
Direct and indirect effects in the regulation of overlapping promoters
Upload
khangminh22Category
view
0download
0
Evaluation of promoters and visual markers fortransformation of eastern white pine
ALLAN ZIPF’. ALEX M. DINER’, RUFINA WARD’. SUtiAN.BHARARA-I. GEORGE BROWN. JR.‘. R. NAGMANI’, L.K. PAREEK’ancl GOWIND C. SHARMA”I DeJmrtmet# of Pkmt und Soil Science. Aldwmu A&M Uikwdty. Nmmul. AL 35262. USA(r-ntuif: uwn~pO1@cedu): 1 USDA Ftsst &p&v. SInttJtem Jnsritutr of FIIIPS~ Getterics.G~tinesville, FL 326 J 1. USA: 3 Dep~rtmtw f tfSume+ Unisersity c~Alu~tnrr-Binnill~h~ni.Birmbt&nn. AL 35294, USA: JDe~~rtn~ent r# Botany, University cf Rujawhun. In&u
Aaqted 29 September 20 ’
Mshaet. This report .serv& to evahate possible promo&s ‘for use in the production oftrmsgenic eastern white pine (P&H stmbus L.). Embryogenic cuttwrrs of eastern whitepine were bombarded with gold.ptuticles coated separately with a variety of gene cons-containinp the Ui& ,&glwcwroni&se (GUS) or green fl~omsc~~ profein (GFP) reportergne. Transient expression of the UidA gene. driven by ;t novet al&11 virus adenine methyltransferuse gene promoter. as well as live other promoters q.sed in imgiosperm tmnsfornmtion.were evulwuted. The maize alcohol- dehydrogenase promoter was not effective in easternwhite pine cultures. The constrwct with th& doubled Cauliflower Mosaic Virus 35s prorhoterply Alfalfa Mosaic Virus enhancer showed the highest levels of expression. GUS expressionwas detected within 14 hours. but deereused after 5 days imd.was MN detectable I5 daysafter bombardment. Expression of GUS activity wa.s recorded’muiniy in sorrutic embryonriheads of wwious stages of development and acasionulty in suspensor cells. Similar to GUSexpression. moditied green Huoresccnt protein (GFP) wi~*.detected in the embryonul headceils 7-4 hours after bombardment. GFP-expressing susp&or cells were both more infrequentand difficult tu detect. w their highly vilcuolatc nature rendered the GFP presence less visibleagainst the yellow buckgruwnd ;rutoliworesc~nce.
Key words: hiolistics. GFP:GUS. Pirm .W&I.V L.. transient gene expression
Ahhreviations: hcnzylnminopurin. BAP: cawlillower musuic virus 3% pmmotcr. 3%:
1.CJiuhlorophcnoxy;lcclic acid. X-I-0: green Iluor~~n~ pnacin. GFPz P-glucunmidase.GUS: plttnt gn)wth rcgulntor. PGR
Introduction
Micn)projcctile-mediated DNA tr;lnsi’er has ken shown to be affective for avariety oi’ woody angiosperms and gymnosptxms. Gymnosperm tissues usedas biolistic targets huvl: included cotyledons (Stomp et al. I99 I 1. vegctutive

4 6
buds and bud-derived calluses (Aronen et al. 1994), mature poilen (Hay et al.1994: Li et al. 1994), xylem (Loops&a et al. .1992). and somatic embryogeniccultures (e.g. Duchesne and Charest 1991; Robertson et al. 1992; Bommineniet al. 1994; Walter et al. 1994; Clapham et al. 1995). Expression has beenmost commonly transient, for several possible reasons (Clapham et al. 1995);however, stable incorporation has been documented in a few conifer species(e.g. Ellis et al. 1993; Charest et al. 1996)..
Surveys are necessary for finding promoters appropriate during the selec-tion process as well as for driving successful expression of the gene of interestin the plant. Early reports of conifer transformation utilized promoters thatwere most effective in dicotyledonous systems, especially the CauliflowerMosaic Virus 35s promoter or modified versions (e.g. Stomp et al. 1991;Campbell et al. 1992; Shin et al. 1994). Monocot promoters have also heeninvestigated, and have shown varying degrees of effectiveness (Ellis et al.1991 vs. Loopstra et al. 1992; Walter et al. 1994).
