
Lactose Synthase AN INVESTIGATION OF THE INTERACTION SITE OF a-LACTALBUMIN FOR GALACTOSYLTRANSFERASE...
9
Lactose Synthase AN INVESTIGATION OF THE INTERACTION SITE OF a-LACTALBUMIN FOR GALACTOSYLTRANSFERASE BY DIFFERENTIAL KINETIC LABELING* (Received for publication, April 24, 1979, and in revised form, December 4, 1979) Richard H. Richardson and Keith Brew$ From the Department of Biochemistry, University of Miami, School of Medicine, Miami, Florida 33101 The region of bovine a-lactalbumin which interacts with bovine colostrum galactosyltransferase in the lac- tose synthasecomplex has been examined by differen- tial labeling of the amino groups with acetic anhydride. a-Lactalbumin was acetylated in the free state and in a 1:l complex with galactosyltransferase in the pres- ence of Mn2+, UDP-glucose, and N-acetylglucosamine with trace amounts of high specific activity [3H]acetic anhydride and was separated from each solution by gel filtration. The sample modified in the presence of ga- lactosyltransferase contained 74% as much 3H label as that modified in the free state; both samples contained less than 0.2 acetyl groups/protein molecule. A portion of each sample was mixed with an aliquot of a-lactal- bumin that had been acetylated with excess [‘4C]acetic anhydride under denaturing conditions to facilitate the determination of the relative reactivities of individual amino groups through 3H/’4C ratios. The protein sam- ples were completely acetylated with unlabeled acetic anhydride, reduced, carboxymethylated, cleaved with cyanogen bromide to give the two cyanogen bromide fragments (CN1 and CN2), which were further sub- cleaved enzymically. Radioactive peptides each con- taining each of the 12 lysines and the a-amino group were purified by column chromatography and located in the sequence by amino acid analysis. Examination of the 3H/14C ratios from the a-lactalbumin modified in free solution revealed a range of reactivities of the amino groups, which are related to their environments in previously proposed models for the structure of the protein. By comparison with this control sample, the protein modified in thecomplex showed major pertur- bations in reactivity in only two groups, lysine 5, which was reduced in reactivity by a factor of 3, and lysine 114, which was increased in reactivity by a factorof 2. The amino groups of these residues are proximally situated in an area of the a-lactalbumin molecule neigh- boring the region corresponding to one end of the active site cleft region in the homologous lysozymes. The ori- gin of the differential reactivities and their relevance to the structure, function, and evolution of a-lactalbu- min are discussed. The lactose synthase enzyme system (see Refs. 1 and 2 for * This work was supported by Grant GM 21363 from the National Institute of General Medical Sciences, National Institutes of Health. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. $Recipient of Research Career Development Award No. KO4 GM00147 from the National Institute of General Medical Sciences, National Institutes of Health. review) consists of two proteins, galactosylt,ransferase and a- lactalbumin, which together catalyze the synthesis of lactose in the lactating mammarygland (Reaction 1). UDP-D-galactose + D-glucose + lactose + UDP (1) Galactosyltransferase isfound in a number of tissues besides the mammary gland and is an intrinsic component of the membranes of the Golgi apparatus. In these other tissues, galactosyltransferase catalyzes the transfer of galactose from UDP-galactose to terminal GlcNAc’ residues in the oligosac- charide moieties of glycoproteins. The enzyme can also cata- lyze the transfer of galactose to the monosaccharide GlcNAc and tooligomers of GlcNAc, but glucose is anextremely poor substrate with a K,,, of about 2 M. @-Lactalbumin, which is produced in significant amounts only by the lactating mam- mary gland, effectively reduces the K, of galactosyltransferase for glucose by 3 orders of magnitude, so that lactose synthesis is catalyzed at physiological concentration of glucose. Kinetic and binding studies indicate that a-lactalbumin affects the binding of glucose to galactosyltransferase through its ability to bind synergistically with the monosaccharide to the enzyme to form 1:l complexes. Thus, glucose (and other monosaccha- rides) tighten the binding of a-lactalbumin and vice versa. The elucidation of the primary structures of bovine and a number of other a-lactalbumins has clearly established that they arehomologous with lysozymes of the chicken egg white type (3). However, little resemblance can be seen in functional properties of a-lactalbumins and lysozymes, a-lactalbumins having no catalytic activity, and lysozymes having no ability to affect monosaccharide binding with galactosyltransferase. Despite a considerable number of previous chemical modi- fication studies (see Ref. 2), little specific information about the nature of functional areas in the a-lactalbumin molecule has been obtained. Such information is needed to develop an understanding of its regulatory effects on galactosyltransfer- ase and of the structural basis of its functional differences from lysozyme. We report here the results of an alternative chemical ap- proach to theidentification of functional areas in a-lactalbu- min based on a procedure developed by Kaplan et al. (4) and Bosshard et al. (5). In this, a-lactalbumin in free solution and ina complex with galactosyltransferasewas modified with trace amounts of [“Hlacetic anhydride. By determining the extent of labeling of the individual amino groups of a-lactal- bumin (a-NH2 and 12 lysyl €-amino groups) under these two conditions, two NH2 groups whose reactivities are perturbed on complex formation have been identified. These appear to be in close proximity in the three-dimensional structure of u- lactalbumin. The results suggest that a region in a-lactalbumin corresponding to part of the active site cleft region in lysozyme I The abbreviation used is: GlcNAc, N-acesylglucosamine. 3377
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Lactose Synthase AN INVESTIGATION OF THE INTERACTION SITE OF a-LACTALBUMIN FOR GALACTOSYLTRANSFERASE...

