
Large-scale purification and characterisation of pea globulins
10
J. Sci. Food Agric. 1984,35, 1984,35, 1024-1033 Large-scale Purification and Characterisation of Pea Globulins Jacques Gueguen, Anh T. Vu and FranGois Schaeffer Ministere de I'Agriculture, Institut National de la Recherche Agronomique, Laboratoire de Biochimie et Technologie des Prottines, Centre Agro-Alimentaire, Chemin de la Glraudiere, 44072 Nantes Cedex, France (Manuscript received 25 November 1983) Pea globulins, vicilin and legumin were isolated by chromatography on DEAE Sepharose and Ultrogel ACA 34. The procedure was carried out on a preparative scale and used to purify about 5 g of each globulin for a separation cycle. The purity of the vicilin and legumin was verified by immunoelectrophoretic and ultracentrifuge analysis. Keywords: Pea; globulin; vicilin; legumin; preparative chromatography. 1. Introduction Processes for protein isolates production from pea (Pisum sativum L.) flour have been extensively studied. Potential uses of these isolates seem to be linked to their functional proper tie^.'-^ The bulk of the work which has been done in this field is relative to the whole isolates but in order to acquire sufficient knowledge of functional properties and of their variation with processing conditions, it is necessary to understand the relations between the physicochemical and the functional properties of the major individual proteins contained in these isolates. Few studies4 have been devoted to this topic in the case of the two major pea globulins, legumin and vicilin, and most of the work has been restricted to the basic aspects of their composition and ~tructure.~-~ One reason why little is known about the physicochemical and conformational basis of the functional properties of vicilin and legumin, may be that such studies are generally very material consuming. The authors have therefore attempted to devise large-scale fractionation and purification procedures for vicilin and legumin as a first step in the investigation of their functional properties. Many methods have been used to isolate pea proteins, but most of them cannot be easily scaled up for preparative procedures. The method widely used for this purpose is the sequenced precipitation of the proteins from the extract by ammonium ~ u l p h a t e . ~ ~ ~ ~ ' ~ - ' * H owever, this method can modify the conformation of the protein, producing partly insoluble protein aggregates, which are not suitable for basic studies on functional properties. To avoid this phenomenon, a chromatographic fractionation of the whole protein solution was used, obtained by direct extraction from the flour. But few chromatographic methods are convenient for preparative uses. Gel filtration is unsuitable because the load of the column has to be low to keep the resolution as good as possible. Zonal precipitation, which is commonly used for the purification of legume seeds protein," at this scale leads to the formation of a plug in the upper part of the gel bed by the protein precipitate. Ion exchange," and hydro~yapatite~-~ and imm~noaffinity~ chromatography are convenient methods for a scale-up from the analysis to the preparation level. Immunoaffinity chromatogra- phy is, however, limited by the production of immunoserum, and when a hydroxyapatite bed in a large column is used, adsorption chromatography does not run well because of the decrease in the flow rate after each separation cycle. This last method is therefore generally used only to improve the purity of a pre-purified ample.^ Preparative h.p.1.c. may also be used, but has not 1024
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Large-scale purification and characterisation of pea globulins

J . Sci. Food Agric. 1984,35, 1984,35, 1024-1033
Large-scale Purification and Characterisation of Pea Globulins
Jacques Gueguen, Anh T. Vu and FranGois Schaeffer
Ministere de I'Agriculture, Institut National de la Recherche Agronomique, Laboratoire de Biochimie et Technologie des Prottines, Centre Agro-Alimentaire, Chemin de la Glraudiere, 44072 Nantes Cedex, France
(Manuscript received 25 November 1983)
Pea globulins, vicilin and legumin were isolated by chromatography on DEAE Sepharose and Ultrogel ACA 34. The procedure was carried out on a preparative scale and used to purify about 5 g of each globulin for a separation cycle. The purity of the vicilin and legumin was verified by immunoelectrophoretic and ultracentrifuge analysis.
Keywords: Pea; globulin; vicilin; legumin; preparative chromatography.
