
Functional Background to Acupuncture Effects in Pain and ...
Upload
sorbonne-frCategory
view
0download
0
UNCORRECTED PROOF
ARTICLE IN PRESS
1Pain modulation triggered by high-intensity
2stimulation: implication for acupuncture analgesia?
3Daniel Le Bars *, Jean-Claude Willer
4INSERM U-161, 2, rue d’Alesia, 75014 Paris, France
5C.H.U. Pitie-Salpetriere Laboratoire de Neurophysiologie, 91 Boulevard de l’Hopital, 75013 Paris, France6
7Abstract
8The spinal transmission of nociceptive signals is modulated by both spinal and supraspinal
9systems. Spinal networks underlie segmental inhibitions, which are mediated via myelinated
10cutaneous afferent fibres activated by mechanical stimuli (e.g. rubbing, etc.) Diffuse noxious
11inhibitory controls (DNIC) are triggered by peripheral Ay- and C-fibres, involve brain structures
12confined to the caudal-most part of the medulla including the subnucleus reticularis dorsalis (SRD)
13and are mediated by descending pathways in the dorsolateral funiculi. Combined psychophysical
14measurements and recordings of nociceptive reflexes (RIII) in both normal volunteers and patients
15with lesions in the spinal cord or brain suggest the existence of identical or equivalent inhibitory
16processes in man. Both these mechanisms could be triggered to produce hypoalgesia by acupuncture
17needles or moxibustion. When applied near a painful focus, the techniques have similarities to high-
18frequency, low-intensity transcutaneous nerve stimulation, which brings segmental inhibitory
19mechanisms into play. When applied to remote areas, the techniques might be related to ‘‘counter-
20irritation’’ or ‘‘counter-stimulation’’ phenomena which, in addition to involving complex autonomic
21functions, seem to involve DNIC. Experimental data in both animals and humans support these
22conclusions. D 2002 Elsevier Science B.V. All rights reserved.23
24Keywords: Acupuncture; Pain; Dorsal horn; Nociceptive reflexes; DNIC
25262728Painful stimuli can diminish, or even mask, pain elicited by stimulation of a remote
29(extrasegmental) part of the body [1–3]. This phenomenon has been known of since
30ancient times as illustrated by the Hippocrates’ aphorism: ‘‘If a patient be subject to two
31pains arising in different parts of the body simultaneously, the stronger blunts the other’’. It
0531-5131/02 D 2002 Elsevier Science B.V. All rights reserved.
PII: S0531 -5131 (02 )00412 -0
* Corresponding author. INSERM U-161, 2, rue d’Alesia, 75014, Paris, France. Tel.: +33-1-40-78-93-50; fax:
+33-1-45-88-13-04.
E-mail address: [email protected] (D. Le Bars).
International Congress Series 1077 (2002) xxx

UNCORRECTED PROOF
ARTICLE IN PRESS
32has even been used during surgical procedures on both human patients and domestic
33animals. In the latter category, two examples are the use of the twitch in horses and of
34nasal forceps in cattle for performing caudectomies or castrations, both of which are
35potentially painful operations. The working hypothesis was developed that some of the
36neurones involved in the transmission of nociceptive signals can be inhibited by
37nociceptive stimulation of peripheral territories outside their own excitatory receptive
38fields. That such an hypothesis is correct at as early a stage as the spinal cord was revealed
39by the finding that some dorsal horn neurones are strongly inhibited when a nociceptive
40stimulus is applied to any part of the body, distinct from their excitatory receptive fields.
41For convenience, this phenomenon was termed ‘‘diffuse noxious inhibitory controls’’
42(DNIC).
431. Diffuse noxious inhibitory controls (DNIC) in animal
44In the rat (see references below), the cat [4,5] and, probably, the monkey [6,7], most
45wide-dynamic-range and some nociceptive-specific neurones can be strongly inhibited by
46noxious inputs applied outside their receptive field. Such effects do not appear to be
47somatotopically organised but apply to the whole body. Conversely, they apply to
48neurones in the dorsal horn of various segments of the spinal cord [8–17] or in either
49nucleus caudalis or nucleus oralis of the trigeminal system [18–21]. The neurones affected
50include some projecting to the thalamus [22]. It should be noted that the inhibitions
51triggered by heterotopic noxious stimuli are highly sensitive to the anaesthetic regimen, an
52observation that could explain some reports of lesser inhibitory effects [6,9,16,23].
53DNIC are triggered by conditioning stimuli applied to any part of the body distant from
54the excitatory receptive field of the neurone under study, provided that the stimuli are
55clearly noxious. There is a clear relationship between the intensity of the conditioning
56stimulus and the strength of the resultant DNIC [24,25]. With strong stimuli, the inhibitory
57effects are powerful indeed and are followed by long-lasting post-stimulus effects, which
58can persist for several minutes. In some cases, the inhibitory effects can involve a complete
59abolition of activity for a long period of time following removal of the conditioning stimuli
60(‘‘switch-off’’) and the activity can be restored to preconditioning levels by further
61manipulations of the excitatory receptive field (‘‘switch-on’’) [26].
62With regard to the viscera, some differences should be noted: visceral stimuli, e.g.
63distension of the colon or urinary bladder, generally produce inhibitions with slower rates
64of onset and recovery but starting with intensities below a painful level [15]. It was
65proposed that these differences may have reflected different amounts and patterns of
66activity in the relevant primary afferent fibres rather than being due to different central
67neural mechanisms.
68In any case, these data suggest that DNIC are triggered specifically by the activation of
69peripheral nociceptors whose signals are carried by A- and C-fibres. In order to further
70investigate the types of peripheral fibres involved in DNIC, we took advantage of the
71facts: (1) that trigeminal and spinal dorsal horn neurones respond with relatively steady
72discharges to the electrophoretic application of excitatory amino-acids, and (2) that DNIC
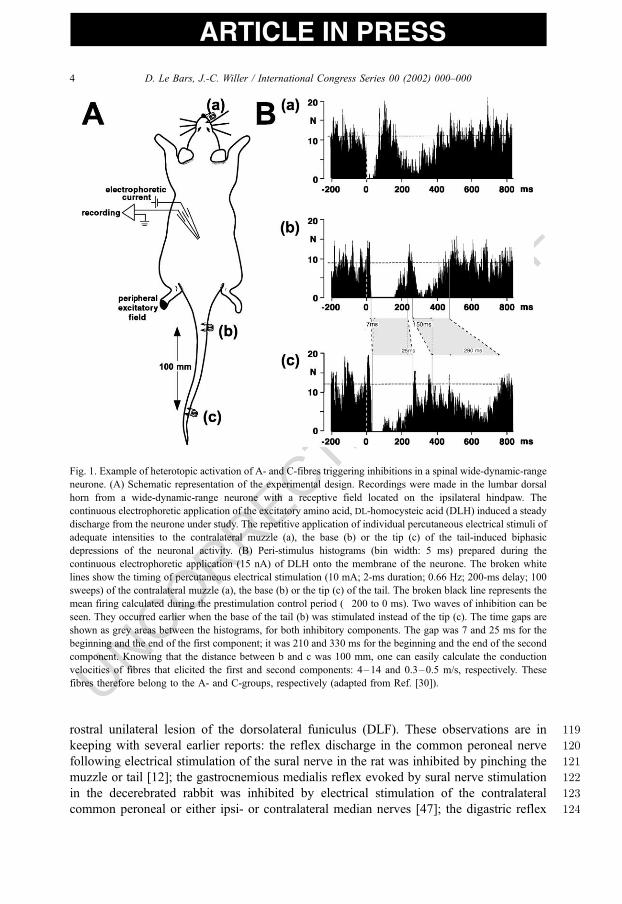
73act by a final postsynaptic inhibitory mechanism involving hyperpolarisation of the
D. Le Bars, J.-C. Willer / International Congress Series 00 (2002) 000–0002

