
Aflatoxin Contamination in Foods and Feeds: A Special Focus on Africa
Upload
independentCategory
view
1download
0
Larval rearing of the Asian Catfish, Pangasius
bocourti (Siluroidei, Pangasiidae): alternative feeds
and weaning time
Le Thanh Hung a,b, Nguyen Anh Tuan c, Philippe Cacot a,Jerome Lazard a,*
aCentre de Cooperation Internationale en Recherche Agronomique pour le Developpement, Departement
Elevage et Medecine Veterinaire/Groupe Aquaculture Continentale Mediterraneenne et Tropicale
(CIRAD–EMVT/GAMET), TA 30/01, 34398 Montpellier cedex 5, FrancebUniversity of Agriculture and Forestry (UAF), Thu Duc, Ho Chi Minh City, Viet Nam
cCan Tho University (CTU), Can Tho City, Viet Nam
Received 25 November 1999; received in revised form 1 July 2001; accepted 7 July 2001
Abstract
Three experiments were carried out to evaluate Pangasius bocourti larval rearing with different
feeding regimes. In the first experiment, survival rates of larvae fed on Artemia nauplii, cladocereans
(Moina sp.), or red-blood worms (Tubifex) were not significantly different, 91–93%. However,
Artemia nauplii and Tubifex gave similar growth rates (35–36% day� 1) while Moina sp. resulted in
lower growth. Commercial trout-starter feed resulted in the lowest survival (67%) and growth rates
(20% day� 1) when compared to live feed.
In the second experiment, feeding dry feed based on yeast and beef liver or feeding decapsulated
Artemia cysts improved survival rates. These were 90% and 87%, respectively, not significantly
different from Artemia nauplii. Regarding growth, dry feed and decapsulated Artemia cysts resulted
in lower growth rates than Artemia nauplii. These were 23% and 29%, respectively. Stomach content
analysis showed that reduced growth in decapsulated-cyst fed larvae compared to live Artemia
reflected a difference in feed ingestion and preference rather than a difference in nutritional quality of
the feed.
0044-8486/02/$ - see front matter D 2002 Elsevier Science B.V. All rights reserved.
PII: S0044 -8486 (01 )00737 -2
* Corresponding author.
E-mail address: [email protected] (J. Lazard).
www.elsevier.com/locate/aqua-online
Aquaculture 212 (2002) 115–127

In the third experiment, it was shown that larvae require 3-day feeding on Artemia nauplii before
weaning them on to trout-starter feed to prevent negative effects on growth. Three days after initial
feeding, fish stomach attains its biological and physiological functions.
D 2002 Elsevier Science B.V. All rights reserved.
Keywords: Larval rearing; Pangasius bocourti; Weaning time; Dry diet; Live feed
1. Introduction
The Asian catfish, Pangasius bocourti Sauvage 1880, is cultured in cages in the
Mekong delta, Vietnam, and annual production is around 27,000 tonnes (Phuong, 1996).
The seed supply has been dependent on catching fry in natural waters. Artificial
propagation and larval rearing represent the major bottleneck for the production and cage
culture of the species. Induced spawning of the species was successfully carried out for the
first time in 1995 (Cacot, 1998). Currently, information on the larval rearing of this species
is lacking.
Brine shrimp nauplii (Artemia sp.) are widely recognized to be an excellent starter feed
for freshwater and marine fish species (Leger et al., 1986), but cyst price is high.
Successful rearing of fish larvae using live zooplankton has been reported for several
species (Watanabe and Fujita, 1983). Among the various species of zooplankton, the genus
Moina (cladocereans) is known to be suitable as an initial feed for Chanos chanos
(Villegas, 1990) and Clarias macrocephalus (Fermin and Bolivar, 1991). Red-blood
worms, Tubifex, have also been used successfully for European catfish (Silurus glanis)
larval rearing (Ronyai and Ruttkay, 1990). The use of decapsulated Artemia cysts rather
than live nauplii, has several advantages: eliminating the inconvenience of producing live
food (Pector et al., 1994), decapsulated cysts are disinfected and have a higher dry weight
and energy content (Vanhaecke et al., 1983). Moreover, they do not leach nutrients into the
water like formulated feed, and their particle size is appropriate for many fish species. As a
consequence, they have been considered as a reference diet for the nutritional study of C.
gariepinus (Verreth et al., 1987). It has also been reported that some freshwater fish
species can be exclusively reared on artificial diets from the start of exogenous feeding.
These include C. gariepinus, Cyprinus carpio and Heterobranchus longifilis (Appelbaum
and Van Damme, 1988; Charlon et al., 1986; Legendre et al., 1995).
