
Investigation of the flow field downstream of an artificial heart valve by means of PIV and PTV
Upload
independentCategory
view
1download
0
Lack of CIITA Expression Is Central to theAbsence of Antigen Presentation Functionsof Trophoblast Cells and Is Caused byMethylation of the IFN-g InduciblePromoter (PIV) of CIITA
Peter J. van den Elsen, Nienke van der Stoep,Henk E. Vietor, Louis Wilson, Marlijn van Zutphen,and Sam J. P. Gobin
ABSTRACT: Lack of MHC-mediated antigen presentingfunctions of fetal trophoblast cells is an important mecha-nism to evade maternal immune recognition. In this studywe demonstrated that the deficiency in MHC expressionand antigen presentation in the trophoblast cell lines JEG-3and JAR is caused by lack of class II transactivator (CIITA)expression due to hypermethylation of its interferon-g (IFN-g)-responsive promoter (PIV). Circumvention of this lack ofCIITA expression by introduction of exogenous CIITA in-duced cell surface expression of HLA-DR, -DP, and -DQ,leading to an acquired capacity to present antigen to antigen-specific T cells. Transfection of CIITA in JEG-3 cells also
upregulated functional HLA-B and HLA-C expression. Note-worthy, this lack of IFN-g-mediated induction of CIITAwas also found to exist in normal trophoblast cells expandedfrom chorionic villus biopsies. Together, these observationsdemonstrate that lack of CIITA expression is central to theabsence of antigen presentation functions of trophoblastcells. Human Immunology 61, 850–862 (2000). © Amer-ican Society for Histocompatibility and Immunogenetics,2000. Published by Elsevier Science Inc.
KEYWORDS: transcription; MHC; CIITA; trophoblastcells; IFN-g
ABBREVIATIONSbp base pairsCIITA class II transactivatorEBV Epstein Barr virus
IFN-g interferon-gmAb monoclonal antibodyPBMC peripheral blood mononuclear cellsRFX regulatory factor X
INTRODUCTIONExpression of MHC genes is tightly controlled duringpregnancy, especially in trophoblast cells of the placenta,which are in direct contact with the maternal immunesystem. As a result of the haplo-disparity between the
mother and the developing fetus, successful placentationis dependent on the trophoblast that forms a barrier forputative destruction by the maternal immune system. Inthis regard, the lack of expression of MHC class IImolecules on trophoblast cells prevents initiation of amaternal alloimmune response, whereas lack of HLA-Aand HLA-B expression impairs the effector phase of thealloimmune response [1, 2]. Expression of HLA-G andHLA-C is thought to protect trophoblast cells fromdestruction by decidual natural killer cells in this other-wise MHC class I-deficient environment [1–3].
The exact immunological mechanisms by which themammalian mother accepts the implanting fetus as a
From the Department of Immunohematology and Blood Transfusion,Leiden University Medical Center, Leiden, The Netherlands (P.J.V.D.E.,N.V.D.S., L.W., M.V.Z., S.J.P.G.); and Department of Clinical Genetics,University Hospital Nijmegen, Nijmegen, The Netherlands (H.E.V.).
Address reprint requests to: Dr. Peter J. Van den Elsen, Division ofMolecular Biology, Department of Immunohematology and Blood Transfu-sion, Building 1, E3-Q, Leiden University Medical Center, Albinusdreef 2,POBox 9600, 2300 RC Leiden, The Netherlands; Tel: 31-71-5263831;Fax: 31-71-5216751; E-Mail: [email protected].
Received and accepted May 19, 2000.
Human Immunology 61, 850–862 (2000)0198-8859/00/$–see front matter© American Society for Histocompatibility and Immunogenetics, 2000
Published by Elsevier Science Inc. PII S0198-8859(00)00159-2

semi-allograft remain to be elucidated. These mecha-nisms may include secretion of soluble factors such asinterleukin-10 and transforming growth factor-b, whichdisplay an immunosuppressive effect [4] and down-mod-ulation of MHC class II and class I expression. Previ-ously, it has been shown that trophoblast cells have thecapacity to present peptides through MHC class I [1, 2].This finding has been corroborated by the observationthat HLA-G associates with the TAP-complex [5, 6] andthat peptides can be eluted from its peptide bindinggroove [7, 8]. In addition, trophoblast cells transfectedwith HLA-A*0201 can present a flu-virus-encoded pep-tide after virus infection [6]. Together, these observa-tions argue that trophoblast cells have the capacity topresent peptide, provided that the genes encoding MHCclass I heavy chains are expressed.
Several conserved DNA elements in the proximalpromoter of MHC class I and II genes mediate thecontrol of constitutive and inducible expression [9]. Re-cently, it has been demonstrated that the promoters ofMHC class I and class II genes share several of theseelements, which act in concert and therefore can beviewed as a regulatory module [10, 11]. This regulatorymodule consists of S-, X- (comprising the X1 and X2sites), and Y-box elements. The SXY regulatory modulealso exists in the promoters of the b2m, invariant chain(Ii), and HLA-DM genes [10]. A number of DNA bind-ing protein complexes, which act through the commonSXY regulatory module, was found to be critical forMHC expression. The RFX complex (consisting ofRFX5, RFXAP, and RFX-B/RFXANK) binds to theX1-box of the SXY module [12–15], and is crucial forMHC class II and class I expression as is clearly illus-trated in patients suffering from bare lymphocyte syn-drome [10, 11]. The subunits of the RFX complex areexpressed ubiquitously. The X2-box is bound by CREB/ATF [11, 16]. The Y-box is the target sequence forNF-Y [17, 18]. Together, the RFX, CREB/ATF andNF-Y protein complexes form a highly stable multipro-tein complex on XY DNA [19–22, Gobin et al., unpub-lished]. For MHC class II, the higher order multiproteincomplex is not sufficient for this route of induction ofMHC class II gene expression, but needs the activity ofa coactivator, the MHC class II transactivator, CIITA[23–25]. For MHC class I and b2m, CIITA fulfills anancillary function in the transcriptional control of thesegenes [11, 26, 27]. CIITA is only constitutively ex-pressed in antigen-presenting cells of the immune systemand can be induced by IFN-g in other cell types [28–30]. Transcription of CIITA is directed by several prox-imal promoters, of which the principal IFN-g responsivepromoter is PIV, and is activated predominantly in cellsthat lack MHC class II expression [31–33]. The PIIIpromoter is predominantly used in B cells for the con-
stitutive expression of CIITA and subsequent MHC classII expression [33, 34].
In this study we investigated whether the lack ofMHC expression in trophoblast cells correlates with theabsence of key factors in MHC transcription. We showedthat the absence of MHC class II expression is caused bylack of CIITA expression and is due to hypermethylationof the PIV promoter of CIITA. Introduction of exoge-nous CIITA could reconstitute cell surface expression ofMHC class II molecules in both trophoblast cell lines. Inaddition, introduction of exogenous CIITA in JEG-3cells also resulted in induction of functional HLA-B andHLA-C cell surface expression. It was also demonstratedthat normal trophoblast cells lack IFN-g-mediated in-duction of CIITA.
MATERIALSCell Lines and CellsThe trophoblast cell lines JEG-3 (ATCC HTB36) andJAR (ATCC HTB144) were obtained from the AmericanType Culture Collection (ATCC; Rockville, MD). Theother cell lines used in this study were: HeLa (cervicalcarcinoma, ATCC CCL2), K562 (erythroleukemia), WSI(dermal fibroblast, 11), Tera-2 (teratocarcinoma, ATCCHTB106), U373 (glioblastoma, ATCC HTB 17), SB (Bcell, ATCC CCRF), and the EBV-transformed B-celllines EKR, QBL, VLIET, MIR/S, JVM, APD, and HUISthat were established from PBMC of healthy donors.PBMC of donors GW and TRI were used as respondercells. The HLA types of the various cell lines and re-sponder cells used in this study are depicted in Table 1.The HLA genotype was determined by serology andDNA typing. The CTL clones used in this study wereobtained from Dr. E. Roosnek [35] and their specificitywas determined to be HLA-Cw4 (RN14) and HLA-A24
TABLE 1 HLA phenotype of responder and targetcells used in this study
Responders:TRI: HLA-A24, A11, B7, B8, DR17, DR4, DQ2, DQ7GW: HLA-A24, B18, B27, DR12, DR13, DQ7, DQ6
Target cells:QBL: HLA-A26, B18, Cw5, DR17, DQ2SB: HLA-A1, A2, B44, B57, DR15, DR13, DQ6VLIET: HLA-A1, A2, B8, B60, Cw3, Cw7, DR17, DR4MIR/S: HLA-A2, A11, B8, B57, Cw6, Cw7, DR17, DR7JVM: HLA-A2, B18, Cw7, DR11APD: A1, B60, DR13HUIS: A3, B7, B35, Cw4, Cw7, DR1, DR13JEG-3: HLA-A1, A11, B8, B35, Cw4, Cw7, G, DR3, DR103JAR: HLA-A3, A30, B7, B47, Cw6, H-DR7, DR13Tera-2: HLA-A1, B8, Cw7, DR17K562: HLA-A11, B18, Cw5, DR17
851CIITA-Mediated Induction of MHC Expression in Trophoblast Cells

(RN16). Cells were cultured in Iscove’s modified DMEM(Life Technologies, Paisley, Scotland), supplementedwith 10% (v/v) heat-inactivated fetal calf serum (LifeTechnologies), penicillin (100 U/ml), and streptomycin(100 mg/ml). For MHC induction experiments cellstreated with 500 units/ml of IFN-g (Boehringer In-gelheim, Alkmaar, The Netherlands) for 48 h.
