
Life-Long Shedding of Puumala Hantavirus in Wild Bank Voles (Myodes glareolus)
34
Journal of General Virology Life-Long Shedding of Puumala Hantavirus in Wild Bank Voles (Myodes glareolus) --Manuscript Draft-- Manuscript Number: JGV-D-14-00201R1 Full Title: Life-Long Shedding of Puumala Hantavirus in Wild Bank Voles (Myodes glareolus) Short Title: Life-long hantavirus shedding by wild rodent hosts Article Type: Standard Section/Category: Animal - Negative-strand RNA Viruses Corresponding Author: Liina Voutilainen, Ph.D. University of Helsinki Helsinki, FINLAND First Author: Liina Voutilainen, Ph.D. Order of Authors: Liina Voutilainen, Ph.D. Tarja Sironen, Ph.D. Elina Tonteri, PhD Anne Tuiskunen Bäck, PhD Maria Razzauti, PhD Malin Karlsson Maria Wahlström Jukka Niemimaa, MSc Heikki Henttonen, PhD Åke Lundkvist, PhD Abstract: The knowledge of viral shedding patterns and viremia in the reservoir host species is a key factor in assessing the human risk of zoonotic viruses. The shedding of hantaviruses (family Bunyaviridae) by their host rodents has widely been studied experimentally, but rarely in natural settings. Here we present the dynamics of Puumala (PUUV) hantavirus shedding and viremia in naturally infected, wild bank voles (Myodes glareolus). In a monthly capture-mark-recapture study, we analyzed 18 bank voles for the presence and relative quantity of PUUV RNA in the excreta and blood from 2 months before up to 8 months after seroconversion. The proportion of animals shedding PUUV RNA in saliva, urine, and feces peaked during the first month after seroconversion, but continued throughout the study period with only a slight decline. The quantity of shed PUUV in RT-PCR positive excreta was constant over time. In blood, PUUV RNA was present for up to 7 months but both the probability of viremia and the virus load declined by time. Our findings contradict the current view of a decline in virus shedding after the acute phase and a short viremic period in hantavirus infection - an assumption widely adopted into current epidemiological models. We suggest the life-long shedding as a means of hantaviruses to survive over host population bottlenecks, and to disperse in fragmented habitats where local host and/or virus populations face temporary extinctions. Our results indicate that the kinetics of pathogens in wild hosts may considerably differ from those observed in laboratory settings. Powered by Editorial Manager® and ProduXion Manager® from Aries Systems Corporation JGV Papers in Press. Published February 20, 2015 as doi:10.1099/vir.0.000076
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Life-Long Shedding of Puumala Hantavirus in Wild Bank Voles (Myodes glareolus)
Journal of General Virology
Life-Long Shedding of Puumala Hantavirus in Wild Bank Voles (Myodes glareolus)--Manuscript Draft--
Manuscript Number: JGV-D-14-00201R1
Full Title: Life-Long Shedding of Puumala Hantavirus in Wild Bank Voles (Myodes glareolus)
Short Title: Life-long hantavirus shedding by wild rodent hosts
Article Type: Standard
Section/Category: Animal - Negative-strand RNA Viruses
Corresponding Author: Liina Voutilainen, Ph.D.University of HelsinkiHelsinki, FINLAND
First Author: Liina Voutilainen, Ph.D.
Order of Authors: Liina Voutilainen, Ph.D.
Tarja Sironen, Ph.D.
Elina Tonteri, PhD
Anne Tuiskunen Bäck, PhD
Maria Razzauti, PhD
Malin Karlsson
Maria Wahlström
Jukka Niemimaa, MSc
Heikki Henttonen, PhD
Åke Lundkvist, PhD
Abstract: The knowledge of viral shedding patterns and viremia in the reservoir host species is akey factor in assessing the human risk of zoonotic viruses. The shedding ofhantaviruses (family Bunyaviridae) by their host rodents has widely been studiedexperimentally, but rarely in natural settings. Here we present the dynamics ofPuumala (PUUV) hantavirus shedding and viremia in naturally infected, wild bankvoles (Myodes glareolus). In a monthly capture-mark-recapture study, we analyzed 18bank voles for the presence and relative quantity of PUUV RNA in the excreta andblood from 2 months before up to 8 months after seroconversion.The proportion of animals shedding PUUV RNA in saliva, urine, and feces peakedduring the first month after seroconversion, but continued throughout the study periodwith only a slight decline. The quantity of shed PUUV in RT-PCR positive excreta wasconstant over time. In blood, PUUV RNA was present for up to 7 months but both theprobability of viremia and the virus load declined by time.Our findings contradict the current view of a decline in virus shedding after the acutephase and a short viremic period in hantavirus infection - an assumption widelyadopted into current epidemiological models. We suggest the life-long shedding as ameans of hantaviruses to survive over host population bottlenecks, and to disperse infragmented habitats where local host and/or virus populations face temporaryextinctions. Our results indicate that the kinetics of pathogens in wild hosts mayconsiderably differ from those observed in laboratory settings.
Powered by Editorial Manager® and ProduXion Manager® from Aries Systems Corporation
JGV Papers in Press. Published February 20, 2015 as doi:10.1099/vir.0.000076

1
Life-Long Shedding of Puumala Hantavirus in Wild Bank Voles (Myodes glareolus) 1
2
3
Liina Voutilainen1,2#
, Tarja Sironen1,2
, Elina Tonteri2,3
, Anne Tuiskunen Bäck3,4
, Maria Razzauti1,2
, 4
Malin Karlsson4, Maria Wahlström
4, Jukka Niemimaa
1, Heikki Henttonen
1, Åke Lundkvist
3,4,5 5
6
1 Natural Resources Institute Finland, Vantaa, Finland 7
2University of Helsinki, Department of Virology, Helsinki, Finland 8
3Department of Microbiology, Tumor and Cell Biology, Karolinska Institutet, Stockholm, Sweden 9
4The Public Health Agency of Sweden, Solna, Sweden 10
5Department of Medical Biochemistry and Microbiology, Uppsala University, Uppsala, Sweden 11
#Corresponding author. Mailing address: Department of Virology, PO Box 21, FI-00014 University of 12
Helsinki, Finland. Phone: +358 9 1912 6548. Fax: +358 9 1912 6491. E-mail: 13
15
Running title: Life-long hantavirus shedding by wild rodent hosts 16
Content category: Animal Negative-strand RNA Viruses 17
Word count for summary 250 18
Word count for text: 5357 19
Number of tables: 2 20
Number of figures: 2 21
22
Manuscript Including References (Word document)Click here to download Manuscript Including References (Word document): Voutilainen et al PUUV shedding JGV revised.docx

2
Summary 23
The knowledge of viral shedding patterns and viremia in the reservoir host species is a key factor in 24
assessing the human risk of zoonotic viruses. The shedding of hantaviruses (family Bunyaviridae) by 25
their host rodents has widely been studied experimentally, but rarely in natural settings. Here we 26
present the dynamics of Puumala (PUUV) hantavirus shedding and viremia in naturally infected, wild 27
bank voles (Myodes glareolus). In a monthly capture-mark-recapture study, we analyzed 18 bank voles 28
for the presence and relative quantity of PUUV RNA in the excreta and blood from 2 months before up 29
to 8 months after seroconversion. 30
The proportion of animals shedding PUUV RNA in saliva, urine, and feces peaked during the first 31
month after seroconversion, but continued throughout the study period with only a slight decline. The 32
quantity of shed PUUV in RT-PCR positive excreta was constant over time. In blood, PUUV RNA was 33
present for up to 7 months but both the probability of viremia and the virus load declined by time. 34
Our findings contradict the current view of a decline in virus shedding after the acute phase and a short 35
viremic period in hantavirus infection – an assumption widely adopted into current epidemiological 36
models. We suggest the life-long shedding as a means of hantaviruses to survive over host population 37
bottlenecks, and to disperse in fragmented habitats where local host and/or virus populations face 38
temporary extinctions. Our results indicate that the kinetics of pathogens in wild hosts may 39
considerably differ from those observed in laboratory settings. 40
41
42

