
Evaluation of promoters and visual markers for transformation ...
Upload
regeneratemdCategory
view
0download
0
Differential Usage of Class II Transactivator Promoters PI andPIV during Inflammation and Injury in Kidney
OKI TAKEUCHI,* TASHA N. SIMS,† YUTAKA TAKEI,† VIDO RAMASSAR,†
KONRAD S. FAMULSKI,† and PHILIP F. HALLORAN†
*Department of Medical Microbiology and Immunology, and †Department of Medicine, Division ofNephrology and Immunology, University of Alberta, Edmonton, Alberta, Canada.
Abstract. Expression of class II transactivator (CIITA), thetranscriptional regulator that controls all class II expression, iscontrolled in cell lines in vitro by three promoters: the dendriticcell promoter PI, the B cell promoter PIII, and the interferon-�(IFN-�)–inducible promoter, PIV. The authors examined thepromoter usage in vivo in mouse kidney in the basal state andin response to IFN-�, endotoxin, allostimulation, and renalinjury. Genetically modified mice were used to examine thedependency of each promoter on IFN-� and on the transcrip-tion factor interferon regulatory factor 1 (IRF-1). Usage ofdistinct CIITA promoters was monitored by real-time reversetranscriptase polymerase chain reaction (RT-PCR) using theunique sequences in the 5' end of the transcript from each
promoter. Kidneys in both control mice and IFN-� knockoutsexpressed chiefly PI- and PIV–related products. Administra-tion of recombinant IFN-� activated only promoter PIV. En-dotoxin or allogeneic stimulation elevated the PIV-relatedmRNA, dependent on IFN-� and on IRF-1. Ischemic renalinjury, however, increased the PI- and PIV–driven mRNAexpression in wild-type but also in IFN-�–deficient mice. Thusthe in vivo control of CIITA promoters in kidney is similar tothat observed in vitro (i.e., basal-state usage of PI and IFN-�–dependent usage of PIV during inflammation), but it alsoshows additional levels of control: IFN-�–independent basalactivity of PIV and IFN-�–independent induction of PIV dur-ing tissue injury.
MHC class II molecules are important in CD4 T cell develop-ment and in antigen presentation during the immune responsein infection, autoimmunity, and graft rejection. Class II pro-teins are expressed on antigen-presenting cells such as B cells,dendritic cells, and macrophages. Class II expression in vivo inperipheral tissues may play a role in T cell homeostasis,maintaining the peripheral CD4 T cell population (1,2). Inaddition, in human kidney in the basal state, a variable numberof epithelial and endothelial cells express class II (3), perhapsdue to basal production of interferon-� (IFN-�) (4). Expressionof class II in mouse kidney endothelium and epithelium can beinduced de novo by IFN-� (5,6).
The master regulator of cell-specific and IFN-�–inducibleclass II expression is the class II transactivator (CIITA) (7–10).CIITA belongs to the NACHT protein family, the members ofwhich are involved in inflammation and apoptosis (11). CIITAmediates its function through specific interaction with DNA-bound proteins rather than by direct binding to DNA (12). Thecloning of the human and mouse CIITA 5' regulatory region
revealed three nonhomologous promoters: PI, PIII, and PIV(13–15). These promoters direct the synthesis of three distincttranscripts with unique 5' regions. Promoter I (PI) is activatedin dendritic cells and directs the constitutive expression of the132-kD isoform. Promoter III (PIII) product, a 124-kD iso-form, is mainly detectable in B cell lines (14,16) and is silencedwith maturation to plasma cells (17). Promoter IV (PIV) is theIFN-�–inducible promoter in vitro (14,18), and it has multipletranscription factor binding sites, including STAT1, E-box, andinterferon regulatory factor 1 (IRF-1) sites. It produces the121-kD isoform that is crucial for the expression of MHC classII in nonhematopoietic cells (19).
Functions of the CIITA promoters have been studied pri-marily in vitro, leaving the in vivo role of each promoter ininflammation or tissue injury unknown. Given the potentiallyimportant role of class II in renal disease and transplantation,we examined in the present studies the control of class IIexpression in kidney by measuring the relative activity of eachpromoter in the basal and stimulated states using real-timereverse transcriptase polymerase chain reaction (RT-PCR).The results confirm that CIITA regulation in kidney in vivofollows the principal rules identified in cell cultures, but theyalso reveal additional levels of control.
Materials and MethodsMice
Mice with disrupted IFN-� gene (GKO) were created in theBALB/c background (20). Heterozygous GKO mice were generouslyprovided by Dr. T. Stewart (Genentech Inc., South San Francisco,CA). Mice with disrupted IRF-1 gene (IRF-1 KO) were generated by
Received July 9, 2003. Accepted August 12, 2003.Oki Takeuchi and Tasha N. Sims contributed equally to this work.Correspondence to Dr. Philip F. Halloran, Division of Nephrology & Immu-nology, University of Alberta, 250-Heritage Medical Research Centre, Edm-onton, Alberta T6G 2S2, Canada. Phone: 780-407-8880; Fax: 780-407-3417;E-mail: [email protected]
1046-6673/1411-2823Journal of the American Society of NephrologyCopyright © 2003 by the American Society of Nephrology
DOI: 10.1097/01.ASN.0000094084.18567.CC
J Am Soc Nephrol 14: 2823–2832, 2003

disrupting the IRF-1 gene in the 129/J background (21). HomozygousIRF-1 knockout (KO) mice were a gift from Dr. T. W. Mak (OntarioCancer Institute, Toronto, ON, Canada). Wild-type (WT) femalemice, 6 to 10 wk old (WT) BALB/c (BALB/cCr/AltBM) were ob-tained from the Health Science Laboratory Animal Services (HSLAS)at the University of Alberta and were used as matched controls forGKO mice. 129/J mice were obtained from The Jackson Laboratory(Bar Harbor, ME) and were used as matched controls for IRF-1 KOmice. Mice were kept on acidified water to inhibit the growth ofbacteria in the water source, a standard practice in mouse colony.Animal maintenance and experiments were in conformity with ap-proved animal care protocols.
