
Fungal laccase: properties and activity on lignin.
43
J. Basic Microbiol. 41 (2001) 3 – 4, 185 – 227 Review (Department of Biochemistry, Maria-Curie-Sklodowska University, Pl-20031 Lublin, Poland; 1 School of Forest Resources, Chungbuk National University, Cheongju 360-763, Korea and 'HSDUWPHQW RI $SSOLHG DQG (RORJLDO 0LURELRORJ\ )ULHGULK6KLOOHU8QLYHUVLW\ RI -HQD ' -HQD *HUPDQ\ Fungal laccase: properties and activity on lignin ANDRZEJ LEONOWICZ, NAM-SEOK CHO 1 , JOLANTA LUTEREK, ANNA WILKOLAZKA, MARIA WOJTAS-WASILEWSKA, ANNA MATUSZEWSKA, MARTIN HOFRICHTER 3 , DIRK WESENBERG 2 and JERZY ROGALSKI (Received 17 October 2000/Accepted 12 April 2001) The sources of ligninocellulose that occur in various forms in nature are so vast that they can only be compared to those of water. The results of several, more recent experiments showed that laccase probably possesses the big ability for “lignin-barrier” breakdown of ligninocellulose. The degradation of this compound is currently understood as an enzymatic process mediated by small molecules, therefore, this review will focus on the role of these mediators and radicals working in concert with enzymes. The fungi having a versatile machinery of enzymes are able to attack directly the “lignin- barrier” or can use a multienzyme system including “feed-back” type enzymes allowing for simulta- neous transformation of lignin and carbohydrate compounds. Fungal laccase (benzenediol:oxygen oxidoreductase, EC 1.10.3.2) is an enzyme secreted into the medium by mycelia of Basidiomycetes, Ascomycetes and Deuteromycetes (BOLLAG and LEONOWICZ 1984, AGEMATU et al. 1993). In some fungal strains, laccase can be in- duced by anilines (FAHRAEUS et al. 1958, BOLLAG and LEONOWICZ, 1984), methoxyphe- nolic acids (ROGALSKI et al. 1992, LEONOWICZ et al. 1997a) lignin preparations (ROGALSKI et al. 1991) or heat shock (FINK-BOOTS et al. 1997 and 1999). However, the most effective inducer of laccase in fungi is 2,5-xylidine. The stimulating effect of this compound was first described in 1958 by FAHRAEUS et al. for the white-rot basidiomycete Trametes (Coriolus) versicolor. The authors selected 2,5-xylidine among various aniline derivatives and found an over 160-fold stimulation of laccase activity when the compound was added to growing fungal cultures. In addition, it was shown that not only in Trametes versicolor (FAHRAEUS and REINHAMMAR 1967), but also in other lignin degrading fungi such as Fomes annosum, Pholiota mutabilis, Pleurotus ostreatus (AGEMATU et al. 1993) and Phlebia radiata (ROGALSKI and LEONOWICZ 1992), laccase levels are enhanced in the presence of 2,5- xylidine. Also certain phenolic compounds were found to stimulate laccase production, e.g., ferulic acid which considerably induces the enzyme in the cultures of Pholiota mutabilis, Pleurotus ostreatus and Trametes versicolor (LEONOWICZ et al. 1978). The highest amounts of laccase are produced by white-rot fungi (wood-decaying basidio- mycetes; LEONOWICZ et al. 1997a). Biochemically, laccase is an enzyme which oxidizes a variety of aromatic hydrogen donors. Thus, it catalyzes the removal of an electron and a proton from phenolic hydroxyl or aromatic amino groups to form free phenoxy radicals and amino radicals, respectively. Moreover, this group of copper-containing laccase having four copper atoms all in the 2+ oxidation state in the active site (i.e. blue oxidase) does not only oxidize phenolic and methoxyphenolic acids, but also decarboxylates them (AGEMATU et al. 1993) and attacks their methoxyl groups through demethylation (LEONOWICZ et al. 1984) or © WILEY-VCH Verlag Berlin GmbH, 13086 Berlin, 2001 0233-111X/01/3-407-0185 $ 17.50+.50/0
Transcript of Fungal laccase: properties and activity on lignin.

J. Basic Microbiol. 41 (2001) 3–4, 185–227
Review
(Department of Biochemistry, Maria-Curie-Sklodowska University, Pl-20031 Lublin, Poland; 1Schoolof Forest Resources, Chungbuk National University, Cheongju 360-763, Korea and ������������ ������� ��� ��������� ���������� ���������������������������� �� ����� �� !!"# ����$�����%
Fungal laccase: properties and activity on lignin
ANDRZEJ LEONOWICZ, NAM-SEOK CHO1, JOLANTA LUTEREK, ANNA WILKOLAZKA,MARIA WOJTAS-WASILEWSKA, ANNA MATUSZEWSKA, MARTIN HOFRICHTER3,DIRK WESENBERG2 and JERZY ROGALSKI
(Received 17 October 2000/Accepted 12 April 2001)
The sources of ligninocellulose that occur in various forms in nature are so vast that they can only becompared to those of water. The results of several, more recent experiments showed that laccaseprobably possesses the big ability for “lignin-barrier” breakdown of ligninocellulose. The degradationof this compound is currently understood as an enzymatic process mediated by small molecules,therefore, this review will focus on the role of these mediators and radicals working in concert withenzymes. The fungi having a versatile machinery of enzymes are able to attack directly the “lignin-barrier” or can use a multienzyme system including “feed-back” type enzymes allowing for simulta-neous transformation of lignin and carbohydrate compounds.
Fungal laccase (benzenediol:oxygen oxidoreductase, EC 1.10.3.2) is an enzyme secretedinto the medium by mycelia of Basidiomycetes, Ascomycetes and Deuteromycetes (BOLLAGand LEONOWICZ 1984, AGEMATU et al. 1993). In some fungal strains, laccase can be in-duced by anilines (FAHRAEUS et al. 1958, BOLLAG and LEONOWICZ, 1984), methoxyphe-nolic acids (ROGALSKI et al. 1992, LEONOWICZ et al. 1997a) lignin preparations (ROGALSKIet al. 1991) or heat shock (FINK-BOOTS et al. 1997 and 1999). However, the most effectiveinducer of laccase in fungi is 2,5-xylidine. The stimulating effect of this compound was firstdescribed in 1958 by FAHRAEUS et al. for the white-rot basidiomycete Trametes (Coriolus)versicolor. The authors selected 2,5-xylidine among various aniline derivatives and foundan over 160-fold stimulation of laccase activity when the compound was added to growingfungal cultures. In addition, it was shown that not only in Trametes versicolor (FAHRAEUSand REINHAMMAR 1967), but also in other lignin degrading fungi such as Fomes annosum,Pholiota mutabilis, Pleurotus ostreatus (AGEMATU et al. 1993) and Phlebia radiata(ROGALSKI and LEONOWICZ 1992), laccase levels are enhanced in the presence of 2,5-xylidine. Also certain phenolic compounds were found to stimulate laccase production, e.g.,ferulic acid which considerably induces the enzyme in the cultures of Pholiota mutabilis,Pleurotus ostreatus and Trametes versicolor (LEONOWICZ et al. 1978).
The highest amounts of laccase are produced by white-rot fungi (wood-decaying basidio-mycetes; LEONOWICZ et al. 1997a). Biochemically, laccase is an enzyme which oxidizes avariety of aromatic hydrogen donors. Thus, it catalyzes the removal of an electron and aproton from phenolic hydroxyl or aromatic amino groups to form free phenoxy radicals andamino radicals, respectively. Moreover, this group of copper-containing laccase having fourcopper atoms all in the 2+ oxidation state in the active site (i.e. blue oxidase) does not onlyoxidize phenolic and methoxyphenolic acids, but also decarboxylates them (AGEMATU et al.1993) and attacks their methoxyl groups through demethylation (LEONOWICZ et al. 1984) or
© WILEY-VCH Verlag Berlin GmbH, 13086 Berlin, 2001 0233-111X/01/3-407-0185 $ 17.50+.50/0

186 A. LEONOWICZ et al.
demethoxylation (POTTHAST et al. 1995). These reactions may represent an important stepin the initial transformation of the lignin polymer (LEONOWICZ et al. 1984, AGEMATU et al.1993, POTTHAST 1995). It has also been reported that the enzyme is involved in the trans-formation and detoxification of reactive lignin degradation products (BOLLAG et al. 1988).Laccase reacts with polyphenols and other lignin-derived aromatic compounds, which, inturn, can be both polymerized and depolymerized or act as low-molecular weight mediators(BOURBONNAIS et al. 1995).&��������������������������'������ ������(������ ����� �� �������)���������������
�*(�(���� ����� ������ ������������������������������� +LUTEREK ������ ,--.%/ ���'������ ��������������(�����������������������(��������������(�����+BOURBONNAIS ��PAICE ,--0�� BOURBONNAIS ������ ,--1%�� ��� �� �������)��� �(���� �(��� ��� 2��� ��������+&���������� ,--�%������ � ����������� �����+CHO ������ ,---�%/� ��� ��������������������2��� ������� �� ������ ��� �������� ���������� ����(������ ��� ��� ���'�������� �������(����� � ����� +BOLLAG ������ ,-..%� �� ��� ���� ������� �� ������� ��� ����������� ���(�� ��� ��(���������(�������2����2������+NANNIPIERI ���BOLLAG ,--,%/3���'��������������� �������� ���'� �������� �� *(���� ��������������2���� ����(���*(�������������� ��������� �� ���(������������+SIMMONS ������ ,-.-� LIU ������,-.1%/� ���� ����������� �(��� �������� ���� �� ������� ������� ��� 2����/4������(����� �� ���� ����� � ����� �� �� '���)�� �� ���� �'�� �������� ��� ��������������)��� � ������ ���� �������� +GIANFREDA �� BOLLAG ,--"%/ ���� ����(��(���*(�� ������������� ��������������������� ��� ������������ � ��� �����(�(�+GIANFREDA ���BOLLAG ,-."%/
Laccase is also involved in various physiological processes of economic and ecologicalrelevance. It is engaged in the development of fungal fruit bodies (LEATHAM andSTAHMANN 1981), pigmentation of fungal spores (CLUTTERBUCK 1972), pathogenicity ofmoulds (RIGLING and VAN ALFEN 1991), sexual differentiation (AISEMBERG et al. 1989),and rhizomorph formation (WORALL et al. 1986).
The biodegradation of the lignocellulose constituents, cellulose, hemicelluloses and lig-nin, is achieved by enzymatic activities. The conversion of cellulose and hemicellulosesinto simple sugars has been studied for a long time. A large number of micro-organisms(bacteria, fungi, protozoans) make use of a whole string of hydrolases which are able toproduce large quantities of mono- and disaccharides from all polysaccharide components inligninocellulose. The degradation, however, is effected by the occurrence of polysaccha-rides in a complex with lignin because the latter forms a barrier against the microbial attackby hydrolytic enzymes. The lignin barrier also complicates cellulose production in the pulpand paper industry. For these ecological, economic and other reasons, research into thebiotransformation of lignin has been carried out for decades.
Wood-rotting basidiomycetes penetrate wood tissues in order to come into contact withthe easily assimilable carbohydrate constituents of the lignocellulosic complex. The white-rot group of these fungi, which has a versatile machinery of enzymes co-operating withcertain secondary metabolites of fungi, is capable of attacking the lignin barrier efficiently.These fungi use a multi-enzyme system including the so-called “feed-back” type enzymesto transform and degrade all structural elements of the lignocellulosic complex (polysaccha-rides and lignin). The currently known enzymes of white-rot fungi involved in wood degra-dation can be divided into three groups. The first can attack the wood constituents or theirprimary degradation products directly; this group includes the cellulase and hemicellulasecomplexes, laccase, different peroxidases, protocatechuate-3,4 dioxygenase, etc. The sec-ond group of enzymes, comprising among others aryl alcohol oxidase and glyoxal oxidase,co-operates with the first group by providing H2O2 for the peroxidases, but these enzymesdo not attack wood components directly. The third enzyme group represented by glucoseoxidase and cellobiose:quinone oxidoreductase (cellobiose dehydrogenase) includes feed-back type enzymes which play a key role in joining the metabolic chains during the bio-

Fungal laccase: properties and activity on lignin 187
transformation of high-molecular mass wood constituents. All these enzymes, includinglaccase, can act separately or in co-operation.
Among the white-rot fungi, the polyporous fungus Trametes versicolor and the corticoidfungus Phanerochaete chrysosporium have been particularly studied in detail due to theirhigh ligninolytic activities. This makes them promising candidates for applications in thepulp and paper industry (e.g., bleaching, pulping, waste water treatment; MANZANARESet al. 1995, PASZCZYNSKI and CRAWFORD 1995). The production of high amounts of lac-case by Trametes versicolor has already been known for decades (FAHRAEUS et al. 1958)while the lignin modifying activity of this enzyme was detected relatively early(LEONOWICZ and TROJANOWSKI 1965). The situation is different in the case of Phanero-chaete chrysosporium, the most-investigated white-rot fungus, since laccase was discoveredin this fungus relatively late and is only produced in certain strains and under special condi-tions (SRINIVASAN et al. 1995). Therefore, ligninolytic activities of this Phanerochaetechrysosporium have mainly been attributed to lignin peroxidase and manganese peroxidase(KUHAD et al. 1997); its laccase has received more attention only recently (DITTMER et al.1997, RODRIGUEZ et al. 1997). Consequently, it has been suggested that two classes ofextracellular oxidative enzymes – peroxidases and laccases – are involved in ligninolysisowing to their ability to catalyze the cleavage of carbon-carbon or carbon-oxygen bonds incomplex lignin polymer or lignin model compounds (KIRK and HAMMEL 1992, HAMMELet al. 1993, MARZULLO et al. 1996).
Both extracellular peroxidases and laccases (also called ligninolytic or lignin-modifyingenzymes; HATAKKA 1994) have the ability to catalyze one-electron oxidations resulting inthe formation of radicals which undergo numerous spontaneous reactions. These, in turn,lead to various bond cleavages including aromatic ring fission (SHOEMAKER and LEISOLA1990, TUOR et al. 1992, AKTHAR et al. 1997, ZAPANTA and TIEN 1997). Manganese peroxi-dase attacks aromatic lignin structures indirectly via Mn3+-chelates acting as a low-molecular mass redox mediator while lignin peroxidase oxidizes aromatic lignin moietiesdirectly (WARIISHI et al. 1991, KUAN and TIEN 1993, ZAPANTA and TIEN 1997). Laccaseoxidizes preferentially phenolic lignin structures to phenoxy radicals which subsequentlyform quinones. This spontaneous rearrangement can also lead to the fission of carbon-carbon or carbon-oxygen bonds inside the lignin phenyl-propane subunits resulting either inthe degradation of both side chains (FUKUZUMI 1960) and aromatic rings (KAWAI et al.1988b), or in demethylation processes (HARKIN and OBST 1974, ANDER et al. 1983,LEONOWICZ et al. 1984, 1991a). In this respect, laccase can co-operate with various FAD-containing oxidases like glucose oxidase (SZKLARZ and LEONOWICZ 1986), veratryl alcoholoxidase (MARZULLO et al. 1995), cellobiose:quinone oxidoreductase (WESTERMARK andERIKSSON 1975), and cellobiose dehydrogenase (AYERS et al. 1978).
Although the actual role of laccase in lignin biodegradation is still under discussion andnot completely understood, several authors have reported that laccase acts on the ligninpolymer in the ways different from ligninolytic peroxidases. In order to present the role oflaccase more clearly several conceptions of biochemical routes in lignin biodegradationhave been reviewed, especially with respect to the co-operation of laccase with other bio-catalysts.
Hististorical outlook
The first attempts to delignify wood by means of white-rot fungi were made already in theearly 1930s. According to CAMPBELL (1930) and, independently, to WIERTELAK (1932),Trametes versicolor and Trametes pini respectively were shown to transform wood effi-ciently. Several years later, a more detailed analysis of the delignification process of beechsawdust by the white-rot fungi Polyporus abietinus, Stereum rugosum, and Marasmiusscorodonius was carried out (FAHRAEUS et al. 1949). However, due to the poor analyticalmethods at the time the analysis was extremely difficult. It was only the development of

188 A. LEONOWICZ et al.
OCH3OCH3
OCH3OCH3
CH2CH2 COOH
H CO3H CO3
OCH3OCH3
OCH3OCH3
OCH3OCH3 OCH3OCH3
OCH3OCH3
OCH3OCH3
OCH3OCH3
OH
OH
OH
OH
OH
OH OH
R=
Lignin ( I )Lignin ( I )
Laccase( III )
Laccase( III )
( IV )( IV ) ( V )( V )
Decayed lignin ( II )Decayed lignin ( II )
ConiferylalkoholConiferylalkohol( IIIIII )
R' =
R'
HemicelluloseGuaiacylpropanolsHemicelluloseGuaiacylpropanols
Guaiacylpropanones( Ketone compounds )Guaiac lpropanones( Ketone compounds )
Guaiacylpropanones
vanilic acid like
compound
Guaiacylpropanones
vanilic acid like
compound
go to the pathway of homogentisic
acid oxidation
go to the pathway of homogentisic
acid oxidation
HC
HC
HC
CHCH
COCO COCO
HC
HCOH
OH
HCHC
CHCH
CH2CH2 CH2CH2
O O
O
O
OC
C
CC
C
CCH OH2CH OH2
CH OH2CH OH2
CH OH2CH OH2R R
H COH2H COH2 H HH H
H COH2H COH2 H H
H COH2H COH2
H COH2H COH2
H COH2H COH2 COOHCOOH
.
Fig. 1Hypothetical scheme of lignin degradation by Poria subacida (adapted from FUKUZUMI 1960)
novel chromatographic methods that helped to explain some stages of lignin biotransforma-tion by the characterization of certain degradation products. Thus, in the 1960s, the firstschemes of lignin degradation by white-rot fungi were published based on the data fromfungal decay of wood mill. Among others, a hypothetical mechanism was suggested byFUKUZUMI (1960), where laccase was presented as acting as a lignin-attacking enzyme(Fig. 1).
In this scheme, however, laccase appeared not at the beginning of but in the latter part ofthe degradation process. That is why the questions arose of how the lignin barrier could beinitially attacked and what the responsible attacking agent was. One of our first reportscovered the earliest attempt at explaining the initial microbial attack on lignin and postu-lated that it could indeed be catalyzed by fungal laccase (and also peroxidase) through thepreliminary demethylation of lignin. In addition, it was suggested that following the de-methylation, the break-down of ether bonds connecting lignin subunits probably occursspontaneously (non-enzymatically) and takes place as a result of the electronophilic effectof the ortho-quinones on the ether bonds between phenylpropane subunits.
The first indication of the reliability of this conception was to be found in the work ofHARKIN and OBST (1974). They showed that laccase and/or peroxidase generate a numberof reactive phenoxy radicals during the process of 2,4,6-trimethoxyphenol oxidation.