1. Introduction
Processes for protein isolates production from pea (Pisum sativum L.) flour have been extensively studied. Potential uses of these isolates seem to be linked to their functional proper tie^.'-^ The bulk of the work which has been done in this field is relative to the whole isolates but in order to acquire sufficient knowledge of functional properties and of their variation with processing conditions, it is necessary to understand the relations between the physicochemical and the functional properties of the major individual proteins contained in these isolates.
Few studies4 have been devoted to this topic in the case of the two major pea globulins, legumin and vicilin, and most of the work has been restricted to the basic aspects of their composition and ~ t r u c t u r e . ~ - ~ One reason why little is known about the physicochemical and conformational basis of the functional properties of vicilin and legumin, may be that such studies are generally very material consuming. The authors have therefore attempted to devise large-scale fractionation and purification procedures for vicilin and legumin as a first step in the investigation of their functional properties.
Many methods have been used to isolate pea proteins, but most of them cannot be easily scaled up for preparative procedures. The method widely used for this purpose is the sequenced precipitation of the proteins from the extract by ammonium ~ u l p h a t e . ~ ~ ~ ~ ' ~ - ' * H owever, this method can modify the conformation of the protein, producing partly insoluble protein aggregates, which are not suitable for basic studies on functional properties.
To avoid this phenomenon, a chromatographic fractionation of the whole protein solution was used, obtained by direct extraction from the flour. But few chromatographic methods are convenient for preparative uses. Gel filtration is unsuitable because the load of the column has to be low to keep the resolution as good as possible. Zonal precipitation, which is commonly used for the purification of legume seeds protein," at this scale leads to the formation of a plug in the upper part of the gel bed by the protein precipitate.
Ion exchange," and hydro~yapat i te~-~ and imm~noaffinity~ chromatography are convenient methods for a scale-up from the analysis to the preparation level. Immunoaffinity chromatogra- phy is, however, limited by the production of immunoserum, and when a hydroxyapatite bed in a large column is used, adsorption chromatography does not run well because of the decrease in the flow rate after each separation cycle. This last method is therefore generally used only to improve the purity of a pre-purified ample.^ Preparative h.p.1.c. may also be used, but has not
1024

Large-scale purification of pea globulins 1025
been performed extensively on legume proteins, even on an analytical scale, and it is a very expensive technique.
For these reasons, ion exchange chromatography (medium pressure) was chosen as the most suitable method.
2. Materials and methods 2.1. Materials All chemicals were obtained from Merck and were of analytical grade. ACA 34 Ultrogel and Trisacryl GF05 were from IBF-Pharmindustrie. DEAE-Sepharose was a product of Pharmacia Fine Chemicals. These gels were chosen because of their mechanical properties, being well adapted to high flow rate and preparative chromatography.
2.2. Preparation of the crude protein extract The pea flour used was obtained by grinding the dehulled seed (cv. Amino) with an industrial roller mill and it had an average particle size of about 150pm. The crude protein extract was prepared by stirring the resulting flour with sodium phosphate (Na2HP04.2H20)-citrate buffers, at the chosen pH and ionic strength, for 2 h at room temperature. The flour:solution ratio was 1 g: 10ml.
The mixture was centrifuged at 3800xg for 30min and the supernatant introduced on the column after filtration. Extraction was carried out without mercaptoethanol because according to C a ~ e y , ~ the presence of mercaptoethanol in the solubilisation buffer has no effect on the analytical ultracentrifuge pattern of the globulins.
2.3. Gel chromatography For all the experiments on DEAE Sepharose, the columns were loaded with 0.5ml of crude extract per ml of gel and eluted at a flow rate of lOcm h-'. At the laboratory scale the column diameter was 2.6cm and the bed gel was 40cm high. The column was washed with the equilibrating buffer to eliminate the non-bound material and then eluted with a sodium chloride gradient.
At the preparative scale, about one litre of crude extract could be applied on the 45xlOcm column. Desorption of the bound proteins was obtained with several steps of increasing NaCl concentration.
The vicilin and legumin fractions collected were pooled and concentrated by ultrafiltration on a Millipore Apparatus (Pellicon PTGC MX 10000 membranes) or on a DC2 Amicon equipment (Hollow fibre HlPlO Amicon) depending on the volume to concentrate.
