
Cyclocondensations of homophthalic anhydrides with 1-aza-1,3-dienes
Upload
independentCategory
view
3download
0
Glycobiology vol. 18 no. 12 pp. 1094–1104, 2008doi:10.1093/glycob/cwn094Advance Access publication on September 29, 2008
Identification of the Drosophila core 1 β1,3-galactosyltransferase gene that synthesizesT antigen in the embryonic central nervous system and hemocytes
Hideki Yoshida2,3,4,5, Takashi J Fuwa3,4,5, Mikiko Arima4,Hiroshi Hamamoto4, Norihiko Sasaki4, Tomomi Ichimiya4,Ken-ichi Osawa4, Ryu Ueda5,6, and Shoko Nishihara1,4,5
4Department of Bioinformatics, Laboratory of Cell Biology, Faculty ofEngineering, Soka University, 1-236 Tangi-cho, Hachioji, Tokyo 192-8577;5Core Research for Evolutional Science and Technology (CREST) of JapanScience and Technology Agency (JST), Kawaguchi Center Building, 4-1-8,Hon-cho, Kawaguchi, Saitama 332-0012; and 6Invertebrate GeneticsLaboratory, National Institute of Genetics, 1111 Yata, Mishima, Shizuoka441-8540, Japan
Received on January 15, 2008; revised on September 6, 2008; accepted onSeptember 23, 2008
T antigen (Galβ1-3GalNAcα1-Ser/Thr), the well-knowntumor-associated antigen, is a core 1 mucin-type O-glycan structure that is synthesized by core 1 β1,3-galactosyltransferase (C1β3GalT), which transfers Gal fromUDP-Gal to Tn antigen (GalNAcα1-Ser/Thr). Three puta-tive C1β3GalTs have been identified in Drosophila. However,although all three are expressed in embryos, their roles dur-ing embryogenesis have not yet been clarified. In this study,we used P-element inserted mutants to show that CG9520,one of the three putative C1β3GalTs, synthesizes T antigenexpressed on the central nervous system (CNS) during em-bryogenesis. We also found that T antigen was expressedon a subset of the embryonic hemocytes. CG9520 mutantembryos showed the loss of T antigens on the CNS andon a subset of hemocytes. Then, the loss of T antigens wasrescued by precise excision of the P-element inserted intothe CG9520 gene. Our data demonstrate that T antigensexpressed on the CNS and on a subset of hemocytes aresynthesized by CG9520 in the Drosophila embryo. In ad-dition, we found that the number of circulating hemocyteswas reduced in third instar larvae of CG9520 mutant. We,therefore, named the CG9520 gene Drosophila core 1 β1,3-galactosyltransferase 1 because it is responsible for the syn-thesis and function of T antigen in vivo.
Keywords: CNS/core 1 β1, 3-galactosyltransferase/Drosophila/hemocyte/T antigen
Introduction
Mucin-type O-glycans contain GalNAc in an α1-linkage to aserine or threonine residue. This posttranslational modification
1To whom correspondence should be addressed: Tel: +81-426-91-8140; Fax:+81-426-91-8140; e-mail: [email protected] address: Terrence Donnelly Centre for Cellular Biomolecular Research,University of Toronto, 160 College Street, Toronto, Ontario, M5S 3E1, Canada.3These authors contributed equally to this work.
is found on many membrane-bound and -secreted pro-teins (Van den Steen et al. 1998). In vertebrates, eightdifferent O-glycan core structures have been described(Brockhausen 1997). There is evidence that one of these corestructures, core 1 structure (Galβ1-3GalNAcα1-Ser/Thr), calledT or Thomsen–Friedenreich antigen, is associated with immuno-suppression, metastasis dissemination, and the proliferation ofseveral types of cancer cells (Springer et al. 1984; Berger 1999;Brockhausen 1999).
The core 1 structure is synthesized from GalNAcα1-Ser/Thr by core 1 β1,3-galactosyltransferase (C1β3GalT).C1β3GalTs have been identified from various organisms, in-cluding rat (Ju, Cummings et al. 2002), human (Ju, Breweret al. 2002), mouse (Xia et al. 2004), and Caenorhabditis ele-gans (Ju et al. 2006). Xia et al. recently reported that C1β3GalT(T-synthase) null mice showed embryonic lethality, defectiveangiogenesis, and fatal embryonic hemorrhage (Xia et al. 2004;Xia and McEver 2006). Thrombocytopenia and kidney dis-ease have been observed in plt1 mice, which have an N-ethyl-N-nitrosourea-induced point mutation in the C1β3GalT geneand very low residual C1β3GalT activity (Alexander et al.2006). However, most mammalian mucin-type core 1 struc-tures are elongated or modified by sialylation or fucosylation(Brockhausen 1999). It has also been reported that T anti-gen is expressed in the normal placenta, seminal plasma, andthe developing and adult kidney in humans (Richter et al.2000; Toma et al. 2000; Yamaguchi et al. 2001). Recently,it was reported that three putative Drosophila C1β3GalTshave C1β3GalT activity in vitro (Muller et al. 2005). Theyshowed activity on glycolipids as well as mucin proteins. How-ever, the in vivo roles of these C1β3GalTs have not yet beenclarified.
In Drosophila, the only mucin-type O-glycans identifiedto date are T antigen and Tn antigen (GalNAcα1-Ser/Thr)(Kramerov et al. 1996). Unlike vertebrates, these structures arenot sialylated in Drosophila. It is known that the Drosophilaα2,6-sialyltransferase cannot transfer sialic acid to T antigen(Koles et al. 2004). In addition, it has been reported that thedistribution of T antigen, identified using peanut agglutinin(PNA) lectin, is regulated in tissue- and stage-specific mannersduring embryonic development (Fristrom DK and Fristrom JW1982; D’Amico and Jacobs 1995). These data remind us thatDrosophila may be a suitable model system for investigatingthe functions of T antigen and the C1β3GalT genes duringdevelopment.
In this study, we demonstrate that the CG9520 gene, oneof the Drosophila orthologs of human C1β3GalT, is involvedin synthesis of T antigen in the central nervous system (CNS)and a subset of hemocytes during the developmental process ofDrosophila.
c© The Author 2008. Published by Oxford University Press. All rights reserved. For permissions, please e-mail: [email protected] 1094
by guest on January 14, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from

Drosophila C1β3GalT synthesizes T antigen on the CNS
Results
Candidate for synthesizing T antigen in Drosophila embryosRecently, the Drosophila genome was reported to have atleast three C1β3GalT genes, CG9520, CG8708, and CG13904(Muller et al. 2005). To estimate which C1β3GalT synthe-sizes T antigen in vivo, we built a phylogenetic tree fromthe sequences of the three Drosophila C1β3GalTs, humanC1β3GalT (Ju, Brewer, et al. 2002), mouse T-synthase (Xiaet al. 2004), C. elegans T-synthase (Ju et al. 2006), rat C1β3GalT(Ju, Cummings, et al. 2002), and putative chicken, Xenopus andzebrafish C1β3GalTs (Figure 1A). The phylogenetic tree indi-cated that CG9520 and CG8708 are the orthologs of mammalianC1β3GalTs responsible for the synthesis of T antigen. Then, weselected the two as the first candidates for synthesizing T antigenin vivo. To distinguish which of the two is responsible for thesynthesis of T antigen, we investigated the expression patternsof the CG9520 and CG8708 genes during embryonic devel-opment and also in several tissues and cultured cell lines byquantitative real-time PCR (Figure 1B). In embryos, the highestlevel of CG9520 transcript was found at 2 h and was maternallyderived. On the other hand, CG8708 transcripts were stronglyexpressed in the larval salivary glands and testis of adult male.Recently, Muller et al. used an in situ hybridization analysis toshow that the CG9520 transcript is maternally expressed andalso expressed in the amnioserosa of late-stage embryos. In ad-dition, they found that the CG8708 transcript is only expressedin the salivary glands of the embryos. They also showed thatCG9520 exhibits more than 100 times higher C1β3GalT activ-ity than CG8708 (Muller et al. 2005). In embryos, the levels ofCG8708 transcripts were less than 10 times of those of CG9520transcripts. Therefore, CG9520 is a good candidate for synthe-sizing T antigen in embryos.
