
Continental fossil vertebrates from the mid-Cretaceous (Albian–Cenomanian) Alcântara Formation,...
15
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
-
Upload
tyrrellmuseum -
Category
Documents
-
view
1 -
download
0
Transcript of Continental fossil vertebrates from the mid-Cretaceous (Albian–Cenomanian) Alcântara Formation,...

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Continental fossil vertebrates from the mid-Cretaceous (Albian–Cenomanian)Alcântara Formation, Brazil, and their relationship with contemporaneousfaunas from North Africa
Carlos Roberto A. Candeiro a,⇑, Federico Fanti b, François Therrien c, Matthew C. Lamanna d
a Laboratório de Geologia, Curso de Geografia, Universidade Federal de Uberlândia, Av. João Naves de Ávilla, 2160, Sta. Mônica, Uberlândia, Minas Gerais, Brazilb Dipartimento di Scienze della Terra e Geologico-Ambientali, Alma Mater Studiorum, Università di Bologna, Via Zamboni 67, I-40127 Bologna, Italyc Royal Tyrrell Museum of Palaeontology, Box 7500, Drumheller, Alberta T0J 0Y0, Canadad Section of Vertebrate Paleontology, Carnegie Museum of Natural History, 4400 Forbes Ave., Pittsburgh, PA 15213, USA
a r t i c l e i n f o
Article history:Received 13 January 2010Received in revised form 8 February 2011Accepted 9 February 2011Available online 19 February 2011
Keywords:PaleobiogeographyBrazilNorth AfricaCretaceousFaunal exchange
a b s t r a c t
The Albian–Cenomanian Alcântara Formation of northeastern Brazil preserves the most diverse continen-tal vertebrate fauna of this age yet known from northern South America. The Alcântara vertebrate assem-blage, consisting of elasmobranchs, actinopterygians, sarcopterygians, turtles, crocodyliforms, pterosaurs,and non-avian dinosaurs, displays close similarities to contemporaneous faunas from North Africa. Theco-occurrence of as many as eight freshwater or estuarine fish taxa (Onchopristis, Bartschichthys, Lepidotes,Stephanodus, Mawsonia, Arganodus, Ceratodus africanus, and possibly Ceratodus humei) and up to seventerrestrial archosaur taxa (Sigilmassasaurus, Rebbachisauridae, Baryonychinae, Spinosaurinae, Carcharod-ontosauridae, possibly Pholidosauridae, and doubtfully Bahariasaurus) suggests that a land route connect-ing northeastern Brazil and North Africa existed at least until the Albian. Interestingly, most componentsof this mid-Cretaceous northern South American/North African assemblage are not shared with coevalsouthern South American faunas, which are themselves characterized by a number of distinct freshwaterand terrestrial vertebrate taxa (e.g., chelid turtles, megaraptoran and unenlagiine theropods). Theseresults suggest that, although mid-Cretaceous faunal interchange was probably possible between north-ern South America and North Africa, paleogeographic, paleoclimatic, and/or paleoenvironmental barriersmay have hindered continental vertebrate dispersal between northern and southern South America dur-ing this time.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
Knowledge of the continental vertebrate faunas of the exten-sive, mid-Cretaceous to ?Cenozoic, coastal to continental depositsof the Itapecuru Group in northern Maranhão State, Brazil(Fig. 1), has improved dramatically over the past 30 years (e.g.,Cunha and Ferreira, 1980; Carvalho and Campos, 1988; Carvalho,1994, 1995; Carvalho and Gonçalves, 1994; Medeiros and Schultz,2001a, 2002, 2004; Nobre and Carvalho, 2002; Carvalho et al.,2003; Castro et al., 2004, 2007; Nobre, 2004; Medeiros, 2006; Eliaset al., 2007a,b; Medeiros et al., 2007; Kellner et al., 2009).Vertebrate fossils from the Itapecuru Group include those of fishes,turtles, plesiosaurs, mosasaurs, crocodyliforms, pterosaurs, andnon-avian dinosaurs. Within the Itapecuru Group, the Albian–Cenomanian Alcântara Formation has yielded the greatest diversityand abundance of continental vertebrates. However, few studies
documenting the variety of taxa known from this unit have beenpublished (e.g., Corrêa Martins, 1997; Medeiros and Schultz,2001a, 2002; Medeiros et al., 2007). In this paper, we review andrevise the continental (i.e., freshwater, estuarine, and terrestrial)vertebrate taxa recovered from the Alcântara Formation, and com-pare the fauna of this unit to those of other mid-Cretaceous WestGondwanan (i.e., African and other South American) deposits, tobetter establish the paleobiogeographic relationships of SouthAmerica and Africa during this interval.
2. Geological setting
Located in northeastern Brazil (Fig. 1), the Itapecuru Groupcomprises intercalated marine, paralic, and continental sedimentsthat range in age from the Early Cretaceous (Albian) to possiblythe early Cenozoic (Pedrão et al., 1993; Rossetti, 1997; Rossettiand Truckenbrodt, 1997). Rifting between South America andAfrica that began during the Early Cretaceous led to rapid marinetransgression and, eventually, the deposition of the Itapecuru
1464-343X/$ - see front matter � 2011 Elsevier Ltd. All rights reserved.doi:10.1016/j.jafrearsci.2011.02.004
⇑ Corresponding author. Tel.: +55 54 34 3268 6118; fax: +55 51 34 3269 2389.E-mail addresses: [email protected], [email protected] (Carlos Ro-
berto A. Candeiro).
Journal of African Earth Sciences 60 (2011) 79–92
Contents lists available at ScienceDirect
Journal of African Earth Sciences
journal homepage: www.elsevier .com/locate / ja f rearsc i

Author's personal copy
Group. Although the lowermost portion of the Itapecuru Groupconsists of undifferentiated deposits (informally termed the‘‘Undifferentiated Unit’’ by some authors [e.g., Castro et al.,2007]), two formal stratigraphic units are recognized in the upperportion: the Alcântara and Cujupe formations (Rossetti, 1997;Rossetti and Truckenbrodt, 1997, 1999). The Alcântara Formationis extensively exposed in the São Luís Basin (Fig. 1), where it con-formably overlies undifferentiated strata of the lower ItapecuruGroup (Fig. 2). Considered Albian–Cenomanian in age by mostworkers (e.g., Rossetti and Truckenbrodt, 1997, 1999), this forma-tion reaches a maximum thickness of 35 m and is composed offine- to medium-grained sandstones interbedded with mudstonesand limestones (Rossetti, 1997). Late Cretaceous through possiblyearly Cenozoic deposits of the Cujupe Formation overlie theAlcântara Formation (Rossetti and Truckenbrodt, 1997; Dias Limaand Rossetti, 1999) (Fig. 2).
2.1. Paleogeographic context
Cretaceous faunal similarities between continental Africa andSouth America are widely documented in the dinosaurian fossil
record (e.g., Calvo and Salgado, 1995; Russell, 1996; Sereno et al.,1996, 1998, 1999, 2004; Forster, 1999; Sues et al., 2002; Upchurchet al., 2002; Candeiro et al., 2004, 2006; Holtz et al., 2004; Coriaand Currie, 2006; Wilson, 2006; Sereno and Brusatte, 2008; Cavinet al., 2010; Le Loeuff et al., 2010; Smith et al., 2010). However, itis important to view paleontological discoveries in light of strati-graphic and paleogeographic data when generating and evaluatingpaleobiogeographic hypotheses. According to current paleogeo-graphic models, the South Atlantic began to open northward inthe Early Cretaceous, with emplacement of oceanic crust betweensouthern Africa and southern South America initiating atapproximately 135 Ma (Viramonte et al., 1999; Jokat et al., 2003;Macdonald et al., 2003). Continental extension between WestAfrica and Brazil resulted in the development of the evaporites ofthe South Atlantic salt basin, Barremian-aged oceanic crust, and riftpropagation from east to west across the West African margin(Karner and Gamboa, 2007). The barrier to southern Atlantic mar-ine incursions relates to the magmatic constructions of a proto-Walvis Ridge and the long-lived anomalous topography of thesoutheastern Brazilian highlands (Gradstein et al., 2004; Karnerand Gamboa, 2007). In the Early Cretaceous, isolated rift/
Fig. 1. Location and schematic geological map of the São Luis Basin in northeastern Maranhão State, Brazil.
80 Carlos Roberto A. Candeiro et al. / Journal of African Earth Sciences 60 (2011) 79–92

Author's personal copy
strike–slip basins provided intermittent shallow-water connec-tions in relatively small and isolated basins, and during the Ap-tian–Albian a major rift phase led to deeper basins withrestricted deep water circulation (Pletsch et al., 2001; Macdonaldet al., 2003). No later than the early Cenomanian, a mature spread-ing stage allowed a continuous and permanent deep-water con-nection between the southern and northern Atlantic, interruptingpossible terrestrial connections and faunal interchanges betweenSouth America and Africa (Petri, 1987; Reyment and Dingle,1987; Pletsch et al., 2001; Tello Saenz et al., 2003; Guedes et al.,2005; Bodin et al., 2010).
3. Fossil vertebrates from the Alcântara Formation, and theirpaleogeographic distributions
Vertebrate remains reported from the Alcântara Formation per-tain to freshwater, estuarine, and terrestrial taxa, including elas-mobranchs, actinopterygians, coelacanths, lungfishes, turtles,plesiosaurs, mosasaurs, crocodyliforms, pterosaurs, and non-aviandinosaurs (Table 1). Putative records of plesiosaurs and mosasaursfrom this unit (Eugênio, 1994; Carvalho et al., 1997, 2000; VilasBôas and Carvalho, 2001; Marinho et al., 2004; Elias et al., 2005;Marinho and Carvalho, 2005) were recently questioned by Kellneret al. (2009, p. 49); consequently, these aquatic reptiles will not beconsidered here.
3.1. Fishes
Fossils of chondrichthyans and actinopterygians are among themost common fish remains found in the Alcântara Formation.Chondrichthyans are known from fin spines of a Tribodus-like hyb-odontid shark (Marques da Silva and Medeiros, 2003; Medeiroset al., 2007) and material referable to Hylaeobatis sp. (Medeiros,2001), Myliobatidae (Ferreira et al., 1995), and the sclerorhynchidsawfish Onchopristis numidus (Pereira and Medeiros, 2003, 2007).Most recently, Pereira and Medeiros (2008) described the newsclerorhynchid taxon Atlanticopristis equatorialis based on rostral
teeth from Cajual Island in northern Maranhão. Actinopterygiansare represented by material referred to several taxa: the polypteridBartschichthys (Pereira and Medeiros, 2003), an indeterminatepycnodontiform (Sousa et al., 2004), the semionotiform Lepidotes(e.g., Ferreira et al., 1995; Medeiros and Schultz, 2001a; Medeiroset al., 2007), and the teleosts Eotrigonodon and Stephanodus(Pereira and Medeiros, 2003) (Table 1). Although the material inquestion is still undescribed, the report of Bartschichthys is ofparticular interest. If confirmed, this material would representthe oldest record of Polypteriformes outside of continental Africa,and one of the most ancient in the world (see Meunier and Gayet,1996; Gayet et al., 2002).
Sarcopterygian remains from the Alcântara Formation includefossils referred to the giant coelacanthiform Mawsonia sp.(Medeiros and Vilas Bôas, 1999; Medeiros and Schultz, 2001a;Medeiros et al., 2007; Amiot et al., 2010) and multiple dipnoan taxa(Arganodus cf. tiguidiensis [Castro et al., 2004], Ceratodus africanus[Cunha and Ferreira, 1979, 1980; Medeiros and Schultz, 2001a;Toledo and Bertini, 2005; Medeiros et al., 2007], Ceratodus humei[Toledo et al., 2005], and an unidentified, possibly new, ceratodontid[Sousa et al., 2005]) (Table 1).
The coelacanthiform genus Mawsonia has been reported fromEarly and mid-Cretaceous deposits of various sedimentary basinsthroughout Brazil and Africa (e.g., Egypt, Algeria, Morocco, Niger,Democratic Republic of the Congo) (Maisey, 2000; Cavin and Forey,2004; Yabumoto and Uyeno, 2005; Carvalho and Maisey, 2008).Cranial remains and fin rays of Mawsonia from the AlcântaraFormation (Medeiros and Vilas Bôas, 1999; Medeiros and Schultz,2001a; Medeiros et al., 2007; Amiot et al., 2010) are among themost geologically recent, if not the most recent, records of thiscoelacanth yet known from South America.
