
Cenomanian-Turonian rudists from Western Sinai, Egypt: Systematic paleontology and paleoecology
Upload
midwesternCategory
view
1download
0
645
Journal of Vertebrate Paleontology 22(3):645–660, September 2002q 2002 by the Society of Vertebrate Paleontology
LIZARDS OF THE MUSSENTUCHIT LOCAL FAUNA (ALBIAN–CENOMANIAN BOUNDARY)AND COMMENTS ON THE EVOLUTION OF THE CRETACEOUS LIZARD FAUNA OF
NORTH AMERICA
RANDALL L. NYDAM*Oklahoma Museum of Natural History, 2401 Chautauqua Ave., Norman, Oklahoma, 73072; and Oklahoma City Community
College, 7777 South May Ave, Oklahoma City, Oklahoma, 73159
ABSTRACT—A diverse fauna of lizards has been recovered from the Cedar Mountain Formation (Albian–Cenomanianboundary) of central Utah. These lizards are a component of the now recognized Mussentuchit local fauna and, likemany of the other vertebrate groups within this fauna, show striking similarities to their North American Late Cretaceous(Campanian–Maastrichtian) counterparts. Unlike the Late Cretaceous lizard faunas, the lizards from the Mussentuchitlocal fauna also include at least one ‘‘paramacellodid,’’ a group common to Late Jurassic and Early Cretaceous lizardfaunas of North America. Comparison with other Cretaceous lizard faunas from both North America and Asia indicatesthat the changes in the North American lizard fauna represented by the lizards of the Mussentuchit local fauna arelikely the result of the introduction of taxa from Asia during the Early Cretaceous. Supposedly herbivorous taxa arefound among the Polyglyphanodontine lizards from the Late Cretaceous of North American and Asia, but the generalchanges in the Cretaceous lizard fauna of North America cannot be convincingly shown to be linked to the concurrentangiosperm radiation. Later influxes of Asian lizard taxa prior to the Campanian may explain the relatively suddenappearance of additional groups of lizards in the early Campanian of North America. Whereas there is good evidenceto support the hypothesis of an Asian influence on the Cretaceous lizard fauna of North America, there is insufficientevidence to determine what, if any, influence may have come from Europe or South America.
INTRODUCTION
The North American record of Mesozoic lepidosauromor-phans includes intermittent fossil horizons from the Late Juras-sic through the K/T boundary. Within this time frame two basicfaunal arrangements have been identified. The first is an older,more primitive fauna composed primarily of sphenodontians,‘‘paramacellodid’’ scincomorphs, and primitive anguimorphansfrom the Jurassic (Kimmeridgian) Morrison Formation (Proth-ero and Estes, 1980; Evans, 1993, 1996; Evans and Chure,1998a, b, 1999; Fraser and Wu, 1998). The composition of thisfauna remained relatively stable through the Early Cretaceous(Aptian–Albian), as indicated by lepidosaurians from: the Trin-ity Group of Texas (Winkler et al., 1990); Cantera Tlayua, Mex-ico (Reynoso, 1995, 1996, 1997, 1998); and the Antlers andCloverly formations of Oklahoma and Montana/Wyoming, re-spectively (Nydam and Cifelli, 2002a). The most notablechanges after the Kimmeridgian and before the Aptian–Albianare the reduction in the presence of both sphenodontians (re-stricted to single occurrences in the Albian of Montana andCantera Tlayua, Mexico; Throckmorton et al., 1981; Reynoso,1997, respectively) and anguimorphans (possibly present in theAntlers Formation; Nydam and Cifelli, 2002a) and the appear-ance of the earliest representatives of Teiidae (Winkler et al.,1990; Nydam and Cifelli, 2002a). In both the Kimmeridgianand Aptian–Albian faunas, however, the major taxonomic com-ponent consists of paramacellodid or ‘‘paramacellodid’’-likescincomorphan lizards (Nydam and Cifelli, in press a).
Following the Aptian–Albian, there is a 301 million yearperiod for which little is known about the North American liz-ard fauna. The second faunal arrangement of the Mesozoic isthe younger (Campanian–Maastrichtian), more derived fauna,which is composed of a wide diversity of taxa referable to nu-
*Present address: Arizona College of Osteopathic Medicine, Mid-west-ern University, 19555 N. 59th. Ave., Glendale, Arizona, email: [email protected]
merous modern groups. These groups include Anguidae, Cor-dylidae, Monstersauria (clade of platynotans that includes Hel-odermatidae, Norell and Gao, 1997), Iguanidae, Scincidae, Tei-idae, Xenosauridae, Xantusiidae, and Varanidae (Estes 1964,1983a; Gao and Fox, 1996). Of these groups, the greatest di-versity is found in the Teiidae, both polyglyphanodontine (Gil-more, 1940, 1943b) and non-polyglyphanodontine (sensu Den-ton and O’Neill, 1995) taxa. Anguids (particularly Odaxosau-rus) also are commonly recovered from most Late Cretaceouslocalities in North America (Estes, 1964, 1969, 1983a; Sullivan,1981; Rowe et al., 1992; Gao and Fox, 1996). Between theCampanian and the Maastrichtian there was little in the way oftaxonomic changes in the lizard fauna (sphenodontians are notfound in North America after the Albian), but there was anapparent loss of diversity. There are 37 lizard taxa (named andunnamed) reported from Campanian-aged strata in North Amer-ica (Estes, 1983b; Rowe et al., 1992; Denton and O’Neill, 1995;Gao and Fox, 1996; McCord, 1998; Eaton et al., 1999; Nydam,1999) versus 20 taxa from Maastrichtian-aged strata (Estes,1983b; Gao and Fox, 1996). Although possibly a taxonomicartifact, a similar loss of diversity in lizards between the Cam-panian and Maastrichtian has been identified for the lizards ofMongolia (Alifanov, 1993a) indicating that the trend was glob-al. Lehman (1997) suggested that at least during the Campanianthere also were latitudinal differences in the lizard fauna, asindicated by a greater diversity of lizard taxa in more southerlydeposits. However, additions to the record of lizards from theCampanian of Canada (Gao and Fox, 1996) indicate that thelatitudinal trend, if real, may be opposite of that proposed byLehman (1997).
Comparison of these two faunal arrangements shows a majorshift in the composition of lepidosaurian taxa between the Ap-tian–Albian and the Campanian–Maastrichtian. However, thefossil record of lizards from North America between these ho-rizons is sparse, giving little indication of the specifics (e.g.,timing, pace) of this change. Some evidence suggests thatchanges had begun in at least two lineages, polyglyphanodon-

646 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 22, NO. 3, 2002
FIGURE 1. Locality map of lizard-producing localities from Mussen-tuchit Member of Cedar Mountain formation (gray stippling) in EmeryCounty, Utah.
tine teiids and monstersaurian (helodermatid-like) platynotans,by the Albian–Cenomanian (Nydam, 1999, 2000). However, itis not clear when other lineages appeared and whether therewas a rapid or gradual transition between the primitive andderived faunas.
Herein I describe the lizards of the Mussentuchit local faunafrom the Cedar Mountain Formation (Albian–Cenomanianboundary), Emery County, Utah. This fauna is important in thatit is temporally intermediate between the Late Jurassic and LateCretaceous faunas, and has been well sampled (Cifelli et al.,1999). I make comparisons to both older and younger lizardfaunas of North America to examine how the Mussentuchit liz-ards fit into the transition between the primitive and derivedfaunas. I also examine the relationship of the Mussentuchit liz-ards to the temporally relevant lizard faunas of Asia, whichhave been previously considered to be a possible source of thetaxonomic diversity of lizards from the Late Cretaceous ofNorth America (Estes, 1983a, 1983b).
Institutional Abbreviations MNA, Museum of NorthernArizona, Flagstaff; OMNH, Sam Noble Oklahoma Museum ofNatural History, Norman.
GEOLOGIC SETTING
The Cedar Mountain Formation is comprised of variegated,terrigenous rocks that are exposed in central and eastern Utah,continuing on into western Colorado as the laterally equivalentBurro Canyon Formation (Stokes, 1944, 1952; Tschudy et al.,1984; Kirkland et al., 1997; Currie, 1998). The lower memberof the Cedar Mountain Formation is the massive, but discontin-uous, sand–pebble Buckhorn Conglomerate (Stokes, 1944,1952). The upper mudstones of the Cedar Mountain Formationhave been subdivided into four additional members based onlithological and paleontological differences (Kirkland et al.,1997). Microvertebrate-producing localities (Fig. 1) are knownonly from the uppermost, or Mussentuchit Member, of whichmajor exposures lie principally on the west side of the SanRafael Swell, a prominent anticline in the region. The Mussen-tuchit Member is 126 m thick at its type locality (Stokes, 1952),but is variable in thickness throughout the region. It is com-posed of highly smectitic gray–green mudstones with interbed-ded channel sandstones (Kirkland, 1987; Cifelli et al., 1999).The environment of deposition of the Mussentuchit Member isinterpreted as having been overbank, or crevasse splay, depositsassociated with multiple flood events in a meandering river sys-tem (Nelson and Crooks, 1987). Microvertebrate localities oc-cur in a narrow stratigraphic interval in the upper part of theunit, 10–20 m below the contact with the overlying DakotaFormation. Multiple, concordant radiometric determinationsfrom two of the localities (in direct association with the fossilhorizon at each) yield an age of 98.39 6 0.07 Ma for the fauna(Cifelli et al., 1997), placing the fauna at the Albian–Cenoman-ian (Early–Late Cretaceous) boundary (sensu Gradstein et al.,1995). This horizon has also been referred to informally as themedial Cretaceous (Cifelli and de Muizon, 1997; Cifelli andMadsen, 1998, 1999; Cifelli et al., 1999) as a method to suc-cinctly describe the horizon as the boundary between the Earlyand the Late Cretaceous. This convention is followed hereinwhen discussing comparisons between this and other horizons.
TAXONOMIC INFORMATION
As outlined in Nydam and Cifelli (2002a) the status of thelizard family ‘‘Paramacellodidae’’ is uncertain. To indicate thisuncertainty the name ‘‘Paramacellodidae’’ will be used inquotes.
SYSTEMATIC PALEONTOLOGY
SQUAMATA Oppel, 1811SCINCOMORPHA Camp, 1923
SCINCOIDEA Oppel, 1811‘‘Paramacellodidae’’ Estes, 1983
cf. PSEUDOSAURILLUS Hoffstetter, 1967
cf. Pseudosaurillus sp.(Figs. 2, 9AA)
Referred Specimen OMNH 27711, broken right maxilla.Locality and Horizon OMNH locality V695; 10–20 m be-
low upper contact of Mussentuchit Member of the Cedar Moun-tain Formation and Dakota Formation, Emery County, Utah.
Description OMNH 27711 is the posterior portion of aright maxilla, preserving 10 teeth. The exterior surface of the

647NYDAM—CRETACEOUS LIZARDS
FIGURE 2. OMNH 27711, broken right maxilla of cf. Pseudosauril-lus sp. Scale bar equals 1 mm.
element is smooth and unornamented. There are three superioralveolar foramina above the lateral parapet. The anterior twoforamina open anterolaterally and the posterior foramen opensposterolaterally. Internally, the posterior interior alveolar fora-men opens posteriorly above the sixth preserved tooth position.The facet for the maxillary process of the jugal is a narrow slitrestricted by the dorsally swollen supradental shelf to the lateraledge of this shelf. The supradental shelf is well developed andwide, but not very deep. The supradental gutter, which is nar-row and shallow, does not extend posterior to the posterior in-terior alveolar foramen. The teeth are homodont, columnar,closely spaced, and strongly pleurodont (only the very tips ofthe teeth extend beyond the lateral parapet). The tips of theteeth are rounded and very weakly recurved. However, thecrowns are not preserved well enough to determine if the me-dial surfaces were striate. Deep replacement pits at the basesof the teeth indicate that tooth replacement was direct.