IFN-� TreatmentRecombinant IFN-� (rIFN-�) was generously provided by Dr. T.
Stewart (Genentech Inc., South San Francisco, CA). Five mice pereach time point received 100,000 IU of rIFN-� in a single intraperi-toneal injection and were killed at 2, 4, 12, 24, 48, 72, or 96 h afterinjection. Kidneys were then immediately removed, decapsulated,snap-frozen in liquid nitrogen, and kept at �70°C.
P815 Tumor AllograftsMurine mastocytoma tumor cell line P815 (ATCC, American Type
Culture Collection, Rockville, MD) was passaged in DBA/2 (H-2d)mice (Jackson Laboratory). After P815 cells were collected from theascites fluid, 20 � 106 cells were injected intraperitoneally into fivemice per experimental group. Kidneys were harvested on day 7.
Endotoxin TreatmentEndotoxin (LPS) from Salmonella minnesota (Sigma, St. Louis,
MO) was dissolved in sterile saline at 250 �g/ml and heated at 56°Cfor 5 h. Five mice per experimental group were injected intraperito-neally with either saline (control) or LPS (25 �g), and kidneys wereharvested at 1, 3, 6, 12, 24, 48, 72, or 96 h after injection.
Ischemic Acute Tubular NecrosisIschemic injury to the kidney was achieved by clamping the left
renal pedicle for 60 min. Sham-operated control mice underwent asimple laparotomy under identical conditions.
Radioactive Antibody-Binding AssayThis technique has been previously reported, and its quantitative
characteristics have been described (22). Briefly, kidney transplantspecimens were homogenized, washed in phosphate-buffered saline(PBS), and centrifuged at 3000 rpm for 20 min. The pellets weresuspended in PBS at 20 mg/ml. Five milligrams of extract was thencentrifuged and resuspended in 100 �l of radiolabeled mAb solution(1000 cpm/�l) and incubated on ice for 60 min. After that time,incubates were diluted with 1 ml of PBS and centrifuged at 3000 rpmfor 20 min. The pellets were counted in a gamma counter and thevalues were corrected for the nonspecific binding.
Analysis of mRNA by Real-Time Quantitative RT-PCRTotal RNA was prepared from tissues as described previously. Two
micrograms of RNA was transcribed using M-MLV reverse transcrip-tase and random primers. Quantification of gene expression wasperformed using the ABI prism 7700 Sequence Detection System (PEApplied Biosystems) as described previously (23). All TaqMan probe/primer combinations were designed using Primer Express softwareversion 1.5 (PE Applied Biosystems) and are listed in Figure 1.
Hypoxanthine phosphoribosyltransferase (mHPRT) primers andprobe sequences were as follows: sense 5'-TGACACTGGTAAAA-CAATGCAAACT-3', antisense 5'-AACAAAGTCTGGCCTGTATC-CAA-3', probe 5'-TTCACCAGCAAGCTTGCAACCTTAACC-3'.
Probes were labeled at the 5'-end with the reporter dye moleculeFAM (6-carboxy-fluorescein) or VIC, respectively, and at the 3'-endwith the quencher dye molecule TAMARA (6-carboxy-tetramethyl-rhodamine). Each 25-�l reaction consisted of 1 � TaqMan UniversalPCR Master Mix (PE Applied Biosystems), 300 nM forward primer,300 nM reverse primer, 200 nM TaqMan quantification probe, and 25ng of cDNA template. PCR amplification conditions were as follows:
Figure 1. Primers and probes designed for quantitative reverse tran-scriptase polymerase chain reaction (RT-PCR) of class II transactiva-tor (CIITA). (Upper panel) positions of the forward primers within thethree different 5' ends of the CIITA mRNAs and position of thecommon probe/common reverse primer in the second exon are shown.Coding regions are represented by wide boxes, and 5'-untranslatedregions by narrow boxes. The non-homologous regions are hatcheddifferently. Open bars represent the size (bp) of each amplicon andalso the synthesized oligo-nucleotides used for the amplification ef-ficiency control. Relevant sequences are given below the scheme.(Lower panel) amplification efficiency control for primer/probe setsdesigned to detect PI, PIII, and PIV products.
2824 Journal of the American Society of Nephrology J Am Soc Nephrol 14: 2823–2832, 2003
50°C for 2 min, 95°C for 10 min, then 40 cycles of 95°C for 15 s and62°C for 1 min. Emitted fluorescence for each reaction well wasmeasured every cycle during both the denaturation and annealing/extension phases. Amplification plots were constructed using ABIPRISM 7700 Sequence Detection System software version 1.7 (PEApplied Biosystems). Threshold cycle numbers (Ct) were determinedand transformed using the �Ct or ��Ct methods as described by themanufacturer, using hypoxanthine phosphoribosyltransferase (mH-PRT) as the calibrator. All PCR assays were performed in duplicatesand reported as the average. To estimate the total CIITA expression,we used a mixture of all three forward primers, the common reverseprimer, and the common probe. All data were expressed as relative tothe total CIITA expression in a control or sham kidney.