Transabdominal chorionic villus biopsy was per-formed under ultrasound guidance. Fetal chorionic villiwere separated and identified manually using microscopyand morphological differences between maternal and fe-tal cells. After separation, more than 95% of the chori-onic villi samples were found to contain trophoblastcells. The cells were subsequently cultured in eitherChang or a-MEM medium.
Fluorescent-Activated Cell Sorter AnalysisCells from subconfluent cultures were harvested fol-
lowing trypsinization and subsequently were stained byindirect immunofluorescence. The mAb W6/32 was usedto detect HLA class I expression. HLA-DR was detectedwith the B8.11.2 mAb [36], HLA-DQ with the B7/21mAb [37], and HLA-DP with the SPV-L3 mAb [38].Costimulatory molecules were detected with anti-CD80(B7-1) and CD86 (B7-2) (Becton-Dickinson, MountainView, CA), whereas cell adhesion molecules were detectedwith anti-CD54 (LFA-3) and CD58 (ICAM) (Becton Dick-inson). Fluorescein isothiocyanate (FITC)- and/or R-phy-coerythrin (PE)-conjugated anti-mouse IgG was used asthe second antibody. The acquisition was performed on aFACScan, using the CELL Quest program (Becton Dick-inson) for analysis.
Generation of CTL LinesPBMCs were isolated from peripheral blood obtainedfrom HLA-typed normal healthy volunteers. Mixed cellcultures were initiated by stimulating of 5 3 106 PBMCwith 2.5 3 106 each of the JEG-3/CIITA transfectants(5,000 rads irradiated) in RPMI containing 10% humanAB serum. CTL lines were generated by weekly stimu-lation of responder PBMCs with a feeder cell mixturecomprising irradiated allogeneic PBMCs (3,000 rads)and irradiated transfectants (5,000 rads), in RPMI con-taining 10% human AB serum and exogenous rIL-2 (20U/ml).
Cellular AssaysProliferation was measured in a standard [3H]thymidineincorporation assay to analyze MHC class II-mediatedantigen presentation functions of CIITA-transfectedJEG-3 cells. A total of 2 3 106 cells irradiated (5,000rad) vector- or CIITA-transfected JEG-3 cells weremixed with 105 T cells of the tetanus-toxoid specificT-cell line RST-TT (HLA-DR3 restricted) in the pres-
ence or absence of 1.9 lf/ml tetanus toxoid (RIVM,Bilthoven, The Netherlands). The EBV-transformed B-cell lines QBL and SB served as control. The stimulationindex was determined as [3H] thymidine incorporationby T cells in the presence of JEG-3 cells with tetanustoxoid divided by [3H]thymidine incorporation by Tcells in the presence of JEG-3 cells only.
Then, 106 vector- or CIITA-transfected JEG-3 cellswere loaded with 100 mCi of Na51CrO4 (Dupont-NEN,Boston, MA) for 1 h at 37°C to be used as target cells forthe allospecific CTL clones RN14 (anti HLA-Cw4) andRN16 (anti HLA-A24). Cell lysis was assessed for 16 hwith effector/target cell ratios at 40:1 [6]. The degree ofcytotoxicity was determined by chromium release in thesupernatant using a Cobra Auto Gamma Counter (Pack-ard). Percentage specific lysis was calculated as (testsample 2 spontaneous release/total release 2 spontane-ous release) 3 100. Maximal release was obtained afteraddition of 2% Triton X-100.
Reverse Transcription-Polymerase ChainReaction (RT-PCR)Total RNA was isolated from subconfluent cell culturesof JEG-3 and JAR cells using the RNAzol extractionmethod (Cinna/Biotecx Laboratories, Houston, TX).RNA samples (5 mg) were transcribed into cDNA usingAMV reverse transcriptase (Promega, Madison, WI). ThecDNA was subjected to a semiquantitative PCR usingoligonucleotide primers specific for CIITA, RFX5,RFXAP, and glyceraldehyde-3-phosphate dehydrogenase(GAPDH). The nucleotide sequence of the oligonucleo-tides used for the amplification is as follows:
CIITA: forward primer 59-GATTCCTACACAATGCGTTGCCTGGCTC-39;
CIITA: reverse primer 59-TTCATCTGGTCCTATGTGCTT-39;
RFX5: forward primer 59-tacaagcttTGGGCATATATGGGCCTGGCGAAG-39;
RFX5: reverse primer 59-tgagcggccGCCTCTACTAGGCAAAGTTAACG-39;
RFXAP: forward primer 59-cagtagaattcGGCCAAGCAGGTGCTAAAAG-39;
RFXAP: reverse primer 59-cagaggatCCATGTAGATGTTCTTGGTAAG-39;
GAPDH: forward primer 59-GGTCGGAGTCAACGGATTTG-39; and
GAPDH: reverse primer 59-ATGAGCCCCAGCCTTCTCCAT-39.
For the CIITA PCR, 2 ml of cDNA was used in a totalreaction volume of 100 ml containing 10 mM Tris-HCl(pH 8.4), 50 mM KCl, 4 mM MgCl2, 0.6 mg/ml BSA,0.5 mM of each dNTP, 20 pmol of 59-primer, and 20pmol of 39-primer and 2.5 U of AmpliTaq DNA poly-
852 P. J. van den Elsen et al.

merase (Perkin-Elmer, Roche Molecular Systems, Branch-burg, NJ). PCR cycles were performed for 30 times inwhich each cycle consisted of a denaturation step at 94°Cfor 1 min, an annealing step at 59°C for 1 min, and anelongation step at 72°C for 1 min. The final elongationstep was performed for an extra 5 min at 72°C. The samePCR was also performed for GAPDH (25 cycles). For theRFX5 PCR, 2 ml of cDNA was used in a total volume of100 ml consisting of XL buffer (Perkin/Elmer, RocheMolecular Systems) supplemented with 0.5 mM of eachdNTP, 1.1 mM Mg(OAc)2, 30 pmol 59-primer, and 30pmol of 39-primer. The samples were heated for 2 min at94°C, cooled to 50°C, after which 4 U of Tth enzyme withXL buffer were added. The PCR consisted of 10 cycles inwhich each cycle consisted of a denaturation step for 1 minat 94°C, an annealing step for 1 min at 50°C, and anelongation step for 2 min at 68°C. After this initial PCRa second PCR was performed consisting of 20 cycles withincreasing elongation steps (20 sec per PCR cycle). Thefinal elongation step included an extra 5 min at 68°C.
The RFXAP RT-PCR was performed in a similarfashion as the RFX5 PCR with the exception that DMSOwas included in the reaction (final concentration 5%).The PCR consisted of 40 cycles, each with a denaturationstep at 94°C for 1 min, an annealing step at 50°C for 1min, and an elongation step at 68°C for 2 min.
For the analysis of CIITA and IRF-1 expression innormal trophoblast cells the following primer pairs wereused:
CIITA-pan forward primer: 59-AGCCGGTGGAGCAGTTCTACCGC-39;
CIITA-pan reverse primer: 59-TGGAAGATCAGCCCAGCCAGAAAGC-39;
IRF-1 forward primer: 59-ATGAGACCCTGGCTAGAGATGC-39; and
IRF-1 reverse primer: 59-GGAGGGCATGGGTGACACCTG-39.
RT-PCR cycles were performed for 35 times (CIITA) or27 times (IRF-1).
For the analysis of levels of HLA-A and -B transcriptsthe RT-PCR cycles were performed for 35 times inwhich each cycle consisted of a denaturation step at94°C, an annealing step at 60°C, and an elongation stepat 72°C each step for 1 min. The primer pairs used were:
HLA-A forward primer: 59-tataaaagcttATGGCCGT-CATGGCGCCCC-39;
HLA-A reverse primer: 59-gcggcctctagaTCACACTTTACAAGCTGTC-39;
HLA-B forward primer: 59-tatgaattcATGCGGGTCACGGCGCCCCGA-39; and
HLA-B reverse primer: 59-ctcgaatTCAAGCTGTGAGAGACACATCAG-39.