This study evaluates several promoters, including an algal virus genepromoter not previously reported for use in the genetic manipulation of treespecies, In addition, two commonly used visual markers, Green FluorescentProtein (GFP) and p-glucuronidase (GUS), were compared for their utility ineastern white pine transformation.
Materials and methods
Preparation of embvogenic tissues for bombardment
Embryogenic cultures of eastern ‘white pine, Pinus strobus L., were initi-ated and maintained for 2 years as described by Kaul (1995). Immatureseeds were surface-sterilized in 10% CloroxT” for 10 min and then rinsedthree times in sterile water. Excised megagametophytes were soaked in 5%CloroxTM for 5 min, rinsed and then transferred to DCR medium (Becwaret al. 1990) containing 12 I’M 2,4-dichlorophenoxyacetic acid (2.4-D) and4 “M benzylaminopurine [BAP], and solidified with 0.2% Gelrite (Sigma,St. Louis, MO). As the cones came from open-pollinated trees, a culturearising from each megagametophyte was designated as an individual geno-type line (e.g. WPIB2. WP/7) and maintained separately on gelled medium.Embryogenic cultures were separated from their respective megagameto-phytes after S-10 weeks and sub-cultured at two or three week intervalsfor 4 months. They were then transferred to Brown and Lawrence medium(Brown and Lawrence 1968) containing 73 mM (2.5%) sucrose. 0.65% agar(Sigma, St. Louis. MO) and modified to contain L-glutamine ( 10 mM) as the

47
sole source of amino nitrogen. The PGRs 2,4-D (8 “M) and BAP (4 ‘“M)were added. and the pH was adjusted to 5.8. All cultures Were incubatedin the dark at 20-22 ‘C, and subcultured every three weeks. Embryogeniccultures were transferred to fresh medium 5-7 days prior to bombardmentand spread uniformly to a circle of approximately 2-3 cm in diameter. Oneplate per treatment was used as a nonbombarded control. For the genotypeexpression experiment. matched replicate cultures of two lines, identified asWP/B2 and WP/7. respectively, were prepared for each bombardment usingcommon pools of DNA-coated particles, as was a nonbombarded control.Three diverse constructs were utilized in this experiment.
Micmpmjectile bombardment
Procedures used for 1 l”rn gold particle microprojectile preparation andcoating with DNA were according to Heiser (1992). Each culture wasbombarded twice with I fig of construct DNA per macrocarrier disk.Each plate was rotated 90” between bombardments. Bombardment with theBiolisticTM particle delivery system PDS-1OOOMe (DuPont, Wilmington,DE) employed a rupture disc pressure of 1100 psi and a sample distance of5 cm. Following bombardment, the cultures were incubated in petri dishes inthe dark at 20-22 “C.
Constructs
The following constructs, all containing the GUS reporter gene (Jeffersonet al. 1987). were used (Figure 1): 1) pAHC25, driven by the maizeubiquitin Ubil promoter and first intron (Christensen et al. 1992), providedby Dr. Peter Quail, USDA/ARS, Plant Gene Expression Center, Albany,CA; 2) pAIl GUS,Dy 10s. driven by the maize alcohol dehydrogenase (adhl)promoter and first intron, provided by Dr. Ann Blechl, USDA/ARS, CropImprovement and Utilization Research Unit, Albany. CA; 3) pAMTGUS25,driven by the Chiorella virus adenine methyl transferase gene promoter(Mitra and Higgins 1994) plus adhl first intron, provided by Dr. Amit Mitra.Department of Plant Pathology. University of Nebraska, Lincoln, NE; 4ajpMON752, driven by a chime& double 35s promoter plus adhl intron 1,and 4b) pMON 18350. driven by a chimeric double 35s promoter. providedby Dr. Tim Conner and Dr. M.E. Fromm. Monsanto Co., St. Louis, MO and 5)pB1426. driven by a double 35s promoter plus an alfalfa mosaic virus (AMV)enhancer sequence (Datla et al. 1991). provided by Dr. Raju Datla, NationalResearch Council Canada. Plant Biotechnology Institute. Saskatoon, Saskat-chewan. The plasmid pHBT-SGFP-TYG-NOS, containing a synthetic GFP

I Y
pBI426
Ubi 1. . . . . . . . . . .
