
Laurus novocanariensis essential oil: Seasonal variation and valorization
10
Laurus novocanariensis essential oil: Seasonal variation and valorization Jesu ´s M. Rodilla a , Maria Teresa Tinoco b , Julio Cruz Morais b , Cristina Gimenez c , Raimundo Cabrera c , Darı ´o Martı ´n-Benito d , Lucia Castillo d , Azucena Gonzalez-Coloma d, * a Departamento de Quı ´mica, Universidade da Beira Interior, 6201-001 Covilh~ a, Portugal b Instituto de Cie ˆncias Agr arias Mediterra ˆ nicas-ICAM e Departamento de Quı ´mica, Universidade de E ´ vora, 7000-671 E ´ vora, Portugal c UDI Fitopatologı ´a, Universidad de la Laguna, Tenerife, Spain d Centro de Ciencias Medioambientales-CSIC, Serrano 115-dpdo, 28006 Madrid, Spain Received 27 February 2007; accepted 16 September 2007 Abstract We investigated the qualitative and quantitative seasonal variation of the leaf and fruit oils of the Macaronesian endemism Laurus novocanariensis and their plant defensive potential. The monoterpene fraction dominated the leaf (74%) and berry essential oils (73e44%, ripeeunripe). The insect antifeedant effects of these oils were species- and season-dependent against the aphids (Myzus persicae and Rhopalosiphum padi). Overall, the biological effects of these oils correlated with the oxygenated terpene fraction. Among the pure components tested, b-caryophyllene and its oxide were strong antifeedants to Leptinotarsa decemlineata and Spodoptera littoralis. The aphids responded to b-ocimene, b-pinene, 1,8-cineole, linalool (antifeedants) and linalool oxide (attractive to M. persicae). The antifungal effects of the leaf oils on Fusarium spp. were season-dependent. b-Caryophyllene oxide proved to be a strong antifungal. L. novocanariensis oils inhibited Lactuca sativa germination and radicle elongation, the leaves being more effective. Linalool also inhibited seed germination. Ó 2007 Elsevier Ltd. All rights reserved. Keywords: Laurus novocanariensis; Essential oils; Season; Antifeedant; Phytotoxic 1. Introduction Laurus novocanariensis (Rivas-Martı ´nez et al., 2002), formerly classified as Laurus azorica Franco (Seub.), is an endemic species native to the laurel forest of Madeira and The Canary Islands and the dominant tree species in the forest of Madeira. The leaves and stem wood are used in traditional cookery for their aromatic properties (Neves and Valente, 1992; Rivera and Obo ´n, 1995). The oil obtained from pressed ripe berries is widely used * Corresponding author. Tel.: þ34 917452500; fax: þ34 915640800. E-mail address: [email protected] (A. Gonzalez-Coloma). 0305-1978/$ - see front matter Ó 2007 Elsevier Ltd. All rights reserved. doi:10.1016/j.bse.2007.09.001 www.elsevier.com/locate/biochemsyseco Available online at www.sciencedirect.com Biochemical Systematics and Ecology 36 (2008) 167e176
Transcript of Laurus novocanariensis essential oil: Seasonal variation and valorization

www.elsevier.com/locate/biochemsyseco
Available online at www.sciencedirect.com
Biochemical Systematics and Ecology 36 (2008) 167e176
Laurus novocanariensis essential oil: Seasonalvariation and valorization
Jesus M. Rodilla a, Maria Teresa Tinoco b, Julio Cruz Morais b,Cristina Gimenez c, Raimundo Cabrera c, Darıo Martın-Benito d,
Lucia Castillo d, Azucena Gonzalez-Coloma d,*
a Departamento de Quımica, Universidade da Beira Interior, 6201-001 Covilh~a, Portugalb Instituto de Ciencias Agr�arias Mediterranicas-ICAM e Departamento de Quımica,
Universidade de Evora, 7000-671 Evora, Portugalc UDI Fitopatologıa, Universidad de la Laguna, Tenerife, Spain
d Centro de Ciencias Medioambientales-CSIC, Serrano 115-dpdo, 28006 Madrid, Spain
Received 27 February 2007; accepted 16 September 2007
Abstract
We investigated the qualitative and quantitative seasonal variation of the leaf and fruit oils of the Macaronesian endemismLaurus novocanariensis and their plant defensive potential. The monoterpene fraction dominated the leaf (74%) and berry essentialoils (73e44%, ripeeunripe). The insect antifeedant effects of these oils were species- and season-dependent against the aphids(Myzus persicae and Rhopalosiphum padi). Overall, the biological effects of these oils correlated with the oxygenated terpenefraction. Among the pure components tested, b-caryophyllene and its oxide were strong antifeedants to Leptinotarsa decemlineataand Spodoptera littoralis. The aphids responded to b-ocimene, b-pinene, 1,8-cineole, linalool (antifeedants) and linalool oxide(attractive to M. persicae). The antifungal effects of the leaf oils on Fusarium spp. were season-dependent. b-Caryophyllene oxideproved to be a strong antifungal. L. novocanariensis oils inhibited Lactuca sativa germination and radicle elongation, the leavesbeing more effective. Linalool also inhibited seed germination.� 2007 Elsevier Ltd. All rights reserved.
Keywords: Laurus novocanariensis; Essential oils; Season; Antifeedant; Phytotoxic
1. Introduction
Laurus novocanariensis (Rivas-Martınez et al., 2002), formerly classified as Laurus azorica Franco (Seub.), isan endemic species native to the laurel forest of Madeira and The Canary Islands and the dominant tree speciesin the forest of Madeira. The leaves and stem wood are used in traditional cookery for their aromatic properties(Neves and Valente, 1992; Rivera and Obon, 1995). The oil obtained from pressed ripe berries is widely used
* Corresponding author. Tel.: þ34 917452500; fax: þ34 915640800.
E-mail address: [email protected] (A. Gonzalez-Coloma).
0305-1978/$ - see front matter � 2007 Elsevier Ltd. All rights reserved.
doi:10.1016/j.bse.2007.09.001

168 J.M. Rodilla et al. / Biochemical Systematics and Ecology 36 (2008) 167e176
by the local population for therapeutic purposes (anti-rheumatic, wound-healing, against respiratory and diges-tive problems and as a homeostatic remedy) and its chemical composition has been reported (Castilho et al.,2005).
The chemical composition of L. azorica essential oil from The Azores (leaves and unripe berries) (Hokwerda et al.,1982; Pedro et al., 2001) and Madeira Islands (leaves) (Pino et al., 2004) has been reported. The composition ofL. novocanariensis spring leaf oil from a group of trees growing in the same location and collected in Madeira hasalso been reported along with its acaricidal effects against the rabbit ear mite Psoroptes cuniculi (Macchioni et al.,2006).
