
Remazol Brilliant Blue R decolourization by the laccase from Trametes trogii
8
Remazol Brilliant Blue R decolourization by the laccase from Trametes trogii Tahar Mechichi * , Nejla Mhiri, Sami Sayadi Laboratoire des Bioproce ´de ´s, Centre de Biotechnologie de Sfax BP ‘‘K’’, 3038 Sfax, Tunisia Received 3 October 2005; received in revised form 22 December 2005; accepted 22 December 2005 Available online 13 February 2006 Abstract The decolourization of the recalcitrant dye RBBR by the culture filtrate of Trametes trogii and its isolated laccase was investigated. Both filtrates from Cu-induced cultures as well as purified laccase decolourized the dye RBBR. The purified laccase decolourized the dye down to 97% of 100 mg l 1 initial concentration of RBBR when only 0.2 U ml 1 of laccase was used in the reaction mixture. The effects of different physicochemical parameters were tested and optimal decolourization rates occurred at pH 5 and at a temperature of 50 °C. Decolourization of RBBR occurred in the presence of metal ions which could be found in textile industry effluents. Of all the metal ions tested, FeCl 2 was the most inhibiting for the decolourization. HBT was shown to have no effect on the decolourization of RBBR at low concentration, while at a concentration of 5 mM it slightly inhibited decolourization. The presence of aromatic compounds was found to be inhibiting for the decolourization at a concentration of 10 mM, but not at 0.1 mM, while at 1 mM only ortho-diphenols were inhibiting. Probing the effect of methanol it was found that higher concentrations caused a decrease in the decolourization rate of RBBR. The effect of laccase inhibitors on the decolourization of RBBR was tested with L-cysteine, SDS and EDTA. It was demonstrated that L-cysteine was the most inhibiting substrate for the decolourization while SDS was only inhibiting at 10 mM concentration and ETDA was not inhibiting at all tested concentrations. Ó 2006 Elsevier Ltd. All rights reserved. Keywords: Laccase; Synthetic dyes; Remazol Brilliant Blue R (RBBR); Trametes trogii; Decolourization 1. Introduction Laccases (benzenediol: oxygen oxidoreductases, EC 1.10.3.2) are widely distributed in nature. Laccase activity has been demonstrated in several organisms including higher plants, some insects and a few bacteria (Claus, 2003). However, most studied laccases are of fungal source. Laccase catalyses the oxidation of a huge variety of phe- nolic and non-phenolic substrates including ortho- and paradiphenols, aminophenols, polyphenols, polyamines, lignins and aryl diamines as well as some inorganic ions coupled to the reduction of molecular dioxygen to water (Solomon et al., 1996). The enzyme exhibits a broad sub- strate specificity, which can be enhanced by addition of redox mediators (Call and Mucke, 1997). Due to the broad variety of reactions catalysed by laccases, this enzyme holds great promise for many potential applications. Lac- case has been tested in many industrial applications such as pulp bleaching (Bajpai, 2004), textile dye decolouriza- tion (Wesenberg et al., 2003) and food industry (Minussi et al., 2002). Synthetic dyes are being increasingly used in the textile, paper, cosmetics, pharmaceutical and food industries. Unfortunately, these compounds chemically classified as azo, anthraquinone, heterocyclic, triphenylmethane or phthalocyanine dyes, cause serious environmental pollu- tion. Most of them are toxic, mutagenic and carcinogenic. Moreover, they are unusually resistant to degradation. De- colourization of these dyes by physical or chemical methods 0045-6535/$ - see front matter Ó 2006 Elsevier Ltd. All rights reserved. doi:10.1016/j.chemosphere.2005.12.061 * Corresponding author. Tel.: +216 74 440 816; fax: +216 74 440 818. E-mail address: [email protected] (T. Mechichi). www.elsevier.com/locate/chemosphere Chemosphere 64 (2006) 998–1005
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Remazol Brilliant Blue R decolourization by the laccase from Trametes trogii

www.elsevier.com/locate/chemosphere
Chemosphere 64 (2006) 998–1005
Remazol Brilliant Blue R decolourization by the laccasefrom Trametes trogii
Tahar Mechichi *, Nejla Mhiri, Sami Sayadi
Laboratoire des Bioprocedes, Centre de Biotechnologie de Sfax BP ‘‘K’’, 3038 Sfax, Tunisia
Received 3 October 2005; received in revised form 22 December 2005; accepted 22 December 2005Available online 13 February 2006
Abstract
The decolourization of the recalcitrant dye RBBR by the culture filtrate of Trametes trogii and its isolated laccase was investigated.Both filtrates from Cu-induced cultures as well as purified laccase decolourized the dye RBBR. The purified laccase decolourized the dyedown to 97% of 100 mg l�1 initial concentration of RBBR when only 0.2 U ml�1 of laccase was used in the reaction mixture. The effectsof different physicochemical parameters were tested and optimal decolourization rates occurred at pH 5 and at a temperature of 50 �C.Decolourization of RBBR occurred in the presence of metal ions which could be found in textile industry effluents. Of all the metal ionstested, FeCl2 was the most inhibiting for the decolourization.
