
Characterisation of the R276A gain-of-function mutation in the ectodomain of murine P2X7

© 2007 The Authors
Genes to Cells (2007)
12
, 329–343
Journal compilation © 2007 by the Molecular Biology Society of Japan/Blackwell Publishing Ltd.
329
DOI: 10.1111/j.1365-2443.2007.01060.x
Blackwell Publishing IncMalden, USAGTCGenes to Cells1356-9597© Blackwell Publishing Ltd? 2007123Original ArticleEctodomain shedding by meltrin
β
(ADAM19) at the Golgi apparatusT Yokozeki et al.
Meltrin ββββ
(ADAM19) mediates ectodomain shedding of Neuregulin ββββ
1 in the Golgi apparatus: fluorescence correlation spectroscopic observation of the dynamics of ectodomain shedding in living cells
Tomoichi Yokozeki
1
, Shuji Wakatsuki
1
, Kiyotaka Hatsuzawa
2
, Roy A. Black
3
, Ikuo Wada
2,
* and Atsuko Sehara-Fujisawa
1,
*
1
Department of Growth Regulation, Institute for Frontier Medical Sciences, Kyoto University, Kyoto 606-8507, Japan
2
Institute of Biomedical Sciences, Fukushima Medical University School of Medicine, Fukushima 960-1295, Japan
3
Amgen, 1201 Amgen Court West, Seattle, WA 98119-3105, USA
Membrane-anchored Neuregulin ββββ
1 sheds its ectodomain as soluble factors. Two proteases thatbelong to a disintegrin and metalloprotease (ADAM) family are known to cleave Neuregulin ββββ
1.One is tumor necrosis factor-αααα
converting enzyme (TACE/ADAM17). The other is Meltrin ββββ
(ADAM19). Against our expectation that shedding by ADAM proteases occurs at the cell surface,here we found that Meltrin ββββ
mediates the ectodomain shedding of Neuregulin ββββ
1 in the Golgiapparatus. Meltrin ββββ
was localized in and around the Golgi apparatus in developing sensory neurons.Subcellular fractionation revealed that Meltrin ββββ
generated soluble Neuregulin ββββ
1 in Golgi-enrichedfractions while TACE-cleaved Neuregulin ββββ
1 was recovered in lighter fractions. To examine whetherMeltrin ββββ
-mediated ectodomain shedding occurs in the Golgi apparatus in living cells, we tookadvantage of different diffusion properties of cleavage products from those of membrane-anchoredprecursor proteins. Fluorescence correlation spectroscopy (FCS) is the most sensitive method todetermine milli∼∼∼∼
submillisecond diffusion
in vivo.
Protease-active Meltrin ββββ
caused a shift in auto-correlation function in FCS of green fluorescent protein (GFP)-tagged Neuregulin ββββ
1 in the Golgiapparatus, suggesting a conversion of Neuregulin ββββ
1 molecules from membrane-anchored to solubleforms in that organelle. The Golgi apparatus is a site of processing Neuregulin ββββ
1 by Meltrin ββββ
.
Introduction
Various intercellular signaling and adhesion moleculesgovern cell–cell interactions during the developmentof multicellular organisms. The actions of these moleculesare regulated not only by transcriptional and translationalcontrols but also by post-translational modifications suchas phosphorylation and proteolytic processing. The latterprocess includes ectodomain shedding, which mayactivate or inactivate the functions of a membraneprotein or substantially change its functional properties(Wong
et al
. 1989; Hooper
et al
. 1997; Tanaka
et al
. 1998;Yamazaki
et al
. 2003). Although ectodomain sheddingof numerous membrane proteins and proteases that act
as “sheddases” has been reported, little is known aboutthe exact subcellular sites of ectodomain shedding and,to our knowledge, the dynamic process of ectodomainshedding in living cells has never been observed.
Membrane anchored Neuregulin is one of membraneproteins that are subject to the ectodomain shedding.Neuregulin is a multifunctional growth factor that playsessential roles in the development of the heart andnervous system (Meyer & Birchmeier 1995; Falls 2003).The core domain of the protein has an epidermal growthfactor (EGF) motif and functions as a ligand of hetero-dimeric ErbB receptors. Although some of Neuregulinisoforms are soluble proteins, the majority are mem-brane anchored, including beta-exon-containing type-INeuregulin-1a (NRG
β
1). As was reported with othermembrane proteins, phorbol 12-myristate 13-acetate(PMA) treatment of cells results in the immediate dis-appearance of the ectodomain of Neuregulin 1 from the
Communicated by
:
Yo-ichi Nabeshima*
Correspondence
:
E-mail: [email protected]; [email protected]

T Yokozeki
et al.
Genes to Cells (2007)
12
, 329–343
© 2007 The AuthorsJournal compilation © 2007 by the Molecular Biology Society of Japan/Blackwell Publishing Ltd.
330
cell surface. The ectodomain shedding of Neuregulin 1also occurs in intracellular compartments. However, theexact subcellular sites of these proteolytic events have notbeen determined (Burgess
et al
. 1995).Two proteases that belong to a disintegrin and metallo-
proteases (ADAM) family are known to participate inthe ectodomain shedding of NRG
β
1. One is the tumornecrosis factor-
α
converting enzyme (TACE/ADAM17),which participates in cleavage of NRG
β
1 upon PMAtreatment of cells. The other is Meltrin
β
(ADAM19),which exerts the ectodomain shedding of NRG
β
1 ina manner dependent on its protease activity (Shirakabe
et al
. 2001; Wakatsuki
et al
. 2004).ADAM family proteases are involved in the ectodomain
shedding of many membrane and extracellular matrixproteins. However, whether individual ADAM proteaseshave strict substrate specificities or whether some ofthem share substrates is not clarified yet. Instead, recentstudies showed that these proteases are apparently distinctfrom each other in terms of their mechanisms of regulationof ectodomain shedding. For example, TACE cleaves theectodomains of various membrane proteins upon PMAtreatment of cells (Peschon
et al
. 1998; Montero
et al
. 2000;Hinkle
et al
. 2004; Horiuchi
et al
. 2005). Kuzbanian/ADAM10 cleaves the ectodomains of various membraneproteins in response to calcium influx (Nagano
et al
. 2004;Reiss
et al
. 2005). Significant differences among ADAMproteases have been also reported in terms of the sub-cellular compartments in which they cleave theectodomain. The activities of TACE and Kuzbanian areenhanced when the cholesterol-rich microdomaincalled the membrane raft is disrupted (Kojro
et al
.2001; Matthews
et al
. 2003; Zimina
et al
. 2005), whileMeltrin
β
is located in the raft fraction together withNRG
β
1, which is required for ectodomain sheddingof the latter (Wakatsuki
et al
. 2004). Moreover, TACEand Kuzbanian cause disappearance of the ectodomainof CD44 at focal contacts and at the rear ends of sometypes of moving cells, respectively, suggesting differentsubcellular sites of ectodomain shedding between thesetwo (Nagano
et al
. 2004). These results raise a possibilitythat each ADAM protease meets and cleaves its substrateat distinct subcellular regions among each other.Determination of exact sites of ectodomain sheddingby individual proteases is meaningful, assuming that sub-cellular sites of growth factor generation could regulateintercellular signaling events temporally and spatially.
On the basis of these previous studies, we aimed toidentify subcellular sites at which Meltrin
β
mediates thecleavage of NRG
β
1. However, determination of sites ofectodomain shedding may not be easy. The major sub-cellular locations of each ADAM protease are not always
in accord with the sites of ectodomain shedding; that is,the disappearance of ectodomain at focal contacts doesnot necessarily mean that proteolysis occurs at the cellsurface at focal contacts. First of all, anterograde andretrograde intracellular traffics of the protease, its sub-strate precursors and cleavage products should be takeninto consideration. Some ADAM proteins, such as TACEand Meltrin
α
/ADAM12, change their localizations inresponse to certain stimuli, making the sites of ectodo-main shedding difficult to determine (Itai
et al
. 2001;Matthews
et al
. 2003). Whether the protease is expressedas an active form at particular intracellular sites should bealso examined.
To overcome such difficulties and to determine theexact sites of cleavage mediated by Meltrin
β
, we setout to determine them in living cells. For this purpose,we applied fluorescence correlation spectroscopy (FCS)to living cells to monitor the ectodomain sheddingof green fluorescent protein (GFP)-tagged NRG
β
1molecules. FCS is the most sensitive method to determinemilli
∼
submillisecond diffusion in an area as small as sub-femtoliter confocal volume
in vivo.
As a result, against ourexpectation that shedding by ADAM proteases occurs atthe cell surface in general, we could identify the Golgiapparatus as a site where Meltrin
β
mediates proteolyticprocessing of Neuregulin. Meltrin
β
was localized in andaround the Golgi apparatus in developing sensory neurons.Subcellular fractionation substantiated the result of FCS.We discuss differential regulation of growth factors byectodomain shedding both in the Golgi apparatus and inother subcellular regions (such as at the cell surface).Effective application of FCS in the study of ectodomainshedding is also discussed.