The purification of these globulins was performed by gel filtration on a column filled with ACA 34 Ultrogel (laboratory scale: 100X2.5 cm; preparative scale: lOOX IOcm), equilibrated with the McIlvaine buffer (phosphate-citrate pH 7, 0 . 1 6 ~ in Na2HP04.2H20). The column was loaded with 0.04 ml of sample per ml of gel (320 ml for 8 litres of gel) and the flow rate was about 4.5 cm h-'. The purified vicilin and legutnin fractions were then pooled, desalted on Trisacryl GF05 and freeze-dried.
2.4. Gel electrophoresis SDS-polyacrylamide gel electrophoresis was carried out after reduction of the protein by p- mercaptoethanol in gradient (10-20%) polyacrylamide slab gels as described by Laemmli. l3 After electrophoresis the gels were fixed by 500ml of trichloroacetic acid (12.5% wtlvol) for 30min and stained overnight by addition of 50ml of Coomassie G at a concentration of 0.25%. Destaining was achieved by washing the gel during 2 h with water : ethanol : acetic acid (4.5 : 4.5 : 1 by vol) and then with a solution of acetic acid (5%).
2.5. Density gradient ultracentrifuging Centrifuging was performed in linear isokinetic sucrose concentration gradients (5 to 20% wthol) established in sodium phosphate (Na2HPO4.2Hz0)-citrate (pH7, 0 . 1 6 ~ in phosphate) buffer.

1026 J. Gueguen et ul.
Aliquots of the protein sample dissolved in the buffer (0.3ml at lOmgml-') were layered on the top of the gradient and centrifuged for 18 h at 285000xg and 20°C, using a Beckman L5 65B preparative centrifuge equipped with a SW40 Ti rotor. After centrifuging, the tubes were emptied by injecting a 30% sucrose solution at the bottom and the protein detected in the effluent by u.v: at 280nm. Fractions (0.5ml) were collected, and those corresponding to the peaks were pooled for further analysis. Sedimentation coefficients were evaluated following the method of Martin and Ames.14 Lysozome (1.9S), Bovine serum albumin (4.4s) (Cohn's fraction V), y-globulin (7S), and catalase (11.2S), obtained from Sigma, were used as sedimentation coefficient standards.
2.6. Amino acid analysis The amino acid analysis of proteins was performed on a Kontron analyser. The purified globulins were hydrolysed in a sealed tube in 6~ HCI, under nitrogen atmosphere at 110°C for 24h. Sulphur amino acid were determined after performic acid oxydation as cysteic acid and methionine sulphone. l5
2.7. Immunochemical techniques Antisera to legumin and vicilin were raised in rabbits. Freund's adjuvant aliquots (0.5ml) were injected subcutaneously into each rabbit. After 19 days, the rabbits were immunised with four intramuscular injections of 1 ml protein solution (1 mgml-') at 10-day intervals. The legumin and vicilin injected were prepared by preparative ultracentrifugation from the purified fractions obtained by the described chromatography procedure.
The rabbits were bled 10 days after the last injection. Under these conditions the antisera were monospecific. For immunochemical analysis of the protein samples, double diffusion was carried out at room temperature for 48h in 1% agarose gel (Agarose A45-IBF) in Verona1 buffer ( 0 . 0 2 5 ~ ~ pH8.2). As described by Ouchterlony," wells were cut in the gel, the centre well filled with the immunoserum and the surrounding wells with the antigenic solutions. After diffusion, plates were washed in physiological water, dried and stained with R. Coomasie blue for 10min.
Immunoelectrophoresis was performed according to Scheidegger," in 1 % agarose gel in the Scheidegger buffer. Wells were cut in the gel and filled with antigenic proteins and the electrophoresis carried out at 6 V cm-' (I.E. 20mA plate-') for 90min. After electrophoresis, a trough was cut and filled with antisera. The diffusion time was usually 2 to 3 days. Plates were coloured as before.
3. Results and discussion
3.1. Preparative chromatography Vicilin and legumin were purified from pea flour as shown in the flow diagram (Figure 1). The whole protein extract was fractionated by ion exchange chromatography. The separated vicilin and legumin enriched fractions were pooled, concentrated by ultrafiltration and then purified by size exclusion chromatography. The resulting purified globulins fractions were desalted through a column of Trisacryl GF05 and freeze-dried.