ResultsPrimers and Probes Used in Quantitative RT-PCR
To prevent amplification of the genomic DNA, we selectedspecific forward primer sequences for transcripts from eachclass II transactivator (CIITA) promoter that spanned exon-intron boundaries between the first and second exon. Se-quences for the common reverse primer and the common probewere selected from the second exon of CIITA that is shared byall isoforms (Figure 1A). We confirmed that the amplificationefficiency of the three primer/probe sets was similar by syn-thesizing three oligonucleotides corresponding to sequencesdetermined by our primer sets and using them as templates inthe assay (Figure 1B).
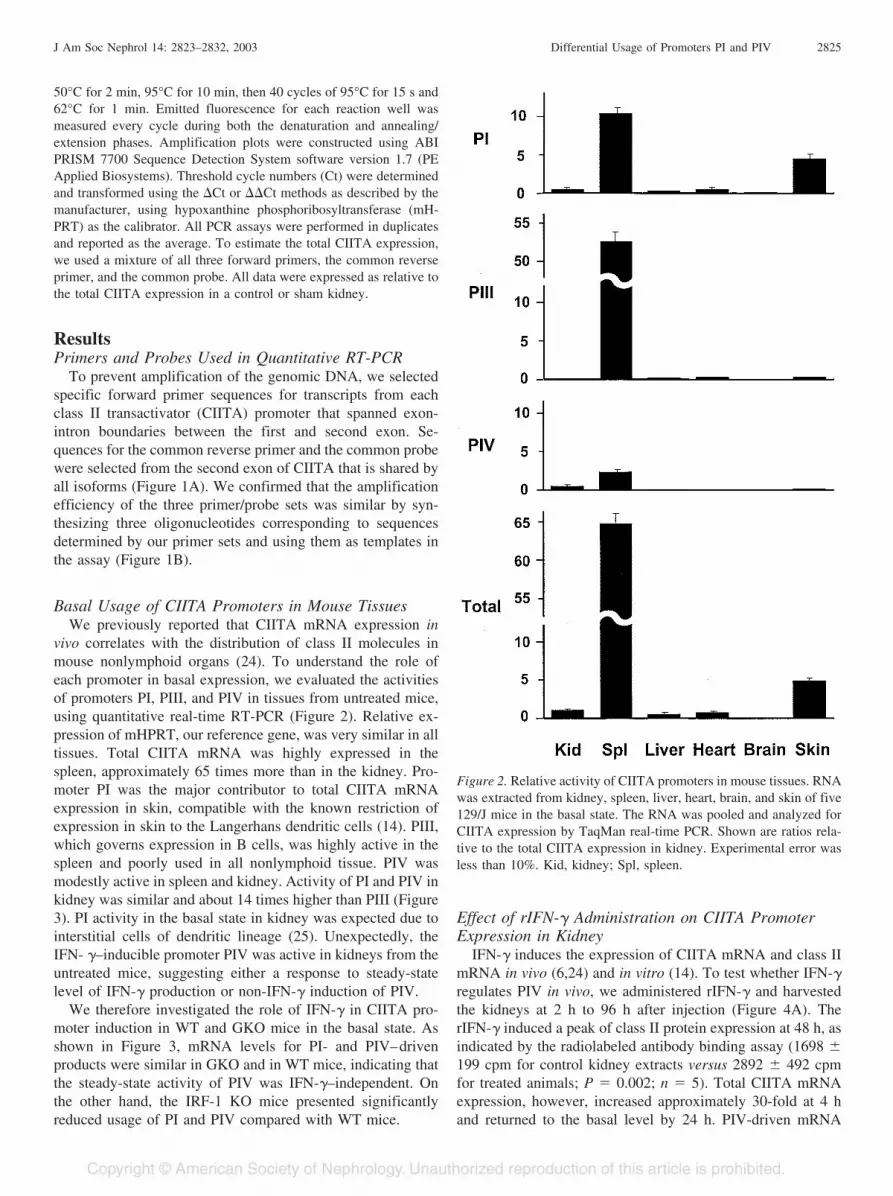
Basal Usage of CIITA Promoters in Mouse TissuesWe previously reported that CIITA mRNA expression in
vivo correlates with the distribution of class II molecules inmouse nonlymphoid organs (24). To understand the role ofeach promoter in basal expression, we evaluated the activitiesof promoters PI, PIII, and PIV in tissues from untreated mice,using quantitative real-time RT-PCR (Figure 2). Relative ex-pression of mHPRT, our reference gene, was very similar in alltissues. Total CIITA mRNA was highly expressed in thespleen, approximately 65 times more than in the kidney. Pro-moter PI was the major contributor to total CIITA mRNAexpression in skin, compatible with the known restriction ofexpression in skin to the Langerhans dendritic cells (14). PIII,which governs expression in B cells, was highly active in thespleen and poorly used in all nonlymphoid tissue. PIV wasmodestly active in spleen and kidney. Activity of PI and PIV inkidney was similar and about 14 times higher than PIII (Figure3). PI activity in the basal state in kidney was expected due tointerstitial cells of dendritic lineage (25). Unexpectedly, theIFN- �–inducible promoter PIV was active in kidneys from theuntreated mice, suggesting either a response to steady-statelevel of IFN-� production or non-IFN-� induction of PIV.