Results
Meltrin ββββ
is localized in the Golgi apparatus and its immediate vicinity
Meltrin
β
is expressed abundantly in the peripheralnervous system and in the heart during its development(Kurisaki
et al
. 1998; Kurohara
et al
. 2004). To determinethe subcellular localization of Meltrin
β
, neurons preparedfrom developing dorsal root ganglia of mouse embryosat embryonic day 16.5 were immunostained with anti-bodies specific for Meltrin
β
, Golgi 58K or
β
COP.Both Golgi 58K (forminotransferase cyclodeaminase)and
β
COP (the beta subunit of a coatomer complex) areGolgi markers. Meltrin
β
was found in intracellularvesicles (Fig. 1). Some of these vesicles were Golgi 58K-or
β
COP-positive, whereas others were not but were inthe vicinity of Golgi 58K-positive or
β
COP-positive

Ectodomain shedding by meltrin
β
(ADAM19) at the Golgi apparatus
© 2007 The Authors
Genes to Cells (2007)
12
, 329–343
Journal compilation © 2007 by the Molecular Biology Society of Japan/Blackwell Publishing Ltd.
331
vesicles. Meltrin
β
was undetectable at the cell surfaceand on Transferrin receptor-positive endosomes (datanot shown). These results indicate that Meltrin
β
waslocalized mainly in the Golgi apparatus.
Golgi-enriched fractions prepared from Meltrin ββββ
-expressing cells contain shed, soluble NRG ββββ
1
Previously, we showed that co-expression of Meltrin
βwith NRG β1 markedly enhanced the ectodomainshedding of NRG β1 (Shirakabe et al. 2001). In addi-tion, the Neuregulin1 gene and the Meltrin β gene areactively transcribed and translated in developing neuronsat similar stages (Meyer et al. 1997; Shirakabe et al. 2001).We thus examined whether the Golgi apparatus is thesite of ectodomain shedding of NRG β1 by Meltrin β.If Meltrin β cleaves NRG β1 in the Golgi apparatus aftertranslation in the endoplasmic reticulum (ER), releasedNRG β1 should be found in the former organelle. On thebasis of this idea, NRG β1 with a hemagglutinin peptide-
tag at its N-terminus (HA-NRG β1) was produced inthe cells and the intracellular distribution of membrane-bound and soluble HA-NRG β1 was determined.
Post-nuclear supernatants of N1E115 neuroblastsand COS7 cells, which expressed HA-NRG β1 togetherwith wild-type Meltrin β (WT), empty vector or Meltrinβ (EQ), a mutant of Meltrin β that has a single aminoacid substitution in the active site of the protease domain,were separated by Nycodenz (Histodenz) gradient cen-trifugation. Each fraction was subjected to immuno-blotting using antibodies against HA, ERp72 (as an ERmarker) or Golgi 58K. ERp72 and Golgi 58K weremainly contained in fractions 6–10 and in fractions 1–5,respectively (Figs. 2 and 3). Golgi 58K(+)ERp72(–)and Golgi 58K(–)ERp72(+) fractions were assessed asGolgi- and ER-enriched fractions, respectively. Fraction6, which often showed Golgi 58K(+)ERp72(+), wasconsidered as a transient or intermediate fraction betweenthe ER and the Golgi apparatus. The bottom Golgi58K(+)ERp72(+) fraction (fraction 10) was assumed
Figure 1 Confocal laser microscopeimages of developing neurons preparedfrom mouse dorsal root ganglia at E16.5immunostained with anti-Meltrin β(red), together with Golgi 58K or β COP(green). Nuclei are indicated by the letterN in dashed circles. Scale bar = 10 µm.Many of the Meltrin β (+) vesicles wereco-localized with Golgi markers, andothers were not co-localized but were nearthe markers.

T Yokozeki et al.
Genes to Cells (2007) 12, 329–343 © 2007 The AuthorsJournal compilation © 2007 by the Molecular Biology Society of Japan/Blackwell Publishing Ltd.
332
to contain cell debris. The top Golgi 58K(+)ERp72(+)fractions (fractions 1 and 2) might include lighter mem-brane fractions such as endosomes, secretory vesiclesand particulated plasma membrane, as well as cytosoliccomponents. In NIE115 neuroblasts, high levels ofendogenous Meltrin β were produced. Although endo-genous or exogenously introduced Meltrin β was foundnot only in the Golgi fractions but also in ER fractions,prodomain-excised, active forms of Meltrin β were foundin the Golgi 58K(+) (fractions 1–5) and intermediate(fraction 6) fractions. The majority of NRG β1 mole-cules in these fractions were soluble forms not only inthe exogenous Meltrin β-treated cells but also in emptyvector-treated cells (Fig. 2A,C). In contrast, expressionof Meltrin β (EQ) resulted in the suppression of ectodo-main shedding in Golgi-enriched and intermediate(fractions 3–6) fractions (Fig. 2B), while fractions 1 and2 contained soluble NRG β1. To exclude the possibility
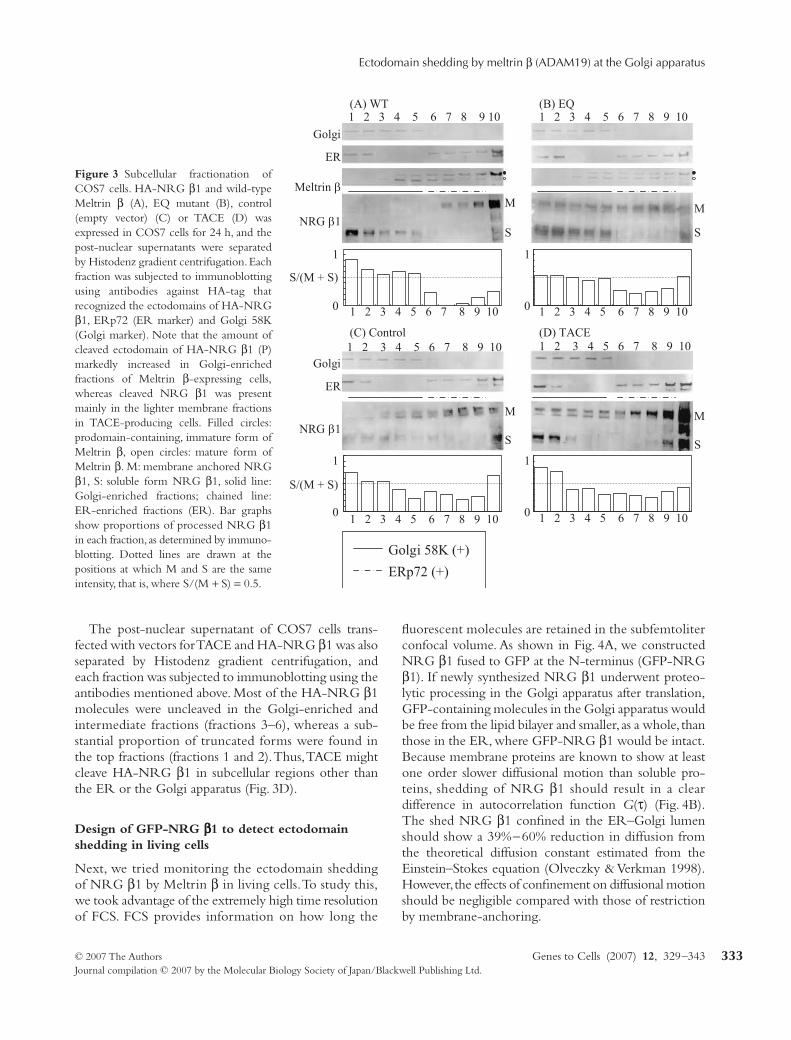
that ectodomain shedding with Meltrin β was mediatedby TACE, the post-nuclear supernatant of TACE protease-deficient (TACE–/–) cells transfected with HA-NRG β1together with Meltrin β (WT) or with Meltrin β (EQ)was analysed similarly. Ectodomain shedding of HA-NRG β1 was enhanced by Meltrin β (WT) in the Golgiapparatus in a TACE-independent manner (Fig. 2D).Similar results were obtained in COS7 cells. Golgi58K(+) (fractions 1–5) fractions of the cells transfectedwith Meltrin β (WT) exclusively contained shed, solubleNRG β1, whereas ER-enriched fractions (fractions7–9) of these cells contained only uncleaved molecules.In contrast, a substantial proportion of NRG β1 remaineduncleaved in Golgi 58K(+) fractions of the cells trans-fected with empty vector or Meltrin β (EQ) vector (Fig. 3).Taken together, these results suggested that Meltrin βgenerated soluble NRG β1 in the Golgi apparatus,which was delivered to the lighter membrane fractions.