As part of our ongoing valorization of Macaronesian endemic species based on their plant protection properties, wehave studied the chemical composition and seasonal variations of the essential oils of L. novocanariensis leaves andberries. We have also evaluated the plant defensive properties of the oils, their main components (a-, b-pinene,b-ocimene, 1,8-cineole) and selected minor ones (linalool, b-caryophyllene and their oxides) on the herbivorousinsect species Spodoptera littoralis, Leptinotarsa decemlineata, Rhopalosiphum padi and Myzus persicae alongwith their antifungal action on Fusarium spp. and their phytotoxicity to Lactuca sativa.
2. Materials and methods
2.1. Chemicals
The pure compounds used for the bioassays were purchased from SigmaeAldrich. Linalool and linalool oxidewere kindly provided by Prof. J. Sanz (IQO-CSIC). Pure commercial essential oil components used as standardsfor GC-FID analyses were obtained from Fluka and Extrasynthese.
2.2. Plant material
Plant material was collected during two seasons from a group of trees located at an altitude of 850 m in the laurelforest of Madeira Island. Leaves were collected during the flowering (spring) and vegetative (fall) periods and theberries at unripe (July) and ripe stages (September). The leaves and berries were dried in the shade for 3 days beforehydrodistillation.
2.3. Extraction of essential oil
Dried leaves (500 g) and berries (100 g) were hand-cut into small fragments and subjected to hydrodistillation ina Clevenger-type apparatus for 2 h. The isolated oils were dried over anhydrous sodium sulphate and stored at 4 �C inthe dark prior to analysis.
2.4. Essential oil analysis
Analyses of essential oils were performed on a HewlettePackard 5890 II gas chromatograph (GC) equippedwith two different polarity-fused silica capillary columns: Supelcowax 10 (polyethylene glycol) and SPB-1 (poly-dimethylsiloxane), both 30 m long� 0.25 mm ID� 0.25 mm film thickness. Temperature program: initial temper-ature, 70 �C; hold 3 min; temperature rate 3 �C min�1, final temperature 220 �C; hold 10 min; column flow rate0.6 ml He/min; injector and detector temperatures 195 �C and 210 �C, respectively. Injection volume was 0.2 mLand split ratio 50:1. GCeMS analyses were performed on a combined GCeMS (HewlettePackard 6890,HewlettePackard 5973) instrument using an HP-1 fused silica capillary column coated with polydimethylsilox-ane (30 m long� 0.25 mm ID� 0.25 mm film thickness), connected to a quadrupole detector operating in elec-tron impact (EI) mode at 70 eV.
Component identification was obtained by comparison of their relative retention indices (to the C8eC23 n-alkaneseries) and their MS spectra with reference compounds, previously reported data (Adams, 1995) and mass spectrafrom Wiley 6 library. The quantitative composition of the oils (percent) was determined using the normalizationmethod from the GC peak areas of the two columns, without correction factors. Percent data shown are the meanvalues of three injections of each oil sample.

169J.M. Rodilla et al. / Biochemical Systematics and Ecology 36 (2008) 167e176
2.5. Insect bioassays
S. littoralis, L. decemlineata, M. persicae and R. padi colonies were reared on artificial diet, potato foliage (Solanumtuberosum), bell pepper (Capsicum annuum) and barley (Hordeum vulgare) plants, respectively, and maintained at22� 1 �C, >70% relative humidity with a photoperiod of 16:8 h (L:D) in a growth chamber.
2.5.1. Choice feeding assayThese experiments were conducted with L. decemlineata, M. persicae and R. padi (apterous) adults and sixth-instar
S. littoralis larvae. Feeding or settling inhibition (%FI or %SI) was calculated as described in Reina et al. (2001). Theactive compounds were tested in a doseeresponse experiment to calculate their relative potency (EC50 values, theeffective dose to give 50% inhibition), which was determined from linear regression analysis (% inhibition on logdose, SPSS.11).
2.6. Phytotoxicity tests
These experiments were conducted with L. sativa var. Carrascoy seeds as described in Moiteiro et al. (2006).Germination was monitored daily for 6 days and radicle length measured at the end of the experiment (20 digitalizedradicles randomly selected for each experiment) using the application Image J Version 1.37r, 2006 (http://rsb.info.nih.gov./ij/). An analysis of variance (ANOVA) was performed on germination and radicle length data.
2.7. Antifungal assays
Fusarium moniliforme (Sheldon) [CECT2152], Fusarium oxysporum fs. lycopersici (Scheldt) [CECT2715] andFusarium solani (Mart) [CECT2199] were provided by the Coleccion Espanola de Cultivos Tipo (CET). Antifungalactivity was analyzed as mycelial growth inhibition by a modified agar-dilution method (Reina et al., 1997), whichincludes the addition of 0.05 mg/ml of methyltetrazolium salts (MTT). Colonies grown on Petri dishes incubatedfor 48 h were digitalized and measured (SIGMASCAN). Percent inhibition (%I) was calculated as: %I¼ (C�T/C)� 100, where C is the diameter of the control colonies and T is the diameter of the test colonies. The active com-pounds were tested in a doseeresponse experiment to calculate their relative potency (EC50 values, the effective doseto give 50% inhibition), which was determined from linear regression analysis (% inhibition on log dose, SPSS.11).