We therefore investigated the role of IFN-� in CIITA pro-moter induction in WT and GKO mice in the basal state. Asshown in Figure 3, mRNA levels for PI- and PIV–drivenproducts were similar in GKO and in WT mice, indicating thatthe steady-state activity of PIV was IFN-�–independent. Onthe other hand, the IRF-1 KO mice presented significantlyreduced usage of PI and PIV compared with WT mice.
Effect of rIFN-� Administration on CIITA PromoterExpression in Kidney
IFN-� induces the expression of CIITA mRNA and class IImRNA in vivo (6,24) and in vitro (14). To test whether IFN-�regulates PIV in vivo, we administered rIFN-� and harvestedthe kidneys at 2 h to 96 h after injection (Figure 4A). TherIFN-� induced a peak of class II protein expression at 48 h, asindicated by the radiolabeled antibody binding assay (1698 �199 cpm for control kidney extracts versus 2892 � 492 cpmfor treated animals; P � 0.002; n � 5). Total CIITA mRNAexpression, however, increased approximately 30-fold at 4 hand returned to the basal level by 24 h. PIV-driven mRNA
Figure 2. Relative activity of CIITA promoters in mouse tissues. RNAwas extracted from kidney, spleen, liver, heart, brain, and skin of five129/J mice in the basal state. The RNA was pooled and analyzed forCIITA expression by TaqMan real-time PCR. Shown are ratios rela-tive to the total CIITA expression in kidney. Experimental error wasless than 10%. Kid, kidney; Spl, spleen.
J Am Soc Nephrol 14: 2823–2832, 2003 Differential Usage of Promoters PI and PIV 2825

expression showed a similar pattern. Promoters PI and PIII didnot contribute to mRNA expression after IFN-� administration.
Role of IRF-1 in Basal and IFN-�–Induced Usage ofCIITA Promoters
Transcription factor IRF-1 is necessary for optimal inductionof CIITA mRNA and class II expression by rIFN-� (26). Toaddress the effect of IRF-1 on the activation of each CIITApromoter in vivo, we injected WT and IRF-1 KO mice withrIFN-� and harvested kidneys 4 h later, at the peak of CIITAmRNA expression. As expected, rIFN-� induced PIV-depen-dent and total CIITA mRNA only in the kidneys of WT mice(Figure 4B).
Class II induction in tubular epithelial cells is less in IRF-1KO kidneys (26). Figure 4B shows that rIFN-� induced littlePIV in kidneys of IRF-1KO mice. Thus the contribution ofIRF-1 to CIITA mRNA induction is attributable to its interac-tion with promoter PIV.
Effect of LPS on CIITA mRNA ExpressionLPS strongly induces class II mRNA and protein expression
in kidney epithelium (6,24). To explore the role of CIITA
promoters in this response, we administered LPS to BALB/cmice and examined their kidneys at various times (Figure 5A).LPS predominantly affected PIV, with distinct peaks of PIVinduction and total CIITA mRNA expression at 12 and 96 h. Asmall increase in PI mRNA usage occurred at 96 h, risingapproximately three-fold to five-fold above basal levels. PIIIwas not affected by LPS administration at any time.
We examined whether the increase in CIITA mRNA andPIV usage caused by LPS was IFN-�–dependent. WT andGKO mice were treated with LPS, and kidneys were analyzedat 12 and 96 h (Figure 5B). PIV-driven CIITA mRNA and totalCIITA mRNA were increased approximately 13-fold in WTmice in response to LPS injection at 12 h and 8-fold to 10-foldat 96 h. Kidneys of GKO mice showed little change in PIV ortotal CIITA expression at either time. GKO mice also failed toshow the small increase in promoter PI mRNA at 96 h. Thusthe major effect of LPS on CIITA and class II expression isprobably due to IFN-�–dependent induction of PIV.
Induction of CIITA Promoters by AllogeneicStimulation
Allostimulation strongly induces class II expression in kid-ney via the systemic effects of IFN-� and other cytokines (27).Unfortunately, it is difficult to study CIITA expression inkidney during kidney rejection because of the extreme diver-sity of cell types in the infiltrate. However, we previouslyshowed that rejection of the allogeneic tumor cells (P815) inthe peritoneal cavity induces a massive class II expression inkidney tubular epithelium and vascular endothelium by day 7,dependent on IFN-� and IRF-1 (6,26,28). We therefore exam-ined the systemic effect of allostimulation on CIITA promot-ers. WT, GKO, and IRF-1 KO mice were injected with P815cells on day 0, and the kidneys were harvested on day 7. In theWT kidney allogeneic stimulation induced total CIITA mRNAexpression and activated PIV 18-fold and 15-fold, respectively(Figure 6A). PI-dependent mRNA synthesis was smaller (ap-proximately 5-fold) (Figure 6A). There was no induction oftotal CIITA mRNA or PI-, PIII-, and PIV–related mRNA in thekidneys harvested from GKO mice, consistent with the previ-ous observations (6,24). IRF-1 KO mice, however, showed theinduction of total CIITA-, PI-, and PIV–dependent mRNA.The extent of induction was at least 50% of that observed inWT mice (Figure 6B). Thus CIITA induction during allostimu-lation correlates mainly with PIV induction but with a smallcomponent of PI induction.