Figure 2 (A–C) Subcellular fractionationof NIE115 neuroblasts. HA-NRG β1and wild-type Meltrin β (A), EQ mutant(B), or control (empty vector) (C) wereexpressed in N1E115 neuroblasts for 24 h,and the post-nuclear supernatants of theN1E115 cells were separated by Histodenzgradient centrifugation. Each fractionwas subjected to immunoblotting usingantibodies against HA, ERp72 and Golgi58K. Note that the amount of ectodomainof HA-NRG β1 was markedly increasedin the Golgi-enriched fractions ofN1E115 cells (C) that express endogenousMeltrin β. (D) Subcellular fractionationof TACE(–/–) mouse embryonic fibroblastsafter production of HA-NRG β1 togetherwith Meltrin β or Meltrin β (EQ). Theeffect of Meltrin β did not require TACEprotease activity. Filled circles: immatureform of Meltrin β, open circles: mature formof Meltrin β. M: membrane anchoredNRG β1, S: soluble form NRG β1, solidline: Golgi-enriched fractions; chained line:ER-enriched fractions. Bar graphs showproportions of processed NRG β1 in eachfraction (described in caption of Fig. 2).
Ectodomain shedding by meltrin β (ADAM19) at the Golgi apparatus
© 2007 The Authors Genes to Cells (2007) 12, 329–343Journal compilation © 2007 by the Molecular Biology Society of Japan/Blackwell Publishing Ltd.
333
The post-nuclear supernatant of COS7 cells trans-fected with vectors for TACE and HA-NRG β1 was alsoseparated by Histodenz gradient centrifugation, andeach fraction was subjected to immunoblotting using theantibodies mentioned above. Most of the HA-NRG β1molecules were uncleaved in the Golgi-enriched andintermediate fractions (fractions 3–6), whereas a sub-stantial proportion of truncated forms were found inthe top fractions (fractions 1 and 2). Thus, TACE mightcleave HA-NRG β1 in subcellular regions other thanthe ER or the Golgi apparatus (Fig. 3D).
Design of GFP-NRG ββββ1 to detect ectodomain shedding in living cells
Next, we tried monitoring the ectodomain sheddingof NRG β1 by Meltrin β in living cells. To study this,we took advantage of the extremely high time resolutionof FCS. FCS provides information on how long the
fluorescent molecules are retained in the subfemtoliterconfocal volume. As shown in Fig. 4A, we constructedNRG β1 fused to GFP at the N-terminus (GFP-NRGβ1). If newly synthesized NRG β1 underwent proteo-lytic processing in the Golgi apparatus after translation,GFP-containing molecules in the Golgi apparatus wouldbe free from the lipid bilayer and smaller, as a whole, thanthose in the ER, where GFP-NRG β1 would be intact.Because membrane proteins are known to show at leastone order slower diffusional motion than soluble pro-teins, shedding of NRG β1 should result in a cleardifference in autocorrelation function G(τ) (Fig. 4B).The shed NRG β1 confined in the ER–Golgi lumenshould show a 39%−60% reduction in diffusion fromthe theoretical diffusion constant estimated from theEinstein–Stokes equation (Olveczky & Verkman 1998).However, the effects of confinement on diffusional motionshould be negligible compared with those of restrictionby membrane-anchoring.
Figure 3 Subcellular fractionation ofCOS7 cells. HA-NRG β1 and wild-typeMeltrin β (A), EQ mutant (B), control(empty vector) (C) or TACE (D) wasexpressed in COS7 cells for 24 h, and thepost-nuclear supernatants were separatedby Histodenz gradient centrifugation. Eachfraction was subjected to immunoblottingusing antibodies against HA-tag thatrecognized the ectodomains of HA-NRGβ1, ERp72 (ER marker) and Golgi 58K(Golgi marker). Note that the amount ofcleaved ectodomain of HA-NRG β1 (P)markedly increased in Golgi-enrichedfractions of Meltrin β-expressing cells,whereas cleaved NRG β1 was presentmainly in the lighter membrane fractionsin TACE-producing cells. Filled circles:prodomain-containing, immature form ofMeltrin β, open circles: mature form ofMeltrin β. M: membrane anchored NRGβ1, S: soluble form NRG β1, solid line:Golgi-enriched fractions; chained line:ER-enriched fractions (ER). Bar graphsshow proportions of processed NRG β1in each fraction, as determined by immuno-blotting. Dotted lines are drawn at thepositions at which M and S are the sameintensity, that is, where S/(M + S) = 0.5.

T Yokozeki et al.
Genes to Cells (2007) 12, 329–343 © 2007 The AuthorsJournal compilation © 2007 by the Molecular Biology Society of Japan/Blackwell Publishing Ltd.
334
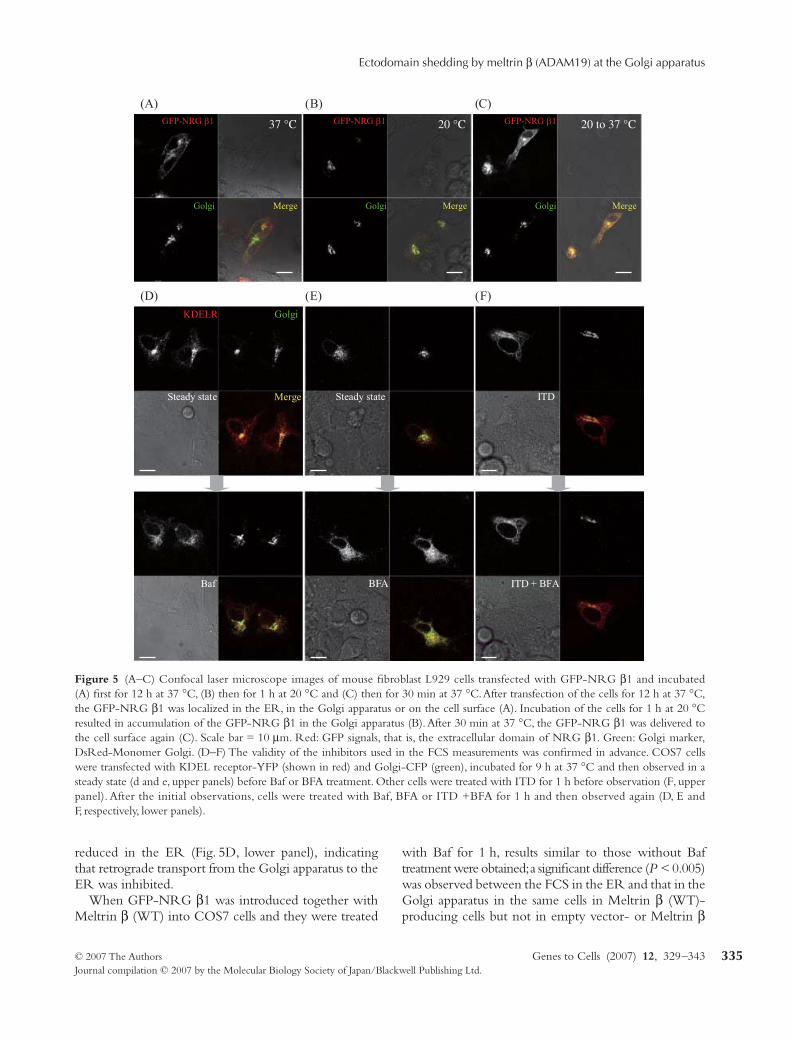
We first determined whether nascent GFP-NRG β1could be trapped early in the secretory pathway andreach the cell surface properly in living cells. GFP-NRGβ1 was expressed in mouse fibroblast L929 cells for 12 hat 37 °C and was observed by confocal microscopy.GFP-NRG β1 was found to be localized at the cellsurface, in the Golgi apparatus and in the ER. Whenthe incubation temperature was subsequently shifted to20 °C for 1 h, GFP-NRG β1 accumulated mainly in theGolgi apparatus. When the incubation temperatureof the cells was then shifted back to 37 °C for 30 min,GFP-NRG β1 appeared at the cell surface again.Thus, newly synthesized GFP-NRG β1 was trans-ported properly (Fig. 5A–C). The ectodomain sheddingof GFP-NRG β1 mediated by Meltrin β generatedN- and C-terminal fragments of the expected sizes(data not shown).
FCS measurements reveal that Meltrin ββββ mediates ectodomain shedding of GFP-NRG ββββ1 in the Golgi apparatus in living cells
GFP-NRG β1 vector was introduced together withMeltrin β (WT) vector into COS7 cells using the glass
microbead method and incubated for 2.5 h. A significantdifference (P < 0.01) was observed between the FCS inthe ER and that in the Golgi apparatus in the same cells(Fig. 6, steady state). A higher proportion of rapid diffu-sional motion was detected in the Golgi apparatus thanin the ER, suggesting that membrane-anchored GFP-NRG β1 molecules were cleaved and showed fasterdiffusional motion in the Golgi apparatus than in theER (Fig. S1, steady state, WT; a clear difference in auto-correlation function G(τ) between ER and Golgi). Incontrast, co-transfection of empty vector or Meltrin β(EQ)vector did not produce such differential composi-tion of diffusion or FCS autocorrelation curves for thetwo organelles (Fig. S1, steady state, EQ and vector).