3
Introduction 43
Hantaviruses (family Bunyaviridae) are three-segmented, negative-stranded RNA viruses transmitted 44
by rodents, insectivores and bats (Guo et al., 2013). More than 50 species (ICTV-approved or tentative) 45
are currently known, and most of them are restrained to a single reservoir host species. Several 46
hantaviruses are human pathogens, causing annually up to 50 000 disease cases worldwide. Pathogenic 47
hantaviruses carried by Old World mice, rats and voles (family Muridae, subfamily Murinae; family 48
Cricetidae, subfamily Arvicolinae) cause Hemorrhagic Fever with Renal Syndrome (HFRS), and those 49
carried by New World rats and mice (family Cricetidae, subfamilies Sigmodontinae and Neotominae) 50
cause Hantavirus Cardiopulmonary Syndrome (HCPS) in humans (Jonsson et al., 2010). Puumala 51
hantavirus (PUUV) causes a mild form of HFRS, nephropathia epidemica (NE) in humans (Brummer-52
Korvenkontio et al., 1980) and accounts for the majority of hantaviral disease incidence in Europe 53
(Vaheri et al., 2013, Vapalahti et al., 2003). The reservoir host of PUUV is the bank vole (Myodes 54
glareolus). 55
Hantaviruses are transmitted among hosts by direct contact (Bernshtein et al., 1999, Botten et al., 2002, 56
Lee et al., 1981, Padula et al., 2004, Yanagihara et al., 1985) and also indirectly via virus-contaminated 57
excreta (Gavrilovskaya et al., 1990, Kallio et al., 2006, Lee et al., 1981, Nuzum et al., 1988) 58
maintaining their infectiousness outside the host for at least two weeks (Kallio et al., 2006). Inhalation 59
of aerosolized rodent excreta is also considered the main route of infection to humans (Hjelle & Glass, 60
2000, Vapalahti et al., 2010). No apparent symptoms have been reported in rodent hosts due to 61
hantavirus infection, but it may impair host survival in nature (Kallio et al., 2007, Luis et al., 2012, 62
Tersago et al., 2012). The hantavirus infection induces a life-long antibody response where IgG 63
appears in blood stream 2 to 3 weeks after infection, but viral RNA and/or infectious virus is found in 64

4
various tissues, especially in the lungs of chronically infected animals (Botten et al., 2003, Hardestam 65
et al., 2008, Meyer & Schmaljohn, 2000, Schountz et al., 2012). 66
The duration of hantavirus shedding by reservoir rodents is a critical factor for understanding both the 67
transmission dynamics in host populations and the risk for human infections. Hence, patterns of 68
hantavirus shedding and/or viremia in the reservoir species have been experimentally studied in 69
numerous host-virus pairs (Billings et al., 2010, Botten et al., 2000, 2002, and 2003, Fulhorst et al., 70
2002, Hardestam et al., 2008, Meyer & Schmaljohn, 2000, Schountz et al., 2012). Many of these 71
studies report a transient viremia of some days to several weeks (Fulhorst et al., 2002, Hutchinson et 72
al., 1998, Lee et al., 1981, Yanagihara et al., 1985) and suggest only occasional viral shedding after an 73
acute phase of few weeks (Botten et al., 2003, Gavrilovskaya et al., 1990, Hardestam et al., 2008, Lee 74
et al., 1981). This pattern of hantavirus shedding by the host has been adopted in a wide array of 75
mathematical models describing hantavirus transmission among rodents and predicting the risk of 76
human infections (Allen et al., 2009, Sauvage et al., 2007, Wolf, 2004). 77
However, the current knowledge of hantavirus shedding patterns is, to a large extent, drawn from 78
experimental infections of laboratory-reared animals, whereas only few studies have entailed the 79
dynamics of shedding in naturally infected, wild rodents (Bernshtein et al., 1999, Korva et al., 2009, 80
McIntyre et al., 2005, Safronetz et al., 2005 and 2008). In contrast to laboratory conditions with ad 81
libitum food and water and thermoneutral temperature, rodents under the limited resources in their 82
natural environment, often in harsh climatic conditions, are subjected to trade-offs between 83
reproductive effort, longevity and immune response (Martin et al., 2008, Mills et al., 2010). These 84
trade-offs may translate to enhanced or prolonged replication of pathogens when resource allocation to 85
immune response is compromised. Indeed, several studies have shown fairly large proportions, i.e. 19 86
to 91% (Boone et al., 1998 and 2002, Kuenzi et al., 2005, McIntyre et al., 2005, Otteson et al., 1996, 87

5
Rowe et al., 1995, Safronetz et al., 2005 and 2006), of wild, hantavirus antibody positive rodents to be 88
RNA viremic compared to what one might expect from the brief viremia observed in laboratory studies 89
(Easterbrook & Klein, 2008). We hypothesized that also hantavirus shedding via saliva, feces and urine 90
could last longer in naturally infected, wild rodents than has been suggested by infection experiments in 91
laboratory. In this study, we present the quantities and prevalence of PUUV shedding and viremia 92
observed in sequential samples of naturally infected, wild bank voles as measured by the presence or 93
PUUV RNA over 5 to 10 month individual follow-up periods. 94
95
Results 96
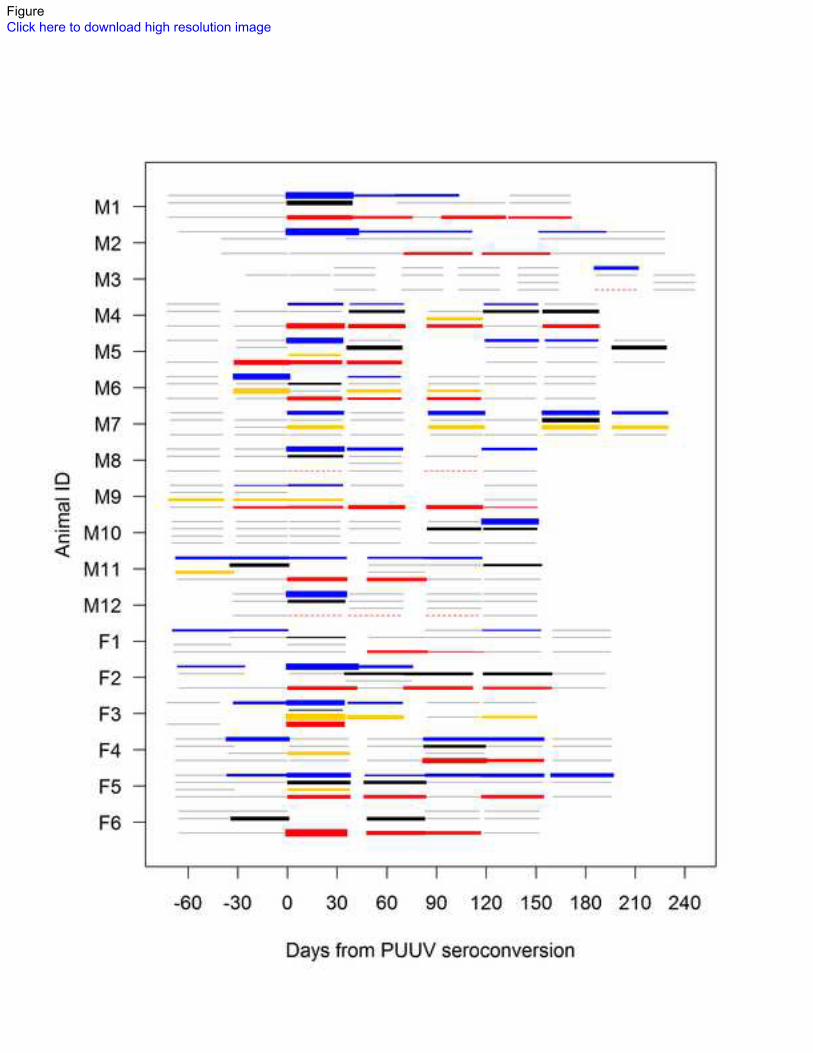
PUUV RNA presence in excreta. PUUV was found in saliva, feces, and urine of bank voles for 97
extensive periods after the approximated time of seroconversion; in each excretion, PUUV RNA was 98
present up to 213 days (±26 days, due to uncertainty of the seroconversion time) after seroconversion 99
(Fig. 1). In some animals, PUUV RNA was also detected long before seroconversion: in saliva, up to 100
50 (±16) days, and in urine, up to 55 (±16) days prior to seroconversion. In feces, PUUV was first 101
detected 16 (±16) days prior to seroconversion. Overall, PUUV RNA was found in approximately half 102
of the saliva and urine samples, whereas only one-fourth of fecal samples were RT-PCR positive 103
(Table 1a). In samples taken after seroconversion, 57%, 28%, and 51% of saliva, fecal, and urine 104
samples were RT-PCR positive, respectively. The proportion of animals shedding PUUV was highest 105
within the first month after seroconversion in all three excreta (Table 1a). Shedding PUUV RNA via 106
any one route appeared to be intermittent; in half of the cases (10/21) where two non-sequential 107
samples of the same animal and excretion showed positive for PUUV RNA, there was a negative 108
sample in between. However, in most of these cases, the animal concurrently shed PUUV via another 109

