
Ligand-binding Characterization of Xanthophyll Carotenoids to Solubilized Membrane Proteins Derived...
12
Ligand-binding Characterization of Xanthophyll Carotenoids to Solubilized Membrane Proteins Derived from Human Retina ALEXANDER YU. YEMELYANOV a , NIKITA B. KATZ ab AND PAUL S. BERNSTEIN a * a Moran Eye Center, University of Utah School of Medicine, Salt Lake City, UT, U.S.A. and b Institute for Natural Resources Inc., Berkeley, CA, U.S.A. (Received Milwaukee 15 September 2000, accepted in revised form 28 November 2000 and published electronically 8 February 2001) The macula of the human retina contains extraordinarily high concentrations of lutein and zeaxanthin, xanthophyll carotenoids that appear to play an important role in protecting against age-related macular degeneration, the leading cause of blindness among the elderly. It is likely that the uptake and stabilization of these carotenoids is mediated by specific xanthophyll-binding proteins. In order to purify and characterize such a binding protein, a carotenoid-rich membrane fraction derived from human macula or peripheral retina was prepared by homogenization, differential centrifugation, and detergent solubilization. Further purification was carried out using ion-exchange chromatography and gel- filtration chromatography coupled with continuous photodiode-array monitoring for endogenously associated xanthophyll carotenoids. The most highly purified preparations contained two major protein bands at 25 and 55 kDa that consistently co-eluted with endogenous lutein and zeaxanthin. The visible absorbance spectrum of the binding protein preparation closely matches the spectral absorbance of the human macular pigment, and it is bathochromically shifted about 10 nm from the spectrum of lutein and zeaxanthin dissolved in organic solvents. Binding of exogenously added lutein and zeaxanthin is saturable and specific with an apparent K d of approximately 1 mM. Canthaxanthin and b-carotene exhibit no significant binding activity to solubilized retinal membrane proteins when assayed under identical conditions. Other potential mammalian xanthophyll-binding proteins such as albumin, tubulin, lactoglobulin and serum lipoproteins possess only weak non-specific binding affinity for carotenoids when assayed under the same stringent binding conditions. This investigation provides the first direct evidence for the existence of specific xanthophyll-binding protein(s) in the vertebrate retina and macula. The possible roles of xanthophyll-binding proteins in normal macular function and in the pathogenesis of age-related macular degeneration remain to be elucidated. # 2001 Academic Press Key words: macular pigment; binding protein; carotenoid; xanthophyll; age-related macular degener- ation; macula; retina; protein purification; membrane proteins; chromatography; lutein; zeaxanthin. 1. Introduction The macula of the human retina contains extra- ordinarily large amounts of two xanthophylls, lutein [(3R,3 0 R,6 0 R)-b,E-carotene-3,3 0 -diol] and zeaxanthin [a mixture of (3R,3 0 R)-b,b-carotene-3,3 0 -diol and (3R,3 0 S-meso)-b,b-carotene-3,3 0 -diol] (Bone, Landrum and Tarsis, 1985; Handelman et al., 1992; Bone et al., 1993). They are members of the much larger class of compounds known as carotenoids, most of which share a common C 40 H 56 isoprenoid backbone. They are classified as xanthophylls by virtue of the presence of at least one oxygen atom in each molecule, making them more polar than purely hydrocarbon carotenes such as b-carotene or lycopene. Humans can carry out only limited metabolic transformations of carotenoids; carotenes cannot be converted to xanthophylls, and xanthophylls cannot be converted to carotenes. All carotenoids in humans are exclusively derived from the diet, and it is possible to raise the levels of the macular xanthophyll pigment through dietary inter- ventions such as eating foods rich in lutein and zeaxanthin or by consuming purified supplements (Hammond et al., 1997; Landrum et al., 1997a). High dietary intakes and high blood levels of lutein and zeaxanthin have been linked epidemiologically to a significantly lower risk of visual loss from exudative age-related macular degeneration (AMD), the leading cause of legal blindness in the elderly of the U.S.A. and other developed countries (Eye Disease Case Control Study Group, 1993; Seddon et al., 1994). Also, a study of human autopsy eyes has reported that lutein and zeaxanthin levels are lower in eyes from patients with a history of AMD relative to eyes from age- matched control individuals without a known history of AMD (Landrum, Bone and Kilburn, 1997b). Approximately 50 % of the entire retina’s caroten- oids are concentrated in the macula, the central region of the retina (approximately 4 % of the total retinal area) responsible for high-resolution visual acuity (Handelman et al., 1992). Anatomically, the macular carotenoid pigment is localized principally to the Henle fiber layer and to the inner plexiform layer (Snodderly, Exp. Eye Res. (2001) 72, 381–392 doi:10.1006/exer.2000.0965, available online at http://www.idealibrary.com on 0014-4835/01/04038112 $35.00/0 # 2001 Academic Press * Address correspondence to: Paul S. Bernstein, Moran Eye Center, University of Utah School of Medicine, 50 North Medical Drive, Salt Lake City, UT, 84132, U.S.A. E-mail: paul.bernstein@hsc. utah.edu
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Ligand-binding Characterization of Xanthophyll Carotenoids to Solubilized Membrane Proteins Derived...

Exp. Eye Res. (2001) 72, 381±392doi:10.1006/exer.2000.0965, available online at http://www.idealibrary.com on
Ligand-binding Characterization of Xanthophyll Carotenoids toSolubilized Membrane Proteins Derived from Human Retina
ALEXANDER YU. YEMELYANOVa, NIKITA B. KATZab AND PAUL S. BERNSTEINa*
aMoran Eye Center, University of Utah School of Medicine, Salt Lake City, UT, U.S.A. andbInstitute for Natural Resources Inc., Berkeley, CA, U.S.A.
(Received Milwaukee 15 September 2000, accepted in revised form 28 November 2000 and published
All carfrom the d
0014-483
* AddressCenter, UniDrive, Salt Lutah.edu
electronically 8 February 2001)
The macula of the human retina contains extraordinarily high concentrations of lutein and zeaxanthin,xanthophyll carotenoids that appear to play an important role in protecting against age-related maculardegeneration, the leading cause of blindness among the elderly. It is likely that the uptake andstabilization of these carotenoids is mediated by speci®c xanthophyll-binding proteins. In order to purifyand characterize such a binding protein, a carotenoid-rich membrane fraction derived from humanmacula or peripheral retina was prepared by homogenization, differential centrifugation, and detergentsolubilization. Further puri®cation was carried out using ion-exchange chromatography and gel-®ltration chromatography coupled with continuous photodiode-array monitoring for endogenouslyassociated xanthophyll carotenoids. The most highly puri®ed preparations contained two major proteinbands at 25 and 55 kDa that consistently co-eluted with endogenous lutein and zeaxanthin. The visibleabsorbance spectrum of the binding protein preparation closely matches the spectral absorbance of thehuman macular pigment, and it is bathochromically shifted about 10 nm from the spectrum of luteinand zeaxanthin dissolved in organic solvents. Binding of exogenously added lutein and zeaxanthin issaturable and speci®c with an apparent Kd of approximately 1 mM. Canthaxanthin and b-caroteneexhibit no signi®cant binding activity to solubilized retinal membrane proteins when assayed underidentical conditions. Other potential mammalian xanthophyll-binding proteins such as albumin,tubulin, lactoglobulin and serum lipoproteins possess only weak non-speci®c binding af®nity forcarotenoids when assayed under the same stringent binding conditions. This investigation provides the®rst direct evidence for the existence of speci®c xanthophyll-binding protein(s) in the vertebrate retinaand macula. The possible roles of xanthophyll-binding proteins in normal macular function and in the
#
pathogenesis of age-related macular degeneration remain to be elucidated. 2001 Academic PressKey words: macular pigment; binding protein; carotenoid; xanthophyll; age-related macular degener-ation; macula; retina; protein puri®cation; membrane proteins; chromatography; lutein; zeaxanthin.
1. Introduction
The macula of the human retina contains extra-ordinarily large amounts of two xanthophylls, lutein[(3R,30R,60R)-b,E-carotene-3,30-diol] and zeaxanthin[a mixture of (3R,30R)-b,b-carotene-3,30-diol and(3R,30S-meso)-b,b-carotene-3,30-diol] (Bone, Landrumand Tarsis, 1985; Handelman et al., 1992; Bone et al.,1993). They are members of the much larger class ofcompounds known as carotenoids, most of whichshare a common C40H56 isoprenoid backbone. Theyare classi®ed as xanthophylls by virtue of the presenceof at least one oxygen atom in each molecule, makingthem more polar than purely hydrocarbon carotenessuch as b-carotene or lycopene. Humans can carry outonly limited metabolic transformations of carotenoids;carotenes cannot be converted to xanthophylls, andxanthophylls cannot be converted to carotenes.
otenoids in humans are exclusively derivediet, and it is possible to raise the levels of the
5/01/040381�12 $35.00/0
correspondence to: Paul S. Bernstein, Moran Eyeversity of Utah School of Medicine, 50 North Medicalake City, UT, 84132, U.S.A. E-mail: paul.bernstein@hsc.
macular xanthophyll pigment through dietary inter-ventions such as eating foods rich in lutein andzeaxanthin or by consuming puri®ed supplements(Hammond et al., 1997; Landrum et al., 1997a).High dietary intakes and high blood levels of luteinand zeaxanthin have been linked epidemiologically toa signi®cantly lower risk of visual loss from exudativeage-related macular degeneration (AMD), the leadingcause of legal blindness in the elderly of the U.S.A. andother developed countries (Eye Disease Case ControlStudy Group, 1993; Seddon et al., 1994). Also, astudy of human autopsy eyes has reported that luteinand zeaxanthin levels are lower in eyes from patientswith a history of AMD relative to eyes from age-matched control individuals without a known historyof AMD (Landrum, Bone and Kilburn, 1997b).