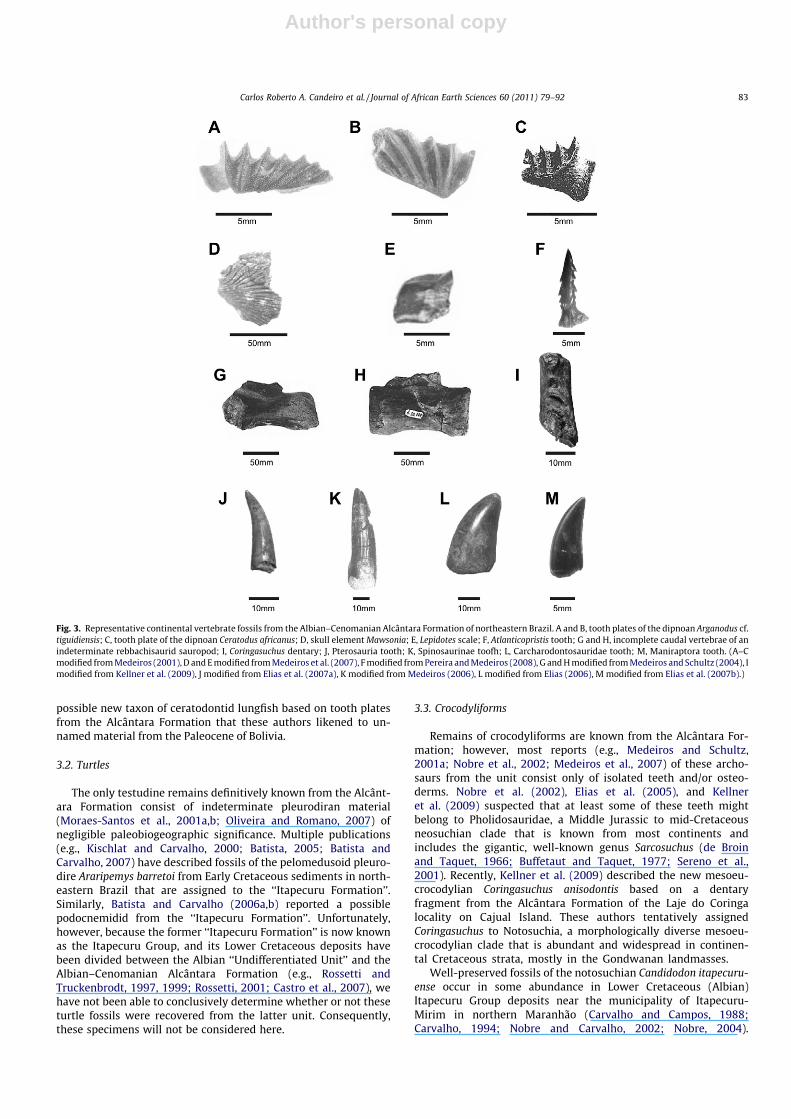
The lungfish Arganodus tiguidiensis was first recognized (asCeratodus tiguidiensis) from the Late Jurassic and Early Cretaceousof North Africa (Tabaste, 1963). Recently, Castro et al. (2004)inferred the presence of Arganodus (as Asiatoceratodus) cf. tiguidien-sis in the Alcântara Formation, on the basis of two tooth plates(Fig. 3A and B). (Kemp (1998) considered the genus Arganodus tobe a junior synonym of Asiatoceratodus, but this proposed synono-my has not been universally accepted [see Cavin et al., 2007;Agnolin, 2010; Soto and Perea, 2010].) Ceratodus africanus (some-times assigned to the genus Neoceratodus [e.g., Martin, 1982,1984], though probably incorrectly [Kemp, 1997]) occurs widelyin the Cretaceous of North Africa (e.g., Haug, 1905; Peyer, 1925;Arambourg and Joleaud, 1943; Tabaste, 1963; Martin, 1984;Churcher and De Iuliis, 2001; Churcher et al., 2006). Tooth platesreferable to this taxon have also been described from the AlcântaraFormation (Cunha and Ferreira, 1979, 1980; Medeiros and Schultz,2001a; Toledo and Bertini, 2005; Medeiros et al., 2007; Fig. 3C).With the exception of a recent report of A. tiguidiensis and C.africanus from the Late Jurassic–?earliest Cretaceous of Uruguay(Soto and Perea, 2010), the Alcântara specimens constitute the onlyreliable occurrences of these species outside of Africa, demonstrat-ing the similarity of the mid-Cretaceous dipnoan faunas of north-eastern South America and North Africa.
Ceratodus humei was originally described on the basis of toothplates from the Late Cretaceous (Campanian) of Quft, Egypt (Priem,1914). Although additional African Cretaceous specimens have beenreferred to C. humei, according to Churcher and De Iuliis (2001), onlythe tooth plates from Upper Cretaceous (probably Campanian) sed-iments of the Dakhleh Oasis, Egypt that these authors described def-initely pertain to this species. Toledo et al. (2005) recently referredtooth plates and fragmentary mandibular elements from the Alcânt-ara Formation to C. humei (as Protopterus humei). If this referral is cor-rect, these Brazilian specimens would constitute the first occurrenceof C. humei outside of Africa, and possibly the oldest record of thespecies wolrdwide. Finally, Sousa et al. (2005) briefly reported a
Fig. 2. Age and stratigraphy of the Itapecuru Group and its subunits (modified fromRossetti and Truckenbrodt (1997)).
Carlos Roberto A. Candeiro et al. / Journal of African Earth Sciences 60 (2011) 79–92 81

Author's personal copy
Table 1Vertebrate fauna of the Alcântara Formation (excluding ichnofossils). Principal references for each record are provided in parentheses.
ChondrichthyesElasmobranchii
Hybodontidae indet. (aff. Tribodus sp. [Marques da Silva and Medeiros, 2003; Medeiros et al., 2007])Ptychodontidae
Hylaeobatis sp. (Medeiros, 2001)Batoidea
Myliobatidae indet. (Ferreira et al., 1995)Sclerorhynchidae
Atlanticopristis equatorialis (Pereira and Medeiros, 2008)Onchopristis numidus (Pereira and Medeiros, 2003, 2007)
OsteichthyesActinopterygii
PolypteridaeBartschichthys sp. (Pereira and Medeiros, 2003)
NeopterygiiPycnodontiformes indet. (Sousa et al., 2004)Semionotiformes
Lepidotes sp. (Ferreira et al., 1995; Medeiros and Schultz, 2001a; Medeiros et al., 2007)Teleostei
Eotrigonodon sp. (Pereira and Medeiros, 2003)Stephanodus sp. (Pereira and Medeiros, 2003)
SarcopterygiiCoelacanthiformes
Mawsonia sp. (Medeiros and Vilas Bôas, 1999; Medeiros and Schultz, 2001a; Medeiros et al., 2007; Amiot et al., 2010)Dipnoi
Arganodus cf. tiguidiensis (Castro et al., 2004)Ceratodus africanus (Cunha and Ferreira, 1979, 1980; Medeiros and Schultz, 2001a; Toledo and Bertini, 2005; Medeiros et al., 2007)Ceratodus humei (Toledo et al., 2005)Ceratodontidae sp. nov.? (Sousa et al., 2005)
TestudinesPleurodira indet. (Moraes-Santos et al., 2001a, 2001b; Oliveira and Romano, 2007)
?Sauropterygia?Plesiosauria indet. (Carvalho et al., 1997, 2000; Vilas Bôas and Carvalho, 2001; Marinho et al., 2004; Elias et al., 2005; Marinho and Carvalho, 2005)
?Lepidosauria?Mosasauridae indet. (Eugênio, 1994; Vilas Bôas and Carvalho, 2001; Marinho and Carvalho, 2005)
CrocodylomorphaCrocodyliformes
Crocodyliformes indet. (Medeiros and Schultz, 2001a; Medeiros et al., 2007)Mesoeucrocodylia
?NotosuchiaCoringasuchus anisodontis (Kellner et al., 2009)
?Pholidosauridae indet. (Nobre et al., 2002; Elias et al., 2005; Kellner et al., 2009)
PterosauriaOrnithocheiroidea
Ornithocheiroidea indet. (Elias et al., 2007a; Lindoso et al., 2008)Anhangueridae indet. (Elias et al., 2007a; Lindoso et al., 2008)
DinosauriaDinosauria incertae sedis
Sigilmassasaurus brevicollis (Medeiros and Schultz, 2001a, 2002; Medeiros et al., 2007)Saurischia
SauropodaSauropoda indet. (Price, 1947; Medeiros et al., 2007; = cf. Astrodon [Medeiros and Schultz, 2001a, 2002])Diplodocoidea
Rebbachisauridae indet. (= Rebbachisaurus sp., = Rebbachisaurus? sp.,=aff. Limaysaurus [‘‘Rayososaurus’’, ‘‘Rebbachisaurus’’] tessonei, = aff.‘‘Rebbachisaurustamesnensis’’ [Medeiros and Schultz, 2001a,b, 2003, 2004; Medeiros and Avilla, 2005; Medeiros et al., 2007])
TitanosauriformesTitanosauriformes indet. (= ‘‘Andesauridae’’ indet. [Medeiros and Schultz, 2001a, 2002; Medeiros et al., 2007])Titanosauria indet. (= Saltasaurinae indet., = ‘‘Titanosauridae’’ indet., = cf. Aegyptosaurus, = cf. ‘‘Titanosauridae’’ indet., = aff. Malawisaurus dixeyi [Medeiros and
Schultz, 2001a, 2002; Medeiros, 2002; Medeiros and Avilla, 2005; Freire et al., 2007; Medeiros et al., 2007])Theropoda
Theropoda indet. (Price, 1960; = aff. ‘‘Elaphrosaurus iguidiensis’’ [Medeiros et al., 2007])Theropoda incertae sedis
?cf. Bahariasaurus ingens (Medeiros and Schultz, 2001a, 2002; Medeiros et al., 2007)Spinosauridae
Spinosauridae indet. (Medeiros, 2006; Medeiros et al., 2007)Spinosauridae sp. nov? (Machado et al., 2009)Baryonychinae indet. (Furtado and Candeiro, 2009a, 2009b)Spinosaurinaecf. Spinosaurus sp. (Medeiros and Vilas Bôas, 1999; Medeiros and Schultz, 2001a, 2002; Medeiros, 2006; Medeiros et al., 2007)
Carcharodontosauridae indet. (= Carcharodontosaurus sp. [Vilas Bôas et al., 1999; Medeiros and Schultz, 2001a, 2002; Medeiros et al., 2007])Maniraptora indet. (= Velociraptorinae indet. [Vilas Bôas, 1999; Elias et al., 2007b])
82 Carlos Roberto A. Candeiro et al. / Journal of African Earth Sciences 60 (2011) 79–92
Author's personal copy
possible new taxon of ceratodontid lungfish based on tooth platesfrom the Alcântara Formation that these authors likened to un-named material from the Paleocene of Bolivia.
3.2. Turtles
The only testudine remains definitively known from the Alcânt-ara Formation consist of indeterminate pleurodiran material(Moraes-Santos et al., 2001a,b; Oliveira and Romano, 2007) ofnegligible paleobiogeographic significance. Multiple publications(e.g., Kischlat and Carvalho, 2000; Batista, 2005; Batista andCarvalho, 2007) have described fossils of the pelomedusoid pleuro-dire Araripemys barretoi from Early Cretaceous sediments in north-eastern Brazil that are assigned to the ‘‘Itapecuru Formation’’.Similarly, Batista and Carvalho (2006a,b) reported a possiblepodocnemidid from the ‘‘Itapecuru Formation’’. Unfortunately,however, because the former ‘‘Itapecuru Formation’’ is now knownas the Itapecuru Group, and its Lower Cretaceous deposits havebeen divided between the Albian ‘‘Undifferentiated Unit’’ and theAlbian–Cenomanian Alcântara Formation (e.g., Rossetti andTruckenbrodt, 1997, 1999; Rossetti, 2001; Castro et al., 2007), wehave not been able to conclusively determine whether or not theseturtle fossils were recovered from the latter unit. Consequently,these specimens will not be considered here.
3.3. Crocodyliforms
Remains of crocodyliforms are known from the Alcântara For-mation; however, most reports (e.g., Medeiros and Schultz,2001a; Nobre et al., 2002; Medeiros et al., 2007) of these archo-saurs from the unit consist only of isolated teeth and/or osteo-derms. Nobre et al. (2002), Elias et al. (2005), and Kellneret al. (2009) suspected that at least some of these teeth mightbelong to Pholidosauridae, a Middle Jurassic to mid-Cretaceousneosuchian clade that is known from most continents andincludes the gigantic, well-known genus Sarcosuchus (de Broinand Taquet, 1966; Buffetaut and Taquet, 1977; Sereno et al.,2001). Recently, Kellner et al. (2009) described the new mesoeu-crocodylian Coringasuchus anisodontis based on a dentaryfragment from the Alcântara Formation of the Laje do Coringalocality on Cajual Island. These authors tentatively assignedCoringasuchus to Notosuchia, a morphologically diverse mesoeu-crocodylian clade that is abundant and widespread in continen-tal Cretaceous strata, mostly in the Gondwanan landmasses.
Well-preserved fossils of the notosuchian Candidodon itapecuru-ense occur in some abundance in Lower Cretaceous (Albian)Itapecuru Group deposits near the municipality of Itapecuru-Mirim in northern Maranhão (Carvalho and Campos, 1988;Carvalho, 1994; Nobre and Carvalho, 2002; Nobre, 2004).
Fig. 3. Representative continental vertebrate fossils from the Albian–Cenomanian Alcântara Formation of northeastern Brazil. A and B, tooth plates of the dipnoan Arganodus cf.tiguidiensis; C, tooth plate of the dipnoan Ceratodus africanus; D, skull element Mawsonia; E, Lepidotes scale; F, Atlanticopristis tooth; G and H, incomplete caudal vertebrae of anindeterminate rebbachisaurid sauropod; I, Coringasuchus dentary; J, Pterosauria tooth; K, Spinosaurinae toofh; L, Carcharodontosauridae tooth; M, Maniraptora tooth. (A–Cmodified from Medeiros (2001), D and E modified from Medeiros et al. (2007), F modified from Pereira and Medeiros (2008), G and H modified from Medeiros and Schultz (2004), Imodified from Kellner et al. (2009), J modified from Elias et al. (2007a), K modified from Medeiros (2006), L modified from Elias (2006), M modified from Elias et al. (2007b).)
Carlos Roberto A. Candeiro et al. / Journal of African Earth Sciences 60 (2011) 79–92 83

Author's personal copy
Candidodon is a peculiar, small-bodied mesoeucrocodylian withlarge orbits and a short snout (Nobre and Carvalho, 2002). As inseveral other notosuchians, it possesses a strikingly heterodontdentition with a surprisingly mammal-like aspect (Carvalho andCampos, 1988; Carvalho, 1994). Regrettably, however, thestratigraphic position(s) of the fossil-bearing sediments inthe Itapecuru-Mirim region within the Itapecuru Group have notyet been established with certainty (Rossetti, pers. comm., 2010);consequently, it is presently unclear whether known material ofCandidodon pertains to the Alcântara Formation (as stated by Eliaset al. (2005)) or the ‘‘Undifferentiated Unit’’ (as stated by Elias(2006, p. 53)). We therefore cannot confidently regard this crocody-liform as a component of the Alcântara Formation fauna, and as such,we will not consider the taxon further here. It is worth noting,however, that most recent phylogenetic analyses that have includedCandidodon (e.g., Turner and Buckley, 2008; Novas et al., 2009;O’Connor et al., 2010) have recovered this notosuchian as the sistertaxon of, or at least very closely related to, Malawisuchusmwakasyungutiensis (Gomani, 1997) from the Early Cretaceous ofMalawi (Africa).