Fungal laccase: properties and activity on lignin 189
OMe OMe
O
HO
+
+ +
OMeOMe
OMeOMeOMeOMe
OMeOMeOMe
O OMe
-MeOH -MeOH
-H.
-H.
H O2
H O2
O
O O
O
O O
OMeOMe O
O
OMe
OMe
OMeOMe
OMeOMe
OMeOMe
OMeOMe OH
OMe
OMeOMe
OMeOMe
OMeOMe O
MeO MeOMeO
MeO
MeO MeOMeO
MeO MeOMeO
O
MeO MeOMeO
II+IV II+III
V VI
IX X
VII VIIIVIII
XI XII
Mesomeric forms of these phenoxy radicals couple with para or orto-quinal dimers whichsubsequently undergo demethylation and decomposition to monomers and are extremelyunstable, which triggers the polymerization effect (Fig. 2).
The same report also demonstrated the ability of laccase (and peroxidase) to cleave etherbonds linking most aromatic subunits in lignin, which pointed to a general significance ofdemethylation and quinone formation in lignin degradation. Subsequent research confirmedthe demethylating activity of laccase and showed the importance of pH on the conversionprocess (LEONOWICZ et al. 1984), e.g., by the use of syringic acid (ISHIHARA and ISHIHARA1976), vanillyl glycol (LUNDQUIST and KRISTERSSON 1985), and vanillic acid (ANDER et al.1983, ANDER and ERIKSSON 1985). The latter authors proposed a possible pathway of va-nillic acid metabolism via demethylation up to the cleavage of the aromatic ring and theformation of ketoacids (ANDER et al. 1985). Later, such an aromatic ring fission was, infact, demonstrated for Trametes (Coriolus) versicolor laccase (KAWAI et al. 1988b) (Fig. 3).The substrate (I) is oxidized by laccase to form phenoxy radicals, which are subsequentlyattached by molecular oxygen. The resulting hydroperoxide reacts with nucleophilic oxygenspecies forming substance (II) via incorporation with H2O (pathway B) or by forming acyclic peroxide (pathway A) which is then converted to a muconate derivative. Subse-quently, the muconate compound undergoes cyclization to yield lactone (II).
Fig. 2Transformation of dimers formed from 2,4,6-trimethoxyphenol during the action of laccase. Decom-position of dimers is accompanied by oxidation anddemethylation processes (adapted from HARKIN andOBST 1974)

190 A. LEONOWICZ et al.
OCH3
O
O O
O
O OH
O2
O2H O2
pathwaypathway A
pathwaypathway B
OOH
OCH3 OCH3
OCH3
OCH3OCH3
OCH3 OCH3
OH
O O
OH
O
O
H
HO
OH
OH2
O HOO
O
O O
laccase
( I ) ( II )
..
.
. ..
.
.
.
Fig. 3Mechanism of mucolactone (II) formation through the aromatic ring cleavage of the lignin modelcompound 4,6-di(t-butyl)guaiacol (I) by laccase of Trametes (Coriolus) versicolor (adapted fromKAWAI et al. 1988b)
Furthermore, it was shown that a phenol oxidase-less (= laccase-less) mutant of Sporo-trichum pulverulentum obtained by UV-mutagenesis did not transform lignin while the wildtype decomposed the polymer effectively (ANDER and ERIKSSON 1976). This findingstrongly pointed to laccase as an important factor in lignin transformation and was laterconfirmed by the use of the regenerated protoplast technique to gain laccase-less mutants(EGUCHI et al. 1994).
OCH3OCH3
OCH3OCH3
OCH3OCH3 OCH3OCH3
OCH3OCH3
OCH3OCH3 OCH3OCH3
OCH3OCH3 OCH3OCH3
OCH3OCH3
OH
OH
OHOH OHOH
HC
HC HCHC
HCHC HCHC
HCHC COCO
COCO
HCOH
CO COOHCOOH
HCOHCO HCOHCO
HCOHHCOH CH2CH2
COOHCOOH
O
O O
O O
OC
C C
C C
C
LaccaseLaccase
LaccaseLaccase AcidolysisAcidolysis
LaccaseLaccase LaccaseLaccase
LaccaseLaccase
Laccase
Laccase
AcidolysisAcidolysis
C
C C
C C
CC
C C
C C
C
H COH2H COH2
H COH2H COH2 H COH2H COH2
H COH2H COH2 H COH2H COH2
COOHCOOH COOHCOOH
CH3CH3
.
Fig. 4A possible pathway for the formation of carboxyl groups in lignin by Trametes (Coriolus) versicolorlaccase (adapted from KONISHI et al. 1974)

Fungal laccase: properties and activity on lignin 191
Significant contributions of laccase to lignin transformation were also reported by manyother authors: ISHIHARA and MIYAZAKI (1972), KONISHI et al. (1974; Fig. 4), ANDER andERIKSSON (1976), LEONOWICZ et al. (1985), KAWAI et al. (1986), SZKLARZ and LEONOWICZ(1986), KAWAI et al. (1988a; Fig. 5). KIRK et al. (1968) proposed a pathway leading toformation of 2,6-Dimethoxy-p-benzoquinone which was indeed found four years later byISHIHARA and MIYAZAKI (1972) as the result of the action of Trametes versicolor laccase onBJÖRKMAN’s lignin. In addition, we have shown in our laboratory that Trametes versicolorlaccase both depolymerizes and polymerizes polymeric lignosulphonates (adequate modelsof the macromolecular lignin structure), which, as soluble in water, are readily available forenzymes (LEONOWICZ et al. 1985; Fig. 6). In similar experiments, laccase also depolymer-ized – although not to a high degree – BJÖRKMAN’s lignin (LEONOWICZ et al. 1997b). Incontrast, GALLIANO et al. (1991), who used 14C-labelled KLASON lignin as the substrate forlaccase from Rigidosporus lignosus, found only slight solubilization of the polymer, proba-bly due to predominant repolymerization reactions.
KONISHI et al. (1974) put forward an interesting hypothesis that the formation of carboxylgroups in the side chains of lignin by laccase “solubilizes” the polymer. The dissolved car-boxylated products were proposed to adhere to the fungal cell-walls, where their conversionand subsequent uptake into the cells might be facilitated. However, this hypothesis – attrac-tive from the point of view of lignin utilization – could not be confirmed in subsequentstudies.
The above route demonstrates that the disproportion of the phenoxy radical of substrate I,formed by the action of laccase, occurs via a cation intermediate, which is further trans-formed to a diphenyl propanone III (pathway A) and to a hydroquinone/quinone (pathway
OCH3OCH3
OCH3OCH3
OCH3OCH3 OCH3OCH3 OH
OCH3OCH3
OCH3OCH3
OCH3OCH3
OCH3OCH3 OCH3OCH3
OCH3OCH3
OCH3OCH3OCH3OCH3OCH3OCH3 CH O3CH O3CH O3CH O3
OCH3OCH3
OCH3OCH3
OCH3OCH3
OCH3OCH3
OCH3OCH3 OCH3OCH3
OCH3OCH3
OCH3OCH3OHOHO
OCH3OCH3
HO
HO
O O
H
O
HO
HO
HO HOHOHO
HOHO
H OOHO
( VI )( VI )( V( VII )( VIII )( VIII )
HOHO
HOOC H2 5OC H2 5
OC H2 5OC H2 5
OC H2 5OC H2 5 OC H2 5OC H2 5
OC H2 5OC H2 5
OC H2 5OC H2 5
OC H2 5OC H2 5
OH
O
O OHO H
H
OHCH O3CH O3
CH O3CH O3
CH O3CH O3 CH O3CH O3
CH O3CH O3
CH O3CH O3
LaccaseLaccase
A B
phenoxy radicalphenoxy radical
disproportionationdisproportionation
( I )
( IIIIII )
( I )OH2OH2
H
.
.
.
.
+
.
Fig. 5Possible mechanism for Ca oxidation (A) and alkyl-aryl cleavage (B) of the lignin model dimer1-(3,5-dimethoxy-4-hydroxyhenyl)-2-(3,5-dimethoxy-4-ethoxyphenyl) propane-1,3-diol (I) by laccaseof Trametes (Coriolus) versicolor (adapted from KAWAI et al. 1988a)

192 A. LEONOWICZ et al.
0
0.8
0.40.5
1.0
1.5
2.0
2.5
3.0B
1.0
2.0
3.0
40 400
A
1
2
3
4
5
80 8060120 100 120160 140 160
Fraction number Fraction number
Ab
so
rb
an
ce
at
28
0n
m
Ab
so
rb
an
ce
at
28
0n
m
Fig. 6Depolymerization (A) and polymerization (B) of Na-lignosuphonate fraction with approximate mo-lecular weights of 97 (A) and 1 (B) kDa. Continuous line: elution profile on the Sephadex G-50 col-umn after treatment with laccase. Dashed line: elution profile of the control (adapted from LEONO-WICZ et al. 1985)
B). It was postulated that laccase is only responsible for the formation of the phenoxy radi-cal whereas both C� and alkyl-aryl cleavages are caused by spontaneous reactions. Thissupports our original hypothesis of the lignin degradation.
Last but not least, it should be mentioned that the so-called “yellow laccases” have beenrecently discovered in straw cultures of Panus tigrinus and Phlebia radiata. These enzymesseem to have a changed oxidation state of copper in the active center, probably caused bythe integration of aromatic lignin degradation products (LEONTIEVSKY et al. 1997a). Inter-estingly, evidence has been provided that yellow laccases are capable of oxidizing non-phenolic lignin models (�-O-4) and veratryl alcohol directly in the presence of O2, but inthe absence of any diffusible mediator (LEONTIEVSKY et al. 1997b).
Enzymes cooperating with laccase in ligninolysis
Extracellular phenoloxidizing activity which indicates the presence of ligninolytic enzymeswas discovered in white-rot fungi already in the 1930s (ERIKSSON et al. 1990). With regardto their typical production patterns of extracellular ligninolytic enzymes, white-rot fungimay be divided into three main groups; (i) LiP-MnP group, (ii) MnP-laccase group, and (iii)LiP-laccase group (HATAKKA 1994). However, several enzymes possessing other than di-rectly lignin-degrading activities, such as H2O2 production, are indispensable in ligninolysisand therefore also reviewed here.
Lignin peroxidase (LIP; EC 1.11.1.14): Lignin peroxidase (at first designated as “ligni-nase”) was discovered in cultures of Phanerochaete chrysosporium after many years ofresearch. The glycosylated enzyme was identified and described independently by twoAmerican teams (GLENN et al. 1983, TIEN and KIRK 1983) and a Japanese group (SHIMADAand HIGUCHI 1983) in supernatants of nitrogen and carbon-limited cultures of the corticoidbasidiomycete Phanerochaete chrysosporium that were flushed with pure oxygen. Later,the enzyme was also found in other wood colonizing, corticoid and polyporous basidiomy-

Fungal laccase: properties and activity on lignin 193
cetes, e.g., Phlebia radiata, Phlebia tremellosa, Trametes versicolor (HATAKKA et al. 1987,LEONOWICZ et al. 1991b, HATAKKA 1994). The only currently known agaric fungi produc-ing lignin peroxidase are Nematoloma frowardii and Clitocybula dusenii (HOFRICHTER andFRITSCHE 1997).
Lignin peroxidase (molecular mass varies between 38 and 47 kDa) necessarily requireshydrogen peroxide (H2O2) generated by other enzymes (e.g., oxidases) to be active. Theenzyme comprises hem in the active site while the catalytic cycle resembles that of horse-radish peroxidase (TIEN et al. 1986). Lignin peroxidase catalyses several oxidations in thealkyl side chains of lignin-related compounds, C-C cleavages in the side chains of ligninsubunits, oxidation of veratryl alcohols and related substances to aldehydes or ketones,intradiol cleavage of phenylglycol structures, and hydroxylation of benzylic methylenegroups (TIEN and KIRK 1983). In Phanerochaete chrysosporium, the enzyme is produced inat least six isozymes (TIEN and KIRK 1988), which are all capable of oxidizing recalcitrantnon-phenolic lignin model substrates (redox potential around 1.5 V) by one-electron ab-straction to form reactive aryl cation radicals, which commonly decay via pathways in-volving C–C and C–O bond-cleaving reactions (KERSTEN et al. 1985).
Reactions of lignin peroxidase have been thought to be mediated by veratryl alcohol, alow-molecular-mass fungal metabolite which is oxidized by the enzyme to veratraldehydethrough an aryl cation radical (KIRK and FARRELL 1987). The latter is highly reactive andshould, in turn, oxidize the lignin polymer. However, this role of veratryl alcohol has beenquestioned since the stability of the radical (its life span) is too short to enable long-distancecharge-transfer. If veratryl alcohol radicals possess mediating properties, this will only berelevant in short distance transfers when the aryl cation radical is somehow complexed tothe enzyme (LUNDELL 1993). Nevertheless, veratryl alcohol stimulates lignin peroxidaseactivity, probably by protecting the sensitive enzyme from the damaging effects of excessH2O2 or phenolics (AKTHAR et al. 1997).
Lignin peroxidase is also capable of cleaving aromatic rings via initial one-electron ab-stractions and subsequent incorporation of oxygen (UMEZAWA and HIGUCHI 1987). Theoptimum activity of lignin peroxidase is extremely low (between pH 2.5 and 3.0; TIEN andKIRK 1988) while the enzyme itself is acidic (pIs 3-5). TIEN and KIRK (1983) have reportedthat in a cell-free system (in vitro) the enzyme is particularly active toward highly-methylated lignins (e.g., lignin isolated from birch wood and additionally methylated withmethyl iodide), which are depolymerized into lower-molecular mass fragments. In contrast,natural lignins containing phenolic rings tend to re-polymerize in vitro in the presence oflignin peroxidase (SARKANEN et al. 1991).
Two years later, it was shown (using 14C-labelled synthetic lignin preparations) that puri-fied Phanerochaete chrysosporium lignin peroxidase is also capable of depolymerizinglignin, which contains phenolic rings, if specific reactions conditions are created: the addi-tion of only low amounts of lignin dissolved in methoxyethanol (around 5 mg/l) and thepresence of veratryl alcohol. Under these conditions, lignin peroxidase is capable of oxi-
Fig. 7Lignin peroxidase catalyzed Ca-Cb cleavage in 14C-labelled synthetic lignin to give a benzoic alde-hyde at the Ca-position (adapted from HAMMEL et al. 1993)

194 A. LEONOWICZ et al.
dizing the lignin polymer directly through cleaving Ca–b bonds yielding benzoic aldehydederivatives of b-O-4 substructures (HAMMEL et al. 1993; Fig. 7).
Since lignin, degraded by lignin peroxidase, provides a substrates for laccase, both en-zymes can be considered as “partners” in certain biotransformation routes of lignin.
Manganese peroxidase (MnP; EC 1.11.1.13): Manganese peroxidase was also discoveredin cultures of Phanerochaete chrysosporium (KUWAHARA et al. 1984). The enzyme resem-bles lignin peroxidase: it is extracellular, glycosylated and contains hem as the prostheticgroup (GLENN and GOLD 1985, PASZCZYNSKI et al. 1986). Manganese peroxidase is alsoexpressed in multiple forms with molecular weights from 40 to 48 kDa and pIs between 2.9and 7.0. Besides Phanerochaete chrysosporium, manganese peroxidase has also been foundin many other white-rot and soil litter-decomposing basidiomycetes, but not in any otherfungi or bacteria (e.g., Trametes versicolor, JOHANSSON and NYMAN 1987; Phlebia radiata,NIKU-PAAVOLA et al. 1988; Ceriporiopsis subvermispora, RÜTTIMANN et al. 1992; Agari-cus bisporus, BONNEN et al. 1994; Nematoloma frowardii, SCHNEEGASS et al. 1997; Stro-pharia rugosoannulata, SCHEIBNER and HOFRICHTER 1998).
The catalytic cycle of manganese peroxidase resembles that of lignin peroxidase, includ-ing native ferric enzyme as well as peroxidase Compound I and Compound II redox states(WARIISHI et al. 1988). However, significant differences appear in the reductive reactionswhere Mn2+ is a required electron donor. Both Compound I and Compound II are reducedby Mn2+ while the latter is oxidized to Mn3+. Mn3+ ions are stabilized to high redox poten-tials via chelation with organic acids such as oxalate, malonate, malate, tartrate, or lactate(WARIISHI et al. 1992, Fig. 8). Chelated Mn3+, in turn, acts as diffusible redox mediator thatoxidizes phenolic lignin structures (WARIISHI et al. 1992).
The oxidative strength of the manganese peroxidase system can be considerably en-hanced in the presence of appropriate additional cooxidants such as thiols (FORRESTER et al.1988, HOFRICHTER et al. 1998c) or lipids (BAO et al. 1994), which results in the cleavedchemical bonds. These bonds are not normally open to attack by manganese peroxidase(e.g., recalcitrant non-phenolic �-O-4 arylethers). Moreover, manganese peroxidase isunique in its ability to degrade aromatic substances, including lignin, directly to carbondioxide (CO2); this process (enzymatic combustion), which is considerably stimulated in thepresence of the above mentioned cooxidants, probably allows ligninolytic fungi to miner-alize lignin - at least in part - outside the cell (HOFRICHTER et al. 1998a, KAPICH et al.1999). The enzyme has been successfully applied to depolymerize 14C-labelled syntheticand natural lignins in vitro (WARIISHI et al. 1991); as a result, low-molecular mass, water-soluble lignin fragments and significant amounts of CO2 have formed (HOFRICHTER et al.1998a, 1998b, 1998c, 1998d, 1998e).
H O2 2
H O2 2
MnP MnP IMnP I
MnP II MnP IIMnP II
k3
k3
k2
k2
Kd
Kd
k1
k1
OMn3 +OMn3 +
OMn3 +OMn3 +
OMn2 +OMn2 +
OMn2 +OMn2 +
OMn2 +OMn2 +
OMn2 +OMn2 +
Fig. 8Mechanism of manganese peroxidasecatalyzed formation of Mn3+-oxalatecomplexes. Mn-oxalate complexes arerepresented by OMn2+ and OMn3+
(adapted from ZAPANTA and TIEN 1997)

Fungal laccase: properties and activity on lignin 195
Manganese peroxidase has been shown to generate H2O2 in the oxidation of certain thiols(e.g., glutathione) and NAD(P)H2; recently, evidence has been provided that the enzyme iseven capable of acting efficiently in the absence of external H2O2 by oxidizing organic acids(e.g., oxalate, malonate, malate) in “oxidase-like,” autocatalytic reactions involving thetransient formation of several radical species (HOFRICHTER et al. 1998e, URZÚA et al.1998).������ ����������������� ���'��������� ��� ���������� ���������� �����������
2����������� �������������5���(������������� � ���������)������,"6��������� �����2��������� ��� ������ ��� 2�� ���� �)����� 2���� ������ �� ���� ������ ��'�(��+GALLIANO ������ ,--,%/� 7�� ��� ���� ���(��� ����� ���� �������� ����� �� ����������'���������������� �����������������������������������8������/
In recent years, an extended role of manganese peroxidase in the lignin degradation hasbeen proposed following the discovery of novel hybrid forms of the enzyme and other per-oxidases in some fungi, e.g., in Pleurotus eryngii and Bjerkandera spp. (MARTINEZ et al.1996, MESTER and FIELD 1998). These enzymes are able to oxidize both Mn2+ and aromaticsubstrates (e.g., veratryl alcohol, various phenolics), thus combining the properties of man-ganese peroxidase and lignin peroxidase.
Other peroxidases (EC 1.11.1.7): A “classic” extracellular peroxidase (similar to horse-radish peroxidase) was identified for the first time in a growing culture of the agaric white-rot fungus Pholiota mutabilis by LEONOWICZ and TROJANOWSKI (1965). Together withlaccase, this peroxidase caused the demethylation of vanillic acid and BJORKMAN's lignin.The enzyme was later purified from culture liquids of two other white-rot fungi, Inonotusradiatus and Trametes versicolor, by LOBARZEWSKI (1981). Its constitution (e.g., molecularweight, hem in the catalytic center, H2O2 requirement) and other properties were similar tothose of plant horseradish peroxidase. Interestingly, the enzyme did not show any effect onveratryl alcohol. Apart from its existing repolymerizing activities, this type of peroxidasefrom Trametes versicolor depolymerized high-molecular weight lignin (water-soluble Na-lignosulphonates) at pH 3 but not to a large extent (Fig. 9).
0.5
1.0
200 200 200600 600 600 ml
A B C
Ab
so
rb
an
ce
at
28
0n
m
Fig. 9Effects of immobilized Trametes versicolor peroxidase on Na-lignosulphonates. The continuous lineindicates the elution profile of lignosulphonates after treatment with peroxidase (A, B and C refer todifferent incubation periods; 3, 12 and 24 hrs respectively). The dashed line indicates the elutionprofile of the control. Radicals and quinones generated by peroxidase (as in the case of laccase) mayreadily undergo polymerization and thus, the depolymerization effect is less clear (adapted fromLOBARZEWSKI et al. 1982)

196 A. LEONOWICZ et al.
A similar type of peroxidase was discovered by de JONG et al. (1992) in Bjerkandera sp.cultured in a nutrient- and nitrogen-rich glucose/yeast extract/peptone medium. Accordingto MESTER et al. (1996), this enzyme – described as manganese independent peroxidase –was inhibited by Mn2+ ions. The enzyme did not oxidize veratryl alcohol at all; however, itoxidized various phenolic substrates, e.g., 2,6-dimethoxyphenol that was also used for theenzyme assay. The essential cosubstrate of this enzyme was hydrogen peroxide and thecatalytic center was found to contain hem as the prosthetic group (KOTTERMAN et al. 1994).Extracellular peroxidases with properties similar to those of Trametes versicolor and Bjer-kandera sp. were also found in a range of other basidiomycetous fungi, e.g., Coprinus spp.(MORITA et al. 1988), Junghuhnia separabilima (VARES et al. 1992) and Panaeolus spp.(HEINZKILL et al. 1998).
�������������� �� ��������������� ����������Dioxygenases, widely distributed amongbacteria and microfungi, have also been identified in ligninolytic basidiomycetes.Protocatechuate 3,4-dioxygenase activity was first observed in Trametes (Polystictus)versicolor by HAIDER et al. (1962) and may serve as an example. In order to examine thecharacteristic steps of dioxygenase catalyzed reactions, radioactively-labelled 14C-vanillicacid was used as test substrate.
Before the discovery of ligninolytic peroxidases, scientists of KIRK’s team (CHEN et al.1983) formulated the hypothesis that among many other reactions occurring during thedecay of lignin by white-rotters, dioxygenase-catalyzed reactions inside the intact polymercould take place. As a result, the peripheric aromatic rings of lignin could be cleaved andaliphatic carboxylic acids formed as the products (Fig. 10).
Protocatechuate 3,4-dioxygenase was also later isolated from the cellulolytic ascomyceteChaetomium piluliferum and the ligninolytic basidiomycete Pleurotus ostreatus, and puri-fied to homogeneity in our laboratory (WOJTAS-WASILEWSKA and TROJANOWSKI 1980,WOJTAS-WASILEWSKA et al. 1983). The effects of this enzyme, including its immobilized
C C
C
C
C
OCH3
OH
COOHCOOH
COOHCOOH
COOHCOOH
OH
CHOCHO
COOHCOOH
COOHCOOH
OCH3
OCH3
OCH3
OCH3
OCH3
O O
O
O
O
O
29
13
O
OH
O
O
CH C
C
C
C
CHOH C
C
C
C
OH
OH
OH
OH
H O2
CH OH2
CH2
COOHCOOH
CHOCHO
CHOCHO
CHOCHO
CHOCHO
HO
L L
L
L
L
Fig. 10Cleavage of a peripheral ring of lignin by Phanerochaete chrysosporium growing on spruce woodchips (adapted from CHEN et al. 1983)