UNCORRECTED PROOF
ARTICLE IN PRESS
74neuronal membrane [27,28]. It was found that when spinal or trigeminal wide-dynamic-
75range neurones were directly excited by the electrophoretic application of glutamate, the
76percutaneous application of single square-wave, electrical stimuli to the tail always
77induced a biphasic depression of the resultant activity [29,30]. Both the early and late
78components of this inhibition occurred with shorter latencies when the base rather than the
79tip of the tail was stimulated. Such differences in latency were used to estimate the mean
80conduction velocities of the peripheral fibres triggering the inhibitions: the means were
81found to be 7.3 and 0.7 m/s, which fall into the A- and C-fibre ranges, respectively. Such
82biphasic inhibitions could be elicited from any part of the body and recorded from any
83wide-dynamic-range neurones. Fig. 1 shows a recording from a lumbar wide-dynamic-
84range neurone with an excitatory receptive field located on the extremity of the ipsilateral
85hind paw: two components of inhibition were induced by the activation of A- and C-fibres,
86respectively, when a single 2-ms duration shock of 10 mA was applied to the muzzle, the
87base or the tip of the tail.
88DNIC are not observed in anaesthetised or decerebrated animals in which the spinal
89cord has been sectioned [5,10,31]. It is therefore obvious that the mechanisms underlying
90DNIC are not confined to the spinal cord and that supraspinal structures must be involved.
91Such a system is therefore completely different from segmental inhibitory systems, which
92work both in intact and in spinal animals, and can be triggered by the activation of low
93threshold afferents [32,33]. DNIC are also very different from the propriospinal inhibitory
94processes which can be triggered by noxious inputs [6,10,34,35].
95The ascending and descending limbs of this loop travel through the ventrolateral and
96dorsolateral funiculi, respectively [36,37]. Since thalamic lesions do not affect DNIC [37],
97it has been proposed that they result from a physiological activation of some of the
98brainstem structures which produce descending inhibition [38]. Surprisingly, DNIC were
99not modified by lesions of the following structures: the PAG, cuneiform nucleus, para-
100brachial area, locus coeruleus/subcoeruleus, RVM [39–41]. By contrast, lesions of
101subnucleus reticularis dorsalis (SRD) in the caudal medulla strongly reduced DNIC
102[42]. The SRD is located ventral to the cuneate nucleus, between trigeminal nucleus
103caudalis and the nucleus of the solitary tract and contains neurones with characteristics
104which suggest that they have a key role in processing specifically nociceptive information
105[43]. Indeed, they are preferentially or exclusively activated by nociceptive stimuli from a
106‘‘whole-body’’ receptive field, they encode the intensity of cutaneous and visceral
107stimulation within noxious ranges and are activated exclusively by activity in cutaneous
108A- or A- and C-fibres. In addition, they send descending projections through the
109dorsolateral funiculus that terminate in the dorsal horn at all levels of the spinal cord.
110The fact that the supraspinal loop sustaining DNIC is confined within the most caudal part
111of the medulla was confirmed in a series of experiments where the potency of DNIC was
112tested in animals with complete transections at different levels of the brainstem [44] (Fig.
1132).
114Interestingly, a C-fibre reflex elicited by electrical stimulation of the sural nerve and
115recorded from the biceps femoris muscle was reported to be strongly inhibited in intact
116anaesthetised rats by both mechanical and thermal noxious heterotopic stimuli whether to
117the muzzle, a paw or the tail, and by colorectal distension [46]. These inhibitory effects
118disappeared when the C-fibre reflex was recorded in spinal animals, or ipsilateral to a
D. Le Bars, J.-C. Willer / International Congress Series 00 (2002) 000–000 3
UNCORRECTED PROOF
ARTICLE IN PRESS
119rostral unilateral lesion of the dorsolateral funiculus (DLF). These observations are in
120keeping with several earlier reports: the reflex discharge in the common peroneal nerve
121following electrical stimulation of the sural nerve in the rat was inhibited by pinching the
122muzzle or tail [12]; the gastrocnemious medialis reflex evoked by sural nerve stimulation
123in the decerebrated rabbit was inhibited by electrical stimulation of the contralateral
124common peroneal or either ipsi- or contralateral median nerves [47]; the digastric reflex
Fig. 1. Example of heterotopic activation of A- and C-fibres triggering inhibitions in a spinal wide-dynamic-range
neurone. (A) Schematic representation of the experimental design. Recordings were made in the lumbar dorsal
horn from a wide-dynamic-range neurone with a receptive field located on the ipsilateral hindpaw. The
continuous electrophoretic application of the excitatory amino acid, DL-homocysteic acid (DLH) induced a steady
discharge from the neurone under study. The repetitive application of individual percutaneous electrical stimuli of
adequate intensities to the contralateral muzzle (a), the base (b) or the tip (c) of the tail-induced biphasic
depressions of the neuronal activity. (B) Peri-stimulus histograms (bin width: 5 ms) prepared during the
continuous electrophoretic application (15 nA) of DLH onto the membrane of the neurone. The broken white
lines show the timing of percutaneous electrical stimulation (10 mA; 2-ms duration; 0.66 Hz; 200-ms delay; 100
sweeps) of the contralateral muzzle (a), the base (b) or the tip (c) of the tail. The broken black line represents the
mean firing calculated during the prestimulation control period (�200 to 0 ms). Two waves of inhibition can be
seen. They occurred earlier when the base of the tail (b) was stimulated instead of the tip (c). The time gaps are
shown as grey areas between the histograms, for both inhibitory components. The gap was 7 and 25 ms for the
beginning and the end of the first component; it was 210 and 330 ms for the beginning and the end of the second
component. Knowing that the distance between b and c was 100 mm, one can easily calculate the conduction
velocities of fibres that elicited the first and second components: 4–14 and 0.3–0.5 m/s, respectively. These
fibres therefore belong to the A- and C-groups, respectively (adapted from Ref. [30]).
D. Le Bars, J.-C. Willer / International Congress Series 00 (2002) 000–0004

UNCORRECTED PROOF
ARTICLE IN PRESS
125evoked by tooth pulp stimulation in the cat was inhibited by toe pinch, percutaneous
126electrical stimulation of a limb or electrical stimulation of the saphenous nerve [48–50].
127These results also fit with the observations in animals of an increase in nociceptive
128thresholds triggered by heterotopic noxious conditioning [1].
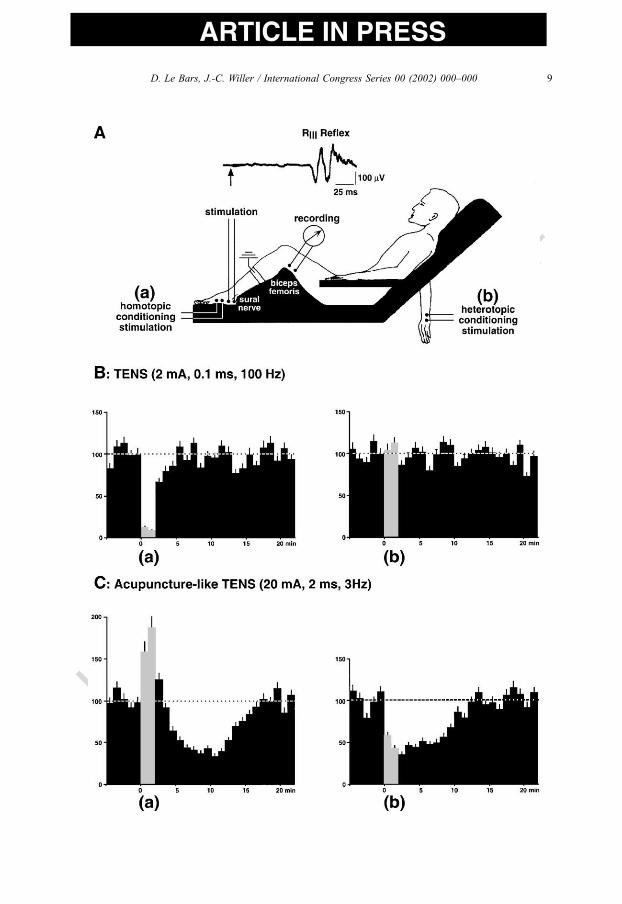
Fig. 2. Triggering of descending inhibitory controls by nociceptive stimulation. When a painful focus appears in a
region of the body, dorsal horn neurones are activated and send an excitatory signal through the ventrolateral
quadrant towards higher centres, including the lower brainstem. This signal activates DNIC, which will inhibit
spinal and trigeminal convergent neurones through the dorsolateral funiculi. The levels of transection that did not
modify DNIC are shown as shaded areas in the upper drawing of a sagittal section of the brainstem. By contrast,
transection caudal to this area blocked DNIC completely, suggesting that the open areas of the drawing
correspond to the parts of the brainstem involved in the upper part of the DNIC circuitry. Abbreviations: 4V,
fourth ventricle; 12, hypoglossal nucleus; AP, area postrema; C, cerebellum; Gi, gigantocellular nucleus; GiA,
gigantocellular nucleus pars alpha; GiV, gigantocellular nucleus ventral; IO, inferior olive; pyx, pyramidal
decussation; RMg, nucleus raphe magnus; RPa, nucleus raphe palidus; ROb, nucleus raphe obscurus (from Ref.
[45], with permission).
D. Le Bars, J.-C. Willer / International Congress Series 00 (2002) 000–000 5