3.4. Pterosaurs
Brazilian pterosaur fossils are known primarily from mid-Cretaceous sediments in the Araripe Basin. Recently, however, Eliaset al. (2007a) and Lindoso et al. (2008) reported the first pterosaurremains from the Alcântara Formation. This record consists of iso-lated teeth from Laje do Coringa that Elias et al. (2007a) referred toOrnithocheiroidea (as Ornithocheiroidea indet. and Anhangueridaeindet.).
3.5. Dinosaurs
Fossils of a variety of non-avian dinosaurs have been collectedfrom the Alcântara Formation (Table 1). More specifically, the dino-saur fauna of this unit presently includes sauropods, theropods,and Sigilmassasaurus brevicollis, an otherwise North African formof controversial taxonomic validity and systematic position.Originally described as a bizarre theropod of uncertain affinities(Russell, 1996), several recent works (Sereno et al., 1996, 1998;Brusatte and Sereno, 2007) have argued that Sigilmassasaurus isactually a junior synonym of the carcharodontosaurid allosauroidCarcharodontosaurus. Other publications (Novas et al., 2005a,b;Canale et al., 2008; Cau and Maganuco, 2009; Novas, 2009) havemaintained the distinction of the two genera; in addition, severalof these latter works (Novas et al., 2005a; Canale et al., 2008;Novas, 2009) have even suggested that at least some materialassigned to Sigilmassasaurus might pertain to the ornithischiandinosaur clade Ornithopoda.
Among the fossils that Russell (1996) allocated to Sigilmassa-saurus are caudal vertebrae from Egypt and Morocco that possessunusual characters, including a transversely compressed centrumwith a subrectangular contour in anteroposterior view, and anelevated, posterodorsally inclined neural spine that expands intransverse dimension towards its apex. Two caudals from theAlcântara Formation of the Laje do Coringa site exhibit the samesuite of features, and have accordingly been referred to thisproblematic genus (Medeiros and Schultz, 2001a, 2002; Medeiroset al., 2007). Provided that, as seems likely, they do not pertainto Carcharodontosauridae (which is already represented in theAlcântara Formation fauna by teeth [see below]), the occurrenceof these vertebrae at Laje do Coringa constitutes yet anothersimilarity between the mid-Cretaceous vertebrate faunas ofnorthern Brazil and North Africa (Novas et al., 2005a; Novas,2009).
3.5.1. OrnithischiansUnless Sigilmassasaurus, or at least the caudal vertebral mor-
photype referred to this taxon, pertains to Ornithopoda (as sug-gested by Novas et al. (2005a), Canale et al. (2008), and Novas(2009)), no ornithischian dinosaur fossils are definitively knownfrom the Alcântara Formation. Avilla et al. (2003) reported a prob-able ornithopod caudal from ‘‘Aptian–Albian strata from the Ita-pecuru Formation’’. However, as stated above, the former‘‘Itapecuru Formation’’ is now known as the Itapecuru Group, andits Lower Cretaceous strata have been partitioned between thelower-middle Albian ‘‘Undifferentiated Unit’’ and the upperAlbian–Cenomanian Alcântara Formation (e.g., Rossetti andTruckenbrodt, 1997, 1999).
3.5.2. SauropodsReasonably abundant but disarticulated and often fragmentary
remains of sauropod dinosaurs have been recovered from theAlcântara Formation (Table 1). Price (1947) provided the first re-port of sauropod fossils from sediments that are now assigned tothis unit. Subsequent works (e.g., Medeiros and Schultz, 2001a,2002, 2004; Medeiros and Avilla, 2005; Freire et al., 2007;Medeiros et al., 2007) have considerably improved our understand-ing of the Alcântara sauropod fauna. Medeiros and Schultz (2001a,2002) referred a single middle or posterior caudal vertebral cen-trum from Laje do Coringa to the titanosauriform Astrodon sp.However, given the fragmentary nature of the specimen, and thefact that the current diagnosis of Astrodon (Carpenter and Tidwell,2005) does not include characters pertaining to the middle or pos-terior caudals, we concur with Medeiros et al. (2007:409) inregarding this Brazilian vertebra as Sauropoda indet.
The presence of rebbachisaurid diplodocoids in the AlcântaraFormation is indicated by disarticulated vertebral remains(Medeiros and Schultz, 2001a,b, 2003, 2004; Medeiros and Avilla,2005; Medeiros et al., 2007; Fig. 3D–F) and possibly by teeth (Fre-ire et al., 2007). Medeiros and Schultz (2004) referred many ofthese fossils to the genus Rayososaurus based on their resemblanceto corresponding skeletal elements of the Patagonian rebbachisau-rid ‘‘Rayososaurus’’ (now known as Limaysaurus) tessonei. More re-cently, however, Medeiros and Avilla (2005) and Medeiros et al.(2007) regarded this Alcântara Formation material as referableonly to Rebbachisauridae indet., an assessment that we followhere. Because rebbachisaurids occurred in at least Europe, SouthAmerica, and North Africa during the Early and mid-Cretaceous(e.g., Mannion, 2009; Apesteguía et al., 2010), the paleobiogeo-graphic utility of the Alcântara material is limited.
Represented by a fragmentary postcranial skeleton from EarlyCretaceous sediments exposed at the Mata locality in northernMaranhão, Amazonsaurus maranhensis is currently the most com-plete sauropod known from the Itapecuru Group and the only dipl-odocoid genus to be named from Brazil (Carvalho et al., 2003). Arecent, comprehensive phylogenetic analysis of Diplodocoidea(Whitlock, in press) suggests that Amazonsaurus may be a basalmember of this clade. Unfortunately, however, as is the case forthe crocodyliform Candidodon, the stratigraphic position of theonly known specimen of Amazonsaurus within the Itapecuru Grouphas not been resolved (Rossetti, pers. comm., 2010); as such, thissauropod may pertain to either the ‘‘Undifferentiated Unit’’ or theAlcântara Formation. As a result, we cannot regard this taxon as amember of the Alcântara fauna and as such do not consider it here.
Titanosauriform sauropods are represented in the AlcântaraFormation fauna by teeth and vertebrae (Medeiros and Schultz,2001a, 2002; Medeiros and Avilla, 2005; Freire et al., 2007;Medeiros et al., 2007). Medeiros and Schultz (2001a, 2002) referredtwo amphiplatyan or shallowly amphicoelous caudal centra fromLaje do Coringa (positioned in the middle-posterior and posteriorregions of the tail, respectively) to ‘‘Andesauridae’’; Medeiros
84 Carlos Roberto A. Candeiro et al. / Journal of African Earth Sciences 60 (2011) 79–92

Author's personal copy
et al. (2007:408) implicitly supported this referral. ‘‘Andesauridae’’was erected by Bonaparte and Coria (1993) to encompass thetitanosaurian genera Andesaurus, Argentinosaurus, and Epachtho-saurus. Nevertheless, as pointed out by Wilson and Upchurch(2003, p. 154), ‘‘. . .Andesauridae is based on primitive charactersthat by definition specify a paraphyletic group. Until taxa are foundsharing synapomorphies with Andesaurus, ‘Andesauridae’ will re-main an informal name.’’ Given that Bonaparte and Coria’s(1993) three ‘‘andesaurid’’ genera are now widely (e.g., Curry Rog-ers, 2005; Wilson, 2006; Filippi et al., in press) regarded as basalmembers of Titanosauria (i.e., non-lithostrotian titanosaurians orbasal lithostrotians), the Alcântara caudals may be those of basaltitanosaurians as well. However, the purportedly ‘‘andesaurid’’characters cited by Medeiros and Schultz (2001a, 2002) and Medei-ros et al. (2007) (amphiplatyan-amphicoelous articular surfaces, ananteriorly-placed neural arch, and other general resemblances tocomparably-positioned vertebrae of Andesaurus) also occur in mid-dle and posterior caudals of non-titanosaurian titanosauriforms.Accordingly, we regard these vertebrae from Laje do Coringa asTitanosauriformes indet.
Despite the uncertainty regarding the affinities of these ‘‘ande-saurid’’ vertebrae within Titanosauriformes, definitive titanosauri-an fossils are known from the Alcântara Formation. Described byMedeiros and Schultz (2001a, 2002), Freire et al. (2007), andMedeiros et al. (2007), these remains consist of dissociated teethand vertebrae. Perhaps the most anatomically informative of thesespecimens is an anterior caudal vertebra, preserving the completecentrum and the base of the neural arch, that was illustrated byMedeiros and Schultz (2001a, 2002) and Medeiros et al. (2007).The centrum of this specimen is strongly procoelous and its neuralarch is anteriorly positioned and anterodorsally inclined, charac-ters that confirm its placement within Titanosauria. Unusuallyamong titanosaurian anterior caudals, the centrum is transverselycompressed (such that it is substantially taller than wide) and sub-rectangular in anteroposterior view.
Medeiros (2002) referred a fragmentary vertebra from Laje doCoringa to the derived titanosaurian subclade Saltasaurinae; how-ever, Medeiros and Avilla (2005) reassigned this material toSauropoda indet.. Other previous studies (e.g., Medeiros andSchultz, 2001a, 2002; Freire et al., 2007; Medeiros et al., 2007) havenoted the resemblances of certain Alcântara Formation titanosau-rian fossils to particular genera (e.g., Aegyptosaurus, Malawisaurus).Nevertheless, due to the fragmentary nature of all describedAlcântara titanosaurian material, and the currently insufficientstate of knowledge regarding character distributions within theclade, we prefer to regard this material as Titanosauria indet. Sincetitanosaurians enjoyed an effectively global distribution during theLate Cretaceous (see, for example, Wilson (2006) or Molnar andWiffen (2007)), these fossils do not allow us to draw any precisepaleobiogeographic conclusions.
Additional sauropod fragments from the Itapecuru Group wererecently described by Castro et al. (2007). Recovered from expo-sures near the municipality of Coroatá in Maranhão, these fossilspertain to the lower-middle Albian ‘‘Undifferentiated Unit’’, notthe Alcântara Formation, and consequently are not taken into ac-count here.
3.5.3. TheropodsLike other kinds of dinosaurs, non-avian theropods are repre-
sented in the Alcântara Formation only by isolated teeth andfragmentary postcranial remains (Table 1). Price (1960) wasthe first to note the occurrence of theropod material in depositsthat modern geologists regard as belonging to the Alcântara For-mation. Decades later, multiple publications (e.g., Vilas Bôaset al., 1999; Medeiros and Schultz, 2001a, 2002; Medeiros,2006; Elias et al., 2007b; Medeiros et al., 2007; Machado et al.,
2009) have reported theropod fossils from this unit, revealing acarnivorous dinosaur assemblage that includes representativesof several major lineages. Some Alcântara theropod remains can-not presently be assigned to particular clades, however. Forexample, Medeiros et al. (2007) described a series of teeth fromthe Laje do Coringa bonebed that these authors likened to thoseof ‘‘Elaphrosaurus iguidiensis’’ from the Cretaceous of North Africa(Lapparent, 1960). Because ‘‘E. iguidiensis’’ is widely considered anomen dubium (e.g., Makovicky et al., 2004; Carrano and Samp-son, 2008), and because material assigned to this taxon is cur-rently regarded as that of one or more indeterminatetheropods (e.g., Carrano and Sampson, 2008; Smith et al.,2010), Medeiros et al. (2007) prudently regarded the Alcântarateeth as Theropoda indet. as well.
Medeiros and Schultz (2001a, 2002) and Medeiros et al. (2007)have compared an isolated ?caudal vertebral centrum from Laje doCoringa to a centrum from In Abangarit, Niger that Lapparent(1960) referred to the enigmatic North African theropod Baharia-saurus ingens. Indeed, in the captions to their illustrations of theBrazilian vertebra, Medeiros and Schultz (2002, Fig. 2E) andMedeiros et al. (2007: Figs. 3.3–3.4) referred to this specimen as‘‘cf. Baharijasaurus (sic)’’ and ‘‘cf Baharijasaurus (sic) ingens’’,respectively. However, in our view, this putative record ofBahariasaurus from the Alcântara Formation is highly doubtful.Although the Laje do Coringa centrum does resemble the vertebrafrom Niger (Lapparent, 1960, pl. V, Fig. 4), the possibility that eitherfossil actually pertains to Bahariasaurus is open to question. Theholotype and all specimens originally referred to B. ingens byStromer (1934) were destroyed in World War II (Rauhut, 1995;Nothdurft et al., 2002; Smith et al., 2006); consequently, compari-sons of more recently collected material to these remains arenecessarily limited to Stromer’s (1934) text and illustrations. Theonly confidently referred caudal vertebrae of Bahariasaurus illus-trated by Stromer (1934, pl. II, Figs. 11, 16, 25, 26)) do not appearclosely similar to either the Alcântara centrum or that from InAbangarit. For example, as also noted by Medeiros and Schultz(2001a, p. 213), the lateral pneumatic fossae (‘‘pleurocoels’’) inthe centrum of the Laje do Coringa vertebra are proportionallymuch larger than those of B. ingens (compare Stromer’s (1934) pl.II, Fig. 25 to Medeiros and Schultz’s (2002) Fig. 2e). We thereforeregard the possible record of Bahariasaurus in the Alcântara Forma-tion as extremely tentative. Indeed, we cannot even be certain thatthis Brazilian vertebra pertains to Theropoda. Interestingly, thespecimen shares several characters with dorsal and sacral centraof juvenile sauropods, including modest size, proportionally enor-mous lateral pneumatic fossae, and unfused neurocentral sutures(compare Medeiros and Schultz’s (2002) Fig. 2e to, for example,Carpenter and McIntosh’s (1994) Fig. 17.2H, Rauhut’s (1999)Fig. 11, Carpenter and Tidwell’s (2005) Figs. 3.5E–I, and/or Schwarzet al.’s (2007) Figs. 5D and E).