Comments Pseudosaurillus was first described from thePurbeck Limestone Formation of England (Hoffstetter, 1967)and placed in the ‘‘Paramacellodidae’’ by Estes (1983b) basedon similarities to Paramacellodus and Becklesius in generalform. Evans and Chure (1998a) noted that the teeth of Pseu-dosaurillus are similar to those of Paramacellodus in havingmedial facetting of the apex. The teeth of OMNH 27711 arepoorly preserved and no facetting or striae can be seen, butthey otherwise fit the same description as for Pseudosaurillusin that they are narrow, cylindrical, closely spaced, and lowcrowned. In the absence of more diagnostic material, I verytentatively refer this specimen to ‘‘Paramacellodidae’’ as cf.Pseudosaurillus. If correct, this assignment would extend therange of this genus from the Berriasian to the Albian–Ceno-manian.
SCINCOIDEA Oppel, 1811Family indet.
DIMEKODONTOSAURUS, gen. nov.
Etymology From the Greek words ‘‘di’’ meaning double,‘‘mekos’’ meaning length, ‘‘odous’’ meaning tooth; and ‘‘sau-ros’’ meaning lizard. Named in reference to the distinct differ-ence in tooth height between the middle and the ends of thetooth row.
Type and Only Known Species Dimekodontosaurus mad-seni, sp. nov.
Diagnosis As for type and only known species.
Dimekodontosaurus madseni, sp. nov.(Figs. 3, 9V)
Etymology In recognition of Scott Madsen who has skill-fully prepared the type of this and many other lizards and mam-mals from the Cedar Mountain Formation.
Holotype OMNH 34560, left dentary.
Referred Specimens OMNH 22105–22106, 28257, jawfragments; 28733, dentary fragment; 28814, partial left maxilla;29634, partial left dentary; 30207, partial right dentary; 32625–32627, 32634–32635, 33431, 33449, jaw fragments; 34558,broken right maxilla; 61243, osteoderm; 61244, osteoderm;61806, jaw fragment.
Localities and Horizon OMNH localities V239, V240,V695 (type locality), and V868; 10–20 m below upper contactof Mussentuchit Member of Cedar Mountain Formation andDakota Formation, Emery County, Utah.
Diagnosis Differs from other known cordyliform scinco-morphs in having distinctly different sizes of teeth along toothrow such that anterior and posterior teeth much taller than teethin middle part of tooth row.
Description OMNH 34560 (Fig. 3A–C) is a complete leftdentary of Dimekodontosaurus madseni with 27 tooth positionsand 23 teeth preserved. The symphysis forms a broad surface,extending only under the anteriormost two tooth positions. Thesubdental gutter is narrow and shallow, and runs the length ofthe tooth row. The subdental shelf is tall anteriorly, but narrowssubstantially posteriorly and turns sharply posterodorsally be-neath the posteriormost three tooth positions. Based on articu-lation scars, an anteromedial process from the coronoid extend-ed anteriorly beneath the posteriormost three tooth positions,separating the splenial from the subdental shelf. The articulationfacet for the splenial on the dentary indicates that the splenialextended anteriorly to end beneath the 9th or 10th tooth positions,leaving a narrow, slit–like symphysial foramen between thesplenial and the symphysis. A lateral lappet of the coronoidonto the dentary is absent as indicated by the lack of a faceton the external edge of the posterior portion of the dentary. Thevery tip of the posterodorsal process of the dentary is broken,and it is not possible to determine if there was a lateral lappetof the dentary onto the coronoid. Within the Meckel’s canal,below the 19th and 20th tooth positions, is the posteriorly di-rected opening of the posterior inferior interior alveolar fora-men. Posterior to this foramen is a remnant of the surangularstill articulated to the surface of the internal lateral wall of thedentary. Along the dorsal edge of this articulation is a smallridge that extends from the roof of the Meckel’s canal and pos-teriorly to the end of the tooth row. Just posterior to the openingof the posterior inferior interior alveolar foramen this ridgeforms a prominent and narrow septum, tapers to short ridge,and becomes a prominent septum again for the rest of its length.The dorsomedial edge of the surangular abuts against this ridge.The exterior surface of the dentary is strongly convex.
The teeth all have a medially directed cuspule (sensu Estes,1983b) at the apex and striae on the lingual surface of thecrowns. There are two well-developed lingual striae that extendfrom the medial cuspule. The anterior and posterior carinae ofeach tooth meet at the apex to form an inward pointing ‘‘V’’in occlusal view. With the exception of the posteriormost twoteeth, the crowns of the teeth are rotated posteriorly such thatthe medial cuspule is directed approximately 308 posteromedi-ally. The last two teeth have almost no rotation of the crown.The anterior 10 teeth (first 12 tooth positions) of the dentaryare tall, narrow, recurved, and have sharp anterior and posteriorcarinae. The 14th tooth is slightly shorter than the 12th tooth(tooth 13 is missing) and has a much wider base than the an-terior teeth. Teeth in positions 15–22 are all blocky teeth thatare distinctly shorter than either the anterior teeth or the teethin the last five tooth positions. Teeth in positions 23–25 aremuch taller and more massive than the seven preceding teeth.Teeth in positions 26 and 27 are not as massive and are muchshorter, but owing to the upward curvature of the tooth-bearingportion of the dentary, their crowns are at the same height asthe preceding three teeth. There are basal replacement pits attooth positions 9, 11, and 27, and missing teeth in positions 1,

648 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 22, NO. 3, 2002
FIGURE 3. Dimekodontosaurus madseni, gen. et sp. nov. A–C, OMNH 34560, holotype left dentary; A, medial view (arrow indicates scar forarticulation of anteromedial flange of coronoid); B, detail of anterior portion of tooth row; C, detail of posterior portion of tooth row. D, scanningelectron micrograph of representative anterior teeth on OMNH 32625; E, scanning electron micrograph of representative posterior teeth on OMNH32626. Scale bars equal 1 mm unless otherwise noted.
FIGURE 4. Other specimens referred to Dimekodontosaurus madseni, gen. et sp. nov. A, medial view of OMNH 28814, broken left maxilla;B, medial view of OMNH 34558, broken right maxilla; C, dorsal view of OMNH 61243, broken osteoderm; D, dorsal view of OMNH 61244,broken osteoderm. Scale bars equal 1 mm.
10, and 13. The replacement pit at tooth position 27 containsthe crown of a developing replacement tooth.
OMNH 32625 and 32626 (Fig. 3D, E) are isolated jaw frag-ments of Dimekodontosaurus madseni that exhibit anterior andposterior tooth morphologies, respectively. OMNH 28814 (Fig.4A) is a partial left maxilla, preserving most of the posterior
maxillary tooth row. The teeth of OMNH 28814 are subpleu-rodont to pleurodont, moderately spaced with bases that arewider than the crowns. The tooth crowns are without medialstriae, and have no distinct carinae, but do have medially di-rected cuspules on the apices. None of the tooth crowns arerotated. Of the six teeth preserved in OMNH 28814, the third

649NYDAM—CRETACEOUS LIZARDS
FIGURE 5. OMNH 28119, holotype left dentary of Bothriagenysmysterion, gen. et sp. nov. A, medial view; B, detail of teeth; C, sketchof specimen. Note narrow opening of Meckel’s groove. Scale bars 1mm.
(counting from anteriormost tooth) is distinctly larger than theother five. The fourth tooth has a large replacement pit at itsbase. This specimen can only be tentatively referred to D. mad-seni because of the lack of lingual striae and anterior and pos-terior carinae on the teeth.
OMNH 34558 (Fig. 4B) is a broken right maxilla preservingtwo teeth from the posterior portion of the maxillary tooth row.Its teeth are subpleurodont, and the posteriormost tooth is dis-tinctly larger than the preceding one. A broken tooth base an-terior of the small tooth indicates this tooth also was relativelysmall. Neither of the complete teeth has discernable striae, butwell-defined, delicate, anterior and posterior carinae are presenton the crowns. Both tooth crowns are similar to those of thedentary in having sharp apices that form inward pointing ‘‘Vs’’in occlusal view. Also like the teeth of the dentary, the crownof the smaller tooth is rotated nearly 308, but the large tooth isrotated nearly 608. There is a small replacement pit at the baseof the smaller tooth. As with OMNH 28814, OMNH 34558 canonly be tentatively referred to D. madseni because of the lackof lingual striae on the teeth.
OMNH 61243 and 61244 (Fig. 4C, D) are imbricating os-teoderms. They are heavily built with a central ridge and aresculptured with a pattern of small pits.
Comments The dentary of D. madseni is almost certainlythat of a scincomorphan with cordyliform affinities, as indicated(sensu Estes et al., 1988) by the large anteromedial process ofthe coronoid that articulated below the posterior portion of thetooth row and the lack of an anterolateral process of the coro-noid. The morphology of the teeth, particularly the striae, cus-pule, and carinae of the crowns, is very much like that of theteeth of some taxa of ‘‘paramacellodids’’ and modern cordylids.However, no known ‘‘paramacellodid’’ (see figures in Estes,1983b; Evans and Chure, 1998a, 1999) or cordylid (Edmund,1969) has a dentition with the heterodonty, particularly withrespect to tooth height, as observed in D. madseni. Withoutmore complete materials it is not possible to make a familialassignment for D. madseni. Sakurasaurus shokawensis from theEarly Cretaceous of Japan (Evans and Manabe, 1999) is a scin-comorphan with heterodonty similar to that of Dimekodonto-saurus in that the anterior teeth are gracile and recurved andthe posterior teeth are blunt and striated. However, all of theteeth in the dentary of Sakurasaurus are the same height. Thereduced height of the teeth in the middle of the tooth row inthe dentary does not occur in any other known scincoids, buta similar condition is found in the maxillae of some Recentteiioids, lacertids, and iguanids (Estes et al., 1988). A similarcondition has been described for the maxillary tooth row of theLate Cretaceous iguanian Cnephasaurus (Gao and Fox, 1996).Such a taxonomically widespread occurrence of this featuresuggests that it may represent a feeding specialization (Gao andFox, 1996) that arose independently in several different lizardlineages.
The osteoderms are referred to Dimekodontosaurus based ontheir similarity to the osteoderms of other ‘‘paramacellodid’’and cordylid scincomorphs. Also, of the other taxa in the faunathat could have possibly been armored, only Dimekodontosau-rus appears to have been large enough to possess osteodermsof the size and structure of OMNH 61243 and 61244.
cf. SCINCOMORPHA Camp, 1923Family indet.
Bothriagenys, gen. nov.
Etymology From the Greek words ‘‘bothrion’’ (diminutivefor ‘‘bothros,’’ meaning trench) and ‘‘genys,’’ meaning jaw. Inreference to narrow Meckel’s groove along length of dentary.
Type and Only Species Bothriagenys mysterion.Distribution Mussentuchit Member of Cedar Mountain
Formation (Albian–Cenomanian boundary), Emery County,Utah.
Diagnosis As for type and only known species.
BOTHRIAGENYS MYSTERION, sp. nov.(Figs. 5, 9J)
Etymology From the Greek word ‘‘mysterion,’’ meaningmystery. In reference to uncertain familial affinities of new tax-on.
Holotype OMNH 28119, left dentary.Hypodigm Holotype and OMNH 22118, dentary fragment.Locality and Horizon OMNH localities V695 (OMNH
22119) and V868 (OMNH 22118); 10–20 m below upper con-tact of the Mussentuchit Member of Cedar Mountain Formationand Dakota Formation, Emery County, Utah.