HBT was shown to have no effect on the decolourization of RBBR at low concentration, while at a concentration of 5 mM it slightlyinhibited decolourization. The presence of aromatic compounds was found to be inhibiting for the decolourization at a concentration of10 mM, but not at 0.1 mM, while at 1 mM only ortho-diphenols were inhibiting. Probing the effect of methanol it was found that higherconcentrations caused a decrease in the decolourization rate of RBBR.
The effect of laccase inhibitors on the decolourization of RBBR was tested with L-cysteine, SDS and EDTA. It was demonstrated thatL-cysteine was the most inhibiting substrate for the decolourization while SDS was only inhibiting at 10 mM concentration and ETDAwas not inhibiting at all tested concentrations.� 2006 Elsevier Ltd. All rights reserved.
Keywords: Laccase; Synthetic dyes; Remazol Brilliant Blue R (RBBR); Trametes trogii; Decolourization
1. Introduction
Laccases (benzenediol: oxygen oxidoreductases, EC1.10.3.2) are widely distributed in nature. Laccase activityhas been demonstrated in several organisms includinghigher plants, some insects and a few bacteria (Claus,2003). However, most studied laccases are of fungal source.Laccase catalyses the oxidation of a huge variety of phe-nolic and non-phenolic substrates including ortho- andparadiphenols, aminophenols, polyphenols, polyamines,lignins and aryl diamines as well as some inorganic ionscoupled to the reduction of molecular dioxygen to water(Solomon et al., 1996). The enzyme exhibits a broad sub-
0045-6535/$ - see front matter � 2006 Elsevier Ltd. All rights reserved.
doi:10.1016/j.chemosphere.2005.12.061
* Corresponding author. Tel.: +216 74 440 816; fax: +216 74 440 818.E-mail address: [email protected] (T. Mechichi).
strate specificity, which can be enhanced by addition ofredox mediators (Call and Mucke, 1997). Due to the broadvariety of reactions catalysed by laccases, this enzymeholds great promise for many potential applications. Lac-case has been tested in many industrial applications suchas pulp bleaching (Bajpai, 2004), textile dye decolouriza-tion (Wesenberg et al., 2003) and food industry (Minussiet al., 2002).
Synthetic dyes are being increasingly used in the textile,paper, cosmetics, pharmaceutical and food industries.Unfortunately, these compounds chemically classified asazo, anthraquinone, heterocyclic, triphenylmethane orphthalocyanine dyes, cause serious environmental pollu-tion. Most of them are toxic, mutagenic and carcinogenic.Moreover, they are unusually resistant to degradation. De-colourization of these dyes by physical or chemical methods

T. Mechichi et al. / Chemosphere 64 (2006) 998–1005 999
including adsorption and precipitation methods, chemicaldegradation or photodegradation is financially and oftenalso methodologically demanding, time-consuming andmostly not very effective. At present, a number of studieshave focused on microorganisms, which are able to deco-lourize and biodegrade these dyes (Wesenberg et al., 2003).
RBBR is one of the most important dyes in the textileindustry. It is frequently used as starting material in theproduction of polymeric dyes. RBBR is an anthracenederivative and represents an important class of toxic andrecalcitrant organopollutants.