GUS gf&&gzg
phHC25
I) adh
,.:.:.:.:.:.~~:.:.:.:.:GUS $~&$gii;. . . . . . . . . . .
pAI1(;US,Dplfk
amt
GFP
Fi,grrrc !. Promoter constructs used IO ~ucly visual mnrkcr cxprcssion in eastern white pintcmbryogcnic cultures. Regions arc not to scnlc rind XC l’or illustrative purposes only. The“cnhanccr” is transcriptional in the cast ot’ the MS cnhanccr ancl translational in the case 01the d11111:,1 mosaic virus f AMV) cnhanccr.

49
mutant gene driven by a hybrid 35s enhancer - maize CJ pyruvate orrh-phosphate dikinase I @p&l) promoter (Sheen 1993). was provided by Dr. J.Sheen. Department of Molecular Biology, Massachusetts General Hospital.Boston. MA.
Visd marker custtys
‘J. Histochemical GUS wwySamples of approximately one quarter of the target circle were assayedaccording to Jefferson et al. (1987) I day after bombardment. In a smallerstudy. select calli bombarded with two promoter constructs were sampled 5and I5 days after bombardment. Both isolated individual cells and clusters ofblue-stained ceils were counted as singfe GUS-expressing sites.
2. GFP$uorescence assuySingle quadrants of those cultures bombarded with the GFP construct wereassayed using a fluorescent microscope (Olympus America, Melville, NY).Samples were excited by blue light (h,,, = 490 nm) and observed via anOlympus BP490 filter cube. Numbers of green fhrorescing cells were countedfor only samples collected I day after bombardment.
Statistics
Analysis of variance and Duncan’s Multiple Range Tests of GUS-positiveFoci (“blue spot”) frequency were performed using the SAS System forWindows, Version 6. I2 (SAS Institute, Cat-y, NC). GUS expression driven bythe different promoters was analyzed in a completely randomized design withunequal replications. Genotype-dependent expression (WP/B2 vs. WP/7) wasanalyzed using a randomized complete block design.
Results and discussion
All embryogenic cultures expressed GUS activity 24 hours after bombard-ment. Photomicroscopy of cells showed both multiple GUS blue-colored“hits” on single embryos, and numerous isolated individual or small clumpsofcells showing the same characteristic blue color (Figures ?A. BJ. Occasion-ally suspensor cells expressed GUS (Figure ?A). Twenty-four-hour assaysshowed intense GUS activity in single cells as well as in small clusters ofj-6 cells, probably arisin, (r from mitosis after successful bombardment and

50
nuclear incorporation. In contrast, a light blue color frequently extended froma transformed cell to the neighboring cells, probably due to GUS gene productdiffusion. Similar observations were also reported in bombarded cotyledonsof loblolly pine (Pilzus me&) (Stomp et al. 1991).