6
route or was viremic (Fig. 1). The bank voles exhibited notable individual variation in temporal 110
patterns of shedding. In some animals, the RT-PCR positive samples were concentrated around the 111
point of seroconversion (Fig. 1; M6, M12, F3); in others, positive samples occurred evenly throughout 112
the study period (M4, M5, M7, M11, F2, F5) and the excreta of some animals did not become RT-PCR 113
positive until a few months after seroconversion (M3, M10). Among the candidate models analyzing 114
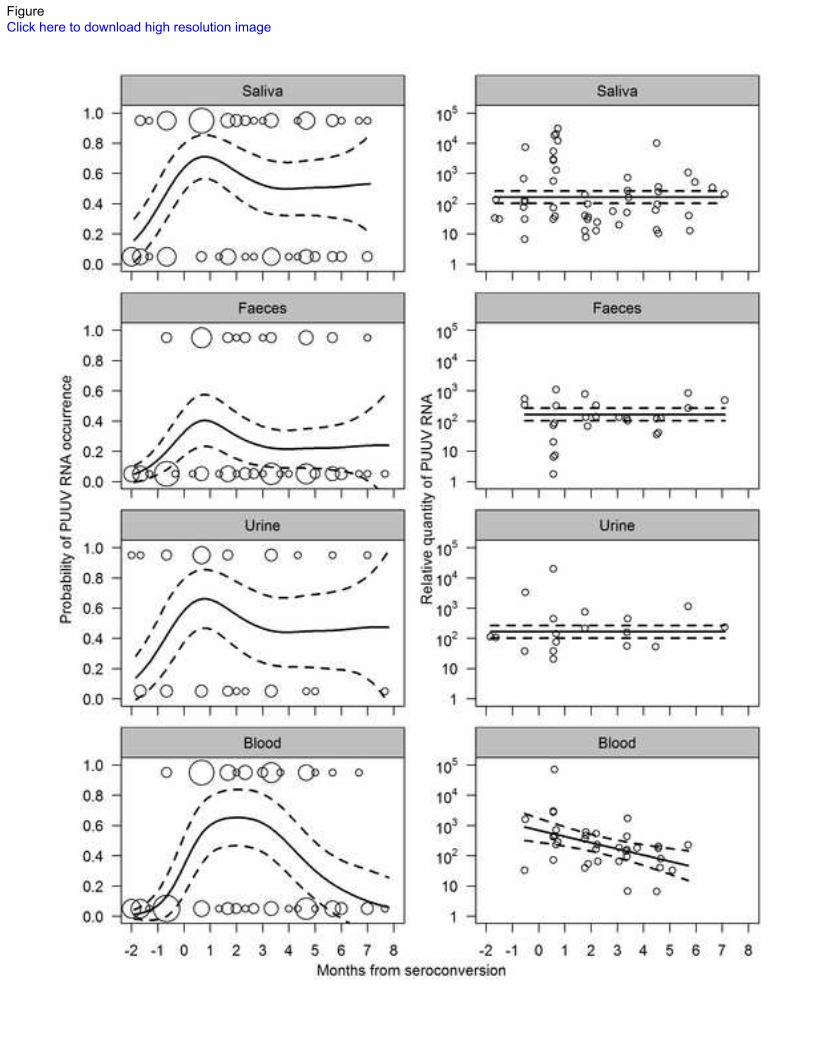
the probability of excreta being PUUV RNA positive, the best supported model was the one with a 115
single smooth term of time from seroconversion (P=0.0014) and different intercepts for each excretion 116
(Table 2). The probability of a bank vole to shed PUUV RNA in feces was significantly lower than the 117
probability to shed in saliva or urine (Wald statistic P<0.0001 for saliva and P=0.016 for urine). The 118
model predicted the probability of a bank vole to shed PUUV RNA to peak within the first month after 119
seroconversion in all three excreta (Fig. 2). During the second and third months after seroconversion, 120
the probability of shedding declined, and from three to eight months after seroconversion, it remained 121
on a constant level. The predicted probability of a bank vole to shed PUUV RNA after 3 months from 122
seroconversion was 71% of the probability predicted for the first month in saliva, 54% in feces, and 123
67% in urine (Fig. 2). The models including sex or its interaction with time as explanatory variables for 124
PUUV presence in excreta were not supported by Akaike information criteria corrected for sample size 125
(AICc, Hurvich & Tsai, 1989, Table 2). 126
PUUV RNA quantity in excreta. The highest single quantities of PUUV RNA (1 096, 19 953, and 30 127
903 relative units/µg RNA in feces, urine, and saliva, respectively) were observed within the first 128
month after seroconversion (Fig. 2). During the first month, also the average quantity in RT-PCR 129
positive saliva samples was highest (Table 1b). In feces, the quantities during the first month averaged 130
lower than in later months. No temporal trends in PUUV RNA quantity could be seen in urine samples. 131
Among the candidate models analyzing the relative quantity of PUUV RNA in RT-PCR positive 132

7
excreta, the model with a constant quantity (164 relative units, 95% CI: 101–263 units) through time 133
and between excreta and sexes was best supported when random individual variation in shedding 134
quantities was assumed (Table 2). The average quantities (±SD) of total RNA in saliva, fecal, urine, 135
and blood RNA elutions were 70±36, 146±105, 47±22, and 78±27 ng/µL, respectively. 136
PUUV in blood. PUUV RNA was detected in bank vole blood from 16 (±16) days before to 199 (±13) 137
days after approximated seroconversion (Fig. 1). Overall, 38% of blood samples were RT-PCR 138
positive, and the highest proportion (86%) of viremic animals was seen 1 month after the 139
seroconversion (Table 1a). Within 1 month before seroconversion, 13% of samples were PUUV RNA 140
viremic, and after seroconversion 52% of blood samples were RT-PCR positive. Among the candidate 141
models analyzing the probability of blood being PUUV RNA positive, the best supported model 142
appeared to be the one with a smooth term of time from seroconversion (P=0.0015, Table 2). The 143
model predicted more than 50% of bank voles to be viremic between 0.5 to 4 months after 144
seroconversion, after which the probability of viremia declined to zero by the 8th
month after 145
seroconversion (Fig. 2). In RT-PCR positive samples, the highest observed quantity (70 795 relative 146
units) occurred 1 month after seroconversion (Fig. 2), when the quantity also averaged as the highest 147
(Table 1b). Among the candidate models analyzing the relative quantity of PUUV RNA in RT-PCR 148
positive blood samples, the model with a declining smooth term of time from seroconversion 149
(P=0.0021) was best supported when random individual variation was assumed (Table 2). The model 150
predicted the PUUV RNA relative quantity to drop from 886 relative units (95% CI: 525–1495 relative 151
units) 1 month before to 47 relative units (95% CI: 26–83 relative units) 6 months after seroconversion 152
(Fig. 2). The models including sex or its interaction with time as explanatory variables for PUUV 153
presence or quantity in blood were not supported by AICc (Table 2). 154
155

8
Discussion 156
Hantavirus infection experiments performed on laboratory-reared rodent hosts have shown a few-week 157
acute phase of intensive virus shedding followed by a chronic phase of absence or occasional shedding 158
in excreta (Botten et al., 2003, Gavrilovskaya et al., 1990, Hardestam et al., 2008, Lee et al., 1981). 159
Our results suggest that this two-phase propagation pattern may not be applicable to describe 160
hantavirus transmission in natural settings, as we showed notably longer periods of PUUV shedding 161
and viremia to be common in wild, naturally infected bank voles. The proportion of shedding 162
individuals was highest during the first month after seroconversion, but declined by less than half 163
during later months of infection. On the other hand, no statistically significant temporal patterns could 164
be distinguished in the quantity of shed virus. However, in contrast to shedding patterns via excreta, 165
both the proportion of viremic individuals and the viral load in blood declined significantly by time 166
from infection. Also in saliva, but not in feces or urine, the relative quantities of PUUV RNA showed a 167
decreasing trend, but this tendency could not be statistically distinguished from overall individual-level 168
variation. The number of saliva samples studied here (N=50) may have been insufficient to show any 169
temporal trends. However, a decreasing trend of PUUV quantity in blood was statistically supported 170
despite the lower number of blood samples (N=43) in the analysis. 171
Bank vole saliva and urine were significantly more often PUUV RNA positive than feces. The lower 172
proportion of positive fecal samples compared to saliva and urine can, to a large extent, be explained 173
by the lower sensitivity of RT-PCR in fecal samples. Bank vole feces include PCR-inhibitory 174
components that reduce the sensitivity of the RT-PCR, so that the actual quantities of PUUV RNA in 175
feces are presumably 100-fold compared to quantities in saliva and urine, and a large proportion of 176
fecal samples were thus likely false-negative in RT-PCR (Hardestam et al., 2008). Also, comparison of 177
PUUV concentrations in different excreta is further complicated by the different volumes of excreta in 178