3. Results and discussion
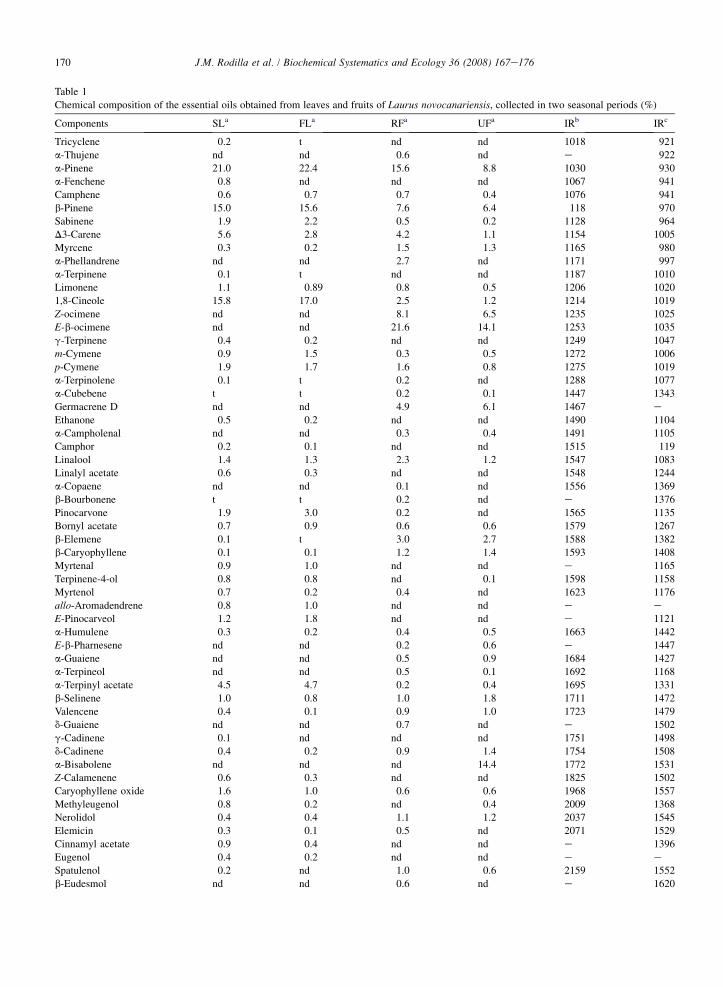
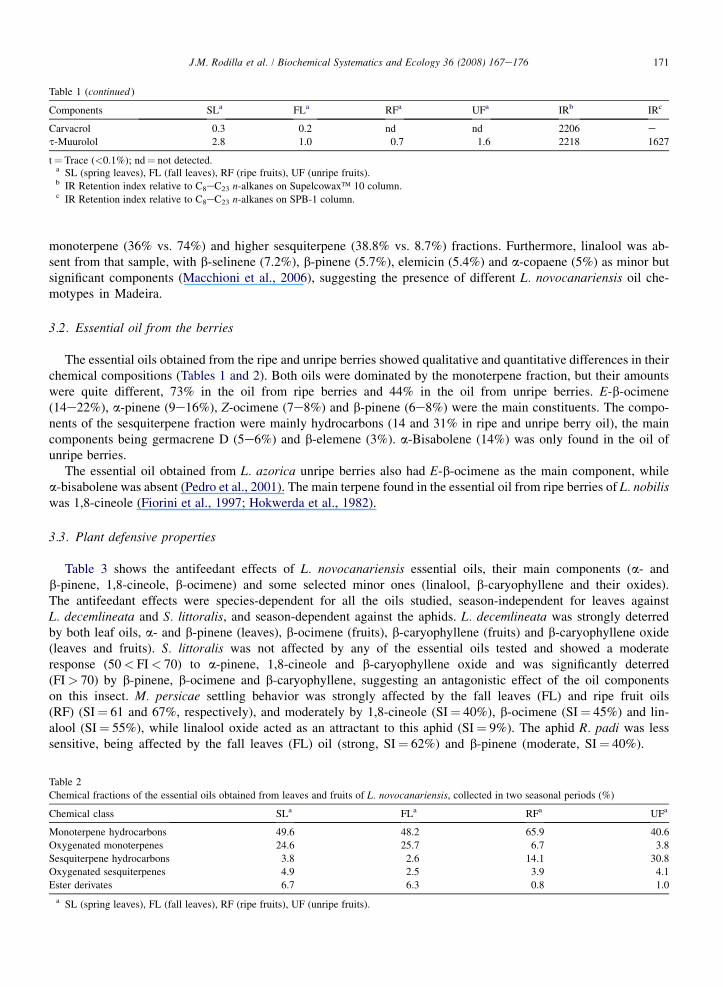
All samples afforded pale yellow essential oils in yields ranging between 0.3 and 0.4% (v/w) from the leavescollected in the spring and fall, respectively. The unripe and ripe berries yielded 0.2 and 0.8% (v/w) essential oil,respectively. A total of 37e44 and 34e41components were identified in the volatile oil of the springefall leavesand ripeeunripe berries, accounting for 85e90% and 80e91% of the volatile oil, respectively. The general chemicalprofile of the essential oils, their quantitative composition and the retention indices of the constituents are summarizedin Tables 1 and 2.
3.1. Essential oil from leaves
The chemical composition of the essential oils from leaves was nearly identical in the two seasons investigated,with the exception of the minor constituents a-fenchene, g-cadinene and spatulenol, which were only detected inthe oil from the spring leaves (Table 1). The monoterpene fraction was the most abundant (74%) with the oxygen-containing monoterpenes accounting for 25e26% (Table 2), the main components being a-pinene (21e22%),1,8-cineole (16e17%), D3-carene (3e6%) and a-terpinyl acetate (4e5%). The sesquiterpene fraction was lessabundant (5e9%) with t-muurolol (1e3%) being its major component.
A previous report showed a-pinene and 1,8-cineole as the main oil components of L. azorica leaves, the mono-terpene fraction dominating the oil (Pedro et al., 2001). The main terpene found in Laurus nobilis leaf oil was also1,8-cineole (Fiorini et al., 1997; Hokwerda et al., 1982; Macchioni et al., 2006). Our results showed qualitative andquantitative differences in leaf oil composition from that reported for L. novocanariensis, which showed lower
170 J.M. Rodilla et al. / Biochemical Systematics and Ecology 36 (2008) 167e176
Table 1
Chemical composition of the essential oils obtained from leaves and fruits of Laurus novocanariensis, collected in two seasonal periods (%)
Components SLa FLa RFa UFa IRb IRc
Tricyclene 0.2 t nd nd 1018 921
a-Thujene nd nd 0.6 nd e 922
a-Pinene 21.0 22.4 15.6 8.8 1030 930
a-Fenchene 0.8 nd nd nd 1067 941
Camphene 0.6 0.7 0.7 0.4 1076 941
b-Pinene 15.0 15.6 7.6 6.4 118 970
Sabinene 1.9 2.2 0.5 0.2 1128 964
D3-Carene 5.6 2.8 4.2 1.1 1154 1005
Myrcene 0.3 0.2 1.5 1.3 1165 980
a-Phellandrene nd nd 2.7 nd 1171 997
a-Terpinene 0.1 t nd nd 1187 1010
Limonene 1.1 0.89 0.8 0.5 1206 1020
1,8-Cineole 15.8 17.0 2.5 1.2 1214 1019
Z-ocimene nd nd 8.1 6.5 1235 1025
E-b-ocimene nd nd 21.6 14.1 1253 1035
g-Terpinene 0.4 0.2 nd nd 1249 1047
m-Cymene 0.9 1.5 0.3 0.5 1272 1006
p-Cymene 1.9 1.7 1.6 0.8 1275 1019
a-Terpinolene 0.1 t 0.2 nd 1288 1077
a-Cubebene t t 0.2 0.1 1447 1343
Germacrene D nd nd 4.9 6.1 1467 e
Ethanone 0.5 0.2 nd nd 1490 1104
a-Campholenal nd nd 0.3 0.4 1491 1105
Camphor 0.2 0.1 nd nd 1515 119
Linalool 1.4 1.3 2.3 1.2 1547 1083
Linalyl acetate 0.6 0.3 nd nd 1548 1244
a-Copaene nd nd 0.1 nd 1556 1369
b-Bourbonene t t 0.2 nd e 1376
Pinocarvone 1.9 3.0 0.2 nd 1565 1135
Bornyl acetate 0.7 0.9 0.6 0.6 1579 1267
b-Elemene 0.1 t 3.0 2.7 1588 1382
b-Caryophyllene 0.1 0.1 1.2 1.4 1593 1408
Myrtenal 0.9 1.0 nd nd e 1165
Terpinene-4-ol 0.8 0.8 nd 0.1 1598 1158
Myrtenol 0.7 0.2 0.4 nd 1623 1176
allo-Aromadendrene 0.8 1.0 nd nd e eE-Pinocarveol 1.2 1.8 nd nd e 1121
a-Humulene 0.3 0.2 0.4 0.5 1663 1442
E-b-Pharnesene nd nd 0.2 0.6 e 1447
