
Evaluation of certain contaminants in food - WHO | World ...
Upload
usherbrookeCategory
view
1download
0
404
Introduction
Laccases (Lac, E.C. 1.10.3.2) have been the subject of extensive research for their potential applications in vari-ous bioremediation fields during the last decades. Many researchers have investigated the use of these enzymes during the treatment of effluents from the pulp and paper industry, the textile industry, and municipal sewage (cf. reviews by Couto & Herrera (2006); Kunamneni et al. (2008); Madhavi & Lele (2009); Majeau et al. (2010); and Strong & Claus (2011)). This increasing interest in lac-cases in wastewater treatment is triggered by the pressing need to use “greener compounds” rather than environ-mentally harmful compounds in treatment processes
and by the necessity to effectively eliminate a wide vari-ety of emerging contaminants that are unsatisfactorily removed by existing effluent treatment technologies.
Along with lignin peroxidase (LiP, E.C. 1.11.1.14), manganese peroxidase (MnP, E.C. 1.11.1.13), and ver-satile peroxidase (VP, E.C. 1.11.1.16), laccases constitute one of the three major enzymes secreted by the white rot fungi (WRF) for the degradation of lignin. Although these enzymes share many properties regarding their oxida-tive capability, the copper-containing oxidases, laccases, possess some specific characteristics for their applica-tion in bioremediation. Laccases catalyze reactions in the presence of O
2 and the substrate (Majeau et al.,
RevIew ARtIcle
Laccase immobilization and insolubilization: from fundamentals to applications for the elimination of emerging contaminants in wastewater treatment
Sidy Ba1, Alexandre Arsenault2, Thanina Hassani2, J. Peter Jones1, and Hubert Cabana2,3
1Department of Chemical Engineering, Université de Sherbrooke, Sherbrooke, Québec, Canada, 2Environmental Engineering Laboratory, Department of Civil Engineering, Université de Sherbrooke, Sherbrooke, Québec, Canada, and 3Étienne-Lebel Clinical Research Centre, Centre Hospitalier Universitaire de Sherbrooke, Sherbrooke, Québec, Canada
AbstractOver the last few decades many attempts have been made to use biocatalysts for the biotransformation of emerging contaminants in environmental matrices. Laccase, a multicopper oxidoreductase enzyme, has shown great potential in oxidizing a large number of phenolic and non-phenolic emerging contaminants. However, laccases and more broadly enzymes in their free form are biocatalysts whose applications in solution have many drawbacks rendering them currently unsuitable for large scale use. To circumvent these limitations, the enzyme can be immobilized onto carriers or entrapped within capsules; these two immobilization techniques have the disadvantage of generating a large mass of non-catalytic product. Insolubilization of the free enzymes as cross-linked enzymes (CLEAs) is found to yield a greater volume ratio of biocatalyst while improving the characteristics of the biocatalyst. Ultimately, novel techniques of enzymes insolubilization and stabilization are feasible with the combination of cross-linked enzyme aggregates (combi-CLEAs) and enzyme polymer engineered structures (EPESs) for the elimination of emerging micropollutants in wastewater. In this review, fundamental features of laccases are provided in order to elucidate their catalytic mechanism, followed by different chemical aspects of the immobilization and insolubilization techniques applicable to laccases. Finally, kinetic and reactor design effects for enzymes in relation with the potential applications of laccases as combi-CLEAs and EPESs for the biotransformation of micropollutants in wastewater treatment are discussed.Keywords: Laccases, combination of cross-linked enzyme aggregates, enzyme polymer engineered structures, bioreactor, emerging contaminants, wastewater treatment
Address for Correspondence: Hubert Cabana, PhD, Environmental Engineering Laboratory, Department of Civil Engineering, Université de Sherbrooke, 2500 boul. de l’Université, Sherbrooke, Québec, J1K 2R1, Canada. Tel: 819-821-8000 ext. 65457. Fax: 819 821-7974. E-mail: [email protected]
(Received 19 March 2012; revised 01 August 2012; accepted 01 August 2012)
Critical Reviews in Biotechnology, 2013; 33(4): 404–418© 2013 Informa Healthcare USA, Inc.ISSN 0738-8551 print/ISSN 1549-7801 onlineDOI: 10.3109/07388551.2012.725390
Critical Reviews in Biotechnology
33
4
404
418
19March2012
01August2012
01August2012
0738-8551
1549-7801
© 2013 Informa Healthcare USA, Inc.
10.3109/07388551.2012.725390
2013
Laccase immobilization and insolubilization
S. Ba et al.
Cri
tical
Rev
iew
s in
Bio
tech
nolo
gy D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsite
De
Sher
broo
ke o
n 11
/13/
14Fo
r pe
rson
al u
se o
nly.

Laccase immobilization and insolubilization 405
© 2013 Informa Healthcare USA, Inc.
2010; Claus et al., 2002; Leonowicz et al., 1988) instead of hydrogen peroxide for LiP, MnP, and VP. Thereby, they are able to carry out a one-electron oxidation of phenolic compounds with a concomitant reduction of oxygen to water without the need for hydrogen peroxide. Moreover, they can oxidize non-phenolic compounds in the pres-ence of electron transfer mediator molecules that enable them to overcome their lower redox potential (450–800 mV) towards these compounds (Bourbonnais et al., 1997, 1998; Li et al., 1999). As a result, their substrate spectra have become even broader making them very appealing for bioremediation applications.
Laccases have proven to have considerable potential for the removal of several pollutants of environmental concern in water bodies and soils. These pollutants include chlorophenols (Roy-Arcand & Archibald, 1991; Ricotta et al., 1996; Gaitan et al., 2011; Ahn et al., 2002; Bollag et al., 1988); dyes (Claus et al., 2002; Abadulla et al., 2000); and some emerging compounds estab-lished or suspected to be endocrine disrupting chemi-cals such as bisphenol A, nonylphenol, and triclosan (Fukuda et al., 2001; Cabana et al., 2007a; Zhang et al., 2009; Nicolucci et al., 2011). However, it is noteworthy that large scale use of free laccase requires an enor-mous quantity of the enzyme produced at relatively low cost which is not an easy task due to the cost of the culture medium (Osma et al., 2011). Furthermore, many factors are known to hinder the full potential catalytic ability of the free enzyme in wastewater (as generally observed for different enzymes in various applications). Inhibition of laccase catalytic properties has been observed through conformational change of the enzyme, modification induced to the amino groups along with the chelation of the copper atoms (Johannes & Majcherczyk, 2000). Some authors demonstrated that copper reducing or chelating agents azide, thiogly-colic acid, diethyl-dithiocarbamate inhibit the activ-ity of laccase (Bollag & Leonowicz, 1984). High ionic strength of the medium due to the presence of certain ions of heavy metals (e.g. Hg2+, Sn2+, Zn2+, Fe2+, Fe3+); halides (e.g. F−, Cl−, Br−); acetates compounds; some organic solvents (e.g. acetone, acetonitrile, dimethyl sulfoxide) and anionic surfactants (e.g. sodium dodec-ylsulfate) are also reported to inhibit laccase activity to some extent (Madhavi & Lele, 2009; Yaropolov et al., 1994; Couto & Toca, 2006). Lack of stability is another major limiting-factor for large scale use of free laccase. Several studies have shown high loss of activity of vari-ous free laccases is a shortcoming to their operational stability (Leonowicz et al., 1988; Zhang et al., 2009; Nicolucci et al., 2011; Cabana et al., 2007b; D’Annibale et al., 1999). Finally, difficult recovery coupled with limited reusability has also been identified as a major limiting-factor for large scale application of the free enzyme (Cabana et al., 2007a).
Lately there have been substantial efforts towards eliminating or mitigating these drawbacks to enable
large scale application of enzymes including laccases. Immobilization is one of the techniques that has been explored in order to reach this objective [cf. review by Duran et al. (2002) and Mateo et al. (2007)]. Enzyme immobilization has been categorized into three methods: support or carrier-binding, encapsulation or entrapment, and cross-linking (Duran et al., 2002; Sheldon, 2007; Roessl et al., 2010). Carrier-binding methods for enzymes are performed through physico-chemical mechanisms. Adsorption occurs via electrostatic interactions and Van der Walls forces between the enzymes and the carrier sur-face. Adsorption via hydrogen bonding is also observed in physical immobilization. Immobilization occurs via ionic or covalent binding. In the former, the enzyme and its support bind depend on the medium pH and its ionic composition whereas in the latter the binding takes place between some functional groups of the carrier and the enzyme. The second method of enzyme immobilization, known as encapsulation (entrapment), is not much differ-ent from the carrier-binding method. In encapsulation, the enzyme is trapped inside the pores of the support in order to preserve the enzyme’s three dimensional structure. The most widely used technique in this category is immobi-lization in silica sol–gel matrices (Hanefeld et al., 2009). Both the carrier-binding and encapsulation methods have the disadvantage of generating large non-catalytic mass (support) for low quantity of catalyst and thus diminishing the efficiency of the obtained biocatalyst. Crosslinking has been developed as an alternative to the carrier-binding and encapsulation methods.
The technique of crosslinking enzymes gained interest in the sixties and seventies (Quiocho & Richards, 1964; Epton et al., 1971) as a route to avoid activity dilution in a carrier. Basically, this immobilization technique is a vari-ant of covalent binding in which functional groups (e.g. amine groups, carboxylic groups) of different enzyme molecules are linked together using a bi- or multi-func-tional reagent. Crosslinking reagents are of prime impor-tance in the immobilization technique as their chemistry enables the inter- and intra-molecular covalent bond-ing and does not contribute to the deactivation of the enzyme (Roy et al., 2005). Carrierless immobilization is performed via several methods. One method consists in direct crosslinking the enzymes in solution known as cross-linked enzymes (CLEs). In turn, support-free bio-catalysts in solution can be spray-dried prior to immo-bilization (cross linked spray dry, CLSD) leading to a low activity retention and low mechanical stability similar to the CLE. A more robust and highly active insolubilized enzyme is found to be feasible by cross-linked enzyme crystals (CLECs). For instance, CLEC of laccase from Trametes versicolor has been proven to be very active in oxidizing several phenolic compounds (guaiacol, cate-chol, pyrogallol, and catechin) and a non-phenolic com-pound (ABTS) (Roy et al., 2005; Bourbonnais & Paice, 1990). However, major drawbacks on CLECs formation reside in the fact that the crystallization process which is
Cri
tical
Rev
iew
s in
Bio
tech
nolo
gy D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsite
De
Sher
broo
ke o
n 11
/13/
14Fo
r pe
rson
al u
se o
nly.

406 S. Ba et al.
Critical Reviews in Biotechnology
very laborious and requires highly pure enzyme. Cross-linked enzymes aggregates (CLEAs), on the other hand, are made with raw enzymes enabling the formed biocat-alyst to be insoluble without major modification of their molecular structure eliminating loss of activity. CLEAs have been the subject of some researches (Schoevaart et al., 2004; Sangeetha & Abraham, 2008; Cabana et al., 2009a) and will be discussed further on in this review.
Novel approaches of enzyme insolubilization and stabilization consisting of a combination of different enzymes as cross-linked enzyme aggregates (combi-CLEAs) (Dalal et al., 2007; Bruggink et al., 2003; Mateo et al., 2004; Schoevaart & Kieboom, 2002a) and single enzyme nanoparticles (SENs) (Kim & Grate, 2003, 2005; Hegedus & Nagy, 2009; Yadav et al., 2011a) have recently emerged as promising biocatalyst synthesis and are gain-ing interest in bioremediation.