UNCORRECTED PROOF
ARTICLE IN PRESS
1292. Comparison with effects triggered by acupuncture manipulation in animal
130The effects of manual acupuncture performed by a traditional Chinese acupuncturist at
131the ‘‘Zusanli’’ point on the right hind limb were compared with the effects elicited by
132acupuncture applied at a non-acupoint, next to ‘‘Zusanli’’, on responses of trigeminal
133wide-dynamic-range neurones [51]. Similar strong inhibitory effects on the C-fibre-evoked
134responses of these neurones were obtained regardless of whether the acupuncture was
135applied at ‘‘Zusanli’’ or the non-acupoint; indeed, such effects were also produced by
136noxious thermal stimulation (Fig. 3). These results were confirmed and extended to
137moxibustion [52,53]. Interestingly, both the acupuncture- and noxious-evoked inhibitions
138were significantly reduced by systemic naloxone.
139The effects on SRD neurones of manual acupuncture at the ‘‘Renzhong’’, ‘‘Sousanli’’,
140‘‘Changqiang’’ and ‘‘Zusanli’’ acupoints, and at a non-acupoint next to ‘‘Zusanli’’ were
Fig. 3. Comparison of noxious- and acupuncture-induced inhibitions of wide-dynamic-range neurones. (A)
Schematic representation of the experimental design. (A) Recordings were made in the left trigeminal nucleus
caudalis, from a wide-dynamic-range neurone with a receptive field located ipsilaterally on the muzzle. The
responses following percutaneous electrical stimulation of their receptive fields were conditioned by manual
acupuncture applied at the ‘‘Zusanli’’ point (a), at a close, non-acupoint on the right hindpaw (b) or by immersion
of the extremity of the left hindpaw in 48 jC water (c). (B) The histograms represent the temporal evolution
(abscissa: time) of the C-fibre responses (ordinate: number of spikes computed within 60–100 ms following each
stimulus) of the neurone to percutaneous electrical stimulation (2 ms; 0.67 Hz). During the period arrowed in the
upper parts of the figures, manual acupuncture was applied either at ‘‘Zusanli’’ (a) or at a non-acupoint (b) on the
right hindpaw or the left hindpaw was immersed in a 48 jC water bath (c). Note that both noxious heat and
acupuncture induced a depression of neuronal activity followed by after-effects of variable duration (adapted from
Ref. [51]).
D. Le Bars, J.-C. Willer / International Congress Series 00 (2002) 000–0006

UNCORRECTED PROOF
ARTICLE IN PRESS
141also studied [54]. Acupuncture stimulation at the acupoints or the non-acupoint strongly
142excited all the neurones tested (Fig. 4). These neurones responded with a discharge of
143rapid onset, which was often followed by post-stimulus discharges. The levels of induced
144activity were of similar magnitude to those evoked by noxious mechanical stimuli applied
145under identical experimental conditions. No differences were found between the capacities
146to activate the neurones from the ‘‘Zusanli’’ point and the adjacent non-acupoint, no matter
147whether these were stimulated ipsi- or contralaterally; this suggests a lack of topographical
148specificity in the activation of these neurones.
1493. Control of spinal nociceptive reflexes in man
150
1513.1. ‘‘TENS-like acupuncture’’ and ‘‘acupuncture-like TENS’’
152It appears to us to be very meaningful that Sarlandiere [55], one of the first European
153acupuncturists in the early 19th century, had the idea of connecting needles to a galvanic
Fig. 4. Manual acupuncture activates nociceptive neurones. Individual example of responses of a SRD neurone
elicited by 30 s of acupuncture stimulation at points located in different parts of the body (circled). Note that
needle stimulation at all points, including the non-acupoint, induced strong neuronal activity followed by long-
lasting post-stimulus discharges (adapted from Ref. [54]).
D. Le Bars, J.-C. Willer / International Congress Series 00 (2002) 000–000 7

UNCORRECTED PROOF
ARTICLE IN PRESS
154current source to treat rheumatism and gout. Electrical stimulation was then, and has been
155since, widely used in clinical practice to relieve acute or chronic pain [56]. Schematically,
156two types of current are effective in producing significant relief of experimental or clinical
157pain [57–62]. The first, usually termed transcutaneous electrical nerve stimulation
158(TENS), constitutes high-frequency, low-intensity and short-duration pulses applied to
159the skin or cutaneous nerves supplying the painful focus. This nonpainful stimulus
160activates large diameter cutaneous afferents (A-fibres) which convey nonpainful signals.
161The second, often termed ‘‘acupuncture-like’’ TENS, constitutes lower frequency, higher
162intensity and longer duration pulses to the skin or cutaneous nerves supplying the painful
163focus or remote (heterotopic) regions. These currents activate nociceptive (A- and C-)
164fibres and produce unpleasant sensations.
165The effects of these two different types of stimulation applied either segmentally (Fig.
1665A: a) or heterotopically (Fig. 5A: b) to an experimental painful focus were studied on the
167nociceptive flexion RIII reflex, elicited in the lower limb by electrical stimulation of the
168sural nerve [63]. Sural nerve stimulation at 1.2 times threshold elicited a slightly
169supraliminal RIII reflex response, which was found to be affected in various ways by
170TENS, depending on the type and location of the conditioning stimulus.
171TENS produced an inhibitory effect only when applied segmentally to the sural nerve.
172This inhibition was maximal within the first minute of stimulation, remained stable
173throughout the 2-min conditioning period and was followed by significant post-stimulus
174effects lasting 2–3 min (Fig. 5B, left). The heterotopic application of TENS remained
175without effect on the RIII (Fig. 5B, right). TENS elicited pleasant tickling and vibrating
176sensations locally at the stimulation site and did not produce any noticeable modifications
177of the skin under the stimulating electrodes.
178By contrast (Fig. 5C), ‘‘acupuncture-like’’ TENS resulted in biphasic facilitatory–
179inhibitory effects when applied segmentally and only in an inhibition when applied
180heterotopically. ‘‘Acupuncture-like’’ TENS applied for 2 min to the sural nerve produced a
181facilitation of the RIII reflex, which was maximal by the second minute of conditioning.
182This facilitatory effect decreased rapidly when the conditioning stimulation was removed
183and inhibitory post-stimulus effects were clear for a period of 14–15 min (Fig. 5C, left).
184By contrast, heterotopic application of ‘‘acupuncture-like’’ TENS induced a clear-cut
185inhibition of the RIII reflex both during and following the 2-min period of application (Fig.
1865C, right). ‘‘Acupuncture-like’’ TENS was associated with (1) unambiguous, albeit
Fig. 5. Depressive effects of segmental and heterotopic application of transcutaneous electrical nerve stimulation
on a lower limb nociceptive flexion reflex in man. (A) A nociceptive flexion reflex (RIII reflex) elicited in the
lower limb by electrical stimulation of the sural nerve at the ankle was studied before, during and after a 2-min
application of conventional or acupuncture-like transcutaneous electrical nerve stimulation (TENS). Conventional
TENS consisted of low-intensity (2 mA), short-duration (0.1 ms) and high-frequency (100 Hz) rectangular pulses.
Acupuncture-like TENS consisted of high-intensity (20 mA), long-duration (2 ms) and low-frequency (3 Hz)
rectangular pulses. Each conditioning stimulus was applied either homotopically to the sural nerve itself (a) or
heterotopically on the skin overlying the first interosseous space of the contralateral hand (b). (B) Non-noxious
conventional TENS applied segmentally produced inhibitions of the RIII reflex only during the 2-minute
conditioning period (a). Heterotopic TENS had no effect (b). (C) When given segmentally, the acupuncture-like
TENS produced a facilitatory effect during the 2-min application, followed by significant inhibitory post-stimulus
effects (a). The administration of acupuncture-like TENS heterotopically resulted in inhibitions of the RIII reflex
both during and after the 2-min conditioning period (b) (adapted from Ref. [63]).
D. Le Bars, J.-C. Willer / International Congress Series 00 (2002) 000–0008
UNCORRECTED PROOF
ARTICLE IN PRESS
D. Le Bars, J.-C. Willer / International Congress Series 00 (2002) 000–000 9