Approximately 50 % of the entire retina's caroten-oids are concentrated in the macula, the central regionof the retina (approximately 4 % of the total retinalarea) responsible for high-resolution visual acuity(Handelman et al., 1992). Anatomically, the macular
carotenoid pigment is localized principally to the Henle®ber layer and to the inner plexiform layer (Snodderly,# 2001 Academic Press

Auran and Delori, 1984a), and macular zeaxanthinpredominates over lutein by a ratio of approximately2 : 1 (Handelman et al., 1992; Landrum et al.,1997b). Based on anatomical and psychophysicalmeasurements, it is estimated that the concentrationof carotenoids at the human fovea (the center of themacula) is in the range of 1 mM (Landrum et al.,1999). By contrast, typical xanthophyll carotenoidconcentrations in human serum and tissue (liver,kidney, lung) are approximately 1000-fold lower atapproximately 0.3 and 2 mM, respectively (Schmitzet al., 1993; Hammond et al., 1997; Landrum et al.,1999). In the peripheral retina, the concentration perunit area of xanthophyll carotenoids is lower by afactor of 100 relative to the fovea, and luteinpredominates over zeaxanthin by a ratio of approxi-mately 2 : 1 (Bone et al., 1988; Landrum et al., 1999).
The uptake of lutein and zeaxanthin into the retinais quite speci®c. Unlike most other human tissues,abundant serum carotenoids such as a-carotene,b-carotene, lycopene, and the cryptoxanthins arefound in no more than trace amounts in the retina(Handelman et al., 1992; Schmitz et al., 1993),although oxidative metabolites of lutein and zea-xanthin are present in signi®cant quantities (Khachik,Bernstein and Garland, 1997). The cellular location ofcarotenoids in the macula and peripheral retina is notwell de®ned. In the macula, they are thought to beassociated with the cone axons (Snodderly et al.,1984a) or the Muller glial cells (Gass, 1999), while inthe peripheral retina they may be largely localized tothe rod outer segments (Sommerberg et al., 1999;Rapp, Maple and Choi, 2000).
Various hypotheses have been offered to accountfor the physiological signi®cance of the selectiveaccumulation of lutein and zeaxanthin in the maculaand peripheral retina (Schalch, Dayhaw-Barker andBarker, 1999). These include ®ltration of potentiallydamaging short-wavelength visible light, quenching ofphotochemically-induced free-radicals, attenuation ofchromatic aberration, or inhibition of apoptosis (Doreyet al., 1997; Balashova, Balashov and Bernstein,1998; Schalch et al., 1999; Sujak et al., 1999).
Whenever a tissue exhibits highly selective uptakeof a chemical compound, it is quite likely that speci®cbinding proteins are involved. For example, retinoidsare also heavily concentrated in the retina due inlarge part to the actions of retinoid-binding proteinssuch as serum retinol-binding protein, cellular retinol-binding protein, cellular retinal-binding protein,interstitial retinoid-binding protein, and rhodopsin,all of which play important roles in the uptake andfunction of vitamin A compounds in the eye (Saari,1994). It is therefore reasonable to hypothesize thatcomparable members of a xanthophyll-bindingprotein (XBP) family should be present in thehuman macula and retina to mediate the uptake,
382
stabilization and function of the high concentrationsof lutein and zeaxanthin.
Carotenoid-binding proteins derived from plantsand invertebrates have been puri®ed, cloned and evencrystallized. The plant and algal proteins are usuallyinvolved in photosynthesis or photoprotection. Pro-minent examples include several membrane-associ-ated and water-soluble carotenoid-binding proteinsisolated from cyanobacteria, prochlorophytes andhigher plants that bind various xanthophylls and/orcarotenes (Lakshman and Okoh, 1993; Reddy,Bullerjahn and Sherman, 1993). a-Crustacyanin isthe 350 kDa astaxanthin-binding protein responsiblefor the color of lobster shells (Zagalsky, 1995). Itconsists of 16 20 kDa subunits, each of which carriesone astaxanthin chromophore (Keen et al., 1991a).The complete sequences of its A2 and C1 subunits areknown, demonstrating that it is a member of thehydrophobic-ligand-carrier protein superfamily thatincludes some retinoid-binding proteins (Keen et al.,1991a,b). A lutein-binding protein that may beinvolved in the speci®c uptake of dietary lutein inthe midgut of the silkworm has been puri®ed andpartially characterized (Jouni and Wells, 1996). Theamino acid sequence of this 35 kDa protein has notyet been reported.
Much less is known about carotenoid-bindingproteins derived from vertebrates. A 67 kDa solubleb-carotene-binding protein has been isolated fromferret liver (Rao, Ghosh and Lakshman, 1997). Itexhibits a binding preference for carotenes overxanthophylls, but its sequence has not been reported.In the deep-sea hatchet®sh and the diurnal gecko,unidenti®ed carotenoids are bound to crystallins ofthe lens (McFall-Ngai et al., 1986; RoÈll, 2000). Inmilk, b-lactoglobulin may be a carotenoid carrier(Dufour and Haertle, 1991). In the human blood-stream, high-density lipoprotein (HDL) is the majorcarrier of lutein and zeaxanthin, while carotenes arepreferentially carried by low-density lipoprotein (LDL)(Clevidence and Bieri, 1993). In the mammalian eye,it has been reported that retinal tubulin bindsmacular carotenoids (Bernstein et al., 1997), possiblyas a site for passive deposition in the tissue in thesame manner as actin which binds astaxanthin insalmon muscle (Hemmi, Hata and Takeuchi, 1991).
Previous studies on mammalian carotenoid-bindingproteins have focused on the water-soluble fractionslargely due to the relative ease of puri®cation, but ithas been suggested that a substantial portion of thelutein and zeaxanthin of the human macula andretina is membrane-associated (Bone, Landrum andCains, 1992). We report in this article that thephysiologically relevant ocular carotenoids lutein andzeaxanthin bind saturably and speci®cally to solubil-ized membrane proteins derived from human retinaand macula. This investigation provides the ®rst directevidence for the existence of speci®c xanthophyll-
A. Y. YEMELYANOV ET AL.
binding protein(s) in the vertebrate retina andmacula.

molecular weight cut-off 10 kDa) and further puri®ed
2. Materials and Methods
Laboratory Reagents
Laboratory reagents were usually purchased fromSigma (St. Louis, MO, U.S.A.), Calbiochem (La Jolla,CA, U.S.A.), or Amresco (Solon, OH, U.S.A.). Rare andunusual carotenoids were gifts from Kemin Foods ofDes Moines, IA, U.S.A. (lutein), Hoffmann-La Rocheof Basel, Switzerland (zeaxanthin, meso-zeaxanthin,isozeaxanthin, b-cryptoxanthin, astaxanthin,canthaxanthin), or Dr Fred Khachik of College Park,MD, U.S.A. (30-epilutein). These carotenoids were usedwithout further puri®cation, and purity and stabilityof stock solutions stored at ÿ708C was con®rmed byHPLC with photodiode-array detection. Extinctioncoef®cients and absorbance maxima for the com-pounds were obtained from a published source(Britton, 1995). Whenever possible, procedures invol-
XANTHOPHYLL BINDING TO RETINAL PROTEINS
ving carotenoids were performed under subdued light
with samples kept on ice or in a coldroom.Preparation of Human Maculae
Human donor globes were obtained within 24 hr ofdeath from the Utah Lions Eye Bank after corneas hadbeen harvested for transplantation. Ninety percentwere obtained between 10 and 24 hr after death.Extremely fresh tissue harvested 6 hr after deathexhibited no detectable differences during chromato-graphic puri®cations or carotenoid-binding assays vshuman donor tissue stored up to 72 hr at 48C. Tissueprocurement and usage complied with institutionalpolicies and the tenets of the Declaration of Helsinki.The iris, lens, ciliary body and anterior sclera weredissected out under subdued light, and most of thevitreous was removed. The macula was localized andtrephined with a 5 mm circular punch. Globes withobservable macular pathology were excluded. Har-vested maculae and peripheral retinas were separatedfrom underlying retinal pigment epithelium andoverlying vitreous and stored at ÿ208C for up to 1
month with no apparent loss of carotenoid-bindingbuffer conditions were 20 mM Tris buffer (pH 7.8)
activity.