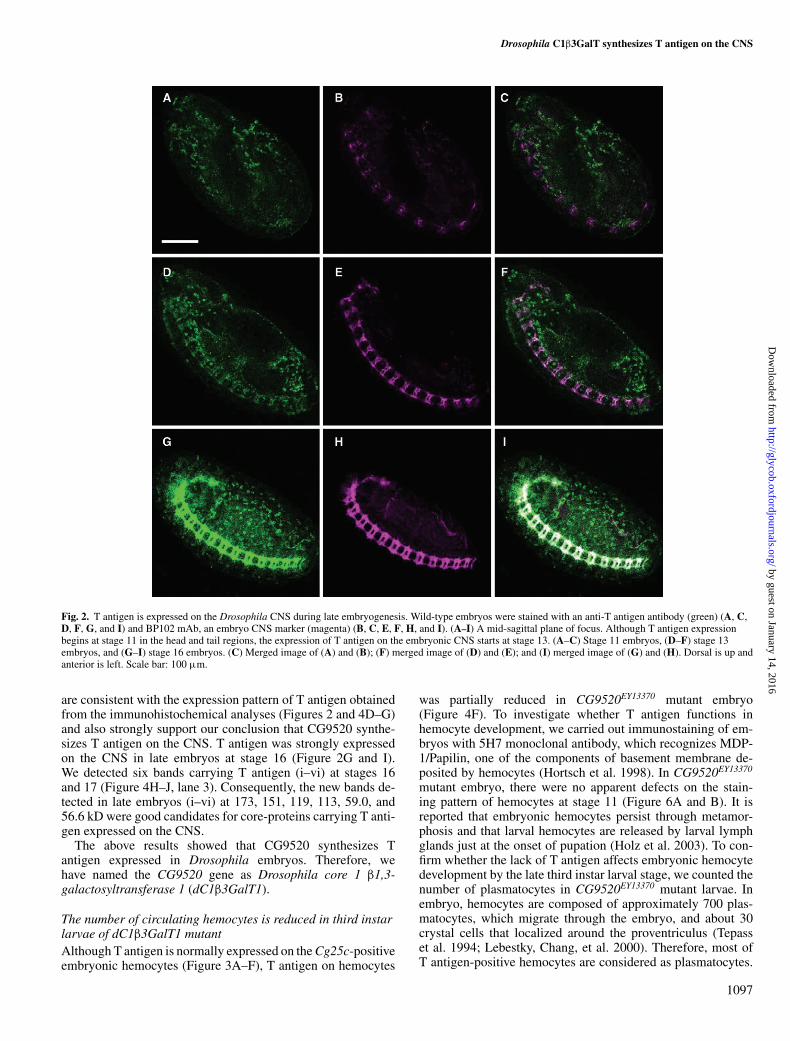
T antigen is present on the embryonic CNS of DrosophilaTo investigate the expression patterns of T antigen during de-velopment, we used an anti-T antigen monoclonal antibody toimmunostain Drosophila embryos. T antigen was first clearlydetected at the head and tail region of stage 11 embryos(Figure 2A). By stage 16, T antigen expressing cells wereevenly dispersed throughout the embryos (Figure 2D and G).Interestingly, T antigen was strongly expressed on the CNS dur-ing late embryogenesis (Figure 2G). To confirm that T antigenis expressed on the CNS, we coimmunostained embryos withan anti-T antigen antibody and mAb BP102, a CNS marker(Figure 2B, E and H). We did not detect colocalization of Tantigen and BP102 on the CNS at stage 11 (Figure 2C), butdid find colocalization after stage 13 (Figure 2F and I). Theseobservations show that T antigen is expressed in the developingCNS during embryogenesis.
T antigen is also expressed on a subset of embryonic hemocytesThe expression pattern of T antigen during embryogenesis wassimilar to the distribution of Drosophila embryonic hemocytes(Tepass et al. 1994; Cho et al. 2002), except for the CNS.Drosophila embryonic hemocytes derive exclusively from themesoderm of the head region of stage 8 embryos and dispersealong several invariant migratory paths throughout the embryoduring late embryogenesis (Cho et al. 2002). To investigatewhether T antigen is expressed in embryonic hemocytes, we
carried out coimmunostaining of embryos carrying Cg25C-lacZwith an anti-T antigen antibody and an anti-β-galactosidase an-tibody. This fly line has the enhancer trap insertion, A109.1F2,inserted into the promoter region of the collagen IV gene, locatedcytologically at 25C (Cg25C), and expresses β-galactosidase inembryonic hemocytes (Bellen et al. 1989; Wilson et al. 1989).A subset of hemocytes expressing Cg25C-lacZ coexpressedT antigen in their cytoplasm; there were also indications thatthe antigen might also be expressed on hemocyte cell surfaces(Figure 3A–F). In Figure 3A–F, T antigen is simultaneously ex-pressed on the CNS but, because of the narrow focal plane ofconfocal microscopy, it is out of focus.
The S2 cell line was derived from a primary culture of late-stage Drosophila embryos (Schneider 1972). It has macrophage-like phagocytic properties and is believed to be derived fromembryonic hemocytes (Ramet et al. 2001). Interestingly, ourreal-time PCR data showed that the CG9520 transcript is highlyexpressed in the S2 cells. To investigate whether T antigenwas expressed in the S2 cells, we stained them using an anti-T antigen antibody. T antigen expression was detected on theS2 cells; it is also worth noting that a large focus of T antigenwas often observed at the contact point between two S2 cells(Figure 3G).
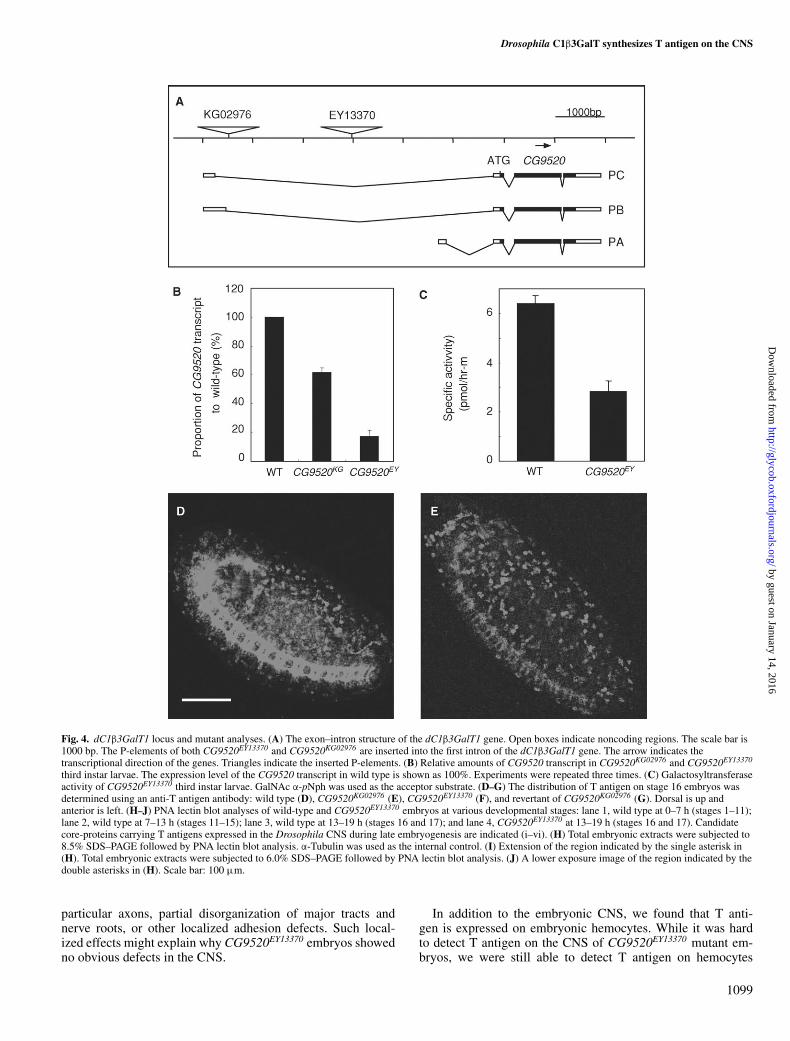
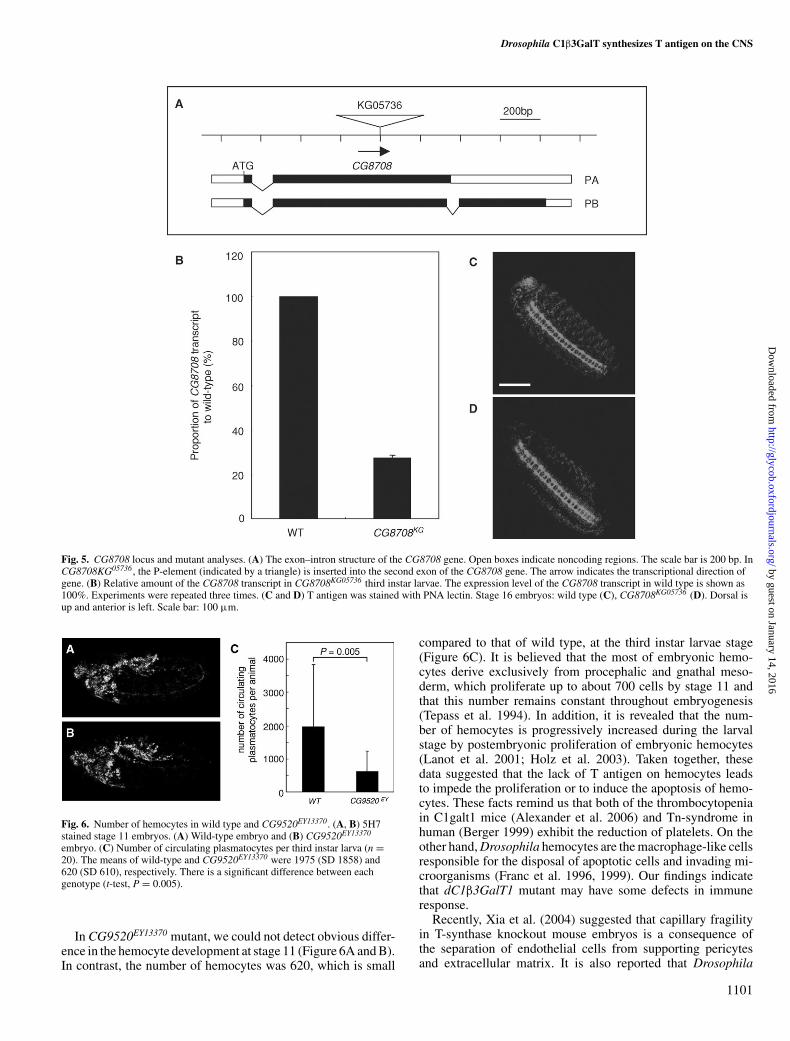