UNCORRECTED PROOF
ARTICLE IN PRESS
187bearable, painful sensations (muscular cramp, pinprick and burning) at the stimulation site
188and radiating into the receptive field of the stimulated nerve, and (2) the appearance of a
189slight cutaneous erythema, localised in the area surrounding the stimulating electrodes.
190None of the conditioning stimuli elicited any significant fluctuations in heart rate or
191respiration—two very sensitive parameters of emotional reactions.
192
1933.2. Diffuse noxious inhibitory controls in man
194Results reminiscent of what had been found in animals were obtained in man during
195studies which combined psychophysical measurements with the recording of nociceptive
196reflexes [64–68]. Electrical stimulation of the sural nerve at the ankle simultaneously
197induces a nociceptive reflex in a flexor muscle of the thigh (the RIII reflex) and a painful
198sensation in the territory of the nerve [69]. Painful heterotopic cutaneous conditioning
199stimuli, no matter whether it is thermal, mechanical or chemical in nature, increase the
200thresholds of both the reflex and the associated painful sensations, with stronger effects
201being observed with more intense conditioning stimuli. These inhibitory phenomena are
202exerted selectively on nociceptive responses and do not affect monosynaptic reflexes of
203proprioceptive origin. These results demonstrate that in man, a painful conditioning
204stimulus can depress both a preexisting pain and its associated nociceptive reflex at as
205early a stage as the first spinal relays for the transmission of nociceptive information.
206Interestingly, the R2-component of the blink reflex is also inhibited by remote painful heat
207[70,71].
208This prompts the following question: Are the inhibitory mechanisms purely spinal or do
209they involve supraspinal structures? To answer this question, the effects on the RIII reflex
210in the right leg of nociceptive conditioning stimuli applied to the fourth and fifth fingers of
211the left hand were compared in normal subjects and tetraplegic patients with lesions of
212traumatic origin at the C5–C7 level [72]. In the normal subjects, as previously, the painful
213conditioning stimuli caused a strong depression of both the RIII reflex and the associated
Fig. 6. Triggering of DNIC in a patient with a Brown–Sequard syndrome. A patient with Brown–Sequard
syndrome due to a 4-year-old spinal cord hemisection produced by a knife-wound in the back was carefully
investigated. The neurological observations and drawing of the lesion from MRI (left side, T6 level) are
summarised on the left. RIII reflexes elicited by stimulation of cutaneous afferents in the ulnar or sural nerves were
studied in the upper and lower limbs by recording from the biceps brachialis and biceps femoris muscles,
respectively. For each limb, the RIII reflex was elicited regularly by stimuli of 1.2 times threshold, before, during
and after periods of nociceptive electrical conditioning stimulation (15 mA; 4 Hz; 1 min) applied successively to
the other three limbs. The experimental set-up is represented in the centre of each insert, while the results are
shown in three histograms, each of which is adjacent to where the appropriate conditioning stimulus was applied.
In the histograms, each bar represents a single reflex response expressed as a percentage of the mean control (0–2
min) values; reflex responses observed during the conditioning period are shown as grey bars. (A) RIII reflex in
the upper right limb (URL). (B) RIII reflex in the upper left limb (ULL). (C) RIII reflex in the lower right limb
(LRL). (D) RIII reflex in the lower left limb (LLL). Strong inhibitions followed by post-conditioning effects were
observed in all situations except that (1) no inhibition could be obtained when the conditioning stimuli were
applied to the lower right limb (contralateral to the spinal lesion) and (2) the RIII reflex in the lower left limb was
completely insensitive to any of the conditioning stimuli. These results suggest that (1) the ascending part of the
loop subserving DNIC is completely crossed at the spinal level and (2) the descending part is confined to the
white matter ipsilateral to the limb being tested (adapted from Ref. [74]).
D. Le Bars, J.-C. Willer / International Congress Series 00 (2002) 000–00010

UNCORRECTED PROOF
ARTICLE IN PRESS
D. Le Bars, J.-C. Willer / International Congress Series 00 (2002) 000–000 11

UNCORRECTED PROOF
ARTICLE IN PRESS
214pain. By contrast, in the tetraplegic patients, nociceptive stimulation of the same cutaneous
215territories, which, being in the C8 and T1 dermatomes, were clinically unaffected by the
216spinal lesion, did not produce any depression of the RIII reflex recorded in the contralateral
217leg. These results demonstrate that in man, as in animals, the inhibitory effects triggered by
218heterotopic nociceptive stimuli are most likely sustained by a loop which includes
219supraspinal structures.
220These results also lead to a subsidiary question: Is it possible to identify, or at least
221localise, these supraspinal structures? Tentative answers were obtained by observations
222made on patients with cerebral lesions causing contralateral hemianalgesia [73]. These
223were patients with either a unilateral thalamic lesion (identified and delineated by a CT
224scan) or a lesion of the retro-olivary part of the medulla (Wallenberg’s syndrome). In the
225former group, the RIII reflex on the normal side was strongly depressed, as in normal
226subjects, by nociceptive conditioning stimuli applied to the affected side which were not
227felt as painful. By contrast, in the patients with Wallenberg’s syndrome, no inhibitions
228were observed when the nociceptive conditioning stimuli were applied to the affected side,
229whereas if these stimuli were applied to the normal side, they triggered inhibitory effects
230and after-effects very similar to those seen in normal subjects. These results show that in
231humans, thalamic structures and, consequently, spinothalamic pathways are not involved
232in DNIC, whereas brainstem—probably reticular—structures seem to play a key role in
233these phenomena.
234We also reported an exceptional case of a patient with (Fig. 6) Brown–Sequard
235syndrome due to a 4-year-old spinal cord hemisection (left side, T6 level) produced by a
236knife-wound in the back [74]. The RIII reflexes elicited by stimulation of cutaneous
237afferents in the ulnar and sural nerves were studied in the upper and lower limbs by
238recording from the biceps brachialis and biceps femoris muscles, respectively. For each
239limb, the RIII reflex was elicited regularly by stimuli of 1.2 times threshold before, during
240and after periods of nociceptive electrical conditioning stimulation applied successively to
241the other three limbs. Inhibitions of around 90% followed by after-effects were observed in
242all situations except that: (1) no inhibition could be obtained when the conditioning stimuli
243were applied to the lower right limb (contralateral to the spinal lesion), and (2) the RIII
244reflex in the lower left limb was completely insensitive to any of the conditioning stimuli.
245These results suggest that in humans: (1) the ascending part of the loop subserving DNIC
246is completely crossed at the spinal level, and (2) the descending part is confined to the
247white matter, ipsilateral to the limb being tested.
248Thus, the following features are shared by the RIII reflex and associated painful
249sensations in humans and by dorsal horn wide-dynamic-range neurones in the rat spinal
250cord. (1) The RIII reflex and the responses of wide-dynamic-range neurones to electrical
251stimulation of their cutaneous receptive fields are similarly inhibited by various hetero-
252topic nociceptive stimuli. (2) The extent of the inhibitions is directly related to the intensity
253of the conditioning stimulus. (3) The inhibitions are followed by after-effects, which can
254last for several minutes. (4) The inhibitions are mediated by a spino–bulbo–spinal loop,
255the ascending part of which is composed of the spinoreticular tract and synaptic relays in
256the brainstem. (5) The ascending pathways of the loop are mainly crossed while the
257descending pathways run ipsilaterally to the recording site. (6) There is at least one
258opioidergic link in this loop both in the rat and in man [75,76]. Such similarities allow one
D. Le Bars, J.-C. Willer / International Congress Series 00 (2002) 000–00012