Preparation of Solubilized Membrane Extracts fromHuman Macula or Peripheral Retina
Thawed human macular or peripheral retinaltissues originating from two to ten eyes werehomogenized on ice with brief sonication (510 sec)using a model F60 Sonic Dismembrator ®tted with atitanium probe (Fisher Scienti®c, Pittsburgh, PA,U.S.A.) at 2±5 W into 0.5±5.0 ml of a cell-lysingsolution containing protease inhibitors (200 mM leu-peptin, 2 mg mlÿ1 aprotinin, 0.2 mg mlÿ1 a2-macro-globulin, and 1 mM phenylmethylsulfonyl¯uoride) in20 mM Tris buffer (pH 7.4) supplemented with 1 mM
CaCl2 , 2 mM MgCl2 , and 1 mM dithiothreitol (DTT).The homogenate was then centrifuged at 5000 g for
10 min at 28C, and the pellet was discarded. A high-speed centrifugation of the low-speed supernatant wasperformed at 200 000 g for 60 min at 28C. The high-speed supernatant was discarded, and the yellowhigh-speed pellet was solubilized with brief sonicationon ice into 500 ml of 25 mM 3-[(3-cholamidopropyl)-dimethylammonio]-1-propanesulfonate (CHAPS) in20 mM 4-morpholineethanesulfonic acid (MES) buffer(pH 5.5) containing 1 mM CaCl2 , 2 mM MgCl2 , and1 mM DTT, followed by centrifugation at 100 000 gfor 60 min at 28C. The CHAPS insoluble pellet wasresolubilized into CHAPS/MES several more timesfollowed by high-speed centrifugation as describedabove. The pale yellow CHAPS-solubilized supernatantwas concentrated using an Ultrafree centrifugal®ltration system (Millipore, Woburn, MA, U.S.A.;
383
as described below.
Ion-exchange Chromatography
Protein chromatography was performed on aBioLogic liquid chromatography system (Bio-Rad,Hercules, CA, U.S.A.) ®tted with nonmetallic bio-inerttubing and couplings. The chromatograph and alleluting solvents were kept in a coldroom to maximizeprotein stability during puri®cation. Detergent-solubil-ized human macular membrane extracts were loadedimmediately after preparation onto a Bio-Rad UNO-Scation exchange column (7 � 35 mm, SOÿ3 exchan-ging groups). Initial buffer conditions were 20 mM MESbuffer containing 8 mM CHAPS at pH 5.5 run at0.5 ml minÿ1 at 48C for 15 min followed by a lineargradient from 0 to 1 M NaCl over 15 min. The eluatewas monitored by a UV6000LP photodiode-arrayspectrophotometer equipped with a high sensitivity50 mm pathlength light-pipe ¯owcell with an internalvolume of 10 ml (Thermo Separations, San Jose, CA,U.S.A.), and 300 ml fractions were collected. The peakfractions containing carotenoids were identi®ed byanalysis of their characteristic photodiode-array spec-tra (peak absorbance at approximately 450 nm withshoulder peaks at approximately 425 and 475 nm). Atlater puri®cation steps, anion-exchange chromatog-raphy was performed on a Bio-Rad UNO-Q column[7 � 35 mm, N(CH3)�3 exchanging groups]. Initial
containing 8 mM CHAPS.
Gel-®ltration Chromatography
The peak carotenoid-containing fractions from theion-exchange step were combined, concentrated to100 ml using a centrifugal concentrator with a 10 kDacut-off, and loaded onto a 300 � 7.5 mm BioSep-SEC-S3000 bio-inert silica-based gel-®ltration column(Phenomenex, Torrance, CA, U.S.A.; separation
range 5±700 kDa) eluted with 50 mM sodiumphosphate buffer (pH 6.8) containing 8 mM CHAPS
run at 0.5 ml minÿ1 at 48C. Photodiode-arraymonitoring and fraction collection were as described
384
above. In order to improve protein purity, the ®nal gel-
®ltration step was usually repeated two or three times.Protein Analysis
Protein assays were performed by the Micro BCAmethod with bovine serum albumin as the standard(Pierce, Rockford, IL, U.S.A.). SDS±PAGE on centri-fugally concentrated portions of fractions was doneusing Bio-Rad pre-cast 10±20 % Tris±tricine gels withappropriate buffers. Gel staining was with Pierce
colloidal coomassie or with a Bio-Rad Silver Stain PlusPrism version 3.00 package for Windows (GraphPad
kit.
Carotenoid Analysis
Samples for carotenoid analysis were diluted to200 ml with an appropriate amount of deionizedwater followed by the addition of 200 ml of methanolto denature proteins. Carotenoids were then extractedinto 200 ml of hexane containing 0.5 % butylated-hydroxytolune (BHT). The organic layer was removedby pipette, and the aqueous layer was extracted asecond time with 200 ml of hexane. No residualcarotenoids were detectable when the aqueous layerwas extracted a third time. The two organic extractswere combined, and 200 ml portions were injectedonto a Waters HPLC system (Milford, MA, U.S.A.)®tted with a 4.6 � 250 mm Microsorb-MV (5 mm,100 AÊ ) silica-based nitrile column (Varian, WalnutCreek, CA, U.S.A.). The eluant was 80 % hexane,19.4 % dichloromethane, 0.5 % methanol, and 0.1 %N,N0-diisopropylethylamine (v/v/v/v) at 1 ml minÿ1
at room temperature, and monitoring was at 450 nm.Peak identities were con®rmed by photodiode-arrayspectra and by co-elution with authentic standards asnecessary. Lutein and zeaxanthin peaks were inte-grated using BORWIN software (JMBS Developpe-ments, Le Fontanil, France), and integrated areas
were converted to nanograms using externalstandardization with pure lutein and zeaxanthin.TABLE
Partial puri®cation of XBP fro
Puri®cation stepLutein
(ng)Zeaxanthin
(ng)Pr
Macular homogenate 80.1 111.1 5Low-speed pellet 2.5 2.9High-speed supernatant 28.9 44.1 1CHAPS supernatant 23.6 31.1 1CHAPS pellet 0.4 0.6Uno-S peak 20.1 29.2Gel-®ltration peak 3.6 2.5
See Fig. 1 for XBP puri®cation protocol. Macular punches were 5 mmintermediate steps for protein and carotenoid analysis. N.D. � not determi
Carotenoid-binding Assays
Carotenoid-binding assays were performed by amodi®cation of the method used for the b-carotene-binding protein derived from ferret liver, a procedurethat used multiple organic solvent extractions andgel-®ltration chromatography to remove unboundcarotenoids (Rao et al., 1997). Two ml of carotenoiddissolved in tetrahydrofuran (THF), the preferredorganic solvent for delivery of carotenoids to aqueousenvironments (Cooney et al., 1993), was added to200 ml of an XBP preparation (typically 250±500 ngof protein) containing 8 mM CHAPS, brie¯y mixed,and then incubated overnight in the dark at 48C.Kinetic binding studies with lutein and zeaxanthindemonstrated that equilibrium binding was reachedwithin 2±4 hr (data not shown). Unbound carotenoidwas removed by four to six cycles of extraction with200 ml of hexane, followed by centrifugation at10 000 rpm for 15 sec in a microcentrifuge andremoval of the organic layer. Residual hexane wasremoved from the extracted sample under a stream ofargon or under vacuum, and the sample was thenanalysed by gel-®ltration chromatography asdescribed above. The peak A460/A280 ratio wasrecorded as a measure of speci®cally bound carotenoid(460 nm absorbance) relative to protein (280 nmabsorbance). The validity of this spectroscopic methodwas con®rmed by protein assays and HPLC analyses ofextracted carotenoids from collected peak fractions inseveral experiments (see Table I). Computerized non-linear regression analyses of binding parameters usingone- and two-site models were performed using the
A. Y. YEMELYANOV ET AL.
Software, San Diego, CA, U.S.A.).
3. Results
XBP Puri®cation Summary
Table I summarizes the protein and carotenoidcontent of a typical partial puri®cation of XBP from
seven human maculae through the ®rst gel-®ltrationstep using the protocol outlined in Fig. 1. The startingI
m seven human maculae
otein(mg) L/Z ratio
(L � Z)/protein(ng mgÿ1) A460/A280
100 0.72 0.037 N.D.
N.D. 0.86 N.D. N.D.
800 0.65 0.040 N.D.
700 0.76 0.032 N.D.
N.D. 0.67 N.D. N.D.
330 0.69 0.16 0.3323 1.44 0.27 0.49
in diameter. Approximately 10 % of the sample was used at thened. L � lutein. Z � zeaxanthin.