Lactose Synthase AN INVESTIGATION OF THE INTERACTION SITE OF a-LACTALBUMIN FOR GALACTOSYLTRANSFERASE BY DIFFERENTIAL KINETIC LABELING*
(Received for publication, April 24, 1979, and in revised form, December 4, 1979)
Richard H. Richardson and Keith Brew$ From the Department of Biochemistry, University of Miami, School of Medicine, Miami, Florida 33101
The region of bovine a-lactalbumin which interacts with bovine colostrum galactosyltransferase in the lac- tose synthase complex has been examined by differen- tial labeling of the amino groups with acetic anhydride. a-Lactalbumin was acetylated in the free state and in a 1:l complex with galactosyltransferase in the pres- ence of Mn2+, UDP-glucose, and N-acetylglucosamine with trace amounts of high specific activity [3H]acetic anhydride and was separated from each solution by gel filtration. The sample modified in the presence of ga- lactosyltransferase contained 74% as much 3H label as that modified in the free state; both samples contained less than 0.2 acetyl groups/protein molecule. A portion of each sample was mixed with an aliquot of a-lactal- bumin that had been acetylated with excess [‘4C]acetic anhydride under denaturing conditions to facilitate the determination of the relative reactivities of individual amino groups through 3H/’4C ratios. The protein sam- ples were completely acetylated with unlabeled acetic anhydride, reduced, carboxymethylated, cleaved with cyanogen bromide to give the two cyanogen bromide fragments (CN1 and CN2), which were further sub- cleaved enzymically. Radioactive peptides each con- taining each of the 12 lysines and the a-amino group were purified by column chromatography and located in the sequence by amino acid analysis. Examination of the 3H/14C ratios from the a-lactalbumin modified in free solution revealed a range of reactivities of the amino groups, which are related to their environments in previously proposed models for the structure of the protein. By comparison with this control sample, the protein modified in the complex showed major pertur- bations in reactivity in only two groups, lysine 5, which was reduced in reactivity by a factor of 3, and lysine 114, which was increased in reactivity by a factor of 2. The amino groups of these residues are proximally situated in an area of the a-lactalbumin molecule neigh- boring the region corresponding to one end of the active site cleft region in the homologous lysozymes. The ori- gin of the differential reactivities and their relevance to the structure, function, and evolution of a-lactalbu- min are discussed.
The lactose synthase enzyme system (see Refs. 1 and 2 for * This work was supported by Grant GM 21363 from the National
Institute of General Medical Sciences, National Institutes of Health. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$Recipient of Research Career Development Award No. KO4 GM00147 from the National Institute of General Medical Sciences, National Institutes of Health.
review) consists of two proteins, galactosylt,ransferase and a- lactalbumin, which together catalyze the synthesis of lactose in the lactating mammary gland (Reaction 1).
UDP-D-galactose + D-glucose + lactose + UDP (1)
Galactosyltransferase is found in a number of tissues besides the mammary gland and is an intrinsic component of the membranes of the Golgi apparatus. In these other tissues, galactosyltransferase catalyzes the transfer of galactose from UDP-galactose to terminal GlcNAc’ residues in the oligosac- charide moieties of glycoproteins. The enzyme can also cata- lyze the transfer of galactose to the monosaccharide GlcNAc and to oligomers of GlcNAc, but glucose is an extremely poor substrate with a K,,, of about 2 M. @-Lactalbumin, which is produced in significant amounts only by the lactating mam- mary gland, effectively reduces the K, of galactosyltransferase for glucose by 3 orders of magnitude, so that lactose synthesis is catalyzed at physiological concentration of glucose. Kinetic and binding studies indicate that a-lactalbumin affects the binding of glucose to galactosyltransferase through its ability to bind synergistically with the monosaccharide to the enzyme to form 1:l complexes. Thus, glucose (and other monosaccha- rides) tighten the binding of a-lactalbumin and vice versa.
The elucidation of the primary structures of bovine and a number of other a-lactalbumins has clearly established that they are homologous with lysozymes of the chicken egg white type (3). However, little resemblance can be seen in functional properties of a-lactalbumins and lysozymes, a-lactalbumins having no catalytic activity, and lysozymes having no ability to affect monosaccharide binding with galactosyltransferase.
Despite a considerable number of previous chemical modi- fication studies (see Ref. 2 ) , little specific information about the nature of functional areas in the a-lactalbumin molecule has been obtained. Such information is needed to develop an understanding of its regulatory effects on galactosyltransfer- ase and of the structural basis of its functional differences from lysozyme.
We report here the results of an alternative chemical ap- proach to the identification of functional areas in a-lactalbu- min based on a procedure developed by Kaplan et al. (4) and Bosshard et al. ( 5 ) . In this, a-lactalbumin in free solution and in a complex with galactosyltransferase was modified with trace amounts of [“Hlacetic anhydride. By determining the extent of labeling of the individual amino groups of a-lactal- bumin (a-NH2 and 12 lysyl €-amino groups) under these two conditions, two NH2 groups whose reactivities are perturbed on complex formation have been identified. These appear to be in close proximity in the three-dimensional structure of u- lactalbumin. The results suggest that a region in a-lactalbumin corresponding to part of the active site cleft region in lysozyme
I The abbreviation used is: GlcNAc, N-acesylglucosamine.
3377

3378 Interaction Site of a-Lactalbumin for Galactosyltransferase
may function in interacting with galactosyltransferase and monosaccharides.
EXPERIMENTAL PROCEDURES'
Materials-Bovine a-lactalbumin and bovine colostrum galacto- syltransferase were prepared as described previously (9). Galactosyl- transferase was estimated by measurement of activity using a specific activity of 18 units/mg (6).
['HIAcetic anhydride (5.3 Ci/mmol) was purchased from Amer- sham/Searle and [l-"C]acetic anhydride (8.6 mCi/mmol) from New England Nuclear. Cyanogen bromide and pyridine were the products of J. T. Baker, iodoacetic acid was the product of Aldrich, and N - acetyl-D-glucosamine and 2,5-diphenyloxazole (PPO) were Sigma products. Triton X-100, 1,4-bis[2-(5-phenyloxazolyl)]benzene (PO- POP), and toluene were obtained from Research Products Interna- tional. Reagents for manual Edman degradation (phenylisothiocyan- ate and anhydrous heptafluorobutyric acid, both of Sequanal grade) were purchased from Pierce.
Thermolysin (A grade) was purchased from Calbiochem and tryp- sin-TPCK (L-1-tosylamido-2-phenylethyl chloromethyl ketone) (228 units/mg) and a-chymotrypsin (65 units/mg) from Worthington.
Aminex A-5 of particle size 13 f 2 p was purchased from Bio-Rad Laboratories and Partisil-10 ODS-2 from Reeve Angel.