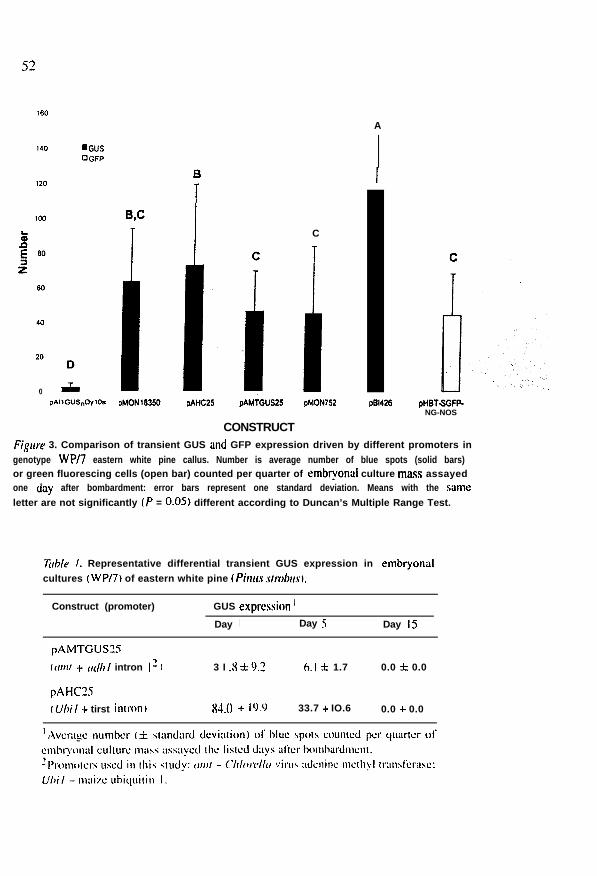
All promoters tested showed some level of GUS expression, based onnumbers of blue-colored sites (Figure 3). while endogenous activity wasnever detected in any bombarded or nonbombarded control. Based onthe average number of GUS-expressing sites. the most effective of thosepromoters tested in white pine embryogenic tissue was the double-3%plus AMV enhancer (pBI426). This was followed by the Ubil promoter +first intron (pAHC25) and the double 3% promoter (pMONl8350) whichwas not significantly different from the arnt promoter plus u&Z intron1 (pAMTGUS25) and the double 35s plus a&l intron 1 (pMON752)which was significantly different from the adhl promoter + first intron(pAIiGUS,DylOs) (Figure 3). The adh promoter exhibited extremely lownumbers of GUS foci as was also seen in bombarded white spruce (Picagfuuca) embryogenic calluses (Ellis et al. 1991). Several double 3%promoters and their derivatives have been tested previously in other coniferspecies (e.g. Bekkaoui et al. 1990; Newton et al. 1992; Charest et al. 1993;Bommineni et al. 1994: Walter et al. 1994). In general 3% and double 35stypes were characterized by medium to strong expression, similar to ourstudy, with double 3% types outperforming the single promoter constructs. Adicot (sunflower) polyubiquitin promoter achieved the highest GUS expres-sion in pollen of Norway spruce (Picen a&es) and Scats pine (Pinussylvesrris) (Haggman et al. 1997). Although the functional promoter regionis organized differently, the maize ubiquitin promoter induced similarly highlevels of GUS expression in our system. Apparently the presence of a 5’ intronhas little effect on gene expression in gymnospermous systems.
The novel promoter. am& is an 85 1 bp upstream region from a Chlorellavirus methyl transferase gene that has been shown to be effective for expres-sion in transformants of a small number of both monocots and dicots (Mitraet al. 1994). No applications of this promoter to tree transformation have beenreported in the literature.
Typically, transient GUS activity decreased within 5 days after bombard-ment and whole tissue assays showed no GUS activity after 15 days (TableI). There was no indication of plasmid gene incorporation without chemical(antibiotic. herbicide) selection. as has been seen in transformations of otherplant species (McCabe and Martinell 1993).

.
0~AliGUS,OylOs
=GUSOGFP
B,C
ipMONlS350
A
C
ipAHC25 PAMTGUSZS WON752 pm426
CONSTRUCT
pHST.SGFP-NG-NOS
Fign’r 3. Comparison of transient GUS and GFP expression driven by different promoters ingenotype WP/7 eastern white pine callus. Number is average number of blue spots (solid bars)or green fluorescing cells (open bar) counted per quarter of embryonai culture rna$s assayedone day after bombardment: error bars represent one standard deviation. Means with the sameletter are not significantly (P = 0.05) different according to Duncan’s Multiple Range Test.
Tirhle 1. Representative differential transient GUS expression in embryonulcultures (WPL’) of eastern white pine (Pinus s/,ahtrst.
Construct (promoter)
pAMTGUS’1.5
I(tr,~f + &I/ intron I2 1
pAHC3.5
(iJIG + tirst inn-on)
GUS expression ’
Day I Day 5
3 I .x rt 0.1 6.1 * 1.7
x-L.0 + 19.5, 33.7 + IO.6
Day I5
0.0 * 0.0
0.0 + 0.0

53
T&& It/. Analysis of variance (ANOVA) for transient GUS expression intwo eastern white pine genotypes bomburded with pAHC75 (I/k/ + intron).pMON 18350 (double 35s) or pBI-I26 [double 35s + AMV enhancer).