In contrast to Bahariasaurus, spinosaurids are unquestionablypresent in the fauna of the Alcântara Formation, with the teethof these aberrant theropods being particularly abundant in the unit(Medeiros and Vilas Bôas, 1999; Medeiros and Schultz, 2001a,2002; Medeiros, 2006; Medeiros et al., 2007; Furtado and Candeiro,2009a, 2009b). Furtado and Candeiro (2009b) recently assigned sixof these teeth to the spinosaurid subclade Baryonychinae. This re-port is of paleobiogeographic significance in that, although spino-saurids as a whole are well-documented from South America,Europe, Asia, and Africa (e.g., Milner et al., 2007; Buffetaut et al.,2008; Buffetaut, 2009; Furtado and Candeiro, 2009a; Hone et al.,2010), baryonychine fossils were previously known only from thelatter three continents. The Alcântara teeth therefore constitutethe first baryonychine occurrence in South America, and possiblythe geologically youngest record of this clade in the Gondwananlandmasses.
Carlos Roberto A. Candeiro et al. / Journal of African Earth Sciences 60 (2011) 79–92 85

Author's personal copy
Most other spinosaurid teeth from the Alcântara Formation arereferable to the subclade Spinosaurinae. Some of these possiblypertain to the genus Spinosaurus itself (e.g., Medeiros and Schultz,2001a, 2002), while others may indicate the presence of a secondspinosaurine taxon in the fauna (Medeiros, 2006). Machado et al.(2009) recently reported what is probably the most anatomicallyand systematically informative spinosaurid fossil yet discoveredfrom the Alcântara Formation. Consisting of paired premaxillae,the specimen exhibits probable synapomorphies of Spinosaurinae(e.g., unserrated teeth) and represents a very large animal, seem-ingly comparable in size to Spinosaurus (see Dal Sasso et al.,2005). With a possible exception from the Barremian of Spain(Sánchez-Hernández et al., 2007), spinosaurines are known only
from mid-Cretaceous sediments of Brazil and North Africa, thusconstituting another notable faunal commonality between thetwo areas.
Carcharodontosaurid theropods are also known from theAlcântara Formation, represented by their distinctive teeth.Although most previous authors (Vilas Bôas et al., 1999; Medeirosand Schultz, 2001a, 2002; Medeiros et al., 2007) have referredthese teeth to Carcharodontosaurus, Candeiro and Martinelli(2005, p. 12) reinterpreted this material as pertaining to indetermi-nate carcharodontosaurids due to its lack of genus-level diagnosticfeatures. Since carcharodontosaurids were widespread during themid-Cretaceous (occurring in North America, Asia, South America,Africa, and possibly Europe [e.g., Brusatte et al., 2010; Ortega et al.,
Fig. 4. Geographic distribution of selected continental vertebrate taxa in northwestern Gondwana during the mid-Cretaceous. (Not all northwestern Gondwanan mid-Cretaceous localities are shown.) (Paleogeographical map modified from Maisey (2000).)
86 Carlos Roberto A. Candeiro et al. / Journal of African Earth Sciences 60 (2011) 79–92

Author's personal copy
Tabl
e2
Dis
trib
utio
nof
cont
inen
talv
erte
brat
eta
xash
ared
betw
een
the
Alc
ânta
raFo
rmat
ion
and
sele
cted
Nor
thA
fric
anan
dSo
uth
Am
eric
anm
id-C
reta
ceou
sst
rati
grap
hic
unit
s.D
ata
sour
ces
are
asfo
llow
s:Ba
hari
yaFo
rmat
ion
(Str
omer
,193
6;Sm
ith
etal
.,20
01);
Chen
iniF
orm
atio
n(B
ento
net
al.,
2000
;A
nder
son
etal
.,20
07);
Kem
Kem
beds
(Dut
heil,
1999
;Ca
vin
etal
.,20
10);
Elrh
azFo
rmat
ion
(Taq
uet,
1976
;Se
reno
and
Brus
atte
,200
8);
Wad
iMilk
Form
atio
n(W
erne
r,19
94;
Wer
ner
and
Gay
et,1
997;
Rauh
ut,1
999)
;A
lcân
tara
Form
atio
n(t
his
pape
r,an
dre
fere
nces
here
in);
Mis
são
Vel
haan
dM
arfi
mfo
rmat
ions
(Mai
sey,
2000
);A
çuFo
rmat
ion
(Ass
ine,
1992
);Sa
ntan
aFo
rmat
ion
(Mai
sey,
2000
;M
arti
ll,20
07);
Loha
nCu
raFo
rmat
ion
(Mar
tine
lliet
al.,
2007
).
Nor
thA
fric
aSo
uth
Am
eric
a
Egyp
tTu
nis
iaM
oroc
coN
iger
Suda
nB
razi
lA
rgen
tin
a
Bah
ariy
aFm
.(C
enom
ania
n)
Ch
enin
iFm
.(A
ptia
n?–
Alb
ian
)
Kem
Kem
beds
(Cen
oman
ian
)
Elrh
azFm
.(A
ptia
n–
Alb
ian
)
Wad
iM
ilk
Fm.
(Cen
oman
ian
)
Alc
ânta
raFm
.(A
lbia
n–
Cen
oman
ian
)
Mis
são
Vel
ha
Fm.
(Apt
ian
)
Mar
fim
Fm.
(Hau
teri
vian
–B
arre
mia
n)
Açu
Fm.
(Alb
ian
–C
enom
ania
n)
San
tan
aFm
.(A
lbia
n)
Loh
anC
ura
Fm.(
Apt
ian
–A
lbia
n)
Can
dele
ros
Fm.
(Cen
oman
ian
)
Hyb
odon
tida
e�
��
��
��
��
Myl
ioba
tida
e�
�O
ncho
pris
tis
��
�Ba
rtsc
hich
thys
��
�Py
cnod
onti
form
es�
��
��
Lepi
dote
s�
��
��
��
��
Step
hano
dus
��
Maw
soni
a�
��
��
��
�A
rgan
odus
tigu
idie
nsis
��
�?
Cera
todu
saf
rica
nus
��
��
�Ce
rato
dus
hum
ei�
?X
?�
�Pl
euro
dira
��
��
��
��
Phol
idos
auri
dae
��
?O
rnit
hoc
hei
roid
ea�
��
��
Sigi
limas
sasa
urus
��
�R
ebba
chis
auri
dae
�?
��
��
�Ti
tan
osau
ria
��
��
��
�Ba
hari
asau
rus
��
?B
aryo
nyc
hin
ae�
�Sp
inos
auri
nae
��
��
�C
arch
arod
onto
sau
rida
e�
��
��
��
Man
irap
tora
��
��
Carlos Roberto A. Candeiro et al. / Journal of African Earth Sciences 60 (2011) 79–92 87

Author's personal copy
2010]), the occurrence of these large-bodied predatory dinosaursin the Albian–Cenomanian of northeastern Brazil is not surprising.Finally, Vilas Bôas (1999) and Elias et al. (2007b) described teethpurportedly belonging to velociraptorine dromaeosaurids fromthe Alcântara Formation. Nevertheless, Gianechini et al. (in press)recently reinterpreted this material as Maniraptora indet., asystematic interpretation that we accept here.
4. Paleobiogeographic implications of the Alcântara Formationvertebrate fauna
As noted by many previous authors (e.g., Medeiros and Schultz,2001a, 2002; Medeiros et al., 2007), numerous similarities existbetween the continental vertebrate assemblages of the AlcântaraFormation of Brazil and multiple mid-Cretaceous units of NorthAfrica, supporting the hypothesis that faunal interchange occurredbetween western Gondwanan landmasses during the Early (andpossibly the early Late) Cretaceous. This interpretation is also sup-ported by the presence of mid-Cretaceous abelisauroid, carchar-odontosaurid, rebbachisaurid, and titanosaurian dinosaurs in bothNorth Africa (e.g., Stromer, 1931; Lavocat, 1954; Russell, 1996;Sereno et al., 1996, 1999, 2004; Smith et al., 2001; Sereno andBrusatte, 2008; Contessi, 2009) and Argentina (e.g., Calvo andSalgado, 1995; Coria and Salgado, 1995; Bonaparte, 1996; Lamannaet al., 2002; Martínez et al., 2004; Salgado et al., 2004; Novas et al.,2005b; Coria and Currie, 2006). Among the many African mid-Cretaceous units that preserve faunas similar to that of theAlcântara Formation are the Bahariya Formation of Egypt, theChenini Formation of Tunisia, the Elrhaz Formation of Niger, theWadi Milk Formation of Sudan, and the Kem Kem beds of Morocco(Fig. 4; Table 2). With the possible exceptions of the Chenini andElrhaz formations, which have been dated to the Aptian?–Albianand the Aptian–Albian, respectively, all of these units are approxi-mately equivalent in age to the Alcântara Formation (Pedrãoet al., 1993; Werner, 1994; Rossetti, 1997; Rossetti and Truckenb-rodt, 1997; Smith et al., 2001; Anderson et al., 2007; Sereno andBrusatte, 2008; Cavin et al., 2010). As is also the case for thisBrazilian formation, each of these African units preserves remainsof many of the following low-level (i.e., ‘‘family-level’’ or less inclu-sive) continental vertebrate taxa: Hybodontidae, Myliobatidae,Onchopristis, Bartschichthys, Lepidotes, Stephanodus, Mawsonia,Arganodus, Ceratodus africanus, Pholidosauridae, Sigilmassasaurus,Rebbachisauridae, Bahariasaurus, Baryonychinae, Spinosaurinae,Carcharodontosauridae, and possibly Ceratodus humei (though, asnoted above, the alleged pre-Campanian African records of this dip-noan species were questioned by Churcher and De Iuliis (2001)).
Several other mid-Cretaceous Brazilian units preserve a conti-nental vertebrate fauna similar to that of the Alcântara Formation.For example, the fauna of the Albian Santana Formation includesHybodontidae, Lepidotes, Mawsonia, and the spinosaurine spino-saurid Irritator challengeri (Maisey, 2000; Sues et al., 2002; Martill,2007), while the Aptian Missão Velha Formation has yielded thesethree fish taxa plus Ceratodus (Brito et al., 1994; Maisey, 2000).Additionally, the Albian–Cenomanian Açu Formation has producedLepidotes remains, while the Hauterivian–Barremian Marfim For-mation yields this semionotiform genus plus Hybodontidae andMawsonia (Fig. 4, Table 2). Thus, the presence of closely relatednonmarine vertebrate taxa in middle Cretaceous deposits of NorthAfrica and Brazil (Sereno et al., 1996, 1998, 1999, 2004; Forster,1999; Candeiro et al., 2004, 2006; Holtz et al., 2004; Wilson,2006; Sereno and Brusatte, 2008; this study), as well as paleogeo-graphic reconstructions, supports the hypothesis that a land routeconnecting the western Gondwanan landmasses may have per-sisted until the early Late Cretaceous, possibly in the equatorial re-
gion (Petri, 1987; Reyment and Dingle, 1987; Pitman et al., 1993;Pletsch et al., 2001; Tello Saenz et al., 2003; Guedes et al., 2005).
Interestingly, certain continental vertebrate taxa that occur inboth the Alcântara Formation and approximately coeval NorthAfrican strata, such as Bartschichthys, Mawsonia, Arganodus,Ceratodus africanus, Sigilmassasaurus, and Spinosauridae, have notbeen discovered in contemporaneous deposits in Argentina. (Weconcur with Buffetaut (2009) in regarding a recent report of aspinosaurid tooth from the Turonian of Patagonia [Salgado et al.,2009] with caution, as this tooth exhibits morphologies that areunknown in definitive spinosaurid taxa.) Similarly, mid-Cretaceousterrestrial tetrapod faunas from Argentina include several taxa thathave not been found in Brazil or North Africa, such as chelid turtles(e.g., Lapparent de Broin and de la Fuente, 2001), dicraeosauridsauropods (Salgado and Bonaparte, 1991), and megaraptoran(Novas, 1998; Benson et al., 2009) and unenlagiine (e.g., Makovickyet al., 2005; Gianechini et al., in press) theropods. These differencesin faunal composition suggest that the mid-Cretaceous continentalvertebrate assemblages of northern South America may have beenmore similar to coeval North African faunas than to those fromsouthern South America.
Although further paleontological discoveries may falsify thisinterpretation, the distinction observed between the northernSouth American/North African and southern South American fau-nal assemblages supports the idea that paleogeographic, paleocli-matic, and/or paleoenvironmental barriers existed betweennorthern South America/North Africa and southern South Americaduring the middle stages of the Cretaceous. Geological features ofSouth America have previously been hypothesized to have playeda role in acting as physical barriers between the northern andsouthern portions of the continent (Arid, 1977; Garcia et al.,2005; Candeiro et al., 2006; Candeiro, 2010). One of the most plau-sible candidates is the Paranaíba High (=Alto Paranaíba Arch), anelevated ridge between central and southern South America thatcould have acted as a structural barrier preventing or hinderingfaunal exchange during the mid-Cretaceous. The emplacement ofthis barrier could have resulted in the divergence of northernand southern South American faunas and in the formation of dis-tinct paleobiogeographic provinces in both areas of the continent.