Generation of Stable TransfectantsOne microgram of EBO-CIITA was transfected into106 JEG-3 and JAR cells by the calcium phosphatecoprecipitation method [39]. Transfected cells wereselected in culture medium containing 50 mg/ml ofhygromycin-B (Boehringer Mannheim, Mannheim,Germany) and evaluated for MHC class II expressionpatterns. The MHC class II-positive cells in the CIITA-transfected pool were enriched for by cell sorting usingDynabeads (Dynal, Oslo, Norway) coated with HLA-DRmAb.
CIITAPIV-Promoter Reporter AssayThe 2461/175 CIITAPIV promoter fragment wasisolated by PCR from Cos14 (a kind gift of C. Alcaide-Loridan), and its nucleotide sequence was confirmed byDNA sequencing. In pGL3-CIITAPIV, the 2461/175CIITAPIV promoter fragment was cloned upstream ofthe luciferase gene of pGL3. Transient transfection ofJEG-3 and JAR cells were performed with pGL3-CIITAPIV essentially as described previously [11].JEG-3 and JAR cells were treated with 500 U/ml ofIFN-g for 48 h. In each of four wells of a six-well plate,0.2 3 106 cells were transfected with 2.5 mg reporterplasmid and 1 mg of RSV-Renilla luciferase controlplasmid. Luciferase values were normalized with theRenilla luciferase values and are expressed as mean 6 SD(n 5 4).
Southern Blot AnalysisFor the hypermethylation analysis of the PIV CIITApromoter, total genomic DNA (gDNA) was isolated bystandard protocols. Then 15 mg of this gDNA fromJEG-3 and JAR was digested with HindIII only, orcombined with AvaI, HpaII, MspI, and SmaI. gDNA ofHeLa cells served as a control. After digestion, frag-ments were separated by electrophoresis on a 1% aga-rose gel containing 0.5 3 TBE buffer and were subse-quently blotted onto Hybond N1 membranes(Amersham, Little Chalfont, England). The blots werehybridized with a [32P]-labeled PIV CIITA-probe con-taining nucleotides 2461 to 175 and analyzed by phos-phor imaging (PhosphorImager, Molecular Dynamics,Sunnyvale, CA).
RESULTSCell Surface MHC Expression on JEG-3 and JARTrophoblast Cell LinesJEG-3 and JAR trophoblast cells are characterized by ageneral lack of MHC class II and classical MHC class Iexpression. JEG-3 trophoblast cells lack constitutive cellsurface expression of all MHC class II isotypes (Fig. 1).As previously established, JEG-3 cells lack expression of
853CIITA-Mediated Induction of MHC Expression in Trophoblast Cells
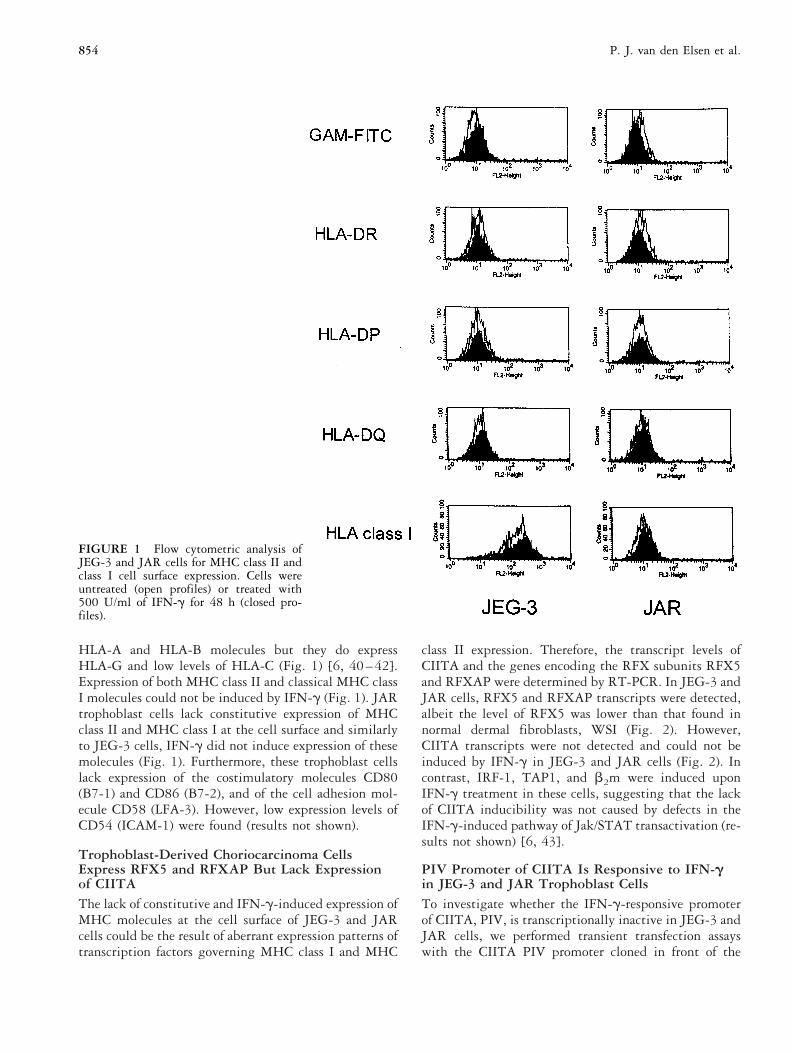
HLA-A and HLA-B molecules but they do expressHLA-G and low levels of HLA-C (Fig. 1) [6, 40–42].Expression of both MHC class II and classical MHC classI molecules could not be induced by IFN-g (Fig. 1). JARtrophoblast cells lack constitutive expression of MHCclass II and MHC class I at the cell surface and similarlyto JEG-3 cells, IFN-g did not induce expression of thesemolecules (Fig. 1). Furthermore, these trophoblast cellslack expression of the costimulatory molecules CD80(B7-1) and CD86 (B7-2), and of the cell adhesion mol-ecule CD58 (LFA-3). However, low expression levels ofCD54 (ICAM-1) were found (results not shown).
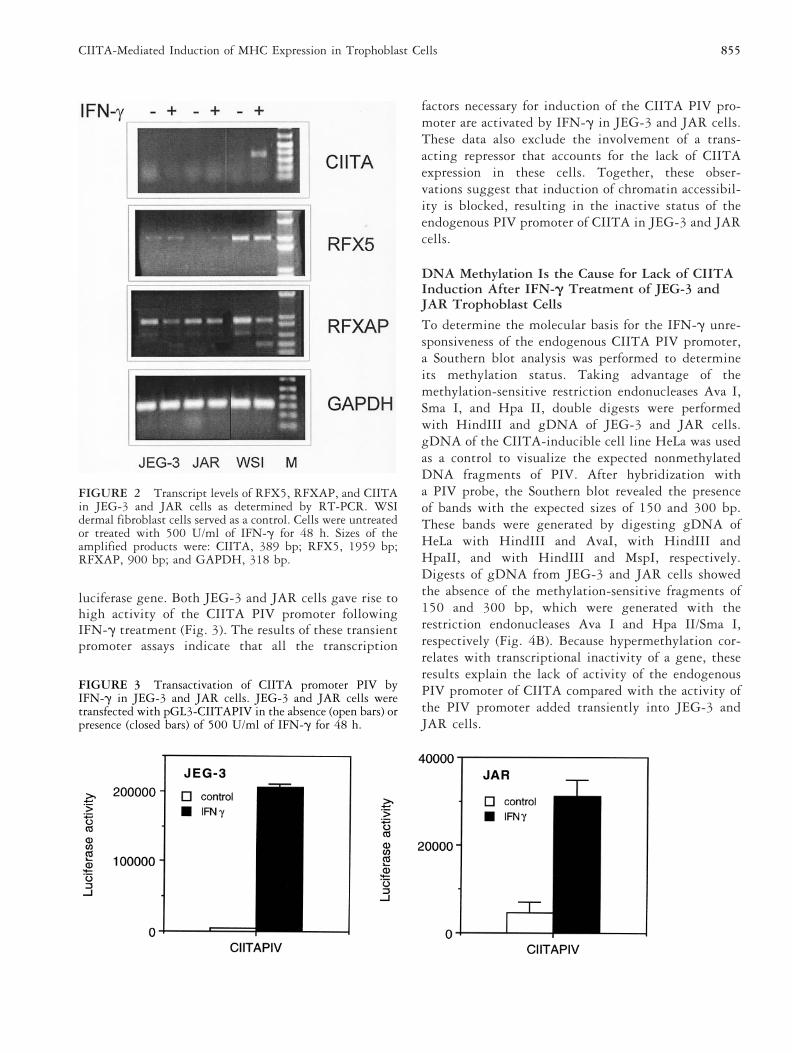
Trophoblast-Derived Choriocarcinoma CellsExpress RFX5 and RFXAP But Lack Expressionof CIITAThe lack of constitutive and IFN-g-induced expression ofMHC molecules at the cell surface of JEG-3 and JARcells could be the result of aberrant expression patterns oftranscription factors governing MHC class I and MHC
class II expression. Therefore, the transcript levels ofCIITA and the genes encoding the RFX subunits RFX5and RFXAP were determined by RT-PCR. In JEG-3 andJAR cells, RFX5 and RFXAP transcripts were detected,albeit the level of RFX5 was lower than that found innormal dermal fibroblasts, WSI (Fig. 2). However,CIITA transcripts were not detected and could not beinduced by IFN-g in JEG-3 and JAR cells (Fig. 2). Incontrast, IRF-1, TAP1, and b2m were induced uponIFN-g treatment in these cells, suggesting that the lackof CIITA inducibility was not caused by defects in theIFN-g-induced pathway of Jak/STAT transactivation (re-sults not shown) [6, 43].
