter J, Eschrich K. Fructose inhibits apoptosisinduced by reoxygenation in rat hepatocytes by decreasingreactive oxygen species via stabilization of the glutathionepool. Biochim Biophys Acta 2002;1542:82–94.
[13] Spasojevic I, Bajic A, Jovanovic K, SpasicM, Andjus P. Protectiverole of fructose in themetabolismof astroglial C6 cells exposedto hydrogen peroxide. Carbohydr Res 2009;344:1676–81.
[14] Semchyshyn HM, Lozinska LM. Fructose protects baker'syeast against peroxide stress: potential role of catalase andsuperoxide dismutase. FEMS Yeast Res 2012;12:761–73.

652 N U T R I T I O N R E S E A R C H 3 4 ( 2 0 1 4 ) 6 4 6 – 6 5 2
[15] Vasiljevic A, Velickovic N, Bursac B, Djordjevic A, MilutinovicDV, Nestorovic N, et al. Enhanced prereceptor glucocorticoidmetabolism and lipogenesis impair insulin signaling in theliver of fructose-fed rats. J Nutr Biochem 2013;24:1790–7.
[16] Velickovic N, Djordjevic A, Vasiljevic A, Bursac B, MilutinovicDV, Matic G. Tissue-specific regulation of inflammation bymacrophage migration inhibitory factor and glucocorticoidsin fructose-fed Wistar rats. Br J Nutr 2013;110:456–65.
[17] Ventura EE, Davis JN, Goran MI. Sugar content of popularsweetened beverages based on objective laboratory analysis:focus on fructosecontent. Obesity (Silver Spring) 2011;19:868–74.
[18] Misra HP, Fridovich I. The generation of superoxide radicalduring the autoxidation of hemoglobin. J Biol Chem1972;247:6960–2.
[19] Claiborne A. Catalase activity. In: Greenwald RA, editor. CRChandbook of methods for oxygen radical research. BocaRaton, FL: CRC Press; 1985. p. 283–4.
[20] Paglia DE, Valentine WN. Studies on the quantitative andqualitative characterization of erythrocyte glutathioneperoxidase. J Lab Clin Med 1967;70:158–69.
[21] Glatzle D, Vuilleumier JP, Weber F, Decker K. Glutathionereductase testwithwholeblood, a convenientprocedure for theassessment of the riboflavin status in humans. Experientia1974;30:665–7.
[22] Rehncrona S, Smith DS, Akesson B, Westerberg E, Siesjo BK.Peroxidative changes in brain cortical fatty acids andphospholipids, as characterized during Fe2+- and ascorbicacid-stimulated lipid peroxidation in vitro. J Neurochem1980;34:1630–8.
[23] Levine RL, Garland D, Oliver CN, Amici A, Climent I, Lenz AG,et al. Determination of carbonyl content in oxidativelymodified proteins. Methods Enzymol 1990;186:464–78.
[24] SongM, Schuschke DA, Zhou Z, Chen T, Pierce JrWM,Wang R,et al. High fructose feeding induces copper deficiency inSprague-Dawley rats: a novel mechanism for obesity relatedfatty liver. J Hepatol 2012;56:433–40.
[25] Douard V, Sabbagh Y, Lee J, Patel C, Kemp FW, Bogden JD, et al.Excessive fructose intake causes 1,25-(OH)(2)D(3)-dependentinhibition of intestinal and renal calcium transport in growingrats. Am J Physiol Endocrinol Metab 2013;304:E1303–13.
[26] Pereira CD, Severo M, Araujo JR, Guimaraes JT, Pestana D,Santos A, et al. Relevance of a hypersaline sodium-richnaturally sparkling mineral water to the protection againstmetabolic syndrome induction in fructose-fedSprague-Dawleyrats: a biochemical, metabolic, and redox approach. Int JEndocrinol 2014;2014:384583.
[27] Song M, Schuschke DA, Zhou Z, Chen T, Shi X, Zhang J, et al.Modest fructose beverage intake causes liver injury and fataccumulation in marginal copper deficient rats. Obesity(Silver Spring) 2013;21:1669–75.