In this review, the fundamental aspects of laccases relevant to their catalytic reaction for the removal of some emerging contaminants in wastewater are pre-sented. The chemistry of crosslinkers and their mecha-nism of reaction with enzymes including laccases in the technique of insolubilization are reviewed. Finally, the potential use of laccases in the form of novel biocatalysts (CLEAs/combi-CLEAs and Enzyme Polymer Engineered Structures (EPESs)) to treat micropollutants of emerging concern in wastewater are discussed.
laccases
CompositionLaccase (benzenediol: oxygen oxidoreductase, EC 1.10.3.2) is a multicopper oxidase (MCO) glycoprotein. First reported to be present in the juice of the milky secre-tion (urushi) of the Japanese tree Rhus vernicifera (Yoshida, 1883), the enzyme is now well-known to be ubiquitous in nature and found in plants, insects, bacteria and fungi. Fungal laccases (the most widely encountered in the lit-erature and the focus of this review) catalyze the oxidation of a broad number of environmental pollutants (Xu, 1996; Aust & Benson, 1993). In order to decipher the oxidation mechanism of laccases, it is critical to understand its three dimensional structure and amino acid (AA) residue sequence. Several studies on laccase from different spe-cies of fungi have recently provided a significant insight of its crystalline structure and AA residues sequence (Ducros et al., 1998; Ferraroni et al., 2007; Hakulinen et al., 2002; Piontek et al., 2002; Garavaglia et al., 2004; Kallio et al., 2011; Bertrand et al., 2002) that is summarized hereafter. Basically, laccases are found to be made of a sequence of polypeptide containing about 500 AA residues and linked to saccharide molecules. The enzyme is generally a monomer containing at least one copper atom with, frequently, three additional coppers bound to specific sites of its structure. The AA residues are arranged in three cupredoxin-like β-barrel domains similar to that of ascorbate oxidase (AO) described by Messerschmidt and Huber (1990). On average, the three domains encase each
140–200 residues with overall structural dimensions of about 70 Å × 55 Å × 50 Å. From UV/visible and electron paramagnetic resonance (EPR) spectroscopy, the four copper atoms have been conventionally classified into three types: types 1 and 2 which comprise one copper atom each and type 3 with two copper atoms. The type 1 copper (T1) occupies a mononuclear copper site embed-ded 6–7 Å below the surface of the enzyme in domain 3 where it is bound to two histidines and a cysteine residues (and often to a methionine as well). T1 is found to have an absorption maximum at around 600 nm, which confers the intense blue color to the enzyme, and is EPR detect-able. T2 is reported as normal copper exhibiting only weak absorption in the visible region and is EPR-active. The binuclear T3 is characterized by an absorption peak at about 300 nm with no EPR signal due to an antiferromag-netic coupling mediated by a hydroxyl or oxygen bridging ligand between the two copper atoms. T2 and T3 form a trinuclear copper cluster which center is sited at the inter-face of domains 1 and 3 at 12–13 Å beneath the surface of the enzyme where they are coordinated to eight histidine residues (two for T2 and three for each of the double cop-per of T3). The coordinate system also includes a histidine-cysteine-histidine tripeptide connection between T1 and the trinuclear cluster T2/T3, two interdomain disulfide bridging (i.e. cysteine–cysteine between domains 1 and 2 and between domains 1 and 3) and an extensive loop bridging domains 2 and 3. These complex labyrinthine interconnections contribute to rendering the overall struc-ture of the enzyme stable. Understanding the coordination complex of laccases established the basis for elucidating their catalysis mechanism that has mostly derived from its MCO family member, AO, comprehensively described (Messerschmidt, 1997; Solomon et al., 1996). Accordingly, the oxidation process is initiated at the T1 site where suc-cessive individual electrons from the reducing substrate are extracted and transferred to the T2/T3 cluster via the His-Cys-His ligand described earlier. At the latter site, accumulated electrons enable the reduction of oxygen to water.
Most fungal laccases are extracellular proteins and have a molecular weight of approximately 60–70 kDa (Baldrian, 2006; Giardina et al., 2010). However, weights not in this range are commonly found as illustrated for Phanerochaete chrysosporium, 47 kDa (Srinivasan et al., 1995) and Podospora anserine, 390 kDa (Minuth et al., 1978) laccases. For these variable molecular masses, the glycosylation state accounts for 10–25% carbohydrate of the protein total molecule by weight. The glycosylation extent also varies significantly from species to species and within the same species with different strains. This is demonstrated for Myceliophthora thermophila, 40–60% (Berka et al., 1997) and Botrytis cinerea, 86–91% (Zouari et al., 1987). Conversely, lower carbohydrate contents were found for Pleurotus eryngii, 1–7% (Muñoz et al., 1997). The glycans commonly found to be bound, usually through an N-linkage, to the polypeptide of the enzyme include mannose, glucose, N-acetyl-d-glucosamine,
Cri
tical
Rev
iew
s in
Bio
tech
nolo
gy D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsite
De
Sher
broo
ke o
n 11
/13/
14Fo
r pe
rson
al u
se o
nly.

Laccase immobilization and insolubilization 407
© 2013 Informa Healthcare USA, Inc.
hexosamine, galactose, fructose, arabinose, and xylose (Minuth et al., 1978; Gigi et al., 1981; Marbach et al., 1984). The role of glycosylation for the biocatalyst is very important. In binding to its molecule, the carbohydrate moiety chains, obviously, contribute to defining the structure of the enzyme with a specific folding which is proven to provide its protective functions against proteo-lytic and elevated temperature degradations (Yoshitake et al., 1993).
Fungal laccases have an acidic isoelectric point (pI) around pH = 4.0 (varying from 2.6 to 6.9) (Strong & Claus, 2011). These enzymes were found to exhibit their higher activity at pH optima in the acidic range (depending on test conditions). This may be attributed to the fact that the microorganisms producing enzymes grow better under acidic medium conditions (Madhavi & Lele, 2009) and to the occurrence at high pH of strong bonding of T2 Cu-OH− that hampers the reduction of oxygen ion to water via the reaction O2− + 2H+ → H
2O (Andréasson &
Reinhammar, 1979). The pH optima are very much sub-strate-dependent. While the oxidation of ABTS occurs at an optimal pH of 4 or lower, the optimal pH is between 4 and 7 for 2,6-dimetoxyphenol (DMP), guaiacol, syrin-galdazine with a bell-shaped curve expressing activ-ity versus pH (Strong & Claus, 2011). However, it is not uncommon to find a non-bell-shaped curve for activity-pH as the optimum pH may show up in a narrow peak or in pseudo-linear forms.
Non-specificityLaccases are non-specific enzymes in regards to their substrates. They are able to catalyze the oxidation of a large number of organic compounds including phenols. As such, they are recognized as versatile biocatalysts oxidizing natural and industrial compounds (Reiss et al., 2011) of environmental interest in effluents treatment. Additionally, laccases are found to be able to oxidize non-phenol compounds with higher redox potential. For instance, in the presence of redox co-catalysts or media-tors, laccases break down the carbon-hydrogen bonds to enable the oxidation process of aliphatic compounds (ten Have & Teunissen, 2001; Call & Mucke, 1997). These mediators include synthetic compounds such as ABTS, violuric acid and 1-hydrobenzotriazole (1-HBT), as well as natural products such as 4-hydroxybenzoic acid and 4-hydroxybenzyl alcohol (Li et al., 1999; Johannes & Majcherczyk, 2000). Unlike most (if not all known) biocatalysts, laccases use four single-electron oxidation steps for their catalysis with readily available molecular oxygen as the unique stoichiometric oxidant of the reac-tions where mediators are not needed. Subsequently, water is found to be the sole by-product of the reactions which eliminates any concern for other end-products when using the biocatalyst. Also, as these oxidations occur in water at ambient temperature (20–40°C) laccases have become good candidates for green and an environ-mentally friendly oxidation method for the elimination of xenobiotic molecules (Wells et al., 2006). Furthermore, it
is known that more and more phenolic compounds, hav-ing harmful effects, are widespread in the environment and that the existing conventional technologies have limited capabilities to treat some of these compounds, therefore the use of laccases is a promising alternative.
Applications of free laccases in wastewater treatment
Laccases are useful biocatalysts for a wide range of biotech-nological (including bioremediation) applications due to their highly nonspecific oxidation ability and their use of readily available molecular oxygen as an electron accep-tor (Giardina et al., 2010). Laccases have been applied for the decolorization and detoxification of effluents from textile, dyestuff, and pulp and paper industries (Harms et al., 2011). The application of laccase in municipal wastewater has also been largely explored. For instance, in a batch study, natural and synthetic estrogens (estrone, 17β-estradiol, estriol, and 17α-ethinylestradiol) were completely removed from synthetic and municipal waste-water when treated with laccases from T. versicolor (Auriol et al., 2007, 2008). In another experiment, laccase from T. versicolor, applied in primary effluents, was able to remove 95–100% of oxybenzone (Garcia et al., 2011). Indeed, amongst the many micropollutants that are not removed from the liquid phase by wastewater treatment plants, some portion of these contaminants are transferred into the sludge and may end up in soil matrices (e.g. through agricultural application). Thus, it is important that lac-case biocatalysts become effective in treating the targeted micopollutants in soil bioremediation processes as well. Several studies have shown the potential of laccases to do so. For instance, laccase from Trametes sp. was used to remove more than 80% of phenolic endocrine-disrupting chemicals (bisphenol A, nonylphenol, octylphenol, and ethynylestradiol) from soils (Tanaka et al., 2001). There are other reported applications of laccases for the detoxifica-tion of soils (Ahn et al., 2002; Bollag, 1992).
However, as the free enzyme is not currently suitable for large scale use in bioremediation, several immobilization techniques have been attempted. For instance, Nicolucci and coworkers, using laccase from T. versicolor immo-bilized on polyacrylonitrile beads in a FBR, were able to completely remove bisphenol A, bisphenol B, bisphenol F and to convert 96% of tetrachlorobisphenol A after 90 min of enzyme treatment of contaminated waters (Nicolucci et al., 2011). In a batch reactor, Zhang and coworkers were able to remove up to 88.8% of 2,4-dichlorophenol in aqueous solution using chitosan-immobilized laccase from Coriolus versicolor (Zhang et al., 2009). In batch con-ditions, total phenols and ortho-diphenols were elimi-nated by 67 and 72%, respectively, from olive-mill effluent after 24 h of application of chitosan-immobilized laccase from Lentinula edodes (D’Annibale et al., 2000). Although application of immobilized laccase for the elimination of pollutants in aqueous solution appears limited in the literature, these previous examples are illustrative of the
Cri
tical
Rev
iew
s in
Bio
tech
nolo
gy D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsite
De
Sher
broo
ke o
n 11
/13/
14Fo
r pe
rson
al u
se o
nly.

408 S. Ba et al.
Critical Reviews in Biotechnology
potential of immobilized/insolubilized laccase in waste-water bioremediation.