unlikely to be the only ocular protein that interacts
anion-exchange column was seen, but the protein
XANTHOPHYLL BINDING TO RETINAL PROTEINS
homogenate contains an average of 27 ng of totalxanthophylls per macula. The low-speed pellet, whichwould be expected to contain nuclei, mitochondria,large cellular fragments and other debris, has onlyminimal amounts of lutein and zeaxanthin associatedwith it. A substantial portion of the endogenousmacular carotenoids is associated with the high-speedsupernatant. This soluble cytosolic fraction has beenthe focus of previously published work on retinalcarotenoid-binding proteins from our laboratorywhich showed that a portion of the carotenoids inthis fraction is associated with tubulin (Bernsteinet al., 1997). After high-speed centrifugation, a visiblyyellow pellet remained. This yellow pellet could besolubilized by brief sonication into CHAPS, a zwitter-ionic detergent known to effectively solubilize manymembrane-associated proteins with minimaldenaturation (Hjelmeland and Chrambach, 1984).After multiple cycles of high-speed centrifugation ofthe CHAPS solubilized material, a small white pelletvirtually devoid of carotenoids remained, while thesupernatant was faint yellow. It is readily apparentthat approximately 25 % of the starting macularcarotenoids are associated with the CHAPS super-natant and that the lutein to zeaxanthin (L/Z) ratio issimilar to the starting homogenate. This fraction isthe focus of all further studies described here. Overall
FIG. 1. XBP puri®cation protocol.
puri®cation as measured by carotenoid to proteinratios was approximately seven-fold, but this should
be considered a minimum estimate because some ofthe carotenoids in the crude homogenate may not bebound to protein, some carotenoids initially bound toXBP may bind nonspeci®cally to surfaces and columnmatrices during the puri®cation process, and XBP is
385
with the macular carotenoids.
Ion-exchange Chromatography of XBP
We explored the use of ion-exchange chromatog-raphy as a ®rst step of puri®cation since it is wellsuited for use on relatively crude homogenates. Wefound it critical to maintain the presence of adetergent (CHAPS) throughout the chromatographicprocess to prevent excessive loss of protein andcarotenoids. At pH 5.5 on an UNO-S cation-exchangecolumn under low salt conditions, macular caroten-oids and their associated proteins do not bind to thecolumn, while many other macular proteins exhibitedstrong binding to the column. Fig. 2(A) shows thephotodiode-array chromatogram of an UNO-S puri®-cation. The photodiode-array spectrum of the peak ofunbound protein at 3 min from the cation-exchangecolumn demonstrates a typical carotenoid patternaround 460 nm as well as some absorbance at410 nm, a wavelength characteristic of porphyrin-binding proteins such as hemoglobin and some cyto-chromes. The fractions eluted with the salt gradientcontain other apparent prophyrin-binding proteinswith high 410 nm absorbance at 28 min, but there isno spectroscopic evidence for the presence of caroten-oids. HPLC analysis of the fractions con®rmed thatphotodiode-array monitoring for characteristiccarotenoid absorption is a sensitive nondestructivemeans to detect elution of the macular carotenoids[Fig. 2(C)]. At this point in the puri®cation, the L/Zratio is still similar to the starting homogenate. Atlater steps in the puri®cation process, we alsoemployed an anion-exchange step using a Bio-RadUNO-Q column with a 20 mM Tris buffer (pH 7.8).Chromatographic behavior was generally similar toUNO-S in that the carotenoid-containing fractions didnot bind to the ion-exchange column, indicating thatXBP probably has a high degree of hydrophobicity, acommon characteristic of many membrane-associatedproteins (van Renswoude and Kempf, 1984). At highpH conditions (49.0), some binding of XBP to the
became quite unstable.
Gel-®ltration Chromatography of XBP
The peak carotenoid-containing fractions from thecation-exchange step were combined and furtherpuri®ed by gel-®ltration chromatography. Macularcarotenoids eluted between the void volume and the
included volume [Fig. 2(B)]. Control experiments withlutein or zeaxanthin incubated without protein in
FIG. 2. Chromatographic puri®cation of XBP from ten human maculae. (A) Photodiode-array chromatogram of the cation-exchange puri®cation of a CHAPS solubilized human macular homogenate on UNO-S. The dashed line denotes a gradient from0 to 1 M NaCl. (B) Gel-®ltration chromatogram of peak carotenoid-containing fractions from the UNO-S step on a silica-basedcolumn. See the Materials and Methods section for detailed chromatographic conditions. The void volume was at 10 min, theincluded volume was at 22 min, and the peak of 460 nm absorbance at 18 min eluted with an apparent molecular weight ofapproximately 30 kDa according to gel-®ltration calibration standards. Insets in (A) and (B) are the photodiode-array visibleabsorption spectra of the peak carotenoid-containing fractions for each run. In (C) and (D), lutein (± ± ±) and zeaxanthin (ÐÐ)content was measured by HPLC on individual fractions with signi®cant carotenoid absorbance from the experiments shown in(A) and (B). Every ®fth fraction was analysed in the regions without signi®cant carotenoid absorbance. (C) HPLC analysis of theUNO-S step (A), and (D) HPLC analysis of the gel-®ltration step (B). The fractions with highest total amounts of lutein andzeaxanthin coincided exactly with the peak absorbance at 460 nm in the chromatograms. Insets in (C) and (D) are HPLCchromatograms of the peak fraction in each experiment, demonstrating the relative amounts of lutein and zeaxanthin present.
FIG. 3. Visible photodiode-array absorption spectrum ofpuri®ed human macular XBP (ÐÐ) in gel-®ltration elutionbuffer compared to the photodiode-array absorption spectraof lutein (± ± ±) and zeaxanthin (. . . . .) in 20 % dichloro-methane in hexane. The l for each spectrum is shown.
386 A. Y. YEMELYANOV ET AL.
CHAPS buffer demonstrated that these carotenoidsare poorly soluble in this detergent. The carotenoid/CHAPS micelles that do form, as expected, elute in theincluded volume at the end of the chromatogram,since CHAPS micelles typically have a molecularweight of approximately 6 kDa (Hjelmeland andChrambach, 1984).
The photodiode-array spectrum of the peakcarotenoid-containing fraction clearly shows acharacteristic carotenoid pro®le with minimalporphyrin contamination at 410 nm. A bathochro-mic shift of approximately 10 nm and ¯attening of thespectral ®ne structure is seen when XBPs absorptionspectrum is compared to the spectra of lutein andzeaxanthin dissolved in organic solvents (Fig. 3).
HPLC analysis of the peak fractions after gel-®ltration con®rmed the presence of lutein andzeaxanthin, but at this point the ratio of lutein tozeaxanthin was inverted [Fig. 2(D) and Table I]. No
L � lutein. Z � zeaxanthin.
other carotenoids such as b-carotene or lycopene weredetected. The splitting of the carotenoid peak in
maxAll spectra were normalized so that the absorbance at eachpeak maximum was 1.0.
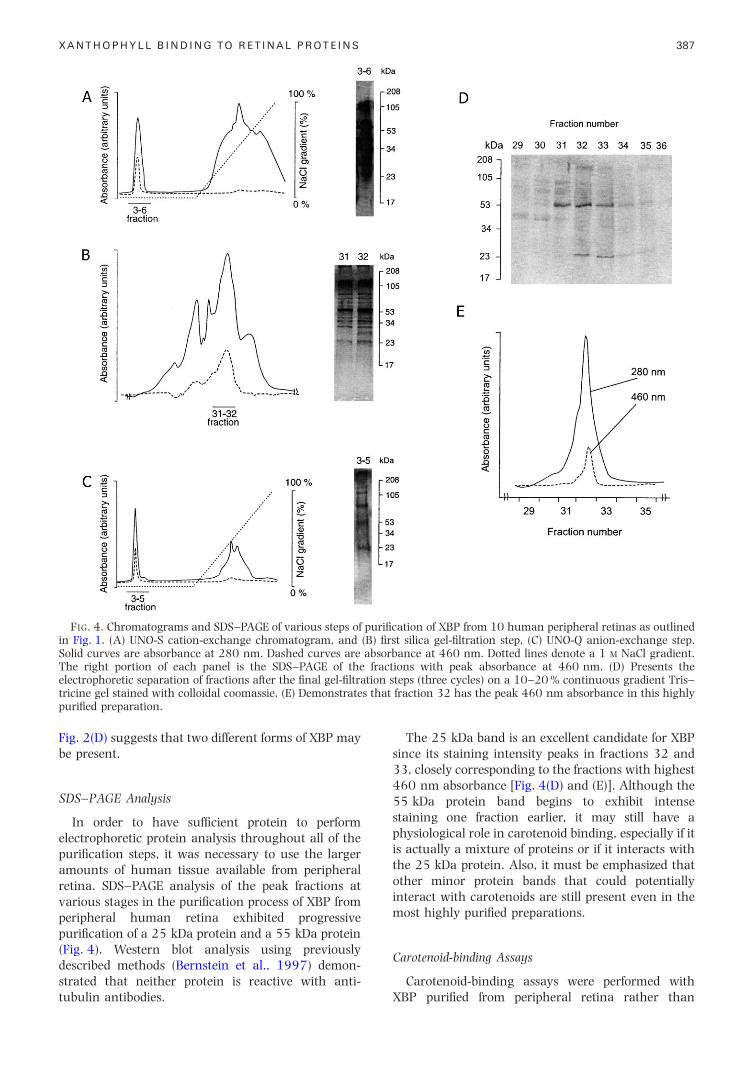
FIG. 4. Chromatograms and SDS±PAGE of various steps of puri®cation of XBP from 10 human peripheral retinas as outlinedin Fig. 1. (A) UNO-S cation-exchange chromatogram, and (B) ®rst silica gel-®ltration step, (C) UNO-Q anion-exchange step.Solid curves are absorbance at 280 nm. Dashed curves are absorbance at 460 nm. Dotted lines denote a 1 M NaCl gradient.The right portion of each panel is the SDS±PAGE of the fractions with peak absorbance at 460 nm. (D) Presents theelectrophoretic separation of fractions after the ®nal gel-®ltration steps (three cycles) on a 10±20 % continuous gradient Tris±
that
XANTHOPHYLL BINDING TO RETINAL PROTEINS 387
tricine gel stained with colloidal coomassie. (E) Demonstratespuri®ed preparation.