3.1.1. lon exchange chromatography Before running on a preparative scale, the conditions for ion exchange chromatography were precisely defined at the analytical scale. The chromatography was performed on DEAE- Sepharose with citrate-phosphate buffer (pH7, 0 . 1 6 ~ ) . About 60% of the proteins loaded were eluted without retention as a large initial peak. The adsorbed proteins were eluted as three different peaks by a linear concentration gradient of NaCl (0 to 0 . 5 ~ ) .
From the electrophoresis pattern, it may be seen that vicilin was eluted both in the adsorbed and non-adsorbed fractions, whereas legumin was desorbed as two peaks by the salt gradient. The separation of various vicilin and legumin types was in good agreement with the results of

Large-scale purification of pea globulins 1027
Pea flour
I I
Extract with phosphate buffer
Centrifuge I
I Discard
Supernatant I
Filtration I
DEAE sepharose
Vicilin-rich (€3) Legurnin-rich (D) fraction fraction
I
Ultrafiltrati'on c +
Ultrogel ACA 34
\ /
1 c Purified legumin (d,) Purified vicilin (b,)
Desalting Trisacryl GF05
Freeze-drying
c 7
Figure 1. Flow diagram for the purification of vicilin and legurnin.
Thomson et al.8'19 and of Gatehouse et ~ l . , ~ who established the molecular and charge heterogeneity of these globulins.
At the preparative scale, this DEAE Sepharose chromatography was greatly simplified compared to the analytical scale. A step gradient of increasing NaCl concentration was used instead of a linear one in order to reduce the elution volume and to shorten the chromatography cycle, and also to improve the separation yield of the column.
As shown in Figure 2, adsorbed proteins were eluted by a sequential elution composed of three different citrate-phosphate buffers (pH 7) with NaCl concentrations of 0.05, 0.25 and O S M , respectively. These molarities were inferred from the linear gradient used in the analytical experiments and were chosen in order to desorb successively the vicilin and the legumin fractions. In these conditions about 60% of the protein were washed out without retention. From the electrophoresis pattern (Figure 2b) it may be seen that the leading edge of the resulting peak (fraction A) was mainly composed of albumins, whereas in the trailing part most of the vicilin (fraction B) was eluted. These results were very close, quantitatively and qualitatively, to those obtained at the analytical scale.
The fraction C (14.6% of the sample) desorbed at 0 . 0 5 ~ NaCl was eluted as a peak which presented a shoulder in its leading edge. As shown by SDS-PAGE, this shoulder was mainly composed of vicilin (PAGE-pattern no. 5: Figure 2b) whereas the last fraction (PAGE-pattern no. 6: Figure 2b) contained most of the convicilin characterised by its 71000 mol. wt ~ u b u n i t . ~
Fraction D eluted by the 0 . 2 5 ~ NaCl buffer corresponded to legumin. It represented about 23% of the protein loaded on the column. From the PAGE pattern (no. 7) the leading edge of
61

1028 J. Gueguen et al.
NaCl 0 5 M
NoCl 0 25M
NaC I 0 0 5 M
I I I I I - I 5 9 13 17 21 L
Figure 2. (a) Ion exchange preparative chromatography of a total protein extract of pea seeds on a column of DEAE Sepharose. Numbers relate to the sample used for electrophoresis. (b) SDS-polyacrylamide gel electrophoresis of the chtomatographic fractions.
the peak was shown to be contaminated by a remaining part of vicilin and convicilin not eluted by the 0 . 0 5 ~ NaCl buffer.
As in the case of DEAE-Sepharose, chromatography carried on at the laboratory scale, non-protein material was also eluted in the peak E , as shown by U.V. spectra, protein determination and PAGE.
The purity of vicilin and legumin contained in each of these fractions, A , B, C and D separated by DEAE Sepharose chromatography was then improved by gel filtration.
3.1.2. Gel filtration The fractions A, B, C and D resulting from ion exchange chromatography were concentrated by ultrafiltration and fractionated on a Pharmacia KlOOllOO column packed with Ultrogel ACA 34.