IFN-� Independent Activation of CIITA Promoters inthe Kidney by Ischemic Injury
Renal injury induces expression of class II and CIITA,involving both IFN-�–dependent and IFN-�–independentmechanisms (6,29). We examined the effect of renal injury onindividual CIITA promoters. Left kidney was cross-clampedfor 60 min to induce ischemic acute tubular necrosis (ATN),while the untreated right kidney served as a control. When thekidneys were examined after 7 d, the injured left kidneyshowed approximately 3-fold increase in total CIITA mRNA inboth WT and GKO mice (Figure 7), consistent with the pre-
Figure 3. Basal activity of CIITA promoters in wild-type (WT),interferon-� (IFN-�)–deficient (GKO), and interferon regulatory fac-tor 1 (IRF-1)–deficient (IRF-1KO) mouse kidneys. Ten mice wereanalyzed in each group. Data are shown as means � SD. Expressionof CIITA in GKO mice was relative to BALB/c mice, whereasIRF-1KO mice were compared with 129/J mice.
2826 Journal of the American Society of Nephrology J Am Soc Nephrol 14: 2823–2832, 2003

viously reported increases in class II products (3,6). ATNinduced not only PIV-dependent mRNA but also PI-dependentmRNA. Even in GKO mice, the kidney with ATN displayedincreased levels of PI and PIV mRNA (Figure 7). Thus inischemic injury the induction of CIITA mRNA was due to theincrease of PI and PIV usage independent of IFN-�.
DiscussionClass II induction in kidney accompanies many disease
states and is one of the first examples of regulated geneexpression in kidney. In this study, we explored the expressionof individual CIITA promoters in kidney in the basal state andin response to stimuli known to induce class II. The behaviorof the CIITA promoters in vivo was consistent with their roles
previously suggested by studies in cell lines, but it also pointedto additional levels of control. The relative levels of promoter-specific mRNAs varied in different mouse tissues in the basalstate. Spleen expressed significant levels of mRNA from PIand PIII promoters, consistent with its content of dendritic cellsand B cells, whereas skin displayed only PI mRNA, consistentwith its high content of Langerhans cells of dendritic lineage.Similarly, kidney expressed promoters PI as expected from itscontent of class II–positive interstitial cells of dendritic lin-eage. On the other hand, both spleen and kidney showedexpression of the IFN-�–inducible PIV mRNA. At least inkidney this was independent of IFN-�, indicating that factorsother than IFN-� can induce PIV. Treatment with rIFN-� orstimuli that trigger endogenous IFN-� production induced high
Figure 4. Effect of rIFN-� on PIV activity in kidney. (A) Time course of recombinant IFN-� (rIFN-�)–dependent expression of CIITA.BALB/c mice were injected with 100,000 U of rIFN-�, and their kidneys were harvested at indicated times. Sh, sham kidney. (B) Dependenceof CIITA expression on IRF-1. 129/J and IRF-1 KO mice were injected with 100,000 U of rIFN-�, and the kidneys were harvested at 4 h.Expression of CIITA in IRF-KO mice was relative to 129/J mice. Five mice per each group were used, and the extracted RNA was pooled.Assays were done in duplicates. Experimental error was less than 10%.
J Am Soc Nephrol 14: 2823–2832, 2003 Differential Usage of Promoters PI and PIV 2827

levels of PIV mRNA in kidney. B cell–specific promoter PIII,on the other hand, showed negligible basal or induced mRNAin kidney. There were, however, novel forms of control in vivonot previously described: IFN-�–independent basal expressionof PI and PIV mRNA in kidney and IFN-�–independent acti-vation of PIV after ischemic injury.
Basal expression of CIITA mRNA in kidney was IFN-�–independent and was driven by promoters PI and, surprisingly,PIV. Immunostaining of mouse kidney in the basal state showsthat class II is confined to the interstitial cells, and is largelyIFN-�- and IRF-1–independent (6,30). Thus PI and PIV areprobably activated in the interstitial cells. Renal interstitialcells are related to dendritic cells, but most lack maturationmarkers such as DEC 205, �881/87 and �87–2 negative (datanot shown) (31,32). Moreover, mature dendritic cells are rarein normal kidney in the basal or induced states (33,34). Otherspecies may differ from mice in their use of the CIITA pro-
moters in nonlymphoid organs; for example, some humans andsome rat strains express class II in epithelial and endothelialcells (35).
It was surprising to find that the expression of class II inkidney in the basal state was independent of IFN-�, given thatexpression of class I molecules in mouse and human nonlym-phoid tissues is partially dependent on the steady-state produc-tion of IFN-� (3,4,6). Tissues such as kidney are exposedlocally to low levels of IFN-� in the basal state, but class II isnot induced. This is probably a dose issue; class I is morereadily induced by IFN-� than is CIITA and therefore class II.Nevertheless, PIV expression in kidneys in the basal state waspartially IRF-1–dependent, and it will be of interest to identifythis additional control mechanism and its relationship to theIFN-�–independent induction of PIV after tissue injury.
Transcriptional activation of the promoter PIV emerges asthe crucial step in CIITA induction in response to allostimu-
Figure 5. LPS-induced CIITA mRNA expression in WT and IFN-�–deficient (GKO) kidney. (A) BALB/c mice were injected with LPS (25�g), and their kidneys were harvested at the times shown. (B) BALB/c and GKO mice were injected with LPS, and the kidneys were harvestedat 12 h and 96 h. Five mice per each group were, used and the extracted RNA was pooled. Assays were done in duplicates. Experimental errorwas less than 10%. Sh, sham kidney.