Diagnosis Meckel’s groove moderately wide at posteriorend of dentary, but open as narrow slit for most of length ofdentary; teeth tall, narrow, conical, closely spaced, crownsweakly medially striate, with short carinae wrapping ventro-medially around crown; differs from Recent scincids (e.g., Eu-meces), cordylids (e.g., Cordylus), in more posterior narrowingof Meckel’s groove. Differs from Penemabuya (Lancian; Gaoand Fox, 1996) in having more fully developed carinae on toothcrown, teeth set higher on lateral parapet of dentary, and an-terior end of dentary that points anteriorly rather than antero-dorsally.
Description OMNH 28119 (Fig. 5) is a left dentary that islightly built, with 16 tooth spaces, only the last two of which

650 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 22, NO. 3, 2002
FIGURE 6. Teiidae of Mussentuchit local fauna. A, OMNH 33889,holotype right dentary of Harmodontosaurus emeryensis, gen. et sp.nov; B, detail of second to last tooth of holotype dentary of Harmo-dontosaurus emeryensis, gen. et sp. nov; C, OMNH 29771, holotypetooth of Dicothodon moorensis, arrow indicates shoulder-like anteriorprocess of crown; D, OMNH 26743, holotype left maxilla of Bicuspidonnumerosus (after Nydam and Cifelli, 2002b). Scale bars equal 1 mm.
have complete teeth. Comparison to lizards with similar dentarymorphology indicates that only the last three to four teeth aremissing. The exterior surface is smooth with five inferior al-veolar foramina opening below the lateral parapet. The subden-tal gutter is narrow but well developed along the entire toothrow. The subdental shelf is well developed and maintains auniform height along the tooth row except at the last two toothpositions, where it narrows in concert with the widening of theMeckel’s groove. The Meckel’s groove is open ventrally as anarrow slit along most of the preserved portion of the dentaryand widens only at the posteriormost part of the element, whereit turns somewhat medial. The anterior extent of the splenialwould have terminated posterior to the midpoint of the toothrow. The teeth appear to have been homodont, as indicated bythe uniform shape and size of the tooth bases, but this cannotbe confirmed without more complete material. The posterior-most two teeth are conical, narrow, and tall, with bases slightlycompressed anteroposteriorly, and crowns slightly laterallycompressed and shallowly medially concave. The medial sur-faces of the crowns are weakly striate and bordered by twoshort carinae that wrap ventromedially around the crown. Deepreplacement pits at the bases of the teeth indicate direct replace-ment.
Comments The presence of a well-developed subdentalgutter and shelf in OMNH 28119 suggests that this taxon isclosely allied to scincomorphans (Gao and Fox, 1996:74). Iconsider the presence of an open Meckel’s groove as furthersupport for such an affiliation. Among scincomorphans, a re-duced anterior extent of the splenial and slit-like opening of theMeckel’s canal is not found in any known ‘‘paramacellodid’’taxa (Estes, 1983b), but is present in some taxa of Cordylidaeand Scincidae (Estes et al., 1988). Of these last two familiesthere are no features preserved on the specimens of Bothri-agenys that diagnose the genus or clearly indicate to whichfamily it might belong. Pending the recovery of more diagnosticmaterial, Bothriagenys can only tentatively be referred to Scin-comorpha (sensu Estes et al., 1988).
TEIIDAE Gray, 1827
HARMODONTOSAURUS, gen. nov.
Etymology From the Greek words ‘‘harmos,’’ meaningshoulder, ‘‘odous,’’ meaning tooth, and ‘‘sauros,’’ meaning liz-ard. Named for the shoulder-like anterior expansion of teeth.
Type and Only Species Harmodontosaurus emeryensis,sp. nov.
Distribution Mussentuchit Member of Cedar MountainFormation (Albian–Cenomanian boundary), Emery County,Utah.
Diagnosis As for type and only known species.
HARMODONTOSAURUS EMERYENSIS, sp. nov.(Figs. 6A, B, 9R)
Etymology Named in reference to Emery County, Utah,where holotype was found.
Holotype OMNH 33889, right dentary broken posteriorly.Referred Specimens OMNH 22109, partial left maxilla;
27753, 28067, partial right dentaries; 28068, 32627, 60195, jawfragments.
Locality and Horizon OMNH localities V868 (type local-ity), V240, and V695, 10–20 m below upper contact of Mus-sentuchit Member of Cedar Mountain Formation and DakotaFormation, Emery County, Utah.
Diagnosis Differs from other known Cretaceous teiids (Gaoand Fox, 1991, 1996) in following combination of characters:anterior teeth gracile, posterior teeth comparatively much more
robust, teeth more closely spaced, and approximately 1/2 oftooth height below lateral parapet.
Description The holotype dentary, OMNH 33889, of Har-modontosaurus emeryensis (Fig. 6A) is heavily built with 15tooth spaces and 14 teeth in the preserved portion of the den-tary. The mandibular symphysis is large and extends posteriorlyto the seventh tooth position. The subdental gutter is narrowand runs the length of the tooth row. The subdental shelf nar-rows posteriorly and is well-developed, weakly concave dor-sally, and deep anteriorly. The splenial is not preserved withany of the dentaries, but it is clear from the attachment scarson the ventral surfaces of the subdental shelves of OMNH28067, 27753, and 33889 that the splenial extended anteriorlyto just short of the symphysis, leaving a short symphysial fo-ramen. In accordance with the hypertrophied splenial, theMeckel’s groove is widely open ventromedially in the posteriorportion of the dentary and narrows anteriorly to a ventrallydirected slit that is open to the symphysis. The exterior surfaceof the dentary is smooth, deeply convex, and bears five smallinferior alveolar foramina. It is not possible to determine theexact tooth count of the element, but comparison with otherteiids indicates that only one to three teeth are missing fromthe holotype specimen. The anterior teeth bear sharp anteriorand posterior carinae and are columnar, procumbent, and weak-

651NYDAM—CRETACEOUS LIZARDS
FIGURE 7. Indeterminate Scincomorpha of Mussentuchit local fauna.A, lateral and B, medial views of OMNH 28905; C, lateral view ofOMNH 30208, partial right maxilla. Abbreviations: AAF, anterior in-terior alveolar foramen; PAF, posterior interior alveolar foramen. Scalebars equal 1 mm.
ly recurved. At about the eighth tooth position successive pos-terior teeth become substantially more massive and widelyspaced, such that the four posterior-most tooth positions areoccupied by heavily built, conical teeth with wide bases. Theseposterior teeth also have more robustly built anterior and pos-terior carinae, with the anterior carina forming a shoulder-likeprocess (Fig. 6B). Starting at the sixth tooth position, all suc-cessively posterior teeth have swollen tooth shafts that extendmedially such as to obscure the subdental gutter in occlusalview. The anterior and posterior carinae on all teeth join at aposteromedially directed apex, forming a similarly directed‘‘V.’’ Harmodontosaurus lacks the medial striae on the crownsof its teeth. Replacement pits form as subcircular pits at thebases of the teeth.
Comments Harmodontosaurus emeryensis is most similarin tooth structure to the Judithian teiid Socognathus unicuspisfrom the Oldman formation of Alberta (Gao and Fox, 1991,1996). Both of these taxa have posteriorly rotated crowns anda well-developed posteromedially directed apex. Unlike Socog-nathus, the teeth of Harmodontosaurus show a greater disparityin size between the front and back of the tooth row, with theposterior teeth nearly twice the size of the anterior teeth, where-as in Socognathus the posterior teeth are roughly only halfagain as large as the anterior teeth.
POLYGLYPHANODONTINAE Estes, 1983DICOTHODON Nydam, 1999
DICOTHODON MOORENSIS Nydam, 1999(Figs. 6C, 9M)
Description and Comments The teeth are bicuspid, bul-bous, transversely oriented in the jaw, and oval in occlusalview. The crowns of the teeth are distinct in having a central,transverse V-shaped blade, as well as anterior and posteriorsemicircular accessory blades that border small basins on eitherside of the V-shaped blade. Dicothodon moorensis is a raretaxon in the Mussentuchit local fauna. A description and dis-cussion of the taxon has already been published (Nydam, 1999).
cf. POLYGLYPHANODONTINAE
BICUSPIDON Nydam and Cifelli, 2002b
BICUSPIDON NUMEROSUS Nydam and Cifelli, 2002b(Figs. 6D, 9M)
Description and Comments Bicuspidon numerosus is asmall teiid with conical, recurved anterior teeth and with trans-versely bicuspid, unrecurved posterior teeth. There is a short,transverse, V-shaped blade between the medial and lateral cuspsof the posterior teeth (Nydam and Cifelli, 2002b). This taxonis the most common lizard in the Mussentuchit local fauna andlikely is a sister taxon to the North American lineage of poly-glyphanodontine teiids (Nydam and Cifelli, 2002b).
SCINCOMORPHA Camp, 1923
Family, Genus, and Species indet.(Fig. 7A, B)
Referred Specimens OMNH 22107, 22108 jaw fragments;22109, partial left maxilla; 22110, jaw fragment; 27405, brokenright maxilla; 27408, partial left maxilla; 28905, left maxillamissing nasal process; 30208, broken right maxilla; 32626, jawfragment; 33455, jaw fragment.
Localities and Horizon OMNH localities V235 (OMNH32628), V695 (OMNH 27405, 30208), V868 (OMNH 22105–22110, 32626, 33455); 10–20 m below upper contact of Mus-sentuchit Member of Cedar Mountain Formation and DakotaFormation, Emery County, Utah.
Description The left maxilla, OMNH 28905 (Fig. 7A, B),has 21–22 tooth spaces (wear at posterior end makes exactcount difficult), with 10 teeth having crowns that extend pastthe parapet. The tooth row is bowed dorsally. Although thisbowing in part can be attributed to the preservation of the el-ement, some bowing was present during life and is consideredhere to be characteristic of the maxillary tooth row. The dorsalpart of the nasal process is missing, but the remaining portionshows that it extended posteriorly to about the 18th tooth po-sition. The facet for the maxillary process of the jugal opensdorsolaterally posterior to the posterior interior alveolar fora-men. The preserved exterior surface is smooth and has eightsuperior alveolar foramina along the parapet above the toothrow. The anteriormost of these foramina is the largest and opensanterolaterally just below the anterior edge of the nasal process.The second through fourth foramina are smaller and open an-terolaterally, but gradually become more medially directed withthe more posterior positions. The fifth through eighth foraminaopen posterolaterally, with the last three closely spaced andposterior to the posterior edge of the nasal process. Internallythe anterior interior alveolar foramen opens anteriorly at thebase of the nasal process above the third tooth position, and theposterior interior alveolar foramen opens posteriorly above the12th tooth position (Fig. 7B). The supradental shelf is narrowwhere preserved and borders a weak supradental gutter. Theteeth are homodont, pleurodont, columnar, weakly recurved an-teriorly, and have tapering tips and striated medial surfaces. Inocclusal view the crowns have a posteromedially pointing ‘‘V,’’indicative of posterior rotation of the tooth crowns. Tooth re-placement was direct, as evidenced by the replacement pits atthe bases of some teeth.
