In vitro galactosylation of human IgG at 1 kg scale using recombinant galactosyltransferase
12
In Vitro Galactosylation of Human IgG at 1 kg Scale Using Recombinant Galactosyltransferase Dale Warnock, Xiaomei Bai, Katie Autote, Johnny Gonzales, Kyle Kinealy, Boxu Yan, Jun Qian, Tom Stevenson, David Zopf, Robert J. Bayer Neose Technologies, Inc., 102 Witmer Road, Horsham, Pennsylvania 19044; telephone: 858-552-2700, fax: 858-452-1009; e-mail: [email protected] Received 12 January 2005; accepted 20 June 2005 Published online 26 September 2005 in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/bit.20658 Abstract: The Fc effector functions of immunoglobulin G (IgG) antibodies are in part determined by structural features of carbohydrates linked to each of the paired gamma heavy chains in the antibody constant domain (C H 2). One glycoform that has been shown to be advantageous is G2, where both arms of complex bi- antennary N-glycans terminate in galactose. In vitro treatment with glycosyltransferases can remodel hetero- geneous IgG glycoforms, enabling preparation of IgG molecules with homogeneous glycan chains. Here we describe optimization of conditions for use of a soluble recombinant galactosyltransferase in vitro to remodel glycans of human serum IgG, and we demonstrate a scaled-up reaction in which >98% of neutral glycans attached to 1 kg IgG are converted to the G2 glycoform. Removal of glycosylation reagents from the product is achieved in one step by affinity chromatography on immobilized Protein A. ß 2005 Wiley Periodicals, Inc. Keywords: glycoengineering; glycoprotein remodeling; glycosylation; glycosyltransferase; antibody; antibody effector function INTRODUCTION Antibody based immunotherapy is a rapidly developing segment of pharmaceutical drug discovery efforts. Several antibody based treatments for various cancers (Clynes et al., 2000; White et al., 2001) and inflammatory diseases such as rheumatoid arthritis (Maini et al., 1999; Rastetter et al., 2004; Sandborn and Hanauer, 1999) are currently approved and many more are currently in clinical and pre-clinical development (Presta, 2002; Souriau and Hudson, 2003). Whereas early efforts to develop therapeutic monoclonal antibodies (mAbs) focused primarily upon maximizing specificity of antigen binding and minimizing immunogeni- city of the immunoglobulin G (IgG) protein, more recent engineering of IgG mAbs has expanded to include custo- mized up- or down-regulation of Fc effector functions (Shriver et al., 2004). The glycans most commonly associated with human serum IgG (Fujii et al., 1990; Hamako et al., 1993; Raju et al., 2000) or therapeutic mAbs (Jefferis, 2001, 2005) are complex bi-antennary chains N-linked to Asn-297 of the g- h-chain. These branched sugar chains are situated within a cleft formed by the paired heavy chains in the C H 2 domain such that they may undergo extensive non-covalent interac- tions with the adjacent polypeptide (Krapp et al., 2003). Pharmacologic properties that may be affected by modifica- tions in IgG glycans include: (i) plasma residence time (ii) rate of antigen clearance from the plasma, (iii) potential for activation of the complement pathway, (iv) efficiency in promoting cell killing via antibody-dependent cellular cytotoxicity (ADCC), and (v) capacity to trigger inflamma- tory responses (Dijstelbloem et al., 2001; Jefferis and Lund, 2002; Krapp et al., 2003). The precise structural basis for glycan-related modulation of antibody efficacy in vivo remains to be fully elucidated, but it has become clear that binding to FcgRII and FcgRIII receptors requires the N- linked glycan at Asp-297 in the C H 2 domain (Coloma et al., 2000; Leatherbarrow et al., 1985; Walker et al., 1989), and that while the minimal trimannosyl-chitobiose core structure (see Fig. 1) is sufficient to allow FcgRIIb binding (Mimura et al., 2001), more extended glycan structures exert effects on the Fc tertiary structure that could serve to modulate specific FcgR–IgG interactions (Jefferis et al., 1998; Mimura et al., 2001). The key mechanism by which glycans may affect antibody binding to Fc receptors appears to be through stabilization of a polypeptide conformation of the Fc domain with increased affinity for Fc receptors (Lund et al., 1995; Radaev and Sun, 2001; Wormald et al., 1997). The presence or absence of galactose on IgG glycans correlates with modified Fc effector function in some (Kumpel et al., 1994) but not all (Boyd et al., 1995; Lund et al., 1995; Wright and Morrison, 1998) monoclonal IgG antibodies, suggesting that the effects observed may be, in ß 2005 Wiley Periodicals, Inc. Correspondence to: R. J. Bayer
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of In vitro galactosylation of human IgG at 1 kg scale using recombinant galactosyltransferase

In Vitro Galactosylation of Human IgGat 1 kg Scale Using RecombinantGalactosyltransferase
Dale Warnock, Xiaomei Bai, Katie Autote, Johnny Gonzales, Kyle Kinealy,Boxu Yan, Jun Qian, Tom Stevenson, David Zopf, Robert J. Bayer
Neose Technologies, Inc., 102 Witmer Road, Horsham, Pennsylvania 19044;telephone: 858-552-2700, fax: 858-452-1009; e-mail: [email protected]
Received 12 January 2005; accepted 20 June 2005
Published online 26 September 2005 in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/bit.20658
Abstract: The Fc effector functions of immunoglobulinG (IgG) antibodies are in part determined by structuralfeatures of carbohydrates linked to each of the pairedgamma heavy chains in the antibody constant domain(CH2). One glycoform that has been shown to beadvantageous is G2, where both arms of complex bi-antennary N-glycans terminate in galactose. In vitrotreatment with glycosyltransferases can remodel hetero-geneous IgG glycoforms, enabling preparation of IgGmolecules with homogeneous glycan chains. Here wedescribe optimization of conditions for use of a solublerecombinant galactosyltransferase in vitro to remodelglycans of human serum IgG, and we demonstrate ascaled-up reaction in which >98% of neutral glycansattached to 1 kg IgG are converted to the G2 glycoform.Removal of glycosylation reagents from the product isachieved in one step by affinity chromatography onimmobilized Protein A. � 2005 Wiley Periodicals, Inc.
Keywords: glycoengineering; glycoprotein remodeling;glycosylation; glycosyltransferase; antibody; antibodyeffector function
INTRODUCTION
Antibody based immunotherapy is a rapidly developing
segment of pharmaceutical drug discovery efforts. Several
antibody based treatments for various cancers (Clynes et al.,
2000; White et al., 2001) and inflammatory diseases such as
rheumatoid arthritis (Maini et al., 1999; Rastetter et al., 2004;
Sandborn and Hanauer, 1999) are currently approved and
many more are currently in clinical and pre-clinical
development (Presta, 2002; Souriau and Hudson, 2003).
Whereas early efforts to develop therapeutic monoclonal
antibodies (mAbs) focused primarily upon maximizing
specificity of antigen binding and minimizing immunogeni-
city of the immunoglobulin G (IgG) protein, more recent
engineering of IgG mAbs has expanded to include custo-
mized up- or down-regulation of Fc effector functions
(Shriver et al., 2004).
The glycans most commonly associated with human
serum IgG (Fujii et al., 1990; Hamako et al., 1993; Raju et al.,
2000) or therapeutic mAbs (Jefferis, 2001, 2005) are
complex bi-antennary chains N-linked to Asn-297 of the g-h-chain. These branched sugar chains are situated within a
cleft formed by the paired heavy chains in the CH2 domain
such that they may undergo extensive non-covalent interac-
tions with the adjacent polypeptide (Krapp et al., 2003).
Pharmacologic properties that may be affected by modifica-
tions in IgG glycans include: (i) plasma residence time (ii)
rate of antigen clearance from the plasma, (iii) potential for
activation of the complement pathway, (iv) efficiency in
promoting cell killing via antibody-dependent cellular
cytotoxicity (ADCC), and (v) capacity to trigger inflamma-
tory responses (Dijstelbloem et al., 2001; Jefferis and Lund,
2002; Krapp et al., 2003). The precise structural basis for
glycan-related modulation of antibody efficacy in vivo
remains to be fully elucidated, but it has become clear that
binding to FcgRII and FcgRIII receptors requires the N-
linked glycan at Asp-297 in the CH2 domain (Coloma et al.,
2000; Leatherbarrow et al., 1985; Walker et al., 1989), and
that while the minimal trimannosyl-chitobiose core structure
(see Fig. 1) is sufficient to allow FcgRIIb binding (Mimura
et al., 2001), more extended glycan structures exert effects on
the Fc tertiary structure that could serve to modulate specific
FcgR–IgG interactions (Jefferis et al., 1998; Mimura et al.,
2001). The key mechanism by which glycans may affect
antibody binding to Fc receptors appears to be through
stabilization of a polypeptide conformation of the Fc domain
with increased affinity for Fc receptors (Lund et al., 1995;
Radaev and Sun, 2001; Wormald et al., 1997).