To check the reliability of these FCS measurements,we compared the autocorrelation functions and the best-fit two-component simulation profiles (Fig. S2). Thegood fit between the experimental and theoretical valuesindicates that the autocorrelation functions of these datawere reliable, assuming that there were two main mobilecomponents (rapidly diffusing and slowly diffusing).According to previous observations on the diffusion ofsoluble and membrane-anchored proteins (Saito et al.2003), the diffusion constants of membrane proteins aregenerally one order lower than the soluble forms. In thisstudy, because most of FCS data were best-fitted withthe two components model in which the fast componentwas ∼300 µs and the slow was ∼12 000 µs, we assumedthat there are two populations whose diffusional motionis distinct.
The calculated proportions of the rapidly and slowlydiffusing components, which would represent soluble(cleaved) forms and membrane-bound (not cleaved) formsof NRG β1, respectively, are shown in Fig. 6 (steady state).
Although the glass microbead method enabled us tomonitor GFP-NRG β1 in 2–3 h after transfection,retrograde transport might bring intact or cleavedGFP-NRG β1 from the cell surface or the trans-Golginetwork to the Golgi apparatus or from the Golgiapparatus to the ER. To avoid contribution of theseretrograde transports, we examined the effect of Bafilo-mycin A1 (Baf). Baf inactivates endosomes and inhibitsretrograde transport from the Golgi apparatus to the ER(Palokangas et al. 1998), enabling us to use FCS to detectnewly synthesized NRG β1 and NRG β1 carried fromthe ER to the Golgi apparatus by anterograde transport.We confirmed the effects of this reagent on the retrogradetransport from the Golgi apparatus to the ER by analysingthe distribution of the YFP-tagged KDEL receptor intro-duced together with CFP-Golgi (Fig. 5D). When COS7cells were treated with Baf (1 µm final concentration) for1 h, the amount of KDEL receptor molecules was
Figure 4 (A and B) Detection of ectodomain shedding by FCSmeasurements of Brownian motion of GFP-tagged NRG β1.(A) Cleavage of GFP-NRG β1 by Meltrin β frees the GFP-taggedextracellular domain from the lipid bilayer; the cleaved domaindiffuses more rapidly than the membrane-bound precursor form.(B) The change in mobility of the molecule due to the releasefrom the lipid bilayer and decrease in molecular weight is reflectedin a shift in the autocorrelation function, G(τ).
Ectodomain shedding by meltrin β (ADAM19) at the Golgi apparatus
© 2007 The Authors Genes to Cells (2007) 12, 329–343Journal compilation © 2007 by the Molecular Biology Society of Japan/Blackwell Publishing Ltd.
335
reduced in the ER (Fig. 5D, lower panel), indicatingthat retrograde transport from the Golgi apparatus to theER was inhibited.
When GFP-NRG β1 was introduced together withMeltrin β (WT) into COS7 cells and they were treated
with Baf for 1 h, results similar to those without Baftreatment were obtained; a significant difference (P < 0.005)was observed between the FCS in the ER and that in theGolgi apparatus in the same cells in Meltrin β (WT)-producing cells but not in empty vector- or Meltrin β
Figure 5 (A–C) Confocal laser microscope images of mouse fibroblast L929 cells transfected with GFP-NRG β1 and incubated(A) first for 12 h at 37 °C, (B) then for 1 h at 20 °C and (C) then for 30 min at 37 °C. After transfection of the cells for 12 h at 37 °C,the GFP-NRG β1 was localized in the ER, in the Golgi apparatus or on the cell surface (A). Incubation of the cells for 1 h at 20 °Cresulted in accumulation of the GFP-NRG β1 in the Golgi apparatus (B). After 30 min at 37 °C, the GFP-NRG β1 was delivered tothe cell surface again (C). Scale bar = 10 µm. Red: GFP signals, that is, the extracellular domain of NRG β1. Green: Golgi marker,DsRed-Monomer Golgi. (D–F) The validity of the inhibitors used in the FCS measurements was confirmed in advance. COS7 cellswere transfected with KDEL receptor-YFP (shown in red) and Golgi-CFP (green), incubated for 9 h at 37 °C and then observed in asteady state (d and e, upper panels) before Baf or BFA treatment. Other cells were treated with ITD for 1 h before observation (F, upperpanel). After the initial observations, cells were treated with Baf, BFA or ITD +BFA for 1 h and then observed again (D, E andF, respectively, lower panels).

T Yokozeki et al.
Genes to Cells (2007) 12, 329–343 © 2007 The AuthorsJournal compilation © 2007 by the Molecular Biology Society of Japan/Blackwell Publishing Ltd.
336
Figure 6 Summary of FCS measurements. The ratios of molecules with fast (cross-hatching in bars) and slow (clear areas of bars) diffusiontimes were estimated from each FCS measurement (Fig. S1). The fast and slow diffusion times were assessed to be 300 and 12 000 µs,respectively, and were, respectively, typical of free and membrane-bound proteins. Mean values ± SE of estimated diffusion times arepresented.

Ectodomain shedding by meltrin β (ADAM19) at the Golgi apparatus
© 2007 The Authors Genes to Cells (2007) 12, 329–343Journal compilation © 2007 by the Molecular Biology Society of Japan/Blackwell Publishing Ltd.
337
(EQ)-introduced cells (Fig. 6, Baf treatment; Fig. S1,Baf treatment). Thus, these measurements furtherconfirmed that cleavage of GFP-NRG β1 dependent onMeltrin β occurred in the Golgi apparatus.
To verify our hypothesis in another way, we used cellstreated with BFA for 1 h. In cells treated with BFA for1 h, anterograde transport from the ER to the Golgiapparatus is inhibited and part of the Golgi cisterna isfused to the ER (Schweizer et al. 1988; Klausner et al.1992). If GFP-NRG β1 is shed in the Golgi apparatusby Meltrin β, rapid diffusional motion would be gener-ated in the ER by the fusion of the Golgi apparatus withBFA treatment. We confirmed the effects of BFA byanalysing the distribution of the YFP-tagged KDELreceptor introduced together with CFP-Golgi. COS7cells treated with BFA (25 µm final concentration) for1 h showed fusion of the Golgi apparatus and the ER,confirming the effect of the BFA (Fig. 5E). GFP-NRGβ1, together with Meltrin β (WT), empty vector orMeltrin β (EQ), was expressed in the COS7 cells asabove and diffusional motion in the ER was observed byFCS. After the initial measurement, cells were treatedwith BFA for 1 h and diffusional motion in the ER of thesame cells was observed again (Fig. 6, BFA treatment;Fig. S1, BFA treatment). Treatment of cells that expressedMeltrin β (WT) with BFA resulted in the appearanceof a higher proportion of molecules with fast diffusionalmotion than in steady-state cells. Because BFA causesfusion of the Golgi apparatus and the ER, we interpretedthese results as evidence of enhanced movement ofsoluble GFP-NRG β1 generated in the Golgi apparatusor of enhanced movement of active Meltrin β into theER or both, resulting in an increased number of solubleGFP-NRG β1 molecules in the fused ER.
To obtain further evidence of the cleavage of GFP-NRG β1 in the Golgi apparatus, we exploited isotetrandrine(ITD), an inhibitor of BFA-induced tubule formationfrom the Golgi complex and retrograde trafficking tothe ER (Chan et al. 2004). Cells treated with ITD showinhibition of the transport of proteins from the Golgiapparatus to the ER induced by BFA. If the increase indiffusional motion in the ER of cells treated with BFAwere caused by the fusion of part of the Golgi cisternato the ER, ITD would inhibit this increase. To checkthe effect of ITD, COS7 cells were treated first with ITD(20 µm final concentration) for 1 h and then with ITDand BFA (25 µm final concentration) for an additional1 h. This treatment inhibited fusion of the Golgi apparatusand the ER (compare Fig. 5E,F). GFP-NRG β1together with Meltrin β (WT), empty vector or Meltrinβ (EQ) was expressed in COS7 cells as above; the cellswere cultured for 1 h in medium containing ITD and
then diffusional motion in the ER was observed by FCS.After the initial observation, cells were treated with BFAand ITD and incubated for 1 h, and diffusional motionin the ER of the same cells was observed (Fig. S1, ITD +BFA). A much smaller proportion of rapid diffusionalmotion was observed in the ITD- and BFA-treated cellsexpressing Meltrin β (WT) than in the cells treatedwith BFA only (Fig. 6, ITD + BFA treatment). The FCSmeasurements in the Golgi apparatus of the cells treatedwith ITD and BFA are shown in Fig. S4. Resultsessentially opposite to those in the ER were obtained.That is, the fast population increased only in the WTMeltrin β-expressing cells treated with ITD + BFA,suggesting that ITD caused inhibition of constitutiverecycling between the ER and Golgi in addition to theblockage of BFA-induced redistribution of Golgi com-ponents. The distribution of Meltrin β treated with BFA,ITD or ITD + BFA was monitored by ligating GFP-tagat the C-terminus in Fig. S3. Meltrin β-GFP was mainlylocalized in the Golgi apparatus and its vicinity. Meltrinβ-GFP redistributed in the ER together with Ds-Red-Golgi-marker after the treatment of BFA, which wasfurther blocked by the cotreatment of ITD. These resultsindicate that rapidly diffusing molecules appeared in theER when the Golgi components of Meltrin β (WT)-producing cells, including matured Meltrin β itself, weredelivered into the ER by treatment with BFA.