OMNH 30208 (Fig. 7C) is an anterior portion of a right max-
652 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 22, NO. 3, 2002
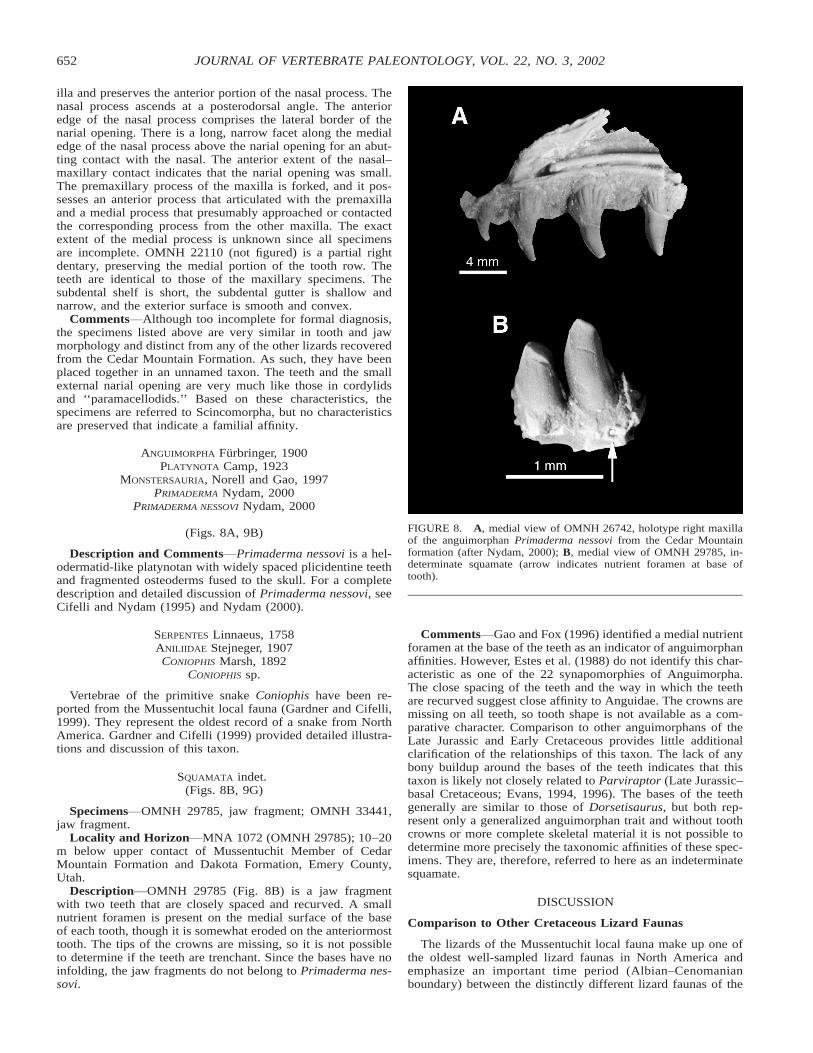
FIGURE 8. A, medial view of OMNH 26742, holotype right maxillaof the anguimorphan Primaderma nessovi from the Cedar Mountainformation (after Nydam, 2000); B, medial view of OMNH 29785, in-determinate squamate (arrow indicates nutrient foramen at base oftooth).
illa and preserves the anterior portion of the nasal process. Thenasal process ascends at a posterodorsal angle. The anterioredge of the nasal process comprises the lateral border of thenarial opening. There is a long, narrow facet along the medialedge of the nasal process above the narial opening for an abut-ting contact with the nasal. The anterior extent of the nasal–maxillary contact indicates that the narial opening was small.The premaxillary process of the maxilla is forked, and it pos-sesses an anterior process that articulated with the premaxillaand a medial process that presumably approached or contactedthe corresponding process from the other maxilla. The exactextent of the medial process is unknown since all specimensare incomplete. OMNH 22110 (not figured) is a partial rightdentary, preserving the medial portion of the tooth row. Theteeth are identical to those of the maxillary specimens. Thesubdental shelf is short, the subdental gutter is shallow andnarrow, and the exterior surface is smooth and convex.
Comments Although too incomplete for formal diagnosis,the specimens listed above are very similar in tooth and jawmorphology and distinct from any of the other lizards recoveredfrom the Cedar Mountain Formation. As such, they have beenplaced together in an unnamed taxon. The teeth and the smallexternal narial opening are very much like those in cordylidsand ‘‘paramacellodids.’’ Based on these characteristics, thespecimens are referred to Scincomorpha, but no characteristicsare preserved that indicate a familial affinity.
ANGUIMORPHA Furbringer, 1900PLATYNOTA Camp, 1923
MONSTERSAURIA, Norell and Gao, 1997PRIMADERMA Nydam, 2000
PRIMADERMA NESSOVI Nydam, 2000
(Figs. 8A, 9B)
Description and Comments Primaderma nessovi is a hel-odermatid-like platynotan with widely spaced plicidentine teethand fragmented osteoderms fused to the skull. For a completedescription and detailed discussion of Primaderma nessovi, seeCifelli and Nydam (1995) and Nydam (2000).
SERPENTES Linnaeus, 1758ANILIIDAE Stejneger, 1907CONIOPHIS Marsh, 1892
CONIOPHIS sp.
Vertebrae of the primitive snake Coniophis have been re-ported from the Mussentuchit local fauna (Gardner and Cifelli,1999). They represent the oldest record of a snake from NorthAmerica. Gardner and Cifelli (1999) provided detailed illustra-tions and discussion of this taxon.
SQUAMATA indet.(Figs. 8B, 9G)
Specimens OMNH 29785, jaw fragment; OMNH 33441,jaw fragment.
Locality and Horizon MNA 1072 (OMNH 29785); 10–20m below upper contact of Mussentuchit Member of CedarMountain Formation and Dakota Formation, Emery County,Utah.
Description OMNH 29785 (Fig. 8B) is a jaw fragmentwith two teeth that are closely spaced and recurved. A smallnutrient foramen is present on the medial surface of the baseof each tooth, though it is somewhat eroded on the anteriormosttooth. The tips of the crowns are missing, so it is not possibleto determine if the teeth are trenchant. Since the bases have noinfolding, the jaw fragments do not belong to Primaderma nes-sovi.
Comments Gao and Fox (1996) identified a medial nutrientforamen at the base of the teeth as an indicator of anguimorphanaffinities. However, Estes et al. (1988) do not identify this char-acteristic as one of the 22 synapomorphies of Anguimorpha.The close spacing of the teeth and the way in which the teethare recurved suggest close affinity to Anguidae. The crowns aremissing on all teeth, so tooth shape is not available as a com-parative character. Comparison to other anguimorphans of theLate Jurassic and Early Cretaceous provides little additionalclarification of the relationships of this taxon. The lack of anybony buildup around the bases of the teeth indicates that thistaxon is likely not closely related to Parviraptor (Late Jurassic–basal Cretaceous; Evans, 1994, 1996). The bases of the teethgenerally are similar to those of Dorsetisaurus, but both rep-resent only a generalized anguimorphan trait and without toothcrowns or more complete skeletal material it is not possible todetermine more precisely the taxonomic affinities of these spec-imens. They are, therefore, referred to here as an indeterminatesquamate.
DISCUSSION
Comparison to Other Cretaceous Lizard Faunas
The lizards of the Mussentuchit local fauna make up one ofthe oldest well-sampled lizard faunas in North America andemphasize an important time period (Albian–Cenomanianboundary) between the distinctly different lizard faunas of the

653NYDAM—CRETACEOUS LIZARDS
Late Jurassic–Early Cretaceous and the Late Cretaceous. Assuch, the lizards of the Mussentuchit local fauna fill an impor-tant gap in the fossil record of lizards and provide an oppor-tunity to examine changes in the North American lizard faunaduring the Early and medial Cretaceous, especially the shiftfrom the primitive lizard fauna of the Late Jurassic–Early Cre-taceous to the more modern-type lizard fauna of the Late Cre-taceous.
North American Early Cretaceous The lizards of the Ear-ly Cretaceous (Aptian–Albian) of North America consist pri-marily of primitive taxa referable to ‘‘Paramacellodidae’’(Winkler et al., 1990; Reynoso, 1995, 1996, 1997, 1998; Ny-dam and Cifelli, 2002a), but also include the earliest records ofteiids (Winkler et al., 1990; Nydam and Cifelli, 2002a). ‘‘Par-amacellodid’’ lizards are common constituents of the Jurassicand basal Cretaceous lizard assemblages from North America,Europe, and many parts of Asia (Prothero and Estes, 1980;Evans, 1993, 1998). The systematic validity of ‘‘Paramacellod-idae’’ has been questioned, with some suggesting that that thefamily may be a junior synonym of Cordylidae (Estes, 1983b;Gao and Fox, 1996). However, phylogenetic analysis of Para-macellodus (Evans and Chure, 1998a) indicates that it is thesister taxon of the Scincoidea (Cordylidae 1 Scincidae). Theteiids from the Early Cretaceous are considered to be primitivebased on their relatively simple dentitions (Winkler et al.,1990). However, at least one of the teiids has some modificationof its teeth, though not modified to the degree found in mostof the Late Cretaceous teiids (Nydam and Cifelli, 2002a).
At least one tentative ‘‘paramacellodid’’ lizard (cf. Pseudo-saurillus) has been identified as part of the Mussentuchit localfauna. In addition, some of the specimens of the indeterminatetaxa (e.g., OMNH 28905 and 30208) are ‘‘paramacellodid’’-like, though too incomplete for diagnosis. In comparison to therest of the lizards from the Mussentuchit local fauna, however,the ‘‘paramacellodid’’ and ‘‘paramacellodid’’-like taxa are rare.Dimekodontosaurus madseni is problematic in that it may rep-resent either a ‘‘paramacellodid’’ or a cordylid. Neither groupincludes characteristics of the lower jaw or dentition in theirmost current diagnoses (Estes et al., 1988; Evans and Chure,1998a). In either case this taxon possesses a dentition that ismore derived than is found in either family and makes it uniqueamong the Scincoidea. Among the teiids of the Mussentuchitlocal fauna, none of the taxa share the primitive conical toothstructure found in three of the four taxa identified from theEarly Cretaceous of North America. The fourth Early Creta-ceous teiid, a new taxon from the Antlers Formation ofOklahoma (Nydam and Cifelli, 2002a), is more derived in thatit has small anteroposterior expansions of the crown. None ofthe teiids from the Mussentuchit local fauna have this type oftooth design. Overall the lizard component of the Mussentuchitlocal fauna includes some primitive taxa, but most of the taxaare quite distinct from the known lizard fauna of the EarlyCretaceous.
North American Late Cretaceous Nearly all taxa fromlizard faunas of the Late Cretaceous have been referred to mod-ern families (Estes, 1983b; Rowe et al., 1992; Gao and Fox,1996). The most common and diverse lizards of the Late Cre-taceous are the teiids. These consist of both non-polyglyphan-odontine taxa (i.e., Chamops, Leptochamops, Meniscognathus,Haptosphenus, Stypodontosaurus, Sphenosiagon, Glyptogenys,Socognathus, Gerontoseps; Estes, 1964, 1969, 1983b; Gao andFox, 1996; Sankey, 1997) and polyglyphanodontine taxa (i.e.,Polyglyphanodon, Paraglyphanodon, Peneteius, Dicothodon;Gilmore, 1940, 1943b; Estes, 1969; McCord, 1998; Nydam,1999). Other modern groups represented in the Late Cretaceouslizard faunas of North America include Platynota (Monstersau-ria and Varanidae; Estes, 1964, 1983b; Gao and Fox, 1996),Anguidae (Estes, 1964, 1969, 1983a; Sullivan, 1981; Gao and
Fox, 1996), Iguanidae (Gao and Fox, 1996), Scincidae (Estes,1964, 1983a; Rowe et al., 1992; Gao and Fox, 1996), Xeno-sauridae (Estes, 1964, 1969, 1983a; Gao and Fox, 1996), andXantusiidae (Gao and Fox, 1996).