The aim of this study was to evaluate different types of live feed, artificial diet and
decapsulated Artemia cysts in term of acceptability, growth and survival rate of P. bocourti
larvae and to determine the optimal weaning time. The development of the digestive canal
of P. bocourti larvae was also considered.
2. Material and methods
Broodfish was induced to spawn by treatment with Human Chorionic Gonadotropin
hormone (HCG). Twenty-four hours after hatching, 500 larvae were placed into each of the
twelve 50-l aquaria. Feeding started from 48 h post-hatching when the yolk sac was not
L.T. Hung et al. / Aquaculture 212 (2002) 115–127116

completely absorbed.Weight and total length at that time ranged from 3.7 to 4.0mg and from
8.7 to 9.0 mm, respectively.
Water flow through the aquaria was maintained constant at 0.4–0.5 l.min � 1 and
aeration was provided. Dissolved oxygen and pH were measured twice a week with a DO
meter (YSI model 518) and a pH meter (Hana HI 8424). Ammonia and nitrite were
measured by colorimetry (Aquaquant 14423, 14424). Temperature, monitored twice a day
at 0800 and 1500 h, ranged from 28 to 30 jC. Dissolved oxygen concentration was always
higher than 5 mg l � 1. The pH level varied from 7.0 to 7.5. Ammonia and nitrite value
ranges were 0.1–0.3 and 0.01–0.04 mg l � 1, respectively.
Three experiments were carried out to evaluate different feeds on growth and survival
rates in P. bocourti larvae and the weaning time was determined using a trout-starter feed.
Each treatment had three replications.
1. The first experiment tested four diets: Artemia nauplii, Tubifex, Moina sp. and trout-
starter feed.
2. The second experiment tested three diets: Artemia nauplii, decapsulated Artemia
cysts and a dry diet based on beef liver and yeast.
3. The third experiment was designed to determine the most suitable weaning time for
early juvenile P. bocourti. These were eight treatments: 0, 1, 2, 3, 4, 5, 6 and A. The
numbers (1, 2, 3. . ., etc.) stand for number of days that larvae were fed with Artemia
nauplii before shifting directly to trout-starter feed. In treatment 0 and A, the larvae were
fed exclusively on dry diet or Artemia nauplii, respectively.
The San Francisco Bay strain Artemia cysts were incubated in 10 ppt saline water for 24
h at 30 jC. The newly hatched Artemia were kept in aerated saline water and used within 12h. The nauplii were 146–250 Am in width and 411–450 Am in total length. Moina were
cultured in earthen ponds fertilized with pig manure. They were collected daily and treated
with formalin (50 ppm) for 1–2 min to eliminate pathogens. They were still alive and
moved actively after treatment. Some other zooplankton organisms were recorded in the
Moina collection, including Eucyclops. Cladocereans were mostlyMoina sp., 288–300 Amwidth and 850–900 Am length. T. tubifex were collected on riverbanks. They were treated
with formalin (50 ppm) for 1–2 min and then chopped into small pieces of 800–900 Amwidth and 900–1000 Am length. Artemia cysts were decapsulated in 25% hypochloride
solution for 5–10 min and then washed in freshwater until the chlorine odor disappeared.
Decapsulated cysts were then kept at 10 jC for daily feeding. The trout-starter diet
(Aqualim, France) used in the first experiment and weaning time trial, had a proximate
analysis of 55% proteins, 18% lipids and 8% minerals. The size of feed particles was 0.2–
0.4 mm. Another dry diet was used in order to improve the survival and growth rates using
beef-liver and yeast with the component and proximate analysis presented in Table 1.
Fish were fed six times a day at 0800, 1200, 1600, 2000, 2400 and 0400 h. Live feed
such as Artemia nauplii, Moina and Tubifex were fed at 160% fish biomass (wet feed
basis), on the basis of fish weight registered every 3 days. The dry diet was distributed at
20% fish biomass and adjusted every 3 days after weighing.
Thirty larvae were randomly sampled every 3 days from each replicate and placed on
paper towels to absorb water and weighed for each replicate as a batch to 0.1 mg
(Kerdchuen and Legendre, 1994). At the end of the experiment, 50 fish were sampled per
aquarium. On each sampling day, five supplementary larvae were also caught 30 min after
L.T. Hung et al. / Aquaculture 212 (2002) 115–127 117

feeding and fixed in formalin solution (10%) for gut content analysis and mouth size
measurement.
Mouth height was the distance from lower jaw to upper jaw in larvae with the mouth
opening at 45j or 90j (Shirota, 1970). Mouth width was taken as the width of the lower
jaw. Measurement was performed under a binocular lens with an accuracy of 0.01 mm.