5. Conclusions
The continental vertebrate fauna of the Albian–CenomanianAlcântara Formation of northeastern Brazil shows strongsimilarities with penecontemporaneous assemblages from NorthAfrica. The co-occurrence of up to eight fish taxa (Onchopristis,Bartschichthys, Lepidotes, Stephanodus, Arganodus, Ceratodusafricanus, Mawsonia, and possibly Ceratodus humei) and as manyas seven low-level terrestrial archosaur clades (Sigilmassasaurus,Rebbachisauridae, Baryonychinae, Spinosaurinae, Carcharodonto-sauridae, possibly Pholidosauridae, and doubtfully Bahariasaurus)in the Alcântara Formation and coeval African deposits indicatesthat either a significant vicariant event took place when NorthAfrica and northern South America separated, or that a terrestrialcorridor at least intermittently connected these areas until the lateEarly or early Late Cretaceous. Interestingly, many of the faunalcommonalities between Brazil and North Africa are not sharedwith southern South America, which is instead characterized bylargely distinct continental vertebrate assemblages. These resultssuggest that, although mid-Cretaceous faunal interchange wasprobably possible between northern South America and North Afri-ca, paleogeographic, paleoclimatic, and/or paleoenvironmentalbarriers may have impeded faunal dispersal between northernand southern South America until the later stages of the LateCretaceous.
88 Carlos Roberto A. Candeiro et al. / Journal of African Earth Sciences 60 (2011) 79–92

Author's personal copy
Acknowledgments
We thank D.M. Martill (University of Portsmouth, UK), F.Lapparent de Broin (Muséum National d’Histoire Naturelle,France), D.F. Rossetti (Museu Paraense Emílio Goeldi, Brazil), andA.G. Martinelli (Universidade de Uberaba, Brasil) for providingvaluable comments in their reviews of earlier versions of this man-uscript. C.R.A. Candeiro was supported by a Jurassic Foundationgrant and Conselho Nacional de Desenvolvimento Científico eTecnológico (CNPq)/ Bolsista de Produtividade em Pesquisa.
References
Agnolin, F., 2010. A new species of the genus Atlantoceratodus (Dipnoiformes:Ceratodontoidei) from the Uppermost Cretaceous of Patagonia and a briefoverview of fossil dipnoans from the Cretaceous and Paleogene of SouthAmerica. Brazilian Geographical Journal: Geosciences and Humanities researchmedium 1 (2), 162–210.
Amiot, R., Wang, X., Lécuyer, C., Buffetaut, E., Boudad, L., Cavin, L., Ding, Z., Fluteau,F., Kellner, A.W.A., Tong, H., Zhang, F., 2010. Oxygen and carbon isotopecompositions of middle Cretaceous vertebrates from North Africa and Brazil:ecological and environmental significance. Palaeogeography, Palaeocli-matology, Palaeoecology 297, 439–451.
Anderson, P.E., Benton, M.J., Trueman, C.N., Paterson, B.A., Cuny, G., 2007.Palaeoenvironment of vertebrates on the southern shore of Tethys: thenonmarine Early Cretaceous of Tunisia. Palaeogeography, Palaeoclimatology,Palaeoecology 243 (1–2), 118–131.
Apesteguía, S., Gallina, P.A., Haluza, A., 2010. Not just a pretty face: anatomicalpeculiarities in the postcranium of rebbachisaurids (Sauropoda: Diplodo-coidea). Historical Biology 22 (1–3), 165–174.
Arambourg, C., Joleaud, L., 1943. Vertébrés fossiles du bassin du Niger. GovernementGénéral de l’Afrique Occidentale Française Bulletin de la Direction des Mines 7,27–84.
Arid, F.M., 1977. Paleogeographical evidences from Bauru Basin (Upper Cretaceous)of southern Brazil. Naturalia 3, 7–13.
Assine, M.L., 1992. Análise estratigráfica da Bacia do Araripe, nordeste do Brasil.Revista Brasileira de Geociências 22 (3), 289–300.
Avilla, L.S., Candeiro, C.R.A., Abrantes, E.A.L., 2003. Ornithischian remains from theLower Cretaceous of Brazil and its paleobiogeographic implications. In: 3Simpósio Brasileiro de Paleontologia de Vertebrados, Livro de Resumos, p. 14.
Batista, D.L., 2005. Nova ocorrência de Araripemys barretoi (Pelomedusoides:Araripemydidae) na Formação Itapecuru, Bacia do Parnaíba. In: CongressoBrasileiro de Paleontologia 19, Resumos (CD-ROM).
Batista, D.L., Carvalho, I.S., 2006a. Contexto geológico dos quelônios da FormaçãoItapecuru (Aptiano–lbiano), Bacia do Parnaíba. In: Perinotto, J.A.J., Lino, I.C.,Saad, A.R., Etchebehere, M.L.C., Morales, N. (Eds.), Boletim do 7� Simpósio doCretáceo do Brasil/1� Simpósio do Terciário do Brasil, p. 17.
Batista, D.L., Carvalho, I.S., 2006b. Um novo quelônio Pleurodira da FormaçãoItapecuru (Cretáceo Inferior), Bacia do Parnaíba. In: XLIII Congresso Brasileiro deGeologia – Geociências e as Sociedades do Futuro, Anais, p. 326.
Batista, D.L., Carvalho, I.S., 2007. Ogênero Araripemys (Chelonii, Pleurodira) noCretáceo Brasileiro. In: Carvalho, I.S., Cassab, R.C.T., Schwanke, C., Carvalho,M.A., Fernandes, A.C.S., Rodrigues, M.A.C., Carvalho, M.S.S., Arai, M., Oliveira,M.E.Q. (Eds.), Paleontologia: Cenários de Vida. Interciência, Rio de Janeiro,Brazil, pp. 291–297.
Benson, R.B.J., Carrano, M.T., Brusatte, S.L., 2009. A new clade of archaic large-bodiedpredatory dinosaurs (Theropoda: Allosauroidea) that survived to the latestMesozoic. Naturwissenschaften 97 (1), 71–78.
Benton, J.M., Bouaziz, S., Buffetaut, E., Martill, D., Ouaja, M., Soussi, M., Trueman, C.,2000. Dinosaurs and other fossil vertebrates from fluvial deposits in the LowerCretaceous of southern Tunisia. Palaeogeography, Palaeoclimatology,Palaeoecology 157, 227–246.
Bodin, S., Petitpierre, L., Wood, J., Elkanouni, I., Redfern, J., 2010. Timing of Early tomid-Cretaceous tectonic phases along North Africa: new insights from theJeffara escarpment (Libya–Tunisia). Journal of African Earth Sciences 58 (3),489–506.
Bonaparte, J.F., 1996. Cretaceous tetrapods of Argentina. MünchnerGeowissenschaftliche Abhandlungen A, 73–130.
Bonaparte, J.F., Coria, R.A., 1993. Un nuevo y gigantesco saurópodo titanosaurio de laFormación Rio Limay (Albiano–Cenomaniano) de la Provincia del Neuquén,Argentina. Ameghiniana 30 (3), 271–282.
Brito, P.M., Bertini, R.J., Martill, D.M., Salles, L.O., 1994. Vertebrate fauna from theMissão Velha Formation (Lower Cretaceous, N.E. Brazil). Boletim do 3� Simpósiosobre o Cretáceo do Brasil, pp. 139–140.
Brusatte, S.L., Sereno, P.C., 2007. A new species of Carcharodontosaurus (Dinosauria:Theropoda) from the Cenomanian of Niger and a revision of the genus. Journalof Vertebrate Paleontology 27 (4), 902–916.
Brusatte, S.L., Chure, D.J., Benson, R.B.J., Xu, X., 2010. The osteology of Shaochilongmaortuensis, a carcharodontosaurid (Dinosauria: Theropoda) from the LateCretaceous of Asia. Zootaxa 2334, 1–46.
Buffetaut, E., 2009. The biogeography of spinosaurid dinosaurs: a review. In: Fanti,F., Spalletta, C. (Eds.), International Conference on Vertebrate
Palaeobiogeography and Continental Bridges Across Tethys, Mesogea, andMediterranean Sea – Abstracts Book. Museo Geologico Giovanni Capellini,Dipartimento di Scienze della Terra e Geologico-Ambientali, Bologna, Italy, pp.18–20.
Buffetaut, E., Taquet, P., 1977. The giant crocodilian Sarcosuchus in the EarlyCretaceous of Brazil and Niger. Palaeontology 20 (1), 203–208.
Buffetaut, E., Suteethorn, V., Tong, H., Amiot, R., 2008. An Early Cretaceousspinosaurid theropod from southern China. Geological Magazine 145 (5),745–748.
Calvo, J.O., Salgado, L., 1995. Rebbachisaurus tessonei sp. Nov. a new Sauropoda fromthe Albian–Cenomanian of Argentina; new evidence on the origin of theDiplodocidae. Gaia 11, 13–33.
Canale, J.I., Novas, F.E., Haluza, A., 2008. Comments about the cervical vertebraereferred to the African theropods Carcharodontosaurus and Sigilmassasaurus. In:Calvo, J.O., Juárez Valieri, R.D., Porfiri, J.D., dos Santos, D.D. (Eds.), Actas IIICongreso Latinoamericano de Paleontología de Vertebrados. Neuquén 22–25Septiembre, 2008. Universidad Nacional del Comahue, Neuquén, Argentina, p. 45.
Candeiro, C.R.A., 2010. Record of the Late Cretaceous South American genusAeolosaurus (Sauropoda, Titanosauria): Paleogeographical implications.Estudios Geológicos, 66 (2), 243–253.
Candeiro, C.R.A., Martinelli, A.G., 2005. Abelisauroidea and Carcharodontosauridae(Theropoda, Dinosauria) in the Cretaceous of South America.Paleogeographical and geocronological implications. Sociedade e Natureza.Uberlândia 17 (33), 5–19.
Candeiro, C.R.A., Abranches, C.T., Abrantes, E.A., Avilla, L.S., Martins, V.C., Moreira,A.L., Torres, S.R., Bergqvist, L.P., 2004. Dinosaurs remains from western SãoPaulo state, Brazil (Bauru Basin, Adamantina Formation, Upper Cretaceous).Journal of South American Earth Sciences 18 (1), 1–10.
Candeiro, C.R.A., Martinelli, A.G., Avilla, L.S., Rich, T.H., 2006. Tetrapods from theUpper Cretaceous (Turonian–Maastrichtian) Bauru Group of Brazil: areappraisal. Cretaceous Research 27 (6), 923–946.
Carpenter, K., McIntosh, J., 1994. Upper Jurassic sauropod babies from the MorrisonFormation. In: Carpenter, K., Hirsch, K.F., Horner, J.R. (Eds.), Dinosaur Eggs andBabies. Cambridge University Press, Cambridge, UK, pp. 265–278.
Carpenter, K., Tidwell, V., 2005. Reassessment of the Early Cretaceous sauropodAstrodon johnsoni Leidy 1865 (Titanosauriformes). In: Tidwell, V., Carpenter, K.(Eds.), Thunder-Lizards: The Sauropodomorph Dinosaurs. Indiana UniversityPress, Bloomington and Indianapolis, Indiana, USA, pp. 78–114.
Carrano, M.T., Sampson, S.D., 2008. The phylogeny of Ceratosauria (Dinosauria:Theropoda). Journal of Systematic Palaeontology 6 (2), 183–236.
Carvalho, I.S., 1994. Candidodon: um crocodilo com heterodontia (Notosuchia,Cretáceo Inferior–Brasil). Anais da Academia Brasileira de Ciências 66 (3),331–346.
Carvalho, I.S., 1995. As pistas de dinossauros da ponta da Guia (Bacia de São Luís,Cretáceo Superior–Maranhão, Brasil). Anais da Academia Brasileira de Ciências67 (4), 413–431.
Carvalho, I.S., Campos, D.A., 1988. Um mamífero triconodonte do Cretáceo Inferiordo Maranhão, Brasil. Anais da Academia Brasileira de Ciências 60 (4), 437–446.
Carvalho, I.S., Gonçalves, R.A., 1994. Pegadas de dinossauros neocretáceas daFormação Itapecuru, Bacia de São Luís (Maranhão, Brasil). Anais da AcademiaBrasileira de Ciências 66 (3), 279–292.
Carvalho, M.S.S., Maisey, J.G., 2008. New occurrence of Mawsonia (Sarcopterygii:Actinistia) from the Early Cretaceous of the Sanfranciscana Basin, MinasGerais, southeastern Brazil. Geological Society, London, Special Publications295, 109–144.
Carvalho, I.S., Vilas Bôas, I., Bergqvist, L.P., 1997. Plesiossauros da região equatorialBrasileira, Bacia de São Luís (Cretáceo Superior), Brasil. In: Congresso Brasileirode Paleontologia 15, Resumos, p. 93.
Carvalho, I.S., Vilas Bôas, I., Bergqvist, L.P., 2000. Plesiossauros da região equatorialBrasileira Bacia de São Luís (Cretáceo Superior), Brasil. Acta GeologicaLeopoldensia 23 (51), 33–41.
Carvalho, I.S., Avilla, L.S., Salgado, L., 2003. Amazonsaurus maranhensis gen. et sp.nov. (Sauropoda, Diplodocoidea) from the Lower Cretaceous (Aptian–Albian) ofBrazil. Cretaceous Research 24, 697–713.
Castro, D.F., Toledo, C.E.V., Sousa, E.P., Medeiros, M.A., 2004. Nova ocorrência deAsiatoceratodus (Osteichthyes, Dipnoiformes) na Formação Alcântara,Eocenomaniano da Bacia de São Luís, Ma, Brasil. Revista Brasileira dePaleontologia 7 (2), 245–248.
Castro, D.F., Bertini, R.J., Santucci, R.M., Medeiros, M.A., 2007. Sauropods of theItapecuru Group (lower/middle Albian), São Luís-Grajaú Basin, Maranhão State,Brazil. Revista Brasileira de Paleontologia 10 (3), 195–200.
Cau, A., Maganuco, S., 2009. A new theropod dinosaur, represented by a singleunusual caudal vertebra, from the Kem Kem Beds (Cretaceous) of Morocco. Attidella Società Italiana di Scienze Naturali e del Museo Civico di Storia Naturale diMilano 150 (2), 239–257.