Fig. 2(D) suggests that two different forms of XBP may
interact with carotenoids are still present even in the
be present.
SDS±PAGE Analysis
In order to have suf®cient protein to performelectrophoretic protein analysis throughout all of thepuri®cation steps, it was necessary to use the largeramounts of human tissue available from peripheralretina. SDS±PAGE analysis of the peak fractions atvarious stages in the puri®cation process of XBP fromperipheral human retina exhibited progressivepuri®cation of a 25 kDa protein and a 55 kDa protein(Fig. 4). Western blot analysis using previouslydescribed methods (Bernstein et al., 1997) demon-
strated that neither protein is reactive with anti-tubulin antibodies.The 25 kDa band is an excellent candidate for XBPsince its staining intensity peaks in fractions 32 and33, closely corresponding to the fractions with highest460 nm absorbance [Fig. 4(D) and (E)]. Although the55 kDa protein band begins to exhibit intensestaining one fraction earlier, it may still have aphysiological role in carotenoid binding, especially if itis actually a mixture of proteins or if it interacts withthe 25 kDa protein. Also, it must be emphasized thatother minor protein bands that could potentially
fraction 32 has the peak 460 nm absorbance in this highly
most highly puri®ed preparations.
Carotenoid-binding Assays
Carotenoid-binding assays were performed withXBP puri®ed from peripheral retina rather than
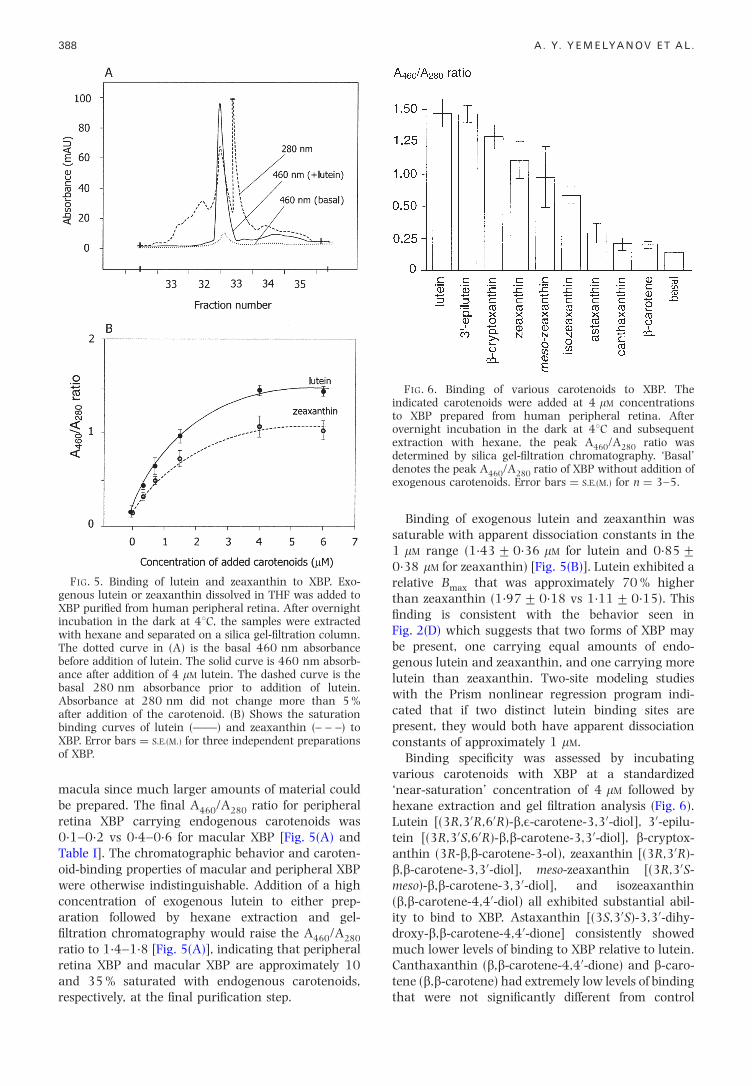
FIG. 5. Binding of lutein and zeaxanthin to XBP. Exo-genous lutein or zeaxanthin dissolved in THF was added toXBP puri®ed from human peripheral retina. After overnightincubation in the dark at 48C, the samples were extractedwith hexane and separated on a silica gel-®ltration column.The dotted curve in (A) is the basal 460 nm absorbancebefore addition of lutein. The solid curve is 460 nm absorb-ance after addition of 4 mM lutein. The dashed curve is thebasal 280 nm absorbance prior to addition of lutein.Absorbance at 280 nm did not change more than 5 %after addition of the carotenoid. (B) Shows the saturationbinding curves of lutein (ÐÐ) and zeaxanthin (± ± ±) to
FIG. 6. Binding of various carotenoids to XBP. Theindicated carotenoids were added at 4 mM concentrationsto XBP prepared from human peripheral retina. Afterovernight incubation in the dark at 48C and subsequentextraction with hexane, the peak A460/A280 ratio wasdetermined by silica gel-®ltration chromatography. `Basal'denotes the peak A /A ratio of XBP without addition of
388 A. Y. YEMELYANOV ET AL.
macula since much larger amounts of material couldbe prepared. The ®nal A460/A280 ratio for peripheralretina XBP carrying endogenous carotenoids was0.1±0.2 vs 0.4±0.6 for macular XBP [Fig. 5(A) andTable I]. The chromatographic behavior and caroten-oid-binding properties of macular and peripheral XBPwere otherwise indistinguishable. Addition of a highconcentration of exogenous lutein to either prep-aration followed by hexane extraction and gel-®ltration chromatography would raise the A460/A280
ratio to 1.4±1.8 [Fig. 5(A)], indicating that peripheralretina XBP and macular XBP are approximately 10
XBP. Error bars � S.E.(M.) for three independent preparationsof XBP.
and 35 % saturated with endogenous carotenoids,respectively, at the ®nal puri®cation step.
Binding of exogenous lutein and zeaxanthin wassaturable with apparent dissociation constants in the1 mM range (1.43+0.36 mM for lutein and 0.85+0.38 mM for zeaxanthin) [Fig. 5(B)]. Lutein exhibited arelative Bmax that was approximately 70 % higherthan zeaxanthin (1.97+0.18 vs 1.11+0.15). This®nding is consistent with the behavior seen inFig. 2(D) which suggests that two forms of XBP maybe present, one carrying equal amounts of endo-genous lutein and zeaxanthin, and one carrying morelutein than zeaxanthin. Two-site modeling studieswith the Prism nonlinear regression program indi-cated that if two distinct lutein binding sites arepresent, they would both have apparent dissociationconstants of approximately 1 mM.