2828 Journal of the American Society of Nephrology J Am Soc Nephrol 14: 2823–2832, 2003

lation, LPS, and IFN-� administration. It probably representsthe major mechanism of class II induction in nonlymphoidtissues in infections and graft rejection. The massive class IIexpression that is induced by systemic and local inflammatorystimuli in vivo is also completely dependent on IFN-� and willprobably prove to reflect PIV activation (6,26). Present dataalso provide evidence that in vivo PIV induction relies mainlyon IRF-1, consistent with our previous conclusion that CIITAmRNA and class II protein expression in kidney is largelydependent on IRF-1 (26,28). The residual PIV induction in theIRF-1KO mice after allogeneic stimulation may reflectSTAT1� binding to its cognate sequence in PIV to activatetranscription even in the absence of IRF-1. Thus phosphory-lated STAT-1 has a substantial contribution to IFN-�–induced
CIITA upregulation (36), although IRF-1 greatly amplifies thiseffect (18,37). Other IRF family members may partially com-pensate for IRF-1 deficiency in activating PIV transcription,particularly IRF-2 (38).
The present results indicate that the major mechanism ofCIITA and class II induction after renal ischemic injury isIFN-�–independent activation of PI and PIV. We have previ-ously shown that acute renal ischemic injury induced theexpression of MHC class II and CIITA, which persisted inGKO mice, indicating at least partial independence of IFN-�(6,29). It is possible that the PI mRNA component is in theinterstitial cells, due to enhanced infiltration (25), and the PIVcomponent is in the epithelium. Under some circumstances,IFN-� production by interstitial infiltrating cells may also
Figure 6. Induction of CIITA promoter IV in the kidney after allogeneic stimulation. (A) CIITA expression in IFN-�–deficient (GKO) and (B)IFR-1–deficient (IRF-KO) mice. BALB/c, GKO, 129/J ,and IRF-1 KO mice were injected with 20 � 106 P815 cells on day 0, and their kidneyswere harvested on day 7. Expression of CIITA in GKO mice was relative to Balb/c mice (A), and IRF-KO mice were compared with 129/Jmice (B). Five mice per each group were used, and the extracted RNA was pooled. Assays were done in duplicates. Experimental error wasless than 10%. Sh, sham kidney.
J Am Soc Nephrol 14: 2823–2832, 2003 Differential Usage of Promoters PI and PIV 2829

contribute (6), although we did not see this component in thisexperiment. We conclude that CIITA PIV promoter can beactivated in injured kidneys by an IFN-independent pathway, aresponse of particular interest in relationship to autoimmunityand allograft rejection. A strong candidate for IFN-independentPIV activation could be the JAK/STAT pathway that is asignificant factor in myocardial ischemia (39) and ischemicbrain injury (40). Injury-related PI activation, on the otherhand, could reflect accumulation of the class II–positive inter-stitial cells that occurs several days after renal injury. In fact,GKO mice show as many class II–positive interstitial cellsafter injury as do WT mice (6). Therefore the amount of PImRNA expressed in the organ can be attributed not only totranscriptional effects, but also to interstitial infiltration (41).One point to keep in mind is that, compared with the MHC
induction that accompanies allogeneic stimulation, LPS, orrIFN-� administration, these injury effects are weak (6).
The present studies in kidney show how the unusual arrange-ment of three promoters for one gene gives remarkable rangeof class II expression in cell types and in response to stimuli.This explains how circulating B cells can be strongly classII–positive regardless of their environment, how cells of thedendritic lineage can be strongly class II–positive but veryresponsive to environmental stimuli, and how epithelial cellscan express class II in times of infection, presumably to helprecognition of infected cells. Given the key role of class II inCD4 T cell recognition, this sophisticated arrangement mayoffer a number of advantages that could not be achieved witha single promoter. It will be of interest as genomics advancesto see how often this strategy has been employed and for whattypes of genes.
Given the role of class II - CD4 T cell interactions inautoimmunity and allograft rejection, the specific CIITA pro-moters could offer targets for selective control of class IIexpression in various cell types and tissue compartments, pos-sibly separating the functions of class II in the afferent andefferent limbs of T cell recognition. Having confirmed thatCIITA promoters operate in kidney as predicted, the next phasewill be to establish the functional importance of these compo-nents of CIITA expression. Strategies to selectively suspendthe functions of each promoter should now be devised, usingmutagenesis, inhibitory RNA, and potentially drugs. The factthat kidneys (42) and hearts (43) lacking CIITA show a reduc-tion in the severity of rejection, despite becoming infiltratedand showing pathologic lesions, support our belief that class IIexpression has functional effects in transplantation. The iden-tification of the individual transcripts could also be useful inmonitoring the components of CIITA transcription and thus ofclass II transcription in disease processes.
AcknowledgmentsThis research has been supported by operating grants from the
Canadian Institutes of Health Research, the Roche Organ TransplantResearch Foundation, the Kidney Foundation of Canada, NovartisPharmaceuticals Canada, Inc., The Muttart Foundation, and TheRoyal Canadian Legion. Dr Halloran holds a Canada Research Chairin Life Sciences Related to Human Health and Disease.