The decolourization of RBBR by white rot fungi hasbeen reported for a number of strains such as Dichomitus
squalens, Ischnoderma resinosum and Pleurotus calyptratus(Eichlerova et al., 2005) and Pleurotus ostreatus (Palmieriet al., 2005). In the case of single enzymes, lignin peroxi-dase produced by T. versicolor (Christian et al., 2005), purefungal laccase, obtained from a commercial formulationused in the textile industry (Soares et al., 2001a) and freeand immobilized laccase from T. versicolor (Peralta-Zamora et al., 2003) were used to study RBBR decolouri-zation. Besides its use as a model compound for the studiesof dye decolourization by white rot fungi, RBBR was alsoused as a redox mediator for laccase during the decolouri-zation of Sella Solid Red and Luganil Green by laccasefrom T. hirsuta cultures (Rodriguez Couto et al., 2005).
In this paper, decolourization of the dye RBBR by thelaccase from Trametes trogii was studied, as it is represen-tative of an important class of recalcitrant anthraquinone-type dyes.
2. Material and methods
2.1. Chemicals
2,6-Dimethoxyphenol (DMP), 1-hydroxybenzotriazole(HBT) and aromatic compounds were obtained fromSigma–Aldrich and Remazol Brilliant Blue R (RBBR)from ICN Biochemicals, Inc.
2.2. Fungal strain, media and culture conditions
T. trogii strain B6J, a newly isolated strain (Dhouibet al., 2005), was selected for this study for its ability to de-colourize RBBR in agar plates and to produce a hightiter of laccase. The culture was maintained on 2% maltextract and 1% agar plates grown at 30 �C and stored at4 �C. The medium used in this study contained the follow-ing (g l�1): glucose, 10; peptone, 5; yeast extract, 1; ammo-nium tartrate, 2; KH2PO4, 1; MgSO4 Æ7H2O, 0.5; KCl, 0.5;trace elements solution, 1 ml. The trace element solutioncomposition was as follows (g l�1): B4O7Na2 Æ10H2O, 0.1;CuSO4 Æ5H2O, 0.01; FeSO4 Æ7H2O, 0.05; MnSO4 Æ7H2O;0.01; ZnSO4 Æ7H2O, 0.07; (NH4)6Mo7O24 Æ4H2O, 0.01.The pH of the solution was adjusted to 5.5. After autoclav-ing, 3 ml of homogenized mycelium were used for the inoc-ulation of 300 ml of culture medium contained in 1000-ml
Erlenmeyer flasks. Cultures were incubated at 30 �C on arotary shaker (160 rpm). 150 lM of CuSO4 were addedto the basal medium to stimulate the production of laccase.
2.3. Enzyme and protein assays
Laccase activity was assayed using 10 mM DMP in100 mM sodium acetate buffer, pH 5 (e469 = 27500M�1 cm�1). The enzymatic reactions were carried out atroom temperature (22–25 �C) and one unit of enzymeactivity was defined as the amount of enzyme oxidizing1 lmol of substrate per min. Protein concentrationswere determined by the Bradford method, using the Bio-Rad protein assay with bovine serum albumin as thestandard.
2.4. Laccase purification
Laccase from T. trogii was purified from basal mediumcontaining 150 lM CuSO4. After incubation for 10 d, theculture liquid (1250 ml with specific activity of 8.1 U mg�1)was separated from mycelia by filtration through Whatmanfilter paper, concentrated and dialyzed against 10 mM tar-trate buffer (pH 5.5) by ultrafiltration (Filtron, 3 kDa cut-off membrane). Samples of 50 ml with a specific activity of8.11 U mg�1 of this crude enzyme preparation were appliedto a Hitrap Q FF cartridge (Amersham Biosciences) equil-ibrated with the same buffer at a flow rate of 1.5 ml min�1.The retained proteins were eluted over 100 min usingthe following NaCl gradient: 0–400 mM, 60 min; 400–1000 mM, 30 min and 1000 mM, 10 min. Fractions of2 ml with the laccase activity were pooled, concentrated(Filtron Microsep, 3 kDa cutoff), and 2 samples of 0.4 mlwere applied to a Superdex 75 (Pharmacia HR 10/30) col-umn equilibrated with 10 mM sodium tartrate buffer, pH5.5 containing 150 mM NaCl, at a flow rate of 0.4 mlmin�1. The laccase peak was pooled, concentrated (FiltronMicrosep, 3 kDa cutoff), dialyzed against 10 mM sodiumtartrate buffer 10 mM pH 5.5, and 1 ml samples appliedto a Mono-Q anion-exchange column (Pharmacia HR 5/5) equilibrated with the same buffer. Retained proteinswere eluted with a linear NaCl gradient from 0 to120 mM over 30 min, at a flow rate of 0.8 ml min�1.
Last step of purification resolved two laccase activitypeaks: a major LacI and a minor LacII. In this study weused the LacI isoenzyme as it is the major laccase producedby the fungus. The laccase isoenzyme was purified 8.7-foldwith a purification yield of 36.8% and a specific activityof 70.66 U mg�1. The kinetic constants against ABTS ofLacI were: Km = 0.050 mM, kcat = 344 s�1 and efficiency =6888 s�1 mM�1.
2.5. Decolourization of RBBR by the culture filtrates
of T. trogii
T. trogii was cultivated in 100 ml Erlenmyer flasks con-taining 25 ml M7 medium which was supplemented or not

1000 T. Mechichi et al. / Chemosphere 64 (2006) 998–1005
by 150 lM of CuSO4. After 12 d of incubation at 30 �C ona rotary shaker (200 rpm), the cultures were filtered toremove the mycelium and the culture filtrates were usedfor the decolourization of RBBR. The dye were added tothe culture filtrate from a stock solution to a final concen-tration of 200 mg l�1 and incubated at 30 �C at 200 rpm ona rotary shaker. The OD (595 nm) of the reaction mixturewas measured at regular time periods of 1 h.
2.6. RBBR decolourization by purified laccase
Unless otherwise indicated all experiments were per-formed using 3 ml-disposable cuvettes with 2 ml finalreaction volume. The reaction mixture was composed of100 mM acetate buffer pH 5, 200 mg l�1 RBBR and0.2 U ml�1 laccase. The reaction was initiated with addi-tion of laccase and further incubated in the dark at30 �C. The decolourization of the RBBR was followed byrecording the spectra of the reaction mixture (between400 and 700 nm) each 30 min or by measuring the OD at595 nm at a regular time period of 15 min. All experimentswere performed in duplicate and controls were performedby using heat inactivated enzymes after incubation at100 �C for 10 min. Decolourization of RBBR by theenzyme was also investigated by successive addition of50 mg l�1 RBBR. The addition of the dye to the reactionmixture was performed 4 times. Decolourization is definedas fellow: Decolourization (%) = 100(ODt0 � ODtf)/ODt0.Where ODt0 is the OD595 nm of the reaction mixture beforeincubation with the enzyme and ODtf is the OD595 nm afterincubation.
2.7. Effect of initial RBBR and enzyme concentration
on RBBR decolourization by purified laccase
The effect of RBBR concentration on the decolouriza-tion by laccase was studied in a first experiment at initialconcentration of 50, 100, 200 or 400 mg l�1 in the reactionmixture and 0.2 U ml�1 laccase in 100 mM acetate bufferpH 4. The effect of enzyme concentration was tested at dif-ferent levels: 0.025, 0.05, 0.1, 0.2, 0.4, and 0.8 U ml�1 witha RBBR concentration of 200 mg l�1.
2.8. Effect of pH and temperature on RBBR
decolourization by purified laccase
To study the effect of pH on the decolourization ofRBBR, 200 mg l�1 of the dye were incubated at 30 �C inthe presence of 0.2 U ml�1 pure laccase at different pH val-ues using the following buffers: 100 mM citrate for pH 3,100 mM acetate for pH 4 and 5, 100 mM phosphate forpH 6, 7 and 8. The effect of temperature was studied bythe incubation of 200 mg l�1 RBBR in the presence of0.2 U ml�1 laccase at the following temperature values:25, 30, 37, 50 and 60 �C. For each pH and temperaturevalue a reaction mixture without the enzyme was prepared
under the same conditions and was used to detect possiblechange of colour which is not due to enzyme activity.