The peak A was separated by this gel filtration chromatography into seven other peaks (al to a7). According to the SDS-PAGE patterns, most of the fractions were albumins fraction. However the peak eluted at about the void volume of the columns contained vicilin and legumin fractions. The globulins were probably eluted there as aggregates because the turbidity of this
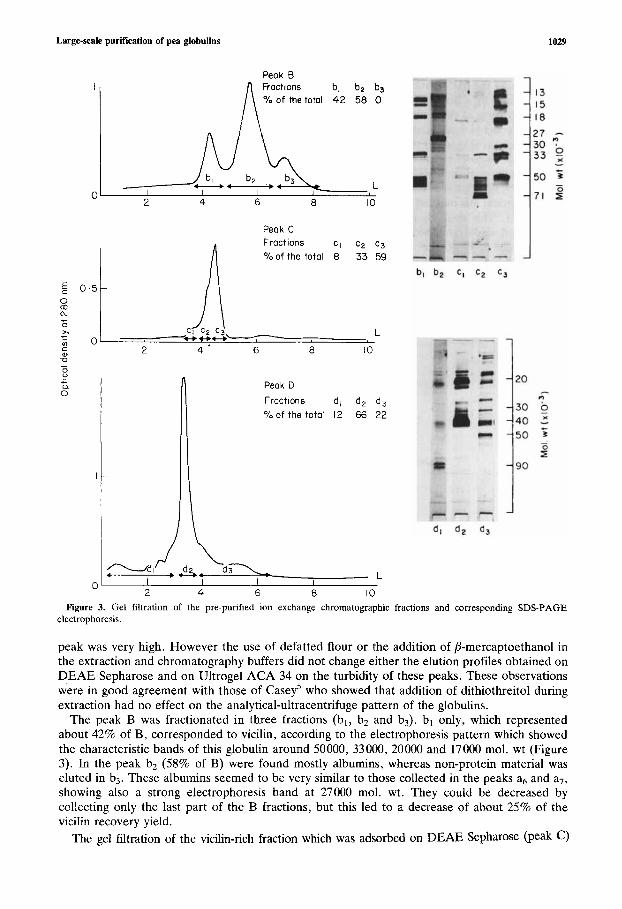
Large-scale purification of pea globulins
I
0 -
1029
bl b2 b3 of the totol 42 58 0
- L I
Peok C
%of the totol 8 33 59
c a 73
0 - u a 0
+
I
2 4 ' 6 8 10
Peok D Froctlons d , d, d, % o f the totol 12 66 22
L I
2 4 6 8 10 0
Figure 3. Gel filtration of the pre-purified ion exchange chromatographic fractions and corresponding SDS-PAGE electrophoresis.
peak was very high. However the use of defatted flour or the addition of P-mercaptoethanol in the extraction and chromatography buffers did not change either the elution profiles obtained on DEAE Sepharose and on Ultrogel ACA 34 on the turbidity of these peaks. These observations were in good agreement with those of Casey' who showed that addition of dithiothreitol during extraction had no effect on the analytical-ultracentrifuge pattern of the globulins.
The peak B was fractionated in three fractions (bl, b2 and b3). bl only, which represented about 42% of B, corresponded to vicilin, according to the electrophoresis pattern which showed the characteristic bands of this globulin around 50000, 33000, 20000 and 17000 mol. wt (Figure 3). In the peak bz (58% of B) were found mostly albumins, whereas non-protein material was eluted in b3. These albumins seemed to be very similar to those collected in the peaks a6 and a7, showing also a strong electrophoresis band at 27000 mol. wt. They could be decreased by collecting only the last part of the B fractions, but this led to a decrease of about 25% of the vicilin recovery yield.
The gel filtration of the vicilin-rich fraction which was adsorbed on DEAE Sepharose (peak c )
1030 J. Gueguen et al.
led to a major peak with two shoulders in its leading edge. The deconvolution of this peak by a HP 25 computer, showed that it was in fact composed of three fractions cl, c2 and c3 (Figure 3). As shown by the SDS-PAGE, the first and the second fractions (c1 and ~ 2 ) contained convicilin, characterised by a band at 71000 mol. wt, whereas the last (c3) was mainly composed of vicilin. The comparison of the vicilin fractions bl and c3 showed that they were both eluted for the same elution volume (4.3 litres), corresponding to a molecular weight of about 160000; but according to electrophoresis (Figure 3) their subunit composition was different. The vicilin c3 showed a very strong electrophoretic band at 30000 mol. wt which seemed to be very minor in the case of vicilin bl. These results confirm the subunit heterogeneity of vicilin and are in good agreement with those reported by Gatehouse et al.9 on pea.