DiscussionWe identified the Golgi apparatus as one of the sub-cellular sites of ADAM protease-mediated ectodomainshedding of membrane-anchored ErbB ligands bymonitoring the ligands in living cells for the first time.Although this study does not exclude roles of Meltrin βin other subcellular regions, we consider the Golgiapparatus to be one of the major subcellular sites that thisprotease acts in judged from its expression patterns in cells.
Although members of the ADAM family proteases areknown to be key players in the ectodomain shedding ofvarious membrane-anchored growth factors and adhe-sion proteins, it has been elusive at which subcellular sitesADAM proteases cleave these membrane proteins. In thecase of Neuregulin 1, a number of researchers have roughlyidentified the subcellular regions where its membrane-anchored forms are proteolytically processed. However,determination of the exact subcellular sites of ectodomainshedding from experiments with cell lysates or immuno-staining of fixed cells alone is extremely difficult.Although pulse-chase analysis can indicate whether thecleaved ectodomains accumulate inside the cell or howfast they are released in the culture medium, complicated

T Yokozeki et al.
Genes to Cells (2007) 12, 329–343 © 2007 The AuthorsJournal compilation © 2007 by the Molecular Biology Society of Japan/Blackwell Publishing Ltd.
338
anterograde and retrograde sorting of intracellular vesiclesmakes the subcellular regions of cleavage difficult todetermine (Burgess et al. 1995; Loeb et al. 1998). Inaddition, the subcellular sites of protease-mediatedectodomain shedding cannot be specified simply fromthe distribution of proteases in cells. The finding thatMeltrin β was localized mainly in the Golgi apparatus ledus to expect that this protease acts on the substrate in theGolgi apparatus. The protease, however, could be in a latentform or it could be physically separated from its substratesor activators there. For example, ADAM proteases haveproprotein domains, and proteolytic removal of thesedomains by furin-like proteases is required for proteaseactivation (Schlondorff et al. 2000). For these reasons,we considered that monitoring the cleavage of substratesin living cells was essential for determining the subcellularsites of ectodomain shedding exactly and conclusively.
We chose NRG β1 as a protease substrate in this studyfor several reasons. Evidence suggests that the ectodomainshedding of NRG β1 occurs both at the cell surfaceand in intracellular compartments (Burgess et al. 1995).Moreover, NRG β1 is reported as a substrate of twodifferent ADAM proteases. We found previously thatMeltrin β is produced in developing neurons at similarstages as Neuregulin 1 and enhances the ectodomainshedding of NRG β1 in a manner dependent on theprotease domain activity of Meltrin β (Shirakabe et al.2001; Wakatsuki et al. 2004). On the other hand, TACEis involved in the PMA-induced ectodomain sheddingof various ErbB ligands, including Neuregulin 1 (Peschonet al. 1998; Montero et al. 2000; Shirakabe et al. 2001;Hinkle et al. 2004; Blobel 2005; Horiuchi et al. 2005).Thus, we initially focused on the question of whetherMeltrin β localized in the Golgi apparatus of cells suchas the developing primary neurons was involved inintracellular ectodomain shedding of NRG β1. Then,we sought to determine whether Meltrin β and TACEshared subcellular sites of ectodomain shedding. Theanswer to the latter question is important, because thesites of ADAM protease-mediated ectodomain sheddingshould be closely associated with the biological roles ofthese proteases.
Simple confocal microscopy cannot be used toobserve the actual moment of the cleavage of N- and C-terminal-tagged proteins (such as YFP-NRG β1-CFP),because the distance between N- and C-termini isapparently below the limit of optical resolution. That is,N-terminal and C-terminal tags merges under theconfocal microscope when segregation of these tags arebelow the resolution of an optical microscope even ifthey exist in separate polypeptides as a result of thecleavage. However, there are two alternative methods
for analyzing proteolytic changes in a single molecule:fluorescence resonance energy transfer (FRET) andFCS. FRET is a powerful method for observing theinteraction of two molecules or the interaction of thedomains of a protein in living cells. The efficiency ofFRET is critical for distinguishing between interactingand non-interacting states in a molecule, and this effi-ciency depends on the distance between the twochromophores and the orientation of their spins. Forour purpose, the chromophores have to be sufficientlydistant from the proteolytic site to avoid steric hindranceof the juxtamembrane proteolytic site within NRG β1.In addition, the orientation of the spins of the twochromophores must be arranged to obtain high FRETefficiency, and this requirement seriously restricts thechoice of constructs for obtaining useful FRET results.
FCS provides information about the dynamics ofmolecular processes by means of measurements of spon-taneous microscopic fluctuations at dilute molecularconcentrations (about 102 molecules in the confocalvolume). FCS detects fluctuations of fluorescence inten-sity in a confocally defined volume with a sharplyfocused laser and can measure translational and rotationaldiffusion coefficients of molecules in solution and inliving cells (Widengren & Rigler 1998; Foldes-Papp et al.2001; Hess et al. 2002; Larson et al. 2003; Gosch et al.2004; Kamada et al. 2004; Bacia et al. 2006). AlthoughFCS is not yet commonly used, several experiments haverevealed the great potential of this method in living cells(Bacia et al. 2006; Kogure et al. 2006). In our experiment,the shed NRG β1 confined in the ER–Golgi lumenshould show a 39%−60% reduction in diffusion from thetheoretical diffusion constant estimated from the Einstein–Stokes equation (Olveczky & Verkman 1998); however,the effects of confinement on diffusional motion shouldbe negligible compared with those of restriction bymembrane-anchoring. Furthermore, although the ERand the Golgi apparatus are structurally and dynamicallydistinct compartments, their structural constraints on thediffusion of luminal proteins should be comparable, sincethe occlusion of space is comparable in both organelles(Voeltz et al. 2002). Hence, we reasoned that comparisonof the time-correlated decay curves obtained by FCSin various compartments along the secretory pathwaywould enable us to identify the site of shedding. Althoughfurther improvements are required for more precisequantitative analysis, this study clearly showed FCS as avaluable tool to assess ectodomain shedding of membraneproteins in living cells.
We fused NRG β1 molecules with GFP at the N-terminus (GFP-NRG β1) to monitor the ectodomainshedding. FCS analysis with this probe was focused on

Ectodomain shedding by meltrin β (ADAM19) at the Golgi apparatus
© 2007 The Authors Genes to Cells (2007) 12, 329–343Journal compilation © 2007 by the Molecular Biology Society of Japan/Blackwell Publishing Ltd.
339
the cleavage within and around the Golgi apparatus,where Meltrin β resides in the primary neurons duringdevelopment. The results of the FCS analysis are sum-marized in Fig. 6. The following three sets of resultsindicated that Meltrin β plays a role in ectodomainshedding in the Golgi apparatus. First, a higher propor-tion of rapidly diffusing forms of GFP-NRG β1 wasfound in the Golgi apparatus than in the ER of Meltrinβ-expressing cells. No such increase was observed by theintroduction of protease-deficient Meltrin β (EQ) orthe empty vector. Similar results were obtained withcells treated with Baf, which inactivates endosomes andinhibits retrograde transport from the Golgi apparatus tothe ER (Palokangas et al. 1998). Baf is a specific inhibitorof vacuolar-type H + ATPase. According to a previousreport, Golgi pH increases approximately 0.6 after thetreatment of Baf (Llopis et al. 1998). Because the metallo-proteases act at a relatively wide range of pH in general,a slight increase in pH may not affect protease activity ofMeltrin β significantly. We therefore interpreted the datathat the proportion of rapidly diffusing molecules slightlyincreased after Baf treatment in the Golgi apparatus ofMeltrin β (WT)-expressing cells, probably because ofthe inhibition of the retrograde transport of shed mole-cules from the Golgi apparatus to the ER. The resultsalso excluded the involvement of endosomal pathway inthis process. Second, BFA treatment caused an increasein the proportion of rapidly diffusing molecules in theER of Meltrin β-producing cells. Third, in contrast,treatment of the cells with ITD in addition to BFAresulted in a decrease in the proportion of rapidly diffus-ing molecules in the ER of Meltrin β-producing cellscompared with treatment of these cells with BFA alone.BFA inhibits anterograde transport from the ER to theGolgi apparatus but stimulates fusion of the Golgiapparatus to the ER (Schweizer et al. 1988; Klausneret al. 1992). ITD inhibits BFA-induced tubule formationfrom the Golgi complex and retrograde trafficking to theER and also inhibits BFA-stimulated tubule formationfrom the Golgi apparatus to the ER (Chan et al. 2004).We therefore interpreted the results of the latter twoexperiments as indicating that retrograde traffic ofMeltrin β from the Golgi apparatus to the ER as a resultof treatment with BFA stimulated ectodomain sheddingin the ER. BFA treatment might have also causedretrograde transport of the rapidly diffusing moleculesgenerated in the Golgi apparatus to the ER. Indeed,inhibition of BFA-induced effects was completely reversedby addition of ITD. Although ITD seems to disturb theconstitutive recycling, at least, we showed that accumu-lation of the fast component by over-expression of WTMeltrin β. Taken together, these results indicate that
Meltrin β is involved in the ectodomain shedding in theGolgi apparatus.