UNCORRECTED PROOF
ARTICLE IN PRESS
259to conclude that the inhibitory processes observed in man, and DNIC in the rat, share
260common mechanisms; the existence of DNIC in humans is therefore more than likely.
2614. Summary and conclusions
262The question arises as to what mechanisms underlie the hypoalgesic effects of
263acupuncture, electroacupuncture and moxibustion, and whether they might share common
264neuronal substrates with other means of relieving pain. As stated above, it seems possible
265to distinguish at least two means of producing hypoalgesia by somatic stimulation. In
266some cases, acupuncture probably activates such processes, especially when the needles
267are applied in the vicinity of the painful focus. The participation of large A-afferent fibres
268is then very likely [32,33,57,77]. However, the mechanisms underlying TENS and
269acupuncture at sites either close or remote to the source of pain cannot be explained on
270the basis of a single mechanism [78–81]. It has been shown that stronger analgesic effects
271can be elicited by using a critical level of stimulation which produces an unpleasant
272sensation [82,83], suggesting that A-fibre activation is important in the production of
273analgesia by somatic stimulation. Indeed, numerous studies have demonstrated that the
274activation of A-fibres: (1) produces the most powerful segmental inhibitions and (2)
275induces extrasegmental inhibitions [84–88]. We have also already mentioned the existence
276of propriospinal inhibitory processes triggered by noxious inputs [6,10,34,35].
277The pain relief produced by acupuncture can have a widespread distribution when the
278stimulation is strong enough to induce the feeling of ‘‘Teh-Chi’’ (or ‘‘de qi’’, ‘‘needling
279sensation’’), which is an unpleasant ‘‘heavy-dull’’ sensation that is probably related to
280activity in thin peripheral fibres [89]. Mann [90] concluded that for effective analgesia, the
281sensation resulting from the stimulation point should be the maximum that the patient can
282support. As stated by MacDonald [91], ‘‘over the two millennia that acupuncture has been
283practised in China, a variety of vigorous needling methods have been codified in
284descriptive traditional terms such as ‘mountain-burning fire’, ‘penetrating heaven cool-
285ness’, ‘dragon and tiger joined in battle’ and ‘green dragon wags its tail’’’ [92].
286Furthermore, it has been shown that antinociceptive effects elicited by acupuncture are
287stronger when the stimulation intensities are sufficient to recruit A- and C-fibres [85,93],
288including Capsaicine-sensitive polymodal nociceptors [94,95].
289A role for endogenous opioid systems in the analgesic effects of acupuncture is very
290likely [96–98]. Naloxone reversed acupuncture analgesia [99,100] and the effects
291triggered by low-frequency but not high-frequency electroacupuncture [101–103]. Enke-
292phalins might be involved in these effects since: (1) manual acupuncture stimulation of the
293‘‘Zusanli’’ point (or an adjacent non-acupoint) induces a heterosegmental release of Met-
294enkephalin-like material in the rat spinal cord [104] and (2) low-frequency electro-
295acupuncture elicits a rise of Met-enkephalin-like material in the cerebrospinal fluid of
296patients [105]. One can emphasise the close relationship between the neural systems
297involved in pain modulation and those involved with autonomic functions [106,107].
298Acupuncture and electroacupuncture produce complex changes in autonomic functions in
299both animals and man [108–110]. This is probably achieved through spinal and supra-
300spinal somato-vegetative reflexes—the latter including spino–bulbo–spinal pathways
D. Le Bars, J.-C. Willer / International Congress Series 00 (2002) 000–000 13

UNCORRECTED PROOF
ARTICLE IN PRESS
301which travel in the dorsolateral funiculi [111]. Such functional connections might play a
302key role in acupuncture analgesia (see Sato, this volume).
303We have also proposed that DNIC may well form a neural basis for the pain-relieving
304effects of procedures related to ‘‘counter-stimulation’’ or ‘‘counter-stimulation’’ phenom-
305ena in which fine afferent fibres play an important role [1,2]. Interestingly, DNIC—
306whether tested on wide-dynamic-range neurones in animals or the nociceptive reflex RIII
307in man—are blocked by naloxone [75,76]. In addition, regardless of whether inhibitions of
308trigeminal wide-dynamic-range neurones are triggered by noxious heat or by manual
309stimulation of the ‘‘Zusanli’’ point, they are reduced to a similar extent by naloxone [51].
310In brief, the inhibitory effects elicited by acupuncture on wide-dynamic-range neurones:
311(1) had a similar magnitude and time-course to those evoked by noxious stimulation, (2)
312exhibited a lack of topographical specificity and (3) involved an opioidergic link. This
313suggests that acupuncture manoeuvres trigger the neuronal mechanisms involved in
314diffuse noxious inhibitory controls, at least under our experimental conditions. Since
315SRD neurones, which are activated exclusively or preferentially by noxious inputs, are
316also activated by manipulation of needles, the conclusion is reached that the signals
317elicited by manual acupuncture travel through pathways responsible for the transmission
318of nociceptive information.
319It is possible that the activation of A-fibres produces a more synchronised input to the
320central nervous system than does the activation of C-fibres, and thus produces a more
321efficacious drive of DNIC for a given stimulus intensity. During electroacupuncture, the
322practitioner generally progressively increases the current intensity applied through the
323needles. This makes the stimulus itself bearable by the patient. The unpleasantness of
324vigorous needling is less in patients complaining of pain that one would expect on the
325basis of corresponding sensations elicited in normal subjects, probably because DNIC is
326already triggered by the preexisting painful focus [91].
327The diffuse nature of the effects of acupuncture has been underlined on several
328occasions. We will not address here the questions of the specificity and size of acupoints
329and their relationship with tender and trigger points (‘‘Ah-Shi points’’). These topics were
330summarised in splendid reviews [91,112–114]. ‘‘One only has a look at modern charts to
331realise there is hardly a site on the body that is not an acupuncture point’’ [91].
332Acknowledgements
333The authors are very grateful to Dr. Cadden for advice in the preparation of the
334manuscript.
335References
336[1] D. Le Bars, B. Calvino, L. Villanueva, S. Cadden, Physiological approaches to counter-irritation phenomena,
337in: M.D. Tricklebank, G. Curzon (Eds.), Stress-induced analgesia, Wiley, Chichester, 1984, pp. 67–101.
338[2] D. Le Bars, J.C. Willer, T. De Broucker, L. Villanueva, Neurophysiological mechanisms involved in the
339pain-relieving effects of counter-irritation and related techniques including acupuncture, in: G. Stux, B.
340Pomeranz (Eds.), Scientific Bases of Acupuncture, Springer-Verlag, Berlin, 1989, pp. 79–112.
D. Le Bars, J.-C. Willer / International Congress Series 00 (2002) 000–00014