The advantage for using of laccases as biocatalysts in the treatment of wastewater lies also in their reac-tion end-products. Indeed, laccases oxidize phenolic compounds to form their corresponding free and very reactive radicals (Bollag, 1992). These phenoxyl radicals then polymerize to form either varieties of oligomers or further degrade the formed oligomers to different fragments. Also, the various metabolites generated have been found to completely or significantly lose their biological activity and toxicity. It is argued that the elimination of the biological activity of the parent compound could be linked to the physical removal by precipitation of the oligomers produced via the laccase catalysis, which implies that these oligomers are insol-uble (Cabana et al., 2007a). In soil matrix, Bollag (1992) claimed that the oxidative coupling of phenolic chemi-cals to the humic substance mediated by laccase leads to a decrease of the amount of material available to interact with the biota, thus reducing the toxicity of the parent phenolic compound. Several studies illustrate the nature of end-products and their resulting activity and toxicity. For instance, it was demonstrated that an application of Trametes villosa laccase on bisphenol A (BPA) catalyzed the oxidative condensation of BPA into its dimer, trimer, tetramer, pentamer and hexamer with C–C and/or C–O bonds between phenol moieties. Subsequently, these oligomers cleaved to fragments of oligomers each with phenol molecules as shown by the release of 4-isopropenylphenol (Fukuda et al., 2004). These authors also found no estrogenic activity for the BPA reaction products and effective BPA’s toxicity removal with sufficient amount of the laccase. Likewise, an elimination of BPA, triclosan (TCS), and nonylphe-nol (NP), three phenolic compounds, with Coriolopsis polyzona laccase produced their respective oligomers associated with fragmentation of some of the formed oligomers; after 8 h of enzymatic treatment, the metab-olites for both BPA and NP had no estrogenic activity (Cabana et al., 2007a). In an experiment for the elimi-nation of BPA and TCS using laccase from Ganoderma stipitatum and Lentinus swartzii fungi, (Arboleda et al., 2012) have shown more than 90% removal of both contaminants with generation of various oligomers as reaction products. Subsequent analysis showed up to a 90% reduction of the estrogenic activity of BPA and an important reduction of the toxicity of both BPA and TCS solutions after enzymatic treatments. This toxicity result is consistent with previous work conducted by Kim and Nicell (2006) in which laccase-catalyzed oxidation of BPA resulted in a significant decrease in toxicity of the aqueous solution.
Immobilization and insolubilization methods of enzymes
In order to optimize their operational performance for industrial and large scale applications, enzymes
have been immobilized/insolubilized through various methods described in several reviews as mentioned in the introduction. Thus, in this section, we focus on the insolubilization technique with regard to the AA residues and crosslinking agents.
Insolubilization is performed without solid support by simply linking enzyme molecules to each other to form large complexes inherently immobilized because indi-vidual enzyme molecules are no longer free to diffuse in solution (Twyman, 2005). As a result, a greater activity ratio to non-catalytic mass than carrier-bound enzymes is obtained assuming that denaturation does not take place during the process. Several methods of enzyme crosslinking have been developed since the second half of the 20th Century for various purposes (Quiocho & Richards, 1964; Epton et al., 1971; Cao, 2005). The tech-niques of crosslinking are a variant of covalent binding involving functional groups (e.g. NH
2) of the enzyme
linked together by the aid of a bi- or multi-functional reagent. Detailed description of these techniques has been comprehensively reviewed (Sheldon, 2007; Roessl et al., 2010; Cao et al., 2003). For these techniques, the chemistry of the cross-linker and the surface properties of enzyme’s amino groups are important features in the process of cross-linking and will be extensively discussed. Novel approaches of enzyme immobilization include the combination of cross-linked enzymes aggregate (combi-CLEAs) and the enzyme polymer engineered structures (EPESs); this latter being an attractive approach to increase CLEAs stability.
Enzymes and amino acid residues surface propertiesAA residues play a major role in enzyme immobiliza-tion/insolubilization. Enzymes (and proteins in gen-eral) are macromolecules formed by polymerization of AA residues. The sequence of these AA residues (also known as the primary structure) forms the backbone of the protein and determines how the protein will fold into a three-dimensional structure. Enzymes also have side chains bound to the AA skeleton. These side chains are also very important features of enzymes as they determine most of their surface properties. The side chains can be classified into four subgroups: the non-polar hydrophobic (generally within the center of the enzyme) unfavorable for covalent bonding, polar hydro-philic (generally on the surface of the enzyme) with hydrogen bonding capability, net positively-charged (strongly hydrophilic acting as base in solution), net negatively-charged (strongly hydrophilic acting as acid in solution). These net charges generally result from solution pH relative to the dissociation constant (pKa) of the AA residue; at pH values above the pKa, AA residue become anionic or neutral, whereas below the pKa AA residue become cationic or neutral. Indeed, the activity level of the side chain is determined by their degree of protonation; a variation in the following order has been reported: –S− > –SH > –O− > –NH
2 > –COO− > –OH > –NH
3+
and their charges can be derived from the solution pH
Cri
tical
Rev
iew
s in
Bio
tech
nolo
gy D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsite
De
Sher
broo
ke o
n 11
/13/
14Fo
r pe
rson
al u
se o
nly.

Laccase immobilization and insolubilization 409
© 2013 Informa Healthcare USA, Inc.
and the pKa of the ionized groups (Cabral & Kennedy, 1991). Also, the AA residue net charges are important in enzyme immobilization/insolubilization by means of adsorption via electrostatic/ionic interactions. In fact, the main factors to consider when immobilizing (or insolubilizing) enzymes are: chemical and physi-cal environment (i.e. solution), charge on enzyme/PI, and polarity of the enzyme (hydrophobic/hydrophilic regions) (Hanefeld et al., 2009).
With regard to fungal laccases, the three most reported AA residues are histidine (His), cysteine (Cys), and methi-onine (Met) (cf. section 2.1). Met is non-polar and contains a sulfur atom not involved in crosslinking contrarily to His and Cys residues both found to be involved in coupling with activated functional groups (Twyman, 2005; Habeeb & Hiramoto, 1968). This has basic side chains positively charged at or near neutral pH or lower. The side chain amino group is attached to an imidazole group which has a pKa of 6.0. However, as pKa values for AA residues depend on the environment and can change significantly within the confines of a protein, this can be protonated or unpro-tonated (Campbell & Farrell, 2006). Cys has a polar neutral side chain at neutral pH which consists of a thiol group (–SH) that can react with other thiol groups to form disulfide (–S–S–) bridges in oxidation reaction. The thiol group can also be deprotonated (Campbell & Farrell, 2006). Along with cysteine and histidine, lysine and tyrosine residues (also present in laccases) are found to be most commonly involved in coupling reaction with activated functional group on support material (Twyman, 2005).
When immobilizing enzymes without support (i.e. insolubilizing), the use of a crosslinking agent, that reacts with the residual functional groups on the surface of the AA residues, is essential. Various compounds, described below, have been used in crosslinking enzymes.
Chemistry of cross-linkersAmongst the numerous crosslinkers, some of them such as dialdehydes have been extensively used over a long period of time.
DialdehydeMany dialdehydes have been used to efficiently cross-link proteins or other biopolymers. The more often, glutaraldehyde is the dialdehyde of choice but glyoxal, malondialdehyde (Kimura et al., 2003) scleraldehyde (Gladstone) and modified polysaccharides have also been used. Excluding the particular case of glutaralde-hyde, most of these dialdehydes will crosslink enzymes in a reaction similar to formaldehyde reacting with an amine (see Figure 1).
Most dialdehydes will produce a Schiff base when reacting with one or more proteins. This Schiff base can then be reduced to yield a stronger link, usually with cyanoborohydride salts (Hermanson, 1996). However, glutaraldehyde reacts differently than other dialdehydes since it undergoes polymerization when in solution (Migneault et al., 2004).
GlutaraldehydeGlutaraldehyde is a synthetic pentacarbon homobifunc-tional aldehyde soluble in water, alcohol, and organic solvents widely used in enzymes technology. It is widely recognized that glutaraldehyde reacts primarily with the amino groups of proteins to form crosslinks analogously to the mechanism of reaction of formaldehyde (Hadorn & Nitschmann, 1944; Fraenkel-Conrat & Olcott, 1948; Fraenkel-Conrat & Mecham, 1949). There also has been a report of glutaraldehyde reaction with various functional groups of proteins including amine, thiol, phenol, and imidazole into crosslinked groups (Habeeb & Hiramoto, 1968); which has been attributed to the nucleophilic character of the side chains (Migneault et al., 2004). Glutaraldehyde has been shown to be very reactive towards laccase-based AA residues as the cases with sulf-hydryl group of cysteine and imidazole group of histidine (Habeeb & Hiramoto, 1968). The ability of glutaraldehyde to react with the primary and side chain structures of pro-teins makes it an efficient reagent candidate in enzyme crosslinking. Moreover, glutaraldehyde is commercially available at low cost and has several reactive forms based on solutions conditions such as pH, concentration, tem-perature, etc. (Migneault et al., 2004) to generate very stable linkages with proteins (Cabral & Kennedy, 1991). Many enzymes were successfully crosslinked by glutar-aldehyde like laccase (Cabana et al., 2007b), tyrosinase (Aytar & Bakir, 2008), lipase (Fernández-Lorente et al. 2006) and many more. These enzymes were bound on solid supports, entrapped in a matrix and formed cross-linked enzyme aggregates. The performances of these treatments varied from one enzyme to another and from a type of immobilization to another. A good example of the efficiency of glutaraldehyde is the CLEAs of laccase produced by Cabana et al. (2007b). During the produc-tion of these CLEAs, they obtained up to 60% of activity recovery and the CLEAs obtained were more resistant to chemical denaturants and to thermal degradation (half-life of 7.7 h instead of 2 h for free laccase in pH3 citrate buffer at 40°C). Some enzymes did not maintain their active form during the immobilization process like the nitrilase which lost most of its activity possibly due to the reaction between glutaraldehyde and an AA residue present in its catalytic site (Sheldon, 2007).
However, glutaraldehyde may cause large losses of enzymatic activity (Schoevaart et al., 2004), a fact attrib-uted to physical attack, chemical modification and/or denaturation of subunits essential for catalytic activity (Roessl et al., 2010; Mateo et al., 2004). Because glutaral-dehyde is highly reactive and small in size, it can diffuse into the internal structure of enzymes and react with the active site’s AA residue resulting to a severe loss of their
Figure 1. Reaction of formaldehyde with amine group.
Cri
tical
Rev
iew
s in
Bio
tech
nolo
gy D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsite
De
Sher
broo
ke o
n 11
/13/
14Fo
r pe
rson
al u
se o
nly.

410 S. Ba et al.
Critical Reviews in Biotechnology
catalytic activity (Mateo et al., 2004). Additionally, glu-taraldehyde may cause adverse health effects to humans (Dimich-Ward et al., 2004; Takigawa & Endo, 2006) such as nasal, throat, and lung irritation, asthma and conjunc-tivitis. This crosslinker is also toxic where it can reduce reproduction and impede the growth of aquatic organ-isms and species (Emmanuel et al., 2005; Leung, 2001; Sano et al., 2005). Due to these drawbacks various cross-linkers have been used as alternative to glutaraldehyde including glyoxal, dextran polyaldehyde, etc.