CG9520 synthesizes T antigen on the CNS during embryonicdevelopmentAs described above, CG9520 and CG8708 were the candi-dates for synthesizing T antigen in vivo. Two mutant lines,CG9520EY13370 and CG9520KG02976, have a P-element inser-tion in the CG9520 gene region. Although the P-elements arelocated in the first intron of the CG9520 gene in both lines(Figure 4A), CG9520KG02976 homozygotes develop normallywhereas some CG9520EY13370 homozygotes die during devel-opment and escaper homozygous CG9520EY13370 adult flieshave abnormal legs (data not shown). One CG8708 mutantline, CG8708KG05736, contains a P-element inserted into the sec-ond exon of the CG8708 gene (Figure 5A). The levels of theCG9520 and CG8708 transcripts in mutant third instar larvaewere measured by quantitative real-time PCR. In third instarlarvae homozygous for CG9520KG02976 or CG9520EY13370, thelevel of the CG9520 transcript was, respectively, reduced to 60%and 15% of that of wild-type third instar larvae (Figure 4B). Inaddition to the decrease in the CG9520 transcript, the extractsof CG9520EY13370 third instar larvae had a galactosyltransferaseactivity of 2.8 pmol/h mg protein (44% of that of wild-type)using GalNAcα-pNph as the acceptor (Figure 4C). Third instarlarvae homozygous for CG8708KG05736 had less than 28% of theCG8708 transcript present in wild type (Figure 5B).
To determine whether CG9520 or CG8708 synthesizes T anti-gen during embryogenesis, we carried out immunostaining ofCG9520 (Figure 4D–F) and CG8708 (Figure 5C and D) mutantembryos with an anti-T antigen monoclonal antibody. Althoughthe expression pattern of T antigen on hemocytes was simi-lar in CG9520KG02976 embryos to that of wild-type embryos,the expression level of T antigen on the CNS was slightly de-creased (Figure 4E). In CG9520EY13370 embryos, we could onlydetect low levels of the expression of T antigen on hemocytesand did not detect any expression on the CNS (Figure 4F).Furthermore, the expression pattern of T antigen was normalin CG9520KG02976 embryos following precise excision of the
1095
by guest on January 14, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from

H Yoshida et al.
B
CG13904
CG8708
CG9520
C. elegans
chicken
human
mouse
rat
Xenopus
zebrafish
0.05
A
0
20
40
60
80
100
120
140
Bra
in
Win
g di
sc
Eye
dis
c
Leg
disc
Sal
ivar
y gl
and
Tes
tis
Ova
ry
BG
3A-C
2
S2
0
50
100
150
200
250
0-2
2-4
4-8
8-12
12-1
6
16-2
2
1st
2nd
3rd
Pup
a
Mal
e
Fem
ale
0
2
4
6
8
10
12
14
16
0
5
10
15
20
25
Embryo (hour) Larva Adult Larva Adult Cell line
Rel
ativ
e am
ount
of t
rans
crip
tR
elat
ive
amou
nt o
f tra
nscr
ipt
(CG
9520
x 10
3/ R
pL32
) (C
G87
08x
103
/ RpL
32)
Fig. 1. Phylogenetic tree of Drosophila, C. elegans, and vertebrate C1β3GalTs and quantitative analyses of the CG9520 and CG8708 transcripts. (A) Dendrogramsshowing the relationship of human, mouse, rat, C. elegans and Drosophila C1β3GalTs, and putative chicken, Xenopus and zebrafish C1β3GalTs. The dendrogramswere constructed using the amino acid sequences with the Clustal X program. The branch length indicates the evolutionary distance between each member. Thescale at the bottom represents evolutionary distance. (B) mRNAs were prepared from embryos and larvae at several developmental stages, from pupae and adultflies (top and bottom left panels) and from tissues and cultured cells (top and bottom right panels). The expression levels of the CG9520 and CG8708 transcriptswere normalized to that of the RpL32 transcript, which was measured in the same cDNAs. Experiments were repeated three times.
P-element (Figure 4G). In contrast, T antigen was not decreasedin CG8708KG05736 embryos (Figure 5D). These results clearlydemonstrated that CG9520 synthesizes T antigen on the CNSin Drosophila embryos, including on hemocytes.