UNCORRECTED PROOF
ARTICLE IN PRESS
341[3] R. Melzack, Folk medicine and the sensory modulation of pain, in: P.D. Wall, R. Melzack (Eds.), Textbook
342of Pain, Churchill Livingstone, Edinburgh, 1989, pp. 897–905.
343[4] C.R. Morton, H.J. Du, H.M. Xiao, B. Maisch, M. Zimmermann, Inhibition of nociceptive responses of
344lumbar dorsal horn neurones by remote noxious afferent stimulation in the cat, Pain 34 (1988) 75–83.
345[5] C.R. Morton, B. Maisch, M. Zimmermann, Diffuse noxious inhibitory controls of lumbar spinal neurons
346involve a supraspinal loop in the cat, Brain Res. 410 (1987) 347–352.
347[6] K.D. Gerhart, R.P. Yezierski, G.J. Giesler Jr., W.D. Willis, Inhibitory receptive fields of primate spinotha-
348lamic tract cells, J. Neurophysiol. 46 (1981) 1309–1325.
349[7] T.J. Brennan, U.T. Oh, S.F. Hobbs, D.W. Garrison, R.D. Foreman, Urinary bladder and hindlimb afferent
350input inhibits activity of primate T2–T5 spinothalamic tract neurons, J. Neurophysiol. 61 (1989) 573–588.
351[8] D. Le Bars, A.H. Dickenson, J.M. Besson, Diffuse noxious inhibitory controls (DNIC): I. Effects on dorsal
352horn wide-dynamic-range neurones in the rat, Pain 6 (1979) 283–304.
353[9] R.W.W. Tomlinson, B.G. Gray, J.O. Dostrovsky, Inhibition of rat spinal cord dorsal horn neurons by non-
354segmental, noxious cutaneous stimuli, Brain Res. 279 (1983) 291–294.
355[10] S.W. Cadden, L. Villanueva, D. Chitour, D. Le Bars, Depression of activities of dorsal horn wide-dynamic-
356range neurones by propriospinal mechanisms triggered by noxious inputs: comparison with diffuse noxious
357inhibitory controls (DNIC), Brain Res. 275 (1983) 1–11.
358[11] B. Calvino, L. Villanueva, D. Le Bars, The heterotopic effects of visceral pain: behavioural and electro-
359physiological approaches in the rat, Pain 20 (1984) 261–271.
360[12] J. Schouenborg, A.H. Dickenson, The effects of a distant noxious stimulation on A and C fibre-evoked
361flexion reflexes and neuronal activity in the dorsal horn of the rat, Brain Res. 328 (1985) 23–32.
362[13] A. Fleischmann, G. Urca, Clip-induced analgesia: noxious neck pinch suppresses spinal and mesencephalic
363neural responses to noxious peripheral stimulation, Physiol. Behav. 46 (1989) 151–157.
364[14] G.D. Sher, D. Mitchell, Intrathecal N-methyl-D-aspartate induces hyperexcitability in rat dorsal horn
365convergent neurones, Neurosci. Lett. 119 (1990) 199–202.
366[15] S.W. Cadden, J.F.B. Morrison, Effects of visceral distension on the activities of neurones receiving cuta-
367neous inputs in the rat lumbar dorsal horn; comparison with effects of remote noxious somatic stimuli,
368Brain Res. 558 (1991) 63–74.
369[16] T.J. Ness, G.F. Gebhart, Interactions between visceral and cutaneous nociception in the rat: I. Noxious
370cutaneous stimuli inhibit visceral nociceptive neurones and reflexes, J. Neurophysiol. 66 (1991) 20–28.
371[17] T.J. Ness, G.F. Gebhart, Interactions between visceral and cutaneous nociception in the rat: II. Noxious
372visceral stimuli inhibit cutaneous nociceptive neurones and reflexes, J. Neurophysiol. 66 (1991) 29–39.
373[18] A.H. Dickenson, D. Le Bars, J.M. Besson, Diffuse noxious inhibitory controls (DNIC): effects on trige-
374minal nucleus caudalis neurones in the rat brain, Brain Res. 200 (1980) 293–305.
375[19] R. Dallel, P. Raboisson, A. Woda, B. Sessle, Properties of nociceptive and non-nociceptive neurons in
376trigeminal subnucleus oralis of the rat, Brain Res. 521 (1990) 95–106.
377[20] J.W. Hu, Response properties of nociceptive and non-nociceptive neurons in the rat’s trigeminal subnu-
378cleus caudalis (medullary dorsal horn) related to cutaneous and deep craniofacial afferent stimulation and
379modulation by diffuse noxious inhibitory controls, Pain 41 (1990) 331–345.
380[21] I.D. Meng, J.W. Hu, A.P. Benetti, D.A. Bereiter, Encoding of corneal input in two distinct regions of the
381spinal trigeminal nucleus in the rat: cutaneous receptive field properties, responses to thermal and chemical
382stimulation, modulation by diffuse noxious inhibitory controls, and projections to the parabrachial area, J.
383Neurophysiol. 77 (1997) 43–56.
384[22] A.H. Dickenson, D. Le Bars, Diffuse noxious inhibitory controls (DNIC) involve trigeminothalamic and
385spinothalamic neurones in the rat, Exp. Brain Res. 49 (1983) 174–180.
386[23] G. Alarcon, F. Cervero, Effects of two anesthetic regimes on the heterotopic inhibition of rat dorsal horn
387neurones, J. Physiol. (London) 416 (1989) 19P.
388[24] D. Le Bars, D. Chitour, A.M. Clot, The encoding of thermal stimuli by diffuse noxious inhibitory controls
389(DNIC), Brain Res. 230 (1981) 394–399.
390[25] L. Villanueva, D. Le Bars, The encoding of thermal stimuli applied to the tail of the rat by lowering the
391excitability of trigeminal wide-dynamic-range neurones, Brain Res. 330 (1985) 245–251.
392[26] S.W. Cadden, The ability of inhibitory controls to ‘switch-off’ activity in dorsal horn convergent neurones
393in the rat, Brain Res. 628 (1993) 65–71.
D. Le Bars, J.-C. Willer / International Congress Series 00 (2002) 000–000 15

UNCORRECTED PROOF
ARTICLE IN PRESS
394[27] L. Villanueva, S.W. Cadden, D. Le Bars, Diffuse noxious inhibitory controls (DNIC): evidence for post-
395synaptic inhibition of trigeminal nucleus caudalis wide-dynamic-range neurones, Brain Res. 321 (1984)
396165–168.
397[28] L. Villanueva, S.W. Cadden, D. Le Bars, Evidence that diffuse noxious inhibitory controls (DNIC) are
398mediated by a final post-synaptic inhibitory mechanism, Brain Res. 298 (1984) 67–74.
399[29] D. Bouhassira, D. Le Bars, L. Villanueva, Heterotopic activation of A?- and C-fibres triggers inhibition of
400trigeminal and spinal convergent neurones in the rat, J. Physiol. (London) 389 (1987) 301–317.
401[30] D. Le Bars, The receptive fields of dorsal horn neurones: a simple question? in: F. Cervero, G.J. Bennett,
402P.M. Headley (Eds.), Processing of Sensory Information in the Superficial Dorsal Horn of the Spinal Cord,
403NATO ASI Series, Series A, vol. 176, Plenum, New York, 1989, pp. 167–176.
404[31] D. Le Bars, A.H. Dickenson, J.M. Besson, Diffuse noxious inhibitory controls (DNIC): II. Lack of effect
405on non wide-dynamic-range neurones, supraspinal involvement and theoretical implications, Pain 6 (1979)
406305–327.
407[32] P. Hillman, P.D. Wall, Inhibitory and excitatory factors influencing the receptive fields of lamina 5 spinal
408cord cells, Exp. Brain Res. 9 (1969) 284–306.
409[33] H.O. Handwerker, A. Iggo, M. Zimmermann, Segmental and supraspinal actions on dorsal horn neurons
410responding to noxious and non-noxious skin stimuli, Pain 1 (1975) 147–165.
411[34] M. Fitzgerald, The contralateral input to the dorsal horn of the spinal cord in the decerebrate spinal rat,
412Brain Res. 236 (1982) 275–287.
413[35] S. McGaraughty, J.L. Henry, Effects of noxious hindpaw immersion on evoked and spontaneous firing of
414contralateral convergent dorsal horn neurons in both intact and spinalized rats, Brain Res. Bull. 43 (1997)
415263–267.
416[36] L. Villanueva, M. Peschanski, B. Calvino, D. Le Bars, Ascending pathways in the spinal cord involved in
417triggering of diffuse noxious inhibitory controls (DNIC) in the rat, J. Neurophysiol. 55 (1986) 34–55.
418[37] L. Villanueva, D. Chitour, D. Le Bars, Involvement of the dorsolateral funiculus in the descending spinal
419projections responsible for diffuse noxious inhibitory controls in the rat, J. Neurophysiol. 56 (1986) 1185–
4201195.
421[38] H.L. Fields, J.M. Besson (Eds.), Pain Modulation, Prog. Brain Res., vol. 77, Elsevier, Amsterdam, 1988.
422[39] D. Bouhassira, Z. Bing, D. Le Bars, Studies of the brain structures involved in diffuse noxious inhibitory
423controls: the mesencephalon, J. Neurophysiol. 64 (1990) 1713–1723.
424[40] D. Bouhassira, Z. Bing, D. Le Bars, Effects of lesions of locus coeruleus/subcoeruleus on diffuse noxious
425inhibitory controls in the rat, Brain Res. 571 (1992) 140–144.
426[41] D. Bouhassira, Z. Bing, D. Le Bars, Studies of the brain structures involved in diffuse noxious inhibitory
427controls: the rostral ventromedial medulla, J. Physiol. (London) 463 (1993) 667–687.
428[42] D. Bouhassira, L. Villanueva, Z. Bing, D. Le Bars, Involvement of the subnucleus reticularis dorsalis in
429diffuse noxious inhibitory controls, Brain Res. 595 (1992) 353–357.
430[43] L. Villanueva, D. Bouhassira, D. Le Bars, The medullary subnucleus reticularis dorsalis (SRD) as a key
431link in both the transmission and modulation of pain signals, Pain 67 (1996) 231–240.
432[44] D. Bouhassira, D. Chitour, L. Villanueva, D. Le Bars, The spinal transmission of nociceptive information:
433modulation by the caudal medulla, Neuroscience 69 (1995) 931–938.
434[45] D. Le Bars, D. Bouhassira, L. Villanueva, Opioids and diffuse noxious inhibitory controls in the rat, in: B.
435Bromm, J.E. Desmedt (Eds.), Pain and the Brain: From Nociceptor to Cortical Activity, Advances in Pain
436Research and Therapy, vol. 22, Raven Press, New York, 1995, pp. 517–539.
437[46] S. Falinower, C. Willer, J.L. Junien, D. Le Bars, A C-fibre reflex modulated by heterotopic somatic stimuli
438in the rat, J. Neurophysiol. 72 (1994) 194–213.
439[47] J.S. Taylor, R.I. Neal, J. Harris, T.W. Ford, R.W. Clarke, Prolonged inhibition of a spinal reflex after
440intense stimulation of distant peripheral nerves in the decerebrated rabbit, J. Physiol. (London) 437 (1991)
44171–83.
442[48] S.W. Cadden, The digastric reflex evoked by tooth-pulp stimulation in the cat and its modulation by stimuli
443applied to the limbs, Brain Res. 336 (1985) 33–43.
444[49] R.W. Clarke, B. Matthews, The effects of anaesthetics and remote noxious stimuli on the jaw-opening
445reflex evoked by tooth-pulp stimulation in the cat, Brain Res. 327 (1985) 105–111.
446[50] D. Banks, M. Kuriakose, B. Matthews, Modulation by peripheral conditioning stimuli of the responses of
D. Le Bars, J.-C. Willer / International Congress Series 00 (2002) 000–00016