Cavin, L., Forey, P.L., 2004. New mawsoniid coelacanth (Sarcopterygii, Actinistia)remains from the Cretaceous of the Kem Kem beds, SE Morocco. In: Arratia, G.,Tintori, A. (Eds.), Mesozoic Fishes III – Systematics, Paleoenvironments andBiodiversity. Verlag Dr. Friedrich Pfeil, Munich, Germany, pp. 493–506.
Cavin, L., Suteethorn, V., Buffetaut, E., Tong, H., 2007. A new Thai Mesozoic lungfish(Sarcopterygii, Dipnoi) with an insight into post-Palaeozoic dipnoan evolution.Zoological Journal of the Linnean Society 149 (2), 141–177.
Cavin, L., Tong, H., Boudad, L., Meister, C., Piuz, A., Tabouelle, J., Aarab, M., Amiot, R.,Buffetaut, E., Dyke, G., Hua, S., Le Loeuff, J., 2010. Vertebrate assemblages fromthe early Late Cretaceous of southeastern Morocco: an overview. Journal ofAfrican Earth Sciences 57, 391–412.
Carlos Roberto A. Candeiro et al. / Journal of African Earth Sciences 60 (2011) 79–92 89

Author's personal copy
Churcher, C.S., De Iuliis, G., 2001. A new species of Protopterus and a revision ofCeratodus humei (Dipnoi: Ceratodontiformes) from the Late Cretaceous MutFormation of eastern Dakhleh Oasis, Western Desert of Egypt. Journal ofPalaeontology 44 (2), 305–323.
Churcher, C.S., De Iuliis, G., Kleindienst, M.R., 2006. A new genus for the dipnoanspecies Ceratodus tuberculatus Tabaste, 1963. Geodiversitas 28 (4), 635–647.
Contessi, M., 2009. Vertebrate remains from the Barremian–Cenomanian (EarlyCretaceous) coastal deposits of South Tunisia. In: Fanti, F., Spalletta, C. (Eds.),International Conference on Vertebrate Palaeobiogeography and ContinentalBridges across Tethys, Mesogea, and Mediterranean Sea – Abstracts Book.Museo Geologico Giovanni Capellini, Dipartimento di Scienze della Terra eGeologico-Ambientali, Bologna, Italy, pp. 31–32.
Coria, R.A., Currie, P.J., 2006. A new carcharodontosaurid (Dinosauria, Theropoda)from the Upper Cretaceous of Argentina. Geodiversitas 28 (1), 71–118.
Coria, R.A., Salgado, L., 1995. A new giant carnivorous dinosaur from the Cretaceousof Patagonia. Nature 377, 224–226.
Corrêa Martins, F.J., 1997. A Laje do Coringa: um ‘‘bone-bed’’ da Formação Itapecuru(Cretáceo Superior da Bacia de São Luís-MA). Anais da Academia Brasileira deCiências 69 (3), 436–437.
Cunha, F.L.S., Ferreira, C.S., 1979. Sur l’evidence des restes de Ceratodus africanusHaug proviennent de gisements crétacées (Cenomanien) de la Baía de SãoMarcos, MA, Brésil. Anais da Academia Brasileira de Ciências 51 (2), 361–362.
Cunha, F.L.S., Ferreira, C.S., 1980. Um Dipnoi da Formação Itapecuru (Cenomaniano),Maranhão, Brasil. In: Congresso Argentino de Paleontología y Biostratigrafía yCongresso Latinoamericano de Paleontología, 2/1, 1980, Actas, pp. 1–9.
Curry Rogers, K., 2005. Titanosauria: a phylogenetic overview. In: Curry Rogers, K.,Wilson, J.A. (Eds.), The Sauropods: Evolution and Paleobiology. University ofCalifornia Press, Berkeley, California, USA, pp. 50–103.
Dal Sasso, C., Maganuco, S., Buffetaut, E., Mendez, M.A., 2005. New information onthe skull of the enigmatic theropod Spinosaurus, with remarks on its size andaffinities. Journal of Vertebrate Paleontology 25 (4), 888–896.
de Broin, F., Taquet, P., 1966. Découverte d’un Crocodilien nouveau dans le Crétacéinférieur du Sahara. Comptes Rendus de l’Academie des Sciences, Paris, Série D262, 2326–2329.
Dias Lima, R., Rossetti, D.F., 1999. Análise faciológica e estratigráfica de depósitos doCretáceo Superior, leste da Bacia do Grajaú, Maranhão. Boletim do 5� Simpósiosobre o Cretáceo do Brasil, 237–241.
Dutheil, D.B., 1999. The first articulated fossil cladistian: Serenoichthys kemkemensis,gen. et sp. nov., from the Cretaceous of Morocco. Journal of VertebratePaleontology 19 (2), 243–246.
Elias, F.A., 2006. Dentes de amniotas da ‘‘Laje do Coringa’’ (Formação Alcântara,Albo-Cenomaniano da Bacia de São Luís-Grajaú). Identificação, descrição,aspectos paleobiológicos, biocronológicos, paleogeográficos e paleobiogeo-gráficos. Master’s thesis. Universidade Estadual Paulista, Rio Claro, Brazil, 223 p.
Elias, F.A., Bertini, R.J., Medeiros, M.A., 2005. Review of the occurrences concerningisolated amniotes teeth, in the Cretaceous deposits from the Maranhão State. In:II Congresso Latino-Americano de Paleontologia de Vertebrados, Boletim deResumos, pp. 99–100.
Elias, F.A., Bertini, R.J., Medeiros, M.A.A., 2007a. Pterosaur teeth from the Laje doCoringa, middle Cretaceous, São Luís-Grajaú basin, Maranhão state, Northern–Northeastern Brazil. Revista Brasileira de Geociências 37 (4), 668–676.
Elias, F.A., Bertini, R.J., Medeiros, M.A.A., 2007b. Velociraptorinae (Manira-ptoriformes) teeth from the Coringa Flagstone outcrop, middle Cretaceous ofthe São Luís-Grajaú Basin, Maranhão State, Northern–Northeastern Brazil. In:Carvalho, I.S., Cassab, R.C.T., Schwanke, C., Carvalho, M.A., Fernandes, A.C.S.,Rodrigues, M.A.C., Carvalho, M.S.S., Arai, M., Oliveira, M.E.Q. (Eds.),Paleontologia: Cenários de Vida. Interciência, Rio de Janeiro, Brazil, pp. 307–317.
Eugênio, W.S., 1994. Caracterização geopaleontológica do Cretáceo da Baía de SãoMarcos, Maranhão, Brasil. Master’s thesis. Universidade Federal do Rio deJaneiro, Rio de Janeiro, Brazil, 74 p.
Ferreira, C.S., Vicalvi, M.A., Carvalho, I.S., 1995. A tafocenose da Ilha Guarapirá (Baciade São Luís, Formação Itapecuru, Cretáceo Superior). Anais da AcademiaBrasileira de Ciências 67 (3), 381–382.
Filippi, L.S., García, R.A., Garrido, A.C., in press. A new titanosaur sauropod dinosaurfrom the Upper Cretaceous of North Patagonia, Argentina. Acta PalaeontologicaPolonica.
Forster, C.A., 1999. Gondwanan dinosaur evolution and biogeographic analysis.Journal of African Earth Sciences 28 (1), 169–185.
Freire, P.C., Medeiros, M.A., Lindoso, R.M., 2007. Sauropod teeth diversity in the Lajedo Coringa fossiliferous site, Eocenomanian of northeastern Brazil. In: Carvalho,I.S., Cassab, R.C.T., Schwanke, C., Carvalho, M.A., Fernandes, A.C.S., Rodrigues,M.A.C., Carvalho, M.S.S., Arai, M., Oliveira, M.E.Q. (Eds.), Paleontologia: Cenáriosde Vida. Interciência, Rio de Janeiro, Brazil, pp. 515–524.
Furtado, M.R., Candeiro, C.R.A., 2009a. The spinosauroid worldwide species andtheir paleogeographic implications. Ameghiniana 46 (Suppl. 4), 26R.
Furtado, M.R., Candeiro, C.R.A., 2009b. First Baryonychinae record in Brazil.Ameghiniana 46 (Suppl. 4), 27R.
Garcia, A.J.V., da Rosa, A.A.S., Goldberg, K., 2005. Paleoenvironmental andpaleoclimatic control on early diagenetic processes and fossil record inCretaceous continental sandstones of Brazil. Journal of South American EarthSciences 19 (3), 243–258.
Gayet, M., Meunier, F.J., Werner, C., 2002. Diversification in Polypteriformes andspecial comparison with the Lepisosteiformes. Palaeontology 45 (2), 361–376.
Gianechini, F.A., Makovicky, P.J., Apesteguía, S., in press. The teeth of theunenlagiine theropod Buitreraptor from the Cretaceous of Patagonia,
Argentina, and the unusual dentition of the Gondwanan dromaeosaurids. ActaPalaeontologica Polonica.
Gomani, E.M., 1997. A crocodyliform from the Early Cretaceous Dinosaur Beds,northern Malawi. Journal of Vertebrate Paleontology 17 (2), 280–294.
Gradstein, F.M., Ogg, J.G., Smith, A., 2004. A Geologic Time Scale 2004. CambridgeUniversity Press, Cambridge, UK. 589 p.
Guedes, E., Heilbron, M., Vasconcelos, P.M., de Morisson Valeriano, C., Horta deAlmeida, J.C., Teixeira, W., Filho, A.T., 2005. K–Ar and 40Ar/39Ar ages of dikesemplaced in the onshore basement of the Santos Basin, Resende area, SE Brazil:implications for the south Atlantic opening and Tertiary reactivation. Journal ofSouth American Earth Sciences 18 (3–4), 371–382.
Haug, E., 1905. Paléontologie. V. Couches crétacées à Ceratodus du Djoua. In:Foureau, F. (Ed.), Documents Scientifiques de la Mission Saharienne (MissionFoureau-Lamy) d’Alger au Congo par le Tchad. Société de Géographie, Paris,France, pp. 814–832.
Holtz Jr., T.R., Chapman, R.E., Lamanna, M.C., 2004. Mesozoic biogeography ofDinosauria. In: Weishampel, D.B., Dodson, P., Osmólska, H. (Eds.), TheDinosauria, second ed. University of California Press, Berkeley, California, USA,pp. 627–642.
Hone, D.W.E., Xu, X., Wang, D.-Y., 2010. A probable baryonychine (Theropoda:Spinosauridae) tooth from the Upper Cretaceous of Henan Province, China.Vertebrata PalAsiatica 48 (1), 19–26.
Jokat, W., Boebel, T., König, M., Meyer, U., 2003. Timing and geometry of earlyGondwana breakup. Journal of Geophysical Research 108 (B9, 2428), 1–15.
Karner, G.D., Gamboa, L.A.P., 2007. Timing and origin of the South Atlantic pre-saltsag basins and their capping evaporites. Geological Society, London, SpecialPublications 285, 15–35.
Kellner, A.W.A., Pinheiro, A.E.P., Azevedo, S.A.K., Henriques, D.D.R., Carvalho, L.B.,Oliveira, G.R., 2009. A new crocodyliform from the Alcântara Formation(Cenomanian), Cajual Island, Brazil. Zootaxa 2030, 49–58.
Kemp, A., 1997. A revision of Australian Mesozoic and Cenozoic lungfish of theFamily Neoceratodontidae (Osteichthyes: Dipnoi), with a description of fournew species. Journal of Paleontology 71 (4), 713–733.
Kemp, A., 1998. Skull structure in post-Paleozoic lungfish. Journal of VertebratePaleontology 18 (1), 43–63.
Kischlat, E.-E., Carvalho, I.S., 2000. A specimen of Araripemys barretoi Price (Chelonii,Pleurodira) from the Itapecuru Formation (Lower Cretaceous of NortheasternBrazil). Boletim de Resumos do 2 Simpósio Brasileiro de Paleontologia deVertebrados, 33.
Lamanna, M.C., Martínez, R.D., Smith, J.B., 2002. A definitive abelisaurid theropoddinosaur from the early Late Cretaceous of Patagonia. Journal of VertebratePaleontology 22 (1), 58–69.
Lapparent, A.F., 1960. Les dinosauriens du ‘Continental Intercalaire’ du Saharacentral. Mémoires de la Société Géologique de France 88A, 5–56.
Lapparent de Broin, F., de la Fuente, M.S., 2001. Oldest world Chelidae (Chelonii,Pleurodira), from the Cretaceous of Patagonia, Argentina. Comptes Rendus del’Académie des Sciences – Serie IIA – Sciences de la Terre et des Planètes 333 (8),463–470.
Lavocat, R.J.M., 1954. Sur les dinosauriens du Continental Intercalaire des Kem-Kemde la Daoura. Comptes Rendus de la Dix-Neuviéme Session, Congrès GéologiqueInternational. Alger 21 (1952), 65–68.
Le Loeuff, J., Métais, E., Dutheil, D.B., Rubino, J.L., Buffetaut, E., Lafont, F., Cavin, L.,Moreau, F., Tong, H., Blanpied, C., Sbeta, A., 2010. An Early Cretaceous vertebrateassemblage from the Cabao Formation of NW Libya. Geological Magazine 147(5), 750–759.
Lindoso, R.M., Elias, F.A., Medeiros, M.A., Barros Santos, R.A., Pereira, A.A., 2008.Paleoecological significance of pterosaurs teeth from the Alcântara Formation,Cretaceous of northeastern Brazil. In: Calvo, J.O., Juárez Valieri, R.D., Porfiri, J.D.,dos Santos, D.D. (Eds.), Actas III Congreso Latinoamericano de Paleontología deVertebrados. Neuquén 22–25 Septiembre, 2008. Universidad Nacional delComahue, Neuquén, Argentina, p. 138.
Macdonald, D., Gomez-Perez, I., Franzese, J., Spalletti, L., Lawver, L., Gahagan, L.,Dalziel, I., Thomas, C., Trewin, N., Hole, M., Paton, D., 2003. Mesozoic break-up ofSW Gondwana: implications for regional hydrocarbon potential of the southernSouth Atlantic. Marine and Petroleum Geology 20 (3–4), 287–308.
Machado, E., Azevedo, S., Carvalho, L., Henriques, D., Kellner, A., 2009. A newspinosaurid from the Cretaceous Alcântara Formation (Maranhão), northeasternBrazil. Journal of Vertebrate Paleontology 29 (Suppl. 3), 138A–139A.
Maisey, J.G., 2000. Continental break up and distribution of fishes in WesternGondwana during the Early Cretaceous. Cretaceous Research 21, 281–314.
Makovicky, P.J., Kobayashi, Y., Currie, P.J., 2004. Ornithomimosauria. In:Weishampel, D.B., Dodson, P., Osmólska, H. (Eds.), The Dinosauria, second ed.University of California Press, Berkeley, California, USA, pp. 137–150.
Makovicky, P.J., Apesteguía, S., Agnolin, F.L., 2005. The earliest dromaeosauridtheropod from South America. Nature 437, 1007–1011.
Mannion, P.D., 2009. A rebbachisaurid sauropod from the Lower Cretaceous of theIsle of Wight, England. Cretaceous Research 30 (3), 521–526.
Marinho, T.S., Carvalho, I.S., 2005. Répteis marinhos da Formação Alcântara(Cretáceo Superior), Maranhão. 1 Congresso Brasileiro de Herpetologia, 11 a16 de Julho–2004. Pontificia Universidade Católica do Paraná, Curitiba,Brazil.
Marinho, T.S., Gasparini, Z.B., Carvalho, I.S., Medeiros, M.A., 2004. Plesiosauria teethfrom Alcântara Formation (Cenomanian), Brazil. Ameghiniana 41 (4), 54R.
Marques da Silva, M.A., Medeiros, M.A., 2003. Ocorrência de Tribodus noEocenomaniano do Maranhão. In: Congresso Brasileiro de Paleontologia 18,Boletim de Resumos, pp. 276–277.
90 Carlos Roberto A. Candeiro et al. / Journal of African Earth Sciences 60 (2011) 79–92

Author's personal copy
Martill, D.M., 2007. The age of the Cretaceous Santana Formation fossil Konservat–Lagerstätte of north-east Brazil: a historical review and an appraisal of thebiochronostratigraphic utility of its biota. Cretaceous Research 28 (6), 895–920.
Martin, M., 1982. Nouvelles données sur la phylogénie et la systématique desdipneustes postpaléozoiques, conséquences stratigraphiques et paléogéo-graphiques. Geobios Mémoire Spéciale 6, 53–64.
Martin, M., 1984. Révision des arganodontidés et des néocératodontidés (Dipnoi,Ceratodontiformes) du Crétacé africain. Neues Jarbuch für Geologie undPaläontologie Abhandlungen 169 (2), 225–260.
Martinelli, A.G., Garrido, A.C., Forasiepi, A.M., Paz, E.R., Gurovich, Y., 2007. Notes onfossil remains from the Early Cretaceous Lohan Cura Formation, NeuquénProvince, Argentina. Gondwana Research 11 (4), 537–552.
Martínez, R.D., Giménez, O., Rodríguez, J., Luna, M., Lamanna, M.C., 2004. Anarticulated specimen of the basal titanosaurian (Dinosauria: Sauropoda)Epachthosaurus sciuttoi from the early Late Cretaceous Bajo Barreal Formationof Chubut Province, Argentina. Journal of Vertebrate Paleontology 24 (1), 107–120.
Medeiros, M.A., 2001. A Laje do Coringa (ilha do Cajual, bacia de São Luís, baía deSão Marcos, MA): conteúdo fossilífero, bioestratinomia, diagênese e implicaçõesna paleobiogeografia do Mesocretáceo do nordeste brasileiro. Doctoraldissertation. Universidade Federal do Rio Grande do Sul, Porto Alegre, Brazil,137 p.
Medeiros, M.A., 2002. Ocorrência de um saltassaurino (Dinosauria, Titanosauridae)no Cenomaniano do nordeste do Brasil. In: Boletim do 6� Simpósio sobre oCretáceo no Brasil, 2� Simpósio sobre el Cretácico de América del Sur, pp. 99–102.
Medeiros, M.A., 2006. Large theropod teeth from the Eocenomanian of northeasternBrazil and the occurrence of Spinosauridae. Revista Brasileira de Paleontologia 9(3), 333–338.
Medeiros, M.A., Avilla, L.S., 2005. Revisão sistemática dos dinosauros saurópodes daFormação Alcântara (Cenomaniano), Maranhão, Brasil. In: Congresso Brasileirode Paleontologia 19, Resumos (CD-ROM).
Medeiros, M.A., Schultz, C.L., 2001a. Uma paleocomunidade de vertebrados doCretáceo medio, Bacia de São Luís. In: Rossetti, D.F., Goés, A.M., Truckenbrodt,W. (Eds.), O Cretáceo na Bacia de São Luís-Grajaú. Museu Paraense EmílioGoeldi, Belém, Brazil, pp. 209–221.
Medeiros, M.A., Schultz, C.L., 2001b. Rebbachisaurus (Sauropoda, Diplodocimorpha)no Mesocretáceo do nordeste do Brasil. In: 13 Encontro de Zoologia doNordeste, Programa e Resumos, p. 166.
Medeiros, M.A., Schultz, C.L., 2002. A fauna dinossauriana da Laje do Coringa,Cretáceo médio do Nordeste do Brasil. Arquivos do Museu Nacional 60 (3), 155–162.
Medeiros, M.A., Schultz, C.L., 2003. Rebbachisaurus? (Sauropoda) na FormaçãoAlcântara, Cenomaniano do Maranhão. In: Congresso Brasileiro dePaleontologia 18, Boletim de Resumos, pp. 187–188.
Medeiros, M.A., Schultz, C.L., 2004. Rayososaurus (Sauropoda, Diplodocoidea) nomeso-Cretáceo do Norte–Nordeste Brasileiro. Revista Brasileira dePaleontologia 7 (2), 255–279.
Medeiros, M.A., Vilas Bôas, I., 1999. A Laje do Coringa: uma paleocomunidadecontinental do Cenomaniano do nordeste do Brasil. In: Congresso Brasileiro dePaleontologia 16, Boletim de Resumos, pp. 70–71.
Medeiros, M.A., Freire, P.C., Pereira, A.A., Santos, R.A.B., Lindoso, R.M., Coêlho, A.F.A.,Passos, E.B., Sousa Jr., E., 2007. Another African dinosaur recorded in theEocenomanian of Brazil and a revision on the paleofauna of the Laje do Coringasite. In: Carvalho, I.S., Cassab, R.C.T., Schwanke, C., Carvalho, M.A., Fernandes,A.C.S., Rodrigues, M.A.C., Carvalho, M.S.S., Arai, M., Oliveira, M.E.Q. (Eds.),Paleontologia: Cenários de Vida. Interciência, Rio de Janeiro, Brazil, pp. 413–423.
Meunier, F.J., Gayet, M., 1996. A new polypteriform from the Late Cretaceous andthe middle Paleocene of South America. In: Arratia, G., Viohl, G. (Eds.), MesozoicFishes – Systematics and Paleoecology. Verlag Dr. Friedrich Pfeil, Munich,Germany, pp. 95–103.
Milner, A., Buffetaut, E., Suteethorn, V., 2007. A tall-spined spinosaurid theropodfrom Thailand and the biogeography of spinosaurs. Journal of VertebratePaleontology 27 (Suppl. 3), 118A.
Molnar, R.E., Wiffen, J., 2007. A Presumed Titanosaurian Vertebra from the LateCretaceous of North Island, New Zealand, vol. 65(4). Arquivos do MuseuNacional, Rio de Janeiro, pp. 505–510.
Moraes-Santos, H.M., Melo, C.C.S., Toledo, P.M., Rossetti, D.F., 2001a. Ocorrência dePleurodira na Formação Alcântara (Albiano-Cenomaniano), Bacia de São Luis,MA. in: Congresso Brasileiro de Paleontologia 17, Resumos, p. 165.
Moraes-Santos, H.M., Melo, C.C.S., Toledo, P.M., Rossetti, D.F., 2001b. Ocorrência dePleurodira na Formação Alcântara (Albiano-Cenomaniano), Bacia de São Luís,MA. In: Rossetti, D.F., Goés, A.M., Truckenbrodt, W. (Eds.), O Cretáceo na Baciade São Luís-Grajaú. Museu Paraense Emílio Goeldi, Belém, Brazil, pp. 151–174.
Nobre, P.H., 2004. Morfologia pós-craniana de Candidodon itapecuruense(Crocodylomorpha, Mesoeucrocodylia) do Cretáceo do Brasil. RevistaBrasileira de Paleontologia 7 (1), 87–92.
Nobre, P.H., Carvalho, I.S., 2002. Osteologia do crânio de Candidodon itapecuruense(Crocodylomorpha, Mesoeucrocodylia) do Cretáceo do Brasil. In: Boletim do 6�Simpósio sobre o Cretáceo no Brasil, 2� Simpósio sobre el Cretácico de Américadel Sur, pp. 77–82.
Nobre, P.H., Medeiros, M.A., Vasconcelos, F.M., 2002. Crocodylomorpha da FormaçãoItapecuru (Cretáceo Superior – Bacia de São Luís) na Laje do Coringa, Ilha doCajual – MA. In: XLI Congresso Brasileiro de Geologia – A Geologia E O Homem,Anais, p. 669.
Nothdurft, W.E., Smith, J.B., Lamanna, M.C., Lacovara, K.J., Poole, J.C., Smith, J.R.,2002. The Lost Dinosaurs of Egypt. Random House, New York, New York, USA.256 p.
Novas, F.E., 1998. Megaraptor namunhuaiquii, gen. et sp. nov., a large-clawed LateCretaceous theropod from Patagonia. Journal of Vertebrate Paleontology 18 (1),4–9.
Novas, F.E., 2009. The Age of Dinosaurs in South America. Indiana University Press,Bloomington and Indianapolis, Indiana, USA. 458 p.
Novas, F.E., Dalla Vecchia, F., Pais, D.F., 2005a. Theropod pedal unguals from the LateCretaceous (Cenomanian) of Morocco, Africa. Revista del Museo Argentino deCiencias Naturales, n.s. 7 (2), 167–175.
Novas, F.E., de Valais, S., Vickers-Rich, P., Rich, T., 2005b. A Large Cretaceoustheropod from Patagonia, Argentina, and the evolution of carcharodontosaurids.Naturwissenschaften 92 (5), 226–230.
Novas, F.E., Pais, D.F., Pol, D., Carvalho, I.S., Mones, A., Scanferla, A., Riglos, M.S.,2009. Bizarre notosuchian crocodyliform with associated eggs from theUpper Cretaceous of Bolivia. Journal of Vertebrate Paleontology 29 (4), 1316–1320.
O’Connor, P.M., Sertich, J.J.W., Stevens, N.J., Roberts, E.M., Gottfried, M.D.,Hieronymus, T.L., Jinnah, Z.A., Ridgely, R., Ngasala, S.E., Temba, J., 2010. Theevolution of mammal-like crocodyliforms in the Cretaceous Period ofGondwana. Nature 466, 748–751.
Oliveira, G.R., Romano, P.S.R., 2007. Histórico dos achados de tartarugas fósseis doBrasil, vol. 65(1). Arquivos do Museu Nacional, Rio de Janeiro, pp. 113–133.
Ortega, F., Escaso, F., Sanz, J.L., 2010. A bizarre, humped Carcharodontosauria(Theropoda) from the Lower Cretaceous of Spain. Nature 467, 203–206.
Pedrão, E., Arai, M., Barrilari, I.M.R., Carvalho, I.S., 1993. Análise palinológica de umaamostra de superfície de Querru (Formação Itapecuru), Município de Itapecuru-Mirim – MA. PETROBRAS/CENPES (Technical Report).
Pereira, A.A., Medeiros, M.A., 2003. Novas ocorrências de peixes no Eocenomanianodo Maranhão. In: Congresso Brasileiro de Paleontologia 18, Boletim de Resumos,pp. 221–222.
Pereira, A.A., Medeiros, M.A., 2007. A new Elasmobranchii form from the AlcântaraFormation (Itapecuru deposits; Eocenomanian of Brazil). In: CongressoBrasileiro de Paleontologia 20, p. 18.
Pereira, A.A., Medeiros, M.A., 2008. A new sclerorhynchiform (Elasmobranchii) fromthe middle Cretaceous of Brazil. Revista Brasileira de Paleontologia 11 (3), 207–212.
Petri, S., 1987. Cretaceous paleogeographic maps of Brazil. Palaeogeography,Palaeoclimatology, Palaeoecology 59, 117–168.
Peyer, B., 1925. Ergebnisse der Forschungsreisen Prof. E. Stromers in den WüstenÄgyptens, II. Wirbeltier–Reste der Baharijestufe (unterstes Cenoman), 6. DieCeratodus-Funde. Abhandlungen der Bayerischen Akademie der Wissen-schaften. Mathemetisch-naturwissenschaftliche Abteilung 30 (5), 1–23.
Pitman III, W.C., Cande, S., LaBrecque, J., Pindell, J., 1993. Fragmentation ofGondwana: the separation of Africa from South America. In: Goldblatt, P.(Ed.), Biological Relationships between Africa and South America. YaleUniversity Press, New Haven, Connecticut, USA, pp. 15–34.
Pletsch, T., Erbacher, J., Holbourn, A.E.L., Kuhnt, W., Moullade, M., Oboh-Ikuenobede,F.E., Söding, E., Wagner, T., 2001. Cretaceous separation of Africa and SouthAmerica: the view from the West African margin (ODP Leg 159). Journal ofSouth American Earth Sciences 14 (2), 147–174.
Price, L.I., 1947. Sedimentos Mesozóicos na Baía de São Marcos, Estado doMaranhão. Ministério da Agricultura, Departamento Nacional de ProduçãoMineral, Divisão de Geologia e Mineralogia, Notas Preliminaires e Estudos 40,pp. 1–7.
Price, L.I., 1960. Dentes de Theropoda num testemunho de sonda no Estado deAmazonas. Anais da Academia Brasileira de Ciências 32 (1), 79–84.
Priem, F., 1914. Sur des vertébrés du Crétacé et de l’Éocène d’Égypte. Bulletin de laSociété Géologique de France 14, 366–382.
Rauhut, O.W.M., 1995. Zur systematischen stellung der afrikanischen TheropodenCarcharodontosaurus Stromer 1931 und Bahariasaurus Stromer 1934. BerlinerGeowissenschaftliche Abhandlungen Reihe E 16, 357–375.
Rauhut, O.W.M., 1999. A dinosaur fauna from the Late Cretaceous (Cenomanian) ofnorthern Sudan. Paleontologia Africana 35, 61–84.
Reyment, R.A., Dingle, R.V., 1987. Palaeogeography of Africa during the CretaceousPeriod. Palaeogeography, Palaeoclimatology, Palaeoecology 59, 93–116.
Rossetti, D.F., 1997. Internal architecture of mixed tide- and storm-influenceddeposits: an example from the Alcântara Formation, northern Brazil.Sedimentary Geology 114 (1–4), 163–169.
Rossetti, D.F., 2001. Arquitetura deposicional da Bacia de São Luís-Grajaú. In:Rossetti, D.F., Goés, A.M., Truckenbrodt, W. (Eds.), O Cretáceo na Bacia de SãoLuís-Grajaú. Museu Paraense Emílio Goeldi, Belém, Brazil, pp. 31–46.
Rossetti, D.F., Truckenbrodt, W., 1997. Revisão estratigráfica para os depósitos doAlbiano–Terciario Inferior (?) na Bacia de São Luís (MA), norte do Brasil. Boletimdo Museu Paraense Emílio Goeldi, Série Ciências da Terra 9, 29–41.
Rossetti, D.F., Truckenbrodt, W., 1999. Nomenclatura estratigrafica de depositosCretáceos expostos na Bacia de São Luís (MA). In: Boletim do 5� Simpósio sobreo Cretáceo do Brasil, pp. 485–490.
Russell, D.A., 1996. Isolated dinosaur bones from the Middle Cretaceous of theTafilalt, Morocco. Bulletin du Muséum National d’Histoire Naturelle, Paris, 4esérie 18 (C, 2–3), 349–402.
Salgado, L., Bonaparte, J.F., 1991. Un nuevo saurópodo Dicraeosauridae,Amargasaurus cazaui gen. et sp. nov., de la Formación La Amarga,Neocomiano de la Provincia del Neuquén, Argentina. Ameghiniana 28 (3–4),333–346.
Carlos Roberto A. Candeiro et al. / Journal of African Earth Sciences 60 (2011) 79–92 91

Author's personal copy
Salgado, L., Garrido, A., Cocca, S.E., Cocca, J.R., 2004. Lower Cretaceousrebbachisaurid sauropods from Cerro Aguada del León (Lohan CuraFormation), Neuquén Province, northwestern Patagonia, Argentina. Journal ofVertebrate Paleontology 24 (4), 903–912.
Salgado, L., Canudo, J.I., Garrido, A.C., Ruiz-Omeñaca, J.I., Garcia, R.A., de la Fuente,M.S., Barco, J.L., Bollati, R., 2009. Upper Cretaceous vertebrates from ElAnfiteatro area, Río Negro, Patagonia, Argentina. Cretaceous Research 30 (3),767–784.
Sánchez-Hernández, B., Benton, M.J., Naish, D., 2007. Dinosaurs and other fossilvertebrates from the Late Jurassic and Early Cretaceous of the Galve area, NESpain. Palaeogeography, Palaeoclimatology, Palaeoecology 249 (1–2), 180–215.
Schwarz, D., Ikejiri, T., Breithaupt, B.H., Sander, P.M., Klein, N., 2007. A nearlycomplete skeleton of an early juvenile diplodocid (Dinosauria: Sauropoda) fromthe Lower Morrison Formation (Late Jurassic) of north central Wyoming and itsimplications for early ontogeny and pneumaticity in sauropods. HistoricalBiology 19 (3), 225–253.
Sereno, P.C., Brusatte, S.L., 2008. Basal abelisaurid and carcharodontosauridtheropods from the Lower Cretaceous Elrhaz Formation of Niger. ActaPalaeontologica Polonica 53 (1), 15–46.
Sereno, P.C., Dutheil, D.B., Iarochene, M., Larsson, H.C.E., Lyon, G.H., Magwene, P.M.,Sidor, C.A., Varricchio, D.J., Wilson, J.A., 1996. Predatory dinosaurs from theSahara and Late Cretaceous faunal differentiation. Science 272, 986–991.
Sereno, P.C., Beck, A.L., Dutheil, D.B., Gado, B., Larsson, H.C.E., Lyon, G.H., Marcot, J.D.,Rauhut, O.W.M., Sadleir, R.W., Sidor, C.A., Varricchio, D.J., Wilson, G.P., Wilson,J.A., 1998. A long-snouted predatory dinosaur from Africa and the evolution ofspinosaurids. Science 282, 1298–1302.
Sereno, P.C., Beck, A.L., Dutheil, D.B., Larsson, H.C.E., Lyon, G.H., Moussa, B., Sadleir,R.W., Sidor, C.A., Varricchio, D.J., Wilson, G.P., Wilson, J.A., 1999. Cretaceoussauropods from the Sahara and the uneven rate of skeletal evolution amongdinosaurs. Science 286, 1342–1347.
Sereno, P.C., Larsson, H.C.E., Sidor, C.A., Gado, B., 2001. The giant crocodyliformSarcosuchus from the Cretaceous of Africa. Science 294, 1516–1519.
Sereno, P.C., Wilson, J.A., Conrad, J.L., 2004. New dinosaurs link southern landmassesin the Mid-Cretaceous. Proceedings of the Royal Society of London B 271 (1546),1325–1330.
Smith, J.B., Lamanna, M.C., Lacovara, K.J., Dodson, P., Smith, J.R., Poole, J.C.,Giegengack, R., Attia, Y., 2001. A giant sauropod dinosaur from an UpperCretaceous mangrove deposit in Egypt. Science 292, 1704–1706.
Smith, J.B., Lamanna, M.C., Mayr, H., Lacovara, K.J., 2006. New information regardingthe holotype of Spinosaurus aegyptiacus Stromer, 1915. Journal of Paleontology80 (2), 400–406.
Smith, J.B., Lamanna, M.C., Askar, A.S., Bergig, K.A., Tshakreen, S.O., Abugares, M.M.,Rasmussen, D.T., 2010. A large abelisauroid theropod dinosaur from the EarlyCretaceous of Libya. Journal of Paleontology 84 (5), 927–934.
Soto, M., Perea, D., 2010. Late Jurassic lungfishes (Dipnoi) from Uruguay, withcomments on the systematics of Gondwanan ceratodontiforms. Journal ofVertebrate Paleontology 30 (4), 1049–1058.
Sousa, E.P., Medeiros, M.A., Bertini, R.J., Pereira, A., Toledo, C.E.V., 2004. Ocorrênciainédita de picnodontiformes para a Laje do Coringa, ilha do Cajual (FormaçãoAlcântara), Eocenomaniano do estado do Maranhão. In: Simpósio Brasileiro dePaleontologia de Vertebrados 4, Boletim de Resumos, pp. 67–68.
Sousa, E.P., Toledo, C.E.V., Bertini, R.J., Medeiros, M.A., 2005. A new morphotype ofCeratodontidae (Dipnoiformes) from the Cretaceous deposits of the CoringaFlagstone, Cajual Island, Maranhão State, north/northeastern Brazil. In: IICongresso Latino-Americano de Paleontologia de Vertebrados, Boletim deResumos, pp. 252–253.
Stromer, E., 1931. Ergebnisse der Forschungsreisen Prof. E. Stromers in den WüstenÄgyptens, II. Wirbeltier-Reste der Baharîjestufe (unterstes Cenoman), 10. EinSkelett-Rest von Carcharodontosaurus nov. gen. Abhandlungen der BayerischenAkademie der Wissenschaften. Mathematisch-naturwissenschaftlicheAbteilung, Neue Folge 9, 1–23.
Stromer, E., 1934. Ergebnisse der Forschungsreisen Prof. E. Stromers in den WüstenÄgyptens, II. Wirbeltier–Reste der Baharîjestufe (unterstes Cenoman), 13.
Dinosauria. Abhandlungen der Bayerischen Akademie der Wissenschaften.Mathematisch-naturwissenschaftliche Abteilung, Neue Folge 22, 1–79.
Stromer, E., 1936. Ergebnisse der Forschungsreisen Prof. E. Stromers in den WüstenÄgyptens, VII. Baharîje-Kessel und -Stufe mit deren Fauna und Flora, Eineergänzende Zusammenfassung. Abhandlungen der Bayerischen Akademie derWissenschaften. Mathematisch-naturwissenschaftliche Abteilung, Neue Folge33, 3–102.
Sues, H.-D., Frey, E., Martill, D.M., Scott, D.M., 2002. Irritator challengeri, aspinosaurid (Dinosauria: Theropoda) from the Lower Cretaceous of Brazil.Journal of Vertebrate Paleontology 22 (3), 535–547.
Tabaste, N., 1963. Étude des restes de poissons du Crétacé saharien, Mémoire IFAN.Mélanges Ichthyologiques 68, 437–485.
Taquet, P., 1976. Geologie et paleontologie de gisement de Gadoufaoua. Cahiers dePaleontologie 1, 191.
Tello Saenz, C.A., Hackspacher, P.C., Hadler Neto, J.C., Iunes, P.J., Guedes, S., Ribeiro,L.F.B., Paulo, S.R., 2003. Recognition of Cretaceous, Paleocene, and Neogenetectonic reactivation through apatite fission-track analysis in Precambrian areasof southeast Brazil: association with the opening of the South Atlantic Ocean.Journal of South American Earth Sciences 15 (7), 765–774.
Toledo, C.E.V., Bertini, R.J., 2005. Occurrences of the fossil Dipnoiformes in Braziland its stratigraphic and chronological distributions. Revista Brasileira dePaleontologia 8 (1), 47–56.
Toledo, C.E.V., Bertini, R.J., Sousa, E.P., Medeiros, M.A., 2005. First occurrence ofProtopterus humei (Dipnoiformes) in South America. In: II Congresso Latino-Americano de Paleontologia de Vertebrados, Boletim de Resumos, pp. 262–263.
Turner, A.H., Buckley, G.A., 2008. Mahajangasuchus insignis (Crocodyliformes:Mesoeucrocodylia) cranial anatomy and new data on the origin of theeusuchian-style palate. Journal of Vertebrate Paleontology 28 (2), 382–408.
Upchurch, P., Hunn, C.A., Norman, D.B., 2002. An analysis of dinosaurianbiogeography: evidence for the existence of vicariance and dispersal patternscaused by geological events. Proceedings of the Royal Society of London B 269,613–621.
Vilas Bôas, I., 1999. Dentes de terópodes e associação fossilífera da Praia daBaronesa, Alcântara, MA (Formação Itapecuru, bacia de São Luís). In: CongressoBrasileiro de Paleontologia 16, Boletim de Resumos, p. 125.
Vilas Bôas, I., Carvalho, I.S., 2001. Répteis marinhos (Mosasauria e Plesiosauria) doCretáceo Superior da Bacia de São Luís (Maranhão, Brasil). In: Rossetti, D.F.,Goés, A.M., Truckenbrodt, W. (Eds.), O Cretáceo na Bacia de São Luís-Grajaú.Museu Paraense Emílio Goeldi, Belém, Brazil, pp. 223–233.
Vilas Bôas, I., Carvalho, I.S., Medeiros, M.A., Pontes, H., 1999. Dentes deCharcarodontosaurus (Dinosauria, Tyrannosauridae) do Cenomaniano, bacia deSão Luís (norte do Brasil). Anais da Academia Brasileira de Ciências 71 (4), 846–847.
Viramonte, J.G., Kay, S.M., Becchio, R., Escayola, M., Novitski, I., 1999. Cretaceous riftrelated magmatism in central–western South America. Journal of SouthAmerican Earth Sciences 12 (2), 109–121.
Werner, C., 1994. Die kontinentale Wirbeltierfauna aus der unteren Oberkreide desSudan (Wadi Milk Formation). Berliner Geowissenschafliche AbhandlungenReihe E 13, 221–249.
Werner, C., Gayet, M., 1997. New fossil Polypteridae from the Cenomanian of Sudan,an evidence of their high diversity in the early Late Cretaceous. Cybium 21 (1),67–81.
Whitlock, J.A., in press. A phylogenetic analysis of Diplodocoidea (Saurischia:Sauropoda). Zoological Journal of the Linnean Society.
Wilson, J.A., 2006. An overview of titanosaur evolution and phylogeny. In: ColectivoArqueológico–Paleontológico de Salas (Eds.), Actas de las III Jornadas sobrePaleontología de Dinosaurios y su Entorno, pp. 169–190.
Wilson, J.A., Upchurch, P., 2003. A revision of Titanosaurus Lydekker (Dinosauria–Sauropoda), the first dinosaur genus with a ‘Gondwanan’ distribution. Journal ofSystematic Palaeontology 1 (3), 125–160.
Yabumoto, Y., Uyeno, T., 2005. New materials of a Cretaceous coelacanth, Mawsonialavocati Tabaste from Morocco. Bulletin of the National Science Museum ofTokyo, Series C 31, 39–49.
92 Carlos Roberto A. Candeiro et al. / Journal of African Earth Sciences 60 (2011) 79–92




