Survival rates were calculated by taking into account the remaining and discarded larvae.
Dead larvae were removed from aquarium twice a day at 1000 and 2000 h. Cannibalism
rates were defined by
Cannibalism rate ð%Þ ¼ 100� ½Survival rate ð%Þ þ Observed mortality ð%Þ�
The development of the alimentary canal was studied in larvae fed exclusively with
Artemia nauplii. The larvae for this study (10 per samples) were collected at 0, 24, 36, and
48 h post-hatching and on the 3rd, 4th, 5th, 7th and 10th days after hatching. The sample
was fixed in buffered formalin. Hollande–Bouin solution was also used to fix samples for
PAS (Periodic Acid Schiff) staining (Humason, 1972). The samples were then dehydrated,
embedded in paraffin wax and were sectioned at 5–10 Am. Sections was stained with
either Hematoxycline–Eosine or PAS, which is used to identify the mucous materials in
tissues (Humason, 1972). The pH level of gastric fluid was measured by injecting
indicators into the digestive gut: Methyl red acting at the pH range of 4.2–4.6 (color
changed from yellow to red) and Congo red acting at the pH range of 3.3–3.5 (color
changed from orange to red).
Mean weight, specific growth rates and survival rates were subjected to one-way
ANOVA, followed by Duncan’s Multiple Range to test the 95% significant difference
among treatments. Data of survival and cannibalism rates were transformed to arcsine�before ANOVA analysis to have homogeneity of variance.
3. Results
3.1. Comparison between Artemia nauplii, Tubifex, Moina sp. and trout-starter feed
Gut content analysis showed that feeding 48 h after hatching at a temperature of 28–30
jC. At this stage the yolk sac was not completely absorbed. At first feeding, the mouth
height at 45j and 90j was 0.55 and 0.95 mm, respectively. The width of the lower jaw was
1.00 mm (Table 2). It showed that the larvae have mouth size large enough to ingest
different types of natural diets including Moina and Tubifex.
Table 1
Component and proximate analysis of artificial diet based on beef liver and yeast
Component of the diet Proximate composition (dry basis)
‘‘Protibel’’ yeast powder 50% Moisture 7.8%
Beef liver 35% Crude proteins 35.7%
Soybean oil 5% Crude lipids 10.9%
Vitamin premix 5% Ash (minerals) 11.7%
Mineral premix 5% Fiber 6.8%
L.T. Hung et al. / Aquaculture 212 (2002) 115–127118
Mean weight, specific growth rates (SGR) and survival rates at the end of the first
experiment indicated that Artemia nauplii and Tubifex are better diets than Moina and trout
starter (P < 0.05) (Table 3). During the first 3 days of feeding, Artemia nauplii led to the
highest growth performances. Nevertheless, by the 8th day the Tubifex fed larvae caught
up with the growth of larvae fed on Artemia nauplii (Fig. 1).
The survival and cannibalism rates of larvae fed with live feed were not significantly
different. The larvae reared on trout starter had the lowest survival rate (67.5%) and the
highest rates of cannibalism (10.4%). Cumulative mortality (Fig. 2) indicated that larvae
fed on live feed, had high mortality during the first 3 days of feeding (2nd to 5th days),
when compared to later days (from 5th to 11th days). By contrast, artificial feed caused an
increased mortality until the end of the experiment. However, the mortality of larvae fed
with the artificial diet tended to diminish after the 5th day.
3.2. Comparison between Artemia nauplii, decapsulated Artemia cysts, and dry yeast and
beef liver diet
In the second experiment, the SGR of larvae fed Artemia nauplii was 35.4% day� 1
higher than that obtained with decapsulated cysts (28.7% day � 1) or dry diet (22.7%
day � 1). However, the survival rates of fish fed on decapsulated cysts or the dry diet, were
90.4% and 86.6%, respectively, which were not significantly different to those fed on
Artemia nauplii (90.5%). Cannibalism rates were not significantly different among the
three treatments (Table 4 and Fig. 3).
Table 3
Mean weight, specific growth rate and survival rate of P. bocourti larvae fed on artificial diet or live feed (Artemia
nauplii, Moina sp., Tubifex) after 9 days of nursing from D2 to D11
Feeding treatments Artemia nauplii Moina sp. Tubifex Trout starter
Initial weight at D2 (mg) 3.7 3.7 3.7 3.7
Weight at D5 (mg) 13.3F 1.7a 9.9F 1.2b 11.0F 0.4b 7.2F 0.4c
Weight at D8 (mg) 41.5F 4.6a 22.9F 0.7b 37.6F 1.3a 14.6F 1.3c
Final weight at D11 (mg) 100.7F 0.7a 60.8F 5.1b 101.5F 11.7a 24.3F 1.7c
SGR (% day� 1) 36.0F 0.4a 31.0F 0.9b 36.7F 1.3a 20.8F 0.8c
Survival rate (%) 91.7F 5.2a 93.7F 2.4a 92.7F 2.1a 67.5F 13.6b
Cannibalism rate (%) 3.9F 1.5a 1.6F 2.4a 2.6F 1.4a 10.40F 2.6b
Figures in the same line having same superscripts are not significantly different by Duncan’s multiple range test
( P< 0.05). MeanF S.D. (based on three replicates).
SGR= 100� (ln(W2)� ln(W1))/(T2�T1).
Table 2
Mouth size of P. bocourti larvae at first feeding. (48 h post-hatching)
Age from Total length Mouth size (mm)hatching (mm)
Opening
at 45jOpening
at 90jWidth of
lower jaw
48 h (D2) 8.72 0.55 0.95 1.00
72 h (D3) 9.65 0.71 1.19 1.28
96 h (D4) 10.60 0.80 1.25 1.31
Distance from the lower to upper jaw was measured for mouth opening at 45j or 90j.
L.T. Hung et al. / Aquaculture 212 (2002) 115–127 119

3.3. Weaning time and digestive tract development
3.3.1. Weaning time
In the third experiment, the growth of larvae fed on Artemia nauplii were not
significantly different from those weaned to dry feed after 4–6 days Artemia feeding
Fig. 2. Cumulative mortality of P. bocourti larvae in response to live feed or trout-starter feed. Feeding started
48 h post-hatching (D2). Vertical bars = S.D.
Fig. 1. Growth curve of P. bocourti larvae fed on live Artemia nauplii, Moina sp., Tubifex and a trout-starter feed
in the first experiment. Feeding started 48 h post-hatching (D2).
L.T. Hung et al. / Aquaculture 212 (2002) 115–127120

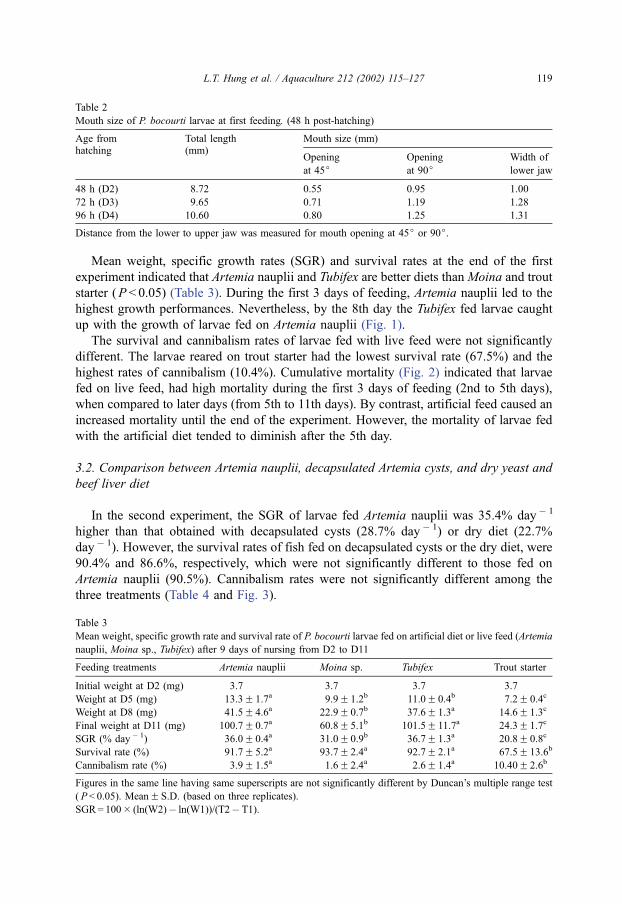
(Fig. 4). However, weaning after 2 days Artemia feeding led to a reduced growth and
fish fed exclusively on dry diets have the lowest growth. In general, the final weight
illustrated a reduced trend if fish were weaned earlier than 3 days Artemia feeding
(Fig. 4).
Concerning survival rates, larvae fed exclusively on a commercial trout diet had the
lowest survival rate (67.2%). Nevertheless, weaning after 1 or 2 days, Artemia feeding
resulted in improving the survival rate (78.0% and 76.0%, respectively). Starting from 3
Fig. 3. Growth curve of P. bocourti larvae fed on Artemia nauplii, decapsulated Artemia cysts or dry diet based on
yeast and beef liver (dry diet 2) in the second experiment. Vertical bars = S.D.
Table 4
Mean weight, specific growth rate and survival rate of P. bocourti larvae fed on Artemia nauplii, decapsulated
Artemia cysts or dry diet based on yeast and beef liver after 9 days of nursing from D2 to D11
Feeding treatments Artemia
nauplii
Decapsulated
Artemia cysts
Dry diet based on
yeast and beef liver
Initial weight (mg) 3.7 3.7 3.7
Final weight (mg) 97.6F 10.7a 53.0F 3.8b 30.9F 2.6c
SGR (% day� 1) 35.3F 2.2a 28.7F 0.8b 22.7F 0.2c
Survival rates (%) 90.5F 4.7a 90.4F 7.4a 86.6F 1.5a
Cannibalism rates (%) 3.2F 4.2a 5.5F 4.8a 7.0F 1.4a
Figures in the same line having same superscripts are not significantly different by Duncan’s multiple range test
( P< 0.05). MeanF S.D. (based on three replicates).
SGR= 100� (ln(W2)� ln(W1))/(T2�T1).
L.T. Hung et al. / Aquaculture 212 (2002) 115–127 121
days Artemia feeding survival rates were consistently high and varied between 88.1% and
96.2%, and they were not significantly different from those fed exclusively on Artemia
nauplii.
3.3.2. Digestive tract development in P. bocourti
The digestive tube is rudimentary and undifferentiated at hatching, and lies on its
yolk sac. Twenty-four hours later, the pancreas and liver differentiated as a clump of
cells in the posterior part of the digestive tract (Fig. 5). At initiation of exogenous
feeding (day 2), the stomach became enlarged with isometric epithelium cells, which are
different from the elongated cells in the intestinal part. Also, at that time, zymogens
were detected in the pancreas, but gastric glands were not yet found in the stomach. Up
to 4 days post-hatching (day 4) these glands were found beneath the gastric epithelium.
On day 5, the section stained with PAS, indicated that the stomach starts secreting
mucopolysaccharides (Fig. 6). Also at that time, the gastric pH level fell to 3.3 by
Fig. 4. Mean weights (mg) and survival rates in P. bocourti larvae weaned to dry diet at different days post-
hatching. Fish were fed either on Artemia nauplii and then shifted directly to a commercial trout-starter feed. The
number (0, 1, 2. . .) in horizontal axis stands for the days that larvae were fed with Artemia nauplii before shifting
directly to trout-starter feed. Treatment ‘‘0’’ and ‘‘A’’ indicate that larvae were fed exclusively on dry diet or
Artemia nauplii, respectively.
L.T. Hung et al. / Aquaculture 212 (2002) 115–127122
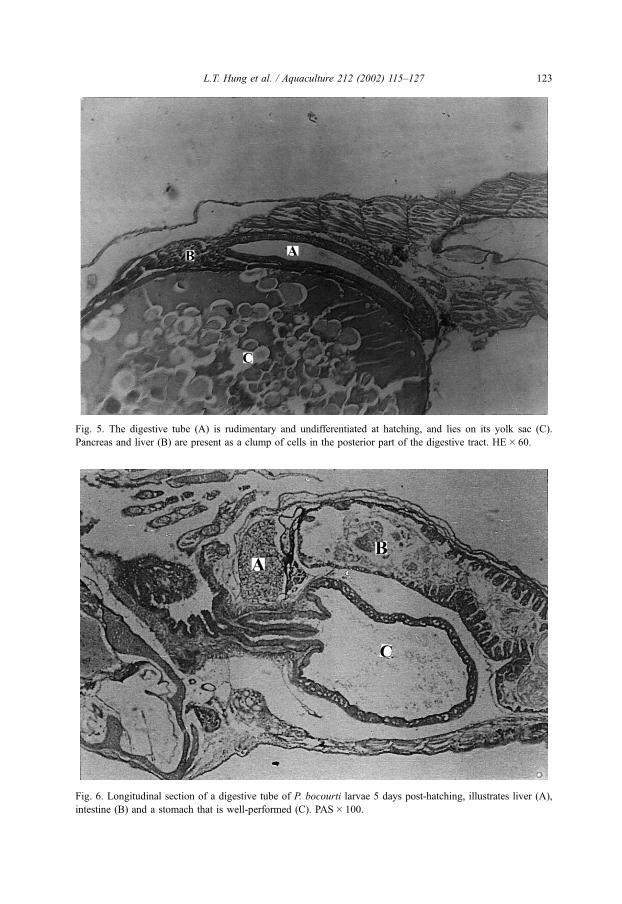
Fig. 5. The digestive tube (A) is rudimentary and undifferentiated at hatching, and lies on its yolk sac (C).
Pancreas and liver (B) are present as a clump of cells in the posterior part of the digestive tract. HE� 60.
Fig. 6. Longitudinal section of a digestive tube of P. bocourti larvae 5 days post-hatching, illustrates liver (A),
intestine (B) and a stomach that is well-performed (C). PAS� 100.
L.T. Hung et al. / Aquaculture 212 (2002) 115–127 123

changing color of Congo red indicator. All these observations indicated that the stomach
then attained its morphological and probably functional completeness.
4. Discussion
4.1. Growth performances and survival rates
In the first and second experiments, Artemia nauplii proved to be an excellent feed in
terms of growth and survival rates. Nevertheless, the survival rate of larvae fed on
Artemia was not significantly different from that fed on Moina or Tubifex, suggesting
that live feeds other than Artemia can be used to rear P. bocourti larvae. Larvae fed on
Moina had lower growth than those fed on Artemia nauplii and Tubifex. The same
conclusion was reported in H. longifilis (Kerdchuen and Legendre, 1994) and in C.
macrocephalus (Fermin and Bolivar, 1991). However, Adeyemo et al. (1994) showed
the opposite for two other catfish, H. bidorsalis and C. gariepinus. Fish fed on M. dubia
had higher growth and survival rates than those fed on Artemia nauplii. In their
experiments the SGRs for Artemia treatment were surprisingly low, 5.1 and 6.1%
day � 1 for a 7-day duration for H. bidorsalis and C. gariepinus, respectively. Numerous
ephippial eggs found in larval stomach content analysis may explain, at least in part, the
lower growth in P. bocourti larvae as ephippial eggs are resistant to digestion in many
fish species (Mellors, 1975). Tubifex have been used economically as a live feed for
nursing European catfish (S. glanis) on large scale in Hungary (Horvath et al., 1981),
but many fish larvae cannot ingest such large prey. Fortunately P. bocourti larvae at
exogenous feeding have a mouth opening large enough permitting the ingestion of
chopped Tubifex, and their growth and survival rates were not different from those fed
on Artemia. Hence, the study confirmed the feasibility of completely replacing Artemia
with Tubifex to nurse P. bocourti larvae.
The lower growth and survival rates of larvae fed on commercial trout-starter feed are
in agreement with findings in other catfish. When fed on trout-starter feed, H. longifilis
larvae (Kerdchuen and Legendre, 1994) and C. gariepinus larvae (Hogendoorn, 1980)
had low survival rates; 32% and 12%, respectively. This may be related to the feed
quality and the digestibility or to the primary development of digestive systems at first
feeding. In the study, P. bocourti apparently showed a better survival rate; 67.5% when
compared to 32% and 12% in the two catfish. That indicates this species has a high
potential to feed on artificial diet. Moreover, yeast and beef-liver dry diet proved
comparable to Artemia nauplii in term of survival rates. The same result was reported in
H. longifilis (Kerdchuen and Legendre, 1994) and in C. gariepinus (Appelbaum and Van
Dame, 1988). The improved survival of dry feed based on yeast and liver could be
attributed to better digestibility of protein. Inactive yeast is roasted at high temperature
during the manufacturing process, thereby, denaturing the proteins to a significant
degree, which in turn facilitates its digestion (Hecht, 1996). Cannibalism was reported in
most larval rearing. Hecht and Appelbaum (1987) demonstrated that cannibalism in C.
gariepinus contributed greater to larval mortality than natural mortality. In C. macro-
cephalus, Fermin and Bolivar (1991) observed a high cannibalism rate in a dry diet
L.T. Hung et al. / Aquaculture 212 (2002) 115–127124
treatment (21.7%), higher than in combination of Artemia with dry diet (9.7%). In the
study the larvae fed trout-starter feed resulted in a higher cannibalism rate (10.4%) than
live food (1.6–4.7%).
Verreth et al. (1987) proposed decapsulated Artemia cysts as a reference diet in larval
nutritional studies. In this study live Artemia nauplii resulted in a higher growth rate than
Artemia cysts. Several workers confirmed a highly nutritional value of decapsulated cysts:
30–50% higher energy than freshly hatched nauplii (Vanhaecke et al., 1983). It is clear
that the nutritive value of feeds does not account for the growth difference observed in
body gain of larvae fed on live Artemia nauplii and on decapsulated cysts. The study
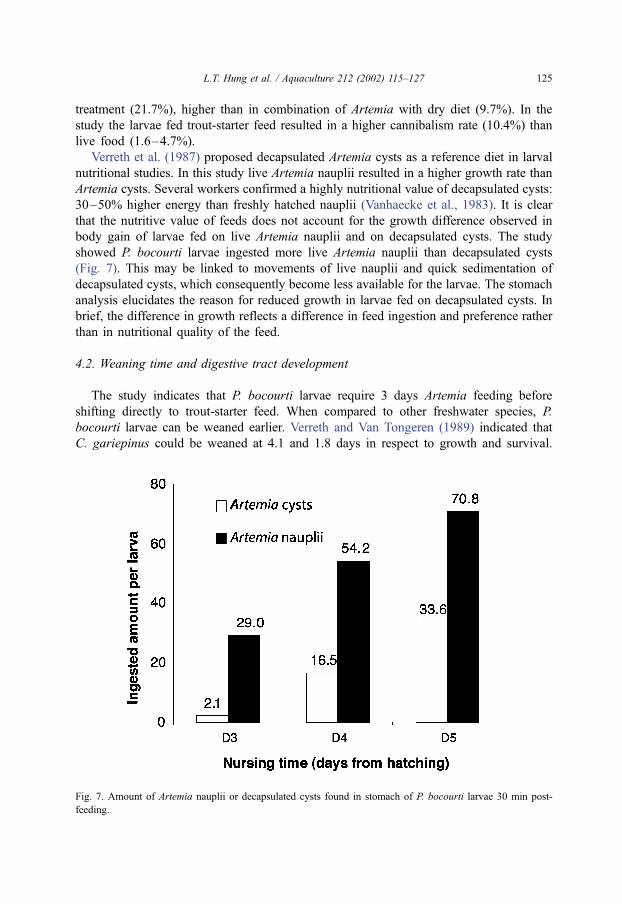
showed P. bocourti larvae ingested more live Artemia nauplii than decapsulated cysts
(Fig. 7). This may be linked to movements of live nauplii and quick sedimentation of
decapsulated cysts, which consequently become less available for the larvae. The stomach
analysis elucidates the reason for reduced growth in larvae fed on decapsulated cysts. In
brief, the difference in growth reflects a difference in feed ingestion and preference rather
than in nutritional quality of the feed.
4.2. Weaning time and digestive tract development
The study indicates that P. bocourti larvae require 3 days Artemia feeding before
shifting directly to trout-starter feed. When compared to other freshwater species, P.
bocourti larvae can be weaned earlier. Verreth and Van Tongeren (1989) indicated that
C. gariepinus could be weaned at 4.1 and 1.8 days in respect to growth and survival.
Fig. 7. Amount of Artemia nauplii or decapsulated cysts found in stomach of P. bocourti larvae 30 min post-
feeding.
L.T. Hung et al. / Aquaculture 212 (2002) 115–127 125

Fermin and Bolivar (1991) reported the weaning time of Asian catfish (C. macro-
cephalus) to be 4 days after live zooplankton feeding. This study shows that P. bocourti
larvae develop a functional stomach earlier than C. gariepinus, which is a possible
reason why the larvae of the species may be weaned onto day feed earlier than other
Siluroidei larvae. Yet, little research has paid attention to the fact that the larval
preference of live feed rather than dry diet is due to the movement of live prey. As a
result, feed intake of the dry diet feeding is constantly lower than that obtained in live
feeding, especially during the initial feeding when larval fins are not yet well developed.
This is possibly an important reason for the unsuitability of the dry diet for larval rearing
in many fish species.
Acknowledgements
CIRAD and IRD supported this work in a co-operative project with Vietnam. The
authors are grateful to Dr. Marc Legendre (IRD), Dr. Pierre Bergot (INRA) for critical
comments on the manuscript.
References
Adeyemo, A.A., Oladosu, G.A., Ayinla, A.O., 1994. Growth and survival of African catfish species, Clarias
gariepinus Burchell, Heterobranchus bidorsalis Geoffroy reared on Moina dubia in comparison with other
first feed sources. Aquaculture 119, 41–45.
Appelbaum, S., Van Damme, P., 1988. The feasibility of using exclusively artificial dry feed for the rearing of
Israeli Clarias gariepinus (Burchell, 1822) larvae and fry. J. Appl. Ichthyol. 4, 105–110.
Cacot, P., 1998. Description of the sexual cycle related to the environment and set up of the artificial propagation
in Pangasius bocourti (Sauvage, 1880) and Pangasius hypophthalmus (Sauvage, 1878), reared in floating
cages and in ponds in the Mekong delta. In: Legendre, M., Pariselle, A. (Eds.), The Biological Diversity and
Aquaculture of Clariids and Pangasiids Catfishes in South-East Asia. Proceedings of the mid-term workshop
of the Catfish Asia project, 11–15 May 1998, Can Tho, Vietnam, pp. 71–90.