We obtained supporting results from Western blottingof N1E115 and COS7 cell lysates after subcellular frac-tionation by Histodenz gradient centrifugation (Figs. 2and 3). Meltrin β (WT), but not empty vector or Melt-rin β (EQ), markedly enhanced generation of the shedectodomain of NRG β1 in Golgi-enriched fractions.These biochemical data not only support the results ofFCS but also indicate that the slowly and rapidly diffus-ing components of GFP-NRG β1 observed in the FCSmeasurements did in fact correspond to GFP-NRG β1molecules before and after ectodomain shedding.Because all the experimental values of the autocorrelationfunction fitted well to the theoretical best-fit two-component simulation profile, the results indicate thereliability of the measurements in this study (Fig. S2).
In addition to determination of a subcellular site ofectodomain shedding that one of ADAM proteasesmediates, our results provided evidence that differentADAM proteases act on the same substrate at differentsubcellular regions. First, subcellular fractionationsuggested that Meltrin β and TACE cleaved the samegrowth factor at different subcellular fractions. More-over, the increase in shed molecules of NRG β1 in thepresence of Meltrin β in the Golgi fraction of TACE–/–
MEFs showed clearly that ectodomain shedding in theGolgi apparatus in Meltrin β-treated cells required theprotease activity of not TACE, but Meltrin β itself.Taken together, our results suggest that Meltrin β isinvolved in intracellular ectodomain shedding in andaround the Golgi apparatus, and that this correspondsto the constitutive ectodomain shedding reportedpreviously (Burgess et al. 1995). The ectodomain shed-ding of Neuregulin 1 is stimulated by PMA or by fetalbovine serum (Loeb et al. 1998). In our experiments,cells were cultured in 1% serum. Further investigation ofwhether ‘constitutive’ shedding in the presence of Meltrinβ requires certain factors in serum is necessary.
TACE and Meltrin β play roles in valve formationduring heart development ( Jackson et al. 2003; Kuroharaet al. 2004; Zhou et al. 2004). Double-knockout micelacking the genes encoding these two proteins showeddilation of the cardiac muscle, which was not found in therespective single-knockout mice (Horiuchi et al. 2005).Thus, these two ADAM proteases have related functionsin heart development. TACE is produced mainly in theendocardial epithelia and is considered to be involvedin the ectodomain shedding of HB-EGF, which is alsoproduced in these cells (Montero et al. 2000; Hinkleet al. 2004). Meltrin β is highly expressed not only inendocardial cell lineage but also in the cardiac neural

T Yokozeki et al.
Genes to Cells (2007) 12, 329–343 © 2007 The AuthorsJournal compilation © 2007 by the Molecular Biology Society of Japan/Blackwell Publishing Ltd.
340
crests adjacent to these epithelia. Although Neuregulin 1is one of the major ErbB ligands produced in these celllineages, further investigation is needed to determinewhether this growth factor is the physiological substrateof Meltrin β in them. Thus, TACE and Meltrin β mayshed different, but related, growth factors, both of whichare required for valve formation and cardiac myogenesis.However, alternative mechanisms are also conceivable.We favor an idea that TACE and Meltrin β, and probablyother ADAMs, play additive or synergistic roles throughproteolytic processing of the same growth factors,including Neuregulin 1, but act in distinct manners.The idea comes from the distribution of the solubleNRGβ1 in the subcellular fractionation experiment. Inthe presence of Meltrin β, soluble NRG β1 was gener-ated in the Golgi-enriched and ER–Golgi intermediatefractions (fractions 3–6). In contrast, when TACE wasco-introduced, soluble NRGβ1 was found only in thelight membrane fractions (fractions 1 and 2). A similarcleavage pattern of NRG β1 was found in N1E115
transfected with the EQ mutant (compare Figs. 2B and3D). Soluble NRG β1 in the light membrane fractionscould be generated by endogenous TACE in N1E115cells transfected with the EQ mutant.
Based on these findings, we propose that Meltrin β isone determinant of the fate of NRG β1 in cells (Fig. 7).Meltrin β might regulate generation of soluble growthfactors temporally or spatially through the ectodomainshedding in the Golgi apparatus. Compared to ecto-domain shedding in response to some stimuli at the cellsurface or in the exosomes, continuous generation ofsoluble ligands in the Golgi apparatus should be impor-tant for the maintenance of growth factor signaling.Alternatively, because the Golgi apparatus plays a centralrole in sorting various membrane and soluble proteins,the soluble and the membrane-anchored forms of thegrowth factor in the Golgi apparatus might be sortedthrough different pathways to different directions fromeach other. Thus, production of soluble growth factorsat different subcellular sites would modulate delivery of
Figure 7 A model on the role of Meltrinβ in the ectodomain shedding of membrane-anchored growth factors in the Golgiapparatus. In the presence of Meltrin β,the growth factor molecules constitutivelyshed their ectodomains in the Golgiapparatus (shown by red arrows). WhenMeltrin β is absent or expressed at lowlevel, in contrast, intact growth factorsare delivered to the cell surface or toother subcellular compartments such as theexosomes (Stoeck et al. 2006) (shown byblue arrows). These pathways may regulateectodomain shedding temporally andspatially (see text).

Ectodomain shedding by meltrin β (ADAM19) at the Golgi apparatus
© 2007 The Authors Genes to Cells (2007) 12, 329–343Journal compilation © 2007 by the Molecular Biology Society of Japan/Blackwell Publishing Ltd.
341
growth factors spatially. Further studies will clarify thebiological significance of the ectodomain shedding inthe Golgi apparatus mediated by Meltrin β.
Experimental proceduresExpression vectors, materials, cell lines, and transfection
Bafilomycin A1 and brefeldin A were purchased from Sigma-Aldrich (Baf: B1793, BFA: B7651, PMA: 79346-1MG, St. Louis,MO, USA). Isotetrandrine was purchased from BIOMOLInternational (G-520, Plymouth Meeting, PA, USA). The construc-tion methods for wild-type Meltrin β (Meltrin β (WT)), E347Qprotease-deficient Meltrin β (Meltrin β (EQ)) and hemagglutinin(HA)-Neuregulin β1 (HA-NRG β1) were previously described(Shirakabe et al. 2001). The construction of ERD-like protein(ELP-1)-YFP (KDEL receptor-YFP) was previously described(Nagaya et al. 2002). DSRed-Monomer Golgi and pCFP-Golgiwere purchased from BD Biosciences (Clontech, Mountain View,CA, USA).
GFP-Neuregulin β1 (GFP-NRG β1) was constructed byligating Acrosin-GFP (kindly provided by G. Kondo (Kondohet al. 1999)) to the N-terminus of NRG β1. The TACE vector waskindly provided by S. Nagata (the TACE vector we used isdescribed as mTACE in Itai et al. (2001)).
The establishment and maintenance of TACE–/– MEFs havebeen previously described (Buxbaum et al. 1998). COS7 andN1E115 neuroblaststoma cells were propagated every 1 or 2 daysby culturing in Dulbecco’s modified Eagle’s medium (DMEM)containing 10% fetal bovine serum. Except for FCS, cells wereplated at a confluence of 70%−80% for 12 h before transfection.For FCS, vectors were introduced directly into the cells usingsiliconized glass microbeads (Nagaya et al. 2002), and the cellswere then cultured in DMEM containing 1% fetal bovine serum.The total amount of introduced DNA was 5 µg, and the DNAconcentration was 1 µg/µL. For subcellular fractionation, the cellswere transfected by the lipofection method using FuGENE6(Roche Applied Science, Mannheim, Germany).
Antibodies
The antibodies used and their sources were as follows: rabbitpolyclonal anti-C-terminus of Meltrin β antibody (Shirakabeet al. 2001), mouse anti-HA epitope antibody (16B12, Babco,Richmond, CA, USA), anti-ERp72 (Stressgen, Victoria, BC,Canada), anti-Golgi 58K (Sigma), anti-β COP antibody (G6160,Sigma), HRP-conjugated secondary antibody (PI-1000, VectorLaboratories, Burlingame, CA, USA), Alexa488-labeled second-ary antibody (A-11029, Invitrogen) and Cy3-labeled secondaryantibody (A-11029, Invitrogen).
Dorsal root ganglia preparation
Dorsal root ganglia were dissected from mice at E16.5 and disso-ciated by treatment with 0.125% collagenase (Worthington
Biomedical, Freehold, NJ, USA; CLS-1) and 40 U/mL Dispase(Godo Shusei Corporation, Tokyo, Japan) for 30 min at 37 °C.Cells were plated on glass-bottomed dishes coated with poly d-lysine and laminin and were cultured in DMEM supplementedwith 2% B27 (Invitrogen) and 50 ng/mL 2.5S nerve growth factor(NGF) (Invitrogen) for 24–48 h.
Confocal microscopy and FCS analysis
FCS analysis with living cells using confocal microscopy has beenpreviously described (Kamada et al. 2004). FCS measurementswere performed 2.5 h after DNA transfection into COS7 cells.The culture temperature of cells on the microscope stage was con-trolled using an objective heater for a planapochromat lens × 63(Bioptechs, Butler, PA, USA) or a silicon heater (Cell MicroCon-trols, Norfolk, VA, USA) for a C-Apochromat × 40 lens in com-bination with a stage heater (Kitazato Supply, Fujinomiya, Japan).To monitor the temperature of the cells under observation, aCT820 infrared thermometer (Citizen Co., Tokyo, Japan) wasused. For confocal microscopy and FCS analysis, a ConfoCor2instrument (Carl Zeiss Microscopy, Jena, Germany) was used.Confocal images were taken with the laser scanning microscopymodule. The excitation light of an argon ion laser at a wavelengthof 514 nm was reflected by a dichroic mirror (HFT 514) andfocused through a C-Apochromat × 40, NA 1.2 water-immersionobjective (pinhole width, 70 µm). A 530- to 560-nm band-passfilter was used to filter out the remaining scattered laser light. Inall the measurements, the minimum laser power available wasused. The fluorescence signal was recorded for three consecutiveperiods of 15 s (time resolution, 200 ns). The autocorrelationfunction and data fitting were performed with the software pro-vided with the ConfoCor2. Indirect immunofluorescence offixed cells and time-lapse analysis of fluorescent molecules in liv-ing cells were carried out and results were processed as previouslydescribed by Nagaya et al. (2002). The confocal images in Fig. 5were taken with an LSM 510 META laser scanning microscopymodule (Carl Zeiss Microscopy, Jena, Germany).
Subcellular fractionation of COS7 cells and N1E115 cells
After 24 h of transfection, cells cultured on 100-mm plates (oneplate per sample) were washed twice with ice-cold phosphate-buffered saline, harvested in homogenization buffer (10 mmTris–HCl, pH 7.5; 250 mm sucrose) at 0.6 mL per plate andhomogenized by passage 10 times through a 27-gauge needle ona 1-mL syringe. All subsequent steps were performed at 4 °C.Unbroken cells and nuclei were removed by centrifugation at1200 g for 5 min. The post-nuclear supernatant was loaded onpreformed Nycodenz (Histodenz, Sigma) gradients (Okumuraet al. 2006). These preformed gradients were prepared for theBeckman MLS-50 rotor from initial discontinuous gradients(24%, 19%, 15% and 10% Nycodenz in 10 mm Tris–HCl, pH 7.5;3 mm KCl; and 1 mm EDTA) that were allowed to diffuse in ahorizontal position for 45 min at room temperature and then cen-trifuged for 4 h at 100 000 g in an Optima Max ultracentrifuge

T Yokozeki et al.
Genes to Cells (2007) 12, 329–343 © 2007 The AuthorsJournal compilation © 2007 by the Molecular Biology Society of Japan/Blackwell Publishing Ltd.
342
(Beckman Coulter, Fullerton, CA, USA) to generate a non-lineardensity gradient profile. The post-nuclear supernatant was loadedon top of the gradient and centrifuged for 3 h at 100 000 g.Ten fractions were collected from the top, and the proteins inaliquots of the fractions were resolved by SDS-PAGE. The distri-butions of HA-NRG β1, Meltrin β, ERp72 and Golgi 58K weredetermined by immunoblotting using ECL Plus (AmershamBiosciences, GE Healthcare, Piscataway, NJ, USA).
AcknowledgementsThis work was supported in part by Grants-in-Aid for ScientificResearch from the Ministry of Education, Culture, Sports,Science and Technology of Japan, by a research grant from CREST(Core Research for Evolutional Science and Technology) withinthe Japan Science and Technology Agency and by a grant fromJapan Space Forum, JAXA (Japan Aerospace Exploration Agency).T. Yokozeki was supported by a research fellowship from the JapanSociety for the Promotion of Science ( JSPS). We thank Dr G.Kondo for his kind provision of Acrosin-GFP vector, and Dr S.Nagata for his kind provision of TACE vector. We also thank DrE. Nishi for technical advice for cell culture, and Mr A. Yaguchiof Carl Zeiss Co., Osaka, Japan, for his help with analysis of theFCS data.
ReferencesBacia, K., Kim, S.A. & Schwille, P. (2006) Fluorescence cross-
correlation spectroscopy in living cells. Nat. Meth. 3, 83–89.
Blobel, C.P. (2005) ADAMs: key components in EGFR signallingand development. Nat. Rev. Mol. Cell. Biol. 6, 32–43.
Burgess, T.L., Ross, S.L., Qian, Y.X., Brankow, D. & Hu, S.(1995) Biosynthetic processing of neu differentiation factor.Glycosylation trafficking, and regulated cleavage from the cellsurface. J. Biol. Chem. 270, 19188–19196.
Buxbaum, J.D., Liu, K.N., Luo, Y., Slack, J.L., Stocking, K.L.,Peschon, J.J., Johnson, R.S., Castner, B.J., Cerretti, D.P. &Black, R.A. (1998) Evidence that tumor necrosis factor alphaconverting enzyme is involved in regulated α-secretase cleavageof the Alzheimer amyloid protein precursor. J. Biol. Chem. 273,27765–27767.
Chan, D., Strang, M., Judson, B. & Brown, W.J. (2004) Inhibitionof membrane tubule formation and trafficking by isotetran-drine, an antagonist of G-protein-regulated phospholipase A2enzymes. Mol. Biol. Cell 15, 1871–1880.
Falls, D.L. (2003) Neuregulins and the neuromuscular system:10 years of answers and questions. J. Neurocytol. 32, 619–647.
Foldes-Papp, Z., Demel, U. & Tilz, G.P. (2001) Ultrasensitivedetection and identification of fluorescent molecules by FCS:impact for immunobiology. Proc. Natl. Acad. Sci. USA 98,11509–11514.
Gosch, M., Serov, A., Anhut, T., Parker, C.E., Stevenson, M.,Russell, W.E. & Lee, D.C. (2004) Parallel single moleculedetection with a fully integrated single-photon 2×2 CMOSdetector array. J. Biomed. Opt. 9, 913–921.
Hess, S.T., Huang, S., Heikal, A.A. & Webb, W.W. (2002)Biological and chemical applications of fluorescence correlationspectroscopy: a review. Biochemistry 41, 697–705.
Hinkle, C.L., Sunnarborg, S.W., Loiselle, D., Parker, C.E.,Stevenson, M., Russell, W.E. & Lee, D.C. (2004) Selective rolesfor tumor necrosis factor α-converting enzyme/ADAM17in the shedding of the epidermal growth factor receptor ligandfamily: the juxtamembrane stalk determines cleavage efficiency.J. Biol. Chem. 279, 24179–24188.
Hooper, N.M., Karran, E.H. & Turner, A.J. (1997) Membraneprotein secretases. Biochem. J. 321, 265–279.
Horiuchi, K., Zhou, H.M., Kelly, K., Manova, K. & Blobel, C.P.(2005) Evaluation of the contributions of ADAMs 9, 12, 15, 17,and 19 to heart development and ectodomain shedding ofneuregulins β1 and β2. Dev. Biol. 283, 459–471.
Itai, T., Tanaka, M. & Nagata, S. (2001) Processing of tumornecrosis factor by the membrane-bound TNF-α-convertingenzyme, but not its truncated soluble form. Eur. J. Biochem.268, 2074–2082.
Jackson, L.F., Qiu, T.H., Sunnarborg, S.W., Chang, A., Zhang,C., Patterson, C. & Lee, D.C. (2003) Defective valvulogenesisin HB-EGF and TACE-null mice is associated with aberrantBMP signaling. EMBO J. 22, 2704–2716.
Kamada, A., Nagaya, H., Tamura, T., Kinjo, M., Jin, H.Y.,Yamashita, T., Jimbow, K., Kanoh, H. & Wada, I. (2004)Regulation of immature protein dynamics in the endoplasmicreticulum. J. Biol. Chem. 279, 21533–21542.
Klausner, R.D., Donaldson, J.G. & Lippincott-Schwartz, J. (1992)Brefeldin A: insights into the control of membrane traffic andorganelle structure. J. Cell Biol. 116, 1071–1080.
Kogure, T., Karasawa, S., Araki, T., Saito, K., Kinjo, M. &Miyawaki, A. (2006) A fluorescent variant of a protein from thestony coral Montipora facilitates dual-color single-laser fluo-rescence cross-correlation spectroscopy. Nat. Biotechnol. 24,577–581.
Kojro, E., Gimpl, G., Lammich, S., Marz, W. & Fahrenholz, F.(2001) Low cholesterol stimulates the nonamyloidogenicpathway by its effect on the α-secretase ADAM 10. Proc. Natl.Acad. Sci. USA 98, 5815–5820.
Kondoh, G., Gao, X.H., Nakano, Y., Koike, H., Yamada, S.,Okabe, M. & Takeda, J. (1999) Tissue-inherent fate of GPIrevealed by GPI-anchored GFP transgenesis. FEBS Lett. 458,299–303.
Kurisaki, T., Masuda, A., Osumi, N., Nabeshima, Y. & Fujisawa-Sehara, A. (1998) Spatially- and temporally-restricted expressionof meltrin α (ADAM12) and β (ADAM19) in mouse embryo.Mech. Dev. 73, 211–215.
Kurohara, K., Komatsu, K., Kurisaki, T., Masuda, A., Irie, N.,Asano, M., Sudo, K., Nabeshima, Y., Iwakura, Y. & Sehara-Fujisawa, A. (2004) Essential roles of Meltrin β (ADAM19) inheart development. Dev. Biol. 267, 14–28.
Larson, D.R., Ma, Y.M., Vogt, V.M. & Webb, W.W. (2003) Directmeasurement of Gag–Gag interaction during retrovirusassembly with FRET and fluorescence correlation spectroscopy.J. Cell Biol. 162, 1233–1244.
Llopis, J., McCaffery, J.M., Miyawaki, A., Farquhar, M.G. &Tsien, R.Y. (1998) Measurement of cytosolic, mitochondrial,

Ectodomain shedding by meltrin β (ADAM19) at the Golgi apparatus
© 2007 The Authors Genes to Cells (2007) 12, 329–343Journal compilation © 2007 by the Molecular Biology Society of Japan/Blackwell Publishing Ltd.
343
and Golgi pH in single living cells with green fluorescentproteins. Proc. Natl. Acad. Sci. USA 95, 6803–6808.
Loeb, J.A., Susanto, E.T. & Fischbach, G.D. (1998) The neu-regulin precursor proARIA is processed to ARIA afterexpression on the cell surface by a protein kinase C-enhancedmechanism. Mol. Cell. Neurosci. 11, 77–91.
Matthews, V., Schuster, B., Schutze, S., Bussmeyer, I., Ludwig,A., Hundhausen, C., Sadowski, T., Saftig, P., Hartmann, D.,Kallen, K.J. & Rose-John, S. (2003) Cellular cholesterol deple-tion triggers shedding of the human interleukin-6 receptorby ADAM10 and ADAM17 (TACE). J. Biol. Chem. 278,38829–38839.
Meyer, D. & Birchmeier, C. (1995) Multiple essential functionsof neuregulin in development. Nature 378, 386–390.
Meyer, D., Yamaai, T., Garratt, A., Riethmacher-Sonnenberg, E.,Kane, D., Theill, L.E. & Birchmeier, C. (1997) Isoform-specific expression and function of neuregulin. Development124, 3575–3586.
Montero, J.C., Yuste, L., Diaz-Rodriguez, E., Esparis-Ogando, A.& Pandiella, A. (2000) Differential shedding of transmembraneneuregulin isoforms by the tumor necrosis factor-alpha-converting enzyme. Mol. Cell. Neurosci. 16, 631–648.
Nagano, O., Murakami, D., Hartmann, D., De Strooper, B.,Saftig, P., Iwatsubo, T., Nakajima, M., Shinohara, M. & Saya,H. (2004) Cell–matrix interaction via CD44 is independentlyregulated by different metalloproteinases activated in responseto extracellular Ca (2+) influx and PKC activation. J. Cell Biol.165, 893–902.
Nagaya, H., Wada, I., Jia, Y.J. & Kanoh, H. (2002) Diacylglycerolkinase delta suppresses ER-to-Golgi traffic via its SAM and PHdomains. Mol. Biol. Cell 13, 302–316.
Okumura, A.J., Hatsuzawa, K., Tamura, T., Nagaya, H., Saeki, K.,Okumura, F., Nagao, K., Nishikawa, M., Yoshimura, A. &Wada, I. (2006) Involvement of a novel Q-SNARE, D12,in quality control of the endomembrane system. J. Biol. Chem.281, 4495–4506.
Olveczky, B.P. & Verkman, A.S. (1998) Monte Carlo analysis ofobstructed diffusion in three dimensions: application tomolecular diffusion in organelles. Biophys. J. 74, 2722–2730.
Palokangas, H., Ying, M., Vaananen, K. & Saraste, J. (1998)Retrograde transport from the pre-Golgi intermediate com-partment and the Golgi complex is affected by the vacuolarH+-ATPase inhibitor bafilomycin A1. Mol. Biol. Cell 9, 3561–3578.
Peschon, J.J., Slack, J.L., Reddy, P., et al. (1998) An essential rolefor ectodomain shedding in mammalian development. Science282, 1281–1284.
Reiss, K., Maretzky, T., Ludwig, A., Tousseyn, T., de Strooper,B., Hartmann, D. & Saftig, P. (2005) ADAM10 cleavage ofN-cadherin and regulation of cell–cell adhesion and beta-catenin nuclear signalling. EMBO J. 24, 742–752.
Saito, K., Ito, E., Takakuwa, Y., Tamura, M. & Kinjo, M. (2003)In situ observation of mobility and anchoring of PKCβI inplasma membrane. FEBS Lett. 541, 126–131.
Schlondorff, J., Becherer, J.D. & Blobel, C.P. (2000) Intracellularmaturation and localization of the tumour necrosis factorα convertase (TACE). Biochem. J. 347 Part 1, 131–138.
Schweizer, A., Fransen, J.A., Bachi, T., Ginsel, L. & Hauri, H.P.(1988) Identification, by a monoclonal antibody, of a 53-kDprotein associated with a tubulo-vesicular compartment atthe cis-side of the Golgi apparatus. J. Cell Biol. 107, 1643–1653.
Shirakabe, K., Wakatsuki, S., Kurisaki, T. & Fujisawa-Sehara, A.(2001) Roles of Meltrin β/ADAM19 in the processing ofneuregulin. J. Biol. Chem. 276, 9352–9358.
Stoeck, A., Keller, S., Riedle, S., Sanderson, M.P., Runz, S.,Le Naour, F., Gutwein, P., Ludwig, A., Rubinstein, E. &Altevogt, P. (2006) A role for exosomes in the constitutive andstimulus-induced ectodomain cleavage of L1 and CD44.Biochem. J. 393, 609–618.
Tanaka, M., Itai, T., Adachi, M. & Nagata, S. (1998) Down-regulation of Fas ligand by shedding. Nat. Med. 4, 31–36.
Voeltz, G.K., Rolls, M.M. & Rapoport, T.A. (2002) Structuralorganization of the endoplasmic reticulum. EMBO Rep. 3,944–950.
Wakatsuki, S., Kurisaki, T. & Sehara-Fujisawa, A. (2004) Lipidrafts identified as locations of ectodomain shedding mediatedby Meltrin β/ADAM19. J. Neurochem. 89, 119–123.
Widengren, J. & Rigler, R. (1998) Fluorescence correlationspectroscopy as a tool to investigate chemical reactions insolutions and on cell surfaces. Cell. Mol. Biol. (Noisy-le-Grand)44, 857–879.
Wong, S.T., Winchell, L.F., McCune, B.K., Earp, H.S., Teixido,J., Massague, J., Herman, B. & Lee, D.C. (1989) The TGF-αprecursor expressed on the cell surface binds to the EGFreceptor on adjacent cells, leading to signal transduction.Cell 56, 495–506.
Yamazaki, S., Iwamoto, R., Saeki, K., et al. (2003) Mice withdefects in HB-EGF ectodomain shedding show severe develop-mental abnormalities. J. Cell Biol. 163, 469–475.
Zhou, H.M., Weskamp, G., Chesneau, V., et al. (2004) Essentialrole for ADAM19 in cardiovascular morphogenesis. Mol. Cell.Biol. 24, 96–104.
Zimina, E.P., Bruckner-Tuderman, L. & Franzke, C.W. (2005)Shedding of collagen XVII ectodomain depends on plasmamembrane microenvironment. J. Biol. Chem. 280, 34019–34024.
Received: 4 October 2006 Accepted: 4 December 2006
Supplementary materialThe following supplementary materials are available for this articleonline:
Figure S1 FCS analysis of GFP-NRG β1 in living cells.
Figure S2 Autocorrelation functions and best-fit two-component simulation profiles.
Figure S3 The distribution of Meltrin β under the condition ofthe inhibitors used in the FCS measurements.
Figure S4 Summary of FCS measurements.
Copyright © 2022 FDOKUMEN




