Binding speci®city was assessed by incubatingvarious carotenoids with XBP at a standardized`near-saturation' concentration of 4 mM followed byhexane extraction and gel ®ltration analysis (Fig. 6).Lutein [(3R,30R,60R)-b,E-carotene-3,30-diol], 30-epilu-tein [(3R,30S,60R)-b,b-carotene-3,30-diol], b-cryptox-anthin (3R-b,b-carotene-3-ol), zeaxanthin [(3R,30R)-b,b-carotene-3,30-diol], meso-zeaxanthin [(3R,30S-meso)-b,b-carotene-3,30-diol], and isozeaxanthin(b,b-carotene-4,40-diol) all exhibited substantial abil-ity to bind to XBP. Astaxanthin [(3S,30S)-3,30-dihy-droxy-b,b-carotene-4,40-dione] consistently showedmuch lower levels of binding to XBP relative to lutein.Canthaxanthin (b,b-carotene-4,40-dione) and b-caro-
460 280exogenous carotenoids. Error bars � S.E.(M.) for n � 3±5.
tene (b,b-carotene) had extremely low levels of bindingthat were not signi®cantly different from control
comparable membrane-associated xanthophyll-bind-
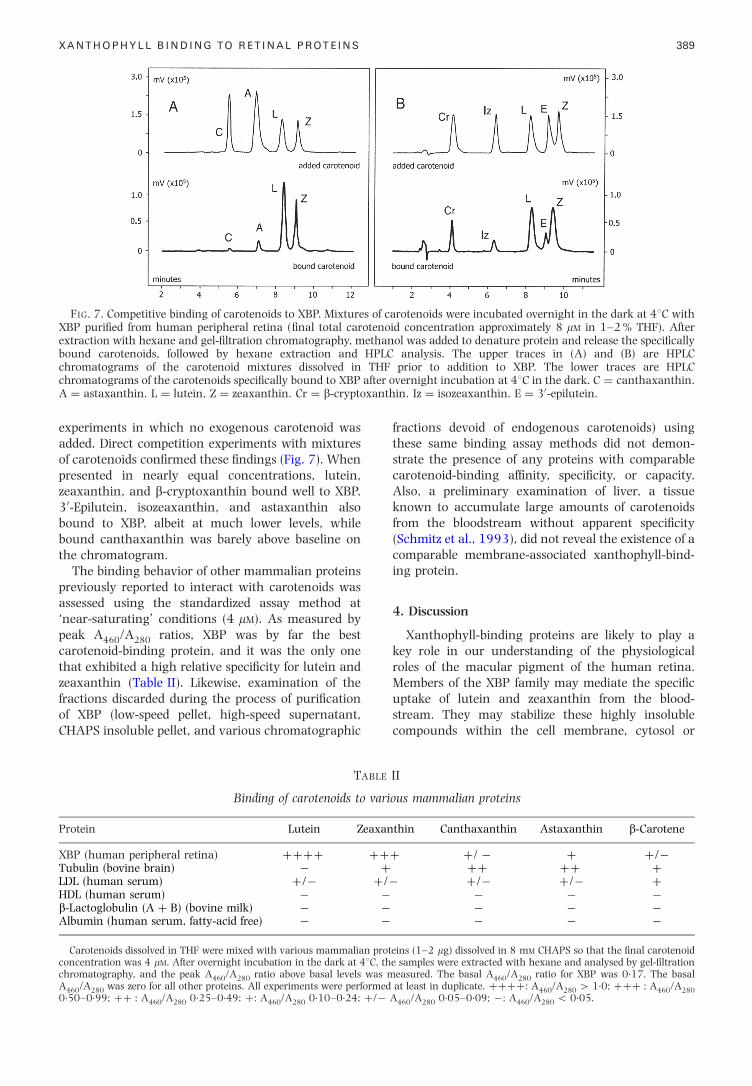
FIG. 7. Competitive binding of carotenoids to XBP. Mixtures of carotenoids were incubated overnight in the dark at 48C withXBP puri®ed from human peripheral retina (®nal total carotenoid concentration approximately 8 mM in 1±2 % THF). Afterextraction with hexane and gel-®ltration chromatography, methanol was added to denature protein and release the speci®callybound carotenoids, followed by hexane extraction and HPLC analysis. The upper traces in (A) and (B) are HPLCchromatograms of the carotenoid mixtures dissolved in THF prior to addition to XBP. The lower traces are HPLC
fterxant
XANTHOPHYLL BINDING TO RETINAL PROTEINS 389
experiments in which no exogenous carotenoid wasadded. Direct competition experiments with mixturesof carotenoids con®rmed these ®ndings (Fig. 7). Whenpresented in nearly equal concentrations, lutein,zeaxanthin, and b-cryptoxanthin bound well to XBP.30-Epilutein, isozeaxanthin, and astaxanthin alsobound to XBP, albeit at much lower levels, whilebound canthaxanthin was barely above baseline onthe chromatogram.
The binding behavior of other mammalian proteinspreviously reported to interact with carotenoids wasassessed using the standardized assay method at`near-saturating' conditions (4 mM). As measured bypeak A460/A280 ratios, XBP was by far the bestcarotenoid-binding protein, and it was the only onethat exhibited a high relative speci®city for lutein andzeaxanthin (Table II). Likewise, examination of thefractions discarded during the process of puri®cation
chromatograms of the carotenoids speci®cally bound to XBP aA � astaxanthin. L � lutein. Z � zeaxanthin. Cr � b-crypto
of XBP (low-speed pellet, high-speed supernatant,CHAPS insoluble pellet, and various chromatographic
TABLE
Binding of carotenoids to vari
Protein Lutein Zeaxan
XBP (human peripheral retina) ���� ��Tubulin (bovine brain) ÿ �LDL (human serum) �/ÿ �/HDL (human serum) ÿ ÿb-Lactoglobulin (A � B) (bovine milk) ÿ ÿAlbumin (human serum, fatty-acid free) ÿ ÿ
Carotenoids dissolved in THF were mixed with various mammalian protconcentration was 4 mM. After overnight incubation in the dark at 48C, thchromatography, and the peak A460/A280 ratio above basal levels was mA460/A280 was zero for all other proteins. All experiments were performed0.50±0.99; �� : A460/A280 0.25±0.49; �: A460/A280 0.10±0.24; �/ÿ
fractions devoid of endogenous carotenoids) usingthese same binding assay methods did not demon-strate the presence of any proteins with comparablecarotenoid-binding af®nity, speci®city, or capacity.Also, a preliminary examination of liver, a tissueknown to accumulate large amounts of carotenoidsfrom the bloodstream without apparent speci®city(Schmitz et al., 1993), did not reveal the existence of a
overnight incubation at 48C in the dark. C � canthaxanthin.hin. Iz � isozeaxanthin. E � 30-epilutein.
ing protein.
4. Discussion
Xanthophyll-binding proteins are likely to play akey role in our understanding of the physiologicalroles of the macular pigment of the human retina.Members of the XBP family may mediate the speci®cuptake of lutein and zeaxanthin from the blood-
stream. They may stabilize these highly insolublecompounds within the cell membrane, cytosol orII
ous mammalian proteins
thin Canthaxanthin Astaxanthin b-Carotene
� �/ ÿ � �/ÿ�� �� �
ÿ �/ÿ �/ÿ �ÿ ÿ ÿÿ ÿ ÿÿ ÿ ÿ
eins (1±2 mg) dissolved in 8 mM CHAPS so that the ®nal carotenoide samples were extracted with hexane and analysed by gel-®ltration
easured. The basal A460/A280 ratio for XBP was 0.17. The basalat least in duplicate. ����: A460/A280 4 1.0; ��� : A460/A280
A460/A280 0.05±0.09; ÿ: A460/A280 5 0.05.

cytoskeleton. They may act as enzymes mediating theinterconversion of lutein, zeaxanthin and theirvarious metabolites within the retina. They may alsofacilitate the antioxidant action of these xanthophyllcarotenoids.
In this investigation, we have taken advantage ofthe high endogenous concentration of lutein andzeaxanthin in the human macula. The quantity ofthese carotenoids is suf®cient to obviate the need toadd exogenous carotenoids during the puri®cationprocess, thus lessening the likelihood of generatingnonphysiological binding interactions. The extremelyhigh visible absorbance properties of the macularcarotenoids in conjunction with sensitive in-linephotodiode-array detection allows for convenientnondestructive monitoring of chromatography eluateson homogenates originating from as few as twomaculae.
As shown in Table I and Fig. 1, the macularcarotenoids are more or less evenly divided betweenthe soluble cytosolic fraction (high-speed supernatant)and the membrane-associated fraction (CHAPS super-natant). We have previously investigated the solublefraction from human and bovine retinas and foundthat a substantial portion of the retinal carotenoids areapparently associated with tubulin (Bernstein et al.,1997). Although it has been hypothesized that at leasta portion of the macular carotenoids may be freelydissolved in lipid bilayers (Bone et al., 1992; Sujaket al., 1999), simple membrane lipid solubilization ofthe macular carotenoids would be unlikely to be able toaccount for the high degree of speci®city of lutein andzeaxanthin uptake into the primate retina. It is notunreasonable to assume that human macular mem-branes could be a rich source of speci®c bindingproteins for the macular carotenoids, especially sincemany plant and invertebrate carotenoid-bindingproteins are known to be membrane-associated(Lakshman and Okoh, 1993; Reddy et al., 1993).
Ion-exchange chromatography in the presence ofthe zwitterionic detergent CHAPS proved to be anexcellent high-capacity ®rst step of puri®cation. Whenwe applied a detergent-solubilized extract to an UNO-Scation-exchange column at pH 5.5, we found that theprotein-associated carotenoids eluted with theunbound fractions and that almost no detectablecarotenoids were associated with the bound proteinseluted by the salt gradient [Fig. 2(A) and (C)].
Gel-®ltration chromatography on the peakcarotenoid-containing fractions after ion-exchangechromatography demonstrated an XBP peak whicheluted between the void volume and the includedvolume [Fig. 2(B) and (D)]. The photodiode-arrayspectrum is clearly characteristic of a carotenoid andclosely matches the known spectrum of the humanmacular pigment (Fig. 3) (Snodderly et al., 1984b;Bone et al., 1992). Its modest bathochromic spectral
390
shift relative to carotenoids in organic solventsindicates that XBP would be best classi®ed as a
`carotenolipoprotein' whose chromophore is boundloosely in a hydrophobic environment (Gomez andMilicua, 1992). By contrast, carotenoid-bindingproteins with rigid chromophore binding pocketssuch as a-crustacyanin usually have much largerbathochromic shifts, while those with multipleaggregated chromophores such as crustochrin typi-cally have hypsochromic shifts (Zagalsky, 1995).
HPLC analysis of the eluted fractions showed thatboth lutein and zeaxanthin were present in the XBPpeak with an L/Z ratio of 1.44 vs a ratio ofapproximately 0.70 in the previous steps of thepuri®cation (Table I and Fig. 2). SDS±PAGE withcolloidal coomassie staining of the puri®ed XBPfractions derived from human peripheral retinademonstrated a relatively small number of mediummolecular-weight proteins (Fig. 4). Comparison of theelution pattern of these proteins to the photodiode-array elution pro®le of the carotenoids from the gel-®ltration column indicates that the most likelymolecular weight for XBP is 25 kDa, although the55 kDa protein band cannot yet be ruled out. It is alsopossible that these two proteins may somehowinteract to form functional XBP or that a minorcomponent protein band is actually responsible for thespeci®c binding of lutein and zeaxanthin. Furtherpuri®cation procedures on much larger amounts ofhuman retinal tissue will be required to settle theissue de®nitively. In the meantime, since ligand-binding studies can be readily carried out on partiallypuri®ed preparations of detergent-solubilized mem-brane proteins (Bellis, Kass-Simon and Rhoads, 1992;Ward, Kopf and Storey, 1994), we proceeded withthat line of investigation in order to begin thebiochemical characterization of ocular XBP.
The hydrophobic nature of carotenoids makes themsusceptible to artifactual nonspeci®c binding inter-actions during biochemical binding assays. Thus, wecarried out our binding assays using stringentconditions with exhaustive hexane extractions andhigh-resolution gel-®ltration analysis to removeunbound and non-speci®cally bound carotenoids.Binding of lutein and zeaxanthin to XBP preparedfrom human peripheral retina was saturable withapparent dissociation constants in the 1 mM range(Fig. 5), midway between the 56 nM and 32 mM
apparent dissociation constants reported for the ferretliver b-carotene-binding protein (Rao et al., 1997).The actual binding af®nity of XBP for xanthophylls islikely to be much tighter than the apparent valuesreported here due to the high hydrophobicity ofcarotenoids. Unbound carotenoids in water/THFsolutions are in a highly ordered (aggregated) state(Cooney et al., 1993), and therefore the `free' concen-tration of carotenoid is substantially lower than thecalculated value. Very tight association of XBP withlutein and zeaxanthin is consistent with the great
A. Y. YEMELYANOV ET AL.
stability of the complex during organic solventextractions and chromatographic separations.

proteins and their possible roles in the pathogenesis
technical assistance of Jill Moore, Garth Muir, and Keith
The number of available binding sites was 70 %higher for lutein relative to zeaxanthin, a ®ndingconsistent with the elution pattern of endogenouscarotenoids with puri®ed XBP which indicated thepossible presence of two different forms of XBP, onecarrying equal amounts of endogenous luteinand zeaxanthin and one carrying more lutein thanzeaxanthin [Fig. 2(D)].
Our binding speci®city and binding competitionstudies showed that a number of mono- anddihydroxycarotenoids bind well to XBP under stan-dardized conditions (Figs 6 and 7). On the other hand,the hydrocarbon carotenoid b-carotene and thediketocarotenoid canthaxanthin do not bind to anysigni®cant extent. Astaxanthin, a diketodihydroxy-carotenoid, has never been detected in human retina,and it binds relatively poorly, suggesting that thepresence of a keto group may be detrimental tobinding to XBP. The ®nding that the monohydroxy-carotenoid b-cryptoxanthin binds to XBP was some-what unexpected since it, like b-carotene, is found inno more than trace amounts in the human retinadespite relatively high serum concentrations, and wespeculate that the human retina might containenzymatic machinery to convert cryptoxanthins toeither lutein, zeaxanthin, or vitamin A. The fact thatisozeaxanthin binds to XBP much better thancanthaxanthin is quite interesting. In a monkeymodel of canthaxanthin toxicity, this carotenoid isreductively metabolized and deposited as 4-hydroxy-echninone and isozeaxanthin in the central macula(Goralczyk et al., 2000), both of which presumablycould be bound to XBP. In the more peripheral areasof the retina, canthaxanthin predominates where itforms large crystals unlikely to be associated with anyspeci®c binding protein.
Comparison of XBP to other potential mammaliancarotenoid-binding proteins using a standardizedbinding assay demonstrates that it is the only one toexhibit an A460/A280 ratio greater than 0.5 and theonly one to have speci®city for lutein and zeaxanthinrelative to the other three carotenoids tested(Table II). Its maximal A460/A280 ratio of approxi-mately 1.8 for highly puri®ed XBP with exogenouslutein bound to it compares favorably with the 2.5A460/A280 ratio reported for the lutein-binding proteinisolated from silkworm midgut (Jouni and Wells,1996) and the 3.3 A630/A280 ratio reported fora-crustacyanin (Keen et al., 1991a). It is interestingto note that well-established mammalian carotenoidexchange proteins such as HDL and LDL (Clevidenceand Bieri, 1993) fared poorly in this binding assay,suggesting that the associated carotenoids are veryloosely bound to these lipoprotein complexes. Tubulincould bind some of the presented carotenoids at low tomoderate levels, but its speci®city differed markedlyfrom XBP. This ®nding is consistent with previously
XANTHOPHYLL BINDING TO RETINAL PROTEINS
published carotenoid photoaf®nity labeling studies inwhich we hypothesized that polymerized tubulin is a
site for non-speci®c passive deposition of carotenoidsin the human retina (Bernstein et al., 1997).
Isolation and characterization of XBP has been anextraordinarily challenging task due to the verylimited availability of human donor eyes. Membrane-associated proteins are notoriously dif®cult to purify tohomogeneity, and puri®cation and detection pro-cedures had to be optimized for small quantities ofprotein throughout the entire protocol. With thepartial puri®cation of a membrane-associated XBPfrom human macula and retina that binds lutein andzeaxanthin saturably and speci®cally, there clearly is aconsiderable amount of further characterization to beperformed. These include de®nitive identi®cation andpuri®cation to homogeneity, additional binding af®-nity and speci®city studies, tissue speci®city andlocalization, and eventual cloning and expression.Nevertheless, this has been an important ®rst step inhelping to de®ne the properties and functions of apotentially important ocular carotenoid-bindingprotein. The physiological functions of XBP and related
391
and treatment of AMD remain to be explored.
Acknowledgements
This work was supported by National Institutes of HealthGrant EY-11600 and by grants from Kemin Foods (DesMoines, IA, U.S.A.) and from Research to Prevent Blindness,Inc. (New York, NY, U.S.A.). The authors thank the UtahLions Eye Bank for supplying postmortem human eyes. The
Neaman is gratefully acknowledged.
References
Balashova, O. A., Balashov, N. A. and Bernstein, P. S.(1998). Lutein and zeaxanthin exhibit photoprotectiveand anti-apoptotic activity in vitro. [ARVO Abstract].Inv. Ophthalmol. Vis. Sci. 39, S129 (Abstract no. 603).
Bellis, S. L., Kass-Simon, G. and Rhoads, D. E. (1992).Partial characterization and detergent solubilization ofthe putative glutathione chemoreceptor from hydra.Biochemistry 31, 9838±43.
Bernstein, P. S., Balashov, N. A., Tsong, E. D. and Rando,R. R. (1997). Retinal tubulin binds macular caro-tenoids. Inv. Ophthalmol. Vis. Sci. 38, 167±75.
Bone, R. A., Landrum, J. T. and Cains, A. (1992). Opticaldensity spectra of the macular pigment in vivo andin vitro. Vision Res. 32, 105±10.
Bone, R. A., Landrum, J. T., Fernandez, L. and Tarsis, S. L.(1988). Analysis of the macular pigment by HPLC:retinal distribution and age study. Inv. Ophthalmol. Vis.Sci. 29, 843±9.
Bone, R. A., Landrum, J. T., Hime, G. W. and Cains, A.(1993). Stereochemistry of the human macularcarotenoids. Inv. Ophthalmol. Vis. Sci. 34, 2033±40.
Bone, R. A., Landrum, J. T. and Tarsis, S. L. (1985).Preliminary identi®cation of the human macularpigment. Vision Res. 25, 1531±5.