References1. Benoist C, Mathis D: Selection for survival? Science 276: 2000–
2001, 19972. Kirberg J, Berns A, Von Boehmer H: Peripheral T cell survival
requires continual ligation of the T cell receptor to major histo-compatibility complex-encoded molecules. J Exp Med 186:1269–1275, 1997
3. Takei Y, Sims TN, Urmson J, Halloran PF: Central role forinterferon-gamma receptor in the regulation of renal MHC ex-pression. J Am Soc Nephrol 11: 250–261, 2000
4. Tellides G, Tereb DA, Kirkiles-Smith NC, Kim RW, Wilson JH,Schechner JS, Lorber MI, Pober JS: Interferon-� elicits arterio-sclerosis in the absence of leukocytes. Nature 403: 207–211,2000
Figure 7. Activation of CIITA promoters after ischemic injury. Isch-emic injury to the left (L) kidney was inflicted for 60 min and wascompared with the unclamped right (R) kidney. Control mice under-went a simple laparotomy under identical conditions. The kidneyswere harvested on day 7. Five mice per each group were used, and theextracted RNA was pooled. Promoter activity is given as a ratiorelative to a sham right kidney in each group. Assays were done induplicates. Experimental error was less than 10%. ATN, acute tubularnecrosis.
2830 Journal of the American Society of Nephrology J Am Soc Nephrol 14: 2823–2832, 2003

5. Skoskiewicz M, Colvin RB, Schneeberger EE, Russell PS: Wide-spread and selective induction of major histocompatibility com-plex-determined antigens in vivo by � interferon. J Exp Med 162:1645–1664, 1985
6. Goes N, Sims T, Urmson J, Vincent D, Ramassar V, HalloranPF: Disturbed MHC regulation in the interferon-� knockoutmouse. J Immunol 155: 4559–4566, 1995
7. Chang CH, Gourley TS, Sisk TJ: Function and regulation of classII transactivator in the immune system. Immunol Res 25: 131–142, 2002
8. Chang CH, Fontes JD, Peterlin M, Flavell RA: Class II transac-tivator (CIITA) is sufficient for the inducible expression of majorhistocompatibility complex class II genes. J Exp Med 180: 1367–1374, 1994
9. Chang CH, Guerder S, Hong SC, van Ewijk W, Flavell RA: Micelacking the MHC class II transactivator (CIITA) show tissue-specific impairment of MHC class II expression. Immunity 4:167–178, 1996
10. Steimle V, Siegrist CA, Mottet A, Lisowska-Grospierre B, MachB: Regulation of MHC class II expression by interferon-� me-diated by the transactivator gene CIITA. Science 265: 106–109,1994
11. Koonin EV, Aravind L: The NACHT family - a new group ofpredicted NTPases implicated in apoptosis and MHC transcrip-tion activation. Trends Biochem Sci 25: 223–224, 2000
12. Ting JP, Trowsdale J: Genetic control of MHC class II expres-sion. Cell 109[Suppl]: 21–33, 2002
13. Lennon AM, Ottone C, Rigaud G, Deaven LL, Longmire J,Fellous M, Bono R, Alcaïde-Loridan C: Isolation of a B-cell-specific promoter for the human class II transactivator. Immuno-genetics 45: 266–273, 1997
14. Muhlethaler-Mottet A, Otten LA, Steimle V, Mach B: Expres-sion of MHC class II molecules in different cellular and func-tional compartments is controlled by differential usage of mul-tiple promoters of the transactivator CIITA. EMBO J 16: 2851–2860, 1997
15. Piskurich JF, Wang Y, Linhoff MW, White LC, Ting JPY:Identification of distinct regions of 5' flanking DNA that mediateconstitutive, IFN-�. STAT 1 and TGF-�-regulated expression ofthe class II transactivator gene. J Immunol 160: 233–240, 1998
16. Ghosh N, Piskurich JF, Wright G, Hassani K, Ting JPY, WrightKL: A novel element and a TEF-2-like element activate themajor histocompatibility complex class II transactivator in B-lymphocytes. J Biol Chem 274: 32342–32350, 1999
17. Landmann S, Muhlethaler-Mottet A, Bernasconi L, Suter T,Waldburger JM, Masternak K, Arrighi JF, Hauser C, Fontana A,Reith W: Maturation of dendritic cells is accompanied by rapidtranscriptional silencing of class II transactivator (CIITA) ex-pression. J Exp Med 194: 379–391, 2001
18. Piskurich JF, Linhoff MW, Wang Y, Ting JPY: Two distinctgamma interferon-inducible promoters of the major histocompat-ibility complex class II transactivator gene are differentiallyregulated by STAT1, interferon regulatory factor 1, and trans-forming growth factor beta. Mol Cell Biol 19: 431–440, 1999
19. Waldburger JM, Suter T, Fontana A, Acha-Orbea H, Reith W:Selective abrogation of major histocompatibility complex class IIexpression on extrahematopoietic cells in mice lacking promoterIV of the class II transactivator gene. J Exp Med 194: 393–406,2001
20. Dalton DK, Pitts-Meek S, Keshav S, Figari IS, Bradley A,Stewart TA: Multiple defects of immune cell function in mice
with disrupted interferon-gamma genes. Science 259: 1739–1742, 1993
21. Matsuyama T, Kimura T, Kitagawa M, Pfeffer K, Kawakami T,Watanabe N, Kündig TM, Amakawa R, Kishihara K, WakehamA, Potter J, Furlonger CL, Narendran A, Suzuki H, Ohashi PS,Paige CJ, Taniguchi T, Mak TW: Targeted disruption of IRF-1 orIRF-2 results in abnormal type I IFN gene induction and aberrantlymphocyte development. Cell 75: 83–97, 1993