a-Guaiene nd nd 0.5 0.9 1684 1427
a-Terpineol nd nd 0.5 0.1 1692 1168
a-Terpinyl acetate 4.5 4.7 0.2 0.4 1695 1331
b-Selinene 1.0 0.8 1.0 1.8 1711 1472
Valencene 0.4 0.1 0.9 1.0 1723 1479
d-Guaiene nd nd 0.7 nd e 1502
g-Cadinene 0.1 nd nd nd 1751 1498
d-Cadinene 0.4 0.2 0.9 1.4 1754 1508
a-Bisabolene nd nd nd 14.4 1772 1531
Z-Calamenene 0.6 0.3 nd nd 1825 1502
Caryophyllene oxide 1.6 1.0 0.6 0.6 1968 1557
Methyleugenol 0.8 0.2 nd 0.4 2009 1368
Nerolidol 0.4 0.4 1.1 1.2 2037 1545
Elemicin 0.3 0.1 0.5 nd 2071 1529
Cinnamyl acetate 0.9 0.4 nd nd e 1396
Eugenol 0.4 0.2 nd nd e eSpatulenol 0.2 nd 1.0 0.6 2159 1552
b-Eudesmol nd nd 0.6 nd e 1620
171J.M. Rodilla et al. / Biochemical Systematics and Ecology 36 (2008) 167e176
monoterpene (36% vs. 74%) and higher sesquiterpene (38.8% vs. 8.7%) fractions. Furthermore, linalool was ab-sent from that sample, with b-selinene (7.2%), b-pinene (5.7%), elemicin (5.4%) and a-copaene (5%) as minor butsignificant components (Macchioni et al., 2006), suggesting the presence of different L. novocanariensis oil che-motypes in Madeira.
3.2. Essential oil from the berries
The essential oils obtained from the ripe and unripe berries showed qualitative and quantitative differences in theirchemical compositions (Tables 1 and 2). Both oils were dominated by the monoterpene fraction, but their amountswere quite different, 73% in the oil from ripe berries and 44% in the oil from unripe berries. E-b-ocimene(14e22%), a-pinene (9e16%), Z-ocimene (7e8%) and b-pinene (6e8%) were the main constituents. The compo-nents of the sesquiterpene fraction were mainly hydrocarbons (14 and 31% in ripe and unripe berry oil), the maincomponents being germacrene D (5e6%) and b-elemene (3%). a-Bisabolene (14%) was only found in the oil ofunripe berries.
The essential oil obtained from L. azorica unripe berries also had E-b-ocimene as the main component, whilea-bisabolene was absent (Pedro et al., 2001). The main terpene found in the essential oil from ripe berries of L. nobiliswas 1,8-cineole (Fiorini et al., 1997; Hokwerda et al., 1982).
3.3. Plant defensive properties
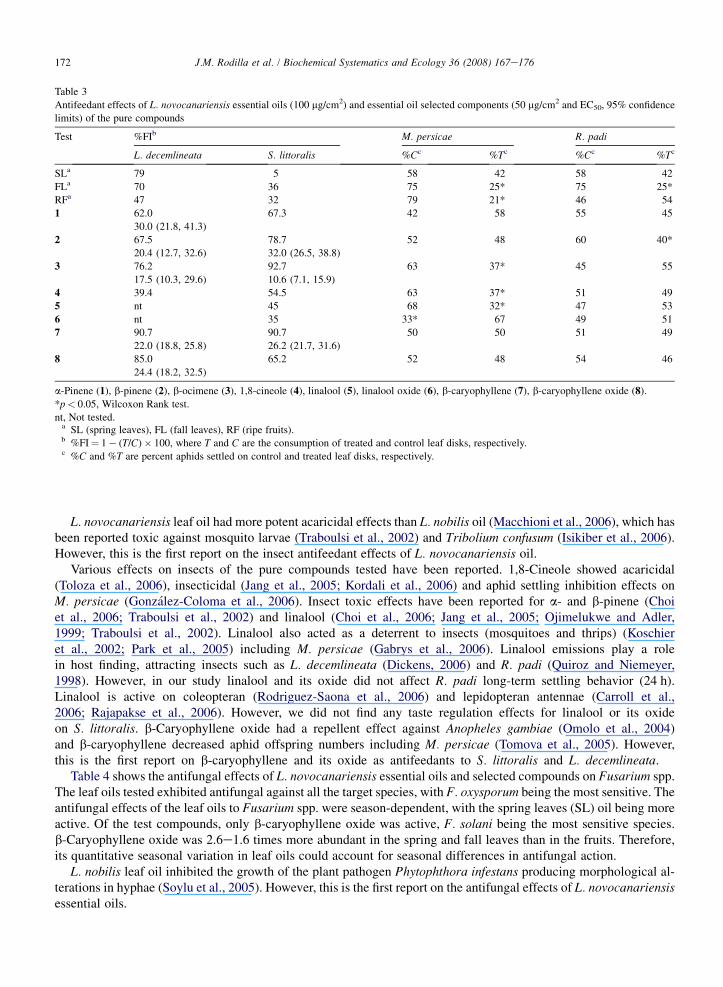
Table 3 shows the antifeedant effects of L. novocanariensis essential oils, their main components (a- andb-pinene, 1,8-cineole, b-ocimene) and some selected minor ones (linalool, b-caryophyllene and their oxides).The antifeedant effects were species-dependent for all the oils studied, season-independent for leaves againstL. decemlineata and S. littoralis, and season-dependent against the aphids. L. decemlineata was strongly deterredby both leaf oils, a- and b-pinene (leaves), b-ocimene (fruits), b-caryophyllene (fruits) and b-caryophyllene oxide(leaves and fruits). S. littoralis was not affected by any of the essential oils tested and showed a moderateresponse (50< FI< 70) to a-pinene, 1,8-cineole and b-caryophyllene oxide and was significantly deterred(FI> 70) by b-pinene, b-ocimene and b-caryophyllene, suggesting an antagonistic effect of the oil componentson this insect. M. persicae settling behavior was strongly affected by the fall leaves (FL) and ripe fruit oils(RF) (SI¼ 61 and 67%, respectively), and moderately by 1,8-cineole (SI¼ 40%), b-ocimene (SI¼ 45%) and lin-alool (SI¼ 55%), while linalool oxide acted as an attractant to this aphid (SI¼ 9%). The aphid R. padi was lesssensitive, being affected by the fall leaves (FL) oil (strong, SI¼ 62%) and b-pinene (moderate, SI¼ 40%).