Fungal laccase: properties and activity on lignin 197
form on sodium lignosulphonates, were examined (WOJTAS-WASILEWSKA and LUTEREK1987, WOJTAS-WASILEWSKA et al. 1987). As the result, a product was obtained whichshowed a significant decrease in the absorbency around 280 nm indicating a loss of aro-matic rings. This may indicate that some peripheric aromatic rings of the tested lignosul-phonates were cleaved by 3,4-protocatechuate dioxygenase.
Other intracellular, ring-cleaving dioxygenases (1,2,4-trihydroxybenzene-1,2 dioxy-genase, catechol-1,2 dioxygenase) were detected in Phanerochaete chrysosporium andPhlebia radiata (RIEBLE et al. 1994, ROGALSKI et al. 1996).
Glucose 1-oxidase (GOD; EC 1.1.3.4): Glucose oxidase is a FAD-dependent oxidase thatcan be found inside and outside fungal cells. During the enzymatic catalysis, glucose – forexample, one originating from cellulose – is oxidized first to the corresponding glucono-lactone with the simultaneous reduction of FAD to FADH2. In the second step, FADH2 isoxidized back to FAD by dioxygen (O2) while hydrogen peroxide is formed (Alberti andKlibanov 1982).
�-D-Glucose + E-FAD � D-Glucono-1,5-lactone + E-FADH2 (1)
E-FADH2 + O2 � E-FAD + H2O2 (2)
Intensive screening studies in our laboratory have shown that glucose oxidase occurs inmany white-rot fungi which attack lignin effectively and secrete high amounts of laccase(LEONOWICZ et al. 1986, 1999d). RAMASAMY et al. (1985) have reported that glucose oxi-dase negative mutants of Phanerochaete chrysosporium exhibited little or no ability todegrade lignin whereas the wild type was able to do so. The same group has, for the firsttime, demonstrated that Phanerochaete chrysosporium also produces laccase. The follow-ing part of this review shows that glucose oxidase co-operates effectively with laccase inlignin degradation; thus, the results of RAMASAMY et al. alone may point to glucose oxidaseas an agent involved in lignin degradation (SRINIVASAN et al. 1995).
Another related sugar oxidizing, intracellular enzyme with FAD as the prosthetic group –pyranose 2-oxidase (EC 1.1.3.10) – was found in the growing mycelium of Phanerochaetechrysosporium. The ultrastructural and immunocytochemical characterization of this en-zyme was reported by DANIEL et al. (1992).
Cellobiose : quinone oxidoreductase (CBQ; EC 1.1.5.1): In 1974, when Trametes (Poly-porus) versicolor was cultured on lignin agar plates supplemented with glucose, cellobioseor cellulose, WESTERMARK and ERIKSSON (1974a) found indications of the existence of anew quinone-reducing enzyme. This enzyme, later designated as cellobiose : quinone oxi-doreductase possessed the ability to stop phenolic coupling during the laccase oxidation ofquaiacol. Further experiments also demonstrated the production of this enzyme by Chryso-sporium lignorum (= Sporotrichum pulverulentum or Phanerochaete chrysosporium;WESTERMARK and ERIKSSON 1974b) from which the enzyme was purified and character-ized as a flavoprotein with FAD as the prosthetic group. The molecular weight of the ho-mogeneous enzyme was 58 kDa (WESTERMARK and ERIKSSON 1975).
Except in Phanerochaete chrysosporium, cellobiose-quinone oxido-reductase was alsofound in 25 other white-rot fungi (ANDER and ERIKSSON 1977). The enzyme reduces a largenumber of quinones produced, for example, by laccase to the corresponding phenols whilecellobiose is simultaneously oxidized to cellobionolactone (WESTERMARK and ERIKSSON1974b). During this reaction hydrogen peroxide is gradually formed from oxygen (MORPETHand JONES 1986).

198 A. LEONOWICZ et al.
Cellobiose dehydrogenase (CDH; EC 1.1.99.18 – earlier called cellobiose oxidase): Theenzyme, for the first time isolated and purified from Sporotrichum pulverulentum byERIKSSON’s team, is a monomeric protein with a molecular mass around 90 kDa (AYERSet al. 1978). It is also a flavoprotein, but with cytochrome-b type heme in the catalytic cen-tre. This group probably transfers two electrons from the electron donor (e.g., cellobiose) toone electron acceptors, e.g., cytochrome-c, Fe3+ or dioxygen. Similarly to cellobi-ose:quinone oxidoreductase, cellobiose dehydrogenase reduces a range of quinones (e.g.,2,6-dichlorophenoloindophenol; SAMEJIMA and ERIKSSON 1991) and oxidizes cellobioseand related oligosaccharides. However, oxygen in this reaction (different from that in thecase of the cellobiose:quinone oxidoreductase), is probably not incorporated into the cello-biose, but reduced to superoxide and/or hydrogen peroxide (ANDER 1994, HENRIKSSON1995). Moreover, cellobiose dehydrogenase also oxidizes the reducing end groups of cello-dextrins and cellulose (AYERS et al. 1978, HABU et al. 1993). Similarities in basic activitiesof CBQ and CDH underlie the assumption that cellobiose : quinone oxidoreductase isprobably a product of the proteolytic cleavage of cellobiose dehydrogenase (HENRIKSSONet al. 1991, WOOD and WOOD 1992, HABU et al. 1993). Apart from the differences in themolecular masses, the enzymes can be distinguished by some differences in their specific-ity; for example, only cellobiose dehydrogenase utilizes cytochrome c as an electron ac-ceptor (SAMEJIMA and ERIKSSON 1991). Recently, it has been unequivocally shown thatboth enzymes reduce not only quinones but also phenoxyl and aryl cation radicals producedfrom aromatic substrates by laccase and peroxidases (ANDER et al. 1990, HENRIKSSON et al.1993, ANDER 1994, ANDER and MARZULLO 1997).
Aryl alcohol oxidases (AAOs; EC 1.1.3.7): Aryl alcohol oxidase is an extracellular, FAD-dependent enzyme, which was discovered in Trametes (Polystictus) versicolor by FARMERet al. (1960). Later the enzyme was also found in Phanerochaete chrysosporium (ASADAet al. 1995) as well as in the culture liquids of other white-rot fungi, e.g., Pleurotus sajor-caju (BOURBONNAIS and PAICE 1988, 1989), Pleurotus eryngii (GUILLÉN et al. 1990), Pleu-rotus ostreatus (SANNIA et al. 1991, PALMIERI et al. 1993) and Bjerkandera adusta(MUHEIM et al. 1990a, 1990b). Aryl alcohol oxidase from Pleurotus eryngii is a glycopro-tein with a molecular weight of 72.6 kDa which oxidizes a number of aromatic alcohols(aryl a- and a-b-unsaturated, g-aromatic alcohols) to the corresponding aldehydes whileoxygen is reduced to hydrogen peroxide (Fig. 11). The optimum pH of the reaction is abovepH 6 (MARZULLO et al., 1995) – this is different from lignin peroxidase which alsoproduces veratraldehyde from veratryl alcohol, but only at pHs below 4 (TIEN and KIRK1988).
Aryl alcohol dehydrogenase (AAD; EC 1.1.91): The NADPH-dependent aryl alcoholdehydrogenase was found as an intracellular enzyme in the white-rot fungus Pleurotuseryngii by GUILLEN and EVANS (1994). The enzyme, reducing aromatic aldehydes intoalcohols, supports the ligninolytic system by providing a suitable amount of aryl alcoholsacting as source of hydrogen peroxide (see above). It has also been suggested that the en-zyme is important for the stability of lignin peroxidase or mediators.
R1 R1R3 R3
R2 R2
CH OH2
CH OH2
CHOO
2O
2
AAO
H O2 2
H O2 2
Fig. 11Mechanism of aromatic alcohol oxida-tion by aryl alcohol oxidases. R1–R3represent various groups characterizingparticular alcohols. In the case of veratrylalcohol, R1 = R2 = OCH3, R3 = H.(adapted from de JONG et al. 1994)

Fungal laccase: properties and activity on lignin 199
Glyoxal oxidase (GLO; EC 1.2.3.5): Glyoxal oxidase is a further FAD-dependent, H2O2-generating enzyme discovered in growing cultures of Phanerochaete chrysosporium(KERSTEN and KIRK 1987). Later, its formation was also demonstrated for Phlebia radiata(VARES et al. 1995). Enzyme purification and characterization revealed that among others itoxidizes glyoxylic acid into oxalic acid while generating hydrogen peroxide. In this way,glyoxal oxidase supports peroxidase activities and supplies chelators for Mn3+ ions(KERSTEN et al. 1990).
Superoxide dismutases (SODs; EC 1.15.1.1): Superoxide dismutase (SODs) is an almostuniversal enzyme of aerobic micro-organisms (HASSAN 1989). It catalyzes the dismutationof the superoxide anion radical to molecular oxygen and hydrogen peroxide in all oxygenmetabolizing organisms and even in certain anaerobes (MCCORT et al. 1971). In thisway, SOD protects organisms from toxic effects of the reactive superoxide radical (O2
. –
= SOR).All currently known SODs are metal proteins containing iron (FeSOD), or manganese
(MnSOD), or copper plus zinc (Cu-ZnSOD) or nickel (NiSOD) as the prosthetic groups.FeSOD commonly occurs in prokaryotic organisms, e.g., Escherichia coli (YOST andFRIDOVICH 1973), but also in various fungi. The crystalline form of FeSOD was obtainedfrom Sulfolobus acidocaldarius by KARDINAHL et al. (1996). MALARCZYK et al. (1995)identified the FeSOD in the white-rot fungus Pleurotus sajor-caju and also in some otherPleurotus spp. Interestingly, the enzyme was produced in increased amounts in fungal fruitbodies and mycelia shortly after the appearance of laccase and peroxidases. MnSODs werepurified to homogeneity from the white-rot fungi Ganoderma microsporium (PAN et al.1997) and Phanerochaete chrysosporium (OZTURK et al. 1999). Recently a Cu-ZnSOD wasidentified in the laccase-producing mould Botrytis cinerea (CHOI et al. 1997). A novelNiSOD dismutase was isolated in 1996 from Streptomyces species (YOUN et al. 1996). Itwas homotetramer protein with little sequence similarity to known SODs (KIM et al. 1996).
The toxic and reactive SOR commonly appears during the quinone redox cycles, e.g., inthe laccase-producing, white-rot fungus Pleurotus eryngii (GUILLEN et al. 1997). During thecycle of this fungus, the cell-bound divalent reduction of quinones (Q) to hydroquinones(Q2–) is followed by the extracellular laccase-mediated oxidation of hydroquinones intosemiquinones (Q. –), which, in turn, are autoxidized to quinones. In both quinone oxidationphases production of SOR occurs:
Q2– + O2 � Q. – + O2. – (3)
Q. – + O2 � Q + O2. – (4)
Subsequently, SOR reacts as a reducing or oxidizing agent with other radicals produced byligninolytic enzymes, thus contributing to various lignin transformation processes, e.g.,aromatic ring fission (KAWAI et al. 1988b) or demethylation (POTTHAST et al. 1995). As anoxidant, SOR is able to generate highly reactive Mn3+ from Mn2+ and hydrogen peroxide:
O2. – + Mn2+ + 2H+ � Mn3+ + H2O2; (5)
as a reducing agent, SOR reacts with Fe3+ reducing it to Fe2+ yielding oxygen:
O2. – + Fe3+ � O2 + Fe2+; (6)
then, the reduced iron and hydrogen peroxide react with each other to form extremely reac-tive hydroxyl radicals via the FENTON reaction (FENTON 1894, WAILING 1975, GOLDSTEINet al. 1993, PRATAP and LEMLEY 1998):
Fe2+ + H2O2 � Fe3+ + HO. + OH– ; (7)

200 A. LEONOWICZ et al.
hydroxyl radicals are also produced from SOR and hydrogen peroxide in the Haber-Weissreaction (HABER and WEISS 1934):
O2. – + H2O2 � HO. + OH– + O2 (8)
As a one-electron reduction product of oxygen, SOR is not strong enough for ligninoly-sis, but produced with a share of oxygen, hydrogen peroxide or hydroxyl radicals, it mightbe an important agent in lignin transformation (FORNEY et al. 1982, GUILLÉN et al. 1997,PARK et al. 1997).
On the other hand, the highly reactive hydroxyl radicals can interact with DNA, proteins,lipids, or other biomolecules resulting in damage to the cells. To prevent the formation ofhydroxyl radicals from SOR, SOD is produced, which catalyzes its rapid dismutation to O2
and H2O2 (GUILLÉN et al. 1997, MCCORMICK et al. 1998) as follows:
2 O2. – + 2H.
� H2O2 + O2 . (9)
Hydrogen peroxide yielded here is either transformed to O2 and H2O by catalase or in-volved in peroxidative reactions. Consequently, the living cells (also of basidiomycetousfungi) commonly possess SOD, catalase and peroxidases, which are able to eliminate harm-ful oxygen species. According to our results, the maximum activity of SOD in lignin de-grading fungi appear when the activities of ligninolytic enzymes diminish indicating aregulative function of this enzyme in ligninolysis (Fig. 12).
Proteolytic activities in the white-rot Basidiomycete fungi: In the wood-decaying basidi-omycetes known for ligninolytic activities, proteolytic enzymes have been occasionallydetected and isolated. As the majority of these enzymes is extracellular (ERIKSSON andPETTERSSON 1982, STASZCZAK et al. 1996 and 2000), they might be of general importancewith respect to their effects on the lignin- or cellulolytic enzymes secreted by the same
25
20
15
10
5
0M P1 P2 P3 F1 F2 F3c F3s Ma
Stages of development
En
zym
eact
ivit
ies
Legend
peroxidase Ulaccase (nkatal/l)
SOD (U)phenol.oxid.(A315nm)
Fig. 12Activity of SOD and of ligninolytic enzymes in Pleurotus sajor-caju during growth in the stages ofprimordia and fruiting bodies. P – primordia; 1,2,3 – stages from small to big forms, F – fruit bodies;c – cap; s – stem of fruit body (adapted from MALARCZYK et al. 1995)

Fungal laccase: properties and activity on lignin 201
fungi. Such a proteolytic enzyme, namely carboxyl proteinase (EC 3.4.23.6), was firstfound in Pycnoporus coccineus and its substrate specificity was determined (ICHISHIMAet al. 1980). Three years later, the same authors (ICHISHIMA et al. 1983) found another pro-teolytic enzyme – acidic carboxypeptidase (EC 3.4.16.1) – in the same fungal genus, but ina different species (Pycnoporus sanguineus). STASZCZAK and NOWAK (1984) pointed to themultiple, extra-and intracellular proteolytic activities of the white-rot fungus Trametes ver-sicolor. Purification and partial characterization of two acid proteases from Sporotrichumpulverulentum grown on cellulose were described by ERIKSSON and PETERSON (1982,1988). The authors stated that the enzymes play a regulatory role in the activation ofthe endo-1,4-�-glucanases. The regulatory effect of proteases on lignin peroxidase and�-amylase was also proposed (DOSORETZ et al. 1990, DEY et al. 1991). HABU et al. (1993)demonstrated the release of a FAD domain from cellobiose oxidase (CBO) and the conver-sion of CBO to cellobiose:quinone oxidoreductase (CBQ) by proteases from cellulolyticcultures of Phanerochaete chrysosporium. The decrease in MnP activity due to the actionof proteases was observed in Phanerochaete chrysosporium (CRUZ-CORDOVA et al. 1999).The same report showed that LiP was less sensitive to proteolytic activities than MnP. Inaddition, reports from our laboratory (STASZCZAK et al. 1996 and 2000, STASZCZAK andGRZYWNOWICZ 1997) suggest both inhibiting and activating functions of Trametes versi-color serine proteinases in the regulation of laccase activity.
The growth of wood decaying fungi, especially under natural conditions, requires controlof their nitrogen economy, which involves the regulation of proteolytic activities for theintracellular protein turnover, the extracellular digestion of protein sources, and the modifi-cation of proteins through limited proteolysis. Lignin modifying enzymes are mainly ex-pressed during the secondary metabolic phase of the growth and be accompanied by in-creased protein turnover. Moreover, ligninolytic enzymes acting outside mycelium aresusceptible to the fungus’ extracellular proteinases; this is why their activity is of signifi-cance when the process of ligninolysis is considered.
Low-molecular mass mediators
According to several publications, the molecular size of wood-attacking enzymes does notpermit penetration of the undegraded plant cell wall (EVANS et al. 1991, 1994, FLUORNOYet al. 1991). On the other hand, there have been studies presenting evidence that some ofthese enzymes (e.g., laccase) are able to bleach hard wood pulp by depolymerizing andsolubilizing lignin in the presence of so-called mediator compounds (BOURBONNAIS andPAICE 1992, BOURBONNAIS et al. 1995, CALL 1994, 1996, CALL and MUCKE 1997,MAJCHERCZYK et al. 1999). Therefore, in recent years research has been focused on suchpotential low-molecular mass mediators of internal or external origin, which possess highenough redox potentials (>900 mV) to attack lignin and can migrate from the enzymes intothe tight lignocellulose complex. Consequently, many low-molecular mass compounds havebeen suggested to permeate wood cell walls and initiate decay. Examples of such sub-stances include veratryl alcohol, oxalate, 3-hydroxyanthranilic acid, and Gt-chelators. Theyare produced as a result of fungal metabolism and their secretion enables fungi to colonizeand degrade the wood cell wall structure more effectively than other organisms. The syn-thesis of veratryl alcohol was first observed in Phanerochaete chrysosporium (LUNDQUISTand KIRK 1978); oxalate and other organic acids were already found in culture liquids of anumber of wood-rotting fungi in 1965 (e.g., Armillaria mellea, Fomes annosus, Pleurotusostreatus, (TAKAO 1965). Later, their secretion was also shown for solid-state cultures(GALKIN et al. 1998, HOFRICHTER et al. 1999). 3-Hydroxyanthranilic acid was isolated andidentified from Pycnoporus cinnabarinus (EGGERT et al. 1996 and 1997), and a specialphenolate derivative, the so-called Gt-chelator (molecular mass <1 kDa), has been isolatedfrom the brown-rot fungus Gleophyllum trabeum (GOODELL et al. 1996).