22. Jephthah-Ochola J, Urmson J, Farkas S, Halloran PF: Regulationof MHC in vivo. Bacterial lipopolysaccharide induces class I andII MHC products in mouse tissues by a T cell independent,cyclosporine sensitive mechanism. J Immunol 141: 792–800,1988
23. Heid CA, Stevens J, Livak KJ, Williams PM: Real time quanti-tative PCR. Genome Res 6: 986–994, 1996
24. Ramassar V, Goes N, Hobart M, Halloran PF: Evidence for thein vivo role of class II transactivator in basal and IFN-� inducedclass II expression in mouse tissue. Transplantationt 62: 1901–1907, 1996
25. Austyn JM, Hankins DF, Larsen CP, Morris PJ, Rao AS, RoakeJA: Isolation and characterization of dendritic cells from mouseheart and kidney. J Immunol 152: 2401–2410, 1994
26. Hobart M, Ramassar V, Goes N, Urmson J, Halloran PF: IFNregulatory factor 1 (IRF-1) plays a central role in the regulationof the expression of class I and II MHC genes in vivo. J Immunol158: 4260–4269, 1997
27. Halloran PF, Autenried P, Ramassar V, Urmson J, Cockfield S:Local T cell responses induce widespread MHC expression.Evidence that IFN-� induces its own expression in remote sites.J Immunol 148: 3837–3846, 1992
28. Hobart M, Ramassar V, Goes N, Urmson J, Halloran PF: Theinduction of class I and II major histocompatibility complex byallogeneic stimulation is dependent on the transcription factorinterferon regulatory factor 1 (IRF-1). Observations in IRF-1knockout mice. Transplantation 62: 1895–1901, 1996
29. Sims TN, Goes NB, Ramassar V, Urmson J, Halloran PF: In vivoclass II transactivator expression in mice is induced by a non-interferon-gamma mechanism in response to local injury. Trans-plantation 64: 1657–1664, 1997
30. Halloran PF, Goes N, Urmson J, Ramassar V, Hobart M, Sims T,Lui SL, Miller LW: MHC expression in organ transplants: les-sons from the knock-out mice. Transplant Proc 29: 1041–1044,1997
31. Sallusto F, Lanzavecchia A: Mobilizing dendritic cells for toler-ance, priming, and chronic inflammation. J Exp Med 189: 611–614, 1999
32. Palucka KA, Taquet N, Sanchez-Chapuis F, Gluckman JC: Den-dritic cells as the terminal stage of monocyte differentiation.J Immunol 160: 4587–4595, 1998
33. Saleem M, Sawyer GJ, Schofield RA, Seymour ND, GustafssonK, Fabre JW: Discordant expression of major histocompatibilitycomplex class II antigens and invariant chain in interstitial den-dritic cells. Implications for self-tolerance and immunity. Trans-plantation 63: 1134–1138, 1997
34. Jevnikar AM, Singer GG, Coffman T, Glimcher LH, Rubin-Kelley VE: Transgenic tubular cell expression of class II issufficient to initiate immune renal injury. J Am Soc Nephrol 3:1972–1977, 1993
35. von Willebrand E, Lautenschlager I, Inkinen K, Lehto VP, Vir-tanen I, Hayry P: Distribution of the major histocompatibilitycomplex antigens in human and rat kidney. Kidney Int 27:616–621, 1985
J Am Soc Nephrol 14: 2823–2832, 2003 Differential Usage of Promoters PI and PIV 2831

36. Barbaro AD, Tosi G, Frumento G, Bruschi E, D’Agostino A,Valle MT, Manca F, Accolla RS: Block of Stat-1 activation inmacrophages phagocytosing bacteria causes reduced transcrip-tion of CIITA and consequent impaired antigen presentation. EurJ Immunol 32: 1309–1318, 2002
37. Xi HK, Eason DD, Ghosh D, Dovhey S, Wright KL, Blanck G:Co-occupancy of the interferon regulatory element of the class IItransactivator (CIITA) type IV promoter by interferon regulatoryfactors 1 and 2. Oncogene 18: 5889–5903, 1999
38. Xi HK, Goodwin B, Shepherd AT, Blanck G: Impaired class IItransactivator expression in mice lacking interferon regulatoryfactor-2. Oncogene 20: 4219–4227, 2001
39. Mascareno E, El Shafei M, Maulik N, Sato M, Guo Y, Das DK,Siddiqui MAQ: JAK/STAT signaling is associated with cardiacdysfunction during ischemia and reperfusion: Circulation 104:325–329, 2001
40. Takagi Y, Harada J, Chiarugi A, Moskowitz MA: STAT1 isactivated in neurons after ischemia and contributes to ischemicbrain injury: J Cereb Blood Flow Metab 22: 1311–1318, 2002
41. Dieu MC, Vanbervliet B, Vicari A, Bridon JM, Oldham E,Aït-Yahia S, Brière F, Zlotnik A, Lebecque S, Caux C: Selectiverecruitment of immature and mature dendritic cells by distinctchemokines expressed in different anatomic sites. J Exp Med188: 373–386, 1998
42. Sims TN, Afrouzian M, Urmson J, Zhu LF, Halloran PF: Therole of the class II transactivator (CIITA) in MHC class I and IIregulation and graft rejection in kidney. Am J Transplant 1:211–221, 2001
43. Brickey WJ, Felix NJ, Griffiths R, Zhang J, Wang B, PiskurichJF, Itoh-Lindstrom Y, Coffman TM, Ting JP: Prolonged survivalof class II transactivator-deficient cardiac allografts. Transplan-tation 74: 1341–1348, 2002
2832 Journal of the American Society of Nephrology J Am Soc Nephrol 14: 2823–2832, 2003
Copyright © 2022 FDOKUMEN




