Source l l f Mean squares
Replication 2 543.1 NS
Construct 2 16207. I”***
Genotype I .5379~Q***”-.
Construct x Genotype 3. 296.2 NS
Error 64 426. I
NS; **+* = not significant and signiticant at (P = O.OOOl j level. respectively.
Table Zh. Effect of eastern white pine genotype on transient GUS expressionafter bombardment with pAHC?S (Uhil + intron). pMON IS350 [double 3%)or pB1426 (double 3.5s + AMV enhancer).
Genotype N Mean number of **hits”
WPff 36 84.2 AWPIB2 36 29.5 B
Means with the same letter are not significantly different (P = 0.05j accordingto Duncan’s Multiple Range Test.
Genotype-dependent expression
Transient expression of the GUS reporter gene consistently varied betweenthe two lines of eastern white pine (Pimu strohus L.) embryogenic cultureswhen either pB1426 or pAHC25 or pMONl8350 were used for bombard-ment. Analysis of variance of data from the randomized complete blockdesign revealed no significant Construct x Genotype interactions, but thatonly the construct and genotype were significant (Table 2a). GUS expres-sion in the bombarded somatic embryo masses of WP/B2 was significantlydifferent from that of bombarded WP/7 cultures. based on Duncan’s MultipleRange Test (Table 2b).
This is the tirst report of genotype differences in transgene expressionin eas tern white pine tissues. Genotype variability in reporter gene expres-sion has been noted in other conifer species. Variation of genotype responseto bombarded GUS-containing plasmids has been noted in mature tissues(Aronen et al. 1995) and cultured cells derived from mature tissues (Aronenet al. 1994) of Scats pint (Pimu s$ve.srris). Embryogenic cell lines ofPicecr mlriuw also showed genotype variability when bombarded. with onefamily showing no GUS gene activity at all (Duchesne and Charest I99 I ).

54
Bombarded embryogenic suspensions of Picea nbies had higher levels ofGUS expression than embryogenic callus or zygotic embryos (Newton etal. 1992). Several lines of embryogenic cultures of L,atft- hricina were alsobombarded with various GUS constructs resulting in varying expressionlevels (Klimaszewska et al. 1997).
The reason for the difference in transgene expression in WP/B2 vs. WP/7must lie at some fundamental level. as GUS expression driven by bothmonocot and dicot promoters was similarly reduced. The cultures of bothlines were morphologically similar in texture. color and state of development.Both cultures grew at similar rates and so there were no obvious differencesto account for the variation in expression.
GFP expression
A construct using a modified green fluorescent protein (GFP) sequence (Chiuet al. 1996) was also employed to test the utility of GFP vs. GUS as a visualmarker system for eastern white pine transformation. Embryogenic easternwhite pine cells autofluoresce a bright yellow when excited with long UVwavelengths. In work reported here, GFP expression was detectable 24 hoursafter bombardment, primarily in cells of the embryonal head (Figures 2C,D). Transiently transformed suspensor cells were more difficult to detect,as their highly vacuolate nature rendered the GFP less detectable. Althoughsuspensor cells occupy a greater volume of the somatic embryo and thereforea greater volume of the embryogenic mass. GFP expression by the suspensorcells within that mass was infrequent. Suspensor cells are a magnitude largerthan the embryonal head cells and thus, the nucleus occupies a much smallerproportion of the cell volume than does a nucleus of an embryonal head cell.In fact, the compact nature of the embryonal head makes it more likely that amicroprojectile will somewhere penetrate a nucleus, accounting for the highernumber of GFP-expressing sites in this region of the somatic embryo.
GFP expression rapidly diminished with time and became very difficult todetect even only 3 days after bombardment. GFP, both wild-type and modi-fied. has been expressed in black spruce (Picea marha). white spruce (Piceaglauca) and eastern white pine (Tian et al. 1997).