Although no taxa were figured, Eaton et al. (1999) describeda series of vertebrate faunas, which include numerous lizardsfrom the Cenomanian, Turonian, Coniacian–Santonian, andCampanian of southern Utah. These are important records inthat little is known of the lizards from North America duringthe Cenomanian–Santonian. Interestingly, Eaton et al. (1999)described the Cenomanian vertebrate fauna from the DakotaFormation as generally consisting of more primitive taxa incomparison to the Turonian–Santonian faunas, as well as withthe well-known faunas of the Campanian–Maastrichtian.Among the seven types of lizards they listed from the Ceno-manian-aged Dakota Formation are one, the problematic ‘‘par-amacellodid’’ ‘‘Saurilodon’’ (which is assumed to be equal tothe Saurillodon of Seiffert, 1973 from which Fig. 9BB wasredrawn), is previously only known from the middle–Late Ju-rassic of Europe. This represents an approximately 45-million-year extension of the temporal record of this taxon. The othernotable record listed by Eaton et al. (1999) is the primitiveanguimorphan Dorsetisaurus (Fig. 9E) in the Turonian-agedfauna of the Smoky Hollow Member of the Straight Cliffs For-mation. This is an extension of approximately 60 million yearsof the record of this taxon in North America, but it is alsoknown from basal Cretaceous of Purbeck (Ensom et al., 1991).Eaton et al. (1999) concluded that the lizards from the Turon-ian–Campanian are generally very similar in composition tothose from other Late Cretaceous (Campanian–Maastrichtian)vertebrate faunas from North America. This includes the pres-ence of cordylids, polyglyphanodontines, teiinines, scincids,xantusiids (Contogenys is listed as a scincid by Eaton et al.,1999, but was shown to be a xantusiid by Gao and Fox, 1996),and varanoids.
The diversity of the lizards of the Mussentuchit local faunais much like that of the lizard faunas of the Late Cretaceous.Most notably, Harmodontosaurus, Dicothodon, and Bicuspidonrepresent the earliest examples of non-polyglyphanodontine andpolyglyphanodontine teiids, and Primaderma represents the ear-liest monstersaurian platynotan (Cifelli and Nydam, 1995; Ny-dam, 1999, 2000). Additionally, Bothriagenys appears to be ascincomorphan allied to either Cordylidae or Scincidae. ShouldDimekodontosaurus prove to be a cordylid, it would be anotherexample of a modern taxon in the fauna. Interestingly, of thethree teiids known from the Mussentuchit local fauna, two (Bi-cuspidon, Dicothodon) are allied with the subfamily Polygly-phanodontinae (sensu Estes, 1983a) and one (Harmodontosau-rus) with the subfamily Teiinae (sensu Estes, 1983a; 5 Cham-opsiinae of Denton and O’Neil, 1995, with respect to only Cre-taceous taxa). In the Late Cretaceous of North America,teiinines exhibited a greater taxonomic diversity (Estes, 1983a;Gao and Fox, 1996) and were more widespread (Estes, 1983a;Rowe et al., 1992; Gao and Fox, 1996) than polyglyphanodon-tine teiids (Estes, 1983a; Nydam, 1999).
The similarities between the lizards of the medial and LateCretaceous of North America indicate that many elements ofthe basic taxonomic structure found in Late Cretaceous lizardfaunas had become established between the Aptian–Albian andthe Albian–Cenomanian (Fig. 10). However, the formation ofthe Late Cretaceous lizard fauna appears to have been a multi-step process, since representatives of the Iguania, Xenosauridae,and Varanidae do not have their first records in North Americauntil the Campanian (Estes, 1983b; Gao and Fox, 1996).
Asian Early Cretaceous The fossil record of lizards from theEarly Cretaceous of Asia, particularly Mongolia, stands in starkcontrast to that from the Early Cretaceous of North America. Inaddition to the primitive scincomorphan family ‘‘Paramacellodi-

654 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 22, NO. 3, 2002
FIGURE 9. Diagrammatic illustration of lizard tooth morphotypes from Late Jurassic (Kimmeridgian) through the end of Cretaceous (Maastrich-tian) of North America; notice the relatively simple tooth types during Kimmeridgian and Aptian–Albian and dramatic increase not only in taxa,but also in the complexity of tooth morphology starting at the Albian–Cenomanian. Position of tooth types should not be interpreted as indicatingdirect evolutionary lineages, but as a record of representatives of groups listed to right. All taxa listed left-to-right, top-to-bottom. A, Parviraptor;B, Primaderma; C, Labrodioctes; D, Paraderma; E, Dorsetisaurus; F, indeterminate anguimorphan; G, taxa indicated by XX for Cenomanian–Santonian are based on the lists of Eaton et al. (1999); H, Odaxosaurus; I, Exostinus; J, Bothriagenys; K, Orthrioscincus, Penemabuya, Aocno-doromeus; L, Contogenys; M, Bicuspidon, Dicothodon; N, Polyglyphanodon, Peneteius; O, Polyglyphanodon, Peneteius; P, indeterminate teiidsof Winkler et al. (1990); Q, Ptilotodon; R, Harmodontosaurus; S, Chamops, T, Sphenosiagon, Leptochamops, Gerontoseps, Glyptogenys; U,Chamops, Leptochamops, Haptosphenus, Stypodontosaurus, Meniscognathus; V, Dimekodontosaurus; W, ?Cordylidae; X, Paramacellodus; Y,inderteminate ‘‘paramacellodid’’-like taxa; Z, Paramacellodus; AA, cf. Pseudosaurillus; BB, cf. Saurillodon; CC, indeterminate iguanians. XXindicates published, but not figured, record of representative of group not referred to a known genus. A after Evans (1994, European taxon,complete teeth not known for North American Parviraptor); B after Nydam (2000); C, D, H, I, K, L, T, U, and W, and CC after Gao and Fox(1996); S and Leptochamops of T and U after Estes (1964); E and X after Evans and Chure (1999); F, Q, Y and Z after Nydam and Cifelli(2002a); J, R, V, and AA from specimens reported herein; M after Nydam (1999) and Nydam and Cifelli (in press b); N and O after Nydam(1999) and Nydam et al. (2000). For clarity and size constraints, figures not drawn to scale, and not all published taxa have been included (notaxon that affects patterns seen here was excluded).
dae’’ and the primitive anguimorphan family Dorsetisauridae, thelizards from Hoovor, Mongolia also include Gekkonidae, Scinci-dae, Necrosauridae, Hoplocercidae, Priscagamidae (probable igu-anians allied with Agamidae; Borsuk-Bialynicka, 1984), Xeno-sauridae, Macrocephalosauridae (5Polyglyphanodontinae, Estes,1983a), and possibly Xantusiidae (Alifanov, 1992, 1993a, c). Theage of this fauna is ?Aptian–Albian (Alifanov, 1992, 1993a), but
the lizard taxa exhibit greater diversity than similarly aged lizardfaunas in North America (Winkler et al., 1990; Nydam and Cifelli,2002a) or Europe (Evans, 1995, 1998). Comparison of the EarlyCretaceous Hoovor lizards to those of the medial Cretaceous Mus-sentuchit local fauna shows a number of similarities. These includethe presence of ‘‘paramacellodids,’’ polyglyphanodontines (5Ma-crocephalosauridae of Sulimski, 1975; Alifanov, 1993b), and pla-

655NYDAM—CRETACEOUS LIZARDS
FIGURE 10. Distributions of lizard taxa during Mesozoic in North America and Asia. Bars above group names represent known temporaldistributions based on references cited herein. Gray bars at end of Albian, end of Turonian, and beginning of Campanian denote likely times ofinflux of Asian taxa into American fauna. Asterisk (*) denotes lineages possibly introduced from Asia upon opening of land bridge. Doubleasterisks (**) indicate lineages possibly introduced from Asia during late Santonian–early Campanian. Arrows represent fossil records for groupsbeyond the Cretaceous–Tertiary boundary. Cladogram adapted from Estes et al. (1988), with addition of Polyglyphanodontinae and Dorsetisauridaefollowing Estes (1983a), ‘‘Paramacellodidae’’ following Evans and Chure (1998b), and Parviraptor following Nydam (2000). Helodermatidae ofEstes et al. (1988) considered here to be conceptually equivalent to Monstersauria of Norell and Gao (1997) and Gao and Norell (1998).
tynotans. These similarities are much like those found between thelizards of the medial and Late Cretaceous within North America.Indeed, five of the groups reported from Hoovor (Polyglyphano-dontinae, Xenosauridae, Xantusiidae, Necrosauridae, and Scinci-dae) have also been reported from the Late Cretaceous of NorthAmerica (Estes, 1983a; Gao and Fox, 1996).
Asian Late Cretaceous The lizards of the Late Cretaceousof Asia are, like their North American counterparts, very di-verse, with agamids, iguanids, gekkonids, teiinines, polygly-phanodontines, anguids, monstersaurs, and varanids (Gilmore,1943a; Sulimski, 1972, 1975, 1984; Estes, 1983a; Borsuk-Bi-alynicka, 1984, 1985, 1991, 1996; Borsul-Bialynicka andMoody, 1984; Nessov, 1988; Borsuk-Bialynicka and Alifanov,1991; Alifanov, 1993a, c; Gao and Hou, 1995a, b; Gao andNorell, 1998). Although some of these groups also are foundin the Cretaceous of North America (Polyglyphanodontinae,Varanidae, Iguanidae, Teiinae, Monstersauria), the individualtaxa are distinct from those in North America. This is mostevident between the large-bodied polyglyphanodontines fromthe Late Cretaceous of Asia (Macrocephalosaurus, Darchan-saurus, Erdenetesaurus, Cherminsaurus, Adamisaurus) and
North America (Polyglyphanodon; see Nydam, 1999 for furtherdiscussion of polyglyphanodontine biogeography). In contrast,the small-bodied polyglyphanodontines from the Late Creta-ceous of Mongolia have been described by Alifanov (1993b) tobe quite similar to some of the small teiids from the Late Cre-taceous of Canada. Indeed, Alifanov (1993b), transferred threeof the teiids from the Late Cretaceous of Canada (Gerontoseps,Socognathus, Sphenosiagon) to the Macrocephalosauridae (sen-su Sulimski, 1975).
European Early Cretaceous The Lizards of the Early Cre-taceous of Europe and surrounding regions have previouslybeen reviewed in detail by Evans (1998) and Evans and Bar-badillo (1997). The studied faunas are widespread both geo-graphically and temporally, but are generally similar in beingcomprised of taxa also found in Late Jurassic faunas and lack-ing taxa common to Late Cretaceous faunas.
European Late Cretaceous Very little is known of thevertebrate faunas of the European Late Cretaceous, particularlywith respect to lizards. A modest fauna has been described fromSpain (Astibia et al., 1990; Rage, 1999) which includes an ig-uanid, scincomorphans, an amphisbaenian, and several snakes.

656 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 22, NO. 3, 2002
Additionally, an indeterminate scincomorphan has been identi-fied from the Campanian of France (Buffetaut et al., 1996).Although limited, these records indicate, at least with respectto Spain, the Early and Late Cretaceous lizards are distinct. Thissupports the trend of a changing faunal identified by Evans(1998). It is also apparent that much more needs to be learnedof the fauna of the Late Cretaceous of Europe; a difficult prob-lem when considering the relative lack of appropriately agedterrestrial deposits.
Biogeographical Implications
Between the Late Jurassic and the Early Cretaceous the lizardfauna of North America appears to have remained relativelystable (Evans, 1998; Nydam and Cifelli, 2002a), exhibiting acomposition dominated by primitive ‘‘paramacellodids’’ thatdefines nearly all of the global faunas during the Late Jurassic-basal Cretaceous (Evans, 1993, 1995, 1998; Evans and Chure,1998b). However, there were some changes during this timeperiod: sphenodontians are reduced from being common andwidespread to single occurrences in the Albian of Montana(Throckmorton et al., 1981) and Mexico (Reynoso, 1995,1997), anguimorphans show an apparent loss in diversity, andteiids appear in North America (Nydam and Cifelli, 2002a). Inthe 8–10 million years from the Early (Aptian–Albian) throughthe medial (Albian–Cenomanian) Cretaceous, there was a dra-matic change in the taxonomic composition of the North Amer-ican lizard fauna (Fig. 10).