9
original samples, and the variation of concentrations of total RNA between excreta. In addition, 179
different shedding routes most likely yield varying amounts of viral particles that end up into contact 180
with new, susceptible hosts; most likely, viral particles in saliva are rather effectively administered into 181
the blood stream via a bite wound, but only a low proportion aerosolized viral particles in a urine or a 182
fecal excretion end up into the nostrils or mouth of susceptible animals. 183
The long shedding period in wild hosts evidenced here may result from the limited amount of resources 184
available for immune response in natural conditions. Nevertheless, besides the deprived immune 185
response in wild compared to laboratory-reared animals, also other factors may explain the lesser 186
propagation of hantavirus in laboratory studies: the adaptation of inoculated virus to cell culture 187
(Lundkvist et al., 1997), higher than natural infecting doses (Hardestam et al., 2008), and the low 188
immunogenetic variability in colonized experimental animals (Guivier et al., 2010a) may all bias the 189
shedding patterns observed in infection trials. 190
We acknowledge that the presence of PUUV RNA in blood or excreta does not necessarily indicate 191
infectiousness to new hosts. With this in mind, we did attempt virus isolation from PUUV RNA 192
positive excreta, but all our attempts failed, most likely since no efficient method exists for the isolation 193
of hantaviruses, and since our samples were not optimally preserved for virus isolation. However, in 194
cotton rats (Sigmodon hispidus) inoculated with Black Creek Canal hantavirus, infectious virus could 195
be recovered from salivary glands as long as they showed positive for viral cRNA, and the titers of cell 196
culture infectious virus in blood were consistent with the copy number levels of genomic S segment 197
RNA (Hutchinson et al., 1998). Furthermore, PUUV RT-PCR positive saliva, urine, and feces of 198
experimentally infected bank voles proved infectious when administered intranasally to naïve bank 199
voles (Hardestam et al., 2008). Although not all PUUV RNA detected in rodent samples is necessarily 200

10
associated within infectious particles, we have no reason to assume that the proportion of infectious 201
virus from detected PUUV RNA would change by time from infection. 202
Interestingly, we found PUUV RNA in saliva and urine of infected voles as early as seven weeks (or 203
five to nine weeks, taking into account the sampling interval) before seroconversion. By contrast, 204
following subcutaneous or intramuscular inoculations in laboratory, hantavirus antibodies have been 205
reported to appear already 1 to 3 weeks dpi (Botten et al., 2000, Fulhorst et al., 2002, Gavrilovskaya et 206
al., 1990, Hutchinson et al., 1998, Yanagihara et al., 1985). The relatively late appearance of antibodies 207
could to some extent reflect the lower sensitivity of IFAT compared to ELISA (Lundkvist et al., 1991) 208
used for antibody detection in some studies, but may also result from relatively low virus doses in bank 209
vole excreta: in line with our present results, when excreta of PUUV infected bank voles were 210
intranasally administered into colonized bank voles, only one out of seven successfully infected 211
animals seroconverted before 3 weeks dpi (Hardestam et al., 2008). 212
Thus far, only few data have been published on the occurrence of hantavirus RNA in the excreta of 213
wild rodents (Korva et al., 2009, McIntyre et al., 2005, Safronetz et al., 2005 and 2008). Our results of 214
overall PUUV RNA positivity of 57% in saliva and 52% in urine collected from seropositive animals 215
are similar to the observations where 53% and 35% of wild, Bayou hantavirus seropositive Orozymus 216
palustris shed viral RNA in saliva and urine, respectively (McIntyre et al., 2005). By contrast, lower 217
proportions of shedding (7–21% in saliva and 7–18% in urine) have been reported for Sin Nombre 218
hantavirus (SNV) seropositive deer mice (Peromyscus maniculatus; Safronetz et al., 2005 and 2008). 219
The difference between these results may arise from several factors, but it is possible that host species 220
of hantaviruses differ in their shedding potential as suggested earlier (Meyer & Schmaljohn, 2000, 221
Padula et al., 2004). The above mentioned studies on wild rodents have also reported temporal trends 222
of hantavirus shedding: SNV occurred more often in the saliva of recently seroconverted than 223

11
chronically infected deer mice (Safronetz et al., 2008), and higher quantities of PUUV, Dobrava, and 224
Saaremaa hantavirus RNA were detected in the urine bladders of recently seroconverted than 225
chronically infected bank voles, yellow-necked mice (Apodemus flavicollis), and striped field mice 226
(Apodemus agrarius), respectively (Korva et al., 2009). However, in these two studies the timing of 227
seroconversion was based on IgG antibody avidity, which has been shown unreliable in low antibody 228
titers (Varner & Dearing., 2011). In the present study, we were able to determine the time of 229
seroconversion with a more reliable accuracy (±13 to ±24 days), followed individual wild hosts much 230
longer than in previous studies, and showed hantavirus shedding to last throughout the typical life span 231
(Innes & Millar, 1994, Prévot-Julliard et al., 1999) of the rodent host species without drastic decline. 232
Our finding of high prevalence of hantavirus RNA viremia in seroconverted bank voles is consistent 233
with several earlier studies on wild rodents (Bagamian et al., 2012, Boone et al., 1998, 2002, Kuenzi et 234
al., 2005, McIntyre et al., 2005, Otteson et al., 1996, Rowe et al., 1995, Safronetz et al., 2005 2006, 235
and 2008) . Most individuals were viremic for four months after seroconversion, which is in 236
concordance with the results by Boone and others (Boone et al., 1998), where SNV RNA viremia 237
persisted for at least three months in wild, seroconverted deer mice. An experimental study on SNV 238
transmission in deer mice in outdoor enclosures showed a tendency of high viremic animals to more 239
often transmit the infection to new hosts within the first two months of infection (Bagamian et al., 240
2012), suggesting that RNA viremia correlates with host infectiousness. Our data show that albeit this 241
might be true within the first months of infection, after 4 months from seroconversion the bank voles 242
shed PUUV in excreta whilst the virus was no longer found in blood. Therefore, host viremia may not 243
be a good indicator of host infectiousness. 244
In the present study, no sex differences in likelihood, duration, or quantity of PUUV shedding were 245
found. This may be due to the fact that most of the bank voles in this study were sexually immature, 246

12
overwintering subadults, whereas an earlier study (Hannah et al., 2008) have shown sex steroids to 247
mediate sex differences in hantavirus shedding. Here, the first months of infection were concentrated in 248
the winter, the most challenging season in terms of thermoregulation and food resources. It remains as 249
a target of our further studies to determine how the more abundant food resources and warmer 250
temperature, and on the other hand, the possible costs of sex steroids, mediate PUUV shedding during 251
the breeding season. 252
In cyclic fluctuations of bank vole populations, phases of low density may last for more than a year and 253
are characterized by a low abundance or absence of infected individuals (Kallio et al., 2009, Tersago et 254
al., 2008). We suggest that the persistent PUUV shedding by the individuals that outlive population 255
bottlenecks facilitates the re-initiation of PUUV transmission as the population again starts to grow. 256
Although PUUV has been demonstrated to remain infectious in the environment for several weeks 257
(Kallio et al., 2006), this time most probably is not long enough for the virus to survive in areas where 258
susceptible hosts are long absent. 259
The genomes of both rodent host species and their endemic viruses show notable geographic variation 260
(Asikainen et al., 2000, Deffontaine et al., 2005) that may translate into differential host-pathogen 261
relationships (Guivier et al., 2010b). It remains to be studied whether the geographical differences in 262
the balance of virus tolerance/resistance are reflected in the shed quantities of PUUV — if true, such a 263
difference could in part explain the higher incidence of NE in Finland and elsewhere in Northern 264
Europe compared to France and temperate Europe in general (Vaheri et al., 2013). 265
The majority of mathematical models describing rodent-hantavirus interactions and/or predicting 266
human hantavirus disease risk assume two discernible phases of shedding in infected rodents — an 267
acute phase versus a chronic phase (Allen et al., 2009, Sauvage et al., 2007, Wolf, 2004). In contrast to 268

13
these model assumptions, we found no notable decrease in virus shedding during the course of a 269
rodent’s life. This finding suggests that the abundance of infectious hosts, the most important driver for 270
human hantavirus epidemics, may be underestimated by current mathematical models. Whichever the 271
underlying reasons for the discrepancy between experimental and natural infections, our results stress 272
the importance of studying zoonotic infections in natural settings. We strongly encourage further 273
studies on hantavirus shedding by wild hosts in other virus-host pairs and impose a need to revise the 274
assumptions of infectiousness in hantavirus transmission models, which in turn will provide improved 275
tools to predict the threat of hantavirus infection to public health. 276
277
Methods 278
Collection of wild rodent samples. The study took place at Konnevesi, Central Finland (62°34’ N, 279
26°24’ E), where PUUV is endemic in bank voles (Kallio et al., 2007, 2009, and 2010, Razzauti et al., 280
2008). The study site is situated in the hospital district of Central Finland, where mean annual NE 281
incidence is 71/100 000, the fourth highest of the 20 hospital districts in Finland (Makary et al., 2010). 282
The excreta and blood samples were collected on 13 trapping periods from January 2008 to May 2009 283
at 27 to 48 day intervals. The sample collection was a part of a long-term capture-mark-recapture study, 284
where bank voles were captured on a 5.8 ha trapping grid consisting of 246 trapping stations at 15 m 285
intervals. One Ugglan Special live-trap (Grahnab, Sweden) was permanently placed on each trapping 286
station under a sheet metal “trap chimney” that enabled an access to the traps during the snowy period 287
and provided shelter for captured animals. Each trapping period lasted 3−4 days, during which the traps 288
were checked ten times. Bank voles were individually marked on first capture with subcutaneous PIT 289
tags (Trovan, U.K.), and their blood, urine, feces, and saliva samples were collected on every trapping 290