Table 1 (continued )
Components SLa FLa RFa UFa IRb IRc
Carvacrol 0.3 0.2 nd nd 2206 et-Muurolol 2.8 1.0 0.7 1.6 2218 1627
t¼ Trace (<0.1%); nd¼ not detected.a SL (spring leaves), FL (fall leaves), RF (ripe fruits), UF (unripe fruits).b IR Retention index relative to C8eC23 n-alkanes on Supelcowax� 10 column.c IR Retention index relative to C8eC23 n-alkanes on SPB-1 column.
Table 2
Chemical fractions of the essential oils obtained from leaves and fruits of L. novocanariensis, collected in two seasonal periods (%)
Chemical class SLa FLa RFa UFa
Monoterpene hydrocarbons 49.6 48.2 65.9 40.6
Oxygenated monoterpenes 24.6 25.7 6.7 3.8
Sesquiterpene hydrocarbons 3.8 2.6 14.1 30.8
Oxygenated sesquiterpenes 4.9 2.5 3.9 4.1
Ester derivates 6.7 6.3 0.8 1.0
a SL (spring leaves), FL (fall leaves), RF (ripe fruits), UF (unripe fruits).
172 J.M. Rodilla et al. / Biochemical Systematics and Ecology 36 (2008) 167e176
L. novocanariensis leaf oil had more potent acaricidal effects than L. nobilis oil (Macchioni et al., 2006), which hasbeen reported toxic against mosquito larvae (Traboulsi et al., 2002) and Tribolium confusum (Isikiber et al., 2006).However, this is the first report on the insect antifeedant effects of L. novocanariensis oil.
Various effects on insects of the pure compounds tested have been reported. 1,8-Cineole showed acaricidal(Toloza et al., 2006), insecticidal (Jang et al., 2005; Kordali et al., 2006) and aphid settling inhibition effects onM. persicae (Gonz�alez-Coloma et al., 2006). Insect toxic effects have been reported for a- and b-pinene (Choiet al., 2006; Traboulsi et al., 2002) and linalool (Choi et al., 2006; Jang et al., 2005; Ojimelukwe and Adler,1999; Traboulsi et al., 2002). Linalool also acted as a deterrent to insects (mosquitoes and thrips) (Koschieret al., 2002; Park et al., 2005) including M. persicae (Gabrys et al., 2006). Linalool emissions play a rolein host finding, attracting insects such as L. decemlineata (Dickens, 2006) and R. padi (Quiroz and Niemeyer,1998). However, in our study linalool and its oxide did not affect R. padi long-term settling behavior (24 h).Linalool is active on coleopteran (Rodriguez-Saona et al., 2006) and lepidopteran antennae (Carroll et al.,2006; Rajapakse et al., 2006). However, we did not find any taste regulation effects for linalool or its oxideon S. littoralis. b-Caryophyllene oxide had a repellent effect against Anopheles gambiae (Omolo et al., 2004)and b-caryophyllene decreased aphid offspring numbers including M. persicae (Tomova et al., 2005). However,this is the first report on b-caryophyllene and its oxide as antifeedants to S. littoralis and L. decemlineata.
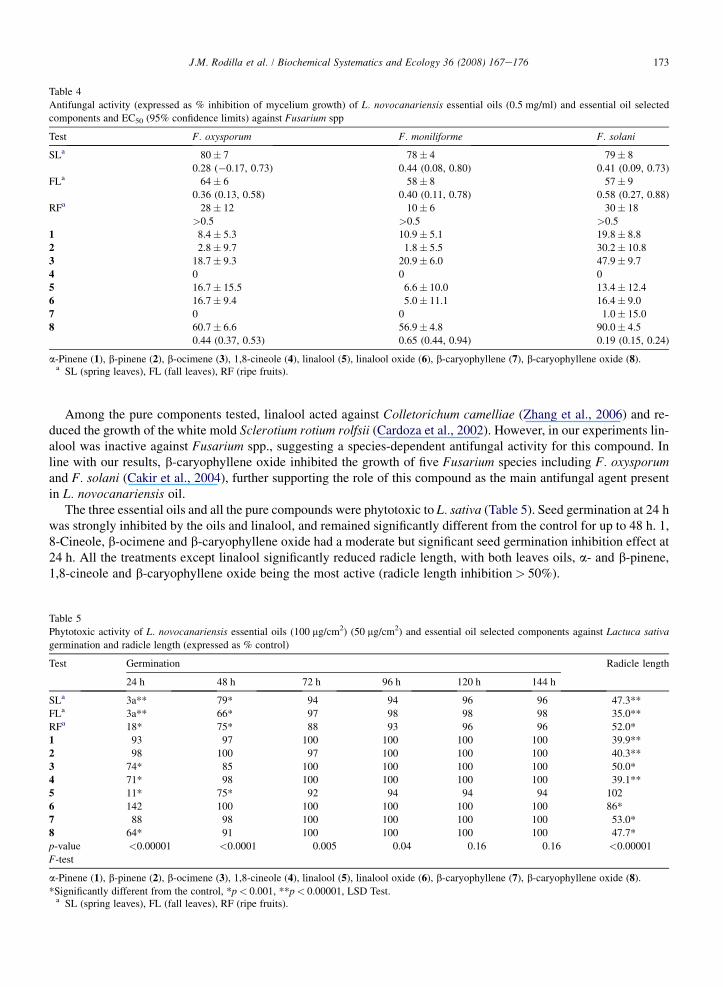
Table 4 shows the antifungal effects of L. novocanariensis essential oils and selected compounds on Fusarium spp.The leaf oils tested exhibited antifungal against all the target species, with F. oxysporum being the most sensitive. Theantifungal effects of the leaf oils to Fusarium spp. were season-dependent, with the spring leaves (SL) oil being moreactive. Of the test compounds, only b-caryophyllene oxide was active, F. solani being the most sensitive species.b-Caryophyllene oxide was 2.6e1.6 times more abundant in the spring and fall leaves than in the fruits. Therefore,its quantitative seasonal variation in leaf oils could account for seasonal differences in antifungal action.
L. nobilis leaf oil inhibited the growth of the plant pathogen Phytophthora infestans producing morphological al-terations in hyphae (Soylu et al., 2005). However, this is the first report on the antifungal effects of L. novocanariensisessential oils.
Table 3
Antifeedant effects of L. novocanariensis essential oils (100 mg/cm2) and essential oil selected components (50 mg/cm2 and EC50, 95% confidence
limits) of the pure compounds
Test %FIb M. persicae R. padi
L. decemlineata S. littoralis %Cc %Tc %Cc %Tc
SLa 79 5 58 42 58 42
FLa 70 36 75 25* 75 25*
RFa 47 32 79 21* 46 54
1 62.0 67.3 42 58 55 45
30.0 (21.8, 41.3)
2 67.5 78.7 52 48 60 40*
20.4 (12.7, 32.6) 32.0 (26.5, 38.8)
3 76.2 92.7 63 37* 45 55
17.5 (10.3, 29.6) 10.6 (7.1, 15.9)
4 39.4 54.5 63 37* 51 49
5 nt 45 68 32* 47 53
6 nt 35 33* 67 49 51
7 90.7 90.7 50 50 51 49
22.0 (18.8, 25.8) 26.2 (21.7, 31.6)
8 85.0 65.2 52 48 54 46
24.4 (18.2, 32.5)
a-Pinene (1), b-pinene (2), b-ocimene (3), 1,8-cineole (4), linalool (5), linalool oxide (6), b-caryophyllene (7), b-caryophyllene oxide (8).
*p< 0.05, Wilcoxon Rank test.
nt, Not tested.a SL (spring leaves), FL (fall leaves), RF (ripe fruits).b %FI¼ 1� (T/C)� 100, where T and C are the consumption of treated and control leaf disks, respectively.c %C and %T are percent aphids settled on control and treated leaf disks, respectively.
173J.M. Rodilla et al. / Biochemical Systematics and Ecology 36 (2008) 167e176
Among the pure components tested, linalool acted against Colletorichum camelliae (Zhang et al., 2006) and re-duced the growth of the white mold Sclerotium rotium rolfsii (Cardoza et al., 2002). However, in our experiments lin-alool was inactive against Fusarium spp., suggesting a species-dependent antifungal activity for this compound. Inline with our results, b-caryophyllene oxide inhibited the growth of five Fusarium species including F. oxysporumand F. solani (Cakir et al., 2004), further supporting the role of this compound as the main antifungal agent presentin L. novocanariensis oil.
The three essential oils and all the pure compounds were phytotoxic to L. sativa (Table 5). Seed germination at 24 hwas strongly inhibited by the oils and linalool, and remained significantly different from the control for up to 48 h. 1,8-Cineole, b-ocimene and b-caryophyllene oxide had a moderate but significant seed germination inhibition effect at24 h. All the treatments except linalool significantly reduced radicle length, with both leaves oils, a- and b-pinene,1,8-cineole and b-caryophyllene oxide being the most active (radicle length inhibition> 50%).
Table 4
Antifungal activity (expressed as % inhibition of mycelium growth) of L. novocanariensis essential oils (0.5 mg/ml) and essential oil selected
components and EC50 (95% confidence limits) against Fusarium spp
Test F. oxysporum F. moniliforme F. solani
SLa 80� 7 78� 4 79� 8
0.28 (�0.17, 0.73) 0.44 (0.08, 0.80) 0.41 (0.09, 0.73)
FLa 64� 6 58� 8 57� 9
0.36 (0.13, 0.58) 0.40 (0.11, 0.78) 0.58 (0.27, 0.88)
RFa 28� 12 10� 6 30� 18
>0.5 >0.5 >0.5
1 8.4� 5.3 10.9� 5.1 19.8� 8.8
2 2.8� 9.7 1.8� 5.5 30.2� 10.8
3 18.7� 9.3 20.9� 6.0 47.9� 9.7
4 0 0 0
5 16.7� 15.5 6.6� 10.0 13.4� 12.4
6 16.7� 9.4 5.0� 11.1 16.4� 9.0
7 0 0 1.0� 15.0
8 60.7� 6.6 56.9� 4.8 90.0� 4.5
0.44 (0.37, 0.53) 0.65 (0.44, 0.94) 0.19 (0.15, 0.24)
a-Pinene (1), b-pinene (2), b-ocimene (3), 1,8-cineole (4), linalool (5), linalool oxide (6), b-caryophyllene (7), b-caryophyllene oxide (8).a SL (spring leaves), FL (fall leaves), RF (ripe fruits).
Table 5
Phytotoxic activity of L. novocanariensis essential oils (100 mg/cm2) (50 mg/cm2) and essential oil selected components against Lactuca sativagermination and radicle length (expressed as % control)
Test Germination Radicle length
24 h 48 h 72 h 96 h 120 h 144 h
SLa 3a** 79* 94 94 96 96 47.3**
FLa 3a** 66* 97 98 98 98 35.0**
RFa 18* 75* 88 93 96 96 52.0*
1 93 97 100 100 100 100 39.9**
2 98 100 97 100 100 100 40.3**
3 74* 85 100 100 100 100 50.0*
4 71* 98 100 100 100 100 39.1**
5 11* 75* 92 94 94 94 102
6 142 100 100 100 100 100 86*
7 88 98 100 100 100 100 53.0*
8 64* 91 100 100 100 100 47.7*
p-value <0.00001 <0.0001 0.005 0.04 0.16 0.16 <0.00001
F-test
a-Pinene (1), b-pinene (2), b-ocimene (3), 1,8-cineole (4), linalool (5), linalool oxide (6), b-caryophyllene (7), b-caryophyllene oxide (8).
*Significantly different from the control, *p< 0.001, **p< 0.00001, LSD Test.a SL (spring leaves), FL (fall leaves), RF (ripe fruits).

174 J.M. Rodilla et al. / Biochemical Systematics and Ecology 36 (2008) 167e176
Essential oils from several plant species (Origanum syriacum, Micromeria fruticosa and Cymbopogon citratus) hadallelopathic effects against several plant species, inhibiting their germination and radicle length (Dudai et al., 1999).However, this is the first report on the phytotoxic effects of L. novocanariensis oil and the pure terpenes tested.
Overall, the effects of the oils on L. decemlineata, R. padi, Fusarium spp. and L. sativa correlated with theoxygenated terpene fraction, abundant in both leaf oils (50e48%) (Table 6), with 1,8-cineole being the main compo-nent of these fractions (16e17%), followed by pinocarvone (2e3%), linalool (1.4e1.3%), a-terpinyl acetate (1e5%),t-muurolol (2.8e1%) and b-caryophyllene oxide (1.6e1%). Furthermore, the oxidation of b-caryophyllene and lin-alool had dramatic effects on their biological effects, with linalool oxide increasing M. persicae settling and b-caryo-phyllene oxide acting as a strong antifungal agent. Recent studies have shown that Tagetes minuta essential oilfractions with a predominant emission of oxygenated monoterpenoids and sesquiterpenes had greater effects on de-creasing aphid populations than other oil fractions (Tomova et al., 2005). Similarly, essential oils with higher levels ofoxygenated monoterpenoids were more effective against Acanthoscelides obtectus (Papachristos et al., 2004).
However, when tested alone, the oxygenated terpenes did not explain some of the observed effects (the lack ofantifeedant effects on S. littoralis, the aphid antifeedant and the germination inhibition effects at24 h) of the leafoils, suggesting synergistic (positive and/or negative) interactions within the components of that fraction andbetween other fractions of the active oils. Essential oil components can have synergistic biological effects. Forexample, the mixture of a- and b-pinene had a stronger toxic effect on Lycoriella mali adults than either onealone (Choi et al., 2006) and the antennal effects of linalool on the blossom weevil Anthonomus rubi interferedwith a-pinene (Bichao et al., 2005).
On the basis of these results, we would suggest that the essential oil from the leaves of L. novocanariensis playsa role in plant protection due to its antifeedant, antifungal and phytotoxic effects, especially when enriched in anoxygenated terpene fraction, and may represent an added value to this endemic species. Since little is known aboutL. novocanariensis chemotypes, plant-to-plant variability should be studied concerning oil composition and plantprotection effects.
Acknowledgements
This work was supported by grants CTQ2006-15597-C02-01/PPQ, CYTED IV-13, INTERREG III-Espanha-Portugal e LABTRANS (2003e2005) and CSIC-GRICES 2006/2007 (2005PT0049). We thank Dr. J. Sanz (IQO-CSIC) for kindly providing linalool and linalool oxide. We gratefully acknowledge S. Carlin for language revision.We also thank to the ICAM for the use of GC equipment.
References
Adams, R.P., 1995. Identification of Essential Oils Components by Gas Chromatography/Mass Spectroscopy. Allured Publishing Corporation,
Carol Stream, IL.
Bichao, H., Borg-Karlson, A.K., Wibe, A., Araujo, J., Mustaparta, H., 2005. Molecular receptive ranges of olfactory receptor neurones responding
selectively to terpenoids, aliphatic green leaf volatiles and aromatic compounds, in the strawberry blossom weevil Anthonomus rubi. Chemo-
ecology 15, 211e226.
Table 6
Pearson Product Moment Correlations (p-value) values between L. novocanariensis essential oil terpene groups and %SI (L. decemlineata,
M. persicae, R. padi, positive correlation expected), Fusarium spp. mycelial growth inhibition (EC50 values, negative correlation), and L. sativa
radicle length (% control, negative correlation)
Tha Toa
L. decemlineata �0.89 (0.28) 0.97 (0.14)
M. persicae 0.48 (0.68) �0.66 (0.53)
R. padi �0.76 (0.44) 0.60 (0.58)
F. oxysporum 0.96 (0.17) �0.99 (0.09)
F. moniliforme 0.98 (0.10) �0.99 (0.04)
F. solani 0.92 (0.24) �0.98 (0.10)
L. sativa 0.82 (0.38) �0.68 (0.52)
a Th (terpene hydrocarbons), To (oxygenated terpenes).

175J.M. Rodilla et al. / Biochemical Systematics and Ecology 36 (2008) 167e176
Cakir, A., Kordali, S., Zengin, H., Izumi, S., Hirata, T., 2004. Composition and antifungal activity of essential oils isolated from Hypericum
hyssopifolium and Hypericum heterophyllum. Flavour Fragrance J. 19, 62e68.
Carroll, M.J., Schmelz, E.A., Meagher, R.L., Teal, P.E.A., 2006. Attraction of Spodoptera frugiperda larvae to volatiles from herbivore-damaged
maize seedlings. J. Chem. Ecol. 32, 1911e1924.
Cardoza, Y.J., Alborn, H.T., Tumlinson, J.H., 2002. In vivo volatile emissions from peanut plants induced by simultaneous fungal infection and
insect damage. J. Chem. Ecol. 28, 161e174.
Castilho, P.C., Costa, M.D., Rodrigues, A., Partidario, A., 2005. Characterization of laurel fruit oil from Madeira Island, Portugal. J. Am. Oil
Chem. Soc. 82, 863e868.
Choi, W.S., Park, B.S., Lee, Y.H., Jang, D.Y., Yoon, H.Y., Lee, S.E., 2006. Fumigant toxicities of essential oils and monoterpenes against
Lycoriella mali adults. Crop Prot. 25, 398e401.
Dickens, J.C., 2006. Plant volatiles moderate response to aggregation pheromone in Colorado potato beetle. J. Appl. Entomol. 130,
26e31.
Dudai, N., Poljakoff-Mayber, A., Mayer, A.M., Putievsky, E., Lerner, H.R., 1999. Essential oils as allelochemicals and their potential as herbi-
cides. J. Chem. Ecol. 25, 1079e1089.
Fiorini, C., Fouraste, I., David, B., Bessiere, J.M., 1997. Composition of the flower, leaf and stem essential oils from Laurus nobilis L. Flavour
Fragrance J. 12, 91e93.
Gabrys, B., Szczepanik, M., Dancewicz, K., Szumny, A., Wawrzenczyk, C., 2006. Environmentally safe insect control: feeding deterrent activity
of alkyl-substituted gamma- and delta-lactones to peach potato aphid (Myzus persicae [Sulz.]) and Colorado potato beetle (Leptinotarsadecemlineata Say). Pol. J. Environ. Stud. 15, 549e556.
Gonz�alez-Coloma, A., Martın, D., Mohamed, N., Garcıa-Vallejo, M.C., Soria, A.C., 2006. Antifeedant effects and chemical composition of
essential oils from different populations of Lavandula luisieri L. Biochem. Syst. Ecol. 34, 609e616.
Hokwerda, H., Bos, R., Tattje, D.H.E., Malingre, T.H.M., 1982. Composition of essential oils of Laurus nobilis, L. nobilis Var. angustifolia and
Laurus azorica. Planta Med. 44, 116e119.