Britton, G. (1995). UV/visible spectroscopy. In Carotenoids:Spectroscopy, Vol. 1B. (Britton, G., Liaaen-Jensen, S. and
Pfander, H., Eds.) Pp. 13±62. Birkhauser-Verlag: Basel,Switzerland.
Clevidence, B. A. and Bieri, J. G. (1993). Association ofcarotenoids with human plasma lipoproteins. MethodsEnzymol. 214, 33±46.
Cooney, R. V., Kappock, T. J., Pung, A. and Bertram, J. S.(1993). Solubilization, cellular uptake, and activity ofb-carotene and other carotenoids as inhibitors ofneoplastic transformation in cultured cells. MethodsEnzymol. 214, 55±68.
Dorey, C. K., Toyoda, Y., Thomson, L., Garnett, K. M.,Sapunzatkis, M., Craft, N., Nichols, C. and Cheng, K.(1997). Light induced photoreceptor apoptosis iscorrelated with dietary and retinal levels of 3R,30R-zeaxanthin. [ARVO Abstract]. Inv. Ophthalmol. Vis. Sci.38, S355 (Abstract no. 1656).
Dufour, E. and Haertle, T. (1991). Binding of retinoids andb-carotene to b-lactoglobulin: in¯uence of protein modi-®cations. Biochim. Biophys. Acta 1079, 316±20.
Eye Disease Case Control Study Group. (1993). Antioxidantstatus and age-related macular degeneration. Arch.Ophthalmol. 111, 104±9.
Gass, J. D. M. (1999). MuÈ ller cell cone, an overlooked part ofthe anatomy of the fovea centralis. Arch. Ophthalmol.117, 821±3.
Gomez, R. and Milicua, J. C. G. (1992). Use of borohydridereduction methods to probe carotenoid-protein binding.Methods Enzymol. 213, 100±10.
Goralczyk, R., Barker, F. M., Buser, S., Leichi, H. and Bausch,J. (2000). Dose dependency of canthaxanthin crystals inmonkey retina and spatial distribution of its metabolites.Inv. Ophthalmol. Vis. Sci. 41, 1513±22.
Hammond, B. R., Johnson, E. J., Russell, R. M., Krinsky, N. I.,Yeum, K.-J., Edwards, R. B. and Snodderly, D. M.(1997). Dietary modi®cation of human macular pig-ment density. Inv. Ophthalmol. Vis. Sci. 38, 1795±801.
Handelman, G. J., Snodderly, D. M., Adler, A. J., Russett,M. D. and Dratz, E. A. (1992). Measurement of caro-tenoids in human and monkey retinas. MethodsEnzymol. 213, 220±30.
Hemmi, H., Hata, M. and Takeuchi, M. (1991). Studies onthe carotenoids in the muscle of the salmon: V: com-bination of astaxanthin and canthaxanthin with bovineserum albumin and egg albumin. Comp. Biochem.Physiol. 99B, 609±12.
Hjelmeland, L. M. and Chrambach, A. (1984). Solubilizationof functional membrane proteins. Methods Enzymol.104, 305±18.
Jouni, Z. E. and Wells, M. A. (1996). Puri®cation and partialcharacterization of lutein-binding protein from themidgut of the silkworm Bombyx mori. J. Biol. Chem.271, 14722±6.
Keen, J. N., Caceres, I., Eliopoulos, E. E., Zagalsky, P. F. andFindlay, J. B. C. (1991a). Complete sequence and modelfor the C1 subunit of the carotenoprotein, crustacyanin,and model for the dimer, b-crustacyanin, formedbetween the C1 and A2 subunits with astaxanthin.Eur. J. Biochem. 202, 31±40.
Keen, J. N., Caceres, I., Eliopoulos, E. E., Zagalsky, P. F. andFindlay, J. B. C. (1991b). Complete sequence and modelfor the A2 subunit of the carotenoid pigment complex,crustacyanin. Eur. J. Biochem. 197, 407±14.
Khachik, F., Bernstein, P. S. and Garland, D. L. (1997).Identi®cation of lutein and zeaxanthin oxidationproducts in human and monkey retina. Inv. Ophthalmol.Vis. Sci. 38, 1802±11.
Lakshman, M. R. and Okoh, C. (1993). Carotenoid-proteincomplexes. Methods Enzymol. 214, 74±86.
Landrum, J. T., Bone, R. A., Joa, H., Kilburn, M. D., Moore,
392
L. L. and Sprague, K. E. (1997a). A one year study ofthe macular pigment: the effect of 140 days of a luteinsupplement. Exp. Eye Res. 65, 57±62.
Landrum, J. T., Bone, R. A. and Kilburn, M. D. (1997b). Themacular pigment: a possible role in protection from age-related macular degeneration. Adv. Pharmacol. 38,537±56.
Landrum, J. T., Bone, R. A., Moore, L. L. and Gomez, C. M.(1999). Analysis of zeaxanthin distribution within indi-vidual human retinas. Methods Enzymol. 299, 457±67.
McFall-Ngai, M., Crescitelli, F., Childress, J. and Horwitz, J.(1986). Patterns of pigmentation in the eye lens of thedeep-sea hatchet®sh, Argyropelecus af®nis Garman. J.Comp. Physiol. A. 159, 791±800.
Rao, M. N., Ghosh, P. and Lakshman, M. R. (1997).Puri®cation and partial characterization of a cellularcarotenoid-binding protein from ferret liver. J. Biol.Chem. 272, 24450±60.
Rapp, L. M., Maple, S. S. and Choi, J. H. (2000). Lutein andzeaxanthin concentrations in rod outer segment mem-branes from perifoveal and peripheral human retina.Inv. Ophthalmol. Vis. Sci. 41, 1200±9.
Reddy, K. J., Bullerjahn, G. S. and Sherman, L. A. (1993).Characteristics of membrane-associated carotenoid-binding proteins in cyanobacteria and prochlorophytes.Methods Enzymol. 214, 390±401.
RoÈll, B. (2000). Carotenoid and retinoid-two pigments in agecko eye lens. Comp. Biochem. Physiol. 125A, 105±12.
Saari, J. C. (1994). Retinoids in photosensitive systems. InThe Retinoids: Biology, Chemistry, and Medicine, 2nd edn.(Sporn, M. B., Roberts, A. B. and Goodman, D. S., Eds.)Pp. 351±85. Raven Press: New York, U.S.A.
Schalch, W., Dayhaw-Barker, P. and Barker, F. M. (1999).The carotenoids of the human retina. In Nutritional andEnvironmental In¯uences on the Eye. (Taylor, A., Ed.)Pp. 215±50. CRC Press: Boca Raton, FL, U.S.A.
Schmitz, H. H., Poor, C. L., Gugger, E. T. and Erdman, J. W.(1993). Analysis of carotenoids in human and animaltissue. Methods Enzymol. 214, 102±16.
Seddon, J. M., Ajani, U. A., Sperduto, R. D., Hiller, R., Blair,N., Burton, T. C., Farber, M. D., Gragoudas, E. S., Haller,J., Miller, D. T., Yanuzzi, L. A. and Willett, W. (1994).Dietary carotenoids, vitamins A, C, E and advanced age-related macular degeneration: a multicenter study.JAMA 272, 1413±20.
Snodderly, D. M., Auran, J. D. and Delori, F. C. (1984a). Themacular pigment. II. Spatial distribution in primateretinas. Inv. Ophthalmol. Vis. Sci. 25, 674±85.
Snodderly, D. M., Brown, P. K., Delori, F. C. and Auran, J. D.(1984b). The macular pigment. I. Absorbance spectra,localization, and discrimination from other yellowpigments in primate retinas. Inv. Ophthalmol. Vis. Sci.25, 660±73.
Sommerberg, O. G., Siems, W. G., Hurst, J. S., Lewis, J. W.,Kliger, D. S. and van Kuijk, F. J. (1999). Lutein andzeaxanthin are associated with photoreceptors in thehuman retina. Curr. Eye Res. 19, 491±5.
Sujak, A., Gabrielska, J., Grudzinski, W., Bore, R., Mazurek,P. and Gruszecki, W. I. (1999). Lutein and zeaxanthinas protectors of lipid membranes against oxidativedamage: the structural aspects. Arch. Biochem. Biophys.371, 301±7.
van Renswoude, J. and Kempf, C. (1984). Puri®cation ofintegral membrane proteins. Methods Enzymol. 104,329±39.
Ward, C. R., Kopf, G. S. and Storey, B. T. (1994). Solubilizationand partial puri®cation from mouse sperm membranesof the speci®c binding activity for 3-quinuclidinylbenzilate, a potent inhibitor of the zona pellucida-inducedacrosome reaction. Mol. Reprod. Dev. 39, 423±32.
Zagalsky, P. F. (1995). Carotenoproteins. In Carotenoids:
A. Y. YEMELYANOV ET AL.
Isolation and Analysis, Vol. 1A. (Britton, G., Liaaen-Jensen, S. and Pfander, H., Eds.) Pp. 287±94.Birkhauser-Verlag: Basel, Switzerland.




