202 A. LEONOWICZ et al.
Veratryl alcohol as well as oxalate and other dicarboxylic acids are fungal metabolites ofmany white-rot fungi: the first one is involved in the lignin peroxidase catalysis as a pro-tecting substance and may be involved in short-distance electron transfers while dicarbox-ylic acids stabilize highly reactive Mn3+ generated by manganese peroxidase. Additionally,the Mn3+-chelates can react with each other resulting in the formation of CO2 and carbon-centred radicals which, in turn, can add dioxygen to form peroxyl radicals (KEREM andHADAR 1997, HOFRICHTER et al. 1998e). The latter have recently been proposed to be deci-sive oxidants in lignin degradation (HAMMEL et al. 1999). Also, oxalate chelates other ca-tions such as Ca2+, Fe2+ and NH4
+ (DUTTON et al. 1993). The removal of calcium wouldenlarge the cell wall pore size, which may permit access of enzyme molecules (GREEN et al.1991). Moreover, oxalate regulates the concentration of ferric ions for the FENTON’s reac-tion (FENTON 1894, KOENIGS 1972). This reaction supplies the degradation environmentwith highly reactive hydronium ions (H3O+) and hydroxyl radicals (HO. and HO–) whichmay initiate lignocellulose degradation by certain fungi.
Four groups of benzene derivatives were identified as the components of the Gt-chelatorfrom Gleophyllum trabeum (Fig. 13). The presence of the chelator in this brown-rot fungusallows for the reduction of iron, the generation of oxygen radicals, and the degradation ofcellulosic and phenolic compounds (GOODELL et al. 1996).
The first natural mediator of laccase – 3-hydroxyanthranilic acid (3-HAA) – supportingligninolysis in Pycnoporus cinnabarinus, a white-rot fungus that does not produce manga-nese and lignin peroxidase, was discovered by EGGERT et al. (1996). The laccase/3-HAA
OCH3OCH3
OCH3OCH3
OCH3OCH3
OCH3OCH3 OCH3OCH3
OCH3OCH3 OCH3OCH3
OCH3OCH3
OCH3OCH3 OCH3OCH3 OCH3OCH3 OHOH O
OHOH O
O
O
O
O
O
O O
O
O
OH
OHOH
OHOH
OHOH
OHOH
OHOH
OHOH
OH
OH
OHOH
HO
HOHO
HO
HO
HOHO
HO
CH O3CH O3
H CO3H CO3
HO
H CO3H CO3
H CO3H CO3
H CO3H CO3
H C3H C3 H C3H C3
H CO3H CO3
H CO3H CO3 H CO3H CO3
H CO3H CO3
H CO3H CO3
H CO3H CO3
HydroxyphenylaceticHydroxyphenylacetic acid derivatives
Hydroxybenzoic acidacid derivatives
Hydroxybenzene derivativesderivatives
Dihydroxyphenylpentane - 1,4 - diolDihydroxyphenylpentane - 1,4 - diol
Fig. 13Structures of the selected phenol-ic compounds identified in theGt-chelator mixture of Gleophyl-lum trabeum (adapted fromGOODELL et al. 1996)

Fungal laccase: properties and activity on lignin 203
system mediates the oxidation of non-phenolic lignin model dimers whereas laccase alonecannot act on these compounds.
The dimeric ethers possessing phenolic groups are neither oxidized nor cleaved by thelaccase/3-HAA system. The laccase/3-HAA activity on the non-phenolic dimer, apart fromthe production of veratric acid and guaiacol, also results in a 6-electron oxidation of 3-HAA.This leads to the formation of a phenoxazinone ring containing compound from 3-HAA i.e.cinnabarinic acid (CA) which is a common secondary metabolite of Pycnoporus cinnabari-nus (EGGERT et al. 1995, 1996). However, TEMP and EGGERT (1999) have also demon-strated that in the presence of a suitable cellulose-derived electron donor, cellulose dehydro-genase (CDH) can regenerate the fungal mediator 3-HAA from cinnabarinic acid (Fig. 14).
Several years ago the delignification of kraft pulp by laccase was found to be supportedby a number of synthetic, low-molecular mass dyes or other aromatic hydrogen donors. Thefirst mediator that was shown to be effective in the delignification of kraft pulp and lignintransformation by laccase was ABTS [2,2�azinobis-(3-ethylbenzenthiazoline-6-sulfonicacid)] (BOURBONNAIS and PAICE 1992, 1996, BOURBONNAIS et al. 1997a).
The reaction mechanism mediated by ABTS seems to proceed as follows: oxygen acti-vates laccase and the mediator is oxidized by the enzyme. The oxidized mediator diffusesinto the pulp and oxidizes lignin disrupting it into smaller fragments which are easily re-moved from the pulp through the alkaline extraction. Superoxide dismutase (SOD) inhibitspolymerization of soluble lignin by laccase and accelerates the delignification process(adapted from SEALEY et al. 1997).
MAJCHERCZYK et al. (1999) demonstrated the oxidation of aromatic alcohols, used as non-phenolic lignin models, by laccase of Trametes versicolor in the presence of ABTS. Cationradical and dication formation were proposed to be involved in this laccase mediator system.
Apart from ABTS, 1-hydroxybenzotriazole (HBT) was recently introduced as an effec-tive laccase mediator in pulp processing (CALL 1994). An R-NO. radical is the active formof this mediator; it is also selective for lignin oxidation. The laccase-HBT couple can causeup to 50% of pulp delignification (SEALEY et al. 1997).
HBT is not less effective in the bleaching processes than ABTS. The discovery of HBTintroduced a new class of mediators with N-OH as the functional group, which is oxidizedto a reactive radical. Other mediators of this type, e.g., 4-hydroxy-3-nitroso-1-naphtalenesulfonic acid (HNNS), 1-nitroso-2-naphthol-3,6-disulfonic acid (NNDS), andRemazol brilliant blue (RBB), have also been shown to promote delignification, but theyare not as efficient as HBT and ABTS (BOURBONNAIS et al. 1997b) (Table 1).
2H O2
laccasered
O2
laccaseox
3-HAA
CDHred
CDHOX
cellobiose
induction
cellobiose
cellulose
cellobiono-
-lactone�
cinnabarinic acid
oxidation
intermediate
Fig. 14Model of interaction of CDHs and laccase during CA formation in P.cinnabarinus (TEMP and EGGERT1999)
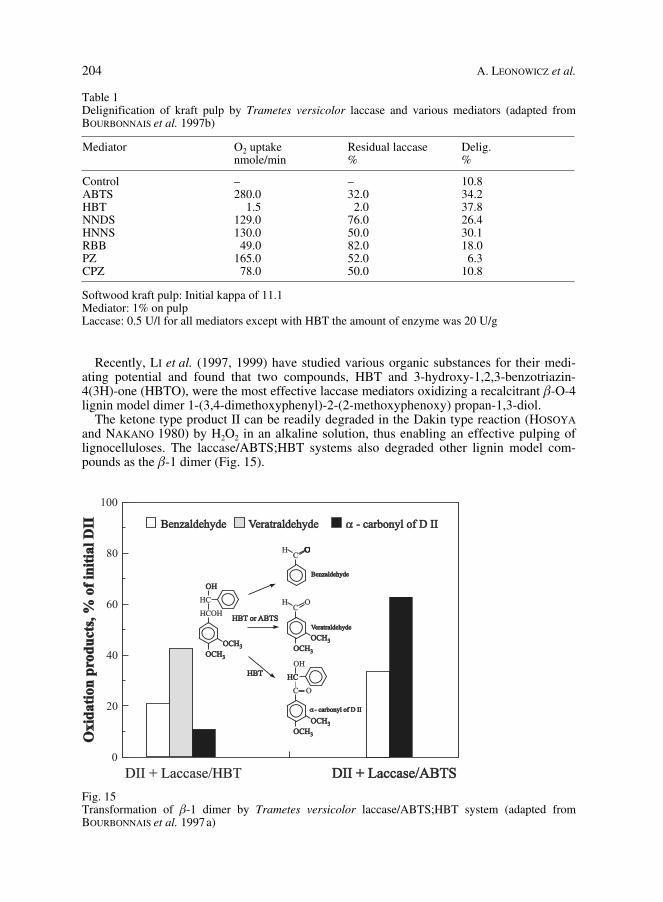
204 A. LEONOWICZ et al.
Table 1Delignification of kraft pulp by Trametes versicolor laccase and various mediators (adapted fromBOURBONNAIS et al. 1997b)
Mediator O2 uptakenmole/min
Residual laccase%
Delig.%
Control – – 10.8ABTS 280.0 32.0 34.2HBT 1.5 2.0 37.8NNDS 129.0 76.0 26.4HNNS 130.0 50.0 30.1RBB 49.0 82.0 18.0PZ 165.0 52.0 6.3CPZ 78.0 50.0 10.8
Softwood kraft pulp: Initial kappa of 11.1Mediator: 1% on pulpLaccase: 0.5 U/l for all mediators except with HBT the amount of enzyme was 20 U/g
Recently, LI et al. (1997, 1999) have studied various organic substances for their medi-ating potential and found that two compounds, HBT and 3-hydroxy-1,2,3-benzotriazin-4(3H)-one (HBTO), were the most effective laccase mediators oxidizing a recalcitrant b-O-4lignin model dimer 1-(3,4-dimethoxyphenyl)-2-(2-methoxyphenoxy) propan-1,3-diol.
The ketone type product II can be readily degraded in the Dakin type reaction (HOSOYAand NAKANO 1980) by H2O2 in an alkaline solution, thus enabling an effective pulping oflignocelluloses. The laccase/ABTS;HBT systems also degraded other lignin model com-pounds as the b-1 dimer (Fig. 15).
OCH3OCH3
BenzaldehydeBenzaldehyde
Oxid
ati
on
pro
du
cts,
%of
init
ial
DII
Oxid
ati
on
pro
du
cts,
%of
init
ial
DII
DII + Laccase/HBT DII + Laccase/ABTSDII + Laccase/ABTS
VeratraldehydeVeratraldehyde
OCH3OCH3
OCH3OCH3
OCH3OCH3
OCH3OCH3
OCH3OCH3
HC
HCHC
HCOH
C
C
C
O
H
H
O
- carbonyl of D II- carbonyl of D II
- carbonyl of D II- carbonyl of D II
VeratraldehydeVeratraldehyde
BenzaldehydeBenzaldehyde
HBT or ABTSHBT or ABTS
HBTHBT
OHOH
OH
0
20
40
60
80
100
Fig. 15Transformation of b-1 dimer by Trametes versicolor laccase/ABTS;HBT system (adapted fromBOURBONNAIS et al. 1997a)

Fungal laccase: properties and activity on lignin 205
Various laccases obtained from six strains of white-rot fungi were compared for theirdelignifying activity with ABTS and HBT as mediators (Table 2).
As it can be observed, there are differences among these laccases both in the delignifica-tion degree and the recovery of residual enzyme activities. Although the authors did notexplain this phenomenon, it seems that such differentiation may be a result of the variedstatus of copper in the activity center of particular laccases as well as of the different redoxpotentials (LI et al. 1999).
Even better results were obtained in pulp delignification using the sequential xylanaseand laccase/mediator system treatments followed by the alkaline peroxide treatment. Inthese experiments, HBT was used as the mediator. The final pulp brightness and reductionof viscosity after the xylanase treatment followed by two “traditional” alkaline peroxidestages were higher than those for laccase/HBT alone followed by two peroxide stages. Thisresult has yielded the greatest brightness of pulp of all biochemical/traditional techniquesapplied so far. Nevertheless, in the case of laccase-containing systems, the darkening effectof the enzyme should be taken into consideration (POPPIUS-LEVLING et al. 1997). It isworth mentioning here that laccase alone, working in the pulp under high oxygen pressure
Table 2Delignification of the kraft pulp with various laccases and ABTS and HBT as mediators (adaptedfrom BOURBONNAIS et al. 1997b)
Pulp Treatment (Laccase source) Residual laccase%
Delig.,%
Phlebia radiata + ABTS+ HBT
32.011.0
34.520.9
Trametes versicolor + ABTS+ HBT
27.013.0
30.030.0
Pleurotus ostreatus + ABTS+ HBT
n.d.1.4
30.012.7
Merulius tremellosus + ABTS+ HBT
86.048.0
32.719.1
Fomes fomentarius + ABTS+ HBT
15.013.0
33.631.8
Ganoderma colossum + ABTS+ HBT
63.024.0
30.040.0
Pulp: 4 g of SW O2 (Kappa = 11.0) at 10% in 50 mM sodium acetate, pH 5Mediator: 1% on pulpTreatment: 2 h, 60 �C, 140 kP of 02 followed by alkaline extraction
SO Na3
SO Na3
COOHCOOHHOHO
SO CH CH OSO Na2 2 2 3
SO CH CH OSO Na2 2 2 3
NH2
NH2
CH3
CH3
OH
CH OH2
CH OH2
O
O
OHH
H HO
H
OH H
O
O
OHHOHO
NH
OH
Fig. 16The chemical structures of Remazol brilliant Blue R (RBBR, left) and carminic acid (CA, right)

206 A. LEONOWICZ et al.
(5 Atm), caused a pulp brightness comparable to that of the laccase/mediator systems with-out increased oxygen pressure (LEONOWICZ et al. 1993). Such an experiment was conducted5 years ago, when both ABTS and other laccase mediators were not known or had just beendiscovered.
Remazol brilliant Blue R (RBBR) and carminic acid (CA) were also used as laccasemediators for bleaching the pulp black liquor (kraft effluent) (Fig. 16).
In the experiment CA used as a mediator caused considerably higher brightness of theblack liquor originating from pulp chlorination than RBBR did (LEONOWICZ et al. 1993).On the other hand, inducible forms of laccase of Kuehneromyces mutabilis and Pleurotusostreatus decolorized both RBBR and CA, which could be considered the ligninolytic ac-tivity of the enzyme degrading the dyes (CHO et al. 1999b). Some authors reported earlierthat decolorization of several polymeric dyes by Phanerochaete chrysosporium usuallyappears under the conditions which favour lignin degradation, thus pointing to interrelationsof dye decolorization and ligninolysis (GLENN and GOLD 1983, SPANDARO et al. 1992).
In the presence of ABTS, Trametes versicolor laccase also oxidized veratryl alcohol toveratraldehyde through the hydrogen abstraction from the �-carbon of the side chain andthe formation of a benzyl radical.
The mechanism of laccase/ABTS electrochemical oxidation was further studied andconfirmed by cyclic voltammetry and bulk electrolysis (BOURBONNAIS et al. 1997b, 1998).The reactions involved are shown in Fig. 17.
Veratryl alcohol is oxidized by the dication of ABTS2+ formed from ABTS via theABTS.+ radical. Thus, the laccase/mediator system oxidizes veratryl alcohol to veratralde-hyde similar to lignin peroxidase, but not via a cation radical intermediate.
The laccase/ABTS couple also cleaved a recalcitrant lignin model dimer (BLANCHETTE1991) while hardwood kraft pulp was bleached by an in vitro laccase/ABTS complex to anextent similar to that in whole fungal cultures (BOYLE et al. 1992). However, since ABTSand other hitherto proposed mediating compounds do not appear in pulp, it is necessary tosearch for novel natural mediators originating from the mycelium or lignocellulose. 3-Hydroxyanthranilate (3-HAA) and syringaldehyde could perform this role. The former isproduced as a natural redox mediator by Pycnoporus cinnabarinus; the latter is a product oflignin degradation. 3-HAA has been reported to be capable of mediating the oxidation ofveratryl alcohol and 14C-ring labelled synthetic lignin (EGGERT et al. 1996).
It was also shown that the laccase:syringaldehyde couple can oxidize veratryl alcohol(KAWAI et al. 1989). The presence of syringaldehyde in the wood degrading environment(also in pulp) is possible since syringic acid (readily reduced by fungi) is a lignin degrada-tion metabolite (CHEN et al. 1982). Thus, syringaldehyde can be considered a naturalequivalent of the ABTS mediator of laccase in wood and pulp. On the other hand, the
-e-
ABTS
Veratryl alcohol Veratraldehyde
In solution
Elektrodesurface
E E472 mV 885 mV
-2H+
-2e-
ABTS ABTS
-e-
. 2 ++: :
Fig. 17Redox reactions of veratryl alcohol andABTS (adapted from BOURBONNAIS et al.1997b)

Fungal laccase: properties and activity on lignin 207
commercialization of these systems for pulp bleaching requires more effective and abun-dant mediators. The first attempt at using laccase/mediator couples for delignification in thepulp industry was made by the development of the LignozymR-Process, which is describedin detail in a review from 1997 (CALL and MUCKE 1997).
To sum up, ligninolytic enzymes – particularly laccase – often carry out their actualligninolytic activities by co-operating with low-molecular weight compounds acting asredox mediators. The ideal mediator should “form a high redox potential oxidation productin a highly reversible reaction” (BOURBONNAIS et al. 1997b). Cheaper bleaching of pulprequires optimization of both factors (BOURBONNAIS et al. 1997a and 1997b).
Cooperation of ligninolytic enzymes in the direct attack on the lignin polymer
Judging from what is already known about the very complex structure of the lignin macro-molecule, it is hard to imagine that this biopolymer can be effectively degraded by oneenzyme alone. It seems logical to accept a hypothesis suggesting the co-operation of severalindividual enzymes in the process of lignin degradation. It is well known, for instance, thatcellulose, a biopolymer of a relatively simple structure (compared to lignin), requires for itstransformation a number of enzymes acting consecutively when the process of hydrolysiscontinues. The situation may be similar in the case of the lignin polymer. As the ligninmacromolecule has a complex structure, researchers have seldom worked with natural lig-nin preparations and used relatively low-molecular weight methoxyphenolic dimers oroligomers which model parts of the lignin structure. After the discovery of lignin peroxi-dase, researchers studied its direct effect on the lignin macromolecule, but with time it hasbecome more and more evident that the reproduction of lignin depolymerization by onlyone enzyme (namely LiP) is a limited approach. On the other hand, it was known at thattime that LiP needs hydrogen peroxide for its activity. Accordingly, researchers searched
OCH3OCH3OCH3OCH3
OCH3OCH3OCH3OCH3
veratrylveratryl
cationradicalcationradical
GLYOXALGLYOXALglyoxic acidglyoxic acidglyoxalglyoxal
OXIDASEOXIDASE
SpontaneonusSpontaneonus
LMW ProductsLMW Products
alcoholalcohol
LIGNIN PEROXIDASELIGNIN PEROXIDASE
MN PEROXIDASEMN PEROXIDASE
OO
HOHOHOHO and Mn2 +and Mn2 +H O2 2H O2 2
H O2 2H O2 2
CO2CO2
H O2 2H O2 2
H O2H O2 O2O2
O2O2
LigninLigninO
OO
HO
OO
O
OHOH
OHOH
+.
Fig. 18Possible co-operation of three enzymes (glyoxal oxidase, manganese peroxidase and lignin peroxi-dase) in ligninolysis Phanerochaete chrysosporium (adapted from KERSTEN et al. 1990)

208 A. LEONOWICZ et al.
for enzymes comprising this compound. Interestingly, one of the enzymes that generateshydrogen peroxide is manganese peroxidase. For this reason, PASZCZYNSKI et al. (1985)considered the possibility of co-operation of both enzymes in the transformation of ligninrelated compounds.
Five years later, KERSTEN et al. (1990) introduced veratryl alcohol as a mediator in ligni-nolysis and suggested the possibility of co-operation of lignin peroxidase with two otherenzymes, namely glyoxal oxidase and manganese peroxidase. Such a system may work incultures of Phanerochaete chrysosporium. For this reason, the lignin depolymerization tolower-molecular weight products (LMW) goes through the cation radicals found in theirlaboratory (Fig. 18).
Glucose oxidase as the hydrogen peroxide donor can also co-operate with lignin peroxi-dase. Such a co-operation in the decomposition of aromatic dimers by a system of bothenzymes was shown by GLENN et al. (1983). Glucose oxidase also co-operates with laccase.As it has already been mentioned, laccase alone can directly depolymerize the lignin mac-romolecule, but the efficiency of depolymerization is low and depends on the rapid removalof reactive radicals or quinones from the media that are formed by the action of laccase(such phenoxy radicals are created, e.g., when laccase acts on 2,4,6-trimethoxyphenol).Spontaneous coupling of the radicals which are formed from phenolic lignin fragmentsleads to products of higher molecular weight (compare Fig. 2). It has turned out that glucoseoxidase is an efficient factor reducing radicals and quinones formed by the action of lac-case. It may function according to the mechanism proposed by GREEN (1977, Fig. 19).
The hydrogen acceptors in glucose oxidation are quinones or radicals (but not molecularoxygen), which are formed from the phenolic compounds by laccase. A similar course ofreaction was noticed by ALBERTI and KLIBANOV (1982) with regard to p-benzoquinone. Thequinoide compound was reduced to hydroquinone by glucose oxidase.
We have investigated the transformation of various phenolic compounds under the se-quential action of laccase and glucose oxidase (LEONOWICZ et al. 1997b). In all cases, thequinones formed by laccase disappeared from the reaction medium when glucose oxidasewas introduced into the reaction mixture. The oxidation of several phenolic compounds andBJÖRKMAN lignin by Pleurotus ostreatus laccase and the reduction of the correspondingquinones may serve as examples of this (Fig. 20).
OH OHOH
H O2 2H O2 2
O2O2
OHOH
OXIDIZEDDEGRADATION
PRODUCTS
OXIDIZEDDEGRADATION
PRODUCTS
POLYMERIZEDQUINOIDS
POLYMERIZEDQUINOIDS
GLUCONOLACTONE
GLUCOSEGLUCOSE
OHOHH HOHOHHO
O
O O OHOHO
+ O2+ O2
O
R R RR
I
IIII H
-2e-2e
R
Fig. 19Cyclic diagram of possible co-operation of laccase and D-glucose oxidase (adapted from GREEN1977)

Fungal laccase: properties and activity on lignin 209
0.0
0.2
0.4
0.6
0.8
1.0
10 20 30 40 50
Time (min.)
Ab
sorb
an
ce
(390
nm
)
More recently, MARZULLO et al. (1995, 1996) reported that not only glucose oxidase, butalso another FAD-dependent oxidase – veratryl alcohol oxidase – can co-operate withlaccase in the transformation of some phenol/quinone derivatives. The above mentionedenzyme is secreted by the white-rot fungus Pleurotus ostreatus. The authors found that theenzyme is able to reduce laccase-generated quinones and phenoxy radicals with concomi-
0.00
0.02
0.04
0.06
0.08
0.10
80 90 100 110 120 130 140
Fraction number
Ab
sorb
an
ce
(28
0n
m)
Fig. 21Elution pattern of Björkman lignin on a Sepharose Cl-6B column after treatment with both laccaseand glucose oxidase (adapted from LEONOWICZ et al. 1997b, 1999d)
Fig. 20Formation and reductionof quinones under se-quential action of lac-case and glucose oxi-dase on BJÖRKMAN lig-nin and several phenoliccompounds (adaptedfrom LEONOWICZ et al.1997b)