14
period. The animals were bled through the retro-orbital sinus using heparinized capillaries 291
(Hirschmann, Germany). Two aliquots of 18 μL were taken, one for RT-qPCR and the other for PUUV 292
immunofluorescent antibody test. Urine samples were collected using plastic buckets that were 293
sterilized between animals by soaking with 2.5% Umonium38
Instruments & Equipments (Huckert’s 294
International) for over five minutes, machine-washed (60°C, 2 min pre-rinse, 4 min wash with 295
detergent, 2 min rinse), sprayed with 70% EtOH and air-dried. The animals were carefully placed at the 296
bottom of a bucket and any generated urine pools were immediately collected with a sterile pipette. 297
Samples that got in contact with the animal’s paws, tail, snout, or fur were discarded. Fresh feces were 298
collected from the bucket or directly from the anus. Saliva was collected using Copan flocked swabs 299
(Copan, U.S.) that were subsequently placed in 500 µL of Universal Transport Medium (Millipore, 300
U.S.). All samples were immediately frozen on dry ice and stored later in –80°C until analyses. All 301
handling procedures of wild bank voles followed the Finnish Act on the Use of Animals for 302
Experimental Purposes (62/2006) and took place with permission from Finnish Animal Experiment 303
Board (license number HY 54-05). 304
Dating the seroconversion and selection of samples for RT-qPCR. As the exact date of infection 305
could not be determined in naturally infected animals, we dated the blood and excretion sampling times 306
in relation to seroconversion, i.e. the appearance of PUUV specific antibodies, instead of days post 307
infection. The presence of PUUV antibodies in each animal on each trapping period was determined 308
from blood samples diluted 1:10 in phosphate-buffered saline using an earlier described 309
immunofluorescent antibody test (Kallio-Kokko et al., 2006). Altogether, we sampled blood and 310
excreta from 352 bank voles, from which 109 had a known time of seroconversion. Based on the 311
antibody results, we selected blood and excretion samples of bank vole individuals where the follow-up 312
time was the longest, i.e., animals whose blood and excreta had been sampled at least once before and 313

15
four times after seroconversion, to be analyzed for the presence and quantity of PUUV RNA. 314
Altogether 349 samples from 121 trapping occasions of 18 bank voles (12 males, 6 females) were thus 315
analyzed, from which 114 were blood, 100 saliva, 98 feces, and 37 urine. In addition, excreta and blood 316
from six trapping occasions of one PUUV antibody negative female bank vole were used as negative 317
controls. The seroconversion of a bank vole had a uniform probability to have happened on any day 318
between the last seronegative and the first seropositive blood sample, but was approximated to have 319
happened on the date exactly between these two sampling dates. As the sampling interval preceding the 320
first seropositive sample varied from 26 to 48 days, the seroconversion could be dated with an accuracy 321
of ±13 to ±24 days. The selected individuals were wintering, non-breeding subadults during most of the 322
study, except for two males (M1, M3) that were sexually mature during the month preceding 323
seroconversion, and three individuals (M7, F1, and F3) that had reached breeding condition when last 324
sampled. The seroconversion took place during fall (September-November) in 16/18 animals. 325
Extraction of viral RNA. Viral RNA was extracted from blood samples by dissolving the 18 µL 326
sample into 450 µL TRIzol (Invitrogen Life TechnologiesTM
), then mixing with 150 µL chloroform and 327
centrifuging for 15 minutes in +4°C, at 12 000 g. RNA extraction was then continued using QIAamp 328
Viral RNA Mini Kit (Qiagen, U.S.) according to manufacturer’s instructions, so that 140 µL of the 329
formed water phase was mixed with 560 µL Buffer AVL (Qiagen, U.S.) and 5.6 µL cRNA. For saliva, 330
urine, and feces, only QIAamp Viral RNA Mini Kit was used. The fecal samples (each consisting of 1 331
to 3 fecal pellets) were first homogenized in 500 µL NaCl and centrifuged with 6000 rpm in +4°C for 332
10 minutes. Then 140 μL supernatant was mixed with 560 μL AVL buffer with 5.6 μL cRNA. For 333
saliva and urine, 70 µL of sample was mixed with 70 µL NaCl, 560 µL Buffer AVL and 5.6 µL cRNA. 334
The RNA elution was stored at −70°C until analysis. The concentration of RNA was measured after 335
extraction by NanoDrop (Thermo Fisher Scientific). 336

16
Reverse transcription quantitative PCR. The amount of PUUV RNA in samples was estimated by 337
relative quantitation of PUUV copies. One-step real-time RT-PCR was performed with qScript One-338
Step Fast MGB qRT-PCR Kit, low ROX (Quanta BioSciences) in a StrataGene Mx3005p. ABI Prism 339
7900 SDS was used instead of StrataGene for the analyses of the 17 blood samples from 3 animals 340
(M3, M8, M12). The primer and probe sequences were optimized to match the PUUV strain circulating 341
at the trapping site (Razzauti et al., 2008): Fw12 - TCC TTG AAA AGC TAC TAC GAG AAA AA, 342
Rev87 - TTC ATG RCG GGT TAT ATC CTC TT, and the probe39 6-FAM-TGG AAT GAG TGA 343
CTT GAC AG-MGB. Five µL of RNA was added to a reaction that contained 0.9 µM primers and 0.2 344
µM probes, and had a total volume of 20 µL. The RT-qPCR was confirmed highly specific since RNA 345
samples of other hantaviruses tested (Dobrava, Saaremaa, Hantaan and Tula) were negative as well as 346
samples from non-infected bank voles. The standard curve for relative quantitation was calculated from 347
10-fold dilutions of RNA extracted from a lung sample of a PUUV-infected bank vole captured at the 348
same site in an earlier study (Razzauti et al., 2008). The last dilution of RNA (10-7
) that was 349
consistently positive was set as relative quantity 1. The analytical sensitivity of the assay was 350
determined for the different sample materials (Supplementary Table S1), and was found equivalent for 351
saliva, urine and blood, but lower for feces. The standard RNA was aliquoted and a new dilution series 352
was added to each RT-qPCR run. All samples were initially screened once to find the positive samples. 353
RNA positive samples were then quantified using triplicate reactions, and the mean quantities of the 354
triplicates were used for further analyses. Reactions where the cycle threshold value deviated by more 355
than 1 cycle from the other two reactions in the triplicate were regarded as artifacts, e.g. pipetting 356
errors, and thus discarded. As the volumes of saliva and feces used for RNA extraction were unknown, 357
the quantity of PUUV RNA in each positive excretion and blood sample was calculated as the quantity 358
per 1 µg total RNA. 359

17
Statistical analyses. PUUV shedding (PUUV in saliva, feces, and urine) and viremia (PUUV RNA in 360
blood) in bank voles were each studied in separate statistical analyses. Generalized additive mixed 361
models (GAMM, Zuur, 2009) in gamm4 package (Wood, 2011) for R statistical software (R 362
Development Core Team, 2011) were used to analyze the presence and the quantity of PUUV RNA in 363
blood and excretion samples in relation to time of seroconversion and sex. More than half of the 364
samples were RT-PCR negative, and thus the relative quantity of PUUV RNA showed a highly zero-365
inflated distribution. When RT-PCR negative samples were excluded, the log10-transformed relative 366
quantities showed a normal distribution. Therefore, we studied both shedding and viremia with two 367
separate models; one to predict PUUV RNA presence/absence with binomial error distributions and a 368
logit link function, and another to predict the log10-transformed relative quantity of PUUV RNA in 369
RT-PCR positive samples with Gaussian error distributions and an identity link function. 370
A set of candidate models with different explanatory variables was constituted for each separate 371
analysis, and AICc scores were compared to find the most parsimonious model in each model set, i.e., 372
the smallest possible set of explanatory variables that explains the variation in the response variable. 373
The model with the lowest AICc score was considered as best supported by the data and used for 374
statistical inference. The candidate model sets analyzing the presence and quantity of PUUV in excreta 375
included two “beyond optimal” models and all their submodels: one with separate smooth terms of 376
time from seroconversion for each sex and different intercepts for each excretion (saliva/feces/urine), 377
and another with separate smooth terms of time for each excretion and different intercepts for each sex. 378
The candidate model sets analyzing the presence and quantity of PUUV in blood included a “beyond 379
optimal” model with separate smooth terms of time from seroconversion for each sex, and all possible 380
models nested therein. As wild rodents were expected to show great variation in immunogenetic 381
properties, physiological status and other characteristics that may influence hantavirus replication and 382