2.9. Effect of metal ions on the decolourization of RBBR
In order to determine the effect of metal ions on the de-colourization of RBBR, 10 mM of: CoCl2, MnCl2, FeCl2,MgSO4, Na2SeO4, Na2MoO4, Na2WO4, CdCl2, CuCl2and NiCl2 were used in the reaction mixture which con-sisted of acetate buffer 100 mM pH 5, 200 mg l�1 RBBRand 0.2 U ml�1 followed by incubation at 30 �C.
2.10. Effect of aromatic compounds and HBT on dye
decolourization by purified laccase
The effect of aromatic compounds on dye decolouriza-tion was tested with catechol, pyrogallol, vanillin, syrin-gate, caffeate, gallate, protocatechuate, vanillate, ferulate,and 3,4-dihydroxyphenylacetate. All compounds were usedat a final concentration of 0.1, 1 or 10 mM. The reactionmixture was as described above.
To determine the effect of HBT concentration on thedecolourization of RBBR, HBT concentration in the reac-tion mixture was varied between 0 and 5 mM. The reactionmixture was as described above.
2.11. Effect of methanol and laccase inhibitors on RBBR
decolourization
The effect of methanol on RBBR decolourization wastested at concentrations ranging from 10% to 40%. The lac-case inhibitors L-cysteine, SDS and EDTA were tested atconcentrations of 0.1, 1 and 10 mM. The reaction mixtureconsisted of 100 mM acetate buffer pH 5, 200 mg l�1
RBBR and 0.2 U ml�1 of laccase.
3. Results and discussion
3.1. RBBR decolourization by culture filtrates of T. trogii
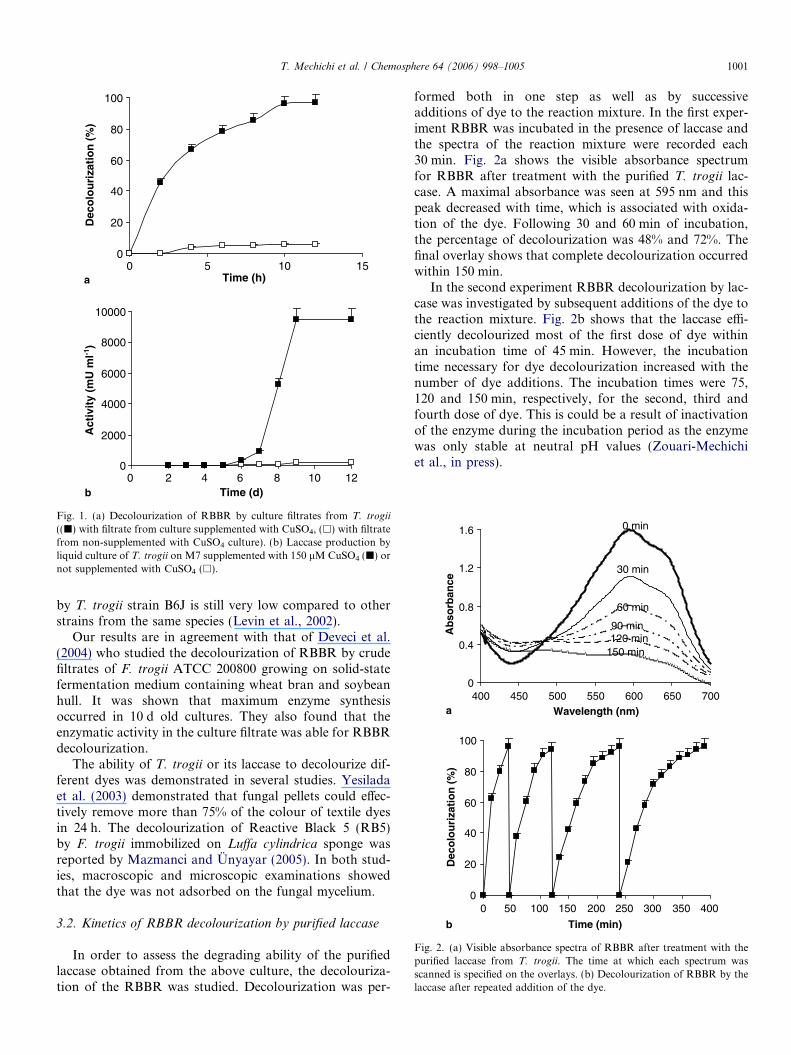