Based on the lizards present in the Mussentuchit local fauna,polyglyphanodontine and teiinine teiids, monstersaurs, and pos-sibly advanced scincomorphs (?Scincidae, ?Cordylidae) wereadded to the North American fauna. ‘‘Paramacellodids’’ wereapparently reduced to a minor constituent. Additionally, thepresence of the two polyglyphanodontines Bicuspidon and Di-cothodon (Nydam, 1999; Nydam and Cifelli, 2002b) shows thatthis group had already undergone some diversification. Also,Primaderma indicates that platynotans had become derived bythe time of the appearance of that genus in the Mussentuchitlocal fauna. Although it is possible that these taxa all evolvedin situ in North America, the 13 million years from the Aptian–Albian to the Albian–Cenomanian requires relatively rapid evo-lution within several lineages. An equally plausible, if not moreparsimonious, hypothesis is that these represent taxa introducedfrom an already derived fauna, such as that described fromMongolia. The existence of a Asian–American corridor late inthe Early Cretaceous is supported by numerous similarities be-tween Asia and North America in the dinosaur and mammalfaunas (Jenkins and Schaff, 1988; Currie, 1992; Cifelli et al.,1997, 1999; Kirkland et al., 1997), as well as by geologicalevidence (Cifelli et al., 1997 and references therein). The lizardfauna from the Early Cretaceous of Mongolia (reviewed above)is both temporally antecedent and taxonomically similar to theMussentuchit local fauna, suggesting that the Early CretaceousAsian lizard fauna could have served as a source for at leastsome of the taxa in the lizard fauna from the Albian–Ceno-manian boundary of North America.
Considering the current record, although limited both geo-graphically and stratigraphically (e.g., a single fauna for Ap-tian–Albian of Mongolia and a single fauna for Albian–Ceno-manian of North American), an Asian–North American inter-change may have occurred more than once during the early LateCretaceous. The initial exchange, which took place after theAptian–Albian and before the Albian–Cenomanian, appears tohave been responsible for the introduction of at least threegroups of lizards (based on taxa in Mussentuchit local fauna)to North America—the Scincidae, Cordylidae, and Monstersau-ria. The earliest North American occurrence of Xantusiidae andNecrosauridae is in the Turonian (Eaton et al., 1999) indicating
a second dispersal from Asia. The Iguania, Xenosauridae, andVaranidae, which all have their earliest North America occur-rence in the Campanian (Estes, 1983a, b; Gao and Fox, 1996),are known from the Santonian of Asia (Borsuk-Bialynicka,1991; Gao and Hou, 1995a, b), suggesting dispersal during thelatest Santonian–earliest Campanian.
Based on the faunal comparisons, there almost certainly wasan Asian influence on the North American Cretaceous lizardfauna. However, during at least the Late Cretaceous, SouthAmerica was another potential source for changes in the NorthAmerican lizard fauna. The sauropod dinosaur Alamosaurus,from the Maastrichtian of New Mexico, Texas, Utah, and Wy-oming is very similar to South American titanosaurs and isbelieved to represent immigration from that continent to NorthAmerica (Kues et al., 1980; Lehman, 1987; but see Lucas andSullivan, 2000). In addition, Estes and Baez (1985 and refer-ences therein) reviewed the evidence (distribution of lower ver-tebrate taxa) of an interchange between North and South Amer-ica and concluded that a limited exchange, possibly over anisland chain, existed during the Late Cretaceous. Alifanov(1993b) proposed the existence of a pre-Cretaceous link be-tween North and South America as a solution to the problematicdistribution of lizards during the Cretaceous and Paleogene. Thelack of fossil taxa from the Mesozoic of South America (theonly record is an iguanian from Brazil; Estes and Price, 1973)makes it difficult to evaluate the possible influences of theSouth American fauna on that of North America. It is likely,however, that teiids entered South America from North Americasometime prior to the end of the Cretaceous (Estes, 1983a). Theconcept of South America as a source for North American ig-uanians (sensu Estes, 1983a) has recently been challenged bynew records of iguanians from the Late Cretaceous of Asia(Gao and Hou, 1995b; Gao, 1997).
The lack of substantial records of lizards from Europe duringthe medial or Late Cretaceous also makes it difficult to assessthe possible role of European taxa on the North American lizardfauna. As mentioned above, the record from the Late Creta-ceous of Europe is limited to sparse remains from France andSpain (Astibia et al., 1990; Buffetaut et al., 1996; Rage, 1999).This material is too limited to be of much use in determininga possible relationship with the North American fauna. Theonly exception might be the iguanid from Spain which sharesan expanded, tri-cuspid tooth crown with the indeterminate ig-uanids from the middle Campanian and Maastrichtian of Can-ada (Gao and Fox, 1996) as well as with the iguanid Anchau-rosaurus gilmorei (Gao and Hou, 1995a, 1996) from the middleCampanian of Asia. The epicontinental sea that bisected NorthAmerica during most of the Late Cretaceous makes dispersalof iguanians between the Western Interior of North Americaand Europe unlikely. It is more likely that the presence of aniguanian in Spain is the result of an additional westward dis-persal of the group out of Asia.
Angiosperm Interactions
The change in the North American lizard fauna from theEarly Cretaceous to the medial Cretaceous coincides with theglobal diversification of angiosperms. Indeed, a moderately di-verse assemblage of angiosperms has been reported for the Ce-dar Mountain Formation, including one of the earliest recordsof angiosperm wood (Thayne et al., 1983; Tschudy et al., 1984).During the Early Cretaceous (Aptian–Albian), angiospermswere relatively simple in both flowering and fruiting structures(Friis and Crepet, 1987), and comprised only a modest part ofthe terrestrial flora (Crane and Lidgard, 1990). Based on paly-nological and macrofossil evidence (Crane and Lidgard, 1990;Lupia et al., 1999), angiosperms had become a more prominentpart of the terrestrial flora by the Albian–Cenomanian, but flow-

657NYDAM—CRETACEOUS LIZARDS
ering and fruiting structures remained relatively primitive untilthe latest Santonian–earliest Campanian (Friis and Crepet,1987:Figs. 6.6, 6.7). The timing of the diversification of angio-sperms coincides with the change in the North American lizardfauna after the Aptian–Albian and before the Albian–Cenoman-ian, raising the possibility that the two events were linked. Ver-tebrate-angiosperm co-evolution has been studied with respectto herbivorous dinosaurs (Swain, 1976; Wing and Tiffney,1987; but see Cifelli et al., 1997), but these studies were basedon herbivorous dinosaurs. However, most lizards are insectiv-orous (Greene, 1982; Zug, 1993) and herbivory (i.e., folivoryand not opportunistic feeding on flowers and fruits) in lizardsis rare (Sokol, 1967; King, 1996). For lizards to be successfulherbivores requires specific physiological and environmentalconditions (Pough, 1973; Zimmerman and Tracy, 1989). Basedon the close association of insects and flowering plants it wouldseem obvious that as angiosperm diversity increased so did thatof insects. There is very little evidence on the prehistoric re-lationships of plants and insects and a review of the fossil re-cord of both (albeit very poor for insects) actually shows a dropin insect diversity when angiosperms show an increase (Laban-deira and Sepkoski, 1993). However, based on molecular datathere appears to be a link between the diversity of beetles andthe Cretaceous increase in angiosperm diversity (Farrel, 1998;but see Labandeira and Sepkoski, 1993). Assuming a parallelincrease in angiosperm and insect diversity, it would be rea-sonable to expect a concomitant increase in lizard diversity. Incontrast to the North American record, the diverse lizard faunafrom the ?Aptian–Albian of Mongolia predates the radiation ofangiosperms, indicating that the diversity of that fauna wasmost likely not tied to angiosperm evolution. Therefore, if theAsian fauna was the source for the derived lizard families thatappear in the North American during the Cretaceous, then in-creasing angiosperm diversity was likely not a major influenceon the changes in the North American lizard fauna. Addition-ally, well represented sequences of faunas and associated florasduring the critical period (mid-Cretaceous: Albian–Santonian)of transition are lacking, making it impossible to investigatesimultaneous changes in vertebrate and angiosperm diversity.
If the presence of angiosperms did not play a major role,however, these plants represented a new resource that may havebeen exploited by the polyglyphanodontine teiids. The NorthAmerican polyglyphanodontine teiids (Bicuspidon, Dicothodon,and Polyglyphanodon) show a temporally progressive trend to-ward larger body size and increasingly medially expanded teeth(Nydam, 1999; Nydam and Cifelli, 2002b) that culminated inthe large, probably herbivorous taxon Polyglyphanodon stern-bergi from the Maastrichtian of Utah (Gilmore, 1942). An anal-ogous scenario was also part of the evolutionary history of theAsian polyglyphanodontine taxa. Macrocephalosaurus, Dar-chansaurus, Erdenetesaurus, and Cherminsaurus all possesslarge, polycuspate teeth very similar to those teeth found in theherbivorous family Iguanidae (Gilmore, 1943a; Sulimski,1975). The similarities in tooth structures between the AsianPolyglyphanodontinae and the Iguanidae indicate that the Asianpolyglyphanodontine taxa were probably herbivorous, or atleast omnivorous (Estes, 1983a; Nydam, 1999).
CONCLUSIONS
The lizards from the Mussentuchit local fauna of central Utahrepresent a diverse fauna composed of both primitive and de-rived taxa. As such, this lizard fauna is both temporally andtaxonomically intermediate between those of the Early (Aptian–Albian) and Late (Campanian–Maastrichtian) Cretaceous. Al-though closer in age to the lizard fauna of the Early Cretaceous,the lizards of the Mussentuchit local fauna show greater taxo-nomic similarity to lizard faunas of the Late Cretaceous. This
relatively rapid change in the North American lizard fauna islikely the result of an influx of Asian taxa some time after theAptian–Albian and before the Albian–Cenomanian. This per-mitted lizards (e.g., helodermatid-like platynotans, polygly-phanodontine teiids, and possibly scincids and xantusiids) fromthe more derived faunas of Asia to become established in NorthAmerica. A Late Cretaceous faunal exchange may have led tothe establishment of Iguanidae, Xenosauridae, and Varanidae inthe early Campanian of North America and also may have beenresponsible for the presence of small-bodied polyglyphanodon-tines in both Asia and North America. Angiosperm evolutionand diversification is coincident with the Early–medial Creta-ceous changes in the North American lizard fauna, but it ap-pears that interaction between lizards and angiosperms was lim-ited to the evolution of a few herbivorous lizards in the LateCretaceous of North America and Asia. Although it is possiblethat taxa from Europe and possibly South America may haveplayed a role in the shaping of the North American lizard fauna,information is too limited to determine what, if any, this rolemay have been. Based on shared lineages of lizards it is clearthat exchange of taxa between Asia and North America wasfundamental to the composition and evolution of the CretaceousNorth American Lizard fauna.
ACKNOWLEDGMENTS
I thank E. Larson, K. Smith, E. Sanders, and J. Hilliard fortheir valuable help in the recovery of the specimens describedherein, and S. K. Madsen for preparation of some of the moredelicate elements. The Judd family in Castledale, Utah providedmany aspects of logistical support during several years of field-work. R. Cifelli, L. Vitt, G. Schnell, and N. Czaplewski pro-vided helpful comments on early versions of the manuscript.Comments provided by M. Caldwell and S. Evans were usefulin improving this paper. Support for this work was provided tothe author through scholarships from the American Federationof Mineralogical Societies and Dinamation International Soci-ety (Ying Chang Scholarship), grants from the University ofOklahoma Graduate Student Senate and monetary support fromthe University of Oklahoma Department of Zoology. The fieldprogram leading to the recovery of fossils described herein wassupported by NGS grant 5021-92 and NSF grants DEB-9401094 and DEB-9870173 to R. Cifelli.