PIV Promoter of CIITA Is Responsive to IFN-gin JEG-3 and JAR Trophoblast CellsTo investigate whether the IFN-g-responsive promoterof CIITA, PIV, is transcriptionally inactive in JEG-3 andJAR cells, we performed transient transfection assayswith the CIITA PIV promoter cloned in front of the
FIGURE 1 Flow cytometric analysis ofJEG-3 and JAR cells for MHC class II andclass I cell surface expression. Cells wereuntreated (open profiles) or treated with500 U/ml of IFN-g for 48 h (closed pro-files).
854 P. J. van den Elsen et al.
luciferase gene. Both JEG-3 and JAR cells gave rise tohigh activity of the CIITA PIV promoter followingIFN-g treatment (Fig. 3). The results of these transientpromoter assays indicate that all the transcription
factors necessary for induction of the CIITA PIV pro-moter are activated by IFN-g in JEG-3 and JAR cells.These data also exclude the involvement of a trans-acting repressor that accounts for the lack of CIITAexpression in these cells. Together, these obser-vations suggest that induction of chromatin accessibil-ity is blocked, resulting in the inactive status of theendogenous PIV promoter of CIITA in JEG-3 and JARcells.
DNA Methylation Is the Cause for Lack of CIITAInduction After IFN-g Treatment of JEG-3 andJAR Trophoblast Cells
To determine the molecular basis for the IFN-g unre-sponsiveness of the endogenous CIITA PIV promoter,a Southern blot analysis was performed to determineits methylation status. Taking advantage of themethylation-sensitive restriction endonucleases Ava I,Sma I, and Hpa II, double digests were performedwith HindIII and gDNA of JEG-3 and JAR cells.gDNA of the CIITA-inducible cell line HeLa was usedas a control to visualize the expected nonmethylatedDNA fragments of PIV. After hybridization witha PIV probe, the Southern blot revealed the presenceof bands with the expected sizes of 150 and 300 bp.These bands were generated by digesting gDNA ofHeLa with HindIII and AvaI, with HindIII andHpaII, and with HindIII and MspI, respectively.Digests of gDNA from JEG-3 and JAR cells showedthe absence of the methylation-sensitive fragments of150 and 300 bp, which were generated with therestriction endonucleases Ava I and Hpa II/Sma I,respectively (Fig. 4B). Because hypermethylation cor-relates with transcriptional inactivity of a gene, theseresults explain the lack of activity of the endogenousPIV promoter of CIITA compared with the activity ofthe PIV promoter added transiently into JEG-3 andJAR cells.
FIGURE 3 Transactivation of CIITA promoter PIV byIFN-g in JEG-3 and JAR cells. JEG-3 and JAR cells weretransfected with pGL3-CIITAPIV in the absence (open bars) orpresence (closed bars) of 500 U/ml of IFN-g for 48 h.
FIGURE 2 Transcript levels of RFX5, RFXAP, and CIITAin JEG-3 and JAR cells as determined by RT-PCR. WSIdermal fibroblast cells served as a control. Cells were untreatedor treated with 500 U/ml of IFN-g for 48 h. Sizes of theamplified products were: CIITA, 389 bp; RFX5, 1959 bp;RFXAP, 900 bp; and GAPDH, 318 bp.
855CIITA-Mediated Induction of MHC Expression in Trophoblast Cells
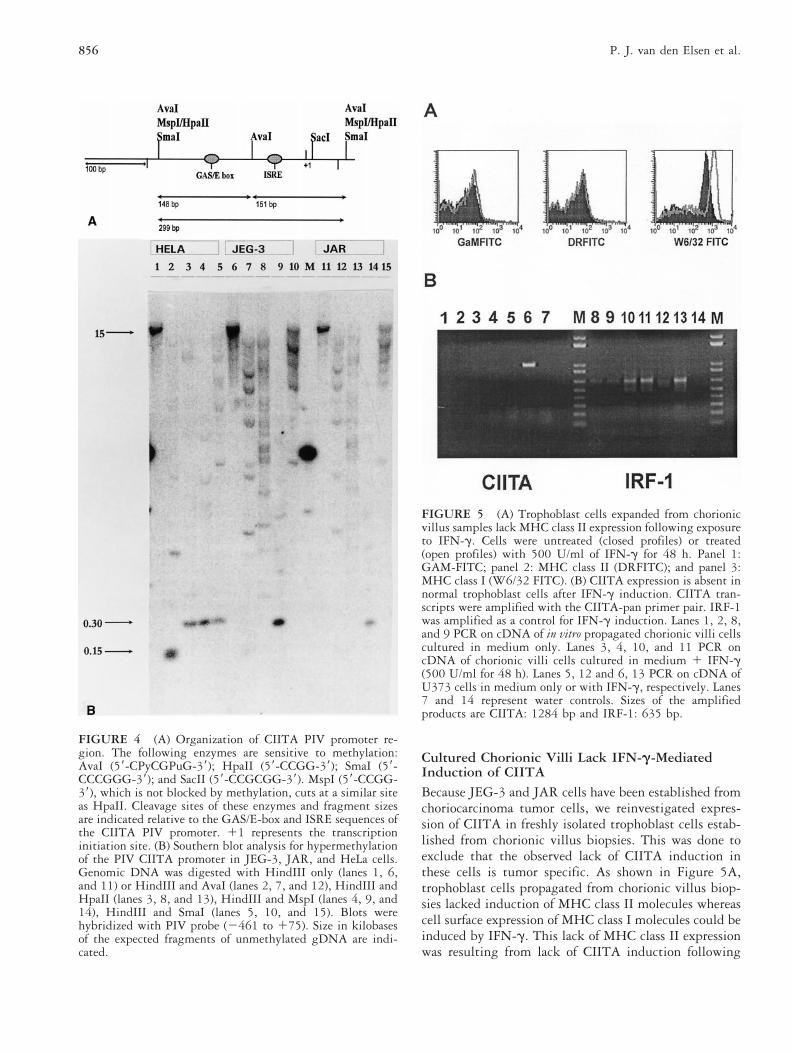
Cultured Chorionic Villi Lack IFN-g-MediatedInduction of CIITABecause JEG-3 and JAR cells have been established fromchoriocarcinoma tumor cells, we reinvestigated expres-sion of CIITA in freshly isolated trophoblast cells estab-lished from chorionic villus biopsies. This was done toexclude that the observed lack of CIITA induction inthese cells is tumor specific. As shown in Figure 5A,trophoblast cells propagated from chorionic villus biop-sies lacked induction of MHC class II molecules whereascell surface expression of MHC class I molecules could beinduced by IFN-g. This lack of MHC class II expressionwas resulting from lack of CIITA induction following
FIGURE 4 (A) Organization of CIITA PIV promoter re-gion. The following enzymes are sensitive to methylation:AvaI (59-CPyCGPuG-39); HpaII (59-CCGG-39); SmaI (59-CCCGGG-39); and SacII (59-CCGCGG-39). MspI (59-CCGG-39), which is not blocked by methylation, cuts at a similar siteas HpaII. Cleavage sites of these enzymes and fragment sizesare indicated relative to the GAS/E-box and ISRE sequences ofthe CIITA PIV promoter. 11 represents the transcriptioninitiation site. (B) Southern blot analysis for hypermethylationof the PIV CIITA promoter in JEG-3, JAR, and HeLa cells.Genomic DNA was digested with HindIII only (lanes 1, 6,and 11) or HindIII and AvaI (lanes 2, 7, and 12), HindIII andHpaII (lanes 3, 8, and 13), HindIII and MspI (lanes 4, 9, and14), HindIII and SmaI (lanes 5, 10, and 15). Blots werehybridized with PIV probe (2461 to 175). Size in kilobasesof the expected fragments of unmethylated gDNA are indi-cated.