GlyoxalGlyoxal is a dialdehyde often used in the process of enzyme crosslinking or immobilization as a surrogate to glutaraldehyde due to its lower toxicity. This three-carbon long crosslinker has been used by Cabana et al. (2009b) to immobilize laccase on Celite® beads leading to improved thermal stability of the enzyme. About 30% of the initial activity was retained after 24 h at 40°C in a pH 3 citrate solution instead of less than 10% for the free laccase. Due to its size, glyoxal generates smaller steric hindrance than glutaraldehyde since the laccase activity recovery after immobilization with the use of glyoxal is higher than with glutaraldehyde. It also has been used by Kiso et al. (2010) to crosslink hydrogels of collagen and chitosan. These hydrogels appeared to be biocompat-ible while those crosslinked by glutaraldehyde showed a high cytotoxicity. This advantage permits the cross-linking simultaneously to the incorporation of the cells in the chitosan and collagen scaffold. However, glyoxal is known to have some adverse effects on human cells especially on the proteins and DNA (Nghiem & Schäfer, 2002).
Dextran polyaldehydeDextrans are commercially available highly hydrophilic carbohydrate polymers. These molecules have been modified to form dextran polyahldehyde as a crosslinker. The synthesis process consists of oxidizing the polysac-charide by dissolution in water and addition of sodium metaperiodate (Mateo et al., 2004) or similar compound to cleave the C–C bonds with subsequent formation of polyaldehydic macromolecules reactive to AA residues (Figure 2). A final reduction of these macromolecules with sodium borohydride results to an irreversibly linked structure.
One of the main advantages of dextran polyaldehyde is its intrinsic macromolecular structure which prevents any possible diffusion to the enzyme active site, thus yielding
greater catalytic activity than with glutaraldehyde unless a steric hindrance takes place. For instance, enzymes (oxynitrilase from Prunus amygdalus, hydroxynitrile lyase, alcohol dehydrogenase from Lactobacillus brevis, Penicillin G acylase from Escherichia coli, and two differ-ent nitrilases) have been synthesized as CLEAs using glu-taraldehyde and dextran polyaldehyde as crosslinkers. The results showed equal (case of oxynitrilase only) to much higher activity recovery with the latter reagent (Mateo et al., 2004). Besides the polysaccharide dextran, the polyaldehydes can be made with several other mono-saccharides such as galactose (Schoevaart & Kieboom, 2001, 2002b), glucose, lactose, and sucrose (Schoevaart et al., 2005).
ChitosanChitosan is a derivative biopolymer of chitin by N-deacetylation (Figure 3). Chitin is one the most abun-dant natural fiber constituting the exoskeleton of crus-taceans and insects, also found in cell walls of fungal mycelia, etc. Chitosan is chemically analogous to cel-lulose in which the hydroxyl at C-2 is substituted by an amino group.
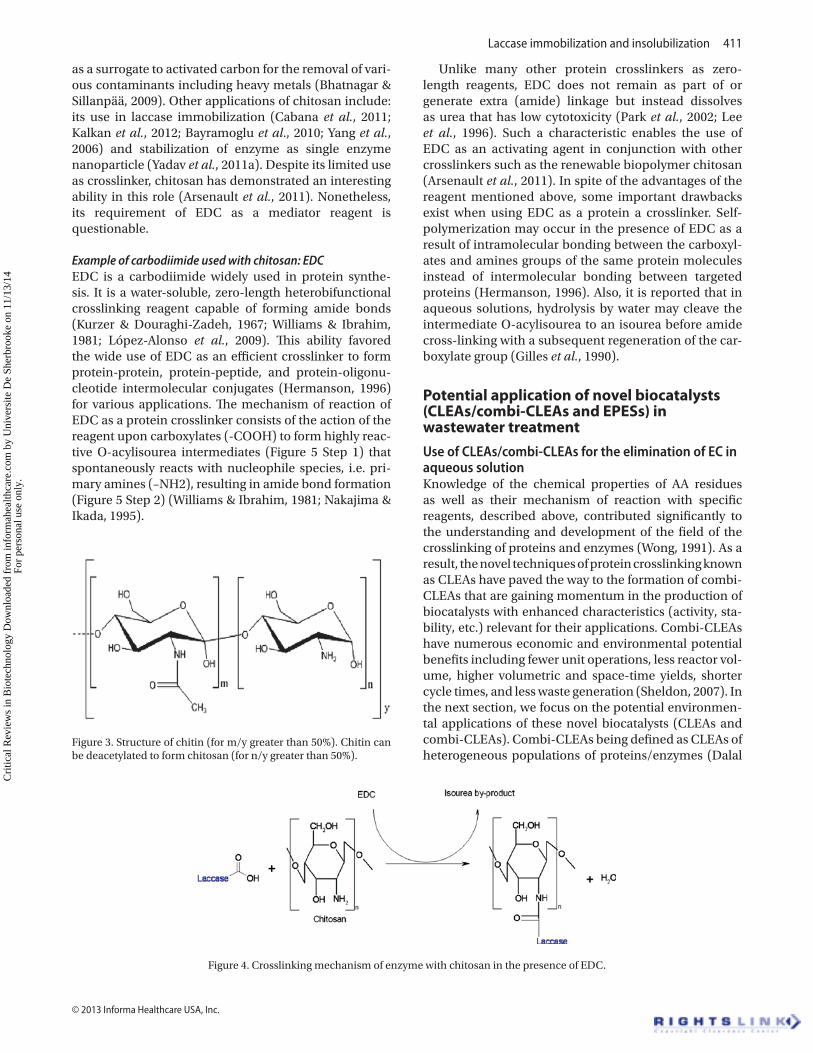
The polyglucosamine is hydrophobic and nearly insol-uble. Its solubility starts at 80% to higher deacetylation (Krajewska, 2004) in pure water but dissolves in dilute acids and is highly basic, unlike most naturally occurring polysaccharides found to be neutral or acidic (Majeti, 2000). Additional properties of chitosan comprise high stability and reactivity with great chelation ability to a wide number of compounds, biodegradability and nontoxicity to the environment, and remarkable affinity to proteins (Krajewska, 2004) rendering it very attractive for various applications including the use as crosslinking agents. In protein engineering, chitosan has been used as a sup-port or matrix to immobilize enzymes in several experi-ments but can also serve as crosslinker with the aid of an agent that activates the carboxylic groups of the enzyme (Arsenault et al., 2011; Cabana et al., 2011). Therefore, the crosslinking mechanism of protein with chitosan is feasible by simply using any of the protein carboxylic-activating agents such as 1-ethyl-3-(3-dimethylamino isopropyl) carbodiimide hydrochloride (EDC) to form amino-reactive derivative. The amino groups of chitosan then react with this derivative to form covalent bindings as crosslinks as illustrated in Figure 4.
Recently, chitosan has gained much interest in water and wastewater treatment due to its high adsorption capability (high content of amino and hydroxyl groups)
Figure 2. Synthesis of dextran to dextran aldehyde by oxidation with sodium periodate [adapted from Liu et al. (2009a)].
Cri
tical
Rev
iew
s in
Bio
tech
nolo
gy D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsite
De
Sher
broo
ke o
n 11
/13/
14Fo
r pe
rson
al u
se o
nly.
Laccase immobilization and insolubilization 411
© 2013 Informa Healthcare USA, Inc.
as a surrogate to activated carbon for the removal of vari-ous contaminants including heavy metals (Bhatnagar & Sillanpää, 2009). Other applications of chitosan include: its use in laccase immobilization (Cabana et al., 2011; Kalkan et al., 2012; Bayramoglu et al., 2010; Yang et al., 2006) and stabilization of enzyme as single enzyme nanoparticle (Yadav et al., 2011a). Despite its limited use as crosslinker, chitosan has demonstrated an interesting ability in this role (Arsenault et al., 2011). Nonetheless, its requirement of EDC as a mediator reagent is questionable.
Example of carbodiimide used with chitosan: EDCEDC is a carbodiimide widely used in protein synthe-sis. It is a water-soluble, zero-length heterobifunctional crosslinking reagent capable of forming amide bonds (Kurzer & Douraghi-Zadeh, 1967; Williams & Ibrahim, 1981; López-Alonso et al., 2009). This ability favored the wide use of EDC as an efficient crosslinker to form protein-protein, protein-peptide, and protein-oligonu-cleotide intermolecular conjugates (Hermanson, 1996) for various applications. The mechanism of reaction of EDC as a protein crosslinker consists of the action of the reagent upon carboxylates (-COOH) to form highly reac-tive O-acylisourea intermediates (Figure 5 Step 1) that spontaneously reacts with nucleophile species, i.e. pri-mary amines (–NH2), resulting in amide bond formation (Figure 5 Step 2) (Williams & Ibrahim, 1981; Nakajima & Ikada, 1995).
Unlike many other protein crosslinkers as zero-length reagents, EDC does not remain as part of or generate extra (amide) linkage but instead dissolves as urea that has low cytotoxicity (Park et al., 2002; Lee et al., 1996). Such a characteristic enables the use of EDC as an activating agent in conjunction with other crosslinkers such as the renewable biopolymer chitosan (Arsenault et al., 2011). In spite of the advantages of the reagent mentioned above, some important drawbacks exist when using EDC as a protein a crosslinker. Self-polymerization may occur in the presence of EDC as a result of intramolecular bonding between the carboxyl-ates and amines groups of the same protein molecules instead of intermolecular bonding between targeted proteins (Hermanson, 1996). Also, it is reported that in aqueous solutions, hydrolysis by water may cleave the intermediate O-acylisourea to an isourea before amide cross-linking with a subsequent regeneration of the car-boxylate group (Gilles et al., 1990).
Potential application of novel biocatalysts (cleAs/combi-cleAs and ePeSs) in wastewater treatment
Use of CLEAs/combi-CLEAs for the elimination of EC in aqueous solutionKnowledge of the chemical properties of AA residues as well as their mechanism of reaction with specific reagents, described above, contributed significantly to the understanding and development of the field of the crosslinking of proteins and enzymes (Wong, 1991). As a result, the novel techniques of protein crosslinking known as CLEAs have paved the way to the formation of combi-CLEAs that are gaining momentum in the production of biocatalysts with enhanced characteristics (activity, sta-bility, etc.) relevant for their applications. Combi-CLEAs have numerous economic and environmental potential benefits including fewer unit operations, less reactor vol-ume, higher volumetric and space-time yields, shorter cycle times, and less waste generation (Sheldon, 2007). In the next section, we focus on the potential environmen-tal applications of these novel biocatalysts (CLEAs and combi-CLEAs). Combi-CLEAs being defined as CLEAs of heterogeneous populations of proteins/enzymes (Dalal
Figure 3. Structure of chitin (for m/y greater than 50%). Chitin can be deacetylated to form chitosan (for n/y greater than 50%).
Figure 4. Crosslinking mechanism of enzyme with chitosan in the presence of EDC.
Cri
tical
Rev
iew
s in
Bio
tech
nolo
gy D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsite
De
Sher
broo
ke o
n 11
/13/
14Fo
r pe
rson
al u
se o
nly.

412 S. Ba et al.
Critical Reviews in Biotechnology
et al., 2007), the potential applications of both biocata-lysts are not discriminated henceforth.
CLEAs/combi-CLEAs have many intrinsic advantages, as mentioned earlier, that favor their potential applications. The circumvention of carriers in the synthesis of these novel biocatalysts yields a higher volumetric catalyst with little non-catalytic mass which in turn results in a reduced volume of reactor design for the usage of the biocatalysts. In fact, reactor design and kinetic parameters are essen-tial features to be taken into account for the application of biocatalysts in the biotransformation of environmen-tal contaminants found at micro- to nano-gram per liter concentrations. In applying immobilized/insolubilized proteins including these novel biocatalysts, various reac-tor designs have been used either in batch or continuous processes. In batch mode, an applied biocatalyst can be recovered by filtration or centrifugation whereas in con-tinuous mode such as packed bed reactor, biocatalyst of fairly large particles can be used in a fashion that large pres-sure drop over the column is prevented while ensuring that minimization of diffusion limitations for efficient catalysis is obtained (Sheldon, 2011).