The legumin was purified as a straight peak d2 from the ion-exchange legumin-rich fraction (D) (Figure 3). This peak dz represented about 66% of D . The edge and. the tail of the peak (fractions dl and d3) had to be discarded because they were contaminated by a protein containing polypeptides of 90000 mol. wt and by vicilin respectively. This phenomenon could be avoided if the ion exchange fraction (C) was desorbed with a 0 . 0 9 ~ NaCl buffer instead of 0 . 5 ~ NaC1, but then the recovery yield of purified legumin was decreased by about 20%. In conclusion, only
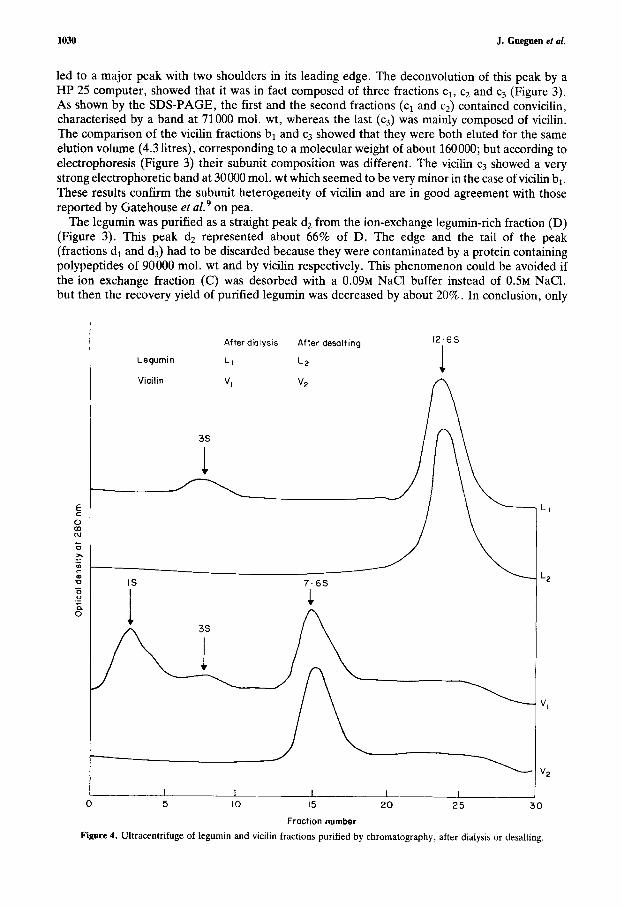
After dialysis After desalting 12-6s
Legurnin L I LP
Vicilin Vl VZ
3s
IS 7 . 6 5
n 1 3s
1 n ?"
I I 1 I 1
Fraction number Figure 4. Ultracentrifuge of legumin and vicilin fractions purified by chromatography, after dialysis or desalting.

Large-sale purification of pea globulins 1031
peaks bl and d2 were kept as purified vicilin and legumin fractions. Fraction c3 was generally discarded because of the low resolution observed between c2 and c3 which led to a high risk of contamination of this vicilin fraction by convicilin. Before freeze-drying, the two purified fractions (bl and d2) were desalted through a column of Trisacryl GF05.
3.1.3. Desalting Desalting was chosen rather than dialysis because it led to the production of more soluble products after freeze-drying. When the collected protein fractions were dialysed at 4°C against distilled water or even against 5 0 m ~ borate buffer pH8, no more than 80% of the freeze-dried protein were soluble. On the other hand when the sample was desalted through the Trisacryl GF05 bed, using distilled water as eluting buffer, and then freeze-dried as before, soluble material was produced.
The long stay of the globulin at 4°C (5-10 days) and low ionic strength probably induced the formation of insoluble aggregates during dialysis. The comparison by ultracentrifuging the desalted sample and the soluble fraction of the dialysed proteins showed that dissociation also occurred during dialysis. Subunits having sedimentation coefficients of 3s in the case of legumin and 3s-1s in the case of vicilin appeared in dialysed samples (Figure 4).