210 A. LEONOWICZ et al.
tant oxidation of veratryl alcohol to veratryl aldehyde. This system also works with syn-thetic quinones, e.g., 2,6-dichlorophenoloindophenol (DCIP). For this reason, DCIP wasused both to assay the quinone reduction and to test the effect of oxygen and veratryl alco-hol on the veratryl alcohol oxidase activity. The cycle given below demonstrates that vera-tryl alcohol oxidase reduces DCIP and consequently prevents the polymerization of guaia-col and ferulic acid by laccase.
This mechanism have regulatory properties, which may, in fact, be applicable in lignintransformation. The first attempt at applying laccase and glucose oxidase was made in ourlaboratory. Two fractions of lignosulphonates (Peritan Na) were applied as laccase sub-strates (SZKLARZ and LEONOWICZ 1986); it was demonstrated that glucose oxidase added tothe reaction mixtures effectively counteracted polymerization and even accelerated thedepolymerization process catalyzed by laccase.
This noticeable effect of laccase and glucose oxidase working in concert during ligni-nolysis was confirmed using Björkman lignin isolated from wheat straw (Fig. 21).
Similar results were reported by GALLIANO et al. (1991). The authors used 14C-labelledKLASON lignin isolated from 14C-labelled ligninocellulose. The latter biopolymer was takenfrom young Hevea twigs grown in a medium with 14C-phenylalanine. The lignin solubiliza-tion in the conditions containing the glucose/glucose oxidase system increases three-foldcompared to the conditions with laccase alone.
Analogously to glucose oxidase, veratryl alcohol oxidase may also co-operate with lac-case (MARZULLO et al. 1995). Its co-operative action prevented the polymerization of phe-nolic lignin fragments and reduced the molecular weight of soluble lignosulphonates to asignificant extent.
Time_(days)
! " # $ %
14C
Solu
biliz
ed(%
ofto
tal14
C)
"&
"!
&
!
Fig. 2214C lignin solubilization by laccase andmanganese peroxidase. (�) control with-out enzymes; (�) laccase and manganeseperoxidase together; (�) laccase, manga-nese peroxidase and glucose oxidasetogether (adapted from GALLIANO et al.1991)

Fungal laccase: properties and activity on lignin 211
POSSIBLEOSSIBLELACCASE AND SUPEROXIDE DISMUTASE (SOD) COOPERATIONLACCASE AND SUPEROXIDE DISMUTASE (SOD) COOPERATIONIN PRODUCTION OF LIGNINOLYTIC FACTORS AND RADICALSIN PRODUCTION OF LIGNINOLYTIC FACTORS AND RADICALS
LACCASELACCASE
SODSOD ( SOR dismutation )( SOR dismutation )
Lignin degradationLignin degradation
Lignin degradationLignin degradation
Q2Q2
Q2Q2 Fe3+Fe3+
Fe3+Fe3+
Fe+2Fe+2
Fe+2Fe+2
Q
Q
+
+
+
+
+
+
+
+
+
++
+
+
+
O2O2
O2O2
O2O2
O2O2
O2O2O2O2Haber - Weiss reaction:Haber - Weiss reaction:
hydroquinonehydroquinone
semiquinonesemiquinone
SOD
superoxide radical (SOR)superoxide radical (SOR)
superoxidesuperoxide dismutasedismutase
OHFenton reaction:Fenton reaction:
OH
OH
OH
( iron reduction )( iron reduction )O2O2
O2O2
O2O2
2O22O2 H O2 2H O2 2
H O2 2H O2 2
H O2 2H O2 2
O2O22H
=
=
=
=
=
=
=
=
=
=
Fig. 23The possible co-operation of laccase and superoxide dismutase in production of ligninolytic factorsand radicals (adapted from LEONOWICZ et al. 1999b)
Manganese peroxidase can also effectively co-operate with laccase, which was unequivo-cally stated in the report by GALLIANO et al. (1991). When manganese peroxidase and lac-case, both isolated from cultures of Rigidosporus lignosus, were added to the same reactionmixture containing radioactive KLASON lignin, lignin degradation was efficient. The addi-tion of glucose oxidase to such an enzyme system again increased the amount of water-soluble, low-molecular weight products. In the presence of either laccase or manganeseperoxidase alone, lignin solubilization was also observed, but at a relatively low level. Thiswas the first demonstration of the synergistic effect of two oxidizing enzymes involved inlignin biodegradation (Fig. 22).
In our more recent results, the maximum of SOD activity during lignin biodegradation bylaccase- and peroxidases-producing fungi appeared when the activities of ligninolytic en-zymes decreased (MALARCZYK et al. 1995, LEONOWICZ et al. 1999a, 1999c). This may becaused by the fact that when the ligninolytic enzymes are not active, SOD protects the or-ganisms from toxic, reactive radicals. On the other hand, during the high activity of theseenzymes, the radicals might be fully involved in the biodeterioration processes. SOD ismost likely involved in the production of these factors from the intermediates of quinoneredox cycling caused by laccase (see Fig. 23). This thesis, however, requires further ex-perimental confirmation.
Cooperation of laccase with other enzymes and mediating factorsin ligninocellulose degradation
It has been unequivocally stated that lignin degradation is accelerated in the presence ofcellulose or its oligomers (HATAKKA and UUSI-RAUVA 1983). The idea of a feedback typeinterdependence of delignification and cellulose degradation was already postulated by

212 A. LEONOWICZ et al.
WESTERMARK and ERIKSSON (1974a, b). This hypothesis is still valid as the reports byGOTTLIEB et al. (1950) concerning the possibility of fungal growth on lignin as the solesource of carbon and energy have not been confirmed; the lignin degradation is now gener-ally thought to be a cometabolic process. The processes of depolymerization of celluloseand of lignin are interrelated in certain points and even accelerate each other (WESTERMARKand ERIKSSON 1974a, 1974b, 1975). The authors discovered the enzyme cello-biose : quinone oxidoreductase, which co-operates with laccase and cellulose hydrolyzingenzymes in the depolymerization of both components of the ligninocellulose complex in afeedback fashion by removing decomposition products. The authors suggested that laccasemight function as a link in an extracellular “electron transport chain” in this system.
It is necessary to mention that ligninolytic peroxidases, nowadays regarded as the crucialenzymes in the disassembly of lignin, had not been known when cellobiose:quinone oxi-doreductase was discovered. For this reason, ANDER and MARZULLO (1997) modifiedWESTERMARK and ERIKSSON’s scheme by introducing the enzymes and radical intermedi-ates discovered in the meantime (Fig. 24).
The DCIP oxidizing and reducing laccase/aryl alcohol oxidase system described byMARZULLO et al. (1995) prevents lignin polymerization as well as considerably reduces themolecular masses. Aryl alcohol oxidase is a part of systems involving not only laccase butalso other enzymes and factors. The hypothetical relationship between aryl alcohol oxidase,laccase, and other enzymes and radicals in the process of ligninocellulose transformation isillustrated in Fig. 25.
In this proposed complex system, laccase, LiP and MnP are secreted by fungal hyphaeclose to the hyphae environment, where they co-operate with each other as well as withmediating factors. The yielded chelators and mediating radicals are exported further to thewood tissue where they work as enzyme “messengers” in ligninocellulose degradation.Therefore, it has been proposed that this feed-back type enzyme system works in lignino-cellulose transformation with a meaningful participation of different ligninolytic enzymes.
native
LiP, MnP, Laccase
native
LiP, MnP, Laccase
PhenolsPhenols
ProteaseProtease
Cellobiose + O2Cellobiose + O2 Cellobiono - 1,5 - lactoneCellobiono - 1,5 - lactone
Cellobionic acidCellobionic acid
+ heme+ hemeMn4+Mn4+ Mn2+Mn2+ Mn3+Mn3+Fe3+Fe3+
Fe2+Fe2+
native LiP, MnPnative LiP, MnPQuinones, radicals
Compound II
Quinones, radicals
Compound II
Lignin
CDH
Cellulose
CBQCBQ
H O or O2 2 2H O or O2 2 2
H O2 2H O2 2 OHOH
H O2H O2
, ,
+
Fig. 24Interaction of the cello-biose dehydrogenase/cellobiose: quinone oxi-doreductase system(CDH/CBQ) with otherenzymes, quinones andphenoxy radicals pro-duced by these enzymes(adapted from ANDERand MARZULLO 1997)

Fungal laccase: properties and activity on lignin 213
FADH2FADH2FADH2FADH2
Fe(II)Fe(II)
Mn(II) Mn(III)Mn(III)Mn(III) ( )Mn(III) ( )
Mn(III) ( )Mn(III) ( )
Mn(III) ( )Mn(III) ( )
Fe(III)Fe(III)
FADFAD
AADAAD
NADPH
NADP
LaccoseLiP or MnPLaccoseLiP or MnP
O2O2
H O2 2H O2 2
H O2 2H O2 2 H O2H O2
OH OH
OH
OHOHOH
MnPMnP
LiPLiP
H O2 2H O2 2
H O2 2H O2 2 H O2H O2
++
+
+ +
+ +
-
-
+
+ +
+
+ +
+
+
+
+
H
Hypho
LignocelluloseLignocellulose
( )
Fig. 25Interaction of aryl alcohol dehydrogenase and other fungal enzymes with mediators and mediatingradicals during transformation of ligninocellulose. From left to right: AAD – aryl alcohol dehydroge-nase with NADP as the prosthetic group; aryl alcohol; aryl aldehyde; LiP – lignin peroxidase; MnP –manganese dependent peroxidase; lignin derived radicals or quinones and their reduced forms, VAO –veratryl alcohol dehydrogenase with FAD as the prosthetic group; LiP with Fe; MnP with Mn; metalchelating agents, e.g. oxalic acid (adapted from ANDER and MARZULLO 1997)
Laccase oxidizes lignin-derived radicals to quinones. These can serve as the oxygensource for glucose oxidase (GOD, SZKLARZ and LEONOWICZ 1986) and aryl alcohol oxidase(MARZULLO et al. 1995) producing H2O2 and preventing polymerization (SZKLARZ andLEONOWICZ 1986, MARZULLO et al. 1995); H2O2 formed serves as the co-substrate forligninolytic peroxidases. The CDH/CBQ system (Fig. 24) requires the presence of dioxygenand hydrogen peroxide for functioning; dioxygen is supplied from air, but external hydro-gen peroxide (needed at least by lignin peroxidase) is only produced at a low rate, by CDHand CBQ, as suggested by the authors. The crucial radical for H2O2 production is superox-ide (O2
. –), probably yielded from quinone cycling (see Superoxide dismutase) by generatingH2O2 through the oxidation of Mn2+. Simultaneously, highly reactive Mn3+ is formed. It isknown from manganese peroxidase to be an efficient lignin attacking agent.
O2. – + Mn2+ + 2H+ � Mn3+ + H2O2 (11)
H2O2 produced here accelerates both the production of Mn3+ from Mn2+ by manganeseperoxidase and the oxidation of lignin and derived fragments (WARIISHI et al. 1992).
However, since the H2O2 production by this system is not sufficient for all H2O2-consuming reactions, it was necessary to look for other natural systems generating enoughH2O2 for the biodeterioration of the ligninocellulose complex. Various possibilities havebeen considered. First of all, the possible involvement of certain enzymes was proven, e.g.,glyoxal oxidase, veratryl alcohol oxidase, manganese peroxidase, or glucose oxidase. Someof these enzymes can function both intracellularly and extracellularly. Other enzymes gen-erating hydrogen peroxide, such as pyranose-2-oxidase (mentioned above), methanol oxi-dase (EC 1.1.3.13), and fatty-acyl-CoA oxidoreductase (EC 1.2.1.42), work only intracel-lularly (DE JONG et al. 1994). This is why they should not be considered as woodtransformation is an exclusively extracellular process. The enzymes listed in the first group

214 A. LEONOWICZ et al.
are relatively stable and resistant to the conditions outside the cells. Among them, only glu-cose oxidase can join both lignin and carbohydrate metabolic chains. This means that glucoseoxidase possesses two possible functions for the acceleration of ligninocellulose degrada-tion: it produces H2O2 by oxidation of glucose (required for peroxidases), and it reduces qui-noids and phenoxy radicals originating from the action of laccase during lignin oxidation.
The remaining above listed enzymes which also reduce quinones and produce H2O2 (e.g.,aryl/veratryl alcohol oxidases) have been taken into consideration, but to a smaller extentbecause their H2O2 does not result from the oxidation of glucose, which is a key metabolitein carbohydrates transformation. Accordingly, probably only glucose oxidase co-operateswith the cellulolytic system by using glucose as the substrate generated by hydrolytic en-zymes from cellulose. The enzyme is produced by all white-rot fungi tested, includingPhanerochaete chrysosporium (KELLEY and REDDY 1988), which secrete laccase(LEONOWICZ et al. 1986) and/or peroxidases (ROGALSKI and LEONOWICZ 1992) and whichshow strong ligninolytic abilities (LEONOWICZ et al. 1986). Phanerochaete chrysosporiumwhich has been deprived of glucose oxidase by metagenesis is not able to decompose ligninat all (RAMASAMY et al. 1985).
In summary, glucose oxidase is a feed-back type enzyme that might prove to be impor-tant in ligninocellulose transformation. When oxygen is indispensable in the second stage ofthe glucose oxidase-glucose reaction, it can be supplied by radicals or quinones generatedwhen phenolic or methoxyphenolic compounds are exposed to laccase (SZKLARZ andLEONOWICZ 1986). On the other hand, when excess quinones produced by laccase inhibitthe enzyme (SZKLARZ and LEONOWICZ 1986), it can be concluded that glucose oxidasecounteracts the poisonous level of quinones in the medium and enables laccase to continueits function. Taking all these reasons into consideration, already more than 10 years ago wepostulated the following hypothetical mechanism of ligninocellulose complex transforma-tion (LEONOWICZ et al. 1986; Fig. 26).
CH OH3CH OH3
O2O2
O2O2
O2O2
CO2CO2
HO2
2
HO2
2
METHOXYPHENYLMETHO YPHENYLDERIVDERIVATIVESTIVES
LIGNIN
DISACCHARIDESDISACCHARIDES
DIPHENOLSDIPHENOLS
KETOACIDSKETOACIDS
KREBSCYCLEKREBSCYCLE
GLUCOSE
( XYLOSE )
OXIDASE
GLUCOSE
( YLOSE )
OXIDASE
QUINONESQUINONES -D-GLUCONO (XYLONO)LACTONE
-D-GLUCONO ( YLONO)LACTONE
�
PENTOSEPATHWAYPENTOSEPATHWAY
GLUCOSE(XYLOSE)GLUCOSE(XYLOSE)
WOODPOLYSACCHARIDESWOODPOLYSACCHARIDES
Dioxygenase
Dioxygenase
Lignindegradingenzyme
Lignindegradingenzyme
Gluco (xylo)
nasesGluco (xylo)
nases
Gluco (xylo)
sidasesGluco (xylo)
sidases
LaccaseLaccase
Fig. 26Hypothetical mechanism of ligninocellulose transformation by enzymes of white-rot fungi. The initialproducts of partial wood hydrolysis distinctly induce enzymatic systems accelerating the degradationprocesses (LEONOWICZ et al. 1986, ROGALSKI 1986)

Fungal laccase: properties and activity on lignin 215
In these mechanisms, glucose oxidase co-operates with lignin and manganese peroxidaseproviding hydrogen peroxide as well as with laccase reducing quinones to phenols. For thisreason, glucose oxidase operates as a feedback system where lignin and manganese peroxi-dase function as the first lignin attacking enzymes and laccase as the demethylating factor.Glucose, which is produced as a result of cellulose hydrolysis by the cellulase complex,becomes the substrate for glucose oxidase. Oxygen needed by glucose oxidase can be re-placed by quinones produced by laccase from lignin oligomers. As a result of glucose oxi-dation, �-D-gluconolactone is formed, which, after certain transformations, reinforces themetabolism of the fungus in the pentose phosphate cycle or in glycolysis. Hydrogen perox-ide produced in the reaction catalyzed by glucose oxidase activates, in turn, lignin andmanganese peroxidase. Lignin exposed to peroxidases undergoes decomposition into lower-molecular weight fragments containing methoxyl groups. Laccase demethylates these oli-gomers while peroxidases degrade them further into even smaller fragments. Generation ofexcess quinones and resulting repolymerization is counterbalanced by glucose oxidase,which reduces them to the respective phenols. The phenols, in turn, are the substrates fordioxygenases present in all fungal cells resulting in ring cleavages; the products obtained inthe form of ketoacids easily find their way to the Krebs cycle.
The above described mechanism could occur in those fungi which possess all the abovementioned enzymes, e.g., Phanerochaete chrysosporium, Phlebia radiata, Trametes versi-color, or Nematoloma frowardii. The mechanism suggested by WESTERMARK and
CH OH3CH OH3
H O2 2H O2 2
H O2 2H O2 2
H O2 2H O2 2
O2O2
CO2CO2
O2O2
Ligm
inase
Ligm
inase
LigninaseLigninase
LigninaseLigninase
Dioxygenase
Ring - cleavageproductsRing - cleavageproducts
Energy +pentosepathway
Energy +pentosepathway
GluconoXylonoCellobionolactones
GluconoXylonoCellobionolactones
PhenolsPhenols
KrebscycleKrebscycle
Ring - cleavageproductsRing - cleavageproducts
de novo
de novo
GlucoseXyloseGlucoseXylose
Glucose(Xylose)oxidases
Glucose(Xylose)oxidases
CBQXBQCBQXBQ
QuinonesPhenoxy radicalsQuinonesPhenoxy radicals
Methoxyphenylderivatives
Methoxyphenylderivatives
Laccase
Peroxidases
Laccase
PeroxidasesO
HO
22
2
OH
O
22
2
Gly
co-
sida
ses
Gly
co-
sida
ses
LIGNIN
LIGNIN
CellobioseXylobioseCellobioseXylobiose
Partlydegradedlignin
Partlydegradedlignin
WOODPOLYSACCHARIDESWOODPOLYSACCHARIDES
Ver. alc. Ver. alc.alc.
Cel
lula
ses
Xyl
anas
es
Cel
lula
ses
Xyl
anas
es
+ .+ .
+
Fig. 27Hypothetical scheme for de-gradation of lignin, celluloseand xylan in wood (adaptedfrom ERIKSSON et al. 1990,ERIKSSON 1993)

216 A. LEONOWICZ et al.
ERIKSSON, where cellobiose : quinone oxidoreductase is the crucial feed back type enzyme,seems to apply quite well to those species in which the presence of lignin peroxidase hasnot been confirmed but which transform ligninocellulose. It is not out of the question thatboth systems also act in concert as it was shown by ERIKSSON et al. (ERIKSSON et al. 1990,LEONOWICZ et al. 1991b, ROGALSKI 1992, ERIKSSON, 1993, Fig. 27).
As it can be observed, our proposed method of ligninocellulose transformation has beenfurther developed by Eriksson’s team by supplementing cellobiose:quinone oxidoreductaseand mediating factors. Its areas of functioning have been greatly extended (not one, but twoenzymes with feedback activity, veratryl alcohol, phenoxy radicals). We are in full agree-ment with this proposition, particularly since it also takes into consideration the contribu-tion of fungal laccase and peroxidases to the demethylation of lignin, and methoxyphenylderivatives, which was postulated by our group many years ago (TROJANOWSKI et al. 1966,LEONOWICZ and TROJANOWSKI 1965). This means that our old conception of the initiallignin transformation by these enzymes still remains valid. Recent discoveries concerningthe role of aryl alcohol oxidase, cellobiose dehydrogenase, aryl alcohol dehydrogenase, and,especially, low-molecular weight mediators and radicals in wood biodeterioration shown inFig. 24 (ANDER and MARZULLO 1997) support the proposed routes by explaining how high-molecular weight enzymes can function in the wood environment which is normally toocompact to allow the penetration of microbial enzymes. However, the existence of other,still-to-be-discovered mechanisms as well as of enzymes also operating as feedback sys-tems in the process of ligninocellulose transformation cannot be ruled out. Detailed investi-gations into these processes would definitely help solve the problems of how to utilizewaste lignocelluloses are accumulated as a result of human activities or how to protect ourforests from damaging microbial attack.
Conclusions
Considering earlier and present results, we can conclude that cellulose and lignin are de-graded simultaneously and that the general outline of the complementary character of car-bohydrates and lignin decomposition as well as the existence of enzymatic systems com-bining these processes are still valid. Demethylation caused by laccase after the initialactivity of lignin and manganese peroxidase appears to be important as the depolymerizingactivity of lignin peroxidase is inhibited by the phenolic groups of native lignin (ANDER andMARZULLO 1997). Also, the role of laccase (induced by toxic lignin degradation products orstress conditions) lies probably in the “activation” of some low-molecular weight mediatorsand radicals produced by fungal cultures. Such activated factors, also produced in coopera-tion with other enzymes, are probably exported into the wood microenvironment wherethey work in degradation processes as the “enzyme messengers.”
In conclusion, it is worth mentioning that the depolymerizing activity of laccase is ratherunusual, because its routine enzyme activity comprises polymerization or coupling reac-tions. However, this routine activity is usually employed in laboratories. Under naturalconditions laccase works in the wood environment in concert with other fungal enzymes,mediators and mediating radicals. Apart from polymerization, splitting processes are alsopossible there. It is in these processes that laccase finds the proper position among otherligninolytic enzymes.
Acknowledgements
Parts of the works cited here were financially supported by the European Community (the RTDE5 ECContract ICA2-CT-2000-10050), KBN SPUB-M.-5PR-UE/DZ 280/2000 and BS/UMCS projects.