Using laccase CLEAs from C. polyzona in a fluid-ized bed reactor (FBR) and a perfusion basket reactor, Cabana et al. (2007b) and Cabana et al. (2009a) were able to eliminate more than 95% of a branched isomer of nonylphenol, triclosan and bisphenol A from an aqueous solution at a 150-min residence time. In batch experiments, combi-CLEAs of versatile peroxidase (VP) and glucose oxidase (GOD) have been shown to elimi-nate several known endocrine disrupting compounds with more than 90% for bisphenol A (BPA), nonylphenol (NP), 17a-ethinylestradiol (EE2), and 17b-estradiol (E2)
and about 25% for triclosan (TCS) (Taboada-Puig et al., 2011). In the same study by Taboada-Puig and coworkers (2011), more than 90% of BPA were eliminated with the VP-GOD combi-CLEAs in a continuous process using a membrane bioreactor.
For the elimination of these micropollutants in actual wastewater, the kinetic parameters and reactor design are very important as explained below.
Kinetics for applied CLEAs/combi-CLEAs in micropollutants eliminationIn the experiments performed by Cabana et al. (2007b) and Cabana et al. (2009a), the Michaelis-Menten kinetics constants, K
M, values for the oxidation of ABTS for free
and insolubilized enzymes were about the same (32.0 and 28.5–31.9 µM, respectively). Likewise, in the experi-ment by Taboada-Puig et al. (2011), the K
M values for
H2O
2 (co-substrate for VP catalysis) were only slightly dif-
ferent (37.8, 36.3 and 27.5 µM, for free VP, free VP–GOD and VP–GOD-CLEAs, respectively). This is in agreement with the two preceding results and indicated that the insolubilization of the free enzymes as combi-CLEAs did not create diffusion limitations for the oxidation of the model substrate (i.e. DMP) in contrast with other immo-bilization techniques (i.e. on beads as carriers) applied elsewhere (Nicolucci et al., 2011; D’Annibale et al., 2000). Additionally, in an experiment by Honda et al. (2005), the biotransformation by hydrolysis of the synthetic sub-strate for α-chymotrypsin, N-glutaryl-L-phenylalanine p-nitroanilide led to kinetic constants K
M and V
max of
0.50 mM and 0.77 mM min−1, respectively. These values are not significantly different with those of solution-phase batchwise reactions using the same enzyme/substrate
Figure 5. Two-step mechanism of carboxylate activation with EDC to form a reactive intermediate (step 1) with subsequent reaction with an amine group to form amide bonds with release of an isourea by-product (step 2) [adapted from Hermanson (1996)].
Cri
tical
Rev
iew
s in
Bio
tech
nolo
gy D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsite
De
Sher
broo
ke o
n 11
/13/
14Fo
r pe
rson
al u
se o
nly.

Laccase immobilization and insolubilization 413
© 2013 Informa Healthcare USA, Inc.
reaction. The results are also consistent with those pre-sented above. In contrast, the hydrolysis of the acylase-based CLEAs for its substrate acetyl-d, l-phenylalanine provided K
M and V
max values of 4.58 mM and 1.10 mM
min−1, respectively compared to 2.33 mM and 4.69 mM min−1 when using free acylase/acetyl-d, l-phenylalanine under the same reaction conditions. These significant dif-ferences in kinetic constants values imply that diffusion of the substrate to the enzyme active site was somehow limited, a fact the authors attributed to partial inhibition of the enzyme hydrolytic capability by the cross-linking agents (glutaraldehyde and paraformaldehyde).
The KM
values of the surrogate ECs to the biocatalysts reported from most experiments for eliminating these ECs result from their high concentration in the order of mg/L (Cabana et al., 2007b; Cabana et al., 2009a). Both these K
M values and the experimental concentrations are
generally far superior to the actual concentration of these micropollutants in real aquatic environments. The actual concentration levels found are in the order of ng/L to µg/L as the case for some endocrine disrupting compounds in wastewaters (Auriol et al., 2006; Servos et al., 2005; Belfroid et al., 1999; Liu et al., 2009b). In experiments, high concentrations of the micropollutants are used as a way to overcome the difficulties associated with the analytical techniques available to accurately assess these micropollutants in solution at very low concentrations. Such difficulties are circumvented by treatment of these micropollutants in solution through a proper CLEAs-based membrane bioreactor without need to exaggerate the concentration levels.
Potential for membrane bioreactors to effectively eliminate micropollutants in wastewaterAs most of the ECs found in waters and wastewaters are in the order of ng/L to µg/L, this concentration scale leads to an ineffective removal of these pollutants from conventional treatment systems (flocculation/coagula-tion, sedimentation, filtration, etc.) (Bolong et al., 2009) in contrast with membrane filtration. Indeed, it is reported that as water contaminated with micropollutants passes through membrane filters a fine layer of micro-particles accumulates at the interface between the membrane and the solution as a result of the clogging process. A phenom-enon due to the colloidal, organic, biological, metal oxide, etc. fouling that leads to a higher concentrations in the solution with these micropollutants in that region effect-ing their mass transfer coefficients (Plakas & Karabelas, 2011) which become comparable to those of particles with higher sizes and are retained (Schäfer et al., 2011). It is therefore envisionable; on the one hand, that an enzyme-based membrane bioreactor would provide the advantage of an apparent high concentration at the membrane-solution interface that would enhance the kinetic of the micropollutants elimination. The high concentration of the micropollutants created at that interface will result in an improvement of their physico-chemical removal that normally would not occur in a classical reactor. On
the other hand, a large surface area contact between the immobilized enzyme membrane and the contaminated solution for the catalysis of the targeted contaminants can be achieved.
There are many examples of membrane-based bioreactors used in the biotransformation of various compounds. Sorgedrager and coworkers were able to biotransform 75–85% of penicillin G to 6-amino peni-cillanic acid with CLEAs of penicillin amidase in con-tinuous mode (Sorgedrager et al., 2008). They found that this CLEAs-based bioreactor combines the advantages of high rates of reaction (negligible diffusion limita-tions) and easy separation of the CLEAs (retained by the membranes) from the substrate and the end-prod-ucts. Likewise, a biotransformation of micropolluants in aqueous solution as targets of insoluble enzymes can be reasonably envisioned in such type of reac-tors. Microchannel reactors (microreactors) have been designed with α-chymotrypsin-based CLEAs (Honda et al., 2005) and acylase-based CLEAs (Honda et al., 2006) immobilized on the surface of the inner wall of polytetrafluoroethylene (PTFE) tubing to form a water-insoluble enzyme-membrane used in hydrolysis reac-tions. Such microreactors provide rapid heat and mass transfer, and large surface/interface area (Honda et al., 2006) favorable to catalytic reactions (Kusakabe et al., 2001) and therefore can be effective for biotransforma-tions. Large surface area to volume ratio of membrane reactors is an important feature of the catalytic process as it enables long contact times of reactants with the enzymes. Bioreactors in which Candida rugosa lipase CLEAs were formed inside both microporous hydro-philic cellulose and hydrophobic PTFE membranes were also used to bio-convert respectively 42 and 35% of oleic acid to butyl oleate (Hilal et al., 2004). Similarly, lipase has been entrapped in the porous structure of hollow-fiber membranes for large scale conversion of methyl ester of 4-methoxy-3-phenylglycidic acid to diltiazem, a drug used in the treatment of hypertension and angina (Lopez & Matson, 1997).
Application of CLEAs in membrane reactors provides the possibility to use membranes with large pore sizes and thus minimizing biofouling and rapid clogging of the membrane, which are two major drawbacks for mem-brane reactors when using smaller size free enzymes (Taboada-Puig et al., 2011). The use of CLEAs also favors the recovery of the biocatalyst and leads to a longer life-time of the reactor, whence the economical benefits.
Although there is no report of laccase-/laccases CLEAs-based bioreactor to our knowledge, from the examples of applications reported above, it becomes obvious that the use of enzymes including laccase CLEAs-based membrane bioreactor for the biotrans-formation of micropollutant of emerging concern in wastewater is a promising route to take. Ultimately, when using CLEAs/combi-CLEAs for the enzyme-cata-lyzed transformation of contaminants in solution, these novel biocatalysts can provide higher performance by
Cri
tical
Rev
iew
s in
Bio
tech
nolo
gy D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsite
De
Sher
broo
ke o
n 11
/13/
14Fo
r pe
rson
al u
se o
nly.

414 S. Ba et al.
Critical Reviews in Biotechnology
increasing their stabilization using surface coating poly-mer engineered structures.
Enzyme polymer engineered structures (EPES)As previously reported, the formation of laccase-based CLEAs significantly increases the stability against harsh conditions such as elevated temperatures and the pres-ence of organic, inorganic or biological denaturants in solution (Cabana et al., 2007b; Arsenault et al., 2011; Matijošyte et al., 2010; Schoevaart et al., 2006). Even if insolublized as crosslinked enzyme aggregates, enzymes generally present higher stability than their free counter-parts (from 1- to 10-fold, depending on the denaturing conditions used), the economic viability of enzymatic bioprocesses requires a drastic improvement of CLEAs stability. This can be achieved by a reengineering of the surface of the enzymes present in CLEAs referred to enzyme polymer engineered structures (EPESs). Essentially, this approach is based on the conjugation of the enzymes with a polymer through the formation of covalent bounds between non-essential AA residues pres-ent at the surface of the CLEAs and the selected polymer. The resulting network should significantly increase the stability of the resulting biocatalysts (Kim & Grate, 2003, 2005; Yadav et al., 2011a,b; Khosravi et al., 2012). This stabilization effect is due to the multipoint attachment of the enzyme and the presence of the polymeric network which enhances the tridimensional conformational sta-bility of enzymes. Also, according to several studies, the protecting network did not induce steric hindrance which could reduce the mass-transfer of the substrate (Hegedus & Nagy, 2009; Yadav et al., 2011a; Khosravi et al., 2012). It appears that the network surrounding the enzyme is sufficiently porous and thick to allow the migration of the substrates into the enzyme’s active site.
To date, this approach has been used only for the modification of free enzymes. However, data from stud-ies in our laboratory (under review for separate publi-cation) have shown the potential for this approach to significantly increase the stability of free laccase- and laccase CLEA-based. Several strategies have been pro-posed for the formation of protecting layers surrounding the enzymes. Essentially, they involved (1) a conjugation step where the selected polymer is bound to the targeted amino acid(s) at the surface of the enzyme, and (2) a maturation step (Figure 6).
Kim and Grate (2003) and Hegedus and Nagy (2009) prepared single enzyme nanoparticles (SENs) with proteolytic enzymes. They used a three-step procedure involving the anchoring of a vinyl group at the surface of the enzyme, the addition of a silane monomer and the orthogonal polymerization of this monomer under UV radiation (Kim et al., 2006). The resulting SENs have shown a half-life of more than 140 days compared to 12 hours for their free counterparts (Kim & Grate, 2003). Additionally, Yadav et al. (2011a,b) have stabilized carbonic anhydrase (CA) through the formation of a chitosan/3-amino propyltriethoxysilane hybrid polymer.
The resulted CA-SENs remained stable for more than 100 days at either −20, 4 or 20°C. In another study, Khosravi et al. (2012) have developed a simple approach for the formation of a dendritic polyester-based network sur-rounding horseradish peroxidase (HRP). The formed dendrozymes present small changes in the protein two- and three-dimensional conformation, which result in a reengineered biocatalyst with catalytic properties similar to free enzyme used. In addition, these novel biocatalysts showed significant stability increase against thermal and chemical denaturation compared to their free counter-parts. HRP dendrozymes maintained more than 75% of their initial activity after 90-min incubation at 65°C and 15-min incubation in organic solvent containing solu-tions (10% v/v) at pH 7.2 and 40°C while the free HRP have lost 90–100% of their initial activity under the same denaturing conditions.
Previous work has shown the beneficial effects of poly-meric modification of different free enzymes. After the bioconjugation of these proteins with various (macro) molecules, their potential for the development of bio-processes has been significantly increased because of the drastic improvements of their stability against operational conditions without altering their kinetic behavior. The techniques of EPESs initially developed for free enzymes appear suitable for the development of a new generation of crosslinked enzyme aggregates with enhanced proper-ties relevant for micropollutant elimination in wastewater.
conclusion
The technique of laccase insolubilization has great application potential in the field of wastewater bioreme-diation targeting large array of contaminants. Indeed, the chemical composition and structural arrangement of the enzyme favoring its simple oxidation capability with non-specific substrate has promoted its use for the elimination of numerous wastewater pollutants in several experiments as described above. An understanding of the chemical properties of proteins and crosslinking agents, as well as their subsequent interactions, translates to the ability to synthesize novel biocatalysts in CLEAs/combi-CLEAs and EPESs of higher characteristics indispensable for their large scale applications. These novel biocatalysts have been shown to possess biotransformation capability
Figure 6. Schematic representation of the formation of a stabilizing network surrounding free enzymes [adapted from Hegedus and Nagy (2009).
Cri
tical
Rev
iew
s in
Bio
tech
nolo
gy D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsite
De
Sher
broo
ke o
n 11
/13/
14Fo
r pe
rson
al u
se o
nly.

Laccase immobilization and insolubilization 415
© 2013 Informa Healthcare USA, Inc.
with respect to several compounds when used in suitable designs of bioreactors. Likewise, we are confident that insoluble laccases in the form of combi-CLEAs and EPESs have great potential to bioconvert contaminants of health and environmental concerns such as emerging contami-nants in wastewaters into innocuous products. Finally, it is possible that an enzyme-based membrane bioreactor would provide an incomparable advantage in biotrans-formation of emerging contaminants in wastewater. New direction may be to develop combi-CLEAs and EPESs and an enzyme-based membrane bioreactor to be used for the elimination of emerging contaminants in wastewater.
Declaration of interest
This project was financially supported by a grant from the Natural Sciences and Engineering Research Council of Canada (NSERC).
ReferencesAbadulla E, Tzanov T, Costa S, Robra KH, Cavaco-Paulo A, Gübitz
GM. 2000. Decolorization and detoxification of textile dyes with a laccase from Trametes hirsuta. Appl Environ Microbiol 66: 3357–3362.
Ahn MY, Dec J, Kim JE, Bollag JM. 2002. Treatment of 2,4-dichlorophenol polluted soil with free and immobilized laccase. J Environ Qual 31: 1509–1515.
Andréasson LE, Reinhammar B. 1979. The mechanism of electron transfer in laccase-catalysed reactions. Biochim Biophys Acta 568: 145–156.
Arboleda C, Cabana H, De Pril E, Jones JP, Jimenez GA, Mejia AI, Agathos SN, Penninckx MJ. 2012. Elimination of bisphenol A and triclosan using the enzymatic system of autochthonous Colombian forest fungi. ISRN Biotechnol 2013: 968241.
Arsenault A, Cabana H, Jones JP. 2011. Laccase-Based CLEAs: Chitosan as a Novel Cross-Linking Agent. Enzyme Res 2011: 376015.
Auriol M, Filali-Meknassi Y, Adams CD, Tyagi RD. 2006. Natural and synthetic hormone removal using the horseradish peroxidase enzyme: temperature and pH effects. Water Res 40: 2847–2856.
Auriol M, Filali-Meknassi Y, Adams CD, Tyagi RD, Noguerol TN, Piña B. 2008. Removal of estrogenic activity of natural and synthetic hormones from a municipal wastewater: efficiency of horseradish peroxidase and laccase from Trametes versicolor. Chemosphere 70: 445–452.
Auriol M, Filali-Meknassi Y, Tyagi RD, Adams CD. 2007. Laccase-catalyzed conversion of natural and synthetic hormones from a municipal wastewater. Water Res 41: 3281–3288.
Aust SD, Benson JT. 1993. The fungus among us: use of white rot fungi to biodegrade environmental pollutants. Environ Health Perspect 101: 232–233.
Aytar BS, Bakir U. 2008. Preparation of cross-linked tyrosinase aggregates. Process Biochemi 43:125–131.
Baldrian P. 2006. Fungal laccases - occurrence and properties. FEMS Microbiol Rev 30: 215–242.
Bayramoglu G, Yilmaz M, Yakup Arica M. 2010. Preparation and characterization of epoxy-functionalized magnetic chitosan beads: laccase immobilized for degradation of reactive dyes. Bioprocess Biosyst Eng 33: 439–448.
Belfroid AC, Van der Horst A, Vethaak AD, Schäfer AJ, Rijs GB, Wegener J, Cofino WP. 1999. Analysis and occurrence of estrogenic hormones and their glucuronides in surface water and waste water in The Netherlands. Sci Total Environ 225: 101–108.
Berka RM, Schneider P, Golightly EJ, Brown SH, Madden M, Brown KM, Halkier T, Mondorf K, Xu F. 1997. Characterization of the gene encoding an extracellular laccase of Myceliophthora thermophila
and analysis of the recombinant enzyme expressed in Aspergillus oryzae. Appl Environ Microbiol 63: 3151–3157.
Bertrand T, Jolivalt C, Briozzo P, Caminade E, Joly N, Madzak C, Mougin C. 2002. Crystal structure of a four-copper laccase complexed with an arylamine: insights into substrate recognition and correlation with kinetics. Biochemistry 41: 7325–7333.
Bhatnagar A, Sillanpää M. 2009. Applications of chitin- and chitosan-derivatives for the detoxification of water and wastewater–a short review. Adv Colloid Interface Sci 152: 26–38.
Bollag JM, Shuttleworth KL, Anderson DH. 1988. Laccase-mediated detoxification of phenolic compounds. Appl Environ Microbiol 54: 3086–3091.
Bollag JM. 1992. Decontaminating soil with enzymes. Environ Sci Technol 26:1876–1881.
Bollag JM, Leonowicz A. 1984. Comparative studies of extracellular fungal laccases. Appl Environ Microbiol 48: 849–854.
Bolong N, Ismail AF, Salim MR, Matsuura T. 2009. A review of the effects of emerging contaminants in wastewater and options for their removal. Desalination 238:229–246.
Bourbonnais R, Leech D, Paice MG. 1998. Electrochemical analysis of the interactions of laccase mediators with lignin model compounds. Biochim Biophys Acta 1379: 381–390.
Bourbonnais R, Paice MG. 1990. Oxidation of non-phenolic substrates. An expanded role for laccase in lignin biodegradation. FEBS Lett 267: 99–102.
Bourbonnais R, Paice MG, Freiermuth B, Bodie E, Borneman S. 1997. Reactivities of various mediators and laccases with kraft pulp and lignin model compounds. Appl Environ Microbiol 63: 4627–4632.
Bruggink A, Schoevaart R, Kieboom T. 2003. Concepts of nature in organic synthesis: cascade catalysis and multistep conversions in concert. Org Process Res Develop 7:622–640.
Cabana H, Ahamed A, Leduc R. 2011. Conjugation of laccase from the white rot fungus Trametes versicolor to chitosan and its utilization for the elimination of triclosan. Bioresour Technol 102: 1656–1662.
Cabana H, Jiwan JL, Rozenberg R, Elisashvili V, Penninckx M, Agathos SN, Jones JP. 2007a. Elimination of endocrine disrupting chemicals nonylphenol and bisphenol A and personal care product ingredient triclosan using enzyme preparation from the white rot fungus Coriolopsis polyzona. Chemosphere 67: 770–778.
Cabana H, Jones JP, Agathos SN. 2007b. Preparation and characterization of cross-linked laccase aggregates and their application to the elimination of endocrine disrupting chemicals. J Biotechnol 132: 23–31.
Cabana H, Jones JP, Agathos SN. 2009a. Utilization of cross-linked laccase aggregates in a perfusion basket reactor for the continuous elimination of endocrine-disrupting chemicals. Biotechnol Bioeng 102: 1582–1592.
Cabana H, Alexandre C, Agathos SN, Jones JP. 2009b. Immobilization of laccase from the white rot fungus Coriolopsis polyzona and use of the immobilized biocatalyst for the continuous elimination of endocrine disrupting chemicals. Bioresour Technol 100: 3447–3458.
Cabral JMS, Kennedy JF. 1991. Covalent and coordination immobilization of proteins. In: Taylor RF, ed. Protein immobilization: fundamentals and applications. New York: Marcel Dekker, pp 73–138.
Call HP, Mucke I. 1997. History, overview and applications of mediated lignolytic systems, especially laccase-mediator-systems (Lignozym(R)-process). J Biotechnol 53:163–202.
Campbell MK, Farrell SO. 2006. Biochemistry. 5th edn. USA: Thomson Brooks/Cole.
Cao L. 2005. Immobilised enzymes: science or art? Curr Opin Chem Biol 9: 217–226.
Cao L, Langen L, Sheldon RA. 2003. Immobilised enzymes: carrier-bound or carrier-free? Curr Opin Biotechnol 14: 387–394.
Claus H, Faber G, König H. 2002. Redox-mediated decolorization of synthetic dyes by fungal laccases. Appl Microbiol Biotechnol 59: 672–678.
Couto SR, Herrera JLT. 2006. Industrial and biotechnological applications of laccases: A review. Biotechnol Adv 24:500–513.
Cri
tical
Rev
iew
s in
Bio
tech
nolo
gy D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsite
De
Sher
broo
ke o
n 11
/13/
14Fo
r pe
rson
al u
se o
nly.

416 S. Ba et al.
Critical Reviews in Biotechnology
Couto SR, Toca JL. 2006. Inhibitors of laccases: A review. Current Enzyme Inhibition 2:343–352.
Dalal S, Kapoor M, Gupta MN. 2007. Preparation and characterization of combi-CLEAs catalyzing multiple non-cascade reactions. J Mol Catal B: Enz 44:128–132.
D’Annibale A, Stazi SR, Vinciguerra V, Di Mattia E, Sermanni GG. 1999. Characterization of immobilized laccase from Lentinula edodes and its use in olive-mill wastewater treatment. Process Biochem (Barking, London, England), 34:697–706.
D’Annibale A, Stazi SR, Vinciguerra V, Giovannozzi Sermanni G. 2000. Oxirane-immobilized Lentinula edodes laccase: stability and phenolics removal efficiency in olive mill wastewater. J Biotechnol 77: 265–273.
Dimich-Ward H, Wymer ML, Chan-Yeung M. 2004. Respiratory health survey of respiratory therapists. Chest 126: 1048–1053.
Ducros V, Brzozowski AM, Wilson KS, Brown SH, Ostergaard P, Schneider P, Yaver DS, Pedersen AH, Davies GJ. 1998. Crystal structure of the type-2 Cu depleted laccase from Coprinus cinereus at 2.2 A resolution. Nat Struct Biol 5: 310–316.
Duran N, Rosa MA, D’Annibale A, Gianfreda L. 2002. Applications of laccases and tyrosinases (phenoloxidases) immobilized on different supports: a review. Enzyme Microb Technol 31:907–931.
Emmanuel E, Hanna K, Bazin C, Keck G, Clément B, Perrodin Y. 2005. Fate of glutaraldehyde in hospital wastewater and combined effects of glutaraldehyde and surfactants on aquatic organisms. Environ Int 31: 399–406.
Epton R, McLaren JV, Thomas TH. 1971. Enzyme insolubilization with cross-linked polyacryloylaminoacetaldehyde dimethylacetal. Biochem J 123: 21P–22P.
Fernández-Lorente G, Palomo JM, Mateo C, Munilla R, Ortiz C, Cabrera Z, Guisán JM, Fernandez-Lafuente R. 2006. Glutaraldehyde cross-linking of lipases adsorbed on aminated supports in the presence of detergents leads to improved performance. Biomacromolecules 7: 2610–2615.
Ferraroni M, Myasoedova NM, Schmatchenko V, Leontievsky AA, Golovleva LA, Scozzafava A, Briganti F. 2007. Crystal structure of a blue laccase from Lentinus tigrinus: evidences for intermediates in the molecular oxygen reductive splitting by multicopper oxidases. BMC Struct Biol 7: 60.
Fraenkel-Conrat H, Mecham DK. 1949. The reaction of formaldehyde with proteins; demonstration of intermolecular cross-linking by means of osmotic pressure measurements. J Biol Chem 177: 477–486.
Fukuda T, Uchida H, Suzuki M, Miyamoto H, Morinaga H, Nawata H, Uwajima T. 2004. Transformation products of bisphenol A by a recombinant Trametes villosa laccase and their estrogenic activity. J Chem Technol Biotechnol 79:1212–1218.
Fukuda T, Uchida H, Takashima Y, Uwajima T, Kawabata T, Suzuki M. 2001. Degradation of bisphenol A by purified laccase from Trametes villosa. Biochem Biophys Res Commun 284: 704–706.
Gaitan IJ, Medina SC, González JC, Rodríguez A, Espejo AJ, Osma JF, Sarria V, Alméciga-Díaz CJ, Sánchez OF. 2011. Evaluation of toxicity and degradation of a chlorophenol mixture by the laccase produced by Trametes pubescens. Bioresour Technol 102: 3632–3635.
Garavaglia S, Cambria MT, Miglio M, Ragusa S, Iacobazzi V, Palmieri F, D’Ambrosio C, Scaloni A, Rizzi M. 2004. The structure of Rigidoporus lignosus Laccase containing a full complement of copper ions, reveals an asymmetrical arrangement for the T3 copper pair. J Mol Biol 342: 1519–1531.
Garcia HA, Hoffman CM, Kinney KA, Lawler DF. 2011. Laccase-catalyzed oxidation of oxybenzone in municipal wastewater primary effluent. Water Res 45: 1921–1932.
Giardina P, Faraco V, Pezzella C, Piscitelli A, Vanhulle S, Sannia G. 2010. Laccases: a never-ending story. Cell Mol Life Sci 67: 369–385.
Gigi O, Marbach I, Mayer AM. 1981. Properties of gallic acid-induced extracellular laccase of Botrytis cinerea. Phytochemistry 20:1211–1213.
Gilles MA, Hudson AQ, Borders CL Jr. 1990. Stability of water-soluble carbodiimides in aqueous solution. Anal Biochem 184: 244–248.
Hadorn H, Nitschmann HH. 1944. Die Umsetzung des Caseins mit Formaldehyd. V. Über das verhalten der ε-aminogruppen des lysins und der peptidgruppen. Helvetica Chimica Acta 27:299–312.
Habeeb AJ, Hiramoto R. 1968. Reaction of proteins with glutaraldehyde. Arch Biochem Biophys 126: 16–26.
Hakulinen N, Kiiskinen LL, Kruus K, Saloheimo M, Paananen A, Koivula A, Rouvinen J. 2002. Crystal structure of a laccase from Melanocarpus albomyces with an intact trinuclear copper site. Nat Struct Biol 9: 601–605.
Hanefeld U, Gardossi L, Magner E. 2009. Understanding enzyme immobilisation. Chem Soc Rev 38: 453–468.
Harms H, Schlosser D, Wick LY. 2011. Untapped potential: exploiting fungi in bioremediation of hazardous chemicals. Nat Rev Microbiol 9: 177–192.
Hegedus I, Nagy E. 2009. Improvement of chymotrypsin enzyme stability as single enzyme nanoparticles. Chem Eng Sci 64:1053–1060.
Heinz Fraenkel-Conrat, Olcott HS. 1948. The reaction of formaldehyde with proteins. V. Cross-linking between Amino and Primary Amide or Guanidyl Groups. J Am Chem Soc 70:2673–2684.
Hermanson GT. 1996. Bioconjugate techniques. San Diego: Academic Press.
Hilal N, Nigmatullin R, Alpatova A. 2004. Immobilization of cross-linked lipase aggregates within microporous polymeric membranes. J Memb Sci 238:131–141.
Honda T, Miyazaki M, Nakamura H, Maeda H. 2006. Facile preparation of an enzyme-immobilized microreactor using a cross-linking enzyme membrane on a microchannel surface. Adv Synth Catal 348:2163–2171.
Honda T, Miyazaki M, Nakamura H, Maeda H. 2005. Immobilization of enzymes on a microchannel surface through cross-linking polymerization. Chem Commun (Camb): 5062–5064.
Johannes C, Majcherczyk A. 2000. Natural mediators in the oxidation of polycyclic aromatic hydrocarbons by laccase mediator systems. Appl Environ Microbiol 66: 524–528.
Kalkan NA, Aksoy S, Aksoy EA, Hasirci N. 2012. Preparation of chitosan-coated magnetite nanoparticles and application for immobilization of laccase. J Appl Poly Sci 123:707–716.
Kallio JP, Gasparetti C, Andberg M, Boer H, Koivula A, Kruus K, Rouvinen J, Hakulinen N. 2011. Crystal structure of an ascomycete fungal laccase from Thielavia arenaria–common structural features of asco-laccases. FEBS J 278: 2283–2295.
Khosravi A, Vossoughi M, Shahrokhian S, Alemzadeh I. 2012. Nano reengineering of horseradish peroxidase with dendritic macromolecules for stability enhancement. Enzyme Microb Technol 50: 10–16.
Kim J, Grate JW. 2005. Single enzyme nanoparticles. 05AIChE: 2005 AIChE Annual Meeting and Fall Showcase, 30 October 2005 through 4 November 2005, Cincinnati, OH, pp. 14286.
Kim J, Grate JW. 2003. Single-enzyme nanoparticles armored by a nanometer-scale organic/inorganic network. Nano Letters 3:1219–1222.
Kim J, Grate JW, Wang P. 2006. Nanostructures for enzyme stabilization. Chem Eng Sci 61:1017–1026.
Kim YJ, Nicell JA. 2006. Laccase-catalyzed oxidation of bisphenol A with the aid of additives. Process Biochem, 41:1029–1037.
Kimura K, Amy G, Drewes JE, Heberer T, Kim T, Watanabe Y. 2003. Rejection of organic micropollutants (disinfection by-products, endocrine disrupting compounds, and pharmaceutically active compounds) by NF/RO membranes. J Memb Sci 227:113–121.
Kiso Y, Muroshige K, Oguchi T, Yamada T, Hirose M, Ohara T, Shintani T. 2010. Effect of molecular shape on rejection of uncharged organic compounds by nanofiltration membranes and on calculated pore radii. J Memb Sci 358:101–113.
Krajewska B. 2004. Application of chitin- and chitosan-based materials for enzyme immobilizations: a review. Enz Microbial Technol 35:126–139.
Kunamneni A, Plou FJ, Ballesteros A, Alcalde M. 2008. Laccases and their applications: a patent review. Recent Pat Biotechnol 2: 10–24.
Cri
tical
Rev
iew
s in
Bio
tech
nolo
gy D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsite
De
Sher
broo
ke o
n 11
/13/
14Fo
r pe
rson
al u
se o
nly.

Laccase immobilization and insolubilization 417
© 2013 Informa Healthcare USA, Inc.
Kurzer F, Douraghi-Zadeh K. 1967. Advances in the chemistry of carbodiimides. Chem Rev 67: 107–152.
Kusakabe K, Miyagawa D, Gu Y, Maeda H, Morooka S. 2001. Development of self-heating microreactor for catalytic reactions. J Chem Eng Jpn 34:441–443.
Lee JM, Edwards HHL, Pereira CA, Samii SI. 1996. Crosslinking of tissue-derived biomaterials in 1-ethyl-3-(3-dimethylaminopropyl)-carbodiimide (EDC). J Mater Sci Mater Med 7:531–541.
Leonowicz A, Sarkar JM, Bollag J. 1988. Improvement in stability of an immobilized fungal laccase. Appl Microbiol Biotechnol 29:129–135.
Leung HW. 2001. Ecotoxicology of glutaraldehyde: review of environmental fate and effects studies. Ecotoxicol Environ Saf 49: 26–39.
Li K, Xu F, Eriksson KE. 1999. Comparison of fungal laccases and redox mediators in oxidation of a nonphenolic lignin model compound. Appl Environ Microbiol 65: 2654–2660.
Liu G, Shi Z, Kuriger T, Hanton LR, Simpson J, Moratti SC, Robinson BH, Athanasiadis T, Valentine R, Wormald PJ, Robinson S. 2009a. Synthesis and characterization of chitosan/dextran-based hydrogels for surgical use. Macromolecular Symposia 279:151–157.
Liu ZH, Kanjo Y, Mizutani S. 2009b. Removal mechanisms for endocrine disrupting compounds (EDCs) in wastewater treatment - physical means, biodegradation, and chemical advanced oxidation: a review. Sci Total Environ 407: 731–748.
Lopez JL, Matson SL. 1997. A multiphase/extractive enzyme membrane reactor for production of diltiazem chiral intermediate. J Memb Sci 125:189–211.
López-Alonso JP, Diez-García F, Font J, Ribó M, Vilanova M, Scholtz JM, González C, Vottariello F, Gotte G, Libonati M, Laurents DV. 2009. Carbodiimide EDC induces cross-links that stabilize RNase A C-dimer against dissociation: EDC adducts can affect protein net charge, conformation, and activity. Bioconjug Chem 20: 1459–1473.
Madhavi V, Lele SS. 2009. Laccase: Properties and Applications. BioResources 4:1694–1717.
Majeau JA, Brar SK, Tyagi RD. 2010. Laccases for removal of recalcitrant and emerging pollutants. Bioresour Technol 101: 2331–2350.
Majeti NVRK. 2000. A review of chitin and chitosan applications. React Funct Poly 46:1–27.
Marbach I, Harel E, Mayer AM. 1984. Molecular properties of extracellular Botrytis cinerea laccase. Phytochemistry 23:2713–2717.
Mateo C, Palomo JM, van Langen LM, van Rantwijk F, Sheldon RA. 2004. A new, mild cross-linking methodology to prepare cross-linked enzyme aggregates. Biotechnol Bioeng 86: 273–276.