3.2. Composition and characterisation of the purified globulins. From the electrophoretic pattern (Figure 3) and the ultracentrifuge results (Figure 4) the vicilin and legumin fractions, bl and d2, obtained by the procedure described were very highly purified. Sedimentation coefficients Szo,w of these two fractions, determined by the method of Martin and Ames14 (Figure 4) were 7.6s and 12.6s respectively for the vicilin and the legumin fractions. These values were very close to those given by Derbyshire et a1.l’ in their review and by Casey,’ but a little higher than those reported by Gwiazda et a1.” who found 6.4s and 11.9s respectively.
The amino acid analyses (Table 1) were very similar to those generally quoted in the paper^.^,'^, l9 Legumin contained more sulphur amino acids and arginine, whereas vicilin was richer in isoleucine, leucine, phenylalanine and lysine. Compared to bl, the other vicilin fraction (c3) was shown to be very similar in amino acid composition. c3 contained a little more aspartic and glutamic acids and arginine. The higher content of diacid amino acids confirms the charge heterogeneity of these two forms of vicilin, and may explain the absorption of c3 vicilin on the anion-exchanger .
Table 1. Amino acid composition (g116 g N) of vicilin and legumin fractions
ASP T h I Ser Glu Pro
Ala Val Met Ile Leu TY r Phe His LY S
Arg CYS
GlY
Vicilin fractions Legumin fraction
b, c3 d,
13.44 13.97 11.22 2.46 2.22 2.38 5.15 5.28 3.91
19.29 21.40 19.67 4.35 4.38 4.78 3.12 3.04 3.83 3.76 3.30 4.03 5.02 4.93 4.30 0.53 0.23 0.59 5.36 5.39 3.76
10.48 10.70 7.60
5.70 6.28 4.41 2.24 2.33 3.01 8.52 8.51 4.84 7.72 8.40 11.35 0.56 0.20 1.54
3.05 3.41 2.86

1032 J. Gueguen et al.
Figure 5. Purity of vicilin and legumin fractions by immunodiffusion. A: centre, vicilin antiserum; 1, ultracentrifuged vicilin; 2, chromatographic vicilin fraction; 3, ultracentrifuged legumin; 4, chromatographic legumin fraction; 5, blank; 6 , total protein extract of pea seeds. B: centre, legurnin antiserum; 1, chromatographic vicilin fraction; 2, ultracentrifuged vicilin; 3, ultracentrifuged legumin; 4, chromatographic legumin fraction; 5, blank; 6 , total protein extract of pea seeds.
Figure 6. Comparative immunoelectrophoresis of (a) purified and native vicilin; and (b) legumin.
The purity of fractions bl and d2 was also verified by immunochemical analysis. Because the ultracentrifuge diagrams of vicilin did not have a steady base line in the 12s region, the cross-contamination between the two globulins was studied. No precipitation lines appeared between vicilin (fraction bl) and anti-legumin serum or between legumin and anti-vicilin serum (Figure 5) . Moreover the precipitation lines obtained by immunoelectrophoresis with the proteins either purified or in the total extract were located at exactly the same place, showing that vicilin and legumin had, after chromatography, a conformation very similar to their native one (Figure 6). This result is very important, and allows the use of purified bl and d2 fractions for basic studies on the functional properties of vicilin and legumin.
4. Conclusion
The simplicity of the procedure described in this paper and the purity of the vicilin and legumin fractions which have been obtained, allow this chromatography method to be used in a preparative manner.

Large-scale purification of pea globulins 1033
As a routine purification process, three DEAE Sepharose chromatography preparations followed by two gel filtrations were used. At the first step, 1 litre of raw extract was fractionated for each ion exchange chromatographic stage. At the second step, the vicilin and legumin fractions bl and d2 were pooled separately, concentrated by ultrafiltration and purified on the Ultrogel ACA 34 bed. Such a separation cycle could be done in a week and produced about 5 g of vicilin and 7 g of legumin. This procedure may also be simplified if only vicilin is needed. In this case, the DEAE chromatography can be shortened by washing the column directly with 0 . 5 ~ NaCl buffer instead of eluting it by the buffer sequence.
References
1. Vose, J. R. Production and functionality of starches and protein isolates from legume seeds (field pea and horsebeans). Cereal Chem., 1980, 57, 406410.
2. Summer, A . K.; Nielson, M. A,; Youngs, C. G. Production and evaluation of pea protein isolate. 1. Food Sci. 1981, 46, 364-372.
3. Gueguen, J. Legume seed protein extraction processing and end product characteristics. In Plant Proteins for Human Food (Bodwell, C. E. ; Petit, L., Eds), Martinus Nijhoff Publishers, 1983, pp. 63-99.
4. Wright, D. J . Comparative physical and chemical aspects of vegetable protein functionality. In Plant Proteim for Human Food (Bodwell, C. E.; Petit, L., Eds), Martinus Nijhoff Publishers, 1983, pp. 185196.
5. Cascy, R. Immunoaffinity chromatography as a means of purifying legumin from Pisum (pea) seeds. Biochem. 1.1979,
6. Gatehouse, J . A , ; Croy, R . R. D.; Boulter, D. Isoelectric focusing properties and carbohydrate content of pea (Pisum sativum) Legurnin. Biochem. J. 1980. 185, 497-503.
7. Croy, R. R. D.: Gatehouse, J. A.; Tyler, M.; Boulter, D. The purification and characterization of a third storage protein (convicilin) from the seeds of pea (Pisum sativurn L.). Biochem. J . 1980, 191, 509-516.
8. Thornson, J. A , ; Schroeder, H. E.; Tassie, A. M. Cotyledonary storage proteins in Pisurn sarivum. V. Further studies on molecular heterogeneity in the vicilin series of holoproteins. Aust. J . Plant. Physiol. 1980, 7, 271-282.
9. Gatehouse, J. A , ; Croy, R. R. D.; Morton, H.; Tyler, M.; Boulter, D. Characterisation and subunit structures of the vicilin storage proteins of pea (Pisurn sativum L.). Eur. 1. Biochem. 1981, 118, 627-633.
10. Danielson, C. E. Seed globulins of the Graminae and Leguminosae. Biochem. 1. 1949, 44, 387-400. 11. Scholz, G.; Richter, J.; Mauteuffel. R. Studies on seed globulins from legumes. I. Separation and purification of
legurnin and vicilin from Vicia faba L. by zone precipitation. Biochem. Physiol. Pflanzen. 1974, 166, 16S172. 12. Grant, D. R.; Lawrence, J . M. Effects of sodium dodecyl sulfate and other disssociation reagents on the globulins of
peas. Arch. Biochem. Biophys. 1964, 108, 552-561. 13. Laemmli, U. K. Cleavage of structural proteins during the assembly of the head of Bacteriophage T4. Nafure (Lond.)
1Y70, 277, 680-685. 14. Martin, R. G.; Ames, B. N. A method for determining the sedimentation bchaviour of enzymes. Application to
protein mixtures. 1. Biol. Chem. 1961, 236, 1372-1379. 15. Moore, S. On the determination of cystine as cysteic acid. J. Biol. Chem. 1963, 238, 235-237. 16. Ouchterlony, 0. Antigen-antibody reaction in gels. Acfa patho/. rnicrobiol. Scnnd. 1949, 26, 507-515. 17. Scheidegger. J. J . Unc micromethode de I’immunoclectrophortse. Int. Arch. Allergy 1955, 7 , lOS110. 18. Derbyshire, E.; Wright, D. J . ; Boulter, D. Review: Legumin and v n, storage proteins of legume seeds.
Phyfochemisrry (Oxf. J 1976, 15, 3-24. 19. Thomson, J. A. ; Schroeder, H. E. ; Dudman, W. F. Cotyledonary storage proteins in Pisum. I. Molecular
heterogeneity. Aust. J . Plant. Physiol. 1978, 5, 26S279. 20. Gwiazda, S.; Schwenke, K . D.; Rutkowski, A . Isolation and partial characterization of proteins from pea (Pisum
safivum L.). Die Nahrung 1980,24,939-950.
177, 509-520.




