"H Acetylation of a-Lactalbumin in Free Solution and in a Com- plex with Galactosyltransferase-Two identical reaction mixtures were prepared except for the presence of galactosyltransferase (2.80 mg; 56 nmol; 2 p ~ ) in one. Both solutions contained 0.795 mg (56 nniol; 2 p ~ ) of bovine a-lactalbumin, 0.3 mM UDP-Glc, 1 mM MnClr, and 0.2 M NaCl in 0.02 M Tris-HCl buffer, pH 8.0. The total volume of each solution was 28 ml. T o each was added ["Hlacetic anhydride (1.23 pmol, 8.3 mCi) in 110 pl of dry, freshly distilled acetonitrile. The modification was performed at 0°C and the reaction was left for 4 h to complete.
"H-acetylated a-lactalbumin was isolated from the mixture by gel filtration. Each reaction mixture was applied, in two separate lots (14.0 ml), to columns (2.5 X 90 cm) of Bio-Gel P-60 equilibrated and eluted with 0.1 M NH.,HCO,I a t 4°C. Fractions of 7.5 ml were collected and aliquots of 10 pl were taken for scintillation counting (see Fig. 1). The combined fractions were freeze-dried and stored at -20'C.
Determination of Radioactiuity-"H/'JC ratios for individual ly- sine residues were determined by counting an aliquot of the purified peptide in 7.5 ml of scintillation fluid and a total of 0.75 ml of water. Samples containing pyridine were first dried in a desiccator with PzOs and redissolved in H,O. The overlap of counts in the 'H channel was 33R, whereas there was no overlap of "H into I4C. Counts were corrected for this overlap before calculation of ratios. Ratios for residues from a-lactalbumin modified in free solution are denoted by r,, whereas ratios for a-lactalbumin modified in complex with galac- tosyltransferase are denoted by r,. A protection factor, R, is defined by R = r,/r<.
Radioactivity in column eluates was measured by counting an aliquot (50 to 100 pl) in 2.5 ml of scintillat,ion fluid with sufficient water to give a total volume of 2.75 ml.
RESULTS
Protective Labeling Procedures-From previous studies, it is known that the association between a-lactalbumin and galactosyltransferase is highly ligand-dependent and is also affected by external conditions such as ionic strength, pH, and temperature (6,7). Of the various complexes whose association constants have been determined, the strongest is a dead end inhibitory complex that resembles a central complex in the catalytic cycle of lactose synthase: galactosyltransferase- Mn'+. UDP-glucose. glucose .a-lactalbumin, which at 16°C and pH 7.4 has a K , value of 6 X 10" M". As the association is strengthened by lower temperatures and higher pH values, the conditions used for modification were chosen to optimize
'' Portions of this paper (including parts of "Experimental Proce- dures" and "Results," Fig. 2 to 8 and Table 111) are presented in miniprint at the end of this paper. Miniprint is easily read with the aid of a standard magnifying glass. Full size photocopies are available from the Journal of Bioiogical Chemistry, 9650 Rockville Pike, Be- thesda, Md. 20014. Request Document No. 79M-790, cite author(s), and include a check or money order for $1.80 per set of photocopies.
these parameters to achieve maximum binding. GlcNAc was also substituted for glucose as galactosyltransferase has a higher affinity for this monosaccharide (9-11) and maximum binding can be achieved at lower concentrations. From kinetic studies, the association constant for a-lactalbumin with a galactosyltransferase Mn2'. UDP-galactose . GlcNAc complex has been calculated to be 3.3 X lo6 M" at pH 7.4 and 37°C (11); allowing for the effect of temperature and pH on binding (6, 7), a value of more than 10' M" would be estimated at 0°C. The exact conditions for the modification are described under "Experimental Procedures." Although the precise value of the association constant for a-lactalbumin and galactosyl- transferase under these conditions cannot be measured by our procedures, because the ultracentrifuge cannot be cooled to O"C, extrapolation suggests that it should be considerably greater than that for the complex containing Mn", UDP- glucose, and glucose at 16°C (8). If the K, for this complex is taken as a minimum value, it can be calculated that at the protein concentrations used in the experiment, a t least 75% of the a-lactalbumin and galactosyltransferase will exist as a complex. Since the labeling procedure is a kinetic one, even this degree of association is adequate for our purposes. How- ever, the extent of association would be expected to be greater than this minimum value. The same ligands were also included in the control samples of a-lactalbumin so as to ensure that the experimental conditions, other than the absence of galac- tosyltransferase from the control solution, were identical. After addition of ['Hlacetic anhydride (1.25 pmol, 8.3 mCi), the acetylation was allowed to proceed with stirring for 4 h a t 0°C to ensure complete reaction, and the products were then separated by gel fdtration with columns of Bio-Gel P-60 (Fig. 1, A and B) . From the reaction mixt,ure containing both proteins, three peaks of radioactivity were obtained. The first corresponded in elution position to galactosyltransferase while the second eluted in the same place as a-lactalbumin; the third peak, which contained 97% of the radioactivity, is as- sumed to be ["Hlacetate formed by hydrolysis of the reagent. From the control reaction mixture, only two peaks, one of [:1H]acetyl a-lactalbumin and one of ["Hlacetate, were ob- tained.
The extent of labeling of a-lactalbumin under these two conditions was determined by counting aliquots from the a- lactalbumin pools, which are indicated in Fig. 1, A and B. The a-lactalbumin modified in free solution contained 1.65 X lo6 cpm, whereas that modified in the presence of galactosyltrans- ferase contained 1.22 X lo6 cpm. By comparing the radioactiv- ity in the a-lactalbumin pools with the total amount of radio- activity recovered from the columns, the percentage of incor- poration into free and complexed a-lactalbumin was calcu- lated to be 0.39 and 0.29%, respectively. These correspond to 0.160 and 0.118 mol of acetyl group incorporated/mol of a- lactalbumin for the a-lactalbumin samples labeled in free solution and in the complex, respectively. Since the bulk of the acetic anhydride under both conditions was hydrolyzed, the difference cannot be attributed to removal of reagent through reaction with galactosyltransferase, because only 2% of the acetic anhydride reacted with the enzyme. It is therefore reasonable to assume that the 26% lower incorporation ob- served in the presence of galactosyltransferase reflects the protection of regions of a-lactalbumin from the reagent when a-lactalbumin is in a complex with galactosyltransferase.
Distribution of 3H Label Among Individual Amino Groups-To determine the pattern of labeling of the individ- ual amino groups in the two a-lactalbumin samples, a portion of each, 4.66 X lo6 cpm of ,'H (complex) and 6.30 X lo6 cpm of 'H (free solution), was mixed with unlabeled carrier a-lactal- bumin, and uniformly labeled N-['4C]acetyl a-lactalbumin was
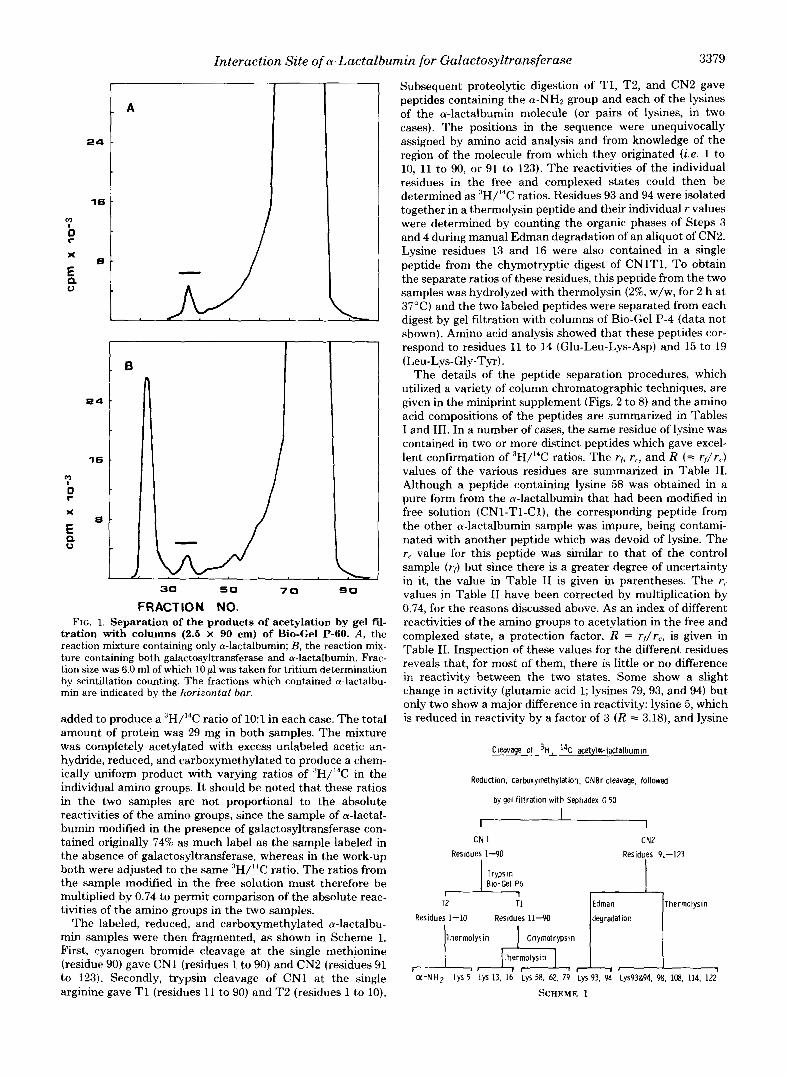
Interaction Site of a-Lactalbumin for Galuctosyltrunsferase 3379
m :
r 0
X
E a 0
16
e
30 5 0 7 0 90
FRACTION NO. FIG. 1. Separation of the products of acetylation by gel fil-
tration with columns (2.5 X 90 cm) of Bio-Gel P-60. A, the reaction mixture containing only a-lactalbumin; B , the reaction mix- ture containing both galactosyltransferase and a-lactalbumin. Frac- tion size was 6.0 ml of which 10 p1 was taken for tritium determination by scintillation counting. The fractions which contained a-lactalbu- min are indicated by the horizontal bar.
added to produce a "H/I4C ratio of 1O:l in each case. The total amount of protein was 29 mg in both samples. The mixture was completely acetylated with excess udabeled acetic an- hydride, reduced, and carboxymethylated to produce a chem- ically uniform product with varying ratios of :'H/14C in the individual amino groups. I t should be noted that these ratios in the two samples are not proportional to the absolute reactivities of the amino groups, since the sample of a-lactal- bumin modified in the presence of galactosyltransferase con- tained originally 74% as much label as the sample labeled in the absence of galactosyltransferase, whereas in the work-up both were adjusted to the same :3H/'4C ratio. The ratios from the sample modified in the free solution must therefore be multiplied by 0.74 to permit comparison of the absolute reac- tivities of the amino groups in the two samples.
The labeled, reduced, and carboxymethylated a-lactalbu- min samples were then fragmented, as shown in Scheme 1. First, cyanogen bromide cleavage at the single methionine (residue 90) gave CNl (residues 1 to 90) and CN2 (residues 91 to 123). Secondly, trypsin cleavage of CN1 at the single arginine gave T1 (residues 11 to 90) and T2 (residues 1 to 10).
Subsequent proteolytic digestion of TI , T2, and CN2 gave peptides containing the a-NH2 group and each of the lysines of the a-lactalbumin molecule (or pairs of lysines, in two cases). The positions in the sequence were unequivocally assigned by amino acid analysis and from knowledge of the region of the molecule from which they originated (i.e. 1 to 10, 11 to 90, or 91 to 123). The reactivities of the individual residues in the free and complexed states could then be determined as .'H/14C ratios. Residues 53 and 94 were isolated together in a thermolysin peptide and their individual r values were determined by counting the organic phases of Steps 3 and 4 during manual Edman degradation of an aliquot of CN2. Lysine residues 13 and 16 were also contained in a single peptide from the chymotryptic digest of CNlT1. To obtain the separate ratios of these residues, this peptide from the two samples was hydrolyzed with thermolysin (2%, w/w, for 2 h a t 37°C) and the two labeled peptides were separated from each digest by gel filtration with columns of Bio-Gel P-4 (data not shown). Amino acid analysis showed that these peptides cor- respond to residues 11 to 14 (Glu-Leu-Lys-Asp) and 15 to 19 (Leu-Lys-Gly-Tyr).
The details of the peptide separation procedures, which utilized a variety of column chromatographic techniques, are given in the miniprint supplement (Figs. 2 to 8) and the amino acid compositions of the peptides are summarized in Tables I and 111. In a number of cases, the same residue of lysine was contained in two or more distinct peptides which gave excel- lent confirmation of 3H/14C ratios. The rf, r,, and R (= rf /rJ values of the various residues are summarized in Table 11. Although a peptide containing lysine 58 was obtained in a pure form from the a-lactalbumin that had been modified in free solution (CN1-TI-Cl), the corresponding peptide from the other a-lactalbumin sample was impure, being contami- nated with another peptide which was devoid of lysine. The r, value for this peptide was similar to that of the control sample (r,) but since there is a greater degree of uncertainty in it, the value in Table I1 is given in parentheses. The r,. values in Table 11 have been corrected by multiplication by 0.74, for the reasons discussed above. As an index of different reactivities of the amino groups to acetylation in the free and complexed state, a protection factor, R = rf /rc , is given in Table 11. Inspection of these values for the different residues reveals that, for most of them, there is little or no difference in reactivity between the two states. Some show a slight change in activity (glutamic acid 1; lysines 79,93, and 94) but only two show a major difference in reactivity: lysine 5, which is reduced in reactivity by a factor of 3 ( R = 3.18), and lysine
Cleavage of 'H , 14C acetyl=-lactalbumin
Reduction. carboxymethylation. CNBr cleavage, followed
by gel filtration with Sephadex G 50
I CN 1 CN2
Residues 1-90 Residues 91-123
T1 Edman Thermolysin
Resldues 1-10 Residues 11-90 degradation
Thermolysin
O(-NH2 LyS 5 LYS 13, 16 LyS 58, 62, 79 Lys 93, 94 Lys93&94, 98, 108. 114, 122
SCHEME 1

3380 Interaction Site of a-Lactalbumin for Galactosyltransferase
TABLE I Amino acid compositions of labeled peptides from a-lactalbumin acetylated in free solution
The peptide compositions given are for the purest form obtained in those cases where two or more forms were obtained. The bottom line indicates the pools from which the peptides were obtained and refers to Fig. 4 to 8. All pools were further purified by gel Ntration with columns of Bio-Gel P-4 eauilibrated and eluted with 0.1 M NHIHCOe before analvsis and determination of 3H/14C ratios.
3-7 11-18 11-14 5-18 69-90 92-94 105-110 110-114 61-68 Residues
Amino group
CMC"
ASP
Thr
Ser
Glu/Homoserine
Pro
GlY
Ala
Val
Ile
Leu
Tyr
Phe
His
LYS
Arg Fig. No. Pool No.
1-2
a-NH2
0.14
2.0 (2)
4 1
54-60
Lys 58
2.06 (2)
1.19 (1)
0.42
1.69 (2)
0.97 (1)
5 1
Lys 13
1.06
1.08
1.07
0.89
>ys 114
0.82 (1)
0.14
0.79 (1) 1 .00 (1)
0.15
1.06 (1)
1.00 (1)
8 2
Lys 93 Lys 94 0.89 (1)
1.08 (1)
2.00 (2)
8 3
1.15
1.04
0.91
0.85
i 'I CMC, S-carboxymethylcysteine.
TABLE I1 rb r , and R values for individual amino groups of a-lactalbumin
Multiple values of rf and rc for individual residues were derived from different peptides which originated from partial enzymic cleavage of fragments (see Table I).
Position of amino group in sequence 5 13 16 58 62 79 93 94 98 108 114 122
rf 7.80 31.5 13.5 31.1 11.2 8.92 8.13 5.91" 17.3" 39.7 5.34 4.18 9.09 32.8 "
12.5 38.6 4.78 - 34.2 11.0
21.Oh 21.8' 12.1' 44.4
Average 32.2 11.9 39.2 5.06 10.0
r, 6.47 10.7 13.9 32.2 9.50 8.55 6.54 5.64" 13.7" 32.3 4.59 8.56 8.36 9.55 "
31.2 8.21 34.5 9.22 u 9.03h 21.1' 8.43'
Average 10.1 31.7 8.38 33.4
R (= rf/rJ 1.21 3.18 0.97 0.98 1.31 1.06 1.24 1.05 1.26 1.17 1.10 0.49 1.14
8.79
" u 2.33h 1.03' 1.43'
Determined by Edman degradation of a sample of CN2. ' Values for CNlT2. '' Values derived from peptides containing two lysines.
114, which is increased in reactivity by a factor of 2 ( R = 0.49). understand the structural basis of its functional divergence from lysozyme and to correlate this with x-ray crystallographic studies of the protein (12).
The present study was prompted by a desire to augment The objective of the investigation was to identify residues our limited knowledge of the regions of a-lactalbumin that are of a-lactalbumin that show differential reactivity with acetic involved in its function in lactose synthesis so as to attempt to anhydride in the free state and in a complex with galactosyl-
DISCUSSION

Interaction Site of a-Lactalbumin for Galactosyltransferase 3381
transferase. The technique of using limiting amounts of high specific activity radioactive acetic anhydride for following changes in the reactivities of amino groups of proteins under different conditions was used originally by Kaplan et al. (4) for determining the pK values of individual amino groups in elastase and has since been used to examine the binding site of tyrosyl-RNA synthetase with tRNA (5) and the interaction site of cytochrome c with cytochrome oxidase (13). It is particularly suitable for studies with lactose synthase because the low levels of chemical modification achieved, in our case, less than 0.2 acetyl groups/protein molecule, avoid the possi- bility of perturbing the reversible association through chemi- cal modification of the two proteins, as each modification reaction involves the reaction of the reagent with an unmod- ified protein molecule. The reactivities of individual amino groups will therefore reflect their microenvironments in the protein in its native state.
The choice of acetic anhydride for modification was based on several considerations. At low concentrations, the reagent is highly specific for amino groups; there are 13 such groups in cY-lactalbumin, which are distributed throughout the amino acid sequence and are also widely spread in the putative conformation (Fig. 9). Thus, whatever parts of the molecule are involved in the function, there is a reasonable chance that a lysyl residue is nearby and could be perturbed. Although previous chemical modification studies have shown that ex- tensive maleylation of cu-lactalbumin has little effect on its activity, substitution with the more bulky trinitrobenzene sulfonate group leads to loss of activity (14). Therefore, while amino groups are unlikely to be involved directly in ionic interactions with groups on galactosyltransferase, some amino
groups are likely to be close to the interaction site. In addition, a reagent that is available in radiochemically labeled form at high specific activity is essential with this system since galac- tosyltransferase can only be obtained in limited amounts, and adequate labeling combined with limited chemical modifica- tion is required in the procedure. Specific labeling of amino groups at high specific activity can also be achieved by reduc- tive methylation (13), but this is not possible in the lactose synthase system since GlcNAc, which is required for adequate complex formation, will react with the borohydride used in the modification reaction.
The results presented here for rr and rf values of individual lysines are all obtained from highly purified peptides, apart from lysine 58 (complex), as noted under “Results” The separate values for residues 93 and 94 were obtained by Edman degradation of CN2; their average is in reasonable agreement with the r values obtained for the thermolysin peptides containing both residues. Residues 13 and 16 were also obtained in a single peptide, which was subjected to further cleavage to give peptides containing the spearate residues. Confirmation of the ratio in lysine 16 was afforded by a peptide containing lysine 16 alone (Lys-Gly-Tyr) isolated from the complex sample (Table III). It should be noted that the most effective procedure that we have found for separating the peptides containing acetylated lysines was reverse phase chromatography (see miniprint supplement).
The final stage in the purification of almost all peptides involved gel filtration, thereby removing contaminants such as pyridine that could affect ‘H/l% ratios by differential quenching. The relatively low proportion of uniformly I%- acetylated a-lactalbumin added to the “H-labeled protein prior
14
FIG. 9. A putative structure for bovine a-la&albumin based on the structure of chicken egg white lysozyme, with appro-
side chains of lysines 5 and 114 are shown together with those of
priate deletions (see Refs. 15 and 16), redrawn with modifica- residues His 32 and Trp 118, which are referred to in the text. The
tions from the method of Dayhoff (21). The (Y carbons are shown positions corresponding to the saccharide binding sites C, D, E, and F are shown (dark lines). Subsites A and B have been omitted
as open circles and the positions in the sequence of the residues bearing amino groups are numbered. The possible orientations of the
because of the presence of a tyrosine residue at position 103 of (Y- lactalbumin which is thought to block access to Site B (15, 16).

3382 Interaction Site of a-Lactalbumin for Galactosyltransferase
to cleavage ratio of 1O:l) was chosen to minimize errors stemming from the correction of "H counts for the overlap of I4C into the "H channel in scintillation counting (33%).
The results are relevant to two aspects of the structure and function of a-lactalbumin: the microenvironments of the in- dividual amino groups of a-lactalbumin in free solution and the perturbation of the environments of these groups when a-lactalbumin is combined in a complex with galactosyltrans- ferase. The activities of the amino groups both in free solution and in the complex can be reasonably assumed to reflect their accessibility to the modifying reagent and their pK values since the acylation reaction involves the unprotonated form of the amino group. I t should be emphasized that the solution of a-lactalbumin modified in the absence of galactosyltrans- ferase contained Mn", UDP-galactose, and GlcNAc, and so interaction between any of these ligands and a-lactalbumin might affect the reactivities of the amino groups. Previous studies have shown that GlcNAc does not bind significantly to a-lactalbumin in free solution (7) and, although binding studies with UDP-sugars have not been performed, the results of kinetic studies do not indicate that they interact with a- lactalbumin (9, ll). However, it is possible that the presence of Mn'+ ions could affect the reactivities of specific groups, e.g. the a-NH2 group, by chelation or by effecting conforma- tional changes, and so conclusions about the environments of the groups in the free a-lactalbumin molecule are not entirely reliable. Present information indicates that a-lactalbumin is closely similar in conformation to the mammalian lysozymes and those of the chicken egg white type (see Refs. 1 and 2). Both model building (15) and energy minimization procedures (16) have been used to obtain models of a-lactalbumin based on the structure of lysozyme, from which the environments of a number of the amino groups can be predicted. I t is interest- ing that these models predict the involvement in ionic inter- action with carboxyl groups of two lysine residues of relatively low reactivity in our study (13 and 93) as well as the a-NH, group of glutamic acid 1, which has a low reactivity if the low pK value of a-NH2 groups relative to eNH2 groups is taken into account.
I t is difficult to predict a satisfactory conformation for the COOH-terminal region of a-lactalbumin based on the struc- ture of lysozyme because of major structural changes, includ- ing deletions, in this region of the molecule (15). From the energy minimization studies of Warme et al. (16), three alter- native conformations for this region have been suggested, designated T2, Tn, and T4. In two of these (T:3 and Ts), the conformation is stabilized by a salt bridge between t h e E - N H ~ group of lysine 5 and the a-C02H group of leucine 123. However, our results indicate that the E - N H ~ group of lysine 5 is among the most reactive of the a-lactalbumin molecule, the other highly reactive groups being the E-NH, groups of lysines 16 and 98. Therefore, this would appear to eliminate conformations T B and Tq as representing the structure of a- lactalbumin under our experimental conditions. Conformation T2, in which tryptophan 118 and other hydrophobic residues are more exposed to solvent, is more consistent with our observations. It is interesting to note that the exposure of tryptophan 118 to solvent is also supported by chemical modification studies using 2-hydroxy-5-nitrobenzyl bromide, dimethyl-(2-hydroxy-5-nitrobenzyl)sulfonium bromide and 2- nitrophenylsulfenyl chloride (17, 18).
Nevertheless, there is considerable evidence that a-lactal- bumin is more flexible in structure than lysozyme (see Ref. 16) and it is possible that the conformation in this region may fluctuate under different conditions. The reactivity of the amino group of lysine 94 is also above average, possibly
reflecting its proximity to another positively charged residue (lysine 93). Lysine 16 is also above average, in keeping with its proposed exposed environment (15), while the other amino groups (lysines 13, 58, 62, 79, 108, 114, and 122) show similar lower levels of reactivity.
Examination of the protection factors of the individual residues (Fig. 10 and Table 11) reveals that 10 are clustered between 0.97 and 1.31. It is conceivable that the differences in R values within the main group of 10 residues (Fig. 10) reflect minor perturbations in the reactivities of some of these resi- dues. However, we consider it more probable that this repre- sents the range of error in our procedures. This error range is comparable to that obtained by Bosshard et al. (5) in previous similar studies. The reactivities of these groups, the a-NH, and the E-NHz groups of lysines 13, 16, 58, 62, 79, 93, 94, 98, and 122, are therefore deduced to be unaffected by association with galactosyltransferase. In contrast, the amino groups of lysines 5 and 114 show major changes, the former being decreased in reactivity by a factor of 3 ( R = 3.18), while the latter is increased in reactivity by a factor of 2 ( R = 0.49). These changes, which show an interesting parallel in their magnitudes, must reflect altered environments of these two amino groups when a-lactalbumin enters a complex with galactosyltransferase. The environments of these residues may have changed either as a result of a conformational change induced in the a-lactalbumin molecule when it forms a complex or as a result of their proximities to regions of galactosyltransferase in such a complex. In either case, the reduced reactivity of lysine 5 could result from an increase in the pK value or through rendering it less accessible to the modifying agent. The increased reactivity of lysine 114 may result either from greater exposure of this group, resulting from a conformational change, or possibly from being placed in proximity with another positively charged group on either a-lactalbumin or galactosyltransferase. I t is not possible from our results to choose between these alternatives.
Although the perturbed residues are in distant parts of the primary structure of a-lactalbumin, they are in close proximity in the predicted conformation based on the structure of b o -
1114
1 2 3 R l = r f / r c l
FIG. 10. Values of protection'factors of isolated peptides. The two lysine residues (5 and 114) which were perturbed in their activities towards acetic anhydride are identified by the numbers in parentheses.

Interaction Site of a-Lactalbumin for Galactosyltransferase 3383
zyme and are situated at the lower end of the cleft region as shown in Fig. 9. Although the conformation of the COOH- terminal region of the a-lactalbumin polypeptide chain as shown in this figure is likely to be incorrect in detail, the presence of a disulfide bond between half-cystine residues 5 and 120 provides substantial evidence that the residues are in the same area in the conformation. The localized nature of the perturbed residues, when the wide distribution of the unperturbed residues is taken into account (Fig. 9), demon- strates the very precise information on the nature of interac- tion sites that can be obtained by differential kinetic labeling. It is interesting that other groups, whose modification has been shown to affect the interaction of a-lactalbumin with galactosyltransferase, are also localized in this area of the molecule. These include histidine 32, whose modification by ethoxyformylation results in a major loss of activity (19), and tryptophan 114, whose modification by sulfenylation also leads to inactivation (18). Reduction of the 5-120 disulfide bond followed by carboxymethylation of the cysteines pro- duces a derivative of a-lactalbumin with reduced binding activity (20). The importance of this region in the function of a-lactalbumin is also supported by the fact that many of the residues in this area (including lysine 5, phenylalanine 31, histidine 32, leucine 110, lysine 114, and tryptophan 118) are conserved in the sequences of all species variants hitherto examined (see Ref. 2). Although the mechanism of the altered reactivities of the two lysines cannot be determined, the combined evidence suggests that this area, to the right of the lower end of the cleft region of a-lactalbumin (in the view of the molecule shown in Fig. 9) may constitute the binding site for galactosyltransferase. I t should be noted that this area overlaps with the region in a-lactalbumin corresponding to the monosaccharide binding region designated F in lysozyme (see Ref. 15).
Kinetic and binding studies have led to the hypothesis that a-lactalbumin binds to galactosyltransferase in such a manner as to bring an area of the a-lactalbumin molecule that can contribute favorable binding interactions with monosaccha- rides into close proximity with the acceptor monosaccharide binding site of galactosyltransferases to form a stronger, hy- brid monosaccharide binding site (8). The proximity of the a-lactalbumin binding site on galactosyltransferase to the acceptor binding site is consistent with the fact that a-lactal- bumin is a competitive inhibitor of the binding of the disac- charide substrate N,N-diacetylchitobiose (11). On this basis, it can be noted that the interaction of a-lactalbumin with galactosyltransferase through the site described above will bring a region of a-lactalbumin corresponding to monosaccha- ride binding subsites in lysozyme (Subsites D and E) close to the surface of the enzyme. One of those regions may therefore be the region that aids in the binding of monosaccharides in the lactose synthase complex. This hypothesis provides a slight functional correlation between a-lactalbumin and lyso-
zyme. We would suggest that a major step in the functional divergence of a-lactalbumin from lysozyme during evolution involved structural changes in an area near Subsite F of lysozyme to produce a region through which the protein is able to interact with galactosyltransferase. The structural changes in this area, which have been discussed previously (see also Ref. E ) , appear to make this region more hydropho- bic than the corresponding area of lysozyme.
Studies are in progress to clarify the functions of these areas of a-lactalbumin. It should be emphasized that the full inter- pretation of these results will be greatly aided by the comple- tion of present crystallographic studies of baboon a-lactalbu- min (12).
REFERENCES 1. Brew, K., and Hill, R. L. (1975) Rev. Physiol. Biochem. Phar-
2. Hill, R. L., and Brew, K. (1975) Adu. Enzymol. Relat. Areas Mol.
3. Brew, K., Castellino, F. J., Vanaman, T. C., and Hill, R. L. (1970)
4. Kaplan, H., Stevenson, K. J., and Hartley, B. S. (1971) Biochem.
5. Bosshard, H. R., Koch, G. L. E., and Hartley, B. S. (1978) J. Mol.
6. Powell, J. T., and Brew, K. (1975) J . Biol. Chem. 250,6337-6343 7. Brew, K., Richardson, R. H., and Sinha, S. K. (1979) in Proceed-
ings of the 12th FEBS Meeting, Dresden, 1978 (Hofman, E., Pfeil, W., and Aurich, H., eds) Pergamon Press, Oxford, pp. 317-330
8. Powell, J. T., and Brew, K. (1976) J . Biol. Chem. 251, 3653-3663 9. Khatra, B. S., Herries, D. G., and Brew, K. (1974) Eur. J.