The presence or absence of galactose on IgG glycans
correlates with modified Fc effector function in some
(Kumpel et al., 1994) but not all (Boyd et al., 1995; Lund
et al., 1995; Wright and Morrison, 1998) monoclonal IgG
antibodies, suggesting that the effects observed may be, in
�2005 Wiley Periodicals, Inc.
Correspondence to: R. J. Bayer

part, antibody specific. Several investigators have assessed
Fc effector functions in vitro after enzymatic removal of
galactose from IgG with b-galactosidase. Tsuchiya et al.
(1989) found that agalacto IgG had reduced C1q and Fc
receptor binding, and Boyd et al. (1995) found that agalacto
Campath1 (monoclonal anti-CD52) had reduced cell-
mediated lysis (CML) but intact capacity to trigger ADCC.
Other reported effects of galactosylation on IgG molecules
include modification of physicochemical properties such as
conformation and surface accessibility (Dwek et al., 1995;
Ghirlando et al., 1999; Krapp et al., 2003; Mimura et al.,
2000).
It would be desirable to convert IgG molecules to a single
(>90%) defined glycoform, both to develop reagents for
studying the impact of glycosylation onFc effector functions,
and to prepare mAb drugs with a desired glycoform at scale.
Fujii et al. (1990) previously demonstrated at analytical scale
that the glycans on intact IgG molecules can function as
acceptors for glycosyltransferases. In this article,we describe
conditions for use of a recombinant glycosyltransferase to
remodel IgG glycans and demonstrate in vitro enzymatic
galactosylation of human IgG at the 1 kg scale, resulting in
nearly complete (>98%) conversion to the G2 glycoform
(Fig. 1).
MATERIALS AND METHODS
Enzymes, Antibodies, and Reagents
A gene construct encoding a truncated form of bovine b-1,4galactosyltransferase-I (NCBI protein no. AAM54035)
(GalT-1) was cloned into a recombinant expression vector
that allows high level expression and secretion from
mammalian derived cell lines (Narimatsu et al., 1986). The
gene product was overexpressed in the mouse myeloma NS0
cell line and recovered as a secreted product from NS0 cell
culture (971 U/L) carried out for 432 h in a 40 L bioreactor.
Clarified and concentrated culture supernatant was buffer
exchanged into 50 mM NaCl, 20 mM 2-(N-morpholi-
no)ethane-sulfonic acid (MES), pH 6.0, then loaded onto a
column of SP sepharose (18� 17 cm). The enzyme was
eluted with 1 M NaCl, 20 mM MES, pH 6.0, concentrated,
and buffer exchanged into 150mMNaCl, 50mMHEPES, pH
7.5 on an Amicon S10Y10 cartridge filter (Millipore Corpo-
ration, Bedford, MA), and sterile filtered. The enzymatic
activity of the purified GalT-1 was assayed by transfer of
[3H]-galactose (0.63 mM uridine 50-diphosphate (UDP)-
galactose) to N-acetylglucosamine (20 mM) in 50 mM
3-(N-morpholino)propanesulfonic acid (MOPS) (pH 7.4),
Figure 1. Typical oligosaccharide structures found in the antibody fragment that crystallizes (Fc) domain of IgG. The sugars highlighted in orange boxes are
variably present, and account for much of the variability observed in Fc glycans.A: Completely galactosylated and sialylated complex bi-antennary structure,
including ‘‘bisecting’’ N-acetyl glucosamine (GlcNAc) linked b1–4 to mannose, and ‘‘core’’ fucose linked a1–6 to the asparagine-linked GlcNAc; (B) G2oligosaccharides havegalactosemoieties onboth antennae; (C)G0 indicates that no galactose is present; (D,E)G1 structures havegalactose on either the 1–6or1–3 arm.
832 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 92, NO. 7, DECEMBER 30, 2005

30 mM MnCl2, 0.2 mg/mL bovine serum albumin (BSA),
during a 3 min incubation at 378C. The radioactive productwas isolated through a 1 mL column of Dowex AG1X8
(Biorad, Hercules, CA) pre-washed in water, and quantified
by scintillation counting. The purity of the GalT-1 prepara-
tion was measured by scanning and integration of an sodium
dodecyl sulfate–polyacrylamide gel electrophoresis (SDS–
PAGE) gel stained with fluorescent SYPRO Tangerine
Protein Gel Stain (Ex/Em 300, 490/640 nm) fromMolecular
Probes according to the manufacturer’s instructions. A
digital image was acquired taking care not to saturate any
of the bands on the gel using a FACE Imaging System from
Glyko (300 nm transilluminator). The image was saved in
TIFF format, and the relative band densities were integrated
by using software (Scion Image for Windows, a modified
version of NIH Image).
Polyclonal antisera raised against bovine GalT-1 was
produced in rabbits. Donkey anti-rabbit horseradish perox-
idase (HRP) conjugate and West-Pico chemiluminescent
reagent were obtained from Pierce (Rockford, IL). All SDS–
PAGE gels and buffers were from Invitrogen (Carlsbad, CA).
UDP-[3H]-galactosewas a product of NewEngland Nuclear/
PerkinElmer (Boston, MA) and UDP-galactose was a
product of Boehringer Mannheim (Indianapolis, IN).
Human gamma globulin was purchased from Serologicals
Corporation (Norcross, GA). Unless otherwise specified, all
other reagent grade chemicals were purchased from Sigma
(St. Louis, MO). Chromatography resins were obtained from
Amersham Biosciences/GE Healthcare (Piscataway, NJ)
unless otherwise noted.
Remodeling Reaction Conditions
Small scale test reactions were carried out in 1.5 mL
microfuge tubes containing 50 mL reaction buffer: 20 mM
MnCl2, 10 mM UDP-galactose, 100 mM MES, pH 6.5. The
concentration of human serum IgG was 20 mg/mL unless
otherwise specified. GalT-1 activity added varied from 10 to
100 mU/mg IgG and reactions were incubated for up to 72 h
at 328C.Large scale remodelingwas performed in a final volume of
33.5 L in a 40 L, temperature controlled, stirred bioreactor
vessel (Wheaton, Inc., Millville, NJ) equipped with pH and
temperature monitors. One kilogram of lyophilized human
IgG was dissolved in 25 L buffer containing 0.1 M MES, 20
mMMnCl2, and 0.02% sodiumazide, pH6.5.UDP-galactose
was added to a concentration of 10 mM, and GalT-1 was
added to a final activity ratio of 25 mU/mg IgG. The reaction
mixture was brought to a volume of 33.5 L (30 mg/mL final
concentration of IgG) and carefully re-adjusted to pH 6.5.
Samples were removed at appropriate time points during the
reaction and stored frozen at �208C until analyzed.
rProtein A-Sepharose Fast Flow (FF) AffinityChromatography
Human IgG (30 mg/mL) remodeled with GalT-1 (48 h
reaction time) was thawed and filtered using a 0.45 mm
Acrodisc HT Tuffryn Syringe filter obtained from Pall
Corporation (AnnArbor,MI). One hundredmilligrams of the
sample was injected onto a 5 mL column of rProtein A-
Sepharose (XK16/60 column on an Akta fast protein liquid
chromatography (FPLC) system, Amersham Biosciences/
GE Healthcare) via a 5 mL sample loop. The loading and
washing buffer was 20 mM Tris, 150 mMNaCl, pH 7.2. The
elution buffer was 0.2 M glycine-HCl, pH 3.0. One-milliliter
fractions were collected and immediately neutralized with
0.1 mL 1 M Tris, pH 9.0.
Sample Preparation for Oligosaccharide Analysis
Twenty-five microliter aliquots of remodeling reaction
samples were diluted to 100 mL with phosphate buffered
saline (PBS), and then centrifuged in a microfuge at 14,000g
for 5 min to remove precipitated manganese phosphate.
Supernatants were applied to protein desalting spin columns
(Pierce) pre-equilibrated with 20 mM sodium phosphate
buffer, pH 7.0, and eluted by centrifugation at 2,000g. Eluted
samples were assayed for protein concentration, diluted to 1
mg/mL, and prepared for 2-anthranilic acid (2-AA) glycan
analysis (Anumula and Dhume, 1998). Glycan release from
100 mm of protein was achieved by incubation overnight at
378C in 50 mM sodium phosphate pH 7.2 containing 0.2%
SDS, 1% NP-40, 0.1 M b-mercaptoethanol, and 50 mU
peptide N-glycosidase F (PNGase F, Calbiochem, La Jolla,
CA). Enzyme reactions were stopped by the addition of 0.6
mL ice-cold ethanol and the resulting mixtures were
incubated on ice for 10 min, centrifuged at 14,000g for 5
min, and concentrated to dryness in a vacuum centrifuge.
Released glycans were labeled with 2-amino anthranilic acid
with a LudgerTag 2-AA glycan labeling kit (Ludger Ltd.,
Oxford, UK) according to the manufacturer’s instructions.
The 2-AA labeled glycanswere desalted on 13mmAP10MF
support pads (Millipore) to remove salts and labeling
reagents, eluted with dH2O, and lyophilized.
HPLC Analysis of 2-AA Labeled Oligosaccharides
Immediately prior to high performance liquid chromatogra-
phy (HPLC) analysis, lyophilized 2-AA labeled glycanswere
re-dissolved at approximately 0.4 mg/mL in dH2O. HPLC
analysis was performed on a Shodex Asahipak NH2P-50 4D
amino column (4.6� 150 mm, Shodex Inc., Tokyo, Japan) at
a flow rate of 0.8 mL/min and a column temperature of 408C.To analyze for neutral glycans, typically a 10 mg sample was
injected in 25 mL and eluted isocratically for 5 min with 70%
mobile phase A (2% acetic acid, 1% tetrahydrofuran in
acetonitrile) and 30% mobile phase B (5% acetic acid, 1%
tetrahydrofuran, 3% triethanolamine in water). Next the
column was eluted with a linear gradient to 50% mobile
phaseAover 60min, 50%mobile phaseA isocratically for 10
min, then a linear gradient to 5% mobile phase A for 10 min,
then re-equilibrated to 70% mobile phase A for 15 min prior
to injection of the next sample. To analyze charged
oligosaccharides, the column was eluted isocratically with
70% mobile phase A for 2.5 min, followed by a linear
WARNOCK ET AL.: GALACTOSYLATION OF HUMAN IGG 833

gradient to 5% mobile phase A for 97.5 min, and 5% mobile
phase A isocratically for 15 min, followed by column re-
equilibration to 70% mobile phase A for 15 min before
injecting the next sample. Eluted glycans were monitored by
fluorescence with excitation and emission wavelengths of
230 and 420 nm, respectively. Peak elution positions were
compared with 2-AA labeled oligosaccharide standards
purchased from Glyko/Prozyme (San Leandro, CA).
MALDI-TOF analysis was performed on the 2AA-labeled
glycan samples after mixing (1:1) with a 2,5-dihydroxyben-
zoic acid solution (10 mg/mL), and then spotting and drying
on a target. An Applied Biosystems DE-Pro MALDI-TOF
mass spectrometer (Foster City, CA) was operated in linear/
negative-ion mode to analyze the samples. Glycan structures
were assigned based on the observed mass-to-charge ratios
and literature precedence (Fujii et al., 1990; Raju et al., 2000).
LC/MS/MS Analysis of SDS–PAGE FractionatedProteins
Coomassie blue stained gel bands were cut out with a clean
razor blade and placed in low-binding 0.5 mL tubes (CLP,
San Diego, CA). The gel pieces were destained by repeated
extraction with 50% acetonitrile containing 50 mM ammo-
nium carbonate for 30 min at 378C, evaporated to dryness byvacuum centrifugation, and rehydrated for 10 min in 10 mL5% acetonitrile, 50 mM ammonium carbonate, 0.1 mg/mL
modified trypsin (Promega, Madison WI). The volume was
then brought to 75 mL with 5% acetonitrile, 50 mM
ammonium carbonate, and the sample incubated at 378Covernight.Acetic acid (10mLof a 10% solution)was added to
stop the digests, and then samples were reduced to a volume
of approximately 10 mL by vacuum centrifugation.
Liquid chromatography electrospray ionization mass
spectrometry (LC/ESI/MS) analysis was performed using a
Haisil 300 reversed-phase C18 column (0.3� 150 mm,
Higgins Analytical, Inc., Mountain View, CA) on an Agilent
1100 system (Palo Alto, CA) coupled with an LCQ Deca XP
MAX (ThermoElectron, Waltham, MA). The column was
eluted with 98%mobile phase A (0.1% formic acid in water)
for 5 min, followed by a linear gradient to 40%mobile phase
B (0.1% formic acid in acetonitrile) over 65min at a flow rate
of 8 mL/min. After each full MS scan from m/z 400 to m/z
2,000 of the column effluent, MS/MS spectra of the twomost
intense MS peaks were recorded. Chromatograms were
analyzed against the April 2004 NCBI human or bovine non-
redundant protein databases with the program SEQUEST
using Bioworks 3.1 (ThermoElectron). SEQUEST cross-
correlation (Xcorr) values and Dcorr values were used to filter
all peptide matches. Acceptable matches had Dcorr values
�0.1 and Xcorr values �1.7 (þ1 ions) or �2.0 (þ2 ions) or
�3.0 (þ3 ions) (Link et al., 1999).
HPLC Analysis of Nucleotides and NucleotideSugars
Ten-microliter samples containingUDP-galactose, UDP, and
uridine 50-monophosphate (UMP) were analyzed by HPLC
on a Vydac 302IC4.6 Ion-Chromatography Column (4.6�250mm,GraceVydac,Hesperia, CA) at a flow rate of 2.0mL/
min and maintained at 288C. The column was eluted as
follows: isocratic elution with 100%mobile phase A (25mM
NaH2PO4/Na2HPO4, 1:1 molar ratio, pH 2.8) for 2 min,
followed by elution with a linear gradient to 100% mobile
phase B (125 mM NaH2PO4/Na2HPO4, 1:1 molar ratio,
adjusted to pH 2.9 with acetic acid) over a period of 17 min.
Eluted nucleotides were detected by absorption at 260 nm.
The method was standardized using a sample matrix
containing bovine gamma globulin at 30 mg/mL and shown
to give accurate and reproducible results under these
conditions.
Protein Determination by Absorbance and BioRadDC Protein Assay
Fractions from rProtein A Sepharose FF affinity chromato-
graphy were diluted into UV clear bottom 96-well plates and
the absorbance read at 280 nm on a Molecular Devices
Spectramax 384 Plus microplate reader (Sunnyvale, CA).
Affinity chromatography fractions and remodeled samples
were quantified with BioRad DC protein determination
reagents in 96-well plate assay format according to the
manufacturer’s instructions. A standard curve of BSA was
included on each plate, and the absorbance was measured at
750 nm.
SDS–PAGE and Western Analysis
Protein fractions from remodeling reactions and rProtein A-
Sepharose FF affinity chromatography were resolved by
SDS–PAGE on 4%–12% NuPAGE gels in MOPS or MES
running buffer. Protein patterns were visualized with Simply
Blue colloidal Coomassie stain (Invitrogen) according to the
manufacturer’s instructions.
For Western blot analysis, proteins were transferred to
0.4 mm nitrocellulose paper using the Novex transfer
blotting module (Invitrogen). Transfer uniformity was
verified by the transfer efficiency of SeeBlue Plus2
prestained markers (Invitrogen). Blots were blocked in
TBS-T20 (20 mM Tris, pH 7.2, 150 mM NaCl, 0.05%
Tween-20) with 5% dry milk for 30 min. The blots were then
placed in TBS-T20 with 5% dry milk containing rabbit anti-
galactosyltransferase anti-sera and incubated at 48C over-
night with rocking. Blots were washed four times for 5 min
with TBS-T20 then incubated for 1 h at room temp with
gentle shaking in a 1:20,000 dilution of donkey anti-rabbit
HRP-conjugated antibody in TBS-SuperBlock (Pierce).
Blots were washed five times for 5 min with TBS-T20 before
incubation with West Pico Chemiluminescent reagent
(Pierce), as per the kit instructions. Blots were exposed on
Bio Max film (Kodak, Rochester, NY). After scanning the
film on a flatbed scanner, uniform reduction of background
was applied to electronic images with Photoshop1 software
(Adobe, San Jose, CA).
834 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 92, NO. 7, DECEMBER 30, 2005

RESULTS
Galactosyltransferase Purification
In total, 32,700Uof purifiedGalT-1with a specific activity of
6 U/mg was recovered in 3.5 L (84% overall yield). The
purity of the final preparation determined by scanning and
integrating an SDS–PAGE gel stained with Sypro Tangerine
was 60%. Sixteen percent of the preparation was serum
albumin, with the balance (24%) being accounted for by a
number of low-abundance bands on the gel.
In Vitro Remodeling of Human IgG Glycans WithRecombinant GalT-1 Enzyme
In order to demonstrate the practicality and scalability of in
vitro enzymatic galactosylation, we undertook the large scale
enzymatic remodeling of the glycans on IgG isolated from
human serum using a recombinant form of the bovine
galactosyltransterase GalT-1. The N-glycans of human IgG
(Fujii et al., 1990; Kornfeld et al., 1971) consist primarily of
neutral bi-antennary complex oligosaccharides. The neutral
bi-antennary structures with zero, one or two terminal b-1,4linked galactosyl residues (Fig. 1) are termed, respectively,
G0, G1, and G2 glycans (Wormald et al., 1997).
The N-glycan profile for commercial pooled human serum
IgG used for the galactosylation experiments reported here
was measured by reversed phase-liquid chromatography
(RP-HPLC) of 2-AA-derivatized glycans released by
PNGase F (Fig. 2). Determinations of total N-glycans species
[mean (�SD) from five separate analyses] revealed that
75.1% (�6.5%) were neutral species, whereas 17.2%
(�3.1%) were singly charged and 7.7% (�3.4%) doubly
charged. The distribution of neutral glycan species calculated
from six separate HPLC glycan analyses of the starting IgG
stock was as follows: G0, 33.2% (�1.8%); G1, 46.7%
(�0.6%); G2, 20.1% (�2.3%).
Incubation of human serum IgG (25 mg/mL) with GalT-1
converted the structurally heterogeneous neutral glycans to
>98%G2 structureswithin 48 h (Fig. 2, Table I). The enzyme
is known to selectively catalyze addition of galactose inb1–4linkage to terminal N-acetyl glucosamine (GlcNAc) on
complexN-linked oligosaccharides, utilizing the high energy
donor UDP-galactose (Khatra et al., 1974). UDP is released
during the reaction. By varying the enzyme/substrate activity
ratio and time of incubation, we determined that conversion
of G0 to G1 occurs rapidly, even at the lowest activity ratio of
GalT-1 tested (10 mU/mg IgG), but conversion of G1 to G2
occurs more slowly, reaching levels¼ 98% after 48 h at
activity ratios of¼ 25 mU GalT-1/mg IgG. Conversion to
100% G2 was achieved only after prolonged incubation (72
h) at a much higher activity ratio (75 mU/mg).
Optimization of Substrate Concentration
To further optimize conditions for large scale IgG glycan
remodeling we carried out galactosylation at increasing
concentrations of IgG. In preliminary experiments, we
observed less than 80% G2 associated with IgG incubated
at 1 mg/mL for 48 h with 50 mU GalT-1/mg IgG. Increasing
the concentration of IgG improved the rate of conversion to
G2 even after decreasing the activity ratio to 25 mU GalT-1/
mg IgG: at 20 mg/mL IgG, neutral glycans were converted to
93%G2 after 24 h (Fig. 2, Table II); at 30mg/mL IgG, 97%of
neutral glycans were converted to G2 after 24 h. At IgG
concentrations¼ 30 mg/mL, conversion to G2 is approxi-
mately 99% in 48 h (Table II). The activity of GalT-1
incubated at 328C for 48 h in the same buffer used for the
remodeling reaction remained constant (data not shown).
Large Scale Remodeling and Glycan AnalysisUsing the Optimized Reaction Conditions
Based on results from smaller scale test reactions (Fig. 2, and
Tables I and II), we chose to remodel 1 kg IgG in a reaction
volume of 33.5 L containing 30 mg/mL IgG, 25 mU GalT-1
Figure 2. Reversed phase-high performance liquid chromatography (RP-
HPLC) analysis of 2-anthranilic acid (2-AA) derivatized glycans from
human IgGbefore and after conversionwith galactosyltransferase-I (GalT-1)
to the G2 glycoform. Human IgG was reacted with GalT-1 and UDP-Gal for
(A) 0 h, (B) 24 h, and (C) 48 h. Glycans eluting at approximately 59 min
terminate in sialic acid on one branch (�1 charge) while those eluting at
approximately 78min terminate in sialic acid on both branches (�2 charges).
The starred peak represents an artifact of glycan preparation.
WARNOCK ET AL.: GALACTOSYLATION OF HUMAN IGG 835

per milligram IgG, and 10 mM UDP-galactose (final
concentrations). The reaction was carried out for 48 h in a
continuously monitored 40 L bioreactor at 328C, pH 6.5 with
constant stirring. Glycan analysis of aliquots at intervals
(Figs. 3 and 4 showed complete conversion of G0 structures
to G1 within 4 h and nearly complete (98.5%) conversion to
G2 structures after 48 h.
Protein A-Sepharose ChromatographyFractionation of Remodeled IgG
To demonstrate removal of remodeling reagents from the
large scale reaction mixture, an aliquot removed at 48 h was
applied to a column of rProtein A-Sepharose and the column
washed with Tris buffer at neutral pH. Most of the human
serum IgG molecules bound to the affinity matrix, whereas
GalT-1 and UDP-galactose and its breakdown products
passed through the column (Fig. 5A). SDS–PAGE analysis
of fractions eluted at neutral pH and fractions eluted at pH 3
(Fig. 5B) showed that neutral flow-through fractions
contained only a trace of IgG plus other protein contaminants
from the original serum IgGpreparation. Themost prominent
contaminating protein bands visualized on SDS–PAGE
gels (marked in Fig. 5B) were excised, digested with trypsin,
and identified by LC/MS/MS as (a) recombinant GalT-1
(approximately 40 kDa), (b) heavy chains of human IgG3
and IgA, immunoglobulins with weak affinity for Protein A
(approximately 55 kDa), (c) human serum albumin
Table I. Impact of galactosyltransferase-I (GalT-1)/IgG ratio on percent G0, G1, and G2 glycans of human
serum IgG after various periods of time.
Time (h) Glycoform
Percent of neutral glycansb
Control
Activity ratio (mU GalT-1/mg IgG)a
10 17 25 50 75
24 G0 33.2 (�1.8) — — — — —
G1 46.7 (�0.6) 28.7 11.9 6.9 2.4 1.2
G2 20.1 (�2.3) 71.3 88.1 93.1 97.6 98.8
48 G0 — — — — —
G1 12.7 4.1 2.0 0.6 0.8
G2 87.3 95.9 98.0 99.4 99.2
72 G0 — — — — —
G1 7.3 2.3 1.7 0.6 —
G2 92.7 97.7 98.3 99.4 100
aReactions were carried out at a concentration of human serum IgG of 20 mg/mL plus GalT-1 at the indicatedactivity ratio.
bPercentages of glycans before (control) and after galactosylation with GalT-1, determined by integration ofRP-HPLC chromatograms of 2-AA labeled glycans analyzed under conditions to resolve neutral glycan structures(see Materials and Methods). Control values are reported as mean (�SD) calculated from six separate analyses.
—, indicates the structure was not detected.
Table II. Impact of increasing IgG concentrations on percent G0, G1,
and G2 glycans after galactosylation by GalT-1 for 24 and 48 h.
Time (h)
Percent of total N-glycansa
Glycoform
IgG concentration (mg/mL)b
Control 20 25 30 35
24 G0 33.2 (�1.8)a — — — —
G1 46.7 (�0.6) 7.2 5.1 3.1 3.5
G2 20.1 (�2.3) 92.8 94.9 96.9 96.5
48 G0 — — — —
G1 2.6 1.4 0.8 1.8
G2 97.4 98.6 99.2 98.7
aPercentages of glycans before (control) and after galactosylation withGalT-1 were determined by integration of RP-HPLC chromatograms of 2-AA labeled glycans analyzed under conditions to resolve neutral glycanstructures (see Materials andMethods). Control values are reported as mean(�SD) calculated from six separate analyses.
bReactionswere carriedout at the indicated concentrations of humanserum IgG in the presence of 25 mU GalT-1/mg IgG (see Materials andMethods).
—, indicates the structure was not detected.
Figure 3. Time course for galactosylation of the neutral glycans of human
serum IgG by recombinant GalT-1 at the kilogram scale. Aliquots were
removed from the bioreactor vessel at the indicated time points and N-
glycans released by peptide N-glycosidase F (PNGase F) were analyzed by
RP-HPLC as 2-AA derivatives (see Materials and Methods).
836 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 92, NO. 7, DECEMBER 30, 2005
(approximately 65 kDa) and (d) a-2 macroglobulin (approxi-
mately 160 kDa) (data not shown).
Western blot analysis with anti-GalT-1 comparing the
enzyme reaction mixture before and after Protein A
chromatography demonstrated removal of at least 90% of
GalT-1 (Fig. 5C) and other non-IgG contaminants (Fig. 5B)
during this single step affinity purification. Analysis of the
same fractions by ion-exchange chromatography demon-
strated a 310-fold reduction in concentration of total
nucleotides. The concentration of UDP-galactose, UDP,
and UMP, and uridine was 9.31 mM in the remodeling
reaction, 10.5 mM in the Protein A Sepharose flow-through,
and 0.03 mM in the Protein A Sepharose low pH eluate. The
primary species present after the remodeling reaction was
uridine, presumably due to the presence of a phosphatase in
the polyclonal antibody preparation, which may have aided
in relieving any possible inhibition by UDP, a known
inhibitor of the galactosyltransferase.
Glycan Analysis of Remodeled IgG
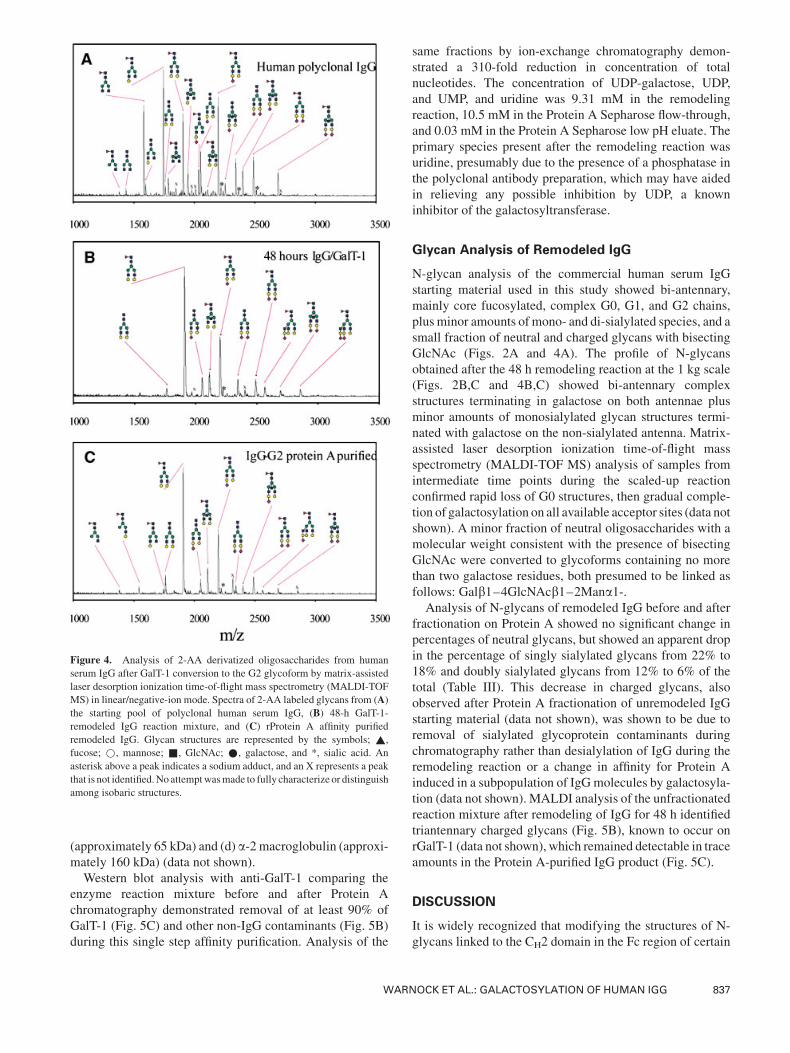
N-glycan analysis of the commercial human serum IgG
starting material used in this study showed bi-antennary,
mainly core fucosylated, complex G0, G1, and G2 chains,
plus minor amounts of mono- and di-sialylated species, and a
small fraction of neutral and charged glycans with bisecting
GlcNAc (Figs. 2A and 4A). The profile of N-glycans
obtained after the 48 h remodeling reaction at the 1 kg scale
(Figs. 2B,C and 4B,C) showed bi-antennary complex
structures terminating in galactose on both antennae plus
minor amounts of monosialylated glycan structures termi-
nated with galactose on the non-sialylated antenna. Matrix-
assisted laser desorption ionization time-of-flight mass
spectrometry (MALDI-TOF MS) analysis of samples from
intermediate time points during the scaled-up reaction
confirmed rapid loss of G0 structures, then gradual comple-
tion of galactosylation on all available acceptor sites (data not
shown). A minor fraction of neutral oligosaccharides with a
molecular weight consistent with the presence of bisecting
GlcNAc were converted to glycoforms containing no more
than two galactose residues, both presumed to be linked as
follows: Galb1–4GlcNAcb1–2Mana1-.Analysis of N-glycans of remodeled IgG before and after
fractionation on Protein A showed no significant change in
percentages of neutral glycans, but showed an apparent drop
in the percentage of singly sialylated glycans from 22% to
18% and doubly sialylated glycans from 12% to 6% of the
total (Table III). This decrease in charged glycans, also
observed after Protein A fractionation of unremodeled IgG
starting material (data not shown), was shown to be due to
removal of sialylated glycoprotein contaminants during
chromatography rather than desialylation of IgG during the
remodeling reaction or a change in affinity for Protein A
induced in a subpopulation of IgGmolecules by galactosyla-
tion (data not shown). MALDI analysis of the unfractionated
reaction mixture after remodeling of IgG for 48 h identified
triantennary charged glycans (Fig. 5B), known to occur on
rGalT-1 (data not shown), which remained detectable in trace
amounts in the Protein A-purified IgG product (Fig. 5C).
DISCUSSION
It is widely recognized that modifying the structures of N-
glycans linked to the CH2 domain in the Fc region of certain
Figure 4. Analysis of 2-AA derivatized oligosaccharides from human
serum IgG after GalT-1 conversion to the G2 glycoform by matrix-assisted
laser desorption ionization time-of-flight mass spectrometry (MALDI-TOF
MS) in linear/negative-ion mode. Spectra of 2-AA labeled glycans from (A)the starting pool of polyclonal human serum IgG, (B) 48-h GalT-1-
remodeled IgG reaction mixture, and (C) rProtein A affinity purified
remodeled IgG. Glycan structures are represented by the symbols; ~,
fucose; *, mannose; &, GlcNAc; *, galactose, and *, sialic acid. An
asterisk above a peak indicates a sodium adduct, and an X represents a peak
that is not identified.No attemptwasmade to fully characterize or distinguish
among isobaric structures.
WARNOCK ET AL.: GALACTOSYLATION OF HUMAN IGG 837

IgG molecules may modulate binding interactions with
FcgRII and FcgRIII receptors (Jefferis and Lund, 2002;
Radaev and Sun, 2001; Tsuchiya et al., 1989) and with
activators of the complement cascade (Boyd et al., 1995).
The N-linked glycans of human IgG consist primarily of
neutral bi-antennary complex oligosaccharides, often a1–6fucosylated on the asparagine-linked core GlcNAc, with a
minor fraction containing bisecting GlcNAc (Fujii et al.,
1990; Kornfeld et al., 1971) (see Fig. 1 for structures).
Increased specific activities of some IgG molecules in
Figure 5. Analysis of chromatography fractions from rProtein A purification of GalT-1-remodeled human serum IgG. Panel A: Protein concentration traceduring fractionation of GalT-1-remodeled human serum IgG on Protein A-Sepharose. pH 3.0 glycine buffer elution was begun at fraction number 19. Panel B:SDS–PAGE analysis of fractions from Protein A-Sepharose chromatography of remodeled IgG. Equivalent volumes of each fraction were resolved on 4%–
12%NuPAGE gels with 3-(N-morpholino)propanesulfonic acid (MOPS) running buffer. Lanemarked ‘‘load’’ represents the IgG remodeling reactionmixture;
lanesmarked 5–10were flow-through fractions, fromwhich protein bandsmarked (a)–(d) were isolated and sequenced as described inMaterials andMethods.
Lanes marked 24–30 represent fractions eluted at pH 3.0. Panel C: Western blot analysis to detect GalT-1 in fractions from the Protein A Sepharose
chromatography fractionation of remodeled IgG. Approximately 3 mg of the remodeled IgG (lane 1 and 3) and Protein A purified remodeled IgG (lane 2 and 4)were resolved by NuPAGE gel electrophoresis. Lanes labeled Std, 1, and 2 were stained with colloidal Coomassie blue and imaged. Equivalent samples plus
prestainedmolecular weight markers (lane Std) were resolved in parallel on the same gel, blotted to nitrocellulose and probedwith antibody specific for GalT-1
as described in Materials and Methods. Lanes 3 and 4 were aligned based on the prestained molecular weight markers.
838 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 92, NO. 7, DECEMBER 30, 2005

triggering Fc receptor mediated ADCC in vitro has been
associated with both the presence of bisecting GlcNAc
(Davies et al., 2001; Rademacher et al., 1986; Umana et al.,
1999) and the absence of core fucose (Shields et al., 2002;
Shinkawa et al., 2003). The neutral bi-antennary sugar chains
containing zero, one, or two galactosyl units are commonly
termed, respectively, G0, G1, and G2 glycans (Dwek et al.,
1995; Wormald et al., 1997) (see Fig. 1). That the relative
abundance of these glycoforms of IgG may be associated
with human disease was described by Parekh et al. (1985)
who observed hypo-galactosylated IgG in patients with
rheumatoid arthritis and osteoarthritis. Malhotra et al. (1995)
found that aggregated agalacto-IgG may activate the
complement pathway through interaction with the man-
nose-binding protein and suggested the potential importance
of this pathway in the genesis of auto-immune diseases.
Placental transport and rescue of antibody from catabolism
are hypothesized to occur via a transcytosis process that
requires binding to the FcgRn (Martin et al., 2001). Although
agalacto-antibody binds efficiently to the FcgRn in vitro,
some speculation and contradictory evidence exists as to IgG
glycan involvement on FcgRn binding in vivo, particularly inplacental transport where more highly galactosylated IgGs
appear to be preferentially transported to the fetal circulation
(Jefferis et al., 1998; Williams et al., 1995).
In the present work, we have explored the possibility of
preparative in vitro conversion of IgG molecules to
homogeneousG2 glycoforms by treatment with recombinant
bovine b1–4 galactosyltransferase (GalT-1, EC 2.4.1.22)
plus the sugar nucleotide donor, UDP-galactose. The
acceptor specificity and reaction products of this enzyme
have been studied extensively (Khatra et al., 1974), and at
analytical scale the enzyme previously has been shown to act
on intact IgG, adding galactose in b1–4 linkage to terminal
GlcNAc residues on both arms of the G0 N-linked bi-
antennary chain (Fujii et al., 1990; Raju et al., 2001). The
human serum IgG starting material used for the present study
contained approximately 33% G0, 47% G1, and 20% G2, a
distribution of neutral glycans that is undergalactosylated
compared with the values, 19% G0, 34% G1, 47% G2,
reported by Routier et al. (1998), for purified human IgG, but
similar to the values, 28%G0, 46%G1, 26%G2, reported for
human IgG purchased from a commercial source (Raju et al.,
2000).
We found that the rate of conversion ofG0 toG1was rapid,
proceeding essentially to completion within 4 h in a reaction
mixture containing IgG at 30 mg/mL and GalT-1 at 25 mU/
mg IgG (Fig. 3). In the same reaction mixture, conversion of
G1 to G2 proceeded more slowly, decreasing dramatically
after 18 h as the concentration of G1 acceptor diminished to
<5% of the starting value. As expected, we observed that the
rate of transfer of galactose increases as the activity ratio (U
GalT-1/mg IG) is increased (Table I), and that galactosylation
can be carried outmore efficiently at higher concentrations of
IgG up to 30mg/mL (Table II). The latter results suggest that
at concentrations of IgG less than 30 mg/mL (187 mM), the
acceptor substrate is below that required for enzyme
saturation. For rat liver GalT-1 the apparent Km for G0 is
0.13 mM, and the Km values are 0.43 and 6.28 mM,
respectively, for a1,3 and a1,6 branched G1 acceptors as
isolated oligosaccharides (Paquet et al., 1984). It has been
reported that the corresponding glycan structures within
native IgG are less accessible to galactosyltransferase (Fujii
et al., 1990; White et al., 1997).
In a production setting, heterogeneity of glycosylation of a
recombinant glycoprotein may be the result of incomplete
Golgi processing in cells due to changing metabolic states
during fermentation and/or the presence of glycosidases
released into the cell culture medium (Goochee et al., 1991;
Jenkins et al., 1996). Recombinant mAbs expressed in
mammalian cell lines exhibit G0, G1, and G2 glycoforms
(Raju et al., 2000), but percentage distributions may vary
depending upon multiple factors, including the species from
which the host expressor cell line was derived and details of
the cell culture conditions (Hills et al., 2001; Kumpel et al.,
1994; Raju et al., 2000; Jefferis, 2005). Thus, it can be a
Table III. Total N-glycan profiles of starting human IgG and of galactosylated IgG before and after purification
on Protein A.
Percent of total glycans
Neutral glycans Charged glycans: number of sialic acids
G0 G1 G2 1 2 3 4
Starting IgG 20.2a 31.2 15.9 21.0 11.7 — —
IgG/GalT-1 reactionmixb — — 65.3 22.6 12.1 — —
IgG/GalT-1 pH 3 eluate — — 75.5 18.5 6.0 — —
IgG/GalT-1 flow-through [26.0] 26.6 45.9 1.5 —
aPercentages of total glycans were determined by integration peaks from single RP-HPLC chromatogramsof 2-AA labeled derivatives analyzed under conditions to optimally resolve charged glycans (see Materials andMethods).
bIgG (1 kg/33.5 L)was incubatedwith 25mUGalT-1/mg IgG for 48 h. An aliquot of the reactionwas applied toProtein A-Sepharose, washed with Tris-buffered saline at pH 7.2, and eluted with 0.2 M glycine-HCl, pH 3.0 (seeMaterials and Methods).
—-, indicates the structure was not detected.
WARNOCK ET AL.: GALACTOSYLATION OF HUMAN IGG 839

significant challenge to achieve uniform batch-to-batch
glycosylation for a recombinant IgG product. Attempts have
beenmade to engineer glycosylation pathways so as to enrich
for specific glycoforms in mAbs secreted by a variety of
expression systems, including cell lines from mammalian
species (Davies et al., 2001), yeast (Bobrowicz et al., 2004;
Vervecken et al., 2004), and plants (Ko et al., 2003; Shriver
et al., 2004; Tekoah et al., 2004).
An emerging alternative is the use of enzymes post-
production to remodel glycan chains, a process made pos-
sible by high-level overexpression of glycosyltransferases in
soluble form (Malissard et al., 2000; Perugino et al., 2004).
We have demonstrated previously the preparative in vitro
remodeling of glycan chains on a therapeutic glycoprotein at
the 10 g scale, including enzymatic sialylation and fucosyla-
tion of complex biantennary N-glycans to homogeneous
sialyl Lewis X (sLex)-active glycoforms (Thomas et al.,
2004). The present work demonstrates the feasibility of using
recombinant GalT-1 to achieve complete (>98%) galacto-
sylation of IgG under mild conditions within 48 h at pilot
production (1 kg) scale. Using Protein A affinity chromato-
graphy, we showed that it is feasible to purify fully galacto-
sylated IgG from the enzyme reactionmixture containing the
glycosylating reagents GalT-1 and UDP-galactose. To
demonstrate scale-up of the galactosylation reaction, we
used a relatively inexpensive and easily available commer-
cial preparation of pooled human IgG containing <5%
contaminating human serum proteins. The fact that this IgG
preparation undoubtedly contains multiple IgG subtypes and
allotypes provides evidence that in vitro enzymatic galacto-
sylation as described here is likely to be robust in its
applicability to multiple forms of the human IgG molecule,
including mAbs.
ABBREVIATIONS
2-AA 2-anthranilic acid
ADCC antigen dependent cellular cytotoxicity
b4GalT-1 b4-galactosyltransferase-IBSA bovine serum albumin
CH2 antibody constant domain
CML cell-mediated lysis
Fc antibody fragment that crystallizes
FF fast flow
FPLC fast protein liquid chromatography
GlcNAc N-acetyl glucosamine
HPLC high performance liquid chromatography
HRP horseradish peroxidase
IgG immunoglobulin G
LC/ESI/MS liquid chromatography electrospray ionization mass
spectrometry
mAb monoclonal antibody
MALDI-TOF MS matrix-assisted laser desorption ionization
time-of-flight mass spectrometry
MES (N-morpholino)ethane-sulfonic acid
MOPS 3-(N-morpholino)propanesulfonic acid
MS mass spectrometry
NSO murine cell line (Lonza Biologics)
PBS phosphate buffered saline
PNGase F peptide N-glycosidase F
RP-HPLC reversed phase liquid chromatography
SDS sodium dodecyl sulfate
SDS–PAGE sodium dodecyl sulfate–polyacrylamide
gel electrophoresis
sLex sialyl Lewis X
TBS tris buffered saline
TBS-T20 20mMTris, pH 7.2, 150mMNaCl, 0.05%Tween-20
UDP uridine 50-diphosphateUMP uridine 50-monophosphate
References
Anumula KR, Dhume ST. 1998. High resolution and high sensitivity
methods for oligosaccharide mapping and characterization by normal
phase high performance liquid chromatography following derivatization
with highly fluorescent anthranilic acid. Glycobiology 8(7):685–694.
Bobrowicz P, Davidson RC, Li H, Potgieter TI, Nett JH, Hamilton SR,
StadheimTA,MieleRG,BobrowiczB,Mitchell T. 2004. Engineering of
an artificial glycosylation pathway blocked in core oligosaccharide
assembly in theyeastPichia pastoris: Productionof complex humanized
glycoproteins with terminal galactose. Glycobiology 14(9):757–766.
Boyd PN, Lines AC, Patel AK. 1995. The effect of the removal of sialic acid,
galactose, and total carbohydrate on the functional activity of
Campath-1H. Mol Immunol 32(17–18):1311–1318.
ClynesRA, Towers TL, Presta LG, Ravetch JV. 2000. Inhibitory Fc receptors
modulate in vivo cytoxicity against tumor targets. Nat Med 6(4):
443–446.
ColomaMJ,CliftA,WimsL,MorrisonSL. 2000.The role of carbohydrate in
the assembly and function of polymeric IgG. Mol Immunol
37(18):1081–1090.
Davies J, Jiang L, Pan LZ, LaBarre MJ, Anderson D, Reff M. 2001.
Expression of GnTIII in a recombinant anti-CD20 CHO production cell
line: Expression of antibodies with altered glycoforms leads to an
increase in ADCC through higher affinity for FC gamma RIII.
Biotechnol Bioeng 74(4):288–294.
DijstelbloemHM, van deWinkel JG, Kallenberg CG. 2001. Inflammation in
autoimmunity: Receptors for IgG revisited. Trends Immunol
22(9):510–516.
Dwek RA, Lellouch AC, Wormald MR. 1995. Glycobiology: ‘The function
of sugar in the IgG molecule’. J Anat 187(Pt 2):279–292.
Fujii S, Nishiura T, Nishikawa A, Miura R, Taniguchi N. 1990. Structural
heterogeneity of sugar chains in immunoglobulin G. Conformation of
immunoglobulin G molecule and substrate specificities of glycosyl-
transferases. J Biol Chem 265(11):6009–6018.
Ghirlando R, Lund J, Goodall M, Jefferis R. 1999. Glycosylation of human
IgG-Fc: Influences on structure revealed by differential scanningmicro-
calorimetry. Immunol Lett 68(1):47–52.
Goochee CF, Gramer MJ, Andersen DC, Bahr JB, Rasmussen JR. 1991. The
oligosaccharides of glycoproteins: Bioprocess factors affecting oligo-
saccharide structure and their effect on glycoprotein properties.
Biotechnology (NY) 9(12):1347–1355.
Hamako J, Matsui T, Ozeki Y, Mizuochi T, Titani K. 1993. Comparative
studies of asparagine-linked sugar chains of immunoglobulin G from
elevenmammalian species. CompBiochemPhysiol B 106(4):949–954.
Hills AE, Patel A, Boyd P, James DC. 2001. Metabolic control of
recombinant monoclonal antibody N-glycosylation in GS-NS0 cells.
Biotechnol Bioeng 75(2):239–251.
Jefferis R. 2001. Glycosylation of human IgG antibodies: Relevance to
therapeutic applications. BioPharm International 14(9):19–27.
Jefferis R. 2005. Glycosylation of recombinant antibody therapeutics.
Biotechnol Prog 21(1):11–16.
Jefferis R, Lund J. 2002. Interaction sites on human IgG-Fc for FcgammaR:
Current models. Immunol Lett 82(1–2):57–65.
Jefferis R, Lund J, Pound JD. 1998. IgG-Fc-mediated effector functions:
Molecular definition of interaction sites for effector ligands and the role
of glycosylation. Immunol Rev 163:59–76.
840 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 92, NO. 7, DECEMBER 30, 2005

Jenkins N, Parekh RB, James DC. 1996. Getting the glycosylation right:
Implications for the biotechnology industry. Nat Biotechnol 14(8):975–
981.
Khatra BS, Herries DG, Brew K. 1974. Some kinetic properties of human-
milk galactosyl transferase. Eur J Biochem 44(2):537–560.
Ko K, Tekoah Y, Rudd PM, Harvey DJ, Dwek RA, Spitsin S, Hanlon CA,
Rupprecht C,DietzscholdB,GolovkinM,KoprowskiH. 2003. Function
and glycosylation of plant-derived antiviral monoclonal antibody. Proc
Natl Acad Sci USA 100(13):8013–8018.
Kornfeld R, Keller J, Baenziger J, Kornfeld S. 1971. The structure of the
glycopeptide of human gamma G myeloma proteins. J Biol Chem
246(10):3259–3268.
Krapp S, Mimura Y, Jefferis R, Huber R, Sondermann P. 2003. Structural
analysis of human IgG-Fc glycoforms reveals a correlation between
glycosylation and structural integrity. J Mol Biol 325(5):979–989.
Kumpel BM, Rademacher TW, Rook GA, Williams PJ, Wilson IB. 1994.
Galactosylation of human IgG monoclonal anti-D produced by EBV-
transformedB-lymphoblastoid cell lines is dependent on culturemethod
and affects Fc receptor-mediated functional activity. Hum Antibodies
Hybridomas 5(3–4):143–151.
Leatherbarrow RJ, Rademacher TW, Dwek RA, Woof JM, Clark A, Burton
DR, Richardson N, Feinstein A. 1985. Effector functions of a
monoclonal aglycosylated mouse IgG2a: Binding and activation of
complement component C1 and interaction with human monocyte Fc
receptor. Mol Immunol 22(4):407–415.
Link AJ, Eng J, Schieltz DM, Carmack E, Mize GJ, Morris DR, Garvik BM,
Yates JR III. 1999. Direct analysis of protein complexes using mass
spectrometry. Nat Biotechnol 17(7):676–682.
Lund J, Takahashi N, Pound JD, Goodall M, Nakagawa H, Jefferis R. 1995.
Oligosaccharide–protein interactions in IgG can modulate recognition
by Fc gamma receptors. FASEB J 9(1):115–119.
Maini R, St. Clair EW, Breedveld F, Furst D, Kalden J, WeismanM, Smolen
J, Emery P, Harriman G, Feldmann M, Lipsky P. 1999. Infliximab
(chimeric anti-tumour necrosis factor alpha monoclonal antibody)
versus placebo in rheumatoid arthritis patients receiving concomitant
methotrexate: A randomised phase III trial. ATTRACT Study Group.
Lancet 354(9194):1932–1939.
Malhotra R,WormaldMR, Rudd PM, Fischer PB, DwekRA, SimRB. 1995.
Glycosylation changes of IgG associated with rheumatoid arthritis can
activate complement via the mannose-binding protein. Nat Med
1(3):237–243.
Malissard M, Zeng S, Berger EG. 2000. Expression of functional soluble
forms of human beta-1, 4-galactosyltransferase I, alpha-2,6-sialyltrans-
ferase, and alpha-1, 3-fucosyltransferase VI in the methylotrophic yeast
Pichia pastoris. Biochem Biophys Res Commun 267(1):169–173.
MartinWL,West AP, Jr., Gan L, Bjorkman PJ. 2001. Crystal structure at 2.8
A of an FcRn/heterodimeric Fc complex: Mechanism of pH-dependent
binding. Mol Cell 7(4):867–877.
Mimura Y, Church S, Ghirlando R, Ashton PR, Dong S, Goodall M, Lund J,
Jefferis R. 2000. The influence of glycosylation on the thermal stability
and effector function expression of human IgG1-Fc: Properties of a
series of truncated glycoforms. Mol Immunol 37(12–13):697–706.
Mimura Y, Sondermann P, Ghirlando R, Lund J, Young SP, Goodall M,
Jefferis R. 2001. Role of oligosaccharide residues of IgG1-Fc in Fc
gamma RIIb binding. J Biol Chem 276(49):45539–45547.
Narimatsu H, Sinha S, Brew K, Okayama H, Qasba PK. 1986. Cloning and
sequencing of cDNA of bovine N-acetylglucosamine (beta 1–
4)galactosyltransferase. Proc Natl Acad Sci USA 83(13):4720–4724.
Paquet MR, Narasimhan S, Schachter H, Moscarello MA. 1984. Branch
specificity of purified rat liver Golgi UDP-galactose: N-acetylglucosa-
mine beta-1,4-galactosyltransferase. Preferential transfer of of galactose
on theGlcNAc beta 1,2-Man alpha 1,3-branch of a complex biantennary
Asn-linked oligosaccharide. J Biol Chem 259(8):4716–4721.
Parekh RB, Dwek RA, Sutton BJ, Fernandes DL, Leung A, Stanworth D,
Rademacher TW, Mizuochi T, Taniguchi T, Matsuta K, Takeuchi P,
Nagano Y, Miyamoto T, Kobata A. 1985. Association of rheumatoid
arthritis and primary osteoarthritis with changes in the glycosylation
pattern of total serum IgG. Nature 316(6027):452–457.
Perugino G, Trincone A, Rossi M, Moracci M. 2004. Oligosaccharide
synthesis by glycosynthases. Trends Biotechnol 22(1):31–37.
Presta LG. 2002. Engineering antibodies for therapy. Curr PharmBiotechnol
3(3):237–256.
Radaev S, Sun PD. 2001. Recognition of IgG by Fcgamma receptor. The role
of Fc glycosylation and the binding of peptide inhibitors. J Biol Chem
276(19):16478–16483.
Rademacher TW, Homans SW, Parekh RB, Dwek RA. 1986. Immunoglo-
bulin G as a glycoprotein. Biochem Soc Symp 51:131–148.
Raju TS, Briggs JB, Borge SM, Jones AJ. 2000. Species-specific variation in
glycosylation of IgG: Evidence for the species-specific sialylation and
branch-specific galactosylation and importance for engineering recom-
binant glycoprotein therapeutics. Glycobiology 10(5):477–486.
Raju TS, Briggs JB, Chamow SM, Winkler ME, Jones AJ. 2001.
Glycoengineering of therapeutic glycoproteins: In vitro galactosylation
and sialylation of glycoproteins with terminalN-acetylglucosamine and
galactose residues. Biochemistry 40(30):8868–8876.
Rastetter W, Molina A, White CA. 2004. Rituximab: Expanding role in
therapy for lymphomas and autoimmune diseases. Annu Rev Med
55:477–503.
Routier FH, Hounsell EF, Rudd PM, Takahashi N, BondA, Hay FC,AlaviA,
Axford JS, Jefferis R. 1998. Quantitation of the oligosaccharides
of human serum IgG from patients with rheumatoid arthritis: A
critical evaluation of different methods. J Immunol Methods
213(2):113–130.
Sandborn WJ, Hanauer SB. 1999. Antitumor necrosis factor therapy for
inflammatory bowel disease: A review of agents, pharmacology, clinical
results, and safety. Inflamm Bowel Dis 5(2):119–133.
Shields RL, Lai J, Keck R, O’Connell LY, Hong K, Meng YG, Weikert SH,
Presta LG. 2002. Lack of fucose on human IgG1 N-linked oligosacchar-
ide improves binding to human Fcgamma RIII and antibody-dependent
cellular toxicity. J Biol Chem 277(30):26733–26740.
Shinkawa T, Nakamura K, Yamane N, Shoji-Hosaka E, Kanda Y, Sakurada
M, Uchida K, Anazawa H, SatohM, Yamasaki M. 2003. The absence of
fucose but not the presence of galactose or bisecting N-acetylglucosa-
mine of human IgG1 complex-type oligosaccharides shows the critical
role of enhancing antibody-dependent cellular cytotoxicity. J Biol Chem
278(5):3466–3473.
Shriver Z, Raguram S, Sasisekharan R. 2004. Glycomics: A pathway to a
class of new and improved therapeutics. Nat Rev Drug Discov
3(10):863–873.
Souriau C, Hudson PJ. 2003. Recombinant antibodies for cancer diagnosis
and therapy. Expert Opin Biol Ther 3(2):305–318.
Tekoah Y, Ko K, Koprowski H, Harvey DJ, Wormald MR, Dwek RA, Rudd
PM. 2004. Controlled glycosylation of therapeutic antibodies in plants.
Arch Biochem Biophys 426(2):266–278.
Thomas LJ, Panneerselvam K, Beattie DT, Picard MD, Xu B, Rittershaus
CW, Marsh HC, Jr., Hammond RA, Qian J, Stevenson T. 2004.
Production of a complement inhibitor possessing sialyl Lewis X
moieties by in vitro glycosylation technology. Glycobiology
14(10):883–893.
TsuchiyaN,EndoT,MatsutaK,YoshinoyaS,AikawaT,KosugeE, Takeuchi
F, Miyamoto T, Kobata A. 1989. Effects of galactose depletion from
oligosaccharide chains on immunological activities of human IgG. J
Rheumatol 16(3):285–290.
Umana P, Jean-Mairet J,MoudryR,AmstutzH, Bailey JE. 1999. Engineered
glycoforms of an antineuroblastoma IgG1 with optimized antibody-
dependent cellular cytotoxic activity. Nat Biotechnol 17(2):176–180.
Vervecken W, Kaigorodov V, Callewaert N, Geysens S, De Vusser K,
Contreras R. 2004. In vivo synthesis of mammalian-like, hybrid-typeN-
glycans in Pichia pastoris. Appl Environ Microbiol 70(5):2639–2646.
Walker MR, Lund J, Thompson KM, Jefferis R. 1989. Aglycosylation of
human IgG1 and IgG3monoclonal antibodies can eliminate recognition
by human cells expressing Fc gammaRI and/or Fc gammaRII receptors.
Biochem J 259(2):347–353.
White KD, Cummings RD, Waxman FJ. 1997. Ig N-glycan orientation can
influence interactions with the complement system. J Immunol
158(1):426–435.
WARNOCK ET AL.: GALACTOSYLATION OF HUMAN IGG 841

White CA,Weaver RL, Grillo-Lopez AJ. 2001. Antibody-targeted immuno-
therapy for treatment of malignancy. Annu Rev Med 52:125–145.
Williams PJ, Arkwright PD, Rudd P, Scragg IG, Edge CJ, Wormald MR,
Rademacher TW. 1995. Short communication: Selective placental
transport of maternal IgG to the fetus. Placenta 16(8):749–756.
WormaldMR,Rudd PM,HarveyDJ, Chang SC, Scragg IG,DwekRA. 1997.
Variations in oligosaccharide–protein interactions in immunoglobulin
G determine the site-specific glycosylation profiles and modulate
the dynamic motion of the Fc oligosaccharides. Biochemistry
36(6):1370–1380.
Wright A, Morrison SL. 1998. Effect of C2-associated carbohydrate
structure on Ig effector function: Studies with chimeric mouse-human
IgG1 antibodies in glycosylation mutants of Chinese hamster ovary
cells. J Immunol 160(7):3393–3402.
842 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 92, NO. 7, DECEMBER 30, 2005