The use of GFP as visual marker for transgenic plants (Leffel et al.1997) can only be used after overcoming several procedural constraints. First,samples may not be directly viewed for GFP while contained in plastic (petridish) vessels, since the plastic reduces the intensity of the effective excitatorywavelengths. Second. few fluorescent microscopes have sufficient stage clear-ance for observation of undisturbed transformed cultures. In our laboratory,the samples had to be flattened in order to spread the cells for visualizationwhich disrupted the culture integrity and any gross morphological structures

5 5
were disaggregated. Third, the GFP fluorescence was easily detectable under100X magnification but was less discernible at 40X. The higher magnificationgreatly lengthened the time for collecting data when compared to assaying forGUS expression which can be visualized under lower magnifications.
However, the detection of GFP is more expedient and inexpensive thantesting for GUS expression, since the latter requires both an overnight incuba-tion as well as an expensive substrate. If GFP expression can be maintained atadequately high levels to be distinguishable from the autofluorescence, it willbe an appropriate selectable marker for eastern white pine transformation.
Distinctive expression in coniferous tissues
As noted by Walter et al. (1994) tissues of Pinus rudiatu exhibit theuncommon property of responding to both monocot and dicot promoters.Expression in eastern white pine embryogenic cultures follows a similarpattern, as strong expression of GUS was found when driven by both amonocot-optimized (pMON752 - 35S-3% + adhl intron) and a dicot-optimized (pB1426 - 35S-35s + AMY enhancer) promoter. In addition, theplant viral promoters (35S, amf) can also be as effective as the angiospermpromoters, suggesting that conifers can serve as tools for deciphering thecontrol and specificity of transcription.
Apparently the conifer transcription/translation machinery may be moretolerant of variability, to still allow expression of introduced genes.For example, the presence of a 5’ intron (Figure I), characteristic ofmonocot constructs. is apparently not a required element or a regulatoryfactor for conifer translation. This is evident from the higher .expressionof the U&l-driven GUS construct (pAHC25) than the adhl-driven one(pAIiGUS,Dy lOs), although both constructs have a similarly positionedintron. Conversely, the presence of the intron did not disrupt expression,as the 35S-35s + adhl intron-driven construct (pMON752) generated highnumbers of GUS foci as did the 35S-35S-driven construct (pMON18350).which lacked such an intron. Promoter deletion analysis could provide addi-tional clues to sequences critical for control of expression in conifer systemsand, possibly, for elements universal to the transcription/translation process.
Acknowledgements
We are grateful to the three anonymous reviewers for their critical commentsand helpful su,,cynestions. We would like to thank Hamidou Sakhanokho andCeleste Bell for help with the statistical analysis and data entry, respectively.

56
Department of Plant and Soil Science. Alabama A&M University,Normal, AL 35762. Contributed by the Agricultural Experiment Station,Alabama A&M University, Journal No. 396. This work was supportedby the U.S.D.A. Historically Black Colleges and Universities Cooper-ative Research Program grants 19-92-O 14. 19-92-049. and 19-95-057 andby U.S.D.A./Forest Service Center of Excellence grant SRS 29-G-96-102.Mention of a trademark. proprietary product or vendor does not constitute a<guarantee or warranty of the product by’the U.S. Department of Agricultureand does not imply its approval to the exclusion of other products or vendorsthat may also be suitable.
References
Aronen, 71. Haggman. H. and Hohtala. A. 1994. Transient beta-giucuronidase expression inScats pine tissues derived from mature trees. Can. J. For. Res. 24: 3006-20 I 1.
Aronen. T.. Hohtola. A.. Laukkanen. H. and Haggman. H. 1995. Seasonal changes in thetransient expression of a 3.5s CaMV-GUS gene construct introduced into Scats pine buds.
Tree Phys. 15: 65-70.Becwar, M.R.. Nagmani. R. and Wann. S.R. 1990. Initiation of embryogenic cultures and
somatic embryo development in loblolly pine (Pburs tuerkr). Can. J. For. Res. 20: 8lO-517.
Bekknoui. F., Dada. R.S.S.. Pilon. M., Tautorus, T.E., Crosby. W.L. and Dunstan. D.I. 1990.The effects of promoter on transient expression in conifer cell lines. Theor. Appl. Gen. 79:353-359.