Isikiber, A.A., Alma, M.H., Kanat, M., Karci, A., 2006. Fumigant toxicity of essential oils from Laurus nobilis and Rosmarinus officinalis against
all life stages of Tribolium confusum. Phytoparasitica 34, 167e177.
Jang, Y.S., Yang, Y.C., Choi, D.S., Ahn, Y.J., 2005. Vapor phase toxicity of marjoram oil compounds and their related monoterpenoids to Blattella
germanica (Orthoptera: Blattellidae). J. Agric. Food Chem. 53, 7892e7898.
Kordali, S., Aslan, I., Calmasur, O., Cakir, A., 2006. Toxicity of essential oils isolated from three Artemisia species and some of their major
components to granary weevil, Sitophilus granarius (L.) (Coleoptera: Curculionidae). Ind. Crops Prod. 23, 162e170.
Koschier, E.H., Sedy, K.A., Novak, J., 2002. Influence of plant volatiles on feeding damage caused by the onion thrips Thrips tabaci. Crop Prot.
21, 419e425.
Macchioni, F., Perrucci, S., Cioni, P., Morelli, I., Castilho, P., Cecchi, F., 2006. Composition and acaricidal activity of Laurus novocanariensis and
Laurus nobilis essential oils against Psoroptes cuniculi. J. Essent. Oil Res. 18, 111e114.
Moiteiro, C., Marcelo Curto, M.J., Mohamed, N., Bailen, M., Martınez-Dıaz, R., Gonz�alez-Coloma, A., 2006. Bio-valorization of friedelane
triterpenes derived from the cork processing industry by-products. J. Agric. Food Chem. 54, 3566e3571.
Neves, H.C., Valente, A.V., 1992. Conheca o Parque Natural da Madeira. Parque Natural da Madeira, Funchal.
Ojimelukwe, P.C., Adler, C., 1999. Potential of zimtaldehyde, 4-allyl-anisol, linalool, terpineol and other phytochemicals for the control of the
confused flour beetle (Tribolium confusum J.D V.) (Col., Tenebrionidae). J. Pest Sci. 72, 81e86.
Omolo, M.O., Okinyo, D., Ndiege, I.O., Lwande, W., Hassanali, A., 2004. Repellency of essential oils of some Kenyan plants against Anopheles
gambiae. Phytochemistry 65, 2797e2802.
Papachristos, D.P., Karamanoli, K.I., Stamopoulos, D.C., Menkissogluspiroudi, U., 2004. The relationship between the chemical composition of
three essential oils and their insecticidal activity against Acanthoscelides obtectus (Say). Pest. Manag. Sci. 60, 514e520.
Park, B.S., Choi, W.S., Kim, J.H., Kim, K.H., Lee, S.E., 2005. Monoterpenes from thyme (Thymus vulgaris) as potential mosquito repellents.
J. Am. Mosq. Control Assoc. 21, 80e83.
Pedro, L.G., Santos, P.A.G., da Silva, J.A., Figueiredo, A.C., Barroso, J.G., Deans, S.G., Looman, A., Scheffer, J.J.C., 2001. Essential oils from
Azorean Laurus azorica. Phytochemistry 57, 245e250.
Pino, J.A., Fernandes, P., Marbot, R., Fontinha, S.S., 2004. Chemical composition of the leaf oil of Laurus azorica (Seub.) Franco var. longifolia(O. Kuntze) Kunk. from Madeira. J. Essent. Oil Res. 16, 22e23.
Quiroz, A., Niemeyer, H.M., 1998. Olfactometer-assessed responses of aphid Rhopalosiphum padi to wheat and oat volatiles. J. Chem. Ecol. 24,
113e124.
Rajapakse, C.N.K., Walter, G.H., Moore, C.J., Hull, C.D., Cribb, B.W., 2006. Host recognition by a polyphagous lepidopteran (Helicoverpaarmigera): primary host plants, host produced volatiles and neurosensory stimulation. Physiol. Entomol. 31, 270e277.
Reina, M., Gonz�alez-Coloma, A., Gutierrez, C., Cabrera, R., Henrıquez, J., Villarroel, L., 1997. Structure elucidation and biological activities of
saturated pyrrolizidine alkaloids from Heliotropium floridum. Phytochemistry 46, 845e853.
Reina, M., Gonz�alez-Coloma, A., Gutierrez, C., Cabrera, R., Rodrıguez, M.L., Fajardo, V., Villarroel, L., 2001. Defensive chemistry of Seneciomiser Hook. J. Nat. Prod. 64, 6e11.
Rivas-Martınez, S., Diaz, T.E., Fern�andez-Gonz�alez, F., Izco, J., Loidi, J., Lous~a, M.E., Penas, A., 2002. Vascular plant communities of Spain and
Portugal. Addenda to syntaxonomical checklist of 2001, Part II. Itinera Geobotanica 15, 703.
Rivera, D., Obon, C., 1995. The ethnopharmacology of Madeira and Porto Santo Islands, a review. J. Ethnopharmacol. 46, 73e93.
Rodriguez-Saona, C., Poland, T.M., Miller, J.R., Stelinski, L.L., Grant, G.G., de Groot, P., Buchan, L., MacDonald, L., 2006. Behavioral and
electrophysiological responses of the emerald ash borer, Agrilus planipennis, to induced volatiles of Manchurian ash, Fraxinus mandshurica.
Chemoecology 16, 75e86.

176 J.M. Rodilla et al. / Biochemical Systematics and Ecology 36 (2008) 167e176
Soylu, E.M., Soylu, S., Kurt, S., 2005. Antimicrobial activities of the essential oils of various plants against tomato late blight disease agent
Phytophthora infestans. Mycopathologia 161, 119e128.
Toloza, A.C., Zygaldo, J., Cueto, G.M., Biurrum, Z., Zerba, E., Picollo, M.I., 2006. Fumigant and repellent properties of essential oils and
component compounds against permethrin-resistant Pediculus humanus capitis (Anoplura: Pediculidae) from Argentina. J. Med. Entomol.
43, 889e895.
Tomova, B.S., Waterhouse, J.S., Doberski, J., 2005. The effect of fractionated Tagetes oil volatiles on aphid reproduction. Entomol. Exp. Appl.
115, 153e159.
Traboulsi, A.F., Taoubi, K., El-Haj, S., Bessiere, J.M., Rammal, S., 2002. Insecticidal properties of essential plant oils against the mosquito Culex
pipiens molestus (Diptera: Culicidae). Pest Manag. Sci. 58, 491e495.
Zhang, Z.Z., Li, Y.B., Qi, L., Wan, X.C., 2006. Antifungal activities of major tea leaf volatile constituents toward Colletorichum camelliae Massea.
J. Agric. Food Chem. 54, 3936e3940.




