FIGURE 5 (A) Trophoblast cells expanded from chorionicvillus samples lack MHC class II expression following exposureto IFN-g. Cells were untreated (closed profiles) or treated(open profiles) with 500 U/ml of IFN-g for 48 h. Panel 1:GAM-FITC; panel 2: MHC class II (DRFITC); and panel 3:MHC class I (W6/32 FITC). (B) CIITA expression is absent innormal trophoblast cells after IFN-g induction. CIITA tran-scripts were amplified with the CIITA-pan primer pair. IRF-1was amplified as a control for IFN-g induction. Lanes 1, 2, 8,and 9 PCR on cDNA of in vitro propagated chorionic villi cellscultured in medium only. Lanes 3, 4, 10, and 11 PCR oncDNA of chorionic villi cells cultured in medium 1 IFN-g(500 U/ml for 48 h). Lanes 5, 12 and 6, 13 PCR on cDNA ofU373 cells in medium only or with IFN-g, respectively. Lanes7 and 14 represent water controls. Sizes of the amplifiedproducts are CIITA: 1284 bp and IRF-1: 635 bp.
856 P. J. van den Elsen et al.

treatment with IFN-g (Fig. 5B). In contrast, IRF-1 wasinduced by IFN-g in these trophoblast cells propagatedfrom chorionic villus biopsies. Together these data revealthat lack of CIITA expression is at the heart of the MHCclass II-deficient character of trophoblast cells.
Antigen Presentation Functions ofCIITA-Transfected JEG-3 and JARTrophoblast CellsBecause CIITA cannot be induced by IFN-g in JEG-3and JAR cells, we investigated the effects of CIITAintroduction on antigen-presenting functions in thesecells. Stable CIITA transfectants of JEG-3 (JEG-3/CIITA) and JAR (JAR/CIITA) were generated to test theMHC expression characteristics. CIITA could restoreMHC class II expression in both trophoblast cell lines(Fig. 6A). Cell surface expression of MHC class I mole-cules was also increased in JEG-3/CIITA cells, whereasJAR/CIITA cells remained MHC class I-deficient at theircell membrane (Fig. 6A). RT-PCR analysis revealed thatJAR/CIITA cells failed to express HLA-A and -B genes,whereas JEG-3/CIITA cells expressed only HLA-B (Fig.6B). There were no significant changes in the levels ofcostimulatory molecules (CD80 and CD86) and adhesionmolecules (CD58 and CD54) expression in JEG-3/CIITAand JAR/CIITA cells (results not shown).
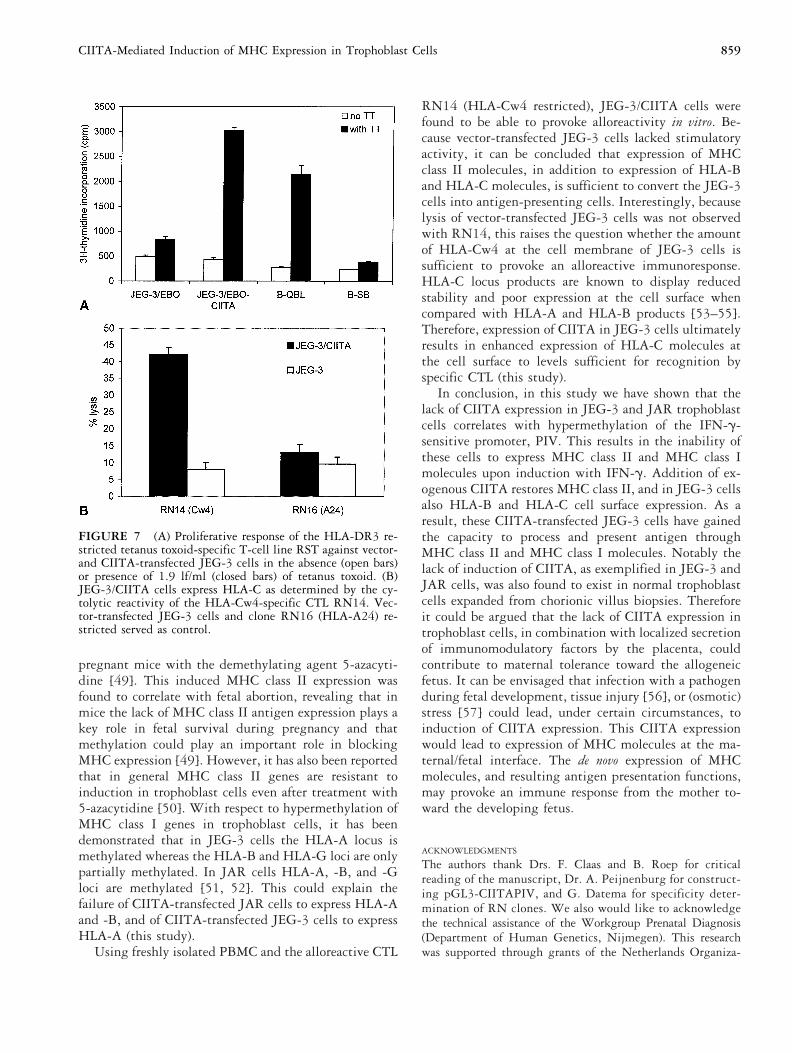
The JEG-3/CIITA cells were tested for antigen pro-cessing and MHC class II-mediated antigen presentation.For those purposes, JEG-3/CIITA cells were pulsed withtetanus toxoid and a proliferation assay was performedwith the TT-specific T-cell line RST-TT (HLA-DR3restricted). The CIITA-transfected JEG-3 cells induced aproliferative response with a stimulation index that wassimilar to those of TT-pulsed EBV-transformed B cells(Fig. 7A). This demonstrates that expression of exoge-nous CIITA in JEG-3 trophoblast cells results in theacquisition of MHC class II-mediated antigen processingand presentation functions.
Previously, JEG-3 cells have been shown to expressHLA-G and HLA-C molecules. Because the HLA-Gpromoter lacks the critical elements for CIITA-mediatedtransactivation [43], we tested the JEG-3/CIITA cells fortheir HLA-C alloreactive capacity. JEG3/CIITA cellswere found to stimulate the HLA-Cw4-specific CTLclone RN14, revealing enhanced cell surface expressionlevels of HLA-C molecules in these CIITA-transfectedJEG-3 cells (Fig. 7B).
To evaluate whether CIITA is capable to stimulatealloreactive responses from PBMC, JEG-3/CIITA cellswere used as stimulator cells to determine whether en-dogenously encoded MHC class I molecules could stim-ulate such an allogeneic T-cell immune response. Basedon serological typing, these JEG-3/CIITA cells werefound to express HLA-B8, -B35, -Cw4, -Cw7, and -G.
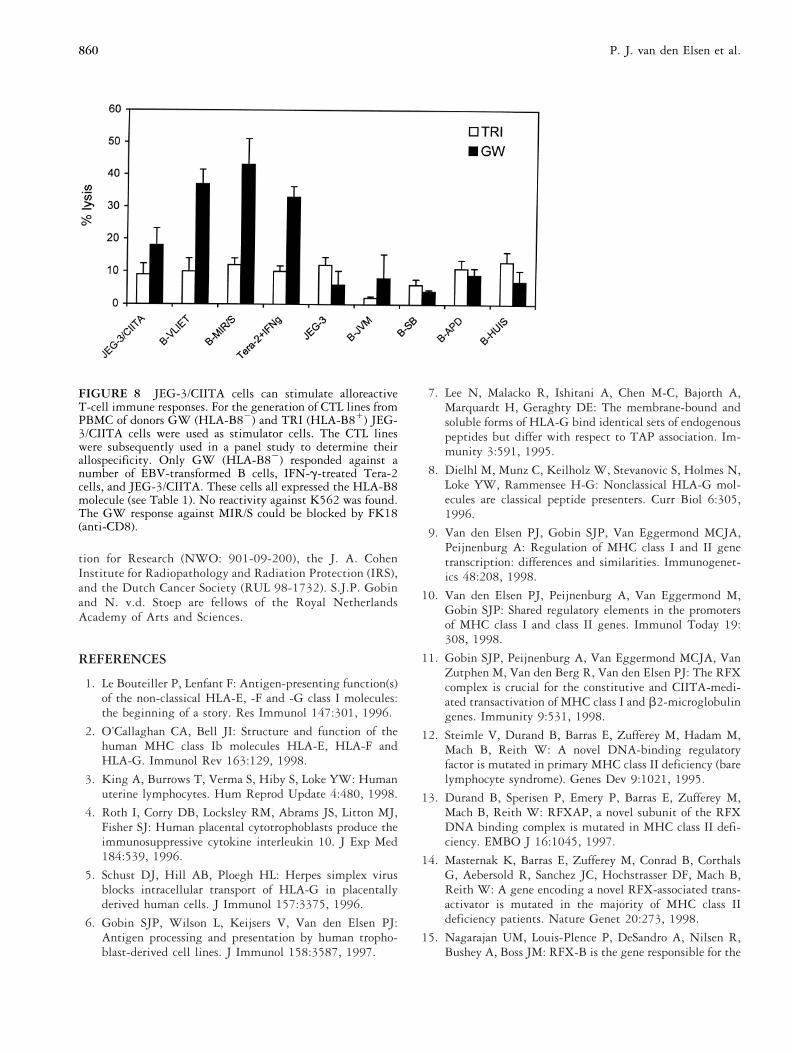
Using PBMC from donors GW (HLA-A24, B18, B27,DR12, DR13, DQ7, DQ6) and TRI (HLA-A24, A11,B7, B8, DR17, DR4, DQ2, DQ7), JEG-3/CIITA cellswere shown to possess low levels of T-cell alloreactivestimulatory activity toward donor GW PBMC only, thatreaction was not observed with untransfected JEG-3 cells(Fig. 8). Subsequently it was shown that the allospecificrestricting element of the CTL line was the HLA-B8allele of JEG-3/CIITA cells. No reactivity of the GW/JEG-3/CIITA CTL line was observed against K562 cellsand cytolysis could be blocked with an anti-CD8 anti-body (results not shown). Together, these results dem-onstrate that expression of CIITA in trophoblast cellsresults in acquisition of antigen presenting functions andcapacity to stimulate allogeneic T-cell responses.