Bommineni. V.R., Dada. R.S.S. and Tsang. E.W.T. 1994. Expression of ,qtcs in somatic embryocultures of black spruce after microprojectile bombardment. J. Exp. Bot. 45: 49 1495.
Brown. CL. and Lawrence. R.H. 1968. Culture of pine callus on a detined medium. For. Sci.14: 62-64.
Campbell. M.A.. Kinlaw. C.S. and Neale. D.B. 199 2 . Expression of luciferase and beta-glucuronidase in Pinr~s rmlirrtrr suspension cells usin g electroporation and particlebombardment. Can. J. For. Rcs. 11: X14-201 8.
Charest. P.J.. Devantier. Y. and Lachnnce. D. 1996. Stable genetic transformation of Picetrmttricurtr (black spruce) via particle bomburdment. In Vitro Cellular Cyr DevelopmentalBiology (Plant) 31: Y I -YY.
Charcst. P.S.. Calem. N.. LaChancc. D.. Datln. R.S.S.. Duchesne. L.C. and Tsang. E.W.T.IY93. Microprojcctilc-DNA &livery in conifer spccios: Factors al’fucting asscssmcnt oftransient gent cxprcssion using p-glucuronidasc rcportcr gcnc. PI. Ccl1 Rsp. 12: 1X9-193.
Chiu. W.L.. Niwa. Y.. Zcny. W.. Hiram). T.. Kohaynshi. H. and Shcon. J. IYYh. EnginecrcdGFP as a vital reporter in plants. Curr. Biol. 6: 325-330.
Chrisrcnscn. A.H.. Sharrock. R.A. and Quail. P. lY(J2. Maize polyuhiquitin gcncs: Structure.thermal pcrturhation ofexprcssion and transcript spliciny. and promouzr activity followingtransfer 10 protoplasts by clcctroporalion. PI. Mol. Biol. 18: 675-6X9.
Clapham. 0.. Mandcrs. G.. Yihrah. H.S. and von Arnold. S. lY9S. Enhanccmcnt of short- andmedium-lcrm cxprcssion ot’ rrnnsgcnes in cmhryogcnic suspcnsittns ol’ P icccc o1Ge.v (I..,.)Kntsl. J. Exp. Hot. -to: 655~662.

57
Dada. R.S.S.. Hammerlindl. J.K.. Pelcher. L.E.. Crosby. W.L. and Selvaruj, G. 1991. Ahifunctional fusion between &glucuronidnse and neomycin phosphotransferae: ;L broadspectrum marker enzyme for plants. Gene IO I : 239-246.
Duchesne. L.C. and Charest. l?J lYYl. Transient expression of fi-glucuronidase gene inembryogenic callus of Picert mcwirrnu. PI. Ceil Rep. IO: 19 I-194.
Ellis. D.D.. McCabe. D.. Russell. D.. Martinell. B. and McGown. B.H. 1991. Expression ofinducible angiosperm promoters in a gymnosperm, Piceu ,&IU~U (white spruce). PI. Mol.Biol. 17: 19-37.
Ellis. D.D.. McCabe. D.E., McInnes. S.. Ramachandran. R.. Russel. D.R.. Wallace. K.M..MartinelI. B.J.. Roberts. D.R.. Raffa, K.F. and McCown. B.H. 1993. Stable transformationof Picecr ~~/u~Lu by particle acceleration. BioTTechnology I I: 84-89.
Haggman, H.M.. Aronen. T.S. and Nikkunen. T.O. lY97. Gene transfer by particle bombard-ment to Norway spruce and Scats pine pollen:Can. J. For. Res. 37: 918-935.
Hay. I.. LaChance, D.. Von Aderka... P. and Charest. P.J. 1994. Transient chimeric gene expres-sion in pollen of live conifer species following microparticle bombardment. Can. 1. For.Res. 74: 14 17-2413.
He&r. W. 1992. Optimization of Biolistic transform;ltion using the helium-driven PDS-IofM)/He system. L’S/EG Bulletin #l688. BIO-RAD. Hercules. CA. 8 pp.
Jefferson. R.A.. Kavnnnh. T.A. and Bevan. M.W. 1987. GUS fusions: /&glucuronida.se a.. asensitive and versatile gene fusion marker in higher plants. EMBO J. 6: 3901-3907.
Kaul. K. 19%. Somatic embryogenesis in eastern white pine If intrs srrdw L.). pp. 257-268.In: Jain. S.M.. Gupta. PK. and Newton. R.J. (Eds.1 Somatic Embryogenesis in WoodyPlants. 3. Gymnosperms. Kluwer Academic Publishers. Dordrecht.
Klimaszewska. K.. Devanticr. Y.. LaChance. D.. Lelu. 1M.A. and Charest. P.J. 1997. LurixIuricincc (tamarack): Somatic embryogenesis and genetic transformation. Can. J. For. Res.77: 538-550.
Leffel. S.M.. Mabon. S.A. and Stewart. Jr.. C.N. l9Y7. Applications of green fluorescentprotein in plants. BioTechniques 23: 911-Y 18.
Li. Y.. Tremblay. F..M. and Seguin. A. 19011. Transient transformation of pollen andcmbryogenic tissues of white spruce ~Picea &lrtrr (Moench.) Vossl resulting frommicroprojectile bombardment. PI. Cell Rep. 13: 661-665.
Loopstra. C.A.. Weissinger. A.K. and Sederoff. R.R. 1992. Transient gene expression indifferentiating pine wood using microprojcctile bombardment. Can. 1. For. Res. 23:YY3-YY6.
NIcC&c. D.E. and .Llartinell. B.J. I YY3.Transformation of elite cottc:n cultivars via particlehomhnrdmcnt of mcristcms. Bio/Tcchnolopy I I: 596-59X.
Mitra. A. and Higgins. D.W. 1004. The Ch/o,r//tr virus adcninc methyltrnnsferase gentpromoter is ;I strong promoter in plants. PI. Mol. Biol. 36: X5-Y3.
Mitra. A.. Higgins. D.W. and Rohc. N.J. IYY4. A C’lhrr//o virus gcnc promoter functionsus a strong promoter both in pkunts and hactcrin. Biochtm. Biophys. Rcs. Comm. 204:I X7- I YJ.
~Ncw~on. R.J.. H.S. Yihmh. N. Don g. D.H. Clapham and S. von Arnold. lYY2. Expression ofan abscisic acid rcsponsivc promoter in /‘icy c~bic#.r (L.) Karst. following hornhardmcntI’rom an olcctric discharge partiolc accclcrator. PI. Coil Rep. I I : I XX-101
Rohcrtscln. D.. Wcishinfcr. A.K.. Acklcy. R.. Glovcr. S. and Scdcrr~ff. R.R. IYYZ. Gsnctic~rans;li~rmution t)l’ Norway spruce 1 IJicrcr crhic,,s t L.) Karst 1 using somatic cmhryo explantshy microprojcctilc homhardmcnt. PI. Mol. Biol. IY: OLYGJ).1S.
Sheen J. lOY.3. Protein phosphatasc activity is rcquircd lirr light-inducihlc ~cnc expression inmaize. EMBO J. 11: 3497-3505.

58
Shin. D.1.; Podila, G.K. and Karnosky, D.F. 1994. Transgenic larch expressing genes forherbicide and insect resistance. Can. J. For. Res. 24: 2059-2067.
Stomp, A-M., Weissinger. A. and Sederoff, R.R. 1991. Transient expression frommicroprojecti le-mediated DNA transfer in Pinus rue&. Pl. Cel l Rep. 10: 187-190.
Tian, L.. Seguin. A. and Charest. P.J. 1997. Expression of the green fluorescent protein genein conifer tissues. Pl. Cell Rep. 16: 267-271.
Walter, C.. Smith, D.R., Connett. M.B.. Grace. L. and White, D.W.R. 1994. A biolisticapproach for the transfer and expression of a gu.sA reporter gene in embryogenic cultures
I of Pinus radiata. PI. Cell Rep. 14: 69-74.
Copyright © 2022 FDOKUMEN




