18
persistence (Easterbrook & Klein, 2008), random intercepts attributable to individual variation were 383
included in all candidate models. 384
385
Acknowledgments 386
We would like to thank Konnevesi Research Station of the University of Jyväskylä, Finland, for 387
laboratory facilities to perform the blood and excretion sampling. This study was funded by EU grant 388
FP7-261504 EDENext and is catalogued by the EDENext Steering Committee as EDENext132 389
(http://www.edenext.eu). The contents of this publication are the sole responsibility of the authors and 390
do not necessarily reflect the views of the European Commission. This study was also supported by 391
grants from Kone Foundation (www.koneensaatio.fi/en), Emil Aaltonen Foundation 392
(www.emilaaltonen.fi/eng.htm), Jenny and Antti Wihuri Foundation 393
(www.wihurinrahasto.fi/foundation.html), and the EU 6th
Framework Programme (GOCE-2003-394
010284 EDEN, www.eden-fp6project.net). 395
396
397
398

19
References 399
Allen, L. J. S., Wesley, C. L., Owen, R. D., Goodin, D. G., Koch, D., Jonsson, C. B., Chu, Y., 400
Hutchinson, J. M. S. & Paige, R. L. (2009). A habitat-based model for the spread of hantavirus 401
between reservoir and spillover species. J Theor Biol 260, 510-522. 402
Asikainen, K., Hänninen, T., Henttonen, H., Niemimaa, J., Laakkonen, J., Andersen, H., Bille, N., 403
Leirs, H., Vaheri, A. & Plyusnin, A. (2000). Molecular evolution of Puumala hantavirus in 404
Fennoscandia: Phylogenetic analysis of strains from two recolonization routes, Karelia and Denmark. 405
J Gen Virol 81, 2833-2841. 406
Bagamian, K. H., Towner, J. S., Kuenzi, A. J., Douglass, R. J., Rollin, P. E., Waller, L. A. & 407
Mills, J. N. (2012). Transmission ecology of Sin Nombre hantavirus in naturally infected North 408
American deermouse populations in outdoor enclosures. PLoS One 7, e47731. 409
Bernshtein, A. D., Apekina, N. S., Mikhailova, T. V., Myasnikov, Y. A., Khlyap, L. A., Korotkov, 410
Y. S. & Gavrilovskaya, I. N. (1999). Dynamics of Puumala hantavirus infection in naturally infected 411
bank voles (Clethrinomys glareolus). Arch Virol 144, 2415-2428. 412
Billings, A. N., Rollin, P. E., Milazzo, M. L., Molina, C. P., Eyzaguirre, E. J., Livingstone, W., 413
Ksiazek, T. G. & Fulhorst, C. F. (2010). Pathology of Black Creek Canal virus infection in juvenile 414
hispid cotton rats (Sigmodon hispidus). Vector Borne Zoonotic Dis 10, 621-628. 415
Boone, J. D., Otteson, E. W., McGwire, K. C., Villard, P., Rowe, J. E. & St Jeor, S. C. (1998). 416
Ecology and demographics of hantavirus infections in rodent populations in the Walker River Basin of 417
Nevada and California. Am J Trop Med Hyg 59. 418
Boone, J., McGwire, K., Otteson, E., Debaca, R., Kuhn, E. & St Jeor, S. (2002). Infection dynamics 419
of Sin Nombre virus after a widespread decline in host populations. Am J Trop Med Hyg 67, 310-318. 420

20
Botten, J., Mirowsky, K., Kusewitt, D., Bharadwaj, M., Yee, J., Ricci, R., Feddersen, R. & Hjelle, 421
B. (2000). Experimental infection model for Sin Nombre hantavirus in the deer mouse (Peromyscus 422
maniculatus). Proc Natl Acad Sci U S A 97, 10578-10583. 423
Botten, J., Mirowsky, K., Ye, C., Gottlieb, K., Saavedra, M., Ponce, L. & Hjelle, B. (2002). 424
Shedding and intracage transmission of Sin Nombre hantavirus in the deer mouse (Peromyscus 425
maniculatus) model. J Virol 76, 7587-7594. 426
Botten, J., Mirowsky, K., Kusewitt, D., Ye, C., Gottlieb, K., Prescott, J. & Hjelle, B. (2003). 427
Persistent Sin Nombre virus infection in the deer mouse (Peromyscus maniculatus) model: Sites of 428
replication and strand-specific expression. J Virol 77, 1540-1550. 429
Brummer-Korvenkontio, M., Vaheri, A., Hovi, T., von Bonsdorff, C. H., Vuorimies, J., Manni, 430
T., Penttinen, K., Oker-Blom, N. & Lähdevirta, J. (1980). Nephropathia epidemica: Detection of 431
antigen in bank voles and serologic diagnosis of human infection. J Infect Dis 141, 131-134. 432
Deffontaine, V., Libois, R., Kotlík, P., Sommer, R., Nieberding, C., Paradis, E., Searle, J. B. & 433
Michaux, J. R. (2005). Beyond the Mediterranean peninsulas: evidence of central European glacial 434
refugia for a temperate forest mammal species, the bank vole (Clethrionomys glareolus). Mol Ecol 435
14, 1727-1739. 436
Easterbrook, J. D. & Klein, S. L. (2008). Immunological mechanisms mediating hantavirus 437
persistence in rodent reservoirs. PLoS Pathog 4, e1000172. 438
Fulhorst, C. F., Milazzo, M. L., Duno, G. & Salas, R. A. (2002). Experimental infection of the 439
Sigmodon alstoni cotton rat with Caño Delgadito virus, a South American hantavirus. Am J Trop Med 440
Hyg 67, 107-111. 441
Gavrilovskaya, I. N., Apekina, N. S., Bernshtein, A. D., Varvara, T. D., Okulova, N. M., 442
Myasnikov, Y. A. & Chomakova, M. P. (1990). Pathogenesis of hemorrhagic fever with renal 443

21
syndrome virus infection and mode of horizontal transmission of hantavirus in bank voles. Arch Virol 444
Suppl 1, 57-62. 445
Guivier, E., Galan, M., Male, P. G., Kallio, E. R., Voutilainen, L., Henttonen, H., Olsson, G. E., 446
Lundkvist, Å., Tersago, K. & other authors (2010a). Associations between MHC genes and 447
Puumala virus infection in Myodes glareolus are detected in wild populations, but not from 448
experimental infection data. J Gen Virol 91. 449
Guivier, E., Galan, M., Salvador, A. R., Xuereb, A., Chaval, Y., Olsson, G. E., Essbauer, S., 450
Henttonen, H., Voutilainen, L. & other authors (2010b). Tnf-alpha expression and promoter 451
sequences reflect the balance of tolerance/resistance to Puumala hantavirus infection in European bank 452
vole populations. Infect Genet Evol 10, 1208-1217. 453
Guo, W., Lin, X., Wang, W., Tian, J., Cong, M., Zhang, H., Wang, M., Zhou, R., Wang, J. & 454
other authors (2013). Phylogeny and origins of hantaviruses harbored by bats, insectivores, and 455
rodents. PLoS Pathog 9, 3159-3159. 456
Hannah, M. F., Bajic, V. B. & Klein, S. L. (2008). Sex differences in the recognition of and innate 457
antiviral responses to Seoul virus in Norway rats. Brain Behav Immun 22, 503-516. 458
Hardestam, J., Karlsson, M., Falk, K. I., Olsson, G., Klingström, J. & Lundkvist, Å. (2008). 459
Puumala hantavirus excretion kinetics in bank voles (Myodes glareolus). Emerg Infect Dis 14, 1209-460
1215. 461
Hjelle, B. & Glass, G. (2000). Outbreak of hantavirus infection in the Four Corners region of the 462
United States in the wake of the 1997-1998 El Niño-southern oscillation. J Infect Dis 181, 1569-1573. 463
Hurvich, C. M. & Tsai, C. L. (1989). Regression and time-series model selection in small samples. 464
Biometrika 76. 465