Fungal laccase: properties and activity on lignin 217
References
AGEMATU, H., SHIBAMOTO, N., NISHIDA, H., OKAMOTO, R., SHIN, T. and MURAO, S., 1993. Oxidativedecarboxylation of 4-hydroxymandelic acid and 2-(4-hydroxyphenyl)glycine by laccase fromTrachyderma tsunodae and Myrothecium verrucaria. Biosci. Biotechnol. Biochem., 57, 1877–1881.
AISEMBERG, G. O., GROREWOLD, E., TACCIOLI, G. E., JUDEWICZ, N. D. and MAJOR, A. 1989.Transcript in the response of Neurospora crassa to protein synthesis inhibition by cycloheximide.Exp. Mycol., 13, 121–128.
AKTHAR, M., BLANCHETTE, R. A. and KIRK, T. K., 1997. Fungal Delignification and BiomechanicalPulping of Wood. Berlin: Springer.
ALBERTI, B. N. and KLIBANOW, A. M., 1982. Preparative production of hydroquinone frombenzoquinone catalyzed by immobilized D-glucose oxidase. Enzyme Microb. Technol., 4, 47–49.
ANDER, P., 1994. The cellobiose-oxidizing enzymes CBQ and CBO as related to lignin and cellulosedegradation – a review. FEMS Microbiol. Rev., 13, 297–312.
ANDER, P. and ERIKSSON, K.-E., 1976. The importance of phenol oxidase activity in lignindegradation by the white-rot fungus Sporotrichum pulverulentum. Arch. Microbiol., 109, 1–8.
ANDER, P. and ERIKSSON, K.-E., 1977. Selective Degradation of wood components by white-rot fungi.Physiol. Plant, 41, 239–248.
ANDER, P. and ERIKSSON, K.-E., 1985. Methanol formation during lignin degradation byPhanerochaete chrysosporium. Appl Microbiol. Biotechnol., 21, 96–102.
ANDER, P., ERIKSSON, K.-E. and YOU, H.-S., 1983. Vanillic acid metabolism by Sporotrichumpulverulentum: evidence for demethylation before ring-cleavage. Arch. Microbiol., 136, 1–6.
ANDER, P., ERIKSSON, M. E. R. and Eriksson, K.-E., 1985. Methanol production from lignin-relatedsubstances by Phanerochaete chrysosporium. Physiol. Plant., 65, 317–321.
ANDER, P. and MARZULLO, L., 1997. Sugar oxidoreductases and veratryl alcohol oxidase as related tolignin degradation. J. Biotechnol., 53, 115–131.
ANDER, P., MISHRA, C., FARELL, R. L. and ERIKSSON, K.-E., 1990. Redox reactions in lignindegradation: interactions between laccase, different peroxidases and cellobiose:quinoneoxidoreductase. J. Biotechnol., 13, 189–198.
ASADA, Y., WATANABE, A., OHTSU, Y. and KUWAHARA, M., 1995. Purification and characterizationof an aryl-alcohol oxidase from the lignin-degrading basidiomycete Phanerochaete chrysosporium.Biosci. Biotech. Biochem., 59, 1339–1341.
AYERS, A. R., AYERS, S. B. and ERIKSSON, K. E., 1978. Cellobiose oxidase, purification and partialcharacterization of a hemoprotein from Sporotrichum pulverulentum. Eur. J. Biochem., 90, 171–181.
BAO, W., FUKUSHIMA, Y., JENSEN, K. A., Jr., MOEN, M. A. and HAMMEL, K. E., 1994. Oxidativedegradation of non-phenolic lignin during lipid peroxidation by fungal manganese peroxidase.FEBS Lett., 354, 297–300.
BLANCHETTE, R., 1991. Delignification by wood-decay fungi. Annu. Rev. Phytopathol., 29, 381–398.BOLLAG, J. M., SHUTTLEWORTH, K. L. and ANDERSON, D. H., 1988. Laccase-mediated detoxification
of phenolic compounds. Appl. Environ. Microbiol., 54, 3086–3091.BOLLAG, J.-M. and LEONOWICZ, A., 1984. Comparative studies of extracellular fungal laccase. Appl.
Environ. Microbiol., 48, 849–854.BONNEN, A. M., ANTON, l. H. and ORTH, A. B., 1994. Lignin-degrading enzymes of the comercial
button mushroom, Agaricus bisporus. Appl. Environ. Microbiol., 60, 960–965.BOURBONNAIS, R., LEECH, D. and PAICE, M. G., 1998. Electrochemical analysis of the interactions of
laccase mediators with lignin model compounds. Biochim. Biophys. Acta, 1379, 381–390.BOURBONNAIS, R. and PAICE, M. G., 1988. Veratryl alcohol oxidases from the lignin-degrading
basidiomycete Pleurotus sajor-caju. Biochem. J., 255, 445–450.BOURBONNAIS, R. and PAICE, M. G., 1989. Oxidative enzymes from the lignin-degrading fungus
Pleurotus sajor-caju. Proc ACS Symp, pp. 472–481.BOURBONNAIS, R. and PAICE, M. G., 1990. Oxidation of non-phenolic substrates. An expanded role
for laccase in lignin biodegradation. FEBS Lett., 267, 99–102.BOURBONNAIS, R. and PAICE, M. G., 1992. Demethylation and delignification of kraft pulp by
Trametes versicolor laccase presence of 2,2�-azinobis-(3-ethylbenzthiazoline-6-sulphonate). Appl.Microbiol. Biotechnol., 36, 823–827.

218 A. LEONOWICZ et al.
BOURBONNAIS, R. and PAICE, M. G., 1996. Enzymatic delignification of kraft pulp using laccase and amediator. TAPPI J., 79, 199–204.
BOURBONNAIS, R., PAICE, M. G., LEECH, D. and FREIERMUTH, B., 1997a. Reactivities of variousmediators and laccases with kraft pulp and lignin model compounds. Appl. Environ. Microbiol.,63, 4627–4632.
BOURBONNAIS, R., PAICE, M. G., LEECH, D. and FREIERMUTH, B., 1997b. Reactivity and mechanismof laccase mediators for pulp delignification. In: TAPPI Proc. Biol. Sci. Symp., TAPPI Press., pp.335–338.
BOURBONNAIS, R., PAICE, M. G., REID, I. D., LANTHIER, P. and YAGUCHI, M., 1995. Lignin oxidationby laccase isozymes from Trametes versicolor and role of the mediator 2,2'-azinobis(3-ethylbenzthiazoline-6-sulfonate) in kraft lignin depolymerization. Appl. Environ. Microbiol., 61,1876–1880.
BOYLE, C. D., KROPP, B. R. and REID, I. D., 1992. Solubilization and mineralization of lignin bywhite rot fungi. Appl. Environ. Microbiol., 58, 3217–3224.
CALL, H. P., 1994. Process for modifying, breaking down or bleaching lignin, materials containinglignin or like substances: PCT Pat. Appl.: WO94/29510.
CALL, H. P., 1996. Mehrkomponentensystem zum Verandern, Abbau oder Bleichen von Lignin, lignin-haltigen Materialien oder anlichen Stoffen sowie Verfahren zu seiner Anwendung. EP 0717143A1.
CALL, H. P. and MUCKE, I., 1997. History, overview and application of mediated lignolytic systems,especially laccase-mediator-system (LignozymR-process). J. Biotechnol., 53, 163–202.
CAMPBELL, W. G., 1930. The chemistry of the white rots of wood. I. The effect on wood substance ofPolystictus versicolor (Linn.) F. Biochem. J. (Cambridge), 24, 1235–1243.
CHEN, C.-L., CHANG, H.-M. and KIRK, T. K., 1982. Aromatic acids produced during degradation oflignin in spruce wood by Phanerochaete chrysosporium. Holzforschung, 36, 3–9.
CHEN, C.-L., CHANG, H.-M. and KIRK, T. K., 1983. Carboxylic acids produced through oxidativecleavage of aromatic rings during degradation of lignin in spruce wood by Phanerochaetechrysosporium. J. Wood Chem. Technol., 3, 35–57.
CHO, N.-S., ROGALSKI, J., JASZEK, M., LUTEREK, J., WOJTAS-WASILEWSKA, M., MALARCZYK, E.,FINK-BOOTS, M. and LEONOWICZ, A., 1999a. Effect of coniferyl alcohol addition on removal ofchlorophenols from water effluent by fungal laccase. J. Wood Sci., 45, 174–178.
CHO, N.-S., PARK, J. M., CHOI, J. M., MATUSZEWSKA, A., JASZEK, M., GRZYWNOWICZ, K., MALAR-CZYK, E., TROJANOWSKI, K. and LEONOWICZ, A., 1999b. The effect of wood rotting fungi andlaccase on destaining of dyes and KP bleaching effluent. Korean J. Wood Sci. Technol., 27, 72–79.
CHOI, G. J., LEE, H. J. and CHO, K. Y., 1997. Involvement of catalase and superoxide dismutase inresistance of Botrytis cinerea to dicarboximide fungicide vinclozolin. Pestic. Biochem. Physiol.,59, 1–10.
CLUTTERBUCK, A. J., 1972. Absence of laccase from yellow-spored mutants of Aspergillus nidulans,J. Gen. Microbiol., 70, 423–435.
CRUZ-CORDOVA, T., ROLDAN-CARRILLO, T. G., DIAZ-CERVANTES, D., ORTEGA-LOPEZ, J., SAUCEDO-CASTANEDA, G., TOMASINI-CAMPOCOSIO, A. and RODRIGUEZ-VAZQUEZ, R., 1999. CO2 evolutionand ligninolytic and proteolytic activities of Phanerochaete chrysosporium grown in solid statefermentation. Resour. Conserv. Recyc., 27, 3–7.
DANIEL, C., VOLC, J., KUBATOVA, E. and NILSSON, T., 1992. Ultrastructural and immunocytochemicalstudies on the H2O2-producing enzyme pyranose oxidase in Phanerochaete chrysosporium grownunder liquid culture conditions. Appl. Environ. Microbiol., 58, 3667–3676.
DAWEL, G., KÄSTNER, M., MICHELS, J., POPPITZ, W., GÜNTHER, W. and FRITSCHE, W., 1997.Structure of a laccase-mediated product of coupling of 2,4-diamino-6-nitrotoluene to guaiacol, amodel for coupling of 2,4,6-trinitrotoluene metabolites to a humic organic soil matrix. Appl.Environ. Microbiol., 63, 2560–2565.
DE JONG, E., FIELD, A. and DE BONT, J. A. M., 1992. Evidence for a new extracellular peroxidase,Manganese-inhibited peroxidase from the white-rot fungus Bjerkandera adusta. FEBS Lett., 299,107–110.
DE JONG, E., FIELD, J. A. and DE BONT, J. A. M., 1994. Aryl alcohols in the physiology of ligninolyticfungi. FEMS Microbiol. Rev., 13, 153–188.
DEY, S., MAITI, T. K., SAHA, N., BANERJEE, R. and BHATTACHARYYA, B. C., 1991. Extracellularprotease and amylase activities in ligninase-producing liquid culture of Phanerochaetechrysosporium. Process. Biochem., 26, 325–329.

Fungal laccase: properties and activity on lignin 219
DITTMER, J. K., PATELA, N. J., DHAWALEB, S. W. and DHAWALEA, S. S., 1997. Production of multiplelaccase isoforms by Phanerochaete chrysosporium grown under nutrient sufficiency. FEMSMicrobiol. Lett., 149, 65–70.
DOSORETZ, C. G., DASS, S. B., REDDY, C. A. and GRETHLEIN, H. E., 1990. Protease-mediateddegradation of lignin peroxidase in liquid cultures of Phanerochaete chrysosporium. Appl.Environ. Microbiol., 56, 3429–3434.
DUTTON, M. V., EVANS, C. S., ATKEY, P. T. and WOOD, D. A., 1993. Oxalate production byBasidiomycetes, including the white-rot species Coriolus versicolor and Phanerochaetechrysosporium. Appl. Microbiol. Biotechnol., 39, 5–10.
EGGERT, C., TEMP, U., DEAN, J. F. and ERIKSSON, K. E., 1995. Laccase-mediated formation of thephenoxazinone derivative, cinnabarinic acid. FEBS Lett., 376, 202–206.
EGGERT, C., TEMP, U., DEAN, J. F. and ERIKSSON, K. E., 1996. A fungal metabolite mediatesdegradation of non-phenolic lignin structures and synthetic lignin by laccase. FEBS Lett., 391,144–148.
EGGERT, C., TEMP, U. and ERIKSSON, K. E., 1997. Laccase is essential for lignin degradation by thewhite-rot fungus Pycnoporus cinnabarinus. FEBS Lett., 407, 89–92.
EGUCHI, F., LEONOWICZ, A., HIGAKI, M. and FUKUZUMI, T., 1994. Laccase-less mutant induced byregeneration of protoplasts of Pleurotus Basidiomycetes. Mokuzai Gakkaishi, 40, 107–110.
ERIKSSON, K.-E., 1993. Concluding remarks: Where do we stand and where are we going? Ligninbiodegradation and practical utilization. J. Biotechnol., 30, 149–158.
ERIKSSON, K. E. and PETTERSSON, B., 1982. Purification and partial characterization of two acidicproteases from the white-rot fungus Sporotrichum pulverulentum. Eur. J. Biochem., 124, 635–642.
ERIKSSON, K.-E., BLANCHETTE, R. A. and ANDER, P., 1990. Biodegradation of lignin. In: Timell, T.E.(ed.): Microbial and Enzymatic Degradation of Wood and Wood Components. Berlin: Springer-Verlag, pp. 225–397.
ERIKSSON, K.-E. and PETTERSSON, B., 1988. Acid proteases from Sporotrichum pulverulentum.Methods Enzymol., 160, 500–508.
EVANS, C. S., GALLAGHER, I. M., ATKEY, P. T. and WOOD, D. A., 1991. Location of degradativeenzymes in white-rot decay of lignocellulose. Biodegradation, 2, 93–106.
EVANS, C. S., DUTTON, M. V., GUILLEN, F. and VENESS, R. G., 1994. Enzymes and small molecularmass agents involved with lignocellulose degradation. FEMS Microbiol. Rev., 13, 235–240.
FAHRAEUS, G. and REINHAMMAR, B., 1967. Large scale production and purification of laccase fromcultures of the fungus Polyporus versicolor and some properties of laccase A. Acta Chem. Scand.,21, 367–2378.
FAHRAEUS, G., NILSSON, R. and NILSSON, G., 1949. Studies in the decomposition of wood by meansof some white rot fungi. Svensk Bot. Tidskr., 43, 343–356.
FAHRAEUS, G., TULLANDER, H. and LJUNGGREN, H., 1958. Production of high laccase yields incultures of fungi. Physiol. Plant., 11, 631–643.
FARMER, V. C., HENDERSON, M. E. K. and RUSSELL, J. D., 1960. Aromatic alcohol-oxidase activity inthe growth medium of Polystictus versicolor. Biochem. J., 74, 257–262.
FENTON, H. J. H., 1894. Oxidation of tartaric acid in presence of iron. J. Chem. Soc., 65, 899–910.FINK-BOOTS, M. D., JASZEK, M. and LEONOWICZ, A., 1997. Heat shock stimulation of laccase in
Abortiporus biennis and Cerrena unicolor. Proc Biol Sci Symp, TAPPI press, pp. 395–396.FINK-BOOTS, M. D., MALARCZYK, E. and LEONOWICZ, A., 1999. Increased enzymatic activities and
levels of superoxide anion and phenolic compounds in cultures of Basidiomycetes aftertemperature stress. Acta Biotechnol., 4, 319–330.
FLOURNOY, D. S., KIRK, T. K. and HIGHLEY, T. L., 1991. Wood decay by brown-rot fungi: changes inpore structure and cell wall volume. Holzforschung, 45, 383–388.
FORNEY, L. J., REDDY, C. A., TIEN, M. and AUST, S. D., 1982. The involvement of hydroxyl radicalderived from hydrogen peroxide in lignin degradation by the white rot fungus Phanerochaetechrysosporium. J. Biol. Chem., 257, 11455–11462.
FORRESTER, I. T., GRABSKI, A. C., BURGESS, R. R. and LEATHAM, G. F., 1988. Manganese, Mn-dependent peroxidases, and the biodegradation of lignin. Biochem. Biophys. Res. Commun., 157,992–999.
FUKUZUMI, T., 1960. Enzymatic degradation of lignin. Part I. Paperchromatographical separation ofintermediate degradation products of lignin by the wood-rotting fungus Poria subacida (Peck).Sacc. Bull. Agr. Soc. Japan, 24, 728–736.

220 A. LEONOWICZ et al.
GALKIN, S., VARES, T., KALSI, M. and HATAKKA, A., 1998. Production of organic acids by differentwhite-rot fungi as detected using capillary zone electrophoresis. Biotechnol. Techniques, 12, 267–271.
GALLIANO, H., GAS, G., SERIS, J. L. and BOUDET, A. M., 1991. Lignin degradation by Rigidoporuslignosus involves synergistic action of two oxidizing enzymes: Mn peroxidase and laccase. EnzymeMicrob. Technol., 13, 478–482.
GIANFREDA, L. and BOLLAG, J-M., 1994. Effect of soils on the behavior of immobilized enzymes. SoilSci. Soc. Am. J., 58, 1672–1681.
GLENN, J. K. and GOLD, M. H., 1983. Decolorization of Several Polymeric Dyes by the Lignin-De-grading Basidiomycete Phanerochaete chrysosporium. Appl. Environ. Microbiol., 45, 1741–1747.
GLENN, J. K. and GOLD, M. H., 1985. Purification and characterization of an extracellular Mn(II)-dependent peroxidase from the lignin-degrading basidiomycete, Phanerochaete chrysosporium.Arch. Biochem. Biophys., 242, 329–341.
GLENN, J. K., MORGAN, M. A., MAYFIELD, M. B., KUWAHARA, M. and GOLD, M. H., 1983. Anextracellular H2O2-requiring enzyme preparation involved in lignin biodegradation by the white rotbasidiomycete Phanerochaete chrysosporium. Biochem. Biophys. Res. Comm., 114, 1077–1083.
GOLDSTEIN, S., MEYERSTEIN, D. and CZAPSKI, G., 1993. The Fenton reagents. Free Rad. Biol. Med.,15, 435–445.
GOODELL, B., LIU, J., JELLISON, L. J., PASZCZYNSKI, A. and FEKETE, F., 1996. Chelation activity andhydroxyl radical production mediated by low molecular weight phenolate compounds isolated fromGloeophyllum trabeum. In: MESSNER, K. and SREBOTNIK, E. (eds.): Biotechnology in the Pulp andPaper Industry, Recent Advances in Applied and Fundamental Research. Vienna, Austria: Facultas-Universitatsverlag, pp. 591–594.
GOTTLIEB, S., DAY, W. C. and PELCZAR, M. J., Jr., 1950. The biological degradation of lignin. II. Theadaptation of white-rot fungi to growth on lignin media. Phytopathology, 40, 926–935.
GREEN, F., 1977. Significance of glucose oxidase in lignin degradation. Nature, 268, 78–80.GREEN, F., LARSEN, M. J., WINARANDY, J. E. and HIGLEY, T. L., 1991. Role of oxalic acid in incipient
brown rot decay. Mat. Organisms, 26, 191–213.GUILLEN, F. and EVANS, C. S., 1994. Anisaldehyde and veratraldehyde acting as redox cycling agents
for H2O2 production by Pleurotus eryngii. Appl. Environ. Microbiol., 60, 2811–2817.GUILLEN, F., MARTINEZ, A. T. and MARTINEZ, M. J., 1990. Production of hydrogen peroxide by aryl-
alcohol oxidase from the ligninolytic fungus Pleurotus eryngii. Appl. Microbiol. Biotechnol., 32,465–469.
GUILLEN, F., MARTINEZ, M. J., MUNOZ, C. and MARTINEZ, A. T., 1997. Quinone redox cycling in theligninolytic fungus Pleurotus eryngii leading to extracellular production of superoxide anionradical. Arch. Biochem.Biophys., 339, 190–199.
HABER, F. and WEISS, J., 1934. The catalytic decomposition of hydrogen peroxide by iron salts. Proc.R Soc. London (A), 147, 332–351.
HABU, N., SAMEJIMA, M., DEAN, J. F. D. and ERIKSSON, K.-E. L., 1993. Release of the FAD domainfrom cellobiose oxidase by proteases from cellulolytic cultures of Phanerochaete chrysosporium.FEBS Lett., 327, 161–164.
HAEMMERLI, S. D., SCHOEMAKER, H. E., SCHMIDT, H. W. H. and LEISOLA, M. S. A., 1987. Oxidationof veratryl alcohol by the lignin peroxidase of Phanerochaete chrysosporium. FEBS Lett., 220,149–154.
HAIDER, K., LIM, S.-U. and FLAIG, W., 1962. Investigations on the effects of microorganisms on 14C-labelled phenolic compounds. Landwirtsch. Forsch., 15, 196–204.
HAMMEL, K. E., JENSEN, K. A., KEREM, Z., SREBOTNIK, E. and KAPITCH, A. N., 1999. Fungal attackon lignocellulose – a radical approach. Proc 7th Intern Fungal Biol. Conf., pp. 57
HAMMEL, K. E., JENSEN, K. A., Jr., MOZUCH, M. D., LANDUCCI, L. L., TIEN, M. and PEASE, E. A.,1993. Ligninolysis by a purified lignin peroxidase. J. Biol. Chem., 268, 12274–12281.
HARKIN, J. M. and OBST, J. R., 1974. Demethylation of 2,4,6-trimethoxyphenol by phenol oxidases.TAPPI J., 57, 118–121.
HASSAN, H. M., 1989. Microbial superoxide dismutases. Adv. Genet., 26, 65–97.HATAKKA and UUSI-RAUVA, 1983. Degradation of 14C-labelled poplar wood lignin by selected white-
rot fungi. Eur. J. Appl.Microbiol. Biotechnol., 17, 235–242.HATAKKA, A., 1994. Lignin-modifying enzymes from selected white-rot fungi: production and role in
lignin degradation. FEMS Microbiol. Revs., 13, 125–135.