We performed PNA-lectin blot analyses of wild-type andCG9520EY13370 embryo extracts at several developmental stages
(Figure 4H–J). The expression level and pattern of T antigenchanged during embryogenesis. In particular, the zygotic ex-pression of T antigen was strong at late embryogenesis between13 and 19 h (stages 16 and 17) (Figure 4H–J, lane 3). More-over, the expression of T antigen was drastically reduced inCG9520EY13370 embryos (Figure 4H–J, lane 4). These results
1096
by guest on January 14, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from
Drosophila C1β3GalT synthesizes T antigen on the CNS
Fig. 2. T antigen is expressed on the Drosophila CNS during late embryogenesis. Wild-type embryos were stained with an anti-T antigen antibody (green) (A, C,D, F, G, and I) and BP102 mAb, an embryo CNS marker (magenta) (B, C, E, F, H, and I). (A–I) A mid-sagittal plane of focus. Although T antigen expressionbegins at stage 11 in the head and tail regions, the expression of T antigen on the embryonic CNS starts at stage 13. (A–C) Stage 11 embryos, (D–F) stage 13embryos, and (G–I) stage 16 embryos. (C) Merged image of (A) and (B); (F) merged image of (D) and (E); and (I) merged image of (G) and (H). Dorsal is up andanterior is left. Scale bar: 100 µm.
are consistent with the expression pattern of T antigen obtainedfrom the immunohistochemical analyses (Figures 2 and 4D–G)and also strongly support our conclusion that CG9520 synthe-sizes T antigen on the CNS. T antigen was strongly expressedon the CNS in late embryos at stage 16 (Figure 2G and I).We detected six bands carrying T antigen (i–vi) at stages 16and 17 (Figure 4H–J, lane 3). Consequently, the new bands de-tected in late embryos (i–vi) at 173, 151, 119, 113, 59.0, and56.6 kD were good candidates for core-proteins carrying T anti-gen expressed on the CNS.
The above results showed that CG9520 synthesizes Tantigen expressed in Drosophila embryos. Therefore, wehave named the CG9520 gene as Drosophila core 1 β1,3-galactosyltransferase 1 (dC1β3GalT1).
The number of circulating hemocytes is reduced in third instarlarvae of dC1β3GalT1 mutantAlthough T antigen is normally expressed on the Cg25c-positiveembryonic hemocytes (Figure 3A–F), T antigen on hemocytes
was partially reduced in CG9520EY13370 mutant embryo(Figure 4F). To investigate whether T antigen functions inhemocyte development, we carried out immunostaining of em-bryos with 5H7 monoclonal antibody, which recognizes MDP-1/Papilin, one of the components of basement membrane de-posited by hemocytes (Hortsch et al. 1998). In CG9520EY13370
mutant embryo, there were no apparent defects on the stain-ing pattern of hemocytes at stage 11 (Figure 6A and B). It isreported that embryonic hemocytes persist through metamor-phosis and that larval hemocytes are released by larval lymphglands just at the onset of pupation (Holz et al. 2003). To con-firm whether the lack of T antigen affects embryonic hemocytedevelopment by the late third instar larval stage, we counted thenumber of plasmatocytes in CG9520EY13370 mutant larvae. Inembryo, hemocytes are composed of approximately 700 plas-matocytes, which migrate through the embryo, and about 30crystal cells that localized around the proventriculus (Tepasset al. 1994; Lebestky, Chang, et al. 2000). Therefore, most ofT antigen-positive hemocytes are considered as plasmatocytes.
1097
by guest on January 14, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from

H Yoshida et al.
Fig. 3. T antigen is expressed on embryonic hemocytes and S2 cells. A109.1F2 line, hemocyte-specific enhancer trap line embryos (A–F), and S2 cells (G) werestained with an anti-T antigen antibody (green). As β-galactosidase is expressed in hemocyte nuclei in A109.1F2 embryos, we could detect embryonic hemocytesby immunostaining with an anti-β-galactosidase antibody (magenta) (B, C, E, and F). (A–F) Stage 16 embryos, (G) S2 cells. Arrow indicates the focus of Tantigen at the focal contact of two cells. (A–C) A superficial plane of focus, and (D–F) higher magnification of a part of (A), (B), and (C), respectively. (C, F)Merged image of (A) and (B), and of (D) and (E), respectively. Dorsal is up and anterior is left (A–F). Scale bar: 100 µm (A), 10 µm (D and G).
The plasmatocytes in CG9520EY13370 mutant larvae was reducedto 620, while the number of plasmatocytes was 1975 in wild-type larvae, which is in good agreement with the previous study(Lanot et al. 2001) (Figure 6C). These data suggested that thelack of T antigen on the hemocytes results in the reduction oftheir number.
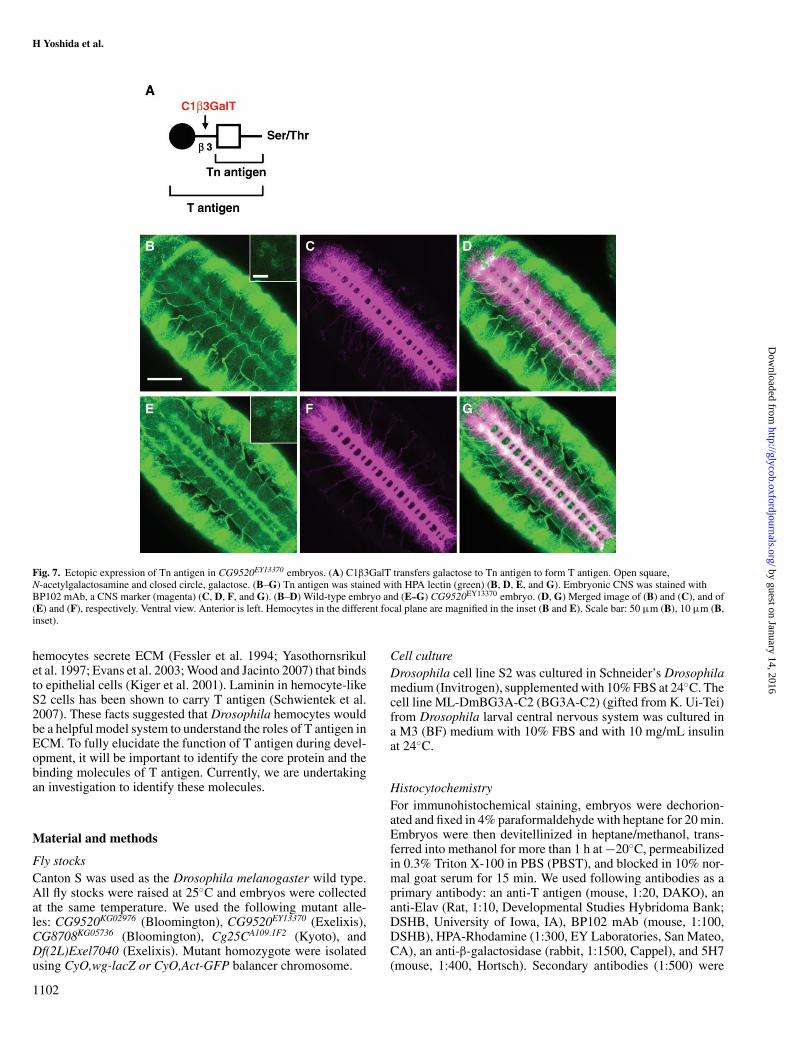
Terminal galactose of T antigen is defective in dC1β3GalT1mutant embryosT antigen is synthesized by C1β3GalT, which transfers Galfrom UDP-Gal to Tn antigen (GalNAcα1-Ser/Thr) (Figure 7A).In wild-type Drosophila embryos, T antigen is expressed onthe CNS and hemocytes (Figure 4D). However, in dC1β3GalT1mutant embryos, T antigen could not be detected on the CNS(Figure 4E). We immunostained CG9520EY13370 embryos withhelix pomatia (HPA) lectin to determine whether Tn antigen,the precursor of T antigen, is expressed on the CNS. We foundthat Tn antigen is weakly expressed on the CNS in wild-typeembryos (Figure 7B). As expected, in dC1β3GalT1 mutant em-bryos, Tn antigen was expressed on the CNS more strongly thanin wild-type embryos (Figure 7E). The same tendency was ob-served on hemocyte-like cells (Figure 7B and E, inset). Theseresults showed that Tn antigens were increased in dC1β3GalT1mutant embryos.