UNCORRECTED PROOF
ARTICLE IN PRESS
447trigeminal brain stem neurones and of the jaw opening reflex to tooth pulp stimulation in chronically
448prepared, anaesthetized cats, Exp. Physiol. 77 (1992) 343–349.
449[51] Z. Bing, L. Villanueva, D. Le Bars, Acupuncture and diffuse noxious inhibitory controls: naloxone
450reversible depression of activities of trigeminal convergent neurones, Neuroscience 37 (1990) 809–818.
451[52] T. Hashimoto, H. Akita, S. Aikawa, Analgesia induced by manual acupuncture: its potency and implica-
452tion, Kitasato Arch. Exp. Med. 65 (1993) 73–82.
453[53] K. Murase, K. Kawakita, Diffuse noxious inhibitory controls in anti-nociception produced by acupuncture
454and moxibustion on trigeminal caudalis neurons in rats, Jpn. J. Physiol. 50 (2000) 133–140.
455[54] Z. Bing, L. Villanueva, D. Le Bars, Acupuncture-evoked responses of subnucleus reticularis dorsalis
456neurons in the rat medulla, Neuroscience 44 (1991) 693–703.
457[55] J.B. Sarlandiere, Memoire sur l’electropuncture, considee comme moyen nouveau de traiter efficacement la
458goutte, les rhumatismes et les affections nerveuses, et sur l’emploi du moxa japonais en France, suivis d’un
459traite de l’acupuncture et du moxa, principaux moyens curatifs chez les peuples de la Chine, de la Coree et
460du Japon. Paris 1825.
461[56] K. Kane, A. Taub, A history of local electrical analgesia, Pain 45 (1975) 125–138.
462[57] P.D. Wall, W.H. Sweet, Temporary abolition of pain in man, Science 155 (1967) 108–109.
463[58] D.M. Long, N. Hagfors, Electrical stimulation in the nervous system: the current status of electrical
464stimulation of the nervous system for the relief of pain, Pain 1 (1975) 109–124.
465[59] J.N. Campbell, A. Taub, Local analgesia from percutaneous electrical stimulation, Arch. Neurol. (Chicago)
46628 (1973) 347–350.
467[60] S.A. Andersson, E. Holmgren, On acupuncture analgesia and the mechanism of pain, Am. J. Chin Med. 3
468(1975) 311–334.
469[61] R.J. Ignelzi, J.K. Nyquist, Direct effect of electrical stimulation on peripheral nerve evoked activity.
470Implications in pain relief, J. Neurosurg. 45 (1976) 159–165.
471[62] P. Hiedl, A. Struppler, A. Gessler, Local analgesia by percutaneous electrical stimulation of sensory nerves,
472Pain 7 (1979) 129–134.
473[63] N. Danziger, S. Rozenberg, P. Bourgeois, G. Charpentier, J.C. Willer, Depressive effects of segmental and
474heterotopic application of transcutaneous electrical nerve stimulation and piezo-electric current on lower
475limb nociceptive flexion reflex in human subjects, Arch. Phys. Med. Rehabil. 79 (1998) 191–200.
476[64] J.C. Willer, A. Roby, D. Le Bars, Psychophysical and electrophysiological approaches to the pain relieving
477effect of heterotopic nociceptive stimuli, Brain 107 (1984) 1095–1112.
478[65] J.C. Willer, A. Barranquero, M.F. Kahn, D. Salliere, Pain in sciatica depresses lower limb cutaneous
479reflexes to sural nerve stimulation, J. Neurol., Neurosurg. Psychiatry 50 (1987) 1–5.
480[66] J.C. Willer, T. De Broucker, D. Le Bars, Encoding of nociceptive thermal stimuli by diffuse noxious
481inhibitory controls in humans, J. Neurophysiol. 62 (1989) 1028–1038.
482[67] C.R. France, S. Suchowiecki, A comparison of diffuse noxious inhibitory controls in men and women, Pain
48381 (1999) 77–84.
484[68] G. Sandrini, I. Milanov, S. Malaguti, M.P. Nigrelli, A. Moglia, G. Nappi, Effects of hypnosis on diffuse
485noxious inhibitory controls, Physiol. Behav. 69 (2000) 295–300.
486[69] J.C. Willer, Comparative study of perceived pain and nociceptive flexion reflex in man, Pain 3 (1977) 69–
48780.
488[70] T. Pantaleo, R. Duranti, F. Bellini, Effects of heterotopic ischemic pain on muscular pain threshold and
489blink reflex in humans, Neurosci. Lett. 85 (1988) 56–60.
490[71] J. Ellrich, R.D. Treede, Characterization of blink reflex interneurons by activation of diffuse noxious
491inhibitory controls in man, Brain Res. 803 (1998) 161–168.
492[72] A. Roby-Brami, B. Bussel, J.C. Willer, D. Le Bars, An electrophysiological investigation into the pain-
493relieving effects of heterotopic nociceptive stimuli: probable involvement of a supraspinal loop, Brain 110
494(1987) 1497–1508.
495[73] T. De Broucker, P. Cesaro, J.C. Willer, D. Le Bars, Diffuse noxious inhibitory controls (DNIC) in man:
496involvement of the spino-reticular tract, Brain 113 (1990) 1223–1234.
497[74] D. Bouhassira, D. LeBars, F. Bolgert, D. Laplane, J.C. Willer, Diffuse noxious inhibitory controls (DNIC)
498in man: a neurophysiological investigation of a patient with a form of Brown–Sequard, Ann. Neurol. 34
499(1993) 536–543.
D. Le Bars, J.-C. Willer / International Congress Series 00 (2002) 000–000 17