22
Hutchinson, K. L., Rollin, P. E. & Peters, C. J. (1998). Pathogenesis of a North American 466
hantavirus, Black Creek Canal virus, in experimentally infected Sigmodon hispidus. Am J Trop Med 467
Hyg 59, 58-65. 468
Innes, D. & Millar, J. (1994). Life-histories of Clethrionomys and Microtus (Microtinae). Mamm Rev 469
24, 179-207. 470
Jonsson, C. B., Figueiredo, L. T. & Vapalahti, O. (2010). A global perspective on hantavirus 471
ecology, epidemiology, and disease. Clin Microbiol Rev 23, 412-441. 472
Kallio, E. R., Klingström, J., Gustafsson, E., Manni, T., Vaheri, A., Henttonen, H., Vapalahti, O. 473
& Lundkvist, Å. (2006). Prolonged survival of Puumala hantavirus outside the host: Evidence for 474
indirect transmission via the environment. J Gen Virol 87, 2127-2134. 475
Kallio, E. R., Voutilainen, L., Vapalahti, O., Vaheri, A., Henttonen, H., Koskela, E. & Mappes, T. 476
(2007). Endemic hantavirus infection impairs the winter survival of its rodent host. Ecology 88, 1911-477
1916. 478
Kallio, E. R., Begon, M., Henttonen, H., Koskela, E., Mappes, T., Vaheri, A. & Vapalahti, O. 479
(2009). Cyclic hantavirus epidemics in humans — predicted by rodent host dynamics. Epidemics 1, 480
101-107. 481
Kallio, E. R., Begon, M., Henttonen, H., Koskela, E., Mappes, T., Vaheri, A. & Vapalahti, O. 482
(2010). Hantavirus infections in fluctuating host populations: The role of maternal antibodies. Proc 483
Biol Sci 277, 3783-3791. 484
Kallio-Kokko, H., Laakkonen, J., Rizzoli, A., Tagliapietra, V., Cattadori, I., Perkins, S. E., 485
Hudson, P. J., Cristofolini, A., Versini, W. & other authors (2006). Hantavirus and arenavirus 486
antibody prevalence in rodents and humans in Trentino, Northern Italy. Epidemiol Infect 134, 830-836. 487
Korva, M., Duh, D., Saksida, A., Trilar, T. & Avsic-Zupanc, T. (2009). The hantaviral load in 488
tissues of naturally infected rodents. Microb Infect 11, 344-351. 489

23
Kuenzi, A., Douglass, R., Bond, C., Calisher, C. & Mills, J. (2005). Long-term dynamics of Sin 490
Nombre viral RNA and antibody in deer mice in Montana. J Wildl Dis 41, 473-481. 491
Lee, H. W., Lee, P. W., Baek, L. J., Song, C. K. & Seong, I. W. (1981). Intraspecific transmission of 492
Hantaan virus, etiologic agent of Korean hemorrhagic fever, in the rodent Apodemus agrarius. Am J 493
Trop Med Hyg 30, 1106-1112. 494
Luis, A. D., Douglass, R. J., Hudson, P. J., Mills, J. N. & Bjørnstad, O. N. (2012). Sin Nombre 495
hantavirus decreases survival of male deer mice. Oecologia 169, 431-439. 496
Lundkvist, Å., Fatouros, A. & Niklasson, B. (1991). Antigenic variation of European hemorrhagic 497
fever with renal syndrome virus strains characterized using bank vole monoclonal antibodies. J Gen 498
Virol 72, 2097-2103. 499
Lundkvist, Å., Cheng, Y., Sjölander, K. B., Niklasson, B., Vaheri, A. & Plyusnin, A. (1997). Cell 500
culture adaptation of Puumala hantavirus changes the infectivity for its natural reservoir, 501
Clethrionomys glareolus, and leads to accumulation of mutants with altered genomic RNA S segment. 502
J Virol 71, 9515-9523. 503
Makary, P., Kanerva, M., Ollgren, J., Virtanen, M. J., Vapalahti, O. & Lyytikäinen, O. (2010). 504
Disease burden of Puumala virus infections, 1995-2008. Epidemiol Infect 138, 1484-1492. 505
Martin, L. B., Weil, Z. M. & Nelson, R. J. (2008). Seasonal changes in vertebrate immune activity: 506
Mediation by physiological trade-offs. Philos Trans R Soc B-Biol Sci 363, 321-339. 507
McIntyre, N. E., Chu, Y. K., Owen, R. D., Abuzeineh, A., De La Sancha, N., Dick, C. W., 508
Holsomback, T., Nisbett, R. A. & Jonsson, C. (2005). A longitudinal study of Bayou virus, hosts, and 509
habitat. Am J Trop Med Hyg 73, 1043-1049. 510
Meyer, B. J. & Schmaljohn, C. S. (2000). Persistent hantavirus infections: Characteristics and 511
mechanisms. Trends Microbiol 8, 61-67. 512

24
Mills, S. C., Grapputo, A., Jokinen, I., Koskela, E., Mappes, T. & Poikonen, T. (2010). Fitness 513
trade-offs mediated by immunosuppression costs in a small mammal. Evolution 64, 166-179. 514
Nuzum, E. O., Rossi, C. A., Stephenson, E. H. & LeDuc, J. W. (1988). Aerosol transmission of 515
Hantaan and related viruses to laboratory rats. Am J Trop Med Hyg 38, 636-640. 516
Otteson, E., Riolo, J., Rowe, J., Nichol, S., Ksiazek, T., Rollin, P. & StJeor, S. (1996). Occurrence 517
of hantavirus within the rodent population of Northeastern California and Nevada. Am J Trop Med Hyg 518
54, 127-133. 519
Padula, P., Figueroa, R., Navarrete, M., Pizarro, E., Cadiz, R., Bellomo, C., Jofre, C., Zaror, L., 520
Rodriguez, E. & Murua, R. (2004). Transmission study of Andes hantavirus infection in wild 521
sigmodontine rodents. J Virol 78, 11972-11979. 522
Prévot-Julliard, A., Henttonen, H., Yoccoz, N. & Stenseth, N. (1999). Delayed maturation in female 523
bank voles: Optimal decision or social constraint? J Anim Ecol 68, 684-697. 524
R Development Core Team (2011). R: A language and environment for statistical computing. R 525
foundation for statistical computing. 2.13.0. 526
Razzauti, M., Plyusnina, A., Henttonen, H. & Plyusnin, A. (2008). Accumulation of point mutations 527
and reassortment of genomic RNA segments are involved in the microevolution of Puumala hantavirus 528
in a bank vole (Myodes glareolus) population. J Gen Virol 89, 1649-1660. 529
Rowe, J., StJeor, S., Riolo, J., Otteson, E., Monroe, M., Henderson, W., Ksiazek, T., Rollin, P. & 530
Nichol, S. (1995). Coexistence of several novel hantaviruses in rodents indigenous to North America. 531
Virology 213, 122-130. 532
Safronetz, D., Lindsay, R., Dibernardo, A., Hjelle, B., Xiao, R., Artsob, H. & Drebot, M. A. 533
(2005). A preliminary study of the patterns of Sin Nombre viral infection and shedding in naturally 534
infected deer mice (Peromyscus maniculatus). Vector Borne Zoonotic Dis 5, 127-132. 535

25
Safronetz, D., Lindsay, R., Hjelle, B., Medina, R. A., Mirowsky-Garcia, K. & Drebot, M. A. 536
(2006). Use of IgG avidity to indirectly monitor epizootic transmisson of Sin Nombre virus in deer 537
mice (Peromyscus maniculatus). Am J Trop Med Hyg 75, 1135-1139. 538
Safronetz, D., Drebot, M. A., Artsob, H., Cote, T., Makowski, K. & Lindsay, L. R. (2008). Sin 539
Nombre virus shedding patterns in naturally infected deer mice (Peromyscus maniculatus) in relation to 540
duration of infection. Vector Borne Zoonotic Dis 8, 97-100. 541
Sauvage, F., Langlais, M. & Pontier, D. (2007). Predicting the emergence of human hantavirus 542
disease using a combination of viral dynamics and rodent demographic patterns. Epidemiol Infect 135, 543
46-56. 544
Schountz, T., Acuña-Retamar, M., Feinstein, S., Prescott, J., Torres-Perez, F., Podell, B., Peters, 545
S., Ye, C., Black, W. C. & Hjelle, B. (2012). Kinetics of immune responses in deer mice 546
experimentally infected with Sin Nombre virus. J Virol 86, 10015-10027. 547
Tersago, K., Schreurs, A., Linard, C., Verhagen, R., Van Dongen, S. & Leirs, H. (2008). 548
Population, environmental, and community effects on local bank vole (Myodes glareolus) Puumala 549
virus infection in an area with low human incidence. Vector Borne Zoonotic Dis 8, 235-244. 550
Tersago, K., Crespin, L., Verhagen, R. & Leirs, H. (2012). Impact of Puumala virus infection on 551
maturation and survival in bank voles: A capture-mark-recapture analysis. J Wildl Dis 48. 552
Vaheri, A., Henttonen, H., Voutilainen, L., Mustonen, J., Sironen, T. & Vapalahti, O. (2013). 553
Hantavirus infections in Europe and their impact on public health. Rev Med Virol 23, 35-49. 554
Vapalahti, O., Mustonen, J., Lundkvist, Å., Henttonen, H., Plyusnin, A. & Vaheri, A. (2003). 555
Hantavirus infections in Europe. Lancet Infect Dis 3, 653-661. 556
Vapalahti, K., Virtala, A. M., Vaheri, A. & Vapalahti, O. (2010). Case-control study on Puumala 557
virus infection: Smoking is a risk factor. Epidemiol Infect 138, 576-584. 558