Charlon, N., Durante, H., Escaffre, A.M., Bergot, P., 1986. Alimentation artificielle des larves de carpe (Cyprinus
carpio L.). Aquaculture 54, 83–88.
Fermin, A.C., Bolivar, M.E.C., 1991. Larval rearing of the Philippine freshwater catfish, Clarias macrocephalus
(Gunther) fed live zooplankton and artificial diet: a preliminary study. Bamidgeh 43, 87–94.
Hecht, T., 1996. An alternative life history approach to the nutrition and feeding of Siluroidei larvae and early
juveniles. In: Legendre, M., Proteau, J.P. (Eds.), The Biology and Culture of Catfishes. Aquat. Living Resour.
vol. 9, pp. 121–133, Hors Serie.
Hecht, T., Appelbaum, S., 1987. Notes on the growth of Israeli sharptooth catfish during the primary nursing
phase. Aquaculture 63, 195–224.
Hogendoorn, H., 1980. Controlled propagation of the African catfish, Clarias lazera (C&V): III. Feeding and
growth of fry. Aquaculture 21, 233–241.
Horvath, L., Tamas, G., Tolg, I., 1981. European catfish sheatfish (Silurus glanis L.) culture in carp farms. In:
Halver, J.F. (Ed.), Special Methods in Pond Fish Husbandry. Akademiai Kiado, Budapest, pp. 100–123.
Humason, G.L., 1972. Animal Tissues Techniques. Freeman, San Francisco, USA, 641 pp.
Kerdchuen, N., Legendre, M., 1994. Larval rearing of an African catfish, Heterobranchus longifilis (Teleostei,
Clariidae): a comparison between natural and artificial diet. Aquat. Living Resour. 7, 247–253.
Legendre, M., Kerdchuen, N., Corraze, G., Bergot, P., 1995. Larval rearing of an African catfish Heterobranchus
longifilis (Teleostei, Clariidae): effect of dietary lipids on growth, survival and fatty acid composition of fry.
Aquat. Living Resour. 8, 355–363.
L.T. Hung et al. / Aquaculture 212 (2002) 115–127126

Leger, P., Bengtson, D.A., Simpson, K.L., Sorgeloos, P., 1986. The use and nutritional value of Artemia as a food
source. Oceanogr. Mar. Biol. Annu. Rev. 24, 521–623.
Mellors, W.K., 1975. Selective predation of ephippial Daphnia and the resistance of ephippial eggs to digestion.
Ecology 56, 974–980.
Pector, R., Tackaert, W., Abelin, P., Olivier, F., Sorgeloos, P., 1994. A comparative study on the use of different
preparations of decapsulated Artemia cysts as food for rearing African catfish (Clarias gariepinus) larvae. J.
World Aquacult. Soc. 25, 366–370.
Phuong, N.T., 1996. On-farm prepared feed and feeding regimes for the Pangasius catfish (P. bocourti) cultured
in cages in the Mekong River in Vietnam. Proceedings of EIFAC Workshop on Fish and Crustacean Nutrition
Methodology and Research for Semi-intensive Pond-based Farming Systems, 3–5 April 1996. Szarvas,
Hungary.
Ronyai, A., Ruttkay, A., 1990. Growth and food utilization of Wels fry (Silurus glanis) fed with Tubifex.
Aquacult. Hung. (Szarvas) VI, 193–202.
Shirota, A., 1970. Study on the mouth size of the fish larvae. Bull. Jpn. Soc. Sci. Fish. 36, 353–368.
Vanhaecke, P., Rutten, H., Ollevier, F., 1983. Energy consumption in cysts and early larval stages of various
geographical strains of Artemia. International Study on Artemia XVII. Ann. Soc. R. Zool. Belg. 113,
155–164.
Verreth, J., Van Tongeren, M., 1989. Weaning time in Clarias gariepinus (Burchell) larvae. Aquaculture 83,
81–88.
Verreth, J., Storch, V., Segner, H., 1987. A comparative studies on the nutritional quality of decapsulated Artemia
cysts, micro-encapsulated egg diets and enriched dry feeds for Clarias gariepinus (Burchell) larvae. Aqua-
culture 63, 269–282.
Villegas, C.T., 1990. The effects on growth and survival of feeding water fleas (Moina macrocopa) and Rotifer
(Brachionus plicatilis) to milkfish (Chanos chanos Forsskal) fry. Bamidgeh 42, 10–17.
Watanabe, T.C., Fujita, S., 1983. Nutritional values of live organisms used in Japan for mass production of fish: a
review. Aquaculture 34, 115–143.
L.T. Hung et al. / Aquaculture 212 (2002) 115–127 127
Copyright © 2022 FDOKUMEN




