UNCORRECTED PROOF
ARTICLE IN PRESS
500[75] D. Le Bars, D. Chitour, E. Kraus, A.H. Dickenson, J.M. Besson, Effect of naloxone upon diffuse noxious
501inhibitory controls (DNIC) in the rat, Brain Res. 204 (1981) 387–402.
502[76] J.C. Willer, T. De Broucker, D. Le Bars, Diffuse noxious inhibitory controls (DNIC) in man: involvement
503of an opioidergic link, Eur. J. Pharmacol. 182 (1990) 347–355.
504[77] R. Melzack, P.D. Wall, Pain mechanisms: a new theory, Science 150 (1965) 971–979.
505[78] R. Satran, M.N. Goldstein, Pain perception: modification and threshold of intolerance and cortical poten-
506tials by cutaneous stimulation, Science 180 (1973) 1201–1202.
507[79] P.W. Nathan, P. Rudge, Testing the gate-control theory of pain in man, J. Neurol., Neurosurg. Psychiatry 37
508(1974) 1366–1372.
509[80] M.E. Jeans, Relief of chronic pain by brief, intense transcutaneous electrical stimulation—a double-blind
510study, in: J.J. Bonica, J.C. Liebeskind, D. Albe-Fessard (Eds.), Advances in Pain Research and Therapy,
511vol. 3, Raven Press, New York, 1979, pp. 601–606.
512[81] V.V. Romita, A. Suk, J.L. Henry, Parametric studies on electroacupuncture-like stimulation in a rat model:
513effects of intensity, frequency, and duration of stimulation on evoked antinociception, Brain Res. Bull. 42
514(1997) 289–296.
515[82] R. Melzack, Prolonged relief of pain by brief, intense transcutaneous somatic stimulation, Pain 1 (1975)
516357–373.
517[83] S.A. Andersson, Pain control by sensory stimulation, in: J. Bonica, J.C. Liebeskind, Albe-Fessard (Eds.),
518Advances in Pain Research and Therapy, vol. 3, Raven Press, New York, 1979, pp. 569–585.
519[84] J.M. Chung, Z.R. Fang, Y. Hori, K.H. Lee, W.D. Willis, Prolonged inhibition of primate spinothalamic
520tract cells by peripheral nerve stimulation, Pain 19 (1984) 259–275.
521[85] K. Kawakita, M. Funakoshi, Suppression of the jawopening reflex by conditioning A-delta fibre stimula-
522tion and electroacupuncture in the rat, Exp. Neurol. 78 (1982) 461–465.
523[86] H.K. Lee, J.M. Chung, W.D. Willis, Inhibition of primate spinothalamic tract cells by TENS, J. Neurosurg.
5242 (1985) 276–287.
525[87] B.H. Sjolund, Peripheral nerve suppression of C-fibre evoked flexion reflex in rats: Part 1. Parameters of
526continuous stimulation, J. Neurosurg. 63 (1985) 612–616.
527[88] C.J. Woolf, D. Mitchell, G.D. Barrett, Antinociceptive effect of peripheral segmental electrical stimulation
528in the rat, Pain 8 (1980) 237–252.
529[89] P. Baldry, Acupuncture, Trigger Points, and Musculoskeletal Pain: A Scientific Approach to Acupuncture
530for Use by Doctors and Physiotherapists in the Diagnosis and Management of Myofascial Trigger Point
531Pain, 2nd edn., Churchill Livingstone, Edinburgh, 1993.
532[90] F. Mann, Acupuncture analgesia, report of 100 experiments, Br. J. Anaesth. 46 (1974) 361–364.
533[91] A.J.R. MacDonald, Acupuncture’s non-segmental and segmental analgesic effects: the points of meridians,
534in: J. Filshie, A. White (Eds.), Medical Acupuncture: A Western Approach, Churchill Livingstone, Edin-
535burgh, 1998, pp. 83–104.
536[92] Shangai College of Traditional Medicine, Acupuncture: A Comprehensive Text, Eastland, Chicago, 1981.
537[93] L. Chen, J. Tang, J. Yan, Analysis of afferent fibres for the impulses of acupuncture analgesia, Kexue
538Tongbao 26 (1981) 564–569.
539[94] K. Kawakita, K. Gotoh, Role of polymodal receptors in the acupuncture-mediated endogenous pain
540inhibitory systems, Prog. Brain Res. 113 (1996) 507–523.
541[95] K. Okada, M. Oshima, K. Kawakita, Examination of the afferent fiber responsible for the suppression of
542jaw-opening reflex in heat, cold, and manual acupuncture stimulation in rats, Brain Res. 740 (1996) 201–
543207.
544[96] H.T. Chang, Neurophysiological interpretation of acupuncture analgesia, Endeavour 4 (1980) 92–96.
545[97] L.F. He, Involvement of endogenous opioid peptides in acupuncture analgesia, Pain 31 (1987) 99–121.
546[98] D.J. Mayer, Biological mechanisms of acupuncture, Prog. Brain Res. 122 (2000) 457–477.
547[99] B. Pomeranz, D. Chiu, Naloxone blockade of acupuncture analgesia: endorphin implicated, Life Sci. 19
548(1976) 1757–1762.
549[100] D.J. Mayer, D.D. Price, A. Rafii, Antagonism of acupuncture analgesia in man by the narcotic antagonist
550naloxone, Brain Res. 121 (1977) 368–372.
551[101] R.S.S. Cheng, B. Pomeranz, Electroacupuncture analgesia could be mediated by at least two pain-relieving
552mechanisms: endorphin and non-endorphin systems, Life Sci. 25 (1979) 1957–1962.
D. Le Bars, J.-C. Willer / International Congress Series 00 (2002) 000–00018

UNCORRECTED PROOF
ARTICLE IN PRESS
553[102] J.S. Han, L. Terenius, Neurochemical basis of acupuncture analgesia, Annu. Rev. Pharmacol. Toxicol. 22
554(1982) 193–220.
555[103] V.V. Romita, J.L. Henry, Spinal mu-, delta-and kappa-opioid receptors mediate intense stimulation-elicited
556inhibition of a nociceptive reflex in the rat, Eur. J. Pharmacol. 357 (1998) 127–138.
557[104] Z. Bing, F. Cesselin, S. Bourgoin, A.M. Clot, M. Hamon, D. Le Bars, Acupuncture-like stimulation
558induces a heterosegmental release of Met-enkephalin-like material in the rat spinal cord, Pain 47 (1991)
55971–77.
560[105] J.S. Han, X.H. Chen, S.L. Sun, X.J. Xu, Y. Yuan, S.C. Yan, J.X. Hao, L. Terenius, Effect of low- and high-
561frequency TENS on Met-enkephalin-Arg-Phe and dynorphin A immunoreactivity in human lumbar CSF,
562Pain 47 (1991) 295–298.
563[106] P. Mason, C.G. Leung, Physiological functions of pontomedullary raphe and medial reticular neurons,
564Prog. Brain Res. 107 (1996) 269–282.
565[107] T.A. Lovick, The medullary raphe nuclei: a system for integration and gain control in autonomic and
566somatomotor responsiveness? Exp. Physiol. 82 (1997) 31–41.
567[108] S. Knardahl, M. Elam, B. Olausson, B.G. Wallin, Sympathetic nerve activity after acupuncture in humans,
568Pain 75 (1998) 19–25.
569[109] L.E. Dyrehag, E.G. Widerstrom-Noga, S.G. Carlsson, S.A. Andersson, Effects of repeated sensory stim-
570ulation sessions (electro-acupuncture) on skin temperature in chronic pain patients, Scand. J. Rehabil. Med.
57129 (1997) 243–250.
572[110] E. Haker, H. Egekvist, P. Bjerring, Effect of sensory stimulation (acupuncture) on sympathetic and para-
573sympathetic activities in healthy subjects, J. Auton. Nerv. Syst. 79 (2000) 52–59.
574[111] A. Sato, Y. Sato, R.F. Schmidt, The impact of somatosensory input on autonomic functions, Rev. Physiol.,
575Biochem. Pharmacol. 130 (1997) 1–328.
576[112] R. Melzack, D.M. Stillwell, E.J. Fox, Trigger points and acupuncture points for pain: correlations and
577implications, Pain 3 (1977) 3–23.
578[113] A.J.R. Macdonald, Acupuncture analgesia and therapy, in: P.D. Wall, R. Melzack (Eds.), Textbook of Pain,
579Churchill Livingstone, Edinburgh, 1989, pp. 906–919.
580[114] F. Mann, Reinventing Acupuncture: A New Concept of Ancient Medicine, 2nd edn., Butterworth-Heine-
581mann, Oxford, 2000.
582
D. Le Bars, J.-C. Willer / International Congress Series 00 (2002) 000–000 19
Copyright © 2022 FDOKUMEN




