The decolourization of RBBR by the culture filtrate ofT. trogii was performed in 100 ml Erlenmeyer flasks con-taining 25 ml M7 medium. The fungus was grown in cul-tures which were supplemented or not with 150 lM ofCu2+ as laccase inducer. After 12 d of incubation the cul-tures were filtered through Whatman filter paper, RBBRwas injected from a stock solution into the culture filtrates(final concentration 100 mg l�1) and then the OD was mea-sured each hour. Fig. 1a shows that with the filtrate fromthe culture supplemented with copper, the almost completedecolourization of RBBR occurred within 8 h compared toonly 6% of the dye was decolourized using the filtrate fromthe culture without Cu in the same time. This could beexplained by the high titer of laccase produced in the Cu-induced culture, whereas, in the non-induced culture verylow levels were produced, as shown in Fig. 1b. Althoughit produces a high titer of laccase, the laccase production
0
2000
4000
6000
8000
10000
0 2 4 6 8 10 12Time (d)
Act
ivit
y (m
U m
l-1)
0
20
40
60
80
100
0 5 10 15Time (h)
Dec
olo
uri
zati
on
(%
)
a
b
Fig. 1. (a) Decolourization of RBBR by culture filtrates from T. trogii
((j) with filtrate from culture supplemented with CuSO4, (h) with filtratefrom non-supplemented with CuSO4 culture). (b) Laccase production byliquid culture of T. trogii on M7 supplemented with 150 lM CuSO4 (j) ornot supplemented with CuSO4 (h).
0
0.4
0.8
1.2
1.6
400 450 500 550 600 650 700Wavelength (nm)
Ab
sorb
ance
0 min
30 min
60 min
90 min120 min
150 min
0
20
40
60
80
100
0 50 100 150 200 250 300 350 400
Time (min)
Dec
olo
uri
zati
on
(%
)
a
b
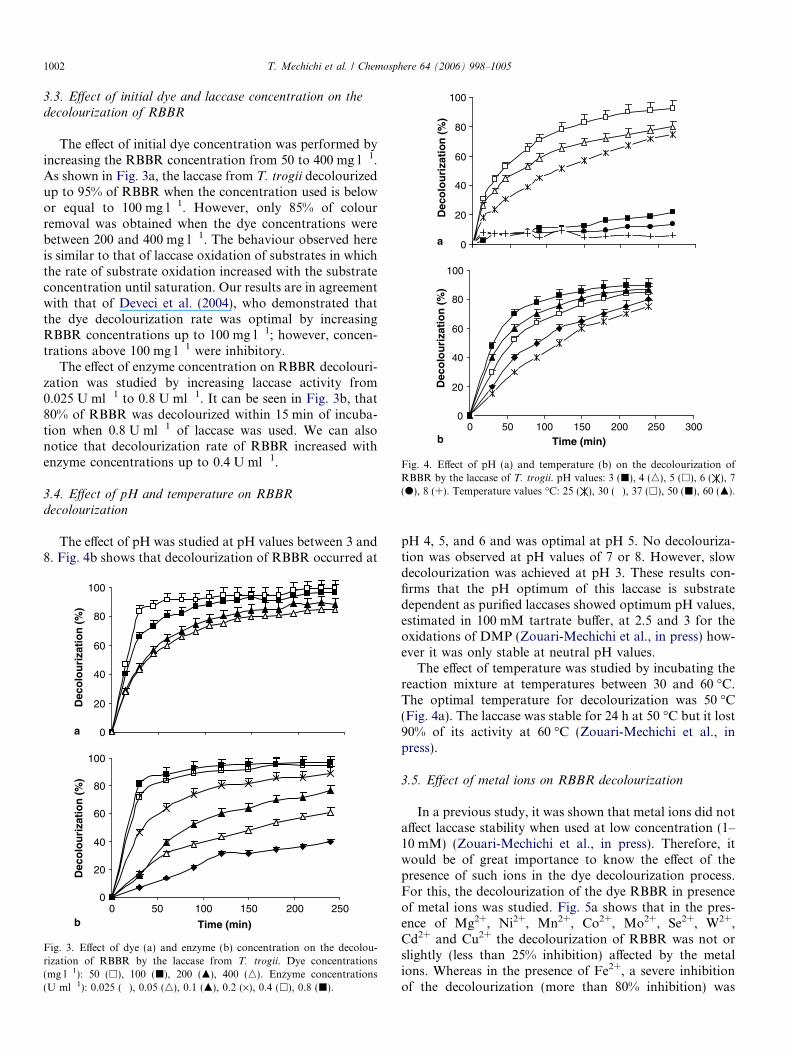
Fig. 2. (a) Visible absorbance spectra of RBBR after treatment with thepurified laccase from T. trogii. The time at which each spectrum wasscanned is specified on the overlays. (b) Decolourization of RBBR by thelaccase after repeated addition of the dye.