DISCUSSIONIn this study we found evidence for a mechanism thatcauses the lack of expression of MHC class II and classicalMHC class I molecules in JEG-3 and JAR trophoblastcell lines. These choriocarcinoma cells lack IFN-g-medi-ated induction of CIITA, a coactivator that plays animportant role in the common regulatory pathway ofMHC class I and class II gene transcription [10, 11, 26,27]. Our observations are in line with recent reports alsorevealing the lack of CIITA expression in trophoblast-derived tumor cells [44, 45]. Notably, we have shown inthis study that trophoblast cells propagated from freshlyisolated chorionic villus biopsies also displayed lack ofCIITA expression following IFN-g treatment. BecauseIRF-1 was induced, it was concluded that the lack ofexpression of endogenous CIITA results from in vivomodifications that prevent transcriptional activation ofthe IFN-g-responsive CIITA promoter, PIV. Here wedemonstrated that this lack of CIITA expression follow-ing IFN-g treatment of choriocarcinoma cells was result-ing from hypermethylation of CIITA promoter PIV.Therefore, the lack of CIITA expression was not due to arepressor or lack of other essential transcription factorsthat affect transcriptional activation of CIITA PIV.
Exogenous CIITA bypasses this defect and transacti-vates MHC class II and class I promoters [44–46] inJEG-3 cells, ultimately resulting in de novo cell surfaceexpression of MHC class II and an increase in expressionof MHC class I molecules. Because trophoblast cellsconstitutively express the TAP and LMP genes [6, 47,48], the newly synthesized heavy and light chains areassembled efficiently and loaded with peptide by thepreexisting proteasome and TAP complexes. It results inan increased expression of the MHC class I moleculesHLA-B and HLA-C on the cell membrane.
It has been shown that MHC class II molecules can beinduced on fetal trophoblast cells following treatment of
857CIITA-Mediated Induction of MHC Expression in Trophoblast Cells
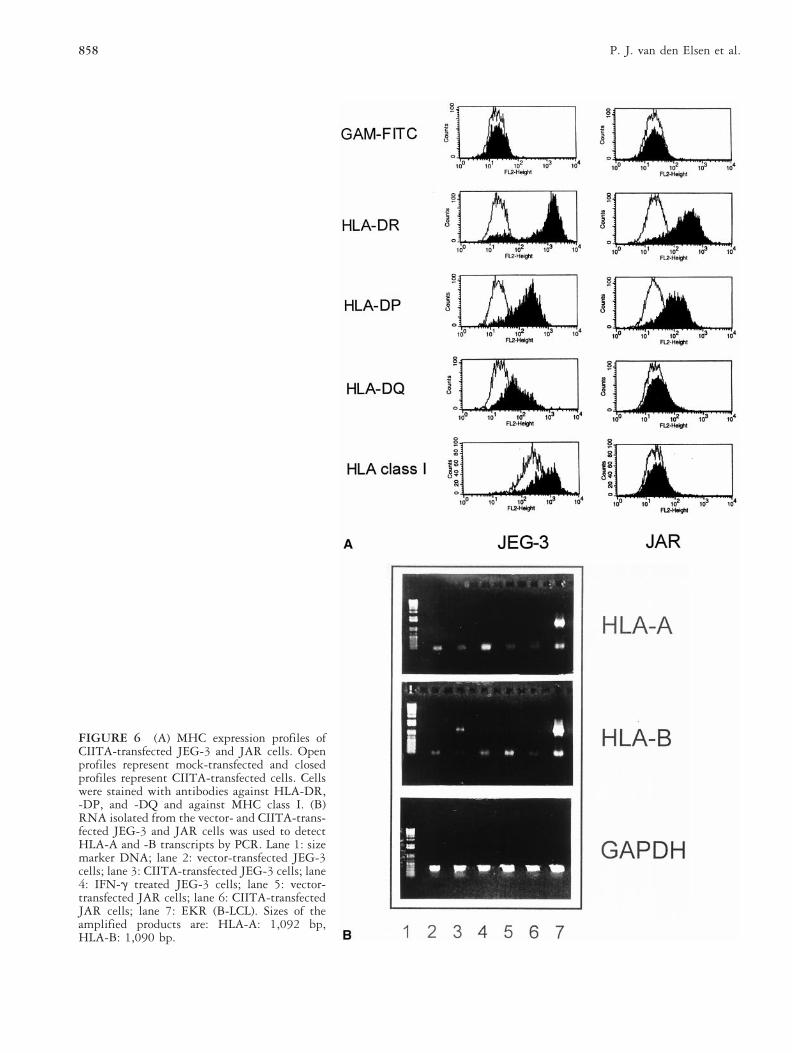
FIGURE 6 (A) MHC expression profiles ofCIITA-transfected JEG-3 and JAR cells. Openprofiles represent mock-transfected and closedprofiles represent CIITA-transfected cells. Cellswere stained with antibodies against HLA-DR,-DP, and -DQ and against MHC class I. (B)RNA isolated from the vector- and CIITA-trans-fected JEG-3 and JAR cells was used to detectHLA-A and -B transcripts by PCR. Lane 1: sizemarker DNA; lane 2: vector-transfected JEG-3cells; lane 3: CIITA-transfected JEG-3 cells; lane4: IFN-g treated JEG-3 cells; lane 5: vector-transfected JAR cells; lane 6: CIITA-transfectedJAR cells; lane 7: EKR (B-LCL). Sizes of theamplified products are: HLA-A: 1,092 bp,HLA-B: 1,090 bp.
858 P. J. van den Elsen et al.
pregnant mice with the demethylating agent 5-azacyti-dine [49]. This induced MHC class II expression wasfound to correlate with fetal abortion, revealing that inmice the lack of MHC class II antigen expression plays akey role in fetal survival during pregnancy and thatmethylation could play an important role in blockingMHC expression [49]. However, it has also been reportedthat in general MHC class II genes are resistant toinduction in trophoblast cells even after treatment with5-azacytidine [50]. With respect to hypermethylation ofMHC class I genes in trophoblast cells, it has beendemonstrated that in JEG-3 cells the HLA-A locus ismethylated whereas the HLA-B and HLA-G loci are onlypartially methylated. In JAR cells HLA-A, -B, and -Gloci are methylated [51, 52]. This could explain thefailure of CIITA-transfected JAR cells to express HLA-Aand -B, and of CIITA-transfected JEG-3 cells to expressHLA-A (this study).
Using freshly isolated PBMC and the alloreactive CTL
RN14 (HLA-Cw4 restricted), JEG-3/CIITA cells werefound to be able to provoke alloreactivity in vitro. Be-cause vector-transfected JEG-3 cells lacked stimulatoryactivity, it can be concluded that expression of MHCclass II molecules, in addition to expression of HLA-Band HLA-C molecules, is sufficient to convert the JEG-3cells into antigen-presenting cells. Interestingly, becauselysis of vector-transfected JEG-3 cells was not observedwith RN14, this raises the question whether the amountof HLA-Cw4 at the cell membrane of JEG-3 cells issufficient to provoke an alloreactive immunoresponse.HLA-C locus products are known to display reducedstability and poor expression at the cell surface whencompared with HLA-A and HLA-B products [53–55].Therefore, expression of CIITA in JEG-3 cells ultimatelyresults in enhanced expression of HLA-C molecules atthe cell surface to levels sufficient for recognition byspecific CTL (this study).