Fungal laccase: properties and activity on lignin 221
HATAKKA, A. I., KANTELINEN, A., TERVILÄ-WILO, A. L. M. and VIIKARI, L., 1987. Production ofligninases by Phlebia radiata in agitated conditions. In: ODIER, E. (ed.), Lignin: Enzymic andMicrobial Degradation. Paris: INRA Publishers, pp. 185–189.
HEINZKILL, M., BECH, L., HALKIER, T., SCHNEIDER, P. and ANKE, T., 1998. Characterization oflaccases and peroxidases from wood-rotting fungi (family Coprinaceae). Appl. Environ.Microbiol., 64, 1601–1606.
HENRIKSSON, G., 1995. Structure, function and applications of cellobiose dehydrogenase fromPhanerochaete chrysosporium. PhD, University, Uppsala.
HENRIKSSON, G., JOHANSSON, G. and PETTERSSON, G., 1993. Is cellobiose oxidase from Phanero-chaete chrysosporium a one-electron reductase? Biochim. Biophys. Acta, 1144, 184–190.
HENRIKSSON, G., PETTERSSON, G., JOHANSSON, G., RUIZ, A. and UZCATEGUI, E., 1991. Cellobioseoxidase from Phanerochaete chrysosporium can be cleaved by papain into two domains. Eur. J.Biochem., 196, 101–106.
HOFRICHTER, M. and FRITSCHE, W., 1997. Depolymerization of low-rank coal by extracellular fungalenzyme systems. II: The ligninolytic enzymes of the coal-humic-acid-depolymerizing fungusNematoloma frowardii b19. Appl. Microbiol. Biotechnol., 47, 419–424.
HOFRICHTER, M., SCHEIBNER, K., HATAKKA, A. and FRITSCHE, W., 1998a. Enzymatic combustion: astrategy to colonize lignocelluloses by Basidiomycetes. Proceedings of Abschlußkolloquiums desDFG-Sonderforschungsbereichs “Struktur- und Funktionsanalyse mikrobieller Lebensgemeinschaf-ten”.
HOFRICHTER, M., SCHEIBNER, K., SCHNEEGASS, I. and FRITSCHE, W., 1998b. Enzymatic combustionof aromatic and aliphatic compounds by manganese peroxidase from Nematoloma frowardii. Appl.Environ. Microbiol., 64, 399–404.
HOFRICHTER, M., SCHEIBNER, K., SCHNEEGASS, I., ZIEGENHAGEN, D. and FRITSCHE, W., 1998c.Mineralization of synthetic humic substances by manganese peroxidase from the white-rot fungusNematoloma frowardii. Appl Microbiol Biotechnol. 49, 584–588.
HOFRICHTER, M., VARES, T., KALSI, M., GALKIN, S., SCHEIBNER, K., FRITSCHE, W. and HATAKKA, A.,1999. Production of manganese peroxidase and organic acids and mineralization of 14C-labelledlignin (14C-DHP) during solid-state fermentation of wheat straw with the white rot fungusNematoloma frowardii. Appl. Environ. Microbiol., 65, 1864–1870.
HOFRICHTER, M., VARES, T., SCHEIBNER, K., GALKIN, S., SIPILA, J. and HATAKKA, A., 1998d.Mineralization and solubilization of synthetic lignin (DHP) by manganese peroxidases fromNematoloma frowardii and Phlebia radiata. J. Biotechnol., 67, 217–228.
HOFRICHTER, M., ZIEGENHAGEN, D., VARES, T., FRIEDRICH, M., JAGER, M. G., FRITSCHE, W. andHATAKKA, A., 1998e. Oxidative decomposition of malonic acid as basis for the action ofmanganese peroxidase in the absence of hydrogen peroxide. FEBS Lett., 434, 362–366.
HOSOYA, S. and NAKANO, J., 1980. Reaction of �-carbonyl group in lignin during alcaline hydrogenperoxide bleaching. Mokuzai Gakkaishi, 26, 97–101.
ICHISHIMA, E., KUMAGAI, H. and TOMODA, K., 1980. Substrate specificity of carboxyl proteinase fromPycnoporus coccineus, a wood-deteriorating fungus. Curr. Microbiol., 3, 333–337.
ICHISHIMA, E., YOSHIMURA, K. and TOMODA, K., 1983. Acid carboxypeptidase from a wood-deteriorating Basidiomycete Pycnoporus sanguineus. Phytochemistry, 22, 825–829.
ISHIHARA, T. and ISHIHARA, M., 1976. Oxidation of syringic acid by fungal laccase. MokuzaiGakkaishi, 22, 371–375.
ISHIHARA, T. and MIYAZAKI, M., 1972. Oxidation of milled wood lignin by fungal laccase. MokuzaiGakkaishi, 18, 415–419.
JOHANSSON, T. and NYMAN, P. O., 1987. A manganese(II)-dependent extracellular peroxidase fromthe white-rot fungus Trametes versicolor. Acta Chem. Scand., B41, 762–765.
KAPICH, A., HOFRICHTER, M., VARES, T. and HATAKKA, A., 1999. Coupling of manganeseperoxidase-mediated lipid peroxidation with destruction of nonphenolic lignin model compoundsand 14C-labelled lignins. Biochem. Biophys. Res. Commun., 259, 212–219.
KARDINAHL, S., SCHMIDT, C. L., PETERSEN, A. and SCHAFER, G. 1996. Isolation, characterization andcrystalization of an iron-superoxide dismutase from the crenarchaeon Sulfolobus acidocaldarius.FEMS Microbiol. Lett., 138, 65–70.
KAWAI, S., UMEZAWA, T. and HIGUCHI, T., 1988a. Degradation mechanisms of phenolic b-1 ligninsubstructure model compounds by laccase of Coriolus versicolor. Arch. Biochem. Biophys., 262,99–110.

222 A. LEONOWICZ et al.
KAWAI, S., UMEZAWA, T. and HIGUCHI, T., 1989. Oxidation of methoxylated benzyl alcohols bylaccase of Coriolus versicolor in the presence of syringaldehyde. Wood Res., 76, 10–16.
KAWAI, S., UMEZAWA, T., SHIMADA, M. and HIGUCHI, T., 1986. Ca-Ca-cleavage of b-1 ligninsubstructure models by the laccase of Coriolus versicolor. Proc 3rd Intern Conf Biotechnol PulpPaper Ind., pp. 39–43
KAWAI, S., UMEZAWA, T., SHIMADA, M. and HIGUCHI, T., 1988b. Aromatic ring cleavage of 4,6-di(tert-butyl)guaiacol, a phenolic lignin model compound, by laccase of Coriolus versicolor. FEBSLett., 236, 309–311.
KELLEY, R. L. and REDDY, C. A., 1988. Glucose oxidase of Phanerochaete chrysosporium. MethodsEnzymol., 161 B, 307–316.
KEREM, Z. and HADAR, Y., 1997. The role of manganese in enhanced lignin degradation by Pleurotusostreatus. Proc Biol Sci Symp, pp. 29–33.
KERSTEN, P. J. and KIRK, T. K., 1987. Involvement of a new enzyme, glyoxal oxidase, in extracellularH2O2 production by Phanerochaete chrysosporium. J. Bacteriol., 169, 2195–2201.
KERSTEN, P. J., STEPHENS, S. K. and KIRK, T. K., 1990. Glyoxal oxidase and the extracellularperoxidase of Phanerochaete chrysosporium. In: KIRK, T. K. and CHANG, H.-M. (eds.):Biotechnology in the Pulp and Paper Manufacture. Stoneham, MA: Butterworth-Heinemann, pp.457–463.
KERSTEN, P. J., TIEN, M., KALYANARAMAN, B. and KIRK, T. K., 1985. The ligninase fromPhanerochaete chrysosporium generates cation radicals from methoxybenzenes. J. Biol. Chem.,260, 2609–2612.
KIM, E.-J., KIM, H. P., HAH, Y. C. and ROE, J. H., 1996. Differential expression of superoxidedismutases containing Ni and Fe/Zn in Streptomyces coelicolor Muller. Mol. Microbiol., 241,178–185.
KIRK, T. K. and FARRELL, R. L., 1987. Enzymatic “combustion”: the microbial degradation of lignin.Ann. Rev. Microbiol., 41, 465–505.
KIRK, T. K. and HAMMEL, K. E., 1992. What is the primary agent of lignin degradation in white-rotfungi? In: KUWAHARA, M. and SHIMADA, M. (eds.): Biotechnology in the Pulp and Paper Industry.Tokyo: Uni Publishers, pp. 535–540.
KIRK, T. K., HARKIN, J. M. and COWLING, E. B., 1968. Degradation of the lignin model compoundsyringilglycol-b-guaiacyl ether by Polyporus versicolor and Stereum frustulatum. Biochim.Biophys. Acta, 165, 145–163.
KOENIGS, J. W., 1972. Production of extracellular hydrogen peroxide and peroxidase by wood-rottingfungi. Phytopathology, 62, 100–110.
KONISHI, K., INOUE, Y. and HIGUCHI, T., 1974. Decomposition of lignin by Coriolus versicolor. V. For-mation of carboxylgroups in the lignin treated with fungal laccase. Mokuzai Gakkaishi, 20, 26–30.
KOTTERMAN, M. J., HEESSELS, E., DE JONG, E., FIELD, J. A. and FIELD, J. A., 1994. The physiology ofanthracene biodegradation by the white rot fungus Bjerkandera sp. strain BOS55. Appl. Environ.Biotechnol., 42, 179–186.
KUAN, I. C. and TIEN, M., 1993. Stimulation of Mn peroxidase activity: a possible role for oxalate inlignin biodegradation. Proc. Natl. Acad. Sci. USA, 90, 1242–1246.
KUHAD, R. C., SINGH, A. and ERIKSSON, K.-E. L., 1997. Microorganisms and enzymes involved in thedegradation of plant fiber cell walls. In: ERIKSSON, K.-E. L. (ed.): Biotechnology in the Pulp andPaper Industry. Berlin: Springer, pp. 45–126.
KUWAHARA, M., GLENN, J. K., MORGAN, M. A. and GOLD, M. H., 1984. Separation andcharacterization of two extracellular H2O2-dependent oxidases from ligninolytic cultures ofPhanerochaete chrysosporium. FEBS Lett., 169, 247–250.
LANTE, A., CRAPISI, A, PASINI, G., ZAMORANI, A. and SPETTOLI, P. 1992. Immobilized laccase formust and wine processing. In: Annals of the New York Academy of Sciences Vol. 672, EnzymeEngineering 11 (CLARK, D. S. and D. A. ESTELL, eds), The New York Academy of Sciences NewYork, New York, pp. 558–561.
LEATHAM, G. F. and STAHMANN, M. A., 1981. Studies on laccase of Lentinus edodes: Specificity,localization and association with the development of fruiting bodies. J. Gen. Microbiol., 125,147–157.
LEONOWICZ, A., EDGEHILL, R. U. and BOLLAG, J.-M., 1984. The effect of pH on the transformation ofsyringic and vanillic acids by the laccases of Rhizoctonia praticola and Trametes versicolor. Arch.Microbiol., 137, 89–96.

Fungal laccase: properties and activity on lignin 223
LEONOWICZ, A., GIANFREDA, L., ROGALSKI, J., JASZEK, M., LUTEREK, J., WOJTAS-WASILEWSKA, M.,MALARCZYK, E., DAWIDOWICZ, A., FINK-BOOTS, M., GINALSKA, G., STASZCZAK, M. and CHO,N. S., 1997a. Appearance of laccase in wood-rotting fungi and its inducibility. Mokchae Konghak,25, 29–36.
LEONOWICZ, A., LUNDELL, T. K., ROGALSKI, J. and HATAKKA, A. I., 1991a. Demethylation andreduction of veratric acid by selected white-rot fungi. Acta. Microbiol. Polon., 40, 205–220.
LEONOWICZ, A., MATSUMOTO, Y., FUJI, R. and ISHIZU, A., 1993. Production of laccase with variousproperties and its application to pulp and paper industry. Proceeedings of 60th Paper and PulpSymposium, JSPPT Research Press, pp. 62–65.
LEONOWICZ, A., MATUSZEWSKA, A., LUTEREK, J., ZIEGENHAGEN, D., WOJTAS-WASILEWSKA, M.,CHO, N.-S., HOFRICHTER, M. and ROGALSKI, J., 1999a. Biodegradation of lignin by white rot fungi.Fungal Gen. Biol., 27, 175–185.
LEONOWICZ, A., MATUSZEWSKA, A., LUTEREK, J., ZIEGENHAGEN, D., WOJTAS-WASILEWSKA, M.,HOFRICHTER, M., ROGALSKI, J. and CHO, N.-S., 1999b. Effect of superoxide dismutase and lowmolecular mediators on lignin degradation. Korean J. Wood Sci. Technol., 27, 1–14.
LEONOWICZ, A., MATUSZEWSKA, A., LUTEREK, J., ZIEGENHAGEN, D., WOJTAS-WASILEWSKA, M.,HOFRICHTER, M., ROGALSKI, J. and CHO, N.-S., 1999c. White-rot Fungi as Lignin Degraders. Proc’99 Internat Conf For Sci, Chungbuk Nat Univ, Cheongju, Korea, pp. 175–193.
LEONOWICZ, A., ROGALSKI, J., JASZEK, M., LUTEREK, J., WOJTAS-WASILEWSKA, M., MALARCZYK, E.,GIANFREDA, L., FINK-BOOTS, M. and CHO, N.-S., 1997b. Feed-back activity of fungal laccase andglucose oxidase in transformation of Bjorkman lignin and some phenolic compounds. Proceeedingsof Internat Sem Forest Scie, Chungbuk Nat Univ, Cheongju, Korea, pp. 59–70.
LEONOWICZ, A., ROGALSKI, J., JASZEK, M., LUTEREK, J., WOJTAS-WASILEWSKA, M., MALARCZYK, E.,GINALSKA, G., FINK-BOOTS, M. and CHO, N.-S., 1999d. Cooperation of fungal laccase and glucose1-oxidase in transformation of BJORKMAN lignin and some phenolic compounds. Holzforschung,53, 376–380.
LEONOWICZ, A., ROGALSKI, J., SZCZODRAK, J. and FIEDUREK, J., 1986. The possible key role ofglucose oxidase in transformation of lignocellulose. Proceeedings of 3rd Intern. Conf. Biotechnol.Pulp Paper Ind, June 16–19, pp. 160–162
LEONOWICZ, A., ROGALSKI, J., WASILEWSKA, M. W.-L. and LUTEREK, J., 1991b. Properties andfunction of lignin peroxidase. In: Biochemical, Molecular and Physoplogical Aspects of PlantPeroxidases, (J. LOBARZEWSKI, H. GREPPIN, C. PENEL and Th. GASPAR eds.), Imperimarie NationalePress, Geneve, pp. 507–516.
LEONOWICZ, A., SZKLARZ, G. and WASILEWSKA, M. W.-L., 1985. The effect of fungal laccase onfractionated lignosulphonates (Peritan-Na). Phytochemistry, 24, 393–396.
LEONOWICZ, A. and TROJANOWSKI, J., 1965. Exoenzymes in fungi degrading lignin. I. Pholiotamutabilis. Acta Microb. Polon., 14, 55–61.
LEONOWICZ, A., TROJANOWSKI, J. and ORLICZ, B., 1978. Induction of laccase in Basidiomycetes.Apparent activity of the inducible and constitutive forms of the enzyme with phenolic substrates.Acta Biochim. Polon., 25, 369–378.
LEONTIEVSKY, A., MYASOEDOVA, N., POZDNYAKOVA, N. and GOLOVLEVA, L., 1997a. ‘Yellow’laccase of Panus tigrinus oxidizes non-phenolic substrates without electron-transfer mediators.FEBS Lett., 413, 446–448.
LEONTIEVSKY, A. A., VARES, T., LANKINEN, P., SHERGILL, J. K., POZDNYAKOVA, N. N., MYASOE-DOVA, N. M., KALKKINEN, N., GOLOVLEVA, L. A., CAMMACK, R., THURSTON, C. F. and HATAKKA,A., 1997b. Blue and yellow laccases of ligninolytic fungi. FEMS Microbiol. Lett., 156, 9–14.
LI, K., BERMEK, H. and ERIKSSON, K. E., 1997. A new technique for screening of laccase-mediators.Proc. Biol. Sci. Symp., pp. 349–353.
LI, K., XU, F. and ERIKSSON, K. E., 1999. Comparison of fungal laccases and redox mediators inoxidation of a nonphenolic lignin model compound. Appl. Environ. Microbiol., 65, 2654–2660.
LIU, S.-Y., FREYER, A. F., MINARD, R. D. and BOLLAG, J.-M. 1985. Enzyme-catalyzed complex for-mation of amino acid esters and phenolic humus constituents. Soil Sci. Soc.Am. J., 49, 337–342.
LOBARZEWSKI, J., 1981. New affinity chromatography method for the separation of vanillic acidinduced forms of fungal peroxidase from Trametes versicolor on porous glass beads activated withvanillin. Int. J. Biol. Macromol., 3, 77–81.
LOBARZEWSKI, J., TROJANOWSKI, J. and WOJTAS-WASILEWSKA, M., 1982. The effect of fungalperoxidase on Na-lignosulphonates. Holzforschung, 36, 173–176.

224 A. LEONOWICZ et al.
LUNDELL, T., 1993. Ligninolytic system of the white-rot fungus Phlebia radiata: lignin modelcompound studies. PhD, University of Helsinki, Helsinki, Finland.
LUNDQUIST, K. and KIRK, T. K., 1978. De novo synthesis and decomposition of veratryl alcohol by alignin-degrading basidiomycete. Phytochemistry, 17, 1676.
LUTEREK, J., GIANFREDA, L., WOJTAS-WASILEWSKA, M., CHO, N. S., ROGALSKI, J., JASZEK, M.,MALARCZYK, E., STASZCZAK, M., FINK-BOOTS, M. and LEONOWICZ, A., 1998. Activity of free andimmobilized extracellular Cerrena unicolor laccase in water miscible organic solvents.Holzforschung, 52, 589–595.
LUNDQUIST, K. and KRISTERSSON, P., 1985. Exhaustive laccase-catalysed oxidation of a lignin modelcompound (vanillyl glycol) produces methanol and polymeric quinoid products. Biochem. J., 229,277–279.
MAJCHERCZYK, A., JOHANNES, C. and HUTTERMANN, A., 1999. Oxidation of aromatic alcohols bylaccase from Trametes versicolor mediated by the 2,2�-azino-bis-(3-ethylbenzothiazoline-6-sulphonic acid) cation radical and dication. Appl. Microbiol. Biotechnol., 51, 267–276.
MALARCZYK, E., NOWAK, G., NOWAK, M., KOCHMANSKA-RDEST, J., FUKUZUMI, T. and LEONOWICZ,A., 1995. Relation between SOD, laccase and other enzymes during the fruiting process ofPleurotus specia growing on a ligninocellulosic medium. In: SREBOTNIK, E. and MESSNER, K.(eds.): Biotechnology in the Pulp and Paper Industry, Recent Advances in Applied andFundamental Research. Vienna, Austria: Facultas-Universitatsverlag, pp. 641–644.
MANZANARES, P., FAJARDO, S. and MARTIN, C., 1995. Production of ligninolytic activities whentreating paper pulp effluents by Trametes versicolor. J. Biotechnol., 43, 115–132.
MARTINEZ, M. J., RUIZ-DUENAS, F. J., GUILLEN, F. and MARTINEZ, A. T., 1996. Purification andcatalytic properties of two manganese peroxidase isoenzymes from Pleurotus eryngii. Eur. J.Biochem., 237, 424–432.
MARZULLO, L., CANNIO, R., GIARDINA, P., PALMIERI, G., AURILIA, E. and SANNIA, G., 1996. Co-operative action of Pleurotus ostreatus enzymes in lignin biodegradation. In: SREBOTNIK, E.and MESSNER, K. (eds.): Biotechnology in the Pulp and Paper Industry, Recent Advancesin Applied and Fundamental Research. Vienna, Austria: Facultas-Universitatsverlag, pp. 351to 356.
MARZULLO, L., CANNIO, R., GIARDINA, P., SANTINI, M. T. and SANNIA, G., 1995. Veratryl alcoholoxidase from Pleurotus ostreatus participates in lignin biodegradation and prevents polymerizationof laccase-oxidized substrates. J. Biol. Chem., 270, 3823–3827.
MCCORD, J. M., KEELE, B. B. and FRIDOVICH, I., 1971. Enzyme-based theory of obligate anaerobiosisphysiological function of superoxide dismutase. Proc. Natl. Acad. Sci. USA, 68, 1024–1027.
MCCORMICK, M. L., BUETTNER, G. R. and BRITIGAN, B. E., 1998. Endogenous superoxide dismutaselevels regulate iron-dependent hydroxyl radical formation in Escherichia coli exposed to hydrogenperoxide. J. Bacteriol., 180, 622–625.
MESTER, T. and FIELD, J. A., 1998. Characterization of a novel manganese peroxidase-ligninperoxidase hybrid isozyme produced by Bjerkandera species strain BOS55 in the absence ofmanganese. J. Biol. Chem., 273, 15412–15417.
MESTER, T., PENA, M. and FIELD, J. A., 1996. Nutrient regulation of extracellular peroxidases in thewhite rot fungus, Bjerkandera sp. strain BOS55. Appl. Microbiol. Biotechnol., 44, 778–784.
MORITA, Y., YAMASHITA, H., MIKAMI, B., IWAMOTO, H., ALBARA, S., TERADA, M. and MINAMI, J.,1988. Purification, crystallization and characterization of peroxidase from Coprinus cinereus. J.Biochem. (Tokyo), 103, 693–699.
MORPETH, F. F. and JONES, G. D., 1986. Resolution, purification and some properties of the multipleforms of cellobiose quinone dehydrogenase from the white-rot fungus Sporotrichum pulveru-lentum. Biochem. J., 236, 221–226.
MUHEIM, A., LEISOLA, M. S. A. and SCHOEMAKER, H. E., 1990a. Aryl-alcohol oxidase and ligninPeroxidase from the white-rot fungus Bjerkandera adusta. J. Biotechnol., 13, 159–167.
MUHEIM, A., WALDNER, R., LEISOLA, M. S. A. and FIECHTER, A., 1990b. An extracellular aryl-alcohol oxidase from the white-rot fungus Bjerkandera adusta. Enzyme Microb. Technol., 12,204–209.
NANNIPIERI, P. and BOLLAG, J.-M., 1991. Use of enzymes to detoxify pesticide-contaminated soilsand waters. J. Environ. Quality, 20, 510–517.
NIKU-PAAVOLA, M. L., KARHUNEN, E., SALOLA, P. and RAUNIO, V., 1988. Ligninolytic enzymes ofthe white-rot fungus Phlebia radiata. Biochem. J., 254, 877–884.

Fungal laccase: properties and activity on lignin 225
OZTURK, R., BOZKAYA, A., ATAV, E., SALAM, N. and TARHAN, L., 1999. Purification andcharacterization of superoxide dismutase from Phanerochaete chrysosporium. Enzyme Microb.Technol., 25, 392–399.
PALMIERI, G., GIARDINA, P., MARZULLO, L., DESIDERIO, B., NITTI, G., CANNIO, R. and SANNIA, G.,1993. Stability and activity of a phenol oxidase from the ligninolytic fungus Pleurotus ostreatus.Appl. icrobiol. Biotechnol., 39, 632–636.
PAN, S.-M., YE, Jr.-S. and HSEU, R.-S., 1997. Purification and characterization of manganesesuperoxide dismutase from Ganoderma microsporium. Biochem. Molecul. Biol. Intern., 42, 1035–1043.
PARK, J. S. B., WOOD, P. M., DAVIES, M. J., GILBERT, B. C. and WHIWOOD, A. C., 1997. A kineticand ESR investigation of iron(II) oxalate oxidation by hydrogen peroxide and dioxygen as a sourceof hydrogen radicals. Free Rad. Res., 27, 447–458.
PASZCZYNSKI, A. and CRAWFORD, R., 1995. Potential for bioremediation of xenobiotic compounds bythe white-rot fungus Phanerochaete chrysosporium. Biotechnol. Prog., 11, 368–379.
PASZCZYNSKI, A., HUYNH, V. B. and CRAWFORD, R., 1985. Enzymatic activities of an extracellular,manganese-dependent peroxidase from Phanerochaete chrysosporium. FEMS Microbiol. Lett., 29,37–41.
PASZCZYNSKI, A., HUYNH, V.-B. and CRAWFORD, R. C., 1986. Comparison of ligninase-1 andperoxidase-M2 from the white rote fingus Phanerochaete chrysosporium. Arch. Biochem.Biophys., 244, 750–765.
POPPIUS-LEVLING, K., WANG, W., RANUA, M., NIKU-PAAVOLA, M. L. and VIIKARI, L., 1997. Bio-bleaching of chemical pulps by laccase/mediator systems. Proceeedings of Biological SciencesSymposium, TAPPI Proceedings, pp. 329–333.
POTTHAST, A., ROSENAU, T., CHEN, C.-L. and GRATZL, J. S., 1995. Selective enzymatic oxidation ofaromatic methyl groups to aldehydes. J. Org. Chem., 60, 4320–4321.
PRATAP, K. and LEMLEY, A.T., 1998. Fenton electrochemical treatment of aqueous atrazine andmetachlor. J. Agric. Food Chem., 46, 3285–3291.
RAMASAMY, K., KELLEY, R. L. and REDDY, C. A., 1985. Lack of lignin degradation by glucoseoxidase-negative mutants of Phanerochaete chrysosporium. Biochem. Biophys. Res. Commun.,131, 436–441.
RIEBLE, S., JOSHI, D. K. and GOLD, M. H., 1994. Purification and characterization of a 1,2,4-trihydroxybenzene 1,2 dioxygenase from the basidiomycete Phanerochaete chrysosporium. J.Bacteriol., 176, 4838–4844.
RIGLING, D. and VAN ALFEN, N. K., 1991. Regulation of laccase biosynthesis in the plant-pathogenicfungus Cryphonectria parasitica by double-stranded RNA. J. Bacteriol. 173, 8000–8003.
RODRIGUEZ, C. S., SANTORO, R., CAMESELLE, C. and SANROMAN, A., 1997. Laccase production insemi-solid cultures of Phanerochaete chrysosporium. Biotechnol. Lett., 19, 995–998.
ROGALSKI, J., 1986. Cooperation of selected hydrolases and phenol oxidases in the degradation ofligninocellulose. Ph. D. thesis, Maria Curie-Sklodowska University, Lublin, pp. 1 –215.
ROGALSKI, J., 1992. Biotransformation of ligninocellulose by white-rot fungi for example of Phlebiaradiata. Habilitation thesis, Maria Curie-Sklodowska University, Lublin, pp. 1–196.
ROGALSKI, J. and LEONOWICZ, A., 1992. Phlebia radiata laccase forms induced by veratric acid andxylidine in relation to lignin peroxidase and manganese-dependent peroxidase. Acta Biotechnol.,12, 213–221.
ROGALSKI, J., LUNDELL, T., LEONOWICZ, A. and HATAKKA, A., 1991. Influence of aromaticcompounds and lignin on production of ligninolytic enzymes by Phlebia radiata. Phytochemistry,30, 2869–2872.
ROGALSKI, J., LUNDELL, T., LEONOWICZ, A. and HATAKKA, A., 1992. Influence of lignin-relatedaromatic compounds on the production of ligninolytic enzymes by Phlebia radiata. In:Ligninocellulosics: Science, Technology, Development and Use (J. F. KENNEDY, G. O. PHIPPIPSand P. A. WILLIAMS eds.), Ellis Horwood Ltd., New York, pp. 55–60.
ROGALSKI, J., WOJTAS-WASILEWSKA, M., BIALY, B., LUTEREK, J. and LEONOWICZ, A., 1996. A study ofaromatic ring cleavage enzymes in Phlebia radiata. In: 6th Intern. Conf. Biotechnol. Pulp Paper Ind.,(E. SREBOTNIK and K. MESSNER eds.), Facultas-Universitatverlag, Vienna, Austria, pp. 447–450.
RUTTIMANN, C., SCHWEMBER, E., SALAS, L., CULLEN, D. and VICUNA, R., 1992. Ligninolytic enzymesof the white rot basidiomycete Phlebia brevispora and Ceriporiopsis subvermispora. Biotech.Appl. Biochem., 16, 64–76.

226 A. LEONOWICZ et al.
SAMEJIMA, M. and ERIKSSON, K. E., 1991. Mechanisms of redox interactions between ligninperoxidase and cellobiose:quinone oxidoreductase. FEBS Lett., 292, 151–153.
SANNIA, G., LIMONGI, P., BUONOCORE, V., NITTI, G. and GIARDINA, P., 1991. Purification andcharacterization of a veratryl alcohol oxidase enzyme from the lignin degrading basidiomycetePleurotus ostreatus. Biochim. Biophys. Acta, 1073, 114–119.
SARKANEN, S., RAZAL, R. A., PICCARIELLO, T., YAMAMOTO, E. and LEWIS, N. G., 1991. Ligninperoxidase: toward a clarification of its role in vivo. J. Biol. Chem., 266, 3636–3643.
SCHEIBNER, K. and HOFRICHTER, M., 1998. Conversion of aminonitrotoluenes by fungal manganeseperoxidase. J. Basic Microbiol., 38, 51–59.
SCHNEEGASS, I., HOFRICHTER, M., SCHEIBNER, K. and FRITSCHE, W., 1997. Purification of the mainmanganese peroxidase isoenzyme MnP2 from the white-rot fungus Nematoloma frowardii b19.Appl. Microbiol. Biotechnol., 48, 602–605.
SEALEY, J. E., RUNGE, T. M. and RAGAUSKAS, A. J., 1997. Biobleaching of kraft pulps with laccaseand hydroxybenzotriazole. Proceeedings of Biological Sciences Symposium, TAPPI Proceedings,pp. 339–342.
SHIMADA, M. and HIGUCHI, T., 1983. Biochemical aspects of the secondary metabolism of xenobioticlignin and veratryl alcohol biosynthesis in Phanerochaete chrysosporium. In: HIGUCHI, T., CHANG,H.-M. and KIRK, T. K. (eds.): Recent Advances in Lignin Biodegradation Research. Tokyo: UniPublishers, pp. 195–208.
SHOEMAKER, H. E. and LEISOLA, M. S. A., 1990. Degradation of lignin by Phanerochaetechrysosporium. J. Biotechnol., 13, 101–109.
SIMMONS, K. E., MINARD, R. D. and BOLLAG, J.-M., 1989. Oxidative co-oligomerization of guaiacoland 4-chloroaniline. Environ. Sci. Technol., 23, 115–121.
SPANDARO, J. T., GOLD, M. H. and RENTGANATHAM, V., 1992. Degradation of azo dyes by the lignin-degrading fungus Phanerochaete chrysosporium. Appl. Environ. Microbiol., 58, 2397–2401.
SRINIVASAN, C., D’SUOZA, T. M., BOOMINATHAN, K. and REDDY, C. A., 1995. Demonstration oflaccase in the white rot Basidiomycete Phanerochaete chrysosporium. Appl. Environ. Microbiol.,61, 4274–4277.
STASZCZAK, M. and GRZYWNOWICZ, K., 1997. Proteolysis in vivo and in vitro in the white rot fungusTrametes versicolor: insight into the control of laccase activity. Proceeedings of BiologicalSciences Symposium, TAPPI Proceedings, pp. 101–104.
STASZCZAK, M. and NOWAK, G., 1984. Proteinase pattern in Trametes versicolor in response tocarbon and nitrogen starvation. Acta Biochim. Polon., 31, 431–437.
STASZCZAK, M., NOWAK, G., GRZYWNOWICZ, K. and LEONOWICZ, A., 1996. Proteolytic activities incultures of selected white-rot fungi. J. Basic Microbiol., 36, 109–203.
STASZCZAK, M., ZDUNEK, E. and LEONOWICZ, A., 2000. Studies on the role of proteases in the white-rot fungus Trametes versicolor. Effect of PMSF and chloroquine on ligninolytic enzymes activity.J. Basic Microbiol., 40, 51–63.
SZKLARZ, G. and LEONOWICZ, A., 1986. Cooperation between fungal laccase and glucose oxidase inthe degradation of lignin derivatives. Phytochemistry, 25, 2537–2539.
TAKAO, S., 1965. Organic acid production by basidiomycetes. I. Screening of acid-production strains.Appl. Microbiol., 13, 732–737.
TEMP, U. and EGGERT, C., 1999. Novel interaction between laccase and cellobiose dehydrogenaseduring pigment synthesis in the white rot fungus Pycnoporus cinnabarinus. Appl. Environ.Microbiol., 65, 389–395.
TIEN, M. and KIRK, T. K., 1983. Lignin-degrading enzyme from the Hymenomycete Phanerochaetechrysosporium Burds. Science, 221, 661–663.
TIEN, M. and KIRK, T. K., 1988. Lignin peroxidase of Phanerochaete chrysosporium. MethodsEnzymol., 161, 238–249.
TIEN, M., KIRK, T. K., BULL, C. and FEE, J. A., 1986. Steady-state and transient-state kinetic studieson the oxidation of 3,4-dimethoxybenzyl alcohol catalyzed by the ligninase of Phanerocheatechrysosporium Burds. J. Biol. Chem., 261, 1687–1693.
TROJANOWSKI, J., LEONOWICZ, A. and HAMPEL, B., 1966. Exoenzymes in fungi degrading lignin. ActaMicrobiol. Polon., 15, 17–22.
TUOR, U., WARIISHI, H., SCHOEMAKER, H. E. and GOLD, M. H., 1992. Oxidation of phenolic aryl-glycerol b-aryl ether lignin model compounds by manganese peroxidase from Phanerochaete chry-sosporium: oxidative cleavage of an �-carbonyl model compound. Biochemistry, 31, 4986–4995.

Fungal laccase: properties and activity on lignin 227
UMEZAWA, T. and HIGUCHI, T., 1987. Mechanism of aromatic ring cleavage of b-O-4 ligninsubstructure models by lignin proxidase. FEBS Lett., 218, 255–260.
URZUA, U., KERSTEN, P. J. and VICUNA, R., 1998. Manganese peroxidase-dependent oxidation ofglyoxylic and oxalic acids synthesized by Ceriporiopsis subvermispora produces extracellularhydrogene peroxide. Appl. Environ. Microbiol., 64, 68–73.
VARES, T., KALSI, M. and HATAKKA, A., 1995. Lignin peroxidases, manganese peroxidases, and otherligninolytic enzvmes produced by Phlebia radiaia during solid-state fermentation of wheat straw.Appl. Environ. Microbiol., 61, 3515–3520.
VARES, T., LUNDELL, T.K. and HATAKKA, A.I., 1992. Novel heme-containing enzyme possiblyinvolved in lignin degradation by the white-rot fungus Junghuhnia separabilima. FEMS Microbiol.Lett., 99, 53–58.
WAILING, C., 1975. Fenton’s reagent revisited. Acc. Chem. Res., 8, 125–131.WARIISHI, H., AKILESWARAN, L. and GOLD, M. H., 1988. Manganese peroxidase from the
basidiomycete Phanerochaete chrysosporium: spectral characterization of the oxidized states andthe catalytic cycle. Biochemistry, 27, 5365–5370.
WARIISHI, H., VALLI, K. and GOLD, M. H., 1991. In vitro depolymerization of lignin by manganeseperoxidase of Phanerochaete chrysosporium. Biochem. Biophys. Res. Commun., 176, 269–275.
WARIISHI, H., VALLI, K. and GOLD, M. H., 1992. Manganese(II) oxidation by manganese peroxidasefrom the basidiomycete Phanerochaete chrysosporium. Kinetic mechanism and role of chelators. J.Biol. Chem., 267, 23688–23695.
WESTERMARK, U. and ERIKSSON, K.-E., 1974a. Carbohydrate-dependent enzymic quinone reductionduring lignin degradation. Acta Chem. Scand., B28, 204–208.
WESTERMARK, U. and ERIKSSON, K.-E., 1974b. Cellobiose:quinone oxidoreductase, a new wood-degrading enzyme from white-rot fungi. Acta Chem. Scand., B28, 209–214.
WESTERMARK, U. and ERIKSSON, K.-E., 1975. Purification and properties of cellobiose:quinoneoxidoreductase from Sporotrichum pulverulentum. Acta Chem. Scand., B29, 419–424.
WIERTELAK, J., 1932. The effect of decay caused by white rot fungi on the chemical composition ofwood. Bull. Int. Acad. Polon. Sci. Lettr. C1 Sci. Math. Nat. Kraków (B), 1, 19–36.
WOJTAS-WASILEWSKA, M. and LUTEREK, J., 1987. The effect of fungal protocatechuate 3,4-dioxygenase on sodium lignosulphonate fractions. Phytochemistry, 26, 2671–2674.
WOJTAS-WASILEWSKA, M., LUTEREK, J., DAWIDOWICZ, A. and LEONOWICZ, A., 1987. Dearoma-tization of lignin derivatives by fungal protocatechuate 3,4-dioxygenase immobilized on porosityglass. Biotechnol. Bioeng., 29, 507–511.
WOJTAS-WASILEWSKA, M. and TROJANOWSKI, J., 1980. Purification and properties of protocatechuate3,4-dioxygenase from Chaetomium piluliferum induced with p-hydroxybenzoic acid. ActaBiochim. Polon., 27, 21–23.
WOJTAS-WASILEWSKA, M., TROJANOWSKI, J. and LUTEREK, J., 1983. Aromatic ring cleavage ofprotocatechuic acid by the white-rot fungus Pleurotus ostreatus. Acta Biochim. Polon., 30, 291 to302.
WOOD, J. D. and WOOD, P. M., 1992. Evidence that cellobiose:quinone oxidoreductase fromPhanerochaete chrysosporium is a breakdown product of cellobiose oxidase. Biochim. Biophys.Acta, 1119, 90–96.
WORRALL, J. J., CHET, I. and HUTTERMANN, A., 1986. Association of rhizomorph formation withlaccase activity in Armillariella spp. J. Gen. Microbiol., 132, 2527–2533.
YOST, F. J. and FRIDOVICH, I., 1973. An iron-containing superoxide dismutase from Escherichia coli.J. Biol. Chem., 248, 4905–4908.
YOUN, H.-D., KIM, E.-J., ROE, J.-H., HAH, Y. C. and KANG, S.-O., 1996. A novel nickel-containingsuperoxide dismutase from Streptomyces spp. Biochem. J., 318, 889–896.
ZAPANTA, L. S. and TIEN, M., 1997. The roles of veratryl alcohol and oxalate in fungal lignindegradation. J. Biotechnol., 53, 93–102.
Mailing address: Prof. Dr. ANDRZEJ LEONOWICZ, Department of Biochemistry, Maria-Curie-Sklodowska University, Pl-20031 Lublin, PolandTel.: +49-81-5375665; Fax: +48-81-5333669;E-mail: [email protected]




