Mateo C, Palomo JM, Fernandez-Lorente G, Guisan JM, Fernandez-Lafuente R. 2007. Improvement of enzyme activity, stability and selectivity via immobilization techniques. Enz Microbial Technol 40:1451–1463.
Mateo C, Palomo JM, Van Langen LM, Van Rantwijk F, Sheldon RA. 2004. A new, mild cross-linking methodology to prepare cross-linked enzyme aggregates. Biotechnol Bioeng 86:273–276.
MatijošyteI, Arends IWCE, de Vries S, Sheldon RA. 2010. Preparation and use of cross-linked enzyme aggregates (CLEAs) of laccases. J Mol Cataly B: Enz 62:142–148.
Messerschmidt A. 1997. Multi-copper Oxidases, World Scientific, Singapore.
Messerschmidt A, Huber R. 1990. The blue oxidases, ascorbate oxidase, laccase and ceruloplasmin. Modelling and structural relationships. Eur J Biochem 187: 341–352.
Migneault I, Dartiguenave C, Bertrand MJ, Waldron KC. 2004. Glutaraldehyde: behavior in aqueous solution, reaction with proteins, and application to enzyme crosslinking. BioTechniques 37: 790–6, 798.
Minuth W, Klischies M, Esser K. 1978. The phenoloxidases of the ascomycete Podospora anserina. Structural differences between laccases of high and low molecular weight. Eur J Biochem 90: 73–82.
Muñoz C, Guillén F, Martínez AT, Martínez MJ. 1997. Laccase isoenzymes of Pleurotus eryngii: characterization, catalytic properties, and participation in activation of molecular oxygen and Mn2+ oxidation. Appl Environ Microbiol 63: 2166–2174.
Nakajima N, Ikada Y. 1995. Mechanism of amide formation by carbodiimide for bioconjugation in aqueous media. Bioconjug Chem 6: 123–130.
Nghiem LD, Schäfer AI. 2002. Adsorption and transport of trace contaminant estrone in NF/RO membranes. Environ Eng Sci 19:441–451.
Nicolucci C, Rossi S, Menale C, Godjevargova T, Ivanov Y, Bianco M, Mita L, Bencivenga U, Mita DG, Diano N. 2011. Biodegradation of bisphenols with immobilized laccase or tyrosinase on polyacrylonitrile beads. Biodegradation 22: 673–683.
Osma JF, Toca-Herrera JL, Rodríguez-Couto S. 2011. Cost analysis in laccase production. J Environ Manage 92: 2907–2912.
Park SN, Park JC, Kim HO, Song MJ, Suh H. 2002. Characterization of porous collagen/hyaluronic acid scaffold modified by 1-ethyl-3-(3-dimethylaminopropyl)carbodiimide cross-linking. Biomaterials 23: 1205–1212.
Piontek K, Antorini M, Choinowski T. 2002. Crystal structure of a laccase from the fungus Trametes versicolor at 1.90-A resolution containing a full complement of coppers. J Biol Chem 277: 37663–37669.
Plakas KV, Karabelas AJ. 2011. A systematic study on triazine retention by fouled with humic substances NF/ULPRO membranes. Sep PurifiTechnol 80:246–261.
Quiocho FA, Richards FM. 1964. Intermolecular cross linking of a protein in the crystalline state: carboxypeptidase-A. Proc Natl Acad Sci USA 52: 833–839.
Reiss R, Ihssen J, Thöny-Meyer L. 2011. Bacillus pumilus laccase: a heat stable enzyme with a wide substrate spectrum. BMC Biotechnol 11: 9.
Ricotta A, Unz RF, Bollag J. 1996. Role of a laccase in the degradation of pentachlorophenol. Bull Environ Contam Toxicol 57: 560–567.
Roessl U, Nahálka J, Nidetzky B. 2010. Carrier-free immobilized enzymes for biocatalysis. Biotechnol Lett 32: 341–350.
Roy JJ, Abraham TE, Abhijith KS, Kumar PV, Thakur MS. 2005. Biosensor for the determination of phenols based on cross-linked enzyme crystals (CLEC) of laccase. Biosens Bioelectron 21: 206–211.
Roy-Arcand L, Archibald FS. 1991. Direct dechlorination of chlorophenolic compounds by laccases from Trametes (Coriolus) versicolor. Enzyme and Microbial Technology 13:194–203.
Sangeetha K, Abraham TE. 2008. Preparation and characterization of cross-linked enzyme aggregates (CLEA) of subtilisin for controlled release applications. Int J Biol Macromol 43: 314–319.
Sano LL, Krueger AM, Landrum PF. 2005. Chronic toxicity of glutaraldehyde: differential sensitivity of three freshwater organisms. Aquat Toxicol 71: 283–296.
Schäfer AI, Akanyeti I, Semião AJ. 2011. Micropollutant sorption to membrane polymers: a review of mechanisms for estrogens. Adv Colloid Interface Sci 164: 100–117.
Schoevaart R, Kieboom T. 2002a. Combined catalytic conversion involving an enzyme, a homogeneous and a heterogeneous catalyst: One-pot preparation of 4-deoxy-D-glucose derivatives from D-galactose. Tetrahedron Lett 43:3399–3400.
Schoevaart R, Kieboom T. 2002b. Galactose dialdehyde as potential protein cross-linker: proof of principle. Carbohydr Res 337: 899–904.
Schoevaart R, Kieboom T. 2001. Galactose dialdehyde: the forgotten candidate for a protein cross-linker? Carbohydr Res 334: 1–6.
Schoevaart R, Siebum A, van Rantwijk F, Sheldon RA, Kieboom T. 2005. Glutaraldehyde cross-link analogues from carbohydrates. Starch 57:161.
Schoevaart R, Wolbers MW, Golubovic M, Ottens M, Kieboom AP, van Rantwijk F, van der Wielen LA, Sheldon RA. 2004. Preparation, optimization, and structures of cross-linked enzyme aggregates (CLEAs). Biotechnol Bioeng 87: 754–762.
Schoevaart WRK, Van Langen LM, Van Den Dool RTM, Boumans JWL. 2006. Method for preparation of crosslinked enzyme aggregates using aldehydes and amines. US Patent 2008/0296231 A1.
Cri
tical
Rev
iew
s in
Bio
tech
nolo
gy D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsite
De
Sher
broo
ke o
n 11
/13/
14Fo
r pe
rson
al u
se o
nly.

418 S. Ba et al.
Critical Reviews in Biotechnology
Servos MR, Bennie DT, Burnison BK, Jurkovic A, McInnis R, Neheli T, Schnell A, Seto P, Smyth SA, Ternes TA. 2005. Distribution of estrogens, 17beta-estradiol and estrone, in Canadian municipal wastewater treatment plants. Sci Total Environ 336: 155–170.
Sheldon RA. 2011. Cross-linked enzyme aggregates as industrial biocatalysts. Org Process Res Develop 15:213–223.
Sheldon RA. 2007. Enzyme immobilization: The quest for optimum performance. Adv Syn Catal 349:1289–1307.
Solomon EI, Sundaram UM, Machonkin TE. 1996. Multicopper Oxidases and Oxygenases. Chem Rev 96: 2563–2606.
Sorgedrager MJ, Verdoes D, van der Meer H, Sheldon RA. 2008. Cross-linked enzyme aggregates in a membrane slurry reactor continuous production of 6-APA by enzymatic hydrolysis of penicillin. Chimica Oggi/Chemistry Today 24:23–25.
Srinivasan C, Dsouza TM, Boominathan K, Reddy CA. 1995. Demonstration of Laccase in the White Rot Basidiomycete Phanerochaete chrysosporium BKM-F1767. Appl Environ Microbiol 61: 4274–4277.
Strong PJ, Claus H. 2011. Laccase: A review of its past and its future in bioremediation. Crit Rev Environ Sci Technol 41:373–434.
Taboada-Puig R, Junghanns C, Demarche P, Moreira MT, Feijoo G, Lema JM, Agathos SN. 2011. Combined cross-linked enzyme aggregates from versatile peroxidase and glucose oxidase: production, partial characterization and application for the elimination of endocrine disruptors. Bioresour Technol 102: 6593–6599.
Takigawa T, Endo Y. 2006. Effects of glutaraldehyde exposure on human health. J Occup Health 48: 75–87.
Tanaka T, Tonosaki T, Nose M, Tomidokoro N, Kadomura N, Fujii T, Taniguchi M. 2001. Treatment of model soils contaminated with phenolic endocrine-disrupting chemicals with laccase from Trametes sp. in a rotating reactor. J Biosci Bioeng 92: 312–316.
ten Have R, Teunissen PJ. 2001. Oxidative mechanisms involved in lignin degradation by white-rot fungi. Chem Rev 101: 3397–3413.
Twyman RM. 2005. Enzymes: Immobilized enzymes. In: Encyclopedia of Analytical Science, 2nd edn. London, UK: Elsevier Science, pp 523–529.
Wells A, Teria M, Eve T. 2006. Green oxidations with laccase-mediator systems. Biochem Soc Trans 34: 304–308.
Williams A, Ibrahim IT. 1981. Carbodiimide chemistry: Recent advances. Chem Rev 81:589–636.
Wong SS. 1991. Chemistry of protein conjugation and cross linking. 1 edition edn. Boca Raton, Florida, USA: CRC Press.
Xu F. 1996. Oxidation of phenols, anilines, and benzenethiols by fungal laccases: correlation between activity and redox potentials as well as halide inhibition. Biochemistry 35: 7608–7614.
Yadav R, Labhsetwar N, Kotwal S, Rayalu S. 2011a. Single enzyme nanoparticle for biomimetic CO
2 sequestration. J Nanoparticle Res
13:263–271.Yadav R, Satyanarayanan T, Kotwal S, Rayalu S. 2011b. Enhanced
carbonation reaction using chitosan-based carbonic anhydrase nanoparticles. Current Sci 100:520–524.
Yang WY, Min DY, Wen SX, Jin L, Rong L, Tetsuo M, Bo C. 2006. Immobilization and characterization of laccase from Chinese Rhus vernicifera on modified chitosan. Process Biochem 41:1378–1382.
Yaropolov AI, Skorobogat’ko OV, Vartanov SS, Varfolomeyev SD. 1994. Laccase - Properties, catalytic mechanism, and applicability. Appl Biochem Biotechnol 49:257–280.
Yoshida H. 1883. Chemistry of Lacquer (Urushi) part I. Chemical Society of Tokio 43:472–486.
Yoshitake A, Katayama Y, Nakamura M, Iimura Y, Kawai S, Morohoshi N. 1993. N-linked carbohydrate chains protect laccase III from proteolysis in Coriolus versicolor. J Gen Microbiol 139:179–185.
Zhang J, Xu Z, Chen H, Zong Y. 2009. Removal of 2,4-dichlorophenol by chitosan-immobilized laccase from Coriolus versicolor. Biochem Eng J 45:54–59.
Zouari N, Romette J, Thomas D. 1987. Purification and properties of two laccase isoenzymes produced by Botrytis cinerea. Appl Biochem Biotechnol 15:213–225.
Cri
tical
Rev
iew
s in
Bio
tech
nolo
gy D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsite
De
Sher
broo
ke o
n 11
/13/
14Fo
r pe
rson
al u
se o
nly.
Copyright © 2022 FDOKUMEN




