[28] Murphy MP. How mitochondria produce reactive oxygenspecies. Biochem J 2009;417:1–13.
[29] Richter K, Haslbeck M, Buchner J. The heat shock response:life on the verge of death. Mol Cell 2010;40:253–66.
[30] Crescenzo R, Bianco F, Falcone I, Coppola P, Liverini G, Iossa S.Increased hepatic de novo lipogenesis and mitochondrialefficiency in a model of obesity induced by diets rich infructose. Eur J Nutr 2013;52:537–45.
[31] Miao L, St Clair DK. Regulation of superoxide dismutasegenes: implications in disease. Free Radic Biol Med2009;47:344–56.
[32] Jabeen R, Saleemuddin M. Polyclonal antibodies inhibit theglycation-induced inactivation of bovine Cu, Zn-superoxidedismutase. Biotechnol Appl Biochem 2006;43:49–53.
[33] MacMillan-CrowLA, Thompson JA. Tyrosinemodifications andinactivation of active site manganese superoxide dismutasemutant (Y34F) by peroxynitrite. Arch Biochem Biophys1999;366:82–8.
[34] Yamakura F, Taka H, Fujimura T, Murayama K. Inactivation ofhuman manganese-superoxide dismutase by peroxynitrite iscaused by exclusive nitration of tyrosine 34 to 3-nitrotyrosine.J Biol Chem 1998;273:14085–9.
[35] Qiu X, Brown K, Hirschey MD, Verdin E, Chen D. Calorierestriction reduces oxidative stress by SIRT3-mediated SOD2activation. Cell Metab 2010;12:662–7.
[36] Chen Y, Zhang J, Lin Y, Lei Q, Guan KL, Zhao S, et al. Tumoursuppressor SIRT3 deacetylates and activates manganesesuperoxide dismutase to scavenge ROS. EMBO Rep2011;12:534–41.
[37] Cavarape A, Feletto F, Mercuri F, Quagliaro L, Daman G,Ceriello A. High-fructose diet decreases catalase mRNA levelsin rat tissues. J Endocrinol Invest 2001;24:838–45.
[38] Girard A, Madani S, Boukortt F, Cherkaoui-Malki M, BellevilleJ, Prost J. Fructose-enriched diet modifies antioxidant statusand lipid metabolism in spontaneously hypertensive rats.Nutrition 2006;22:758–66.
[39] Kizhner T, Shilovizki O, Werman MJ. Long-term fructoseintake reduces oxidative defense and alters mitochondrialperformance in mice. Nutr Res 2007;27:423–31.
[40] Pasko P, Barton H, Zagrodzki P, Izewska A, Krosniak M, GawlikM, et al. Effect of diet supplemented with quinoa seeds onoxidative status in plasma and selected tissues of highfructose-fed rats. Plant Foods Hum Nutr 2010;65:146–51.
[41] Kannappan S, Palanisamy N, Anuradha CV. Suppression ofhepatic oxidative events and regulation of eNOS expressionin the liver by naringenin in fructose-administered rats. Eur JPharmacol 2010;645:177–84.
[42] Botezelli JD, Cambri LT, Ghezzi AC, Dalia RA, Voltarelli FA, deMelloMA. Fructose-rich diet leads to reduced aerobic capacityand to liver injury in rats. Lipids Health Dis 2012;11:78.
[43] Koricanac G, Djordjevic A, Zakula Z, Vojnovic Milutinovic D,Tepavcevic S,VelickovicN, et al.Gendermodulatesdevelopmentof metabolic syndrome phenotype in fructose fed rats. Arch BiolSci 2013;65:455–64.
[44] Bursac BN, Djordjevic AD, Vasiljevic AD, Milutinovic DD,Velickovic NA, Nestorovic NM, et al. Fructose consumptionenhances glucocorticoid action in rat visceral adipose tissue.J Nutr Biochem 2013;24:1166–72.




