26
Varner, J. & Dearing, M. D. (2011). Estimating duration of infection with avidity assays: Potential 559
limitations and recommendations for improvement. EcoHealth 8, 512-518. 560
Wolf, C. (2004). A mathematical model for the propagation of a hantavirus in structured populations. 561
Discrete Contin Dyn Syst -Ser B 4, 1065-1089. 562
Wood, S. (2011). gamm4: Generalized additive mixed models using mgcv and lme4. R package 563
version 0.1-2. 564
Yanagihara, R., Amyx, H. L. & Gajdusek, D. C. (1985). Experimental infection with Puumala virus, 565
the etiologic agent of nephropathia epidemica, in bank voles (Clethrionomys glareolus). J Virol 55, 34-566
38. 567
Zuur A. F. (2009). Mixed effects models and extensions in ecology with R. Springer, NY. 568
569
570

27
Tables 571
Table 1. Bank vole excreta and blood tested and positive for PUUV RNA (a) and the geometric means 572
and 95% confidence intervals of PUUV RNA relative quantities in RT-PCR positive samples (b). 573
Numbers are pooled for each month by the approximated time from seroconversion of each animal. 574
(a) 575
Months from Saliva
Feces
Urine
Blood
seroconversion Positive/N %
Positive/N %
Positive/N %
Positive/N %
-2 3/16 19 0/1 0 2/5 40
0/15 0
-1 7/14 50
2/15 13
2/5 40
2/16 13
1 12/14 86
8/13 62
6/9 67
12/17 71
2 7/13 54
3/10 30
2/5 40
6/12 50
3 3/5 60
2/7 29
0/1 0
4/7 57
4 5/12 42
3/15 20
3/6 50
11/16 69
5 7/12 58
4/12 33
1/2 50
5/15 33
6 4/10 40
2/11 18
1/2 50
2/11 18
7 1/2 50
0/2 0
- -
1/2 50
8 1/2 50
1/2 50
1/2 50
0/3 0
Total 50/100 50 25/98 26 18/37 49
43/114 38
576
577
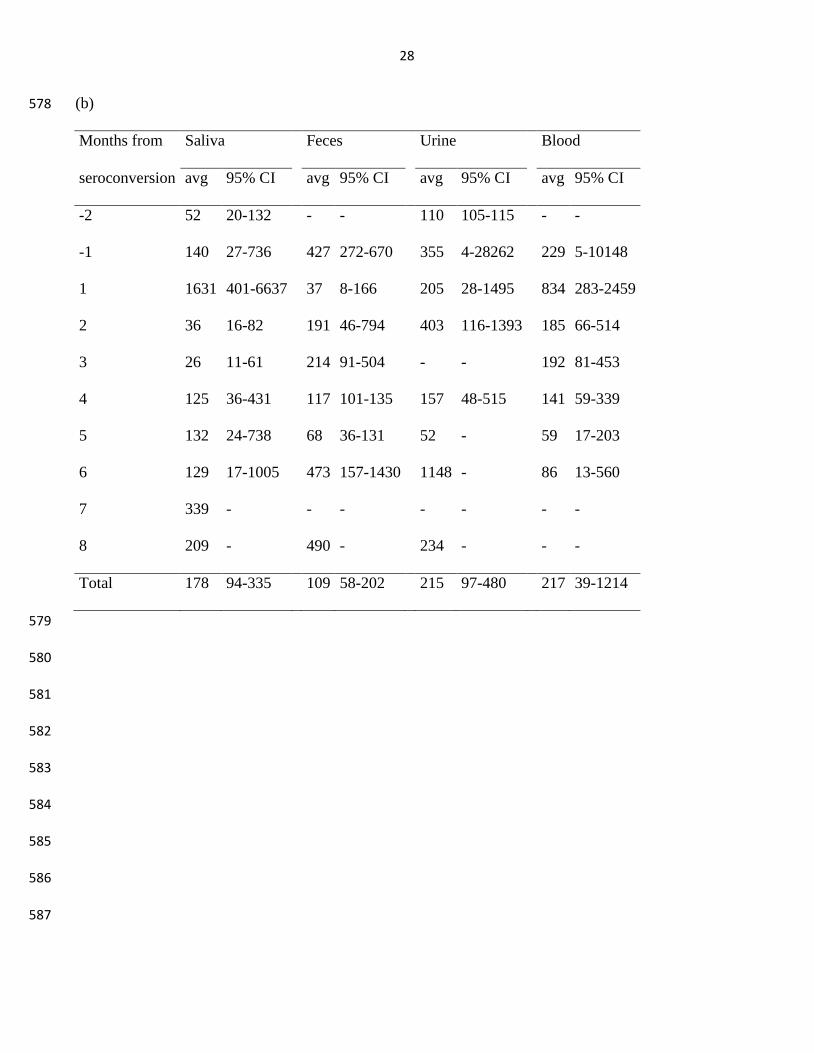
28
(b) 578
Months from Saliva
Feces
Urine
Blood
seroconversion avg 95% CI
avg 95% CI
avg 95% CI
avg 95% CI
-2 52 20-132
- -
110 105-115
- -
-1 140 27-736
427 272-670
355 4-28262
229 5-10148
1 1631 401-6637
37 8-166
205 28-1495
834 283-2459
2 36 16-82
191 46-794
403 116-1393
185 66-514
3 26 11-61
214 91-504
- -
192 81-453
4 125 36-431
117 101-135
157 48-515
141 59-339
5 132 24-738
68 36-131
52 -
59 17-203
6 129 17-1005
473 157-1430
1148 -
86 13-560
7 339 -
- -
- -
- -
8 209 -
490 -
234 -
- -
Total 178 94-335
109 58-202
215 97-480
217 39-1214
579
580
581
582
583
584
585
586
587

29
Table 2. AICc (Akaike's information criteria adjusted for sample size) scores of all candidate models 588
analyzing the probability and quantity of PUUV RNA shedding and viremia in bank voles. 589
Dependent variable
Candidate model Probability Quantity
Shedding Separate time smoothers for each excretion + separate
intercepts for each sex
315.3 264.7
Separate time smoothers for each sex + separate intercepts
for each excretion
309.6 268.5
One time smoother + separate intercepts for each excretion
and sex
300.8 264.0
Separate time smoothers for each excretion 313.8 261.2
Separate time smoothers for each sex 320.9 262.5
One time smoother + separate intercepts for each excretion 299.3* 260.2
One time smoother + separate intercepts for each sex 312.7 256.5
Separate intercepts for each excretion and sex 308.1 255.2
One time smoother 311.2 253.3
Separate intercepts for each excretion 306.5 251.8
Separate intercepts for each sex 318.2 249.7
Constant quantity through time, excreta and sexes 316.7 246.4†
Viremia Separate time smoothers for each sex 146.3 97.1
One time smoother + separate intercepts for each sex 138.0 91.0
One time smoother 135.9‡
88.7§

30
Separate intercepts for each sex 157.1 92.9
Constant quantity through time and sexes 155.0 90.4
“Time smoother” refers to a GAMM smooth term of time (days) to/from PUUV seroconversion. 590
The AICc scores of the best supported models are written in bold. 591
Variance attributable to random individual-level variation in the best supported models: *0.64; †0.05; 592
‡0.45; §0.14. 593
594
Figure Legends 595
596
Figure 1. The presence and relative quantities of PUUV RNA in sequential samples of saliva (blue), 597
feces (black) urine (yellow), and blood (red lines) of individual bank voles in relation to 598
seroconversion. Due to uncertainty in the exact time of seroconversion in wild-caught rodents, intervals 599
of approximated time from seroconversion are shown by lines instead of points. The line width 600
illustrates the log10 relative quantity of PUUV RNA. Gray lines indicate samples where no PUUV 601
RNA was detected and dashed lines RT-PCR positive samples where RNA quantitation was not 602
performed. Letters M and F refer to males and females. 603
604
Figure 2. The probabilities of bank vole excreta and blood being PUUV RNA positive (left panel) and 605
log10-transformed relative PUUV RNA quantities in RT-PCR positive samples (right panel) in relation 606
to the estimated time of seroconversion. Solid lines indicate the model predictions and dashed lines 607
their 95% confidence intervals. Circles denote the observed data. Circle sizes in the left panel indicate 608
the number of overlapping observations (rounded to the nearest 10 days for visual clarity). 609
610
FigureClick here to download high resolution image
FigureClick here to download high resolution image

Supplementary Material FilesClick here to download Supplementary Material Files: Supplementary_material.pdf




