LITERATURE CITED
Alifanov, V. 1992. The oldest Gecko (Lacertilia, Gekkonidae) from theLower Cretaceous of Mongolia. Paleontological Journal (Englishtranslation) 26:124–126.
——— 1993a. Some peculiarities of the Cretaceous and Palaeogenelizard faunas of the Mongolian People’s Republic. Kaupia 3:9–13.
——— 1993b. New lizards of the family Macrocephalosauridae (Sau-ria) from the Upper Cretaceous of Mongolia, critical remarks onthe systematics of Teiidae (sensu Estes, 1983). PaleontologicalJournal (English translation) 27:70–90.
——— 1993c. The Upper Cretaceous lizard fauna from Mongolia andthe problem of the first inter-American contact. PaleontologicalJournal (English translation) 27:100–107.
Astibia, H., E. Buffetaut, A. D. Buscalioni, H. Cappetta, C. Corral, R.Estes, F. Garcia-Garmilla, J. J. Jaeger, E. Jimenez-Fuentes, J. L.Loeuff, J. M. Mazin, X. Orue-Etxebarria, J. Pereda-Suberbiola, J.E. Powell, J. C. Rage, J. Rodriquez-Lazaro, J. L. Sanz, and H.Tong. 1990. The fossil vertebrates from Lano (Basque Country,Spain); new evidence on the composition and affinities of the LateCretaceous continental faunas of Europe. Terra Research 2:460–466.
Borsuk-Bialynicka, M. 1984. Anguimorphan and related lizards fromthe Late Cretaceous of the Gobi Desert, Mongolia. PalaeontologicaPolonica 1984:5–106.
——— 1985. Carolinidae, a new family of xenosaurid-like lizards from

658 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 22, NO. 3, 2002
the Upper Cretaceous of Mongolia. Acta Palaeontologica Polonica30:151–176.
——— 1991. Cretaceous lizard occurrences in Mongolia. CretaceousResearch 12:607–608.
——— 1996. The Late Cretaceous lizard Pleurodontagama and theorigin of tooth permanency in Lepidosauria. Acta PalaeontologicaPolonica 41:231–252.
———, and V. R. Alifanov. 1991. First Asiatic ‘iguanid’ lizards in theLate Cretaceous of Mongolia. Acta Palaeontologica Polonica 36:325–342.
———, and S. M. Moody. 1984. Priscagaminae, a new subfamily ofthe Agamidae (Sauria) from the Late Cretaceous of the Gobi De-sert. Acta Palaeontologica Polonica 29:51–81.
Buffetaut, E., G. Costa, J. Le Loeuff, M. Martin, J.-C. Rage, X. Val-entin, and H. Tong. 1996. An Early Campanian vertebrate faunafrom the Villeveyrac Basin (Herault, Southern France). Neues Jahr-buch fur Geologie und Palaeontologie. Monatshefte 1996:1–16.
Cifelli, R. L., and C. de Muizon. 1997. Dentition and jaw of Kokopelliajuddi, a primitive marsupial or near-marsupial from the medial Cre-taceous of Utah. Journal of Mammalian Evolution 4:241–258.
———, J. I. Kirkland, A. Weil, A. R. Deino, and B. J. Kowallis. 1997.High-precision 40Ar/39Ar geochronology and the advent of NorthAmerica’s Late Cretaceous terrestrial fauna. Proceedings of the Na-tional Academy of Sciences USA 94:11163–11167.
———, and S. K. Madsen. 1998. Triconodont mammals from the me-dial Cretaceous of Utah. Journal of Vertebrate Paleontology 18:403–411.
———, and ——— 1999. Spalacotheriid symmetrodonts (Mammalia)from the medial Cretaceous (upper Albian or lower Cenomanian)Mussentuchit local fauna, Cedar Mountain Formation, Utah, USA.Geodiversitas 21:167–214.
———, and R. L. Nydam. 1995. Primitive, helodermatic-like platyno-tan from the Early Cretaceous of Utah. Herpetologica 51:286–291.
———, ———, J. D. Gardner, A. Weil, J. G. Eaton, J. I. Kirkland, andS. K. Madsen. 1999. Medial Cretaceous vertebrates from the CedarMountain Formation, Emery County: the Mussentuchit local fauna;pp. 219–242 in D. D. Gillette (ed.), Vertebrate Paleontology inUtah. Utah Geological Survey, Salt Lake City.
Crane, P. R., and S. Lidgard. 1990. Angiosperm radiation and patternsof Cretaceous palynological diversity; pp. 377–407 in P. D. Taylorand G. P. Larwood (eds.), Major Evolutionary Radiations. Claren-don Press, Oxford.
Currie, B. S. 1998. Upper Jurassic-Lower Cretaceous Morrison and Ce-dar Mountain formations, NE Utah–NW Colorado: relationshipsbetween nonmarine deposition and early cordilleran foreland-basindevelopment. Journal of Sedimentary Research 68:632–652.
Currie, P. J. 1992. Saurischian dinosaurs of the Late Cretaceous of Asiaand North America; pp. 237–249 in N. J. Mateer and C. Pei-Ji(eds.), Aspects of Nonmarine Cretaceous Geology. China OceanPress, Beijing.
Denton, R. K., and R. C. O’Neill. 1995. Prototeius stageri, gen. et sp.nov., a new teiid lizard from the Upper Cretaceous MarshalltownFormation of New Jersey, with a preliminary phylogenetic revisionof the Teiidae. Journal of Vertebrate Paleontology 15:235–253.
Eaton, J. G., R. L. Cifelli, J. H. Hutchison, J. I. Kirkland, and J. M.Parrish. 1999. Cretaceous vertebrate faunas from the KaiparowitsPlateau, south-central Utah; pp. 345–353 in D. G. Gillette (ed.),Vertebrate Paleontology in Utah; Utah Geological Survey Miscel-laneous Publication 99-1. Utah Geological Survey, Salt Lake City.
Edmund, A. G. 1969. Dentition; pp. 117–200 in C. Gans and T. S.Parsons (eds.), Biology of the Reptilia, 1. Academic Press, London.
Ensom, P. C., S. E. Evans, and A. R. Milner. 1991. Amphibians andreptiles from the Purbeck Limestone Formation (Upper Jurassic) ofDorset; pp. 19–20 in Z. Kielan-Jaworowska, N. Heintz, and H. A.Nakrem (eds.), Fifth Symposium on Mesozoic Terrestrial Ecosys-tems and Biota. University of Oslo, Oslo.
Estes, R. 1964. Fossil vertebrates from the Late Cretaceous Lance For-mation, eastern Wyoming. University of California Publications inGeological Sciences 49:1–169.
——— 1969. Relationships of two lizards (Sauria, Teiidae). Breviora317:1–8.
——— 1983a. Sauria Terrestria, Amphisbaenia. Gustav Fisher Verlag,Stuttgart and New York, 249 pp.
——— 1983b. The fossil record and early distribution of lizards; pp.365–398 in A. G. J. Rodin and K. Kiyata (eds.), Advances in Her-
petology and Evolutionary Biology: Essays in Honor of Ernest E.Williams. Museum of Comparative Zoology, Harvard University,Cambridge.
———, and A. Baez. 1985. Herpetofaunas of North and South Americaduring the Late Cretaceous and Cenozoic: evidence for inter-change?; pp. 139–197 in F. G. Stehli and S. D. Webb (eds.), TheGreat American Biotic Interchange. Plenum Press, New York.
———, P. Berberian, and C. A. M. Meszoely. 1969. Lower Vertebratesfrom the Late Cretaceous Hell Creek Formation, McCone County,Montana. Breviora 337:1–33.
———, and L. Price. 1973. Iguanid lizard from the Upper Cretaceousof Brazil. Science 180:748–751.
———, K. de Quieroz, and J. Gauthier. 1988. Phylogenetic relation-ships within Squamata; pp. 119–281 in R. Estes and G. Pregill(eds.), Phylogenetic Relationships of the Lizard Families: EssaysCommemorating Charles L. Camp. Stanford University Press, Stan-ford.
Evans, S. E. 1993. Jurassic lizard assemblages. Revue de PaleobiologieSpecial 7:55–65.
——— 1994. A new anguimorph lizard from the Jurassic and LowerCretaceous of England. Palaeontology 37:33–49.
——— 1995. Lizards: evolution, early radiation, and biogeography; pp.51–55 in A. Sun and Y. Wang (eds.), Sixth Symposium on Meso-zoic Terrestrial Ecosystems and Biota, Short Papers. China OceanPress, Beijing.
——— 1996. Parviraptor (Squamata: Anguimorpha) and other lizardsfrom the Morrison Formation at Fruita, Colorado; pp. 243–248 inM. Morales (ed.), The Continental Jurassic. Museum of NorthernArizona, Flagstaff.
——— 1998. Lepidosaurian faunas from the Early Cretaceous: a cladein transition. Bulletin of the New Mexico Museum of Natural His-tory and Science 17:193–198.
———, and J. Barbadillo. 1997. Early Cretaceous lizards from LasHoyas, Spain. Zoological Journal of the Linnean Society 119:23–49.
———, and D. C. Chure. 1998a. Paramacellodid lizard skulls from theJurassic Morrison Formation at Dinosaur National Monument,Utah. Journal of Vertebrate Paleontology 18:99–114.
———, and ——— 1998b. Morrison lizards: structure, relationshipsand biogeography. Modern Geology 23:35–48.
———, and ——— 1999. Upper Jurassic lizards from the MorrisonFormation of Dinosaur National Monument, Utah; pp. 151–159 inD. D. Gillette (ed.), Vertebrate Paleontology in Utah. Utah Geo-logical Survey, Salt Lake City.
———, and M. Manabe. 1999. Early Cretaceous lizards from the Oku-rodani Formation of Japan. Geobios 32:889–899.
Farrel, B. D. 1998. ‘‘Inordinate fondness’’ explained: why are there somany beetles? Science 281:555–559.
Fraser, N. C., and X.-C. Wu. 1998. Sphenodontians from the BrushyBasin Member of the Morrison Formation in Dinosaur NationalMonument. Modern Geology 23:17–34.
Friis, E. M., and W. L. Crepet. 1987. Time and appearance of floralfeatures; pp. 145–179 in E. M. Friis, W. C. Chalconer, and P. R.Crane (eds.), The Origins of Angiosperms and Their BiologicalConsequences. Cambridge University Press, Cambridge.
Gao, K. 1997. Sineoamphisbaena phylogenetic relationships discussed:reply. Canadian Journal of Earth Sciences 34:886–889.
———, and R. C. Fox. 1991. New teiid lizards from the Upper Cre-taceous Oldman Formation (Judithian) of southwestern Alberta,Canada, with a review of the Cretaceous record of teiids. Annalsof the Carnegie Museum 60:145–162.
———, and ——— 1996. Taxonomy and evolution of Late Cretaceouslizards (Reptilia: Squamata) from western Canada. Bulletin of theCarnegie Museum of Natural History 33:1–107.
———, and L. Hou. 1995a. Iguanians from the Upper Cretaceous Dja-dochta Formation, Gobi Desert, China. Journal of Vertebrate Pa-leontology 15:57–78.
———, and ——— 1995b. Late Cretaceous fossil record and paleo-biogeography of iguanian squamates; pp. 47–50 in A. Sun and Y.Wang (eds.), Sixth Symposium on Mesozoic Terrestrial Ecosystemsand Biota. China Ocean Press, Beijing.
———, and ——— 1996. Systematics and taxonomic diversity ofsquamates from the Upper Cretaceous Djadochta Formation, BayanMandahu, Gobi Desert, People’s Republic of China. Canadian Jour-nal of Earth Sciences 33:578–598.