T. Mechichi et al. / Chemosphere 64 (2006) 998–1005 1001
by T. trogii strain B6J is still very low compared to otherstrains from the same species (Levin et al., 2002).
Our results are in agreement with that of Deveci et al.(2004) who studied the decolourization of RBBR by crudefiltrates of F. trogii ATCC 200800 growing on solid-statefermentation medium containing wheat bran and soybeanhull. It was shown that maximum enzyme synthesisoccurred in 10 d old cultures. They also found that theenzymatic activity in the culture filtrate was able for RBBRdecolourization.
The ability of T. trogii or its laccase to decolourize dif-ferent dyes was demonstrated in several studies. Yesiladaet al. (2003) demonstrated that fungal pellets could effec-tively remove more than 75% of the colour of textile dyesin 24 h. The decolourization of Reactive Black 5 (RB5)by F. trogii immobilized on Luffa cylindrica sponge wasreported by Mazmanci and Unyayar (2005). In both stud-ies, macroscopic and microscopic examinations showedthat the dye was not adsorbed on the fungal mycelium.
3.2. Kinetics of RBBR decolourization by purified laccase
In order to assess the degrading ability of the purifiedlaccase obtained from the above culture, the decolouriza-tion of the RBBR was studied. Decolourization was per-
formed both in one step as well as by successiveadditions of dye to the reaction mixture. In the first exper-iment RBBR was incubated in the presence of laccase andthe spectra of the reaction mixture were recorded each30 min. Fig. 2a shows the visible absorbance spectrumfor RBBR after treatment with the purified T. trogii lac-case. A maximal absorbance was seen at 595 nm and thispeak decreased with time, which is associated with oxida-tion of the dye. Following 30 and 60 min of incubation,the percentage of decolourization was 48% and 72%. Thefinal overlay shows that complete decolourization occurredwithin 150 min.
In the second experiment RBBR decolourization by lac-case was investigated by subsequent additions of the dye tothe reaction mixture. Fig. 2b shows that the laccase effi-ciently decolourized most of the first dose of dye withinan incubation time of 45 min. However, the incubationtime necessary for dye decolourization increased with thenumber of dye additions. The incubation times were 75,120 and 150 min, respectively, for the second, third andfourth dose of dye. This is could be a result of inactivationof the enzyme during the incubation period as the enzymewas only stable at neutral pH values (Zouari-Mechichiet al., in press).
0
20
40
60
80
100
0 50 100 150 200 250 300
Time (min)D
eco
lou
riza
tio
n (
%)
0
20
40
60
80
100
Dec
olo
uri
zati
on
(%
)b
a
Fig. 4. Effect of pH (a) and temperature (b) on the decolourization ofRBBR by the laccase of T. trogii. pH values: 3 (j), 4 (n), 5 (h), 6 ( ), 7(d), 8 (+). Temperature values �C: 25 ( ), 30 (�), 37 (h), 50 (j), 60 (m).
1002 T. Mechichi et al. / Chