
Lesions in the anterior bed nucleus of the stria terminalis in Syrian hamsters block...
11
BIOLOGY OF REPRODUCTION 57, 796-806 (1997) Lesions in the Anterior Bed Nucleus of the Stria Terminalis in Syrian Hamsters Block Short-Photoperiod-induced Testicular Regression' Martin N. Raitiere, 3 Vasilios T. Garyfallou, 3 and Henryk F. Urbanski 2 , 3 .4 Division of Neuroscience, 3 Oregon Regional Primate Research Center, Beaverton, Oregon 97006 Department of Physiology and Pharmacology,4 Oregon Health Sciences University, Portland, Oregon 97201 ABSTRACT To elucidate the neural circuitry involved in the photoperi- odic control of seasonal reproduction, adult male Syrian ham- sters, previously housed under long photoperiods (LD; 14 h of light per day), received sham or bilateral radiofrequency-current lesions directed towards one of three anterior-to-posterior levels of the bed nucleus of the stria terminalis (BNST; far anterior, anterior, posterior). They were then transferred to a short pho- toperiod (SD; 6 h of light per day) for 12 wk, and their testicular weights and plasma FSH, LH, and testosterone concentrations were determined. All of these parameters became markedly in- hibited in the sham-lesioned SD controls and also in the far anterior and posterior BNST lesioned groups. In contrast, this inhibitory response to SD was completely abolished in 8 of 14 animals that had received anterior BNST lesions; only in these 8 animals did the lesion encompass the lateral aspect of the anterior BNST. In a second experiment, hamsters that had pre- viously been exposed to SD for 12 wk in order to induce testic- ular regression were lesioned in the anterior BNST and for the next 4 weeks were either exposed to LD or further maintained in SD. However, in neither case did the anterior BNST lesions perturb the normal photoperiodic response. Paired testes weights and plasma FSH, LH, and testosterone concentrations at 4 wk did not differ significantly (p > 0.05) between the le- sioned animals and their respective sham-lesioned LD and SD controls, which, respectively, showed recrudescence of the re- productive axis or remained in a regressed condition. Taken to- gether, the results suggest that lateral aspects of the anterior BNST contain a cell group that is critical for perception of the SD neuro-inhibitory signal; obliteration of this cell group inter- rupts the transmission of the inhibitory signal to the reproduc- tive axis but does not directly stimulate it. INTRODUCTION The Syrian hamster (Mesocricetus auratus), a popular model for the study of mammalian photoperiodism, dis- plays a pronounced involution of the reproductive axis in short photoperiods [1, 2]. It is well known that this process requires the generation of melatonin by the pineal gland, that the duration of the melatonin secretory pulse serves to distinguish long days (LD) from short days (SD) [3], and that melatonin receptors in the medial hypothalamus play a key role in transduction of the SD signal [4, 5]. However, the precise steps that are interposed between the processing or "readout" of the melatonin signal by these hypothalamic target cells and the modulation of the reproductive axis re- main unknown. It has often been supposed that the link Accepted May 23, 1997. Received March 24, 1997. 'This work was supported by grants from the National Institutes of Health (HD-29186, HD-24312, and RR-00163) and the National Science Foundation (93-09368). 2Correspondence: Henryk F. Urbanski, Division of Neuroscience, Or- egon Regional Primate Research Center, 505 NW 1 85th Avenue, Beaver- ton, OR 97006. FAX: (503) 690-5384; e-mail: [email protected] between melatonin readout and the GnRH-secreting cells may be direct and intrahypothalamic, e.g., that melatonin- sensitive neurons in the anterior hypothalamic area directly signal GnRH-secreting neurons either at the level of their cell bodies in the medial preoptic area (MPOA) or at the level of their nerve terminals in the median eminence [6]. Yet we recently found evidence suggesting that an extra- hypothalamic region, the bed nucleus of the stria terminalis (BNST), may play a role in communicating the melatonin signal to the GnRH system, for lesions in the hamster BNST impaired the expected regressive response to SD [7]. We suggested that the BNST may enclose a "comparator" which, as Herbert has suggested [8], may be interposed between, and be anatomically distinct from, the tissues re- sponsible for "primary readout" of melatonin and those responsible for GnRH secretion. However, it was not clear from our previous results [7] whether the BNST lesion truly interfered with photoperi- odism proper or whether it simply disinhibited gonadotro- pin secretion in a nonspecific fashion. This question has complicated previous studies of hamster photoperiodism. For example, while it is known that ablation of the hamster olfactory bulbs impairs the expected reproductive response to SD [9, 10], it is not clear that this lesion truly interferes with photoperiodism since bulbectomy disinhibits gonado- tropins to some extent regardless of photoperiod, i.e., in hamsters under both SD and LD [11]; the resulting high gonadotropin levels could thus be offsetting or masking a partially preserved SD-driven decrease in gonadotropins, thereby yielding a false impression of nonresponse to SD [10-12]. A similar argument may thus be invoked to ex- plain the apparent nonresponse to SD of the BNST-lesioned hamster. In the present study, we attempted therefore to clarify whether the previously noted impairment of SD gonadal regression in the Syrian hamster bearing a BNST lesion [7] derives from a true interference with circuits mediating readout of the SD melatonin signal or rather from nonspe- cific disinhibition or stimulation of GnRH cells. We ad- dressed this question by evaluating the effect of a BNST lesion upon the hamster reproductive axis under varying photoperiodic conditions, i.e., we sought to determine whether the BNST lesion not only blocks gonadal regres- sion in a reproductively competent hamster newly exposed to SD but also stimulates gonadotropins in a hamster ren- dered reproductively quiescent by previous exposure to SD. Secondly, we attempted to determine whether specific sub- regions of the BNST might play a role in photoperiodism by placing far smaller lesions than we had achieved in the previous study [7]. We focused in particular on the possible involvement of the anterior division of the BNST in the photoperiodic response to SD. This region not only receives a dense afferent input from the ventromedial nucleus of the hypothalamus [13, 14], a nucleus that plays a key role in readout of the SD melatonin signal [5], but also contains 796
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Lesions in the anterior bed nucleus of the stria terminalis in Syrian hamsters block...

BIOLOGY OF REPRODUCTION 57, 796-806 (1997)
Lesions in the Anterior Bed Nucleus of the Stria Terminalis in Syrian HamstersBlock Short-Photoperiod-induced Testicular Regression'
Martin N. Raitiere,3 Vasilios T. Garyfallou,3 and Henryk F. Urbanski 2,3.4
Division of Neuroscience,3 Oregon Regional Primate Research Center, Beaverton, Oregon 97006Department of Physiology and Pharmacology,4 Oregon Health Sciences University, Portland, Oregon 97201
ABSTRACT
To elucidate the neural circuitry involved in the photoperi-odic control of seasonal reproduction, adult male Syrian ham-sters, previously housed under long photoperiods (LD; 14 h oflight per day), received sham or bilateral radiofrequency-currentlesions directed towards one of three anterior-to-posterior levelsof the bed nucleus of the stria terminalis (BNST; far anterior,anterior, posterior). They were then transferred to a short pho-toperiod (SD; 6 h of light per day) for 12 wk, and their testicularweights and plasma FSH, LH, and testosterone concentrationswere determined. All of these parameters became markedly in-hibited in the sham-lesioned SD controls and also in the faranterior and posterior BNST lesioned groups. In contrast, thisinhibitory response to SD was completely abolished in 8 of 14animals that had received anterior BNST lesions; only in these8 animals did the lesion encompass the lateral aspect of theanterior BNST. In a second experiment, hamsters that had pre-viously been exposed to SD for 12 wk in order to induce testic-ular regression were lesioned in the anterior BNST and for thenext 4 weeks were either exposed to LD or further maintainedin SD. However, in neither case did the anterior BNST lesionsperturb the normal photoperiodic response. Paired testesweights and plasma FSH, LH, and testosterone concentrationsat 4 wk did not differ significantly (p > 0.05) between the le-sioned animals and their respective sham-lesioned LD and SDcontrols, which, respectively, showed recrudescence of the re-productive axis or remained in a regressed condition. Taken to-gether, the results suggest that lateral aspects of the anteriorBNST contain a cell group that is critical for perception of theSD neuro-inhibitory signal; obliteration of this cell group inter-rupts the transmission of the inhibitory signal to the reproduc-tive axis but does not directly stimulate it.
INTRODUCTION
The Syrian hamster (Mesocricetus auratus), a popularmodel for the study of mammalian photoperiodism, dis-plays a pronounced involution of the reproductive axis inshort photoperiods [1, 2]. It is well known that this processrequires the generation of melatonin by the pineal gland,that the duration of the melatonin secretory pulse serves todistinguish long days (LD) from short days (SD) [3], andthat melatonin receptors in the medial hypothalamus playa key role in transduction of the SD signal [4, 5]. However,the precise steps that are interposed between the processingor "readout" of the melatonin signal by these hypothalamictarget cells and the modulation of the reproductive axis re-main unknown. It has often been supposed that the link
Accepted May 23, 1997.Received March 24, 1997.'This work was supported by grants from the National Institutes of
Health (HD-29186, HD-24312, and RR-00163) and the National ScienceFoundation (93-09368).
2Correspondence: Henryk F. Urbanski, Division of Neuroscience, Or-egon Regional Primate Research Center, 505 NW 1 85th Avenue, Beaver-ton, OR 97006. FAX: (503) 690-5384; e-mail: [email protected]
between melatonin readout and the GnRH-secreting cellsmay be direct and intrahypothalamic, e.g., that melatonin-sensitive neurons in the anterior hypothalamic area directlysignal GnRH-secreting neurons either at the level of theircell bodies in the medial preoptic area (MPOA) or at thelevel of their nerve terminals in the median eminence [6].Yet we recently found evidence suggesting that an extra-hypothalamic region, the bed nucleus of the stria terminalis(BNST), may play a role in communicating the melatoninsignal to the GnRH system, for lesions in the hamsterBNST impaired the expected regressive response to SD [7].We suggested that the BNST may enclose a "comparator"which, as Herbert has suggested [8], may be interposedbetween, and be anatomically distinct from, the tissues re-sponsible for "primary readout" of melatonin and thoseresponsible for GnRH secretion.
However, it was not clear from our previous results [7]whether the BNST lesion truly interfered with photoperi-odism proper or whether it simply disinhibited gonadotro-pin secretion in a nonspecific fashion. This question hascomplicated previous studies of hamster photoperiodism.For example, while it is known that ablation of the hamsterolfactory bulbs impairs the expected reproductive responseto SD [9, 10], it is not clear that this lesion truly interfereswith photoperiodism since bulbectomy disinhibits gonado-tropins to some extent regardless of photoperiod, i.e., inhamsters under both SD and LD [11]; the resulting highgonadotropin levels could thus be offsetting or masking apartially preserved SD-driven decrease in gonadotropins,thereby yielding a false impression of nonresponse to SD[10-12]. A similar argument may thus be invoked to ex-plain the apparent nonresponse to SD of the BNST-lesionedhamster.
In the present study, we attempted therefore to clarifywhether the previously noted impairment of SD gonadalregression in the Syrian hamster bearing a BNST lesion [7]derives from a true interference with circuits mediatingreadout of the SD melatonin signal or rather from nonspe-cific disinhibition or stimulation of GnRH cells. We ad-dressed this question by evaluating the effect of a BNSTlesion upon the hamster reproductive axis under varyingphotoperiodic conditions, i.e., we sought to determinewhether the BNST lesion not only blocks gonadal regres-sion in a reproductively competent hamster newly exposedto SD but also stimulates gonadotropins in a hamster ren-dered reproductively quiescent by previous exposure to SD.Secondly, we attempted to determine whether specific sub-regions of the BNST might play a role in photoperiodismby placing far smaller lesions than we had achieved in theprevious study [7]. We focused in particular on the possibleinvolvement of the anterior division of the BNST in thephotoperiodic response to SD. This region not only receivesa dense afferent input from the ventromedial nucleus of thehypothalamus [13, 14], a nucleus that plays a key role inreadout of the SD melatonin signal [5], but also contains
796

BNST LESIONS IN HAMSTERS
cell bodies projecting directly to brainstem noradrenergicsystems [15] that undergo significant seasonal change inactivity preceding and probably driving seasonal changesin gonadotropin levels [2, 16, 17]. We therefore viewed theanterior division of the BNST as an attractive candidate forthe locus of a comparator [8], interposed between the read-out of the SD melatonin signal and the reproductive axis.We reasoned that ablation of such a locus might block thehamster's perception of the SD signal and its subsequentgonadal response without stimulating or disinhibiting its re-productive axis under other photoperiodic conditions.
MATERIALS AND METHODS
Animals
Male Syrian hamsters (Mesocricetus auratus) of the Lak:LVG (SYR) stock (Charles River Laboratories Inc., Wil-mington, MA) were housed under LD photoperiods, com-prising 14 h of light and 10 h of darkness, until 10 wk ofage and were then used in the study. Room temperaturewas maintained at 21-22°C at all times, and pelleted rodentchow and water were available ad libitum. All experimentswere performed in accordance with NIH guidelines andwith the approval of the Institutional Animal Care and Uti-lization Committee.
BNST Terminology
Several detailed descriptions of the rat BNST, each withvarying nomenclature, subdivide the region on the basis ofcytoarchitectural, chemoarchitectural, and connectionaldata [18-20]. While the hamster BNST has not been stud-ied in equivalent detail, recent studies in the hamster [21]suggest a close resemblance to the rat BNST. For the pur-poses of this study, we have focused on the most salientsubnuclei of the BNST and have named them according toMoga et al. [19], but we have segregated these subnucleiinto anterior vs. posterior groups as outlined by Ju andSwanson [20]. Accordingly, the anterior division includesthe antromedial BNST (am), dorsolateral BNST (dl), ven-trolateral BNST (vl), ventromedial BNST (vm), and par-astrial (ps) subnuclei; and the posterior division includesthe posteromedial BNST (pm), posterointermediate BNST(pi), and posterolateral BNST (pl) subnuclei (see Fig. 1 fordepiction of these subnuclei). We have also identified therostral-most extent of the BNST, i.e., rostral to the point atwhich the anterior commissure crosses the midline, as the"far anterior (FAnt)" BNST, thus permitting identificationof three distinct rostral-to-caudal levels (far anterior, ante-rior, posterior; see description of groups below). We haveincluded the ps subnucleus as part of the BNST in view ofits similarities to the BNST in cytoarchitecture and peptidecontent [19].
Experiment 1: Effect of BNST Lesions on TesticularRegression in SD
A Grass LM4 radiofrequency-current lesion maker(Grass Medical Instruments, Quincy, MA) was used to pro-duce fine bilateral lesions in the brains of sexually maturemale hamsters. The animals were anesthetized using tribro-moethanol, and their heads were positioned in a stereotaxicframe (David Kopf Instruments, Tujunga, CA), with breg-ma and lambda in a horizontal plane. A small hole (diam-eter = 1 mm) was drilled in the cranium, and a stainless-steel electrode (diameter = 0.38 mm), insulated with sili-cone coating except for 0.38 mm at the tip, was used to
deliver 2 mA of current (at 95 volts) for 20 sec to one ofthree anterior-posterior regions: FAnt BNST, anterior (Ant)BNST, and posterior (Post) BNST (2.0, 1.1, and 0.7 mmanterior to bregma, respectively); all of the lesions wereplaced bilaterally 1.1-1.2 mm from the mid-sagittal sutureand 5.2-5.7 mm below the dura. Sham-lesioned controlsunderwent surgical procedures identical to those for the le-sioned animals except that the electrode was not loweredinto the brain. Stainless-steel wound clips (MikRon Preci-sion Inc., Gardens, CA) were used to close the skin incisionon the head, and the animals recovered from anesthesia ona heated pad. One to 5 days after surgery, 25 lesioned an-imals and 7 sham-lesioned controls were exposed to SDphotoperiods (6L:18D); 7 sham-lesioned animals also re-mained under LD, serving as additional controls. Caliperswere used to measure testicular width in each of the ani-mals at Week 0 (i.e., day of transfer to SD) and again 4, 8,and 12 wk later. All of the animals received a lethal over-dose of ether at Week 12, and determinations were madeof body and paired testes weights; the testes were fixed inBouin's fluid and subsequently processed for histologicalexamination using Lee's and hematoxylin stains. A bloodsample (1 ml) was collected by cardiac puncture for sub-sequent RIA of plasma FSH, LH, and testosterone [22], andthe animals' brains were then perfusion-fixed using a 4%paraformaldehyde, 50 mM Tris solution (pH 7.6) [23].
Experiment 2: Effect of BNST Lesions on TesticularRecrudescence in LD
Twenty-three adult male hamsters were exposed to SDphotoperiods in order to induce testicular regression. By 12wk all of the animals had shown a decrease in testicularsize, from an initial width measurement of 11-12 mm to< 5 mm, and were randomly assigned to one of four treat-ment groups. One group received bilateral radiofrequency-current lesions in the Ant BNST, using stereotaxic coordi-nates described in experiment 1; 2 mA of current (at 95volts) was administered for 10 sec. One to 5 days afterlesioning, these animals were exposed to LD photoperiods,while a second anterior BNST-lesioned group remained ex-posed to SD. Two additional groups of animals served asLD and SD sham-lesioned controls. After 4 wk, all of theanimals received a lethal dose of ether, and body and pairedtestes weights were determined; the testes were fixed inBouin's fluid and subsequently processed for histologicalexamination using Lee's and hematoxylin stains. A bloodsample (1 ml) was collected by cardiac puncture for sub-sequent RIA of plasma FSH, LH, and testosterone [22], andthe animals' brains were then perfusion-fixed using a 4%paraformaldehyde, 50 mM Tris solution (pH 7.6) [23].
Brain Histology
To verify placement of the lesions, the brains were sec-tioned coronally (25 m) using a freezing-stage microtomeand were stained using thionin. The information containedin these sections provided the primary basis for the lesionmaps depicted in Figures 6 and 11. However, to aid withthe localization of the lesion sites and to determine the ex-tent of associated glial proliferation, some of the sectionswere also processed for immunocytochemistry using an an-tibody to glial fibrillary acidic protein (GFAP; 20 pLg/mldilution; Boehringer-Mannheim Biochemicals, Indianapo-lis, IN) and the standard avidin-biotin-peroxidase system(Vector Laboratories, Burlingame, CA) with 3,3'-diamino-benzidine tetrahydrochloride (DAB) as the chromogen [23].
797
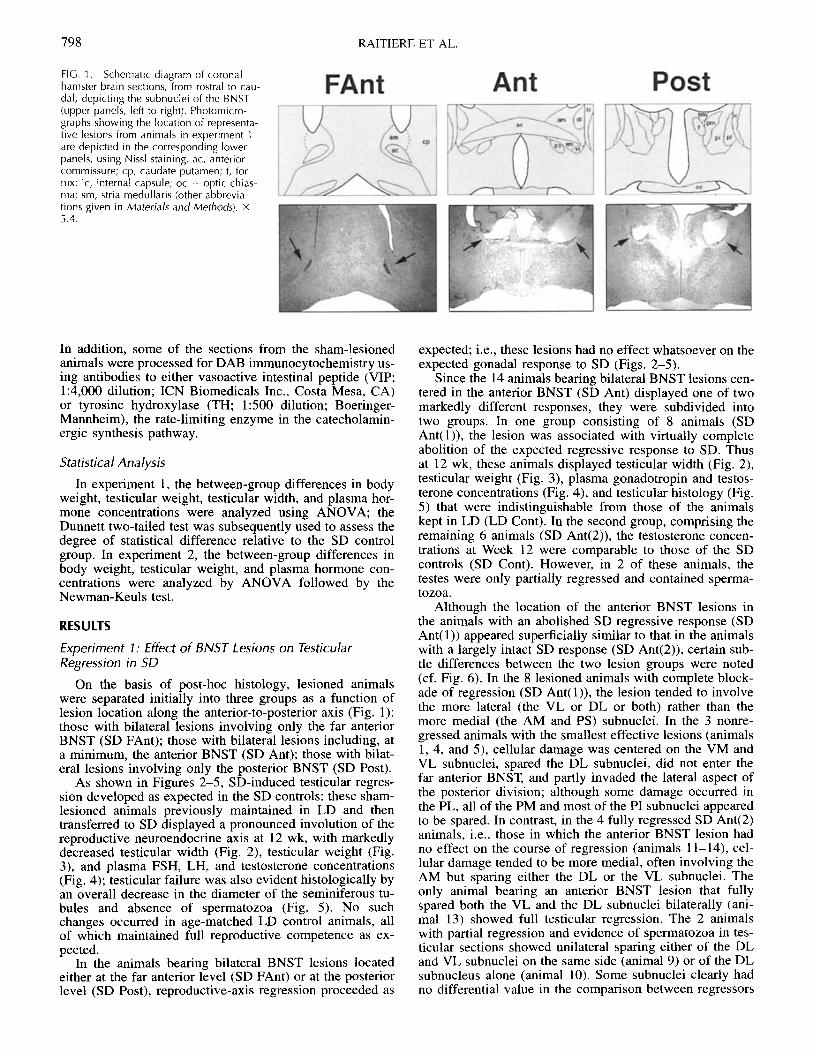
RAITIERE ET AL.
FIG. 1. Schematic diagram of coronalhamster brain sections, from rostral to cau-dal, depicting the subnuclei of the BNST(upper panels, left to right). Photomicro-graphs showing the location of representa-tive lesions from animals in experiment 1are depicted in the corresponding lowerpanels, using Nissl staining. ac, anteriorcommissure; cp, caudate putamen; f, for-nix; ic, internal capsule; oc = optic chias-ma; sm, stria medullaris (other abbrevia-tions given in Materials and Methods). x5.4.
In addition, some of the sections from the sham-lesionedanimals were processed for DAB immunocytochemistry us-ing antibodies to either vasoactive intestinal peptide (VIP;1:4,000 dilution; ICN Biomedicals Inc., Costa Mesa, CA)or tyrosine hydroxylase (TH; 1:500 dilution; Boeringer-Mannheim), the rate-limiting enzyme in the catecholamin-ergic synthesis pathway.
Statistical Analysis
In experiment 1, the between-group differences in bodyweight, testicular weight, testicular width, and plasma hor-mone concentrations were analyzed using ANOVA; theDunnett two-tailed test was subsequently used to assess thedegree of statistical difference relative to the SD controlgroup. In experiment 2, the between-group differences inbody weight, testicular weight, and plasma hormone con-centrations were analyzed by ANOVA followed by theNewman-Keuls test.
RESULTS
Experiment 1: Effect of BNST Lesions on TesticularRegression in SD
On the basis of post-hoc histology, lesioned animalswere separated initially into three groups as a function oflesion location along the anterior-to-posterior axis (Fig. 1):those with bilateral lesions involving only the far anteriorBNST (SD FAnt); those with bilateral lesions including, ata minimum, the anterior BNST (SD Ant); those with bilat-eral lesions involving only the posterior BNST (SD Post).
As shown in Figures 2-5, SD-induced testicular regres-sion developed as expected in the SD controls: these sham-lesioned animals previously maintained in LD and thentransferred to SD displayed a pronounced involution of thereproductive neuroendocrine axis at 12 wk, with markedlydecreased testicular width (Fig. 2), testicular weight (Fig.3), and plasma FSH, LH, and testosterone concentrations(Fig. 4); testicular failure was also evident histologically byan overall decrease in the diameter of the seminiferous tu-bules and absence of spermatozoa (Fig. 5). No suchchanges occurred in age-matched LD control animals, allof which maintained full reproductive competence as ex-pected.
In the animals bearing bilateral BNST lesions locatedeither at the far anterior level (SD FAnt) or at the posteriorlevel (SD Post), reproductive-axis regression proceeded as
expected; i.e., these lesions had no effect whatsoever on theexpected gonadal response to SD (Figs. 2-5).
Since the 14 animals bearing bilateral BNST lesions cen-tered in the anterior BNST (SD Ant) displayed one of twomarkedly different responses, they were subdivided intotwo groups. In one group consisting of 8 animals (SDAnt(l)), the lesion was associated with virtually completeabolition of the expected regressive response to SD. Thusat 12 wk, these animals displayed testicular width (Fig. 2),testicular weight (Fig. 3), plasma gonadotropin and testos-terone concentrations (Fig. 4), and testicular histology (Fig.5) that were indistinguishable from those of the animalskept in LD (LD Cont). In the second group, comprising theremaining 6 animals (SD Ant(2)), the testosterone concen-trations at Week 12 were comparable to those of the SDcontrols (SD Cont). However, in 2 of these animals, thetestes were only partially regressed and contained sperma-tozoa.
Although the location of the anterior BNST lesions inthe animals with an abolished SD regressive response (SDAnt(l)) appeared superficially similar to that in the animalswith a largely intact SD response (SD Ant(2)), certain sub-tle differences between the two lesion groups were noted(cf. Fig. 6). In the 8 lesioned animals with complete block-ade of regression (SD Ant(l)), the lesion tended to involvethe more lateral (the VL or DL or both) rather than themore medial (the AM and PS) subnuclei. In the 3 nonre-gressed animals with the smallest effective lesions (animals1, 4, and 5), cellular damage was centered on the VM andVL subnuclei, spared the DL subnuclei, did not enter thefar anterior BNST, and partly invaded the lateral aspect ofthe posterior division; although some damage occurred inthe PL, all of the PM and most of the PI subnuclei appearedto be spared. In contrast, in the 4 fully regressed SD Ant(2)animals, i.e., those in which the anterior BNST lesion hadno effect on the course of regression (animals 11-14), cel-lular damage tended to be more medial, often involving theAM but sparing either the DL or the VL subnuclei. Theonly animal bearing an anterior BNST lesion that fullyspared both the VL and the DL subnuclei bilaterally (ani-mal 13) showed full testicular regression. The 2 animalswith partial regression and evidence of spermatozoa in tes-ticular sections showed unilateral sparing either of the DLand VL subnuclei on the same side (animal 9) or of the DLsubnucleus alone (animal 10). Some subnuclei clearly hadno differential value in the comparison between regressors
798

BNST LESIONS IN HAMSTERS
7 6 5 8 66 8a
8$1 ii~~~~~~i~~i.:l 3p,1 ..
*~~~~
* ** *
* ----
| XE;x
Va)U)
300
-a 250
E 200
150
> 100'am 50
5
4
3
V)
I-
2
1
0
O u. .D D ] e
../J U) O9l (/a or
FIG. 3. Effect of radiofrequency-current lesions on SD-induced testicularregression in adult Syrian hamsters after 12 wk of photoperiodic treatment(see Results for definitions of treatment groups). Each bar represents themean; the SEMs are depicted by vertical lines and the number of animalsper group is indicated by the adjacent numbers. ** = p < 0.01 (Dunnetttwo-tailed test, relative to the SD Control group).
04'r
a)B.,
0 0 < 0 _ N0 " . C Ca00 00a< -J U) (n ) 0 0a
U) U)
FIG. 2. Effect of radiofrequency-current lesions on SD-induced testicularregression in adult Syrian hamsters. The bilateral lesions were centeredwithin the SD Ant or were placed either in an SD Post or SD FAnt region.Additional animals served either as SD or LD sham-lesioned controls(Cont). The sizes of the testes were determined every 4 wk by measuringtheir width through the scrotal wall, 4 mm representing the minimumpalpable size. Each bar represents the mean; the SEMs are depicted byvertical lines, and the number of animals per group is indicated by theadjacent numbers. ** = p < 0.01 (Dunnett two-tailed test, relative to SDCont).
and nonregressors: the PS subnuclei were fully spared in 5of 8 nonregressors and in 5 of 6 regressors, and the PMsubnuclei were spared in 6 of 8 nonregressors and signifi-cantly damaged in 4 of the 5 regressors in the SD Postgroup, suggesting that the PS and PM subnuclei do notcomprise part of the circuitry mediating regression. In sum-mary, while ablation of no single subnucleus could be iden-tified as a sine qua non for blockade of the SD regressiveresponse, the lesions most effective in abolishing this re-sponse tended to be centered in the anterior BNST, to in-volve the DL or VL subnucleus or both, and on occasion
to extend somewhat into the lateral aspect of the posteriordivision. Lesions involving only the rostral-most BNST, themedial aspects of the anterior division, or the medial as-pects of the posterior division had no effect on the SDregressive response.
Comparison of some sections from sham-lesioned ani-mals processed for antibodies to VIP and TH with sectionsfrom lesioned animals in the SD Ant(l) group aided in thecharacterization of the sites effective in blocking the SDresponse. The effective lesions involved a dense plexus ofTH-positive and VIP-positive fibers most evident in the lat-eral aspects of the anterior BNST (Fig. 7). The location ofthe TH-positive fibers corresponds to that of known nor-adrenergic and dopaminergic projections that terminatemost densely in the VL subnucleus and the DL subnucleus,respectively [24-27], and the location of the VIP-positivefibers corresponds to that of known VIP projections thatterminate in the lateral subnuclei at this level of the BNST[28, 29]; thus the approximate superimposition of the ef-fective lesion on these two fiber plexuses corroborated theview that the lesion centered on the lateral and particularlythe VL and DL subnuclei.
Nonquantitative inspection of sections processed forGFAP, useful in precise delineation of lesion borders (Fig.7), did not reveal any visible differences in glial prolifer-ation at these borders in sections from nonregressed vs.fully regressed animals lesioned in the anterior BNST(groups SD Ant(l) and SD Ant(2), respectively).
Experiment 2: Effect of BNST Lesions on TesticularRecrudescence in LD
As expected, pretreatment of male hamsters with 12 wkof SD resulted in complete testicular regression. Subse-quent exposure of these animals to LD resulted in progres-sive testicular recrudescence during the ensuing 4 wkwhereas animals remaining in SD for an additional 4 wk
14
12
10
8
6
4
14
12
10
8
6
4
14
12
10
8
6
EE
(-
.-
Im
12
4
14
12
10
8
6
4
799

RAITIERE ET AL.
400
300
' 200
inIL 100
200
150
50
ECn
a,
oa,
1-
6
5
4
3
2
1
0O o
o oc] a*1 en
C 0 ;
on 0 < a
00M a aU) U 00Cl) U)
FIG. 4. Effect of radiofrequency-current lesions on SD-induced inhibi-tion of the reproductive neuroendocrine axis in adult Syrian hamsters after12 wk of photoperiodic treatment (see Results for definitions of treatmentgroups). Each bar represents the mean plasma hormone concentration,and the SEMs are depicted by vertical lines. * = p < 0.05, ** = p <0.01 (Dunnett two-tailed test, relative to the SD Control group).
continued to display complete gonadal involution. Lesionsof the anterior BNST in two groups of animals undergoingthe same photoperiodic protocol had no effect on the ex-pected response: those switched to LD recrudesced to thesame extent as the matched controls while those remainingin SD displayed no evidence of recrudescence (Figs. 8-10).Post-hoc analysis of lesion placement revealed that 4 of 6animals in the LD-exposed group (LD BNST) sustainedbilateral destruction of the DL subnucleus, as did 4 of 7animals in the SD-exposed group (SD BNST). The anteriorBNST lesions in experiment 2 (Fig. 11) did not exactlyreplicate those in experiment 1 (in particular, the lesions inexperiment 2 tended to destroy the supracommissural re-gion including the DL subnucleus but to spare the subcom-missural region including the VL subnucleus). Nonetheless,the lesions in experiment 2 regularly involved at least oneof the subnuclei (i.e., DL), ablation of which in experiment1 was associated with blockade of the SD response. Indeed,in certain cases there was hardly any discrepancy betweenlesions in experiment 1 and those in experiment 2. Thuslesions that abolished the regressive response in animals 6and 8 of experiment 1 (see Fig. 6) were nearly identical tolesions that failed to stimulate regression in animals 1, 2,and 3 of the SD group in experiment 2 and to lesions thatfailed to interrupt photostimulation in animals 3, 4, and 5of the LD group in experiment 2 (see Fig. 11).
FIG. 5. Effect of radiofrequency-current lesions on the histological ap-pearance of testes from adult Syrian hamsters. Note the large diameter ofthe seminiferous tubules in the testes of controls maintained under LD (A)and in hamsters that were exposed for 12 wk to SD but received lesionsin the anterior BNST (E). Note also the presence of spermatozoa in thelumen. In contrast, animals in the other groups showed all of the usualsigns of seasonal testicular regression. Their seminiferous tubules showeda marked reduction in diameter, and spermatogonia represented the pre-dominant cell type. A) LD Cont; B) SD Cont; C) SD FAnt; D) SD Post; E)SD Ant(l); F) SD Ant(2) (see Results for definitions of treatment groups).Scale bar = 0.2 mm.
DISCUSSION
Ablation of a relatively small region centered in the an-terior division of the BNST was associated with a pro-nounced impairment in the Syrian hamster's ability to de-velop the expected gonadal response to SD. Syrian ham-sters of the Lak:LVG(SYR) stock ordinarily achieve com-plete testicular regression within 10-12 wk of transfer fromLD to SD [30], a response exemplified in our control SDgroup. On the other hand, 8 of the 14 hamsters that receivedlesions in the anterior BNST, those proving to have lesionsencompassing the lateral aspects of this division, displayedno regression even after 12 wk of SD. Interestingly, abla-tion of other BNST regions had no effect on the normalcourse of regression. It therefore appears that cell groupswithin lateral subnuclei of the anterior BNST may play animportant role in photoperiodism.
While the current results support our previous findingson the role of the BNST in photoperiodism, they suggestthat we were mistaken in assuming that the key BNST lo-cale for perception of the SD signal resides in the posteriordivision [7]. The earlier study's utilization of relativelylarge lesions obscured any reliable conclusions as to selec-tive involvement of BNST subregions, a deficit that weattempted to correct in the current study by making farsmaller lesions, checking their boundaries with GFAP stain-ing, and confirming lesion placement by examining TH andVIP immunohistochemistry in the region of interest.
The results of this study raise two important questions.First, are the effects of the anterior BNST lesions on SD-in-duced gonadal regression apparent rather than real; i.e.,
,*
*=
800

BNST LESIONS IN HAMSTERS
FIG. 6. Schematic diagrams of coronal brain sections from hamster, depicting the location where radiofrequency-current lesions were centered inexperiment 1. Data pertaining to the reproductive axis of these animals are presented in Figures 2-5. Animals in which SD-induced testicular regressionwas either completely or partially blocked are indicated by * and #, respectively.
could they be explained on some basis other than interfer-ence with photoperiodism proper such as nonspecific stim-ulation of the reproductive axis? Second, if the photoperi-odic response to SD as such was indeed abolished by theanterior BNST lesion, at what point in the generation andprocessing of the SD signal does this region exert an influ-ence?
First, it may be argued that the BNST lesion did nottruly impair the response to SD but rather nonspecifically
stimulated the neuroendocrine reproductive axis. The ef-fects of this lesion would thus be analogous to those ofolfactory bulbectomy in the hamster: while bulbectomy im-pairs SD-induced gonadal regression [9, 10], the same le-sion also disinhibits gonadotropins to some extent in ham-sters under both LD and SD [11], and it has therefore beenargued that this disinhibitory effect masks a partially orentirely preserved SD response [10-12]. In the case of theBNST, one mechanism for such nonspecific disinhibition
FIG. 7. Photomicrograph depicting a rep-resentative lesion of the BNST, using im-munocytochemistry for GFAP (right panel);arrows are used to delineate the extent ofthe lesion. Location of the lesion is alsoshown by reference to dense immuno-staining of VIP and TH fibers (left and mid-dle panels, respectively). ac, anterior com-missure. x12.
801

RAITIERE ET AL.
300
c 250
o 200 7
50
,o 100oco 50
t-
co
U4-
4
3
2
1
0
0 Z o Cm 0Q O
FIG. 8. Effect of radiofrequency-current lesions on body and testicularweights in adult hamsters that had been previously maintained under SDphotoperiods for 12 wk. At the start of the experiment (experiment 2), allof the animals had regressed testes, with a diameter of < 5 mm. Theanimals were then either lesioned in the anterior region of the BNST orserved as sham-lesioned controls (Cont). Half of the animals in eachgroup were then either exposed to LD photoperiods or remained underSD photoperiods; all weights were determined 4 wk later. Each bar rep-resents the mean; the SEMs are depicted by vertical lines; the number ofanimals per group is indicated by the adjacent numbers. Values markedwith the same letter indicate that no significant difference was detectedbetween them (p > 0.05; ANOVA followed by Newman-Keuls test).
might involve the trauma-driven release of growth factorsknown to possess a direct stimulatory effect on GnRH cellbodies [31-33]. The proximity of the BNST to several pop-ulations of GnRH neurons, particularly those in the adjoin-ing MPOA [34], lends credence to this possibility.
However, the results of the current study argue stronglyagainst the hypothesis of nonspecific disinhibition, whetherthrough release of progonadotropic growth factors orthrough some other mechanism. First, an anterior BNSTlesion closely resembling one that blocked regression in thehamster transferred from LD to SD (experiment 1) had vir-tually no effect on the reproductive status of the fully in-voluted hamster remaining in SD (experiment 2). Had theanterior BNST lesion been directly stimulatory to GnRHcell bodies, one would have expected a rapid increase ingonadotropins and testicular size over the 4-wk post-lesioninterval, but no increase was noted during that period. Sec-ond, we found no evidence that the size of the lesionsplayed a role in determining their effect on the reproductiveaxis. If trauma-generated progonadotropic growth factorswere stimulating GnRH production and release, one wouldexpect that a large lesion would have stimulated gonado-tropin levels to a greater extent than a small one. The op-posite was found: large lesions rostral or caudal to the an-terior BNST and (in the case of the posterior BNST) equal-ly close to GnRH neurons were associated with less, notmore, impairment of the regressive response than lesionsof the same magnitude or smaller centered in the anteriorBNST Third, no difference was noted in the quantity of
800
E 600
c'400
(/)U.
200
150
125
E 100
: 75
.J 50
25
E
U-
v
0-
i)
e)I.-
6
5
4
3
2
1
0
0 (C (/)Z 0 Z
C) -o
FIG. 9. Effect of radiofrequency-current lesions on the neuroendocrinereproductive axis of adult hamsters previously maintained in a sexuallyquiescent condition under SD photoperiods (experiment 2). The hormonemeasurements were made on terminal plasma samples obtained after 4subsequent weeks of exposure to either SD or LD photoperiods. Each barrepresents the mean plasma hormone concentration from 5-7 animals;the SEMs are depicted by vertical lines. Values marked with the sameletter indicate that no significant difference was detected between them(p > 0.05; ANOVA followed by Newman-Keuls test).
GFAP expression at lesion borders in fully regressed vs.nonregressing hamsters, again suggesting that trauma-driv-en changes (insofar as these are reflected in GFAP expres-sion) bore little relationship to reproductive status. Fourth,it has recently been shown that electrolytic lesions directlywithin the MPOA itself, one of the primary GnRH cell-containing loci, do not interfere significantly with the Syr-ian hamster's reproductive response to SD [5], essentiallyruling out the hypothesis that generation of progonadotrop-ic growth factors by small lesions in immediate proximityto GnRH cells can overcome the hamster's SD photoperi-odic response, and also highlighting the markedly site-spe-cific effect of the lesion in the adjoining anterior BNSTwhich abolishes that response. It should be emphasized thatlack of any obvious nonspecific stimulatory effect resultingfrom the anterior BNST lesion does not necessarily rule outthe possibility that this lesion may have a subtle stimulatoryinfluence in addition to its primary effect of blocking theinhibitory SD response. Overall, we conclude that an an-terior BNST lesion in the present study truly interfered withthe transmission of the SD photoperiodic signal to the re-
b
802

BNST LESIONS IN HAMSTERS
FIG. 10. Effect of radiofrequency-current lesions on the histological ap-pearance of testes from hamsters that had been previously maintained ina sexually quiescent condition under SD photoperiods (experiment 2).Thetestes were examined after 4 subsequent weeks of exposure to either SDor LD photoperiods. Note the enlarged diameter of seminiferous tubulesin the testes of the control and BNST-lesioned hamsters that were exposedto LD photoperiods for 4 wk, as well as the presence of spermatozoainside a well-defined lumen (C and D, respectively). In contrast, the sem-iniferous tubules of the SD control and SD BNST-lesioned hamsters areapproximately half the diameter of those in the corresponding LD ani-mals. The testes of the two SD groups also show very little or no sper-matogenic activity, spermatozoa being completely absent from the sem-iniferous tubules and spermatogonia representing the predominant ger-minal cell type (A and B, respectively). Scale bar = 0.2 mm.
productive axis while having little direct stimulatory effecton GnRH neurons in this or other photoperiodic contexts.
If the expected photoinhibitory response of the Syrianhamster to SD was therefore blocked by a small, site-spe-cific lesion in the anterior BNST, at what point in the se-quence of events linking the perception of the SD signal tothe reproductive axis might this region exert its influence?A lesion abolishing the hamster's response to SD may con-ceivably affect either the generation of the melatonin signalor its readout within the medial hypothalamus and furtherprocessing. Although we did not test the former hypothesisdirectly by examining nocturnal melatonin profiles in le-sioned vs. control SD hamsters, it is unlikely that the BNSTlesion has a significant effect on secretion of melatonin.The BNST is extrinsic to the primary pathway driving mel-atonin secretion, i.e., that leading from retina to pinealgland via the retinohypothalamic tract, suprachiasmatic nu-cleus, hypothalamic paraventricular nucleus, and sympa-thetic fibers arising in the thoracic spinal cord. The BNSTdoes provide an input to the hypothalamic paraventricularnucleus [35], ablation of which blocks SD regression byimpairing melatonin synthesis [36-40]. However, the tra-ditional view that the paraventricular nucleus is a necessaryway-station in the pathway controlling melatonin secretionhas been questioned [41]. Further, if the anterior BNST le-sion were functionally equivalent to pinealectomy, onewould have expected rapid gonadal recrudescence in thegroup of fully regressed hamsters lesioned and left in SD(cf. experiment 2), since pinealectomy of fully regressedhamsters left in SD induces premature recrudescence [42].The observed absence of any post-lesion recrudescence un-der SD (see Figs. 8-10) therefore suggests that the anteriorBNST lesion is not functionally equivalent to pinealectomyand consequently that this lesion's blockade of the SD re-sponse cannot be ascribed to an impairment in the gener-ation of the SD melatonin signal.
FIG. 11. Schematic diagrams of coronal hamster brain sections depict-ing the location where radiofrequency-current lesions were centered inexperiment 2. Data pertaining to the reproductive axis of these animalsare presented in Figures 8-10.
It seems more likely that the anterior BNST plays a rolenot in the generation of melatonin but in the transfer ofinformation distal to the readout of the melatonin signal inthe medial hypothalamus. The BNST is well placed to me-diate between this readout and the GnRH-secreting cell sys-tem. First, several subnuclei of the BNST including the VL,VM, and AM receive a dense efferentation from the ven-tromedial hypothalamus [13, 14], the nucleus that probablybears major responsibility for initial readout of the mela-tonin signal destined for the reproductive axis in this spe-cies [5]. Second, the BNST encloses neurons that modulateGnRH secretion [43]. The substrate for such modulationmay involve either short projections from the BNST to theadjoining MPOA [44] or longer projections from the BNSTto the brainstem. The lateral BNST, comprising part of the"central extended amygdala," has been distinguished from
803

RAITIERE ET AL.
the medial BNST, a component of the "medial extendedamygdala," on the ground that the lateral system subservescentral autonomic regulation through extensive projectionsto brainstem loci including the dorsal vagal complex [45,46] and midbrain central gray [47], whereas the medial sys-tem subserves neuroendocrine regulation through intercon-nections with the medial hypothalamus [48]. This distinc-tion, however, breaks down somewhat in the region impli-cated in photoperiodism by experiment 1: the VL subnu-cleus in particular is strongly associated not only with themedial hypothalamus [13, 14, 19] but also with brainstemcenters involved in both neuroendocrine and autonomicregulation. Most relevant to photoperiodism may be pro-jections from the VL and nearby subnuclei that establishdirect synaptic contact with noradrenergic cell bodies in thebrainstem including the Al, A2, and A6 cell groups [15,45, 49]. The Al noradrenergic cell group comprises the pri-mary source of noradrenergic fibers to GnRH cell bodiesin the MPOA and other basal forebrain regions [50-52],and Al noradrenergic outflow represents a well-known pos-itive influence on GnRH secretion [53, 54]. Since seasonalchanges in noradrenergic activity precede and are believedto drive seasonal changes in gonadotropin levels [2, 16, 17],since the key BNST cell bodies projecting to and modu-lating the noradrenergic system seem to be most denselyconcentrated in the anterior division [15, 45, 49], and sincethe influence exerted by cells in this BNST region on nor-adrenergic function seems to be generally inhibitory [55-58], we propose that the DL, VL, and related anteriorBNST subnuclei may contain cell groups that respond tothe SD melatonin signal by inhibiting Al noradrenergicoutflow, an inhibition that subsequently results in decreasedGnRH secretion and gonadal regression.
However, the results of experiments 1 and 2 taken to-gether suggest that BNST neurons, while responsible forthe initial inhibition of GnRH release in SD, are apparentlynot required for maintenance of the regressed state once itis fully achieved, since lesion of these neurons in experi-ment 2 failed to stimulate fully regressed animals remainingin SD. Since the anterior BNST lesions in experiment 2 didnot exactly replicate those in experiment 1, it may be ar-gued that the slight discrepancy in lesion location accountsfor the nonstimulatory effect of the lesions in experiment2. However, the discrepancy seems minimal, for in somecases lesions that abolished the regressive response in ex-periment 1 are nearly identical to those that failed to stim-ulate fully regressed animals remaining in SD (see Results).It seems, therefore, that the differential response betweenhamsters lesioned before SD exposure and those lesionedafter full regression suggests that the role of the BNST islimited to induction rather than to maintenance of the re-gressive state. Indeed, other aspects of hamster photoperi-odism support the view that the orientation of the repro-ductive axis with respect to the SD signal is a dynamicproperty that changes once regression has been achieved.Just as a distinction can be drawn between the inductionand the maintenance of the refractory state [59], one canbe drawn between the induction and the maintenance ofregression. In a sense, the Syrian hamster's eventual refrac-toriness to melatonin [59] may be equivalent to refractori-ness to the anterior BNST signal (if so, the fully regressedhamster may be viewed as having initiated its own BNST"lesion" before introduction of the experimental lesion).Conceivably, gonadal regression itself, apparently a prereq-uisite for induction of the refractory period ([60], furtherdiscussion in [59]) may lead to structural changes both in
the BNST and in brainstem noradrenergic nuclei that wouldcorrelate with refractoriness to the initiating SD signal, asboth the BNST [61-63] and the Al and A2 noradrenergiccell bodies [64] strongly express sex steroid receptors. Fur-ther experiments would be required to determine whetherthe hamsters' changing responsiveness to melatonin maycorrelate with progressive structural change in the anteriorBNST-noradrenergic circuitry.
Finally, the results of experiment 2 suggest that ablationof the anterior BNST cell groups involved in mediating theinitial response to SD has no effect on the regressed ham-ster's ability to respond appropriately to an LD photostim-ulatory signal; indeed, that the lesion neither blocked noraccelerated the photostimulatory response suggests that thisresponse must involve stimulation by a neural circuit en-tirely extrinsic to the lateral cell groups of the anteriorBNST rather than a disabling of the inhibitory circuit. Inthis respect, the two parts of experiment 2, i.e., the nonres-ponse of the regressed animal to the lesion and the appro-priately stimulatory response of the lesioned regressed an-imal to LD, are compatible with each other in that theyindicate that the BNST inhibitory circuit plays no role afterthe achievement of regression.
In summary, the results of the present study demonstratethat ablation of a relatively restricted region centering onthe anterior BNST and including in particular the VL andDL subnuclei profoundly disturbs the ability of reproduc-tively competent hamsters to initiate an inhibitory gonadalresponse to a nonstimulatory photoperiod. However, sucha lesion does not activate the neuroendocrine reproductiveaxis of hamsters previously rendered reproductively qui-escent by 12 wk of SD. Thus, rather than exerting a tonicnegative influence on GnRH cell bodies regardless of pho-toperiod, the anterior BNST subnuclei targeted by this le-sion seem to respond only to the contingency of the SDmelatonin signal by transmitting an inhibitory message tothe GnRH cell system. This inhibitory message may bemediated in part by an SD-triggered damping of noradren-ergic outflow. However, it remains to be determined wheth-er the anterior BNST cells implicated in the formulationand transmission of the photoinhibitory message are relatedto those anterior BNST cells known to communicate recip-rocally with the Al, A2, and A6 noradrenergic brainstemcell groups.
ACKNOWLEDGMENTS
We thank Dr. David Hess and Verna Russell of the ORPRC P30 CoreFacility (NIH Grant HD-18185) for technical assistance with the testos-terone RIA and testicular histology.
REFERENCES
1. Karsch FJ, Bittman EL, Foster DL, Goodman RL, Legan SJ, RobinsonJE. Neural basis of seasonal reproduction. Recent Prog Horm Res1984; 40:185-225.
2. Bartke A, Steger RW. Seasonal changes in the function of the hypo-thalamic-pituitary-testicular axis in the Syrian hamster. Proc Soc ExpBiol Med 1992; 199:139-148.
3. Maywood ES, Buttery RC, Vance GHS, Herbert J, Hastings MH.Gonadal responses of the male Syrian hamster to programmed infu-sions of melatonin are sensitive to signal duration and frequency butnot to signal phase nor to lesions of the suprachiasmatic nuclei. BiolReprod 1990; 43:74-82.
4. Reiter RJ, Dinh DH, De Los Santos R, Guerra JC. Hypothalamic cutssuggest a brain site for the antigonadotropic action of melatonin inthe Syrian hamster. Neurosci Lett 1981; 23:315-318.
5. Maywood ES, Hastings MH. Lesions of the iodomelatonin-bindingsites of the mediobasal hypothalamus spare the lactotropic, but block
804

BNST LESIONS IN HAMSTERS
the gonadotropic response of male Syrian hamsters to short photope-riod and to melatonin. Endocrinology 1995; 136:144-153.
6. Hastings MH, Roberts AC, Herbert J. Neurotoxic lesions of the an-terior hypothalamus disrupt the photoperiodic but not the circadiansystem of the Syrian hamster. Neuroendocrinology 1985; 40:316-324.
7. Raitiere MN, Garyfallou VT, Urbanski HE Lesions in the bed nucleusof the stria terminalis, but not in the lateral septum, inhibit short-photoperiod-induced testicular regression in Syrian hamsters. BrainRes 1995; 705:159-167.
8. Herbert J. Neural systems underlying photoperiodic time measure-ment: a blueprint. Experientia 1989; 45:965-972.
9. Pieper DR, Tang Y-K, Lipski TP, Subramanian MG, Newman SW.Olfactory bulbectomy prevents the gonadal regression associated withshort photoperiod in male golden hamsters. Brain Res 1984; 321:183-186.
10. Clancy AN, Goldman BD, Bartke A, Macrides E Reproductive effectsof olfactory bulbectomy in the Syrian hamster. Biol Reprod 1986; 35:1202-1209.
11. Pieper DR, Lobocki CA, Thompson M, Subramanian MG. The olfac-tory bulbs tonically inhibit serum gonadotropin and prolactin levelsin male hamsters on long or short photoperiod. J Neuroendocrinol1990; 2:707-714.
12. Pieper DR, Unthank PD, Shuttie DA, Lobocki CA, Swann JM, New-man SW, Subramanian MG. Olfactory bulbs influence testosteronefeedback on gonadotropin secretion in male hamsters on long or shortphotoperiod. Neuroendocrinology 1987; 46:318-323.
13. Cantera NS, Simerly RB, Swanson LW. Organization of projectionsfrom the ventromedial nucleus of the hypothalamus: a Phaseolus vul-garis-leukoagglutinin study in the rat. J Comp Neurol 1994; 348:41-79.
14. Arluison M, Brochier G, Vankova M, Leveil V, Villalobos J, TramuG. Demonstration of peptidergic afferents to the bed nucleus of thestria terminalis using local injections of colchicine: a combined im-munohistochemical and retrograde tracing study. Brain Res Bull 1994;34:319-337.
15. Jannsen SA, Ciriello J. Projections from the cardiovascular responsiveregion of bed nucleus of the stria terminalis (BST) to medullary cat-echolaminergic cell groups. Soc Neurosci Abstr 1991; 17:706.
16. Steger RW, Bartke A, Goldman BD. Alterations in neuroendocrinefunction during photoperiod induced testicular atrophy and recrudes-cence in the golden hamster. Biol Reprod 1982; 26:437-444.
17. Steger RW, Bartke A. Temporal sequence of neuroendocrine eventsassociated with the transfer of male golden hamsters from a stimula-tory to a nonstimulatory photoperiod. Biol Reprod 1991; 44:76-82.
18. de Olmos JS, Alheid GE Beltramino CA. Amygdala. In: Paxinos G(ed.), The Rat Nervous System: Forebrain and Midbrain, vol 1. Syd-ney: Academic Press; 1985: 223-334.
19. Moga MM, Saper CB, Gray TS. Bed nucleus of the stria terminalis:cytoarchitecture, immunohistochemistry and projection to the para-brachial nucleus in the rat. J Comp Neurol 1989; 283:315-332.
20. Ju G, Swanson LW. Studies on the cellular architecture of the bednucleus of the stria terminalis in the rat: I. Cytoarchitecture. J CompNeurol 1989; 280:587-602.
21. Gomez DM, Newman SW. Differential projections of the anterior andposterior regions of the medial amygdaloid nucleus in the Syrian ham-ster. J Comp Neurol 1992; 317:195-218.
22. Urbanski HE A role for N-methyl-D-aspartate receptors in the controlof seasonal breeding. Endocrinology 1990; 127:2223-2228.
23. Garyfallou VT, Kohama SG, Urbanski HE Distribution of NMDA andAMPA receptors in the cerebellar cortex of rhesus monkeys. BrainRes 1996; 716:22-28.
24. Phelix CE Liposits Z, Paull WK. Monoamine innervation of bed nu-cleus of stria terminalis: an electron microscopic examination. BrainRes Bull 1992; 28:949-965.
25. Phelix CE Liposits Z, Paull WK. Catecholamine-CRF synaptic inter-action in a septal bed nucleus: afferents of neurons in the bed nucleusof the stria terminalis. Brain Res Bull 1994; 33:109-119.
26. Freedman LJ, Cassell MD. Distribution of dopaminergic fibers in thecentral division of the extended amygdala of the rat. Brain Res 1994;633:243-252.
27. Roder S, Ciriello J. Collateral axonal projections to limbic structuresfrom ventrolateral medullary Al noradrenergic neurons. Brain Res1994; 638:182-188.
28. Eiden LE, Hokfelt T, Brownstein MJ, Palkovits M. Vasoactive intes-tinal polypeptide afferents to the bed nucleus of the stria terminalis inthe rat: an immunohistochemical and biochemical study. Neuroscience1985; 15:999-1013.
29. Petit J-M, Luppi P-H, Peyron C, Rampon C, Jouvet M. VIP-like im-munoreactive projections from the dorsal raphe and caudal linear ra-phe nuclei to the bed nucleus of the stria terminalis demonstrated bya double immunohistochemical method in the rat. Neurosci Lett 1995;193:77-80.
30. Urbanski HE Doan A, Pierce M. Immunocytochemical investigationof luteinizing hormone-releasing hormone neurons in Syrian hamstersmaintained under long or short days. Biol Reprod 1991; 44:687-692.
31. Junier M-P, Ma YJ, Costa ME, Hill DF, Ojeda SR. TGF-oa contributesto the mechanism by which hypothalamic injury induces precociouspuberty. Proc Natl Acad Sci USA 1991; 88:9743-9747.
32. Junier M-P, Wolff A, Hoffman GE, Ma YJ, Ojeda SR. Effect of hy-pothalamic lesions that induce precocious puberty on the morphologicand functional maturation of the luteinizing hormone-releasing hor-mone system. Endocrinology 1992; 131:787-798.
33. Junier M-P, Hill DF, Costa ME, Felder S, Ojeda SR. Hypothalamiclesions that induce female precocious puberty activate glial expressionof the epidermal growth factor receptor gene: differential regulationof alternatively spliced transcripts. J Neurosci 1993; 13:703-713.
34. Jennes L, Stumpf WE. LHRH systems in the brain of the goldenhamster. Cell Tissue Res 1980; 209:239-256.
35. Cullinan WE, Herman JP, Watson SJ. Ventral subicular interactionwith the hypothalamic paraventricular nucleus: evidence for a relay inthe bed nucleus of the stria terminalis. J Comp Neurol 1993; 332:1-20.
36. Lehman ML, Bittman EL, Newman SW. Role of the hypothalamicparaventricular nucleus in neuroendocrine responses to daylength inthe golden hamster. Brain Res 1984; 308:25-32.
37. Nunez AA, Brown MH, Youngstrom TG. Hypothalamic circuits in-volved in the regulation of seasonal and circadian rhythms in malegolden hamsters. Brain Res Bull 1985; 15:149-153.
38. Brown MH, Badura LL, Nunez AA. Axon-sparing lesions of the hy-pothalamic paraventricular nucleus abolish gonadal responses to pho-toperiod in male Syrian hamsters. J Biol Rhythms 1988; 3:59-69.
39. Bittman EL, Crandell RG, Lehman MN. Influences of the paraven-tricular and suprachiasmatic nuclei and olfactory bulbs on melatoninresponses in the golden hamster. Biol Reprod 1989; 40:118-126.
40. Youngstrom TG, Nunez AA. Hypothalamo-spinal pathways and re-sponses to photoperiod in Syrian hamsters. Brain Res Bull 1992; 29:225-229.
41. Morin LP, Blanchard J. Organization of the hamster paraventricularnucleus. J Comp Neurol 1993; 332:341-357.
42. Hoffmann K. The role of the pineal gland in the photoperiodic controlof seasonal cycles in hamsters. In: Follett BK, Follett DE (eds.), Bi-ological Clocks in Seasonal Reproductive Cycles. Bristol: Wright;1981: 237-250.
43. Beltramino C, Taleisnik S. Dual action of electrochemical stimulationof the bed nucleus of the stria terminalis on the release of LH. Neu-roendocrinology 1980; 30:238-242.
44. Simerly RB, Swanson LW. The organization of neural inputs to themedial preoptic nucleus of the rat. J Comp Neurol 1986; 246:321-342.
45. Sofroniew MV. Direct reciprocal connections between the bed nucleusof the stria terminalis and dorsomedial medulla oblongata: evidencefrom immunohistochemical detection of tracer proteins. J Comp Neu-rol 1983; 213:399-405.
46. Gray TS, Magnuson DJ. Neuropeptide neuronal efferents from the bednucleus of the stria terminalis and central amygdaloid nucleus to thedorsal vagal complex in the rat. J Comp Neurol 1987; 262:365-374.
47. Gray TS, Magnuson DJ. Peptide immunoreactive neurons in theamygdala and the bed nucleus of the stria terminalis project to mid-brain central gray in the rat. Peptides 1992; 13:451-460.
48. Alheid GE Heimer L. New perspectives in basal forebrain organiza-tion of special relevance for neuropsychiatric disorders: the striato-pallidal, amygdaloid, and corticopetal components of substantia in-nominata. Neuroscience 1988; 27:1-39.
49. Luppi P-H, Aston-Jones G, Akaoka H, Chouvet G, Jouvet M. Afferentprojections to the rat locus ceruleus demonstrated by retrograde andanterograde tracing with cholera-toxin subunit and Phaseolus vulgaris-leukoagglutinin. Neuroscience 1995; 65:119-160.
50. McKellar S, Loewy AD. Efferent projections of the Al catecholaminecell group in the rat: an autoradiographic study. Brain Res 1982; 241:11-29.
51. Woulfe JM, Flumerfelt BA, Hrycyshyn AW. Efferent connections ofthe Al noradrenergic cell group: a DBH immunohistochemical andPHA-L anterograde tracing study. Exp Neurol 1990; 109:308-322.
52. Leranth C, MacLusky NJ, Shanabrough M, Naftolin E Catecholamin-
805

RAITIERE ET AL.
ergic innervation of luteinizing hormone-releasing hormone and glu-tamic acid decarboxylase immunopositive neurons in the rat medialpreoptic area. Neuroendocrinology 1988; 48:591-602.
53. Barraclough CA, Wise PM, Selmanoff MK. A role for hypothalamiccatecholamines in the regulation of gonadotropin secretion. RecentProg Horm Res 1984: 40:487-529.
54. Barraclough CA. Neurotransmitter regulation of luteinizing hormone-releasing hormone neuronal function. Acta Biol Hung 1994; 45:189-206.
55. Ciriello J, Jannsen SA. Effect of glutamate stimulation of bed nucleusof the stria terminalis on arterial pressure and heart rate. Am J Physiol(Heart Circ Physiol) 1993; 265:H1516-H1522.
56. Giancola SB, Roder S, Ciriello J. Contribution of caudal ventrolateralmedulla to the cardiovascular responses elicited by activation of bednucleus of the stria terminalis. Brain Res 1993; 606:162-166.
57. Gelsema AJ, Copeland NE, Drolet G, Bachelard H. Cardiovasculareffects of neuronal activation of the extended amygdala in rats. BrainRes 1993; 626:156-166.
58. Dunn JD, Williams TJ. Cardiovascular responses to electrical stimu-lation of the bed nucleus of the stria terminalis. J Comp Neurol 1995;352:227-234.
59. Watson-Whitmyre M, Stetson MH. Reproductive refractoriness inhamsters: environmental and endocrine etiologies. In: Stetson MH(ed.), Processing of Environmental Information in Vertebrates. NewYork: Springer-Verlag; 1988: 219-249.
60. Turek FW, Losee SH. Photoperiodic inhibition of the reproductivesystem: a prerequisite for the induction of the refractory period inhamsters. Biol Reprod 1979; 20:611-616.
61. DeVries GJ, Buijs RM, Van Leeuwen FW, Caffe AR, Swaab DE Thevasopressinergic innervation of the brain in normal and castrated rats.J Comp Neurol 1985; 233:236-254.
62. Swann JM, Newman SW. Testosterone regulates substance P withinneurons of the medial nucleus of the amygdala, the bed nucleus ofthe stria terminalis and the medial preoptic area of the male goldenhamster. Brain Res 1992: 590:18-28.
63. Asmus SE, Newman SW. Tyrosine hydroxylase neurons in the malehamster chemosensory pathway contain androgen receptors and areinfluenced by gonadal hormones. J Comp Neurol 1993; 331:445-457.
64. Everitt BJ, Herbert J, Keverne EB. The neuroendocrine anatomy ofthe limbic system: a discussion with special reference to steroid re-sponsive neurons, neuropeptides, and monoaminergic systems. ProgAnat 1983; 3:235-260.
806
