659NYDAM—CRETACEOUS LIZARDS
———, and M. A. Norell. 1998. Taxonomic revision of Carusia (Rep-tilia: Squamata) from the Late Cretaceous of the Gobi Desert andphylogenetic relationships of the anguimorphan lizards. AmericanMuseum Novitates 3230:1–51.
Gardner, J. D., and R. L. Cifelli. 1999. A primitive snake from theCretaceous of Utah. Special Papers in Palaeontology 60:87–100.
Gilmore, C. W. 1940. New fossil lizards from the Upper Cretaceous ofUtah. Smithsonian Miscellaneous Collections 99:1–3.
——— 1942. Osteology of Polyglyphanodon, an Upper Cretaceous Liz-ard from Utah. Proceedings of the United States National Museum92:229–265.
——— 1943a. Fossil lizards of Mongolia. Bulletin of the AmericanMuseum of Natural History 81:361–384.
——— 1943b. Osteology of Upper Cretaceous Lizards from Utah, witha description of a new species. Proceedings of the United StatesNational Museum 93:209–214.
Gradstein, F. M., F. P. Acterberg, J. G. Ogg, P. van Veen, J. Thierry, andZ. Huang. 1995. A Triassic, Jurassic and Cretaceous time scale;pp. 95–126 in W. A. Berggren, D. V. Kent, M.-P. Aubrey, and J.Hardenbol (eds.), Geochronology Time Scales and Global Strati-graphic Correlation; Society for Sedimentary Geology Special Pub-lication. Society for Sedimentary Geology Special Publication, Tul-sa.
Gray, J. 1827. A synopsis of the genera of saurian reptiles, in whichsome new genera are indicated and the others reviewed by actualexamination. Annals of Philosophy 2:54–58.
Greene, H. W. 1982. Dietary and phenotypic diversity in lizards: whyare some organisms specialized?; pp. 107–128 in D. Mossakowskiand G. Roth (eds.), Environmental Adaptations and Evolution. Gus-tav Fischer, Stuttgart.
Hoffstetter, R. 1967. Coup d’oeil sur les Sauriens (5lacertiliens) descouches de Purbeck (Jurassique superieur d’ Angleterre). ColloquesInternationaux du Centre National de la Recherche Scientifique163:349–371.
Jenkins, F., and C. R. Schaff. 1988. The Early Cretaceous mammalGobiconodon (Mammalia, Triconodonta) from the Cloverly For-mation in Montana. Journal of Vertebrate Paleontology 8:1–24.
King, G. 1996. Reptiles and Herbivory. Chapman and Hall, London,160 pp.
Kirkland, J. I. 1987. Upper Jurassic and Cretaceous lungfish tooth platesfrom the western interior, the last dipnoan faunas of North America.Hunteria 2:1–16.
———, B. Britt, D. L. Burge, K. Carpenter, R. Cifelli, F. Decourten, J.Eaton, S. Hasiotis, and T. Lawton. 1997. Lower to middle Creta-ceous dinosaur faunas of the central Colorado Plateau: a key tounderstanding 35 million years of tectonics, sedimentology, evo-lution and biogeography. Brigham Young University GeologyStudies 42:69–103.
Kues, B. S., T. Lehman, and J. K. J. Rigby. 1980. The teeth of Ala-mosaurus sanjuanensis, a Late Cretaceous sauropod. Journal of Pa-leontology 54:864–869.
Labandeira, C. C., and J. J. J. Sepkoski. 1993. Insect diversity in thefossil record. Science 261:310–315.
Lehman, T. M. 1987. Late Maastrichtian paleoenvironments and dino-saur biogeography in the western interior of North America. Pa-laeogeography, Palaeoclimatology, Palaeoecology 60:189–217.
Lehman, T. H. 1997. Late Campanian dinosaur biogeography in theWestern Interior of North America. Dinofest International Proceed-ings 1997:223–240.
Lucas, S. G., and R. M. Sullivan. 2000. The sauropod dinosaur Ala-mosaurus from the Upper Cretaceous of the San Juan Basin, NewMexico; pp. 147–156 in S. G. Lucas and A. B. Heckert (eds.),Dinosaurs of New Mexico. New Mexico Museum of Natural His-tory and Science Bulletin No. 17. New Mexico Museum of NaturalHistory and Science, Albuquerque.
Lupia, R., S. Lidgard, and P. R. Crane. 1999. Comparing palynologicalabundance and diversity: implications for biotic replacement duringthe Cretaceous angiosperm radiation. Paleobiology 25:305–340.
McCord, R. D. 1998. A new genus and species of Cretaceous polygly-phanodontine lizard (Squamata, Teiidae) from the Kaiparowits Pla-teau, Utah; pp. 281–292 in Y. Tomida, L. J. Flynn, and L. L. Jacobs(eds.), Advances in Vertebrate Paleontology and Geochronology.National Science Museum, Tokyo.
Nelson, M. E., and D. M. Crooks. 1987. Stratigraphy and paleontologyof the Cedar Mountain Formation (Lower Cretaceous), Eastern Em-
ery County, Utah; pp. 55–63 in W. P. Averett (ed.), Paleontologyand Geology of the Dinosaur Triangle: Guidebook for the 1987Field Trip, Sept. 18–20,1987. Museum of Western Colorado, GrandJunction, Colorado.
Nessov, L. A. 1988. Late Mesozoic amphibians and lizards of SovietMiddle Asia. Acta Zoologica Cracoviensia 31:475–486.
Norell, M. A., and K. Gao. 1997. Braincase and phylogenetic relation-ships of Estesia mongoliensis from the Late Cretaceous of the GobiDesert and the recognition of a new clade of lizards. AmericanMuseum Novitates 3211:1–25.
Nydam, R. L. 1999. Polyglyphanodontinae (Squamata: Teiidae) fromthe medial and Late Cretaceous: new records from Utah, U.S.A.and Baja California del Norte, Mexico; pp. 303–317 in D. D. Gil-lette (ed.), Vertebrate Paleontology in Utah. Utah Geological Sur-vey, Salt Lake City.
——— 2000. A new taxon of helodermatid-like lizard from the Albian-Cenomanian of Utah. Journal of Vertebrate Paleontology 20:285.
———, and R. L. Cifelli. 2002a. Lizards from the Lower Cretaceous(Aptian-Albian) Antlers and Cloverly formations. Journal of Ver-tebrate Paleontology. 22:286–298.
———, and ———. 2002b. A new teiid lizard from the medial Cre-taceous of Utah. Journal of Vertebrate Paleontology. 22:276–285.
———, J. A. Gauthier, and J. J. Chiment. 2000. The mammal-like teethof the Late Cretaceous lizard Peneteius aquilonius Estes 1969(Squamata, Teiidae). Journal of Vertebrate Paleontology 20:628–631.
Pough, F. H. 1973. Lizard energetics and diet. Ecology 54:832–844.Prothero, D. R., and R. Estes. 1980. Late Jurassic lizards from Como
Bluff, Wyoming and their palaeobiogeographic significance. Nature286:484–486.
Rage, J.-C. 1999. Squamates (Reptilia) from the Upper Cretaceous ofLano (Basque Country, Spain). Estudios del Museo de CienciasNaturales de Alava 14(Numero Especial 1):121–133.
Reynoso, V.-H. 1995. Lepidosaurian reptiles from the Cantera Tlayua(Albian), Tepexi de Rodrıguez, Puebla, Mexico. Abstract from IIInternational Symposium on Lithographic Limestones, Lleida-Cuenca (Spain), 1995:131–132.
——— 1996. A primitive lizard from the Early Cretaceous of Mexicoand the phylogenetic position of early lizards. Journal of VertebratePaleontology 16(3, suppl.):60A.
——— 1997. A ‘‘beaded’’ sphenodontian (Diapsida: Lepidosauria)from the Early Cretaceous of central Mexico. Journal of VertebratePaleontology 17:52–59.
——— 1998. Huehuecuetzpalli mixtecus gen. et sp. nov.; a basal squa-mate (Reptilia) from the Early Cretaceous of Tepexi de Rodrıgues,central Mexico. Philosophical Transactions of the Royal Society ofLondon, series B, 353:1–23.
Rowe, T. B., R. L. Cifelli, T. M. Lehman, and A. Weil. 1992. TheCampanian Terlingua local fauna, with summary of other verte-brates from the Aguja Formation, Trans-Pecos Texas. Journal ofVertebrate Paleontology 12:472–493.
Sankey, J. T. 1997. Late Cretaceous vertebrate paleontology and paleo-ecology, Upper Aguja Formation, Big Bend National Park, Texas.Journal of Vertebrate Paleontology 17(3, suppl.):73A.
Seiffert, J. 1973. Upper Jurassic lizards from central Portugal. Separatada Memoria dos Servicos Geologicos de Portugal 22:1–85.
Sokol, O. M. 1967. Herbivory in lizards. Evolution 21:192–194.Stokes, W. L. 1944. Morrison Formation and related deposits in and
adjacent to the Colorado Plateau. Bulletin of the Geological Societyof America 55:951–992.
——— 1952. Lower Cretaceous in Colorado Plateau. Bulletin of theAmerican Association of Petroleum Geologists 36:1766–1776.
Sulimski, A. 1972. Adamisaurus magnidentatus n. gen., n. sp. (Sauria)from the Upper Cretaceous of Mongolia. Palaeontologia Polonica27:33–40.
——— 1975. Macrocephalosauridae and Polyglyphanodontidae (Sau-ria) from the Late Cretaceous of Mongolia. Palaeontologia Polonica33:25–102.
——— 1984. A new Cretaceous scincomorph lizard from Mongolia.Palaeontologia Polonica 46:143–155.
Sullivan, R. M. 1981. Fossil lizards from the San Juan Basin, NewMexico; pp. 76–88 in S. G. Lucas, J. K. Rigby, Jr., and B. S. Kues(eds.), Advances in San Juan Basin Paleontology. University ofNew Mexico Press, Albuquerque.
Swain, T. 1976. Angiosperm-reptile co-evolution; pp. 107–122 in A. d.

660 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 22, NO. 3, 2002
A. Bellairs and C. B. Cox (eds.), Morphology and Biology of Rep-tiles. Academic Press, New York.
Thayne, G. F., W. D. Tidwell, and W. L. Stokes. 1983. Flora of theLower Cretaceous Cedar Mountain Formation of Utah and Colo-rado, Part 1. Paraphyllanthoxylon utahense. Great Basin Naturalist43:394–402.
Throckmorton, G., S., J. A. Hopson, and P. Parks. 1981. A redescriptionof Toxolophosaurus cloudi Olson, a Lower Cretaceous herbivoroussphenodontid reptile. Journal of Paleontology 55:586–597.
Tschudy, R. H., B. D. Tschudy, and L. C. Craig. 1984. Palynologicalevaluation of Cedar Mountain and Burro Canyon formations, Col-orado Plateau. United States Geological Survey Professional Paper1281:1–19.
Winkler, D. A., P. A. Murry, and L. L. Jacobs. 1990. Early Cretaceous(Comanchean) vertebrates of central Texas. Journal of VertebratePaleontology 10:95–116.
Wing, S. L., and B. H. Tiffney. 1987. The reciprocal interaction ofangiosperm evolution and tetrapod herbivory. Review of Palaeo-botony and Palynology 50:179–210.
Zimmerman, L. C., and C. R. Tracy. 1989. Interactions between theenvironment and ectothermy and herbivory in reptiles. Physiolog-ical Zoology 62:374–409.
Zug, G. R. 1993. Herpetology, an Introductory Biology of Amphibiansand Reptiles. Academic Press, San Diego, 527 pp.
Received 15 June 2000; accepted 22 June 2001.
Copyright © 2022 FDOKUMEN




