In conclusion, in this study we have shown that thelack of CIITA expression in JEG-3 and JAR trophoblastcells correlates with hypermethylation of the IFN-g-sensitive promoter, PIV. This results in the inability ofthese cells to express MHC class II and MHC class Imolecules upon induction with IFN-g. Addition of ex-ogenous CIITA restores MHC class II, and in JEG-3 cellsalso HLA-B and HLA-C cell surface expression. As aresult, these CIITA-transfected JEG-3 cells have gainedthe capacity to process and present antigen throughMHC class II and MHC class I molecules. Notably thelack of induction of CIITA, as exemplified in JEG-3 andJAR cells, was also found to exist in normal trophoblastcells expanded from chorionic villus biopsies. Thereforeit could be argued that the lack of CIITA expression introphoblast cells, in combination with localized secretionof immunomodulatory factors by the placenta, couldcontribute to maternal tolerance toward the allogeneicfetus. It can be envisaged that infection with a pathogenduring fetal development, tissue injury [56], or (osmotic)stress [57] could lead, under certain circumstances, toinduction of CIITA expression. This CIITA expressionwould lead to expression of MHC molecules at the ma-ternal/fetal interface. The de novo expression of MHCmolecules, and resulting antigen presentation functions,may provoke an immune response from the mother to-ward the developing fetus.
ACKNOWLEDGMENTS
The authors thank Drs. F. Claas and B. Roep for criticalreading of the manuscript, Dr. A. Peijnenburg for construct-ing pGL3-CIITAPIV, and G. Datema for specificity deter-mination of RN clones. We also would like to acknowledgethe technical assistance of the Workgroup Prenatal Diagnosis(Department of Human Genetics, Nijmegen). This researchwas supported through grants of the Netherlands Organiza-
FIGURE 7 (A) Proliferative response of the HLA-DR3 re-stricted tetanus toxoid-specific T-cell line RST against vector-and CIITA-transfected JEG-3 cells in the absence (open bars)or presence of 1.9 lf/ml (closed bars) of tetanus toxoid. (B)JEG-3/CIITA cells express HLA-C as determined by the cy-tolytic reactivity of the HLA-Cw4-specific CTL RN14. Vec-tor-transfected JEG-3 cells and clone RN16 (HLA-A24) re-stricted served as control.
859CIITA-Mediated Induction of MHC Expression in Trophoblast Cells
tion for Research (NWO: 901-09-200), the J. A. CohenInstitute for Radiopathology and Radiation Protection (IRS),and the Dutch Cancer Society (RUL 98-1732). S.J.P. Gobinand N. v.d. Stoep are fellows of the Royal NetherlandsAcademy of Arts and Sciences.
REFERENCES
1. Le Bouteiller P, Lenfant F: Antigen-presenting function(s)of the non-classical HLA-E, -F and -G class I molecules:the beginning of a story. Res Immunol 147:301, 1996.
2. O’Callaghan CA, Bell JI: Structure and function of thehuman MHC class Ib molecules HLA-E, HLA-F andHLA-G. Immunol Rev 163:129, 1998.
3. King A, Burrows T, Verma S, Hiby S, Loke YW: Humanuterine lymphocytes. Hum Reprod Update 4:480, 1998.
4. Roth I, Corry DB, Locksley RM, Abrams JS, Litton MJ,Fisher SJ: Human placental cytotrophoblasts produce theimmunosuppressive cytokine interleukin 10. J Exp Med184:539, 1996.
5. Schust DJ, Hill AB, Ploegh HL: Herpes simplex virusblocks intracellular transport of HLA-G in placentallyderived human cells. J Immunol 157:3375, 1996.
6. Gobin SJP, Wilson L, Keijsers V, Van den Elsen PJ:Antigen processing and presentation by human tropho-blast-derived cell lines. J Immunol 158:3587, 1997.
7. Lee N, Malacko R, Ishitani A, Chen M-C, Bajorth A,Marquardt H, Geraghty DE: The membrane-bound andsoluble forms of HLA-G bind identical sets of endogenouspeptides but differ with respect to TAP association. Im-munity 3:591, 1995.
8. Dielhl M, Munz C, Keilholz W, Stevanovic S, Holmes N,Loke YW, Rammensee H-G: Nonclassical HLA-G mol-ecules are classical peptide presenters. Curr Biol 6:305,1996.
9. Van den Elsen PJ, Gobin SJP, Van Eggermond MCJA,Peijnenburg A: Regulation of MHC class I and II genetranscription: differences and similarities. Immunogenet-ics 48:208, 1998.
10. Van den Elsen PJ, Peijnenburg A, Van Eggermond M,Gobin SJP: Shared regulatory elements in the promotersof MHC class I and class II genes. Immunol Today 19:308, 1998.
11. Gobin SJP, Peijnenburg A, Van Eggermond MCJA, VanZutphen M, Van den Berg R, Van den Elsen PJ: The RFXcomplex is crucial for the constitutive and CIITA-medi-ated transactivation of MHC class I and b2-microglobulingenes. Immunity 9:531, 1998.
12. Steimle V, Durand B, Barras E, Zufferey M, Hadam M,Mach B, Reith W: A novel DNA-binding regulatoryfactor is mutated in primary MHC class II deficiency (barelymphocyte syndrome). Genes Dev 9:1021, 1995.
13. Durand B, Sperisen P, Emery P, Barras E, Zufferey M,Mach B, Reith W: RFXAP, a novel subunit of the RFXDNA binding complex is mutated in MHC class II defi-ciency. EMBO J 16:1045, 1997.
14. Masternak K, Barras E, Zufferey M, Conrad B, CorthalsG, Aebersold R, Sanchez JC, Hochstrasser DF, Mach B,Reith W: A gene encoding a novel RFX-associated trans-activator is mutated in the majority of MHC class IIdeficiency patients. Nature Genet 20:273, 1998.
15. Nagarajan UM, Louis-Plence P, DeSandro A, Nilsen R,Bushey A, Boss JM: RFX-B is the gene responsible for the
FIGURE 8 JEG-3/CIITA cells can stimulate alloreactiveT-cell immune responses. For the generation of CTL lines fromPBMC of donors GW (HLA-B82) and TRI (HLA-B81) JEG-3/CIITA cells were used as stimulator cells. The CTL lineswere subsequently used in a panel study to determine theirallospecificity. Only GW (HLA-B82) responded against anumber of EBV-transformed B cells, IFN-g-treated Tera-2cells, and JEG-3/CIITA. These cells all expressed the HLA-B8molecule (see Table 1). No reactivity against K562 was found.The GW response against MIR/S could be blocked by FK18(anti-CD8).
860 P. J. van den Elsen et al.

most common cause of the bare lymphocyte syndrome, anMHC class II immunodeficiency. Immunity 10:153,1999.
16. Moreno CS, Beresford GW, Louis-Plence P, Morris AC,Boss JM: CREB regulates MHC class II expression in aCIITA-dependent manner. Immunity 10:143, 1999.
17. Zeleznik-Le N, Azizkhan JC, Ting JP-Y: Affinity purifiedCCAAT-box-binding protein (YEBP) functionally regu-lates expression of a human class II major histocompati-bility complex gene and the herpes simplex virus thymi-dine kinase gene. Proc Natl Acad Sci USA 88:1873, 1991.
18. Schoneich J, Lee JL, Mansky P, Sheffery M, Yang SY: Thepentanucleotide ATTGG, the “inverted CCAAT,” is anessential element for HLA class I gene transcription. J Im-munol 158:4788, 1997.
19. Reith W, Siegrist CA, Durand B, Barras E, Mach B:Function of major histocompatibility complex class IIpromoters requires cooperative binding between factorsRFX and NF-Y. Proc Natl Acad Sci USA 91:554, 1994.
20. Wright KL, Vilen BJ, Itoh-Lindstrom Y, Moore TL, Li G,Criscitiello M, Cogswell P, Clarke JB, Ting JP-Y:CCAAT box binding protein NF-Y facilitates in vivorecruitment of upstream DNA binding transcription fac-tors. EMBO J 13:4042, 1994.
21. Fontes JD, Jabrane-Ferrat N, Peterlin BM: Assembly offunctional regulatory complexes on MHC class II promot-ers in vivo. J Mol Biol 270:336, 1997.
22. Louis-Plence P, Moreno CS, Boss JM: Formation of aregulatory factor X/X2 box-binding protein/nuclear fac-tor-Y multiprotein complex on the conserved regulatoryregions of HLA class II genes. J Immunol 159:3899,1997.