10. Andrews, P., Kitchen, B. J., and Winzor, D. J. (1973) Biochem. J.
11. Bell, J. E., Beyer, T. A,, and Hill, R. L. (1976) J. B i d . Chem. 251, 3003-3013
12. Aschaffenberg, R., Fenna, R. E., Phillips, D. C., Smith, S. G., Buss, D. H., Jenness, R., and Thompson, M. P. (1979) J. Mol. B i d . 127, 135-137
13. Rieder, R., and Bosshard, H. R. (1978) J. Biol. Chem. 253,6045- 6053
14. Kitchen, B., and Barman, T. E. (1973) Biochim. Biophys. Acta 298,861-867
15. Brown, W. J., North A. C. T., Phillips, D. C., Brew, K., Vanaman, T. C., and Hill, R. L. (1969) J. Mol. B i d . 42, 65-86
16. Warme, P. K., Momany, F. A., Rumball, S. V., Tuttle, R. W., and Scheraga, H. A. (1974) Biochemistry 13, 768-782
macol. 72, 105-157
B i d . 43,411-490
J. Biol. Chem. 245,4570-4582
J. 124,289-299
B i d . 119, 377-389
Biochem. 44,537-560
135,897-900
17. Barman, T. E., and Bagshaw, W. (1972) Biochim. Biophys. Acta 278,491-500
18. Shechter, Y., Patchornick, A., and Burstein, Y. (1974) J. B i d . Chem. 249,413-419
19. Schindler, M., Sharon, N., and Prieels, J-P. (1976) Biochem. Biophys. Res. Commun. 69, 167-173
20. Shechter, Y., Patchornik, A,, and Burstein, Y. (1973) Biochemis- try 12, 3407-3413
21. Dayhoff, M. 0. (1969) Atlas of Protein Sequence and Structure Vol. 4, National Biomedical Research Foundation, Silver Spring, Md.
3384 Interaction Site of a-Lactalbumin for Galactosyltransferase
" Proteolytic " O l q e r t i a n r . __ Olge~tlons wi th t ryps in chymot ryps in and t h e m l y r i o R T ~ p e r f o m d u n d e r s i m i l a r c o n d i t i & r . P r o t e i n fragnents were d isso lved ~n 0.5 t o 1.0 ml o f 0.1 M NH HCO , p m t e a ~ e was added t o t h e extent Of 2 1 (wlwl and incubated a t 3 f ~ r I hour f o r t r y p r l n 2 n w r s f o r t h e m l y r i n and 4 hours f o r chymt ryps in . T r y p s l n d i 9 e s t l m ( o f CNII yere a p p l i e d d i r e c t l y t o a c a l m of B io -Ge l P-6 equ l l lb ra ted w i th 0.1 M NH HCO ; chymotrypsin and t h e m l y r i n d i g e s t l o n r were t e m i n a t e d b y 4 t h e 3 a d d i t l o n o f glac la l a c e t i c ac?d t o a Concentration of 301. and a p p l l e d t o c o l m n r o f Amne. A-5.
o f CN1 TI and of t h e m l y r i n d i g e s t i o n Of CHZ and CHI T2 were I o n Exchange Chromataqraohy. The p raduc t r o f c h y m t r y p r i n d i g e s t i o n
separated wth c o l v m r (0.4 x 50 cml Of Iminex A-5 a t 55'. Columr were e q u l l l b r a t e d v l t h 304 ( v l v ) acetic acid and were e l u t e d w i t h two separa te g rad len t r o f py r id ine ace ta te . The f i r s t g r a d i e n t was
0.05 M p y r l d i n e aceta te pH 2.6 and 0 .5 M p y r i d i n e acetate prepared concave and compored o f three equal voIurms (50 m l ] O f 301 HAC
as descr ibed PreYioUsly (31 ~n s i m i l a r l n t e r c o n n e c t e d vessels. The second gradrent was l i n e a r and CDmpoIed o f e q u a l v o l m r (50 " 1 ) of 0.5 n p y r l d l n e a c e t a t e pH 3 . 8 and 2M p y r i d i n e a c e t a t e RI 5 0. The f low Fate was approx imate ly 6ml /h and f ract ions o f 1 . 5 ml were c o l l e c t e d . Frdctlona were m n l t o r e d f o r r a d i o a c t i v i t y b y t a k i n g a l l q u o t s , u s u a l l y 50 y l . f o r s c l n t l l l a t l o n c o u n t i n g .
a - lac ta lbumin . A l l there pept ldes had r values that were c l o s e l y s i m l l w IS CN1 (F ig . 21 con ta ins aggregated CN2 or D p r o p o r t i o n o f ""cleaved
t o those pept ides Obta ined frm the d lges t O f CN2, and are included i n Table 11 (main paper). NO C N I pep t ides were i s o l a t e d frm the CNZ peak
F m m the r a m ~ l e O f m - I a c t a l b u m n t h a t had been m d i f i e d i n the
C N 2 1
FRACTION NO.
e -
e -
a t
L - 0 0
FRACTION NO.
Flg. 3 . Seporatlon Of t h e Products of trypsln dlgeation Of CN1 with a column of Bio-Gel P6 11.9 x 80 mi. Fraction B L Z ~ was 2 .75 m 1 and 50 ul was sampled for scintillation c o u n t m g .
Fig. 1. Separatlon Of the prcducts of thernolysin digestlon Of CN1 T2 by ion exchange ChrOmatOgraphy with M i n e l A-5. FOE this Bepacotion a linear gradient from 30\ acetic acid to 0.5 n pyridine acetate was used. The arrow marks the start Of the gradient. Fractions of 1.5 01 were collected and aliquots of 50 u1 were taken for trltium determination by Scintillation counting.
22 c 4
7 P
B
x
E
FRACTION NO.
Fig. 5. Chromatographic separation of chymotryptic digestion Of CN1 T1 with Aminex A-5. The conditiam are given in the *Experimental Procedures' BeCtlOn. The plot s h w s the radioactivity found in fractione collected durina elutlon with the firllt oradlent.

Interaction Site of a-Lactalbumin for Galactosyltransferase 3385
FRACTION NO
1 1M1
FRACTION NO.
f l q . 7. Separatlon of the products of thermolysln d1qestLon of CN2 vlth a column of Amlnex A - 5 .
2.5 -
2 0 -
1.5 .
20 4 0 80 BO
FRACTION NO.
Flg. 8 . Separatlon of the peptides I n CN2 Th 1 from flg. 7 by r e v e r s e phase partltlon Chromatography. See “Experlmental Pcocedures‘ for details.








![Detection of Pancreatic Carcinomas by Imaging Lactose-Binding Protein Expression in Peritumoral Pancreas Using [18F]Fluoroethyl-Deoxylactose PET/CT](https://static.fdokumen.com/doc/165x107/631be6bc93f371de19012dfd/detection-of-pancreatic-carcinomas-by-imaging-lactose-binding-protein-expression.jpg)