Discussion
In this study, we have demonstrated that T antigen is nor-mally expressed on the embryonic CNS and that dC1β3GalT1synthesizes the antigen in Drosophila melanogaster. In thedC1β3GalT1 mutant, CG9520EY13370, we could not detect Tantigen on the CNS (Figure 4F). This defect was rescued byprecise excision of the P-element inserted into the CG9520
gene (Figure 4G). Moreover, CG9520EY13370 embryos that sur-vived later in development showed a dramatic reduction of Tantigen on their core proteins (Figure 4H–J, lane 4). In addi-tion, the galactosyltransferase activity of CG9520EY13370 thirdinstar larvae was 44% of that of wild type (Figure 4C). TheBerkeley Drosophila Genome Project gene expression databaseindicates that the CG9520 transcripts are expressed in the CNS(http://www.flyexpress.net/), while Muller et al. (2005) did notdetect its transcript in the CNS by in situ hybridization. Dueto the weak mRNA expression on CNS, this might have beenoverlooked. A small amount of mRNA might be enough to syn-thesize T antigens on mucin proteins and glycolipids becauseof high enzymatic activity of dC1β3GalT1 (Muller et al. 2005).Taken together, these data demonstrate that CG9520 synthesizesT antigen during development.
Although T antigen disappeared from the CG9520EY13370
CNS (Figure 4F), there was no significant structural de-fect, even in heterozygous combination with CG9520 defi-cient chromosome Df(2L)Exel7040. The ratios of defectiveembryos in the formation of CNS were as follows: 1.4% ofOregon R (n = 648), 3.9% of Canton S (n = 257), 2.2%of CG9520EY13370/CG9520EY13370 (n = 503), and 5.3% ofCG9520EY13370/Df(2L)Exel7040 (n = 376). These data suggestthat dC1β3GalT1 may have redundant functions or only partlyparticipate in the embryonic CNS development. As proteins orlipids modified by T antigen localize on the surfaces of cells, Tantigen may regulate the binding affinity of adhesion molecules.It is also generally assumed that many null mutants having adefect in a cell adhesion protein do not show obvious pheno-types during embryonic CNS development. Mutations in thearmadillo, Fasciclin 2, Fasciclin 3, Nuerotactin, Neurexin IV,or Neuroglian genes (Patel et al. 1987; Peifer and Wieschaus1990; Baumgartner et al. 1996; Schuster et al. 1996; Speicheret al. 1998) have phenotypes restricted to defasciculation of
1098
by guest on January 14, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from
Drosophila C1β3GalT synthesizes T antigen on the CNS
Fig. 4. dC1β3GalT1 locus and mutant analyses. (A) The exon–intron structure of the dC1β3GalT1 gene. Open boxes indicate noncoding regions. The scale bar is1000 bp. The P-elements of both CG9520EY13370 and CG9520KG02976 are inserted into the first intron of the dC1β3GalT1 gene. The arrow indicates thetranscriptional direction of the genes. Triangles indicate the inserted P-elements. (B) Relative amounts of CG9520 transcript in CG9520KG02976 and CG9520EY13370
third instar larvae. The expression level of the CG9520 transcript in wild type is shown as 100%. Experiments were repeated three times. (C) Galactosyltransferaseactivity of CG9520EY13370 third instar larvae. GalNAc α-pNph was used as the acceptor substrate. (D–G) The distribution of T antigen on stage 16 embryos wasdetermined using an anti-T antigen antibody: wild type (D), CG9520KG02976 (E), CG9520EY13370 (F), and revertant of CG9520KG02976 (G). Dorsal is up andanterior is left. (H–J) PNA lectin blot analyses of wild-type and CG9520EY13370 embryos at various developmental stages: lane 1, wild type at 0–7 h (stages 1–11);lane 2, wild type at 7–13 h (stages 11–15); lane 3, wild type at 13–19 h (stages 16 and 17); and lane 4, CG9520EY13370 at 13–19 h (stages 16 and 17). Candidatecore-proteins carrying T antigens expressed in the Drosophila CNS during late embryogenesis are indicated (i–vi). (H) Total embryonic extracts were subjected to8.5% SDS–PAGE followed by PNA lectin blot analysis. α-Tubulin was used as the internal control. (I) Extension of the region indicated by the single asterisk in(H). Total embryonic extracts were subjected to 6.0% SDS–PAGE followed by PNA lectin blot analysis. (J) A lower exposure image of the region indicated by thedouble asterisks in (H). Scale bar: 100 µm.
particular axons, partial disorganization of major tracts andnerve roots, or other localized adhesion defects. Such local-ized effects might explain why CG9520EY13370 embryos showedno obvious defects in the CNS.
In addition to the embryonic CNS, we found that T anti-gen is expressed on embryonic hemocytes. While it was hardto detect T antigen on the CNS of CG9520EY13370 mutant em-bryos, we were still able to detect T antigen on hemocytes
1099
by guest on January 14, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from

H Yoshida et al.
Fig. 4. (Cont.).
(Figure 4F). The CG9520EY13370 allele resulted frominsertion of a P-element into the first intron of the dC1β3GalT1gene (Figure 4A). CG9520EY13370 is not a null allele(Figure 4B) and the low level of the dC1β3GalT1 transcriptthat it produces may be sufficient for synthesis of T anti-gen on hemocytes, but not on the embryonic CNS. The ac-tivity of the dC1β3GalT1 might be different between accep-tor substrates on hemocytes and on the embryonic CNS. Al-ternatively, it is possible that another C1β3GalT may be in-volved in synthesis of T antigen on hemocytes. One of putativeC1β3GalTs, CG8708, could be a candidate for such synthesisas the CG8708 gene is expressed during embryogenesis (Fig-ure 1B). In order to elucidate the reason why T antigen is ex-pressed on hemocytes of CG9520EY13370 mutant embryos, weare currently attempting to isolate a dC1β3GalT1 null mutantby imprecise excision of the P-element from the CG9520EY13370
allele.We found that T antigen is expressed on S2 cells, which are de-
rived from late-stage embryos and have hemocyte cell-like prop-erties. Previously, it was reported that mucin-D carrying mucin-
type O-glycan is expressed in cell lines including the S2 cell line,embryo, imaginal disc, testis, fat body, and larval brain and is lo-calized at cytoplasmic bridges in various germline and somatictissues (Kramerov et al. 1996, 1997; Kramerova and Kramerov1999; Theopold et al. 2001). Here, we showed the expressionof T antigen in the focal contacts between S2 cells (Figure 3G),although we did not confirm whether the cells in Figure 3G weredividing. Mucin-D localized at a contractile ring might carry Tantigen.
Previous studies showed that embryonic hemocytes have arole during CNS condensation in the embryo (Olofsson and Page2005). We investigated whether T antigen plays a role in embry-onic CNS condensation by examining CG9520EY13370 embryos.In wild-type embryos, condensation results in shortening of theVNC from approximately 80% of embryo length at stage 15 toaround 60% at stage 17 (Olofsson and Page 2005). Althoughsome CG9520EY13370 embryos died during embryogenesis, es-caper CG9520EY13370 embryos showed no observable differencein VNC condensation compared to wild-type embryos (data notshown).
1100
by guest on January 14, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from
Drosophila C1β3GalT synthesizes T antigen on the CNS
Fig. 5. CG8708 locus and mutant analyses. (A) The exon–intron structure of the CG8708 gene. Open boxes indicate noncoding regions. The scale bar is 200 bp. InCG8708KG05736 , the P-element (indicated by a triangle) is inserted into the second exon of the CG8708 gene. The arrow indicates the transcriptional direction ofgene. (B) Relative amount of the CG8708 transcript in CG8708KG05736 third instar larvae. The expression level of the CG8708 transcript in wild type is shown as100%. Experiments were repeated three times. (C and D) T antigen was stained with PNA lectin. Stage 16 embryos: wild type (C), CG8708KG05736 (D). Dorsal isup and anterior is left. Scale bar: 100 µm.
Fig. 6. Number of hemocytes in wild type and CG9520EY13370. (A, B) 5H7stained stage 11 embryos. (A) Wild-type embryo and (B) CG9520EY13370
embryo. (C) Number of circulating plasmatocytes per third instar larva (n =20). The means of wild-type and CG9520EY13370 were 1975 (SD 1858) and620 (SD 610), respectively. There is a significant difference between eachgenotype (t-test, P = 0.005).
In CG9520EY13370 mutant, we could not detect obvious differ-ence in the hemocyte development at stage 11 (Figure 6A and B).In contrast, the number of hemocytes was 620, which is small
compared to that of wild type, at the third instar larvae stage(Figure 6C). It is believed that the most of embryonic hemo-cytes derive exclusively from procephalic and gnathal meso-derm, which proliferate up to about 700 cells by stage 11 andthat this number remains constant throughout embryogenesis(Tepass et al. 1994). In addition, it is revealed that the num-ber of hemocytes is progressively increased during the larvalstage by postembryonic proliferation of embryonic hemocytes(Lanot et al. 2001; Holz et al. 2003). Taken together, thesedata suggested that the lack of T antigen on hemocytes leadsto impede the proliferation or to induce the apoptosis of hemo-cytes. These facts remind us that both of the thrombocytopeniain C1galt1 mice (Alexander et al. 2006) and Tn-syndrome inhuman (Berger 1999) exhibit the reduction of platelets. On theother hand, Drosophila hemocytes are the macrophage-like cellsresponsible for the disposal of apoptotic cells and invading mi-croorganisms (Franc et al. 1996, 1999). Our findings indicatethat dC1β3GalT1 mutant may have some defects in immuneresponse.
Recently, Xia et al. (2004) suggested that capillary fragilityin T-synthase knockout mouse embryos is a consequence ofthe separation of endothelial cells from supporting pericytesand extracellular matrix. It is also reported that Drosophila
1101
by guest on January 14, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from
H Yoshida et al.
Fig. 7. Ectopic expression of Tn antigen in CG9520EY13370 embryos. (A) C1β3GalT transfers galactose to Tn antigen to form T antigen. Open square,N-acetylgalactosamine and closed circle, galactose. (B–G) Tn antigen was stained with HPA lectin (green) (B, D, E, and G). Embryonic CNS was stained withBP102 mAb, a CNS marker (magenta) (C, D, F, and G). (B–D) Wild-type embryo and (E–G) CG9520EY13370 embryo. (D, G) Merged image of (B) and (C), and of(E) and (F), respectively. Ventral view. Anterior is left. Hemocytes in the different focal plane are magnified in the inset (B and E). Scale bar: 50 µm (B), 10 µm (B,inset).
hemocytes secrete ECM (Fessler et al. 1994; Yasothornsrikulet al. 1997; Evans et al. 2003; Wood and Jacinto 2007) that bindsto epithelial cells (Kiger et al. 2001). Laminin in hemocyte-likeS2 cells has been shown to carry T antigen (Schwientek et al.2007). These facts suggested that Drosophila hemocytes wouldbe a helpful model system to understand the roles of T antigen inECM. To fully elucidate the function of T antigen during devel-opment, it will be important to identify the core protein and thebinding molecules of T antigen. Currently, we are undertakingan investigation to identify these molecules.
Material and methods
Fly stocksCanton S was used as the Drosophila melanogaster wild type.All fly stocks were raised at 25◦C and embryos were collectedat the same temperature. We used the following mutant alle-les: CG9520KG02976 (Bloomington), CG9520EY13370 (Exelixis),CG8708KG05736 (Bloomington), Cg25CA109.1F2 (Kyoto), andDf(2L)Exel7040 (Exelixis). Mutant homozygote were isolatedusing CyO,wg-lacZ or CyO,Act-GFP balancer chromosome.
Cell cultureDrosophila cell line S2 was cultured in Schneider’s Drosophilamedium (Invitrogen), supplemented with 10% FBS at 24◦C. Thecell line ML-DmBG3A-C2 (BG3A-C2) (gifted from K. Ui-Tei)from Drosophila larval central nervous system was cultured ina M3 (BF) medium with 10% FBS and with 10 mg/mL insulinat 24◦C.
HistocytochemistryFor immunohistochemical staining, embryos were dechorion-ated and fixed in 4% paraformaldehyde with heptane for 20 min.Embryos were then devitellinized in heptane/methanol, trans-ferred into methanol for more than 1 h at −20◦C, permeabilizedin 0.3% Triton X-100 in PBS (PBST), and blocked in 10% nor-mal goat serum for 15 min. We used following antibodies as aprimary antibody: an anti-T antigen (mouse, 1:20, DAKO), ananti-Elav (Rat, 1:10, Developmental Studies Hybridoma Bank;DSHB, University of Iowa, IA), BP102 mAb (mouse, 1:100,DSHB), HPA-Rhodamine (1:300, EY Laboratories, San Mateo,CA), an anti-β-galactosidase (rabbit, 1:1500, Cappel), and 5H7(mouse, 1:400, Hortsch). Secondary antibodies (1:500) were
1102
by guest on January 14, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from

Drosophila C1β3GalT synthesizes T antigen on the CNS
obtained from Molecular Probes. FITC-PNA (1:300, Seikagaku,Tokyo, Japan) was also used for the staining of T antigen.
S2 cells were subcultured at a density of 1.0 × 105 cells/mLonto a chamber slide (Nunc) in Schneider’s Drosophila mediumcontaining 10% FBS. After 48 h, the cells were washed withPBS and fixed with 4% paraformaldehyde for 10 min. The cellswere then washed and permeabilized with PBST, blocked in10% normal goat serum in PBST for 20 min, and incubatedwith primary antibody for 16 h at 4◦C. After being washed, thecells were incubated with secondary antibody for 1 h at roomtemperature. Confocal images were taken on a LSM 5 Pascal(Carl Zeiss).
Quantitative analysis of the CG9520 and CG8708 transcriptsby real-time PCRTotal RNA was extracted from CG9520EY13370, CG9520KG02976,and CG8708KG05736 third instar larvae and from wild-type tissues throughout development. Total RNA wasalso prepared from S2 and BG3A-C2 cells. First-strandcDNA was synthesized using RevaTra Dash (TOYOBO,Osaka, Japan). The following gene-specific primers wereused: CG9520 forward, 5′-GCCTGGCGGAGCTGTTC-3′;reverse, 5′-GGCCATCGTATGGCATGAA-3′; probe, 5′-CTACTCCACGCCGGAGCGAAGTGA-3′ and CG8708forward, 5′-GCATGATTACGACAGGATCTATACGA-3′;reverse, 5′-GGAGTCCATTTCCCGAGCAT-3′; probe, 5′-ACGACCGAAAAGCCAAAGGAGCCC-3′. The amountof Ribosomal protein L32 (RpL32) mRNA in eachcDNA sample was used to normalize the efficiency ofcDNA preparation. The following primers were used:RpL32 forward, 5′-GCAAGCCCAAGGGTATCGA-3′; re-verse, 5′-CGATGTTGGGCATCAGATACTG-3′; probe,5′-AACAGAGTGCGTCGCCGCTTCA-3′. The probes werelabeled at the 5′-end with the reporter dye, 3FAM, and at the3′-end with the quencher dye TAMRA (Nippon EGT, Toyama,Japan). Amplifications were performed using 40 cycles of94◦C for 30 s and 60◦C for 4 min, with an ABI PRISM 7700Sequence Detection System (Applied Biosystems).
Lectin blot analysisWild-type and CG9520EY13370 embryos were collected, dechori-onated, and then homogenized in 50 mM Tris–HCl (pH 7.5),5 mM MgCl2, 150 mM NaCl, 10% glycerol, 0.1% TritonX-100, 0.1% sodium deoxycholate, 5 mM 2-mercaptoethanolwith the protease inhibitors (1 mM PMSF, 10 µg/mL aprotinin,10 µg/mL pepstatin A, 10 µg/mL leupeptin, 1 µg/mL antipain).The supernatant was obtained by centrifugation at 10,000 × g for10 min and used as the total embryonic extracts. These extracts(3 µg per lane) were subjected to 8.5% and 6.0% sodium dodecylsulfate–polyacrylamide gel electrophoresis (SDS–PAGE) fol-lowed by lectin blot analysis. The separated proteins were trans-ferred to an Immobiron-P membrane (Millipore). The mem-brane was probed with horseradish peroxidase-conjugated PNAlectin (Seikagaku) or an anti-α-tubulin antibody and visualizedwith the ECL plus western blotting detection kit (GE HealthcareBio-Science) according to the manufacturer’s instructions.
Assay of galactosyltransferase activityThird instar larvae were homogenized in 100 mM MES(pH 6.8), 2 mM ATP, 20 mM MnCl2, 0.2% Triton X-100
with the protease inhibitors (1 mM PMSF, 10 µg/mL pep-statin A, 1 µg/mL leupeptin, 2 mM benzamidine) (100 µLfor every 10 larvae). The supernatant was obtained by cen-trifugation at 10,000 × g for 10 min and used as the lar-val extracts. Uridine diphosphate-[3H]galactose (UDP-[3H]Gal)(20 Ci/mmol) was supplied by American Radiolabeled Chem-icals Inc. p-Nitrophenyl-N-acetyl-α-galactosaminide (GalNAcα-pNph) was purchased from Calbiochem. An assay of galac-tosyltransferase activity was performed referring to Cummings’paper (Ju, Cummings et al. 2002). The reaction mixture con-tained 15 µg of larvae extracts, 100 mM MES (pH 6.8), 2 mMATP, 20 mM MnCl2, 0.2% triton X-100, 1 mM GalNAc α-pNph,0.4 mM UDP-Gal (including 1.25 µM UDP-[3H]Gal) in volumeof 20 µL. After incubation at 25◦C for 2.5 h, the reaction wasterminated with the addition of 500 µL of water. The reactionmixture was applied to a Sep-Pak C18 cartridges and unreactedUDP-[3H]Gal was washed out with water. The products on thecolumn were eluted with methanol and measured the radioac-tivity using a liquid scintillation counter.
Collection and counting of circulating hemocytesWe drew upon the experimental methodology of Zettervallet al. (2004). Wandering late third instar larvae were washedin PBS, and then bled with fine scissors in 20 µL of PBS for30 s on a siliconized glass plate. Hemocytes-containing PBSwas then loaded onto an improved Neubauer hemocytometer(Digital Bio, Seoul, Korea) by a siliconized tip. Twenty larvaeof each genotype were counted. Student’s t-test was performedwith the statistical package R (http://www.r-project.org/).
Funding
Core Research for Evolutional Science and Technology(CREST) of Japan Science and Technology Agency (JST).“High-Tech Research Center” Project for Private Universities:matching fund subsidy from MEXT (Ministry of Education,Culture, Sports, Science and Technology), 2002–2006.
Acknowledgements
We thank Bloomington Stock Center, Exelixis and DrosophilaGenetic Resource Center in Kyoto Institute of Technology forthe fly strains and Dr. Nakata H., Dr. Hortsch M., and Develop-mental Studies Hybridoma Bank, Iowa University, for antibod-ies.
Conflict of interest statement
None declared.
Abbreviations
C1β3GalT1, core 1 β1,3-galactosyltransferase; CNS, centralnervous system; ECM, extracellular matrix; GalNAcα-pNph,p-nitrophenyl-N-acetyl-α-galactosaminide; HPA, helix poma-tia; PNA, peanut agglutinin; RpL32, Ribosomal protein L32;SDS–PAGE, sodium dodecyl sulfate–polyacrylamide gel elec-trophoresis; VNC, ventral nerve code.
1103
by guest on January 14, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from

H Yoshida et al.
References
Alexander WS, Viney EM, Zhang JG, Metcalf D, Kauppi M, Hyland CD,Carpinelli MR, Stevenson W, Croker BA, Hilton AA, et al. 2006. Thrombo-cytopenia and kidney disease in mice with a mutation in the C1galt1 gene.Proc Natl Acad Sci USA. 103:16442–16447.
Baumgartner S, Littleton JT, Broadie K, Bhat MA, Harbecke R, Lengyel JA,Chiquet-Ehrismann R, Prokop A, Bellen HJ. 1996. A Drosophila neurexin isrequired for septate junction and blood-nerve barrier formation and function.Cell. 87:1059–1068.
Bellen HJ, O’Kane CJ, Wilson C, Grossniklaus U, Pearson RK, Gehring WJ.1989. P-element-mediated enhancer detection: A versatile method to studydevelopment in Drosophila. Genes Dev. 3:1288–1300.
Berger EG. 1999. Tn-syndrome. Biochim Biophys Acta. 1455:255–268.Brockhausen I. 1997. Biosynthesis and functions of O-glycans and regulation
of mucin antigen expression in cancer. Biochem Soc Trans. 25:871–874.Brockhausen I. 1999. Pathways of O-glycan biosynthesis in cancer cells.
Biochim Biophys Acta. 1473:67–95.Cho NK, Keyes L, Johnson E, Heller J, Ryner L, Karim F, Krasnow MA. 2002.
Developmental control of blood cell migration by the Drosophila VEGFpathway. Cell. 108:865–876.
D’Amico P, Jacobs JR. 1995. Lectin histochemistry of the Drosophila embryo.Tissue Cell. 27:23–30.
Evans CJ, Hartenstein V, Banerjee U. 2003. Thicker than blood: Con-served mechanisms in Drosophila and vertebrate hematopoiesis. Dev Cell.5:673–690.
Fessler LI, Nelson RE, Fessler JH. 1994. Drosophila extracellular matrix. Meth-ods Enzymol. 245:271–294.
Franc NC, Dimarcq JL, Lagueux M, Hoffmann J, Ezekowitz RA. 1996. Cro-quemort, a novel Drosophila hemocyte/macrophage receptor that recognizesapoptotic cells. Immunity. 4:431–443.
Franc NC, Heitzler P, Ezekowitz RA, White K. 1999. Requirement forcroquemort in phagocytosis of apoptotic cells in Drosophila. Science.284:1991–1994.
Fristrom DK, Fristrom JW. 1982. Cell surface binding sites for peanut agglutininin the differentiating eye disc of Drosophila. Dev Biol. 92:418–427.
Holz A, Bossinger B, Strasser T, Janning W, Klapper R. 2003. The two originsof hemocytes in Drosophila. Development. 130:4955–4962.
Hortsch M, Olson A, Fishman S, Soneral SN, Marikar Y, Dong R, Jacobs JR.1998. The expression of MDP-1, a component of Drosophila embryonicbasement membranes, is modulated by apoptotic cell death. Int J Dev Biol.42:33–42.
Ju T, Brewer K, D’Souza A, Cummings RD, Canfield WM. 2002. Cloningand expression of human core 1 beta1,3-galactosyltransferase. J Biol Chem.277:178–186.
Ju T, Cummings RD, Canfield WM. 2002. Purification, characterization, andsubunit structure of rat core 1 Beta1,3-galactosyltransferase. J Biol Chem.277:169–177.
Ju T, Zheng Q, Cummings RD. 2006. Identification of core 1 O-glycan T-synthase from Caenorhabditis elegans. Glycobiology. 16:947–958.
Kiger JA, Natzle JE, Green MM. 2001. Hemocytes are essential for wing matura-tion in Drosophila melanogaster. Proc Natl Acad Sci USA. 98:10190–10195.
Koles K, Irvine KD, Panin VM. 2004. Functional characterization of Drosophilasialyltransferase. J Biol Chem. 279:4346–4357.
Kramerov AA, Arbatsky NP, Rozovsky YM, Mikhaleva EA, Polesskaya OO,Gvozdev VA, Shibaev VN. 1996. Mucin-type glycoprotein from Drosophilamelanogaster embryonic cells: Characterization of carbohydrate component.FEBS Lett. 378:213–218.
Kramerov AA, Mikhaleva EA, Rozovsky M, Pochechueva TV, Baikova NA,Arsenjeva EL, Gvozdev VA. 1997. Insect mucin-type glycoprotein: Immun-odetection of the O-glycosylated epitope in Drosophila melanogaster cellsand tissues. Insect Biochem Mol Biol. 27:513–521.
Kramerova IA, Kramerov AA. 1999. Mucinoprotein is a universal constituentof stable intercellular bridges in Drosophila melanogaster germ line andsomatic cells. Dev Dyn. 216:349–360.
Lanot R, Zachary D, Holder F, Meister M. 2001. Postembryonic hematopoiesisin Drosophila. Dev Biol. 230:243–257.
Lebestky T, Chang T, Hartenstein V, Banerjee U. 2000. Specification ofDrosophila hematopoietic lineage by conserved transcription factors. Sci-ence. 288:146–149.
Montell DJ, Goodman CS. 1989. Drosophila laminin: sequence of B2 subunitand expression of all three subunits during embryogenesis. J Cell Biol.109:2441–2453.
Muller R, Hulsmeier AJ, Altmann F, Ten Hagen KG, Tiemeyer M, Hennet T.2005. Characterization of mucin-type core-1 beta1-3 galactosyltransferasehomologous enzymes in Drosophila melanogaster. FEBS J. 272:4295–4305.
Olofsson B, Page DT. 2005. Condensation of the central nervous system inembryonic Drosophila is inhibited by blocking hemocyte migration or neuralactivity. Dev Biol. 279:233–243.
Pace KE, Lebestky T, Hummel T, Arnoux P, Kwan K, Baum LG. 2002. Charac-terization of a novel Drosophila melanogaster galectin. Expression in devel-oping immune, neural, and muscle tissues. J Biol Chem. 277:13091–13098.
Patel NH, Snow PM, Goodman CS. 1987. Characterization and cloning offasciclin III: A glycoprotein expressed on a subset of neurons and axonpathways in Drosophila. Cell. 48:975–988.
Peifer M, Wieschaus E. 1990 The segment polarity gene armadillo encodesa functionally modular protein that is the Drosophila homolog of humanplakoglobin. Cell. 63:1167–1176.
Ramet M, Pearson A, Manfruelli P, Li X, Koziel H, Gobel V, Chung E, KriegerM, Ezekowitz RA. 2001. Drosophila scavenger receptor CI is a patternrecognition receptor for bacteria. Immunity. 15:1027–1038.
Richter DU, Jeschke U, Makovitzky J, Goletz S, Karsten U, Briese V, Friese K.2000. Expression of the Thomsen–Friedenreich (TF) antigen in the humanplacenta. Anticancer Res. 20:5129–5133.
Schneider I. 1972. Cell lines derived from late embryonic stages of Drosophilamelanogaster. J Embryol Exp Morphol. 27:353–365.
Schuster CM, Davis GW, Fetter RD, Goodman CS. 1996. Genetic dissection ofstructural and functional components of synaptic plasticity II: Fasciclin IIcontrols presynaptic structural plasticity. Neuron. 17:655–667.
Schwientek T, Mandel U, Roth U, Muller S, Hanisch FG. 2007. A serial lectinapproach to the mucin-type O-glycoproteome of Drosophila melanogasterS2 cells. Proteomics. 7:3264–3277.
Sears HC, Kennedy CJ, Garrity PA. 2003. Macrophage-mediated corpse engulf-ment is required for normal Drosophila CNS morphogenesis. Development.130:3557–3565.
Speicher S, Garcia-Alonso L, Carmena A, Martin-Bermudo MD, De La EscaleraS, Jimenez F. 1998. Neurotactin functions in concert with other identifiedCAMs in growth cone guidance in Drosophila. Neuron. 20:221–233.
Springer TA, Thompson WS, Miller LJ, Schmalstieg FC, Anderson DC. 1984.Inherited deficiency of the Mac-1, LFA-1, p150,95 glycoprotein family andits molecular basis. J Exp Med. 160:1901–1918.
Tepass U, Fessler LI, Aziz A, Hartenstein V. 1994. Embryonic origin ofhemocytes and their relationship to cell death in Drosophila. Development.120:1829–1837.
Theopold U, Dorian C, Schmidt O. 2001. Changes in glycosylation duringDrosophila development. The influence of ecdysone on hemomucin iso-forms. Insect Biochem Mol Biol. 31:189–197.
Toma V, Zuber C, Sata T, Kmminoth P, Hailemariam S, Eble JN, Heitz PU,Roth J. 2000. Thomsen–Friedenreich glycotope is expressed in developingand normal kidney but not in renal neoplasms. Hum Pathol. 31:647–655.
Van Den Steen P, Rudd PM, Dwek RA, Opdenakker G. 1998. Concepts and prin-ciples of O-linked glycosylation. Crit Rev Biochem Mol Biol. 33:151–208.
Wilson C, Pearson RK, Bellen HJ, O’Kane CJ, Grossniklaus U, Gehring WJ.1989. P-element-mediated enhancer detection: An efficient method for iso-lating and characterizing developmentally regulated genes in Drosophila.Genes Dev. 3:1301–1313.
Wood W, Jacinto A. 2007. Drosophila melanogaster embryonic haemocytes:Masters of multitasking. Nat Rev Mol Cell Biol. 8:542–551.
Xia L, Ju T, Westmuckett A, An G, Ivanciu L, McDaniel JM, Lupu F, CummingsRD, Mcever RP. 2004. Defective angiogenesis and fatal embryonic hemor-rhage in mice lacking core 1-derived O-glycans. J Cell Biol. 164:451–459.
Xia L, McEver RP. 2006. Targeted disruption of the gene encoding core 1beta1-3-galactosyltransferase (T-synthase) causes embryonic lethality anddefective angiogenesis in mice. Methods Enzymol. 416:314–331.
Yamaguchi Y, Otsuka K, Habu Y, Ohyama K, Kurotsu T, Ookawara T,Ohkuma S, Yamakawa T, Suzuki K. 2001. Presence of Vicia gramineaor Vicia unijuga lectin-binding (Vgu) glycoproteins with and withoutThomsen–Friedenreich (T) antigen in normal human seminal plasma. ResCommun Mol Pathol Pharmacol. 110:293–302.
Yasothornsrikul S, Davis WJ, Cramer G, Kimbrell DA, Dearolf CR. 1997.Viking: Identification and characterization of a second type IV collagen inDrosophila. Gene. 198:17–25.
Zettervall CJ, Anderl I, Williams MJ, Palmer R, Kurucz E, Ando I, HultmarkD. 2004. A directed screen for genes involved in Drosophila blood cellactivation. Proc Natl Acad Sci USA. 101:14192–14197.
Zhou Q, Cummings RD. 1993. L-14 lectin recognition of laminin and its pro-motion of in vitro cell adhesion. Arch Biochem Biophys. 300:6–17.
1104
by guest on January 14, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from
Copyright © 2022 FDOKUMEN












![4,4′-Difluoro-2,2′-{[(3a RS ,7a RS )-2,3,3a,4,5,6,7,7a-octahydro-1 H -1,3-benzimidazole-1,3-diyl]bis(methylene)]}diphenol](https://static.fdokumen.com/doc/165x107/63258a217fd2bfd0cb03842e/44-difluoro-22-3a-rs-7a-rs-233a45677a-octahydro-1-h-13-benzimidazole-13-diylbismethylenediphenol.jpg)


![Competing Regiodirecting Effects of Ester and Aryl Groups in [3+3] Cyclocondensations of 1,3-Bis(trimethylsilyloxy)-1,3-butadienes: Regioselective Synthesis of 3-Hydroxyphthalates](https://static.fdokumen.com/doc/165x107/6323a23503238a9ff60a8549/competing-regiodirecting-effects-of-ester-and-aryl-groups-in-33-cyclocondensations.jpg)

![ChemInform Abstract: Competing Regiodirecting Effects of Ester and Aryl Groups in [3 + 3] Cyclocondensations of 1,3-Bis(trimethylsilyloxy)-1,3-butadienes: Regioselective Synthesis](https://static.fdokumen.com/doc/165x107/6323a25a4d8439cb620d0030/cheminform-abstract-competing-regiodirecting-effects-of-ester-and-aryl-groups-in.jpg)

![Synthesis of Pyrrolo[1,3]-Diazepines and Potential Poxvirus ...](https://static.fdokumen.com/doc/165x107/63286568051fac18490eb53f/synthesis-of-pyrrolo13-diazepines-and-potential-poxvirus-.jpg)