23. Steimle V, Otten LA, Zufferey M, Mach B: Complemen-tation cloning of an MHC class II transactivator mutatedin hereditary MHC class II deficiency (or bare lymphocytesyndrome). Cell 75:135, 1993.
24. Riley JL, Westerheide SD, Price JA, Brown JA, Boss JM:Activation of class II MHC genes requires both the X boxregion and the class II transactivator (CIITA). Immunity2:533, 1995.
25. Zhou H, Glimcher LH: Human MHC class II gene tran-scription directed by the carboxyl terminus of CIITA, oneof the defective genes in type II MHC combined immunedeficiency. Immunity 2:545, 1995.
26. Gobin SJP, Peijnenburg A, Keijsers V, Van den Elsen PJ:Site a is crucial for two routes of IFN-g induced MHCclass I transactivation: the ISRE-mediated route and anovel pathway involving CIITA. Immunity 6:601, 1997.
27. Martin B, Chin K-C, Olsen JC, Skinner CA, Dey A, OzatoK, Ting JP-Y: Induction of MHC class I expression by theMHC class II transactivator CIITA. Immunity 6:591,1997.
28. Steimle V, Siegrist C-A, Mottet A, Lisowska-GrospierreB, Mach B: Regulation of MHC class II expression by
interferon-g mediated by the transactivator gene CIITA.Science 265:106, 1994.
29. Chang CH, Fontes JD, Peterlin M, Flavell RA: Class IItransactivator (CIITA) is sufficient for the inducible ex-pression of major histocompatibility complex class IIgenes. J Exp Med 180:1367, 1994.
30. Chin K-C, Mao C, Skinner C, Riley JL, Wright KL,Moreno CS, Stark GR, Boss JM, Ting JP-Y: Molecularanalysis of G1B and G3A IFNg mutants reveals thatdefects in CIITA or RFX result in defective class II MHCand Ii gene induction. Immunity 1:687, 1994.
31. Muhlethaler-Mottet A, Otten LA, Steimle V, Mach B:Expression of MHC class II molecules in different cellularand functional compartments is controlled by differentialusage of multiple promoters of the transactivator CIITA.EMBO J 16:2851, 1997.
32. Muhlethaler-Mottet A, Di Berardino W, Otten LA, MachB: Activation of the MHC class II transactivator CIITA byinterferon-gamma requires cooperative interaction be-tween Stat1 and USF-1. Immunity 8:157, 1998.
33. Piskurich JF, Linhoff MW, Wang Y, Ting JP-Y: Twodistinct gamma interferon-inducible promoters of the ma-jor histocompatibility complex class II transactivator geneare differentially regulated by STAT1, interferon regula-tory factor 1, and transforming growth factor b. Mol CellBiol 19:431, 1999.
34. Lennon AM, Ottone C, Rigaud G, Deaven LL, LongmireJ, Fellous M, Bono R, Alcaıde-Loridan C: Isolation of aB-cell-specific promoter for the human class II transacti-vator. Immunogenetics 45:266, 1997.
35. Barbey C, Irion O, Helg C, Chapuis B, Grand C, Chiz-zolini C, Jeannet M, Roosnek E: Characterisation of thecytotoxic alloresponse of cord blood. Bone Marrow Trans-plant 22(Suppl. 1):S26, 1998.
36. Rebai N, Malissen, Pierres R, Accolla RS, Corte G, MawasC: Distinct HLA-DR epitopes and distinct families ofHLA-DR molecules defined by 15 monoclonal antibodieseither DR or allo anti-Ia crossreacting with human DR.Eur J Immunol 13:106, 1983.
37. Spits H, Borst J, Giphart M, Coligan J, Terhorst C, DeVries JE: HLA-DC antigens can serve as recognition ele-ments for human cytotoxic T lymphocytes. Eur J Immu-nol 14:299, 1984.
38. Vandekerckhove BAE, Datema G, Koning F, Goulmy E,Persijn GG, Van Rood JJ, Claas FHJ, De Vries JE: Anal-ysis of the donor-specific cytotoxic T lymphocyte reper-toire in a patient with a long term surviving allograft.J Immunol 144:1288, 1990.
39. Chen C, Okayama H: High-efficiency transformation ofmammalian cells by plasmid DNA. Mol Cell Biol 7:2745,1987.
40. Rinke de Wit TF, Vloemans S, Van den Elsen PJ, Ha-worth A, Stern PL: Differential expression of the HLAclass I multigene family by human embryonal carcinoma
861CIITA-Mediated Induction of MHC Expression in Trophoblast Cells

and choriocarcinoma cell lines. J Immunol 144:1080,1990.
41. King A, Boocock C, Sharkey AM, Gardner L, Beretta A,Siccardi AG, Loke YW: Evidence for the expression ofHLA-C class I mRNA and protein by human first trimes-ter trophoblast. J Immunol 156:2068, 1996.
42. Schust DJ, Tortorella D, Ploegh HL: HLA-G and HLA-Cat the feto-maternal interface: lessons learned from patho-genic viruses. Semin Cancer Biol 9:37, 1999.
43. Gobin SJP, Van den Elsen PJ: The regulation of HLAclass I expression: is HLA-G the odd one out? SeminCancer Biol 9:55, 1999.
44. Morris AC, Riley JL, Fleming WH, Boss JM: MHC classII gene silencing in trophoblast cells is caused by inhibi-tion of CIITA expression. Am J Reprod Immunol 40:385,1998.
45. Murphy SP, Tomasi TB: Absence of MHC class II antigenexpression in trophoblast cells results from a lack of classII transactivator (CIITA) gene expression. Mol ReprodDev 51:1, 1998.
46. Lefebre S, Moreau P, Dausset J, Carosella ED, Paul P:Downregulation of HLA class I gene transcription inchoriocarcinoma cells is controlled by the proximal pro-moter element and can be reversed by CIITA. Placenta20:293, 1999.
47. Clover LM, Sargent IL, Townsend A, Tampe R, RedmanCW: Expression of TAP1 by human trophoblast. EurJ Immunol 25:543, 1995.
48. Rodriguez AM, Mallet V, Lenfant F, Arnaud J, Girr M,Urlinger S, Bensussan A, Le Bouteiller P: Interferon-gamma rescues HLA class Ia cell surface expression interm villous trophoblast cells by inducing synthesis ofTAP proteins. Eur J Immunol 27:45, 1997.
49. Athanassakis-Vassiliadis I, Galanopoulos VK, GrigoriouM, Papamatheakis J: Induction of class II MHC antigenexpression on the murine placenta by 5-azacytidine cor-relates with fetal abortion. Cell Immunol 128:438, 1990.
50. Gustafsson E, Arvola M, Brunsberg U, Mattsson A,Mattsson R: Lack of detectable major histocompatibilitycomplex class II a beta-chain messenger ribonucleic acidin placentas of interferon-gamma- and 5-azacytidine-treated mice. Biol Reprod 57:715, 1997.
51. Boucraut J, Guillaudeux T, Alizadeh M, Boretto J, Chi-mini G, Malecaze F, Semana G, Fauchet R, Pontarotti P,Le Bouteiller P: HLA-E is the only class I gene thatescapes CpG methylation and is transcriptionally active inthe trophoblast-derived human cell line JAR. Immuno-genetics 38:117, 1993.
52. Guillaudeux T, Rodriguez AM, Girr M, Mallet V, EllisSA, Sargent IL, Fauchet R, Alsat E, Le Bouteiller P:Methylation status and transcriptional expression of theMHC class I loci in human trophoblast cells from termplacenta. J Immunol 154:3283, 1995.
53. Zemmour J, Parham P: Distinctive polymorphisms at theHLA-C locus: implications for the expression of HLA-C.J Exp Med 176:937, 1992.
54. McCutcheon J, Gumperz J, Smith K, Lutz C, Parham P:Low HLA-C expression at the cell surface correlates withincreased turnover of heavy chain mRNA. J Exp Med181:2085, 1995.
55. Gussow D, Rein R, Meijer I, De Hoog W, Seeman G,Hochstenbach F, Ploegh HL: Isolation, expression, andthe primary structure of HLA-Cw1 and HLA-Cw2 genes:evolutionary aspects. Immunogenetics 25:313, 1987.
56. Sims TN, Goes NB, Ramassar V, Urmson J, Halloran PF:In vivo class II transactivator expression in mice is inducedby a non-interferon-gamma mechanism in response tolocal injury. Transplantation 64:1657, 1997.
57. Gatsios P, Terstegen L, Schliess F, Haussinger D, KerrIM, Heinrich PC, Graeve L: Activation of the Januskinase/signal transducer and activator of transcriptionpathway by osmotic shock. J Biol Chem 273:22962,1998.
862 P. J. van den Elsen et al.
Copyright © 2022 FDOKUMEN




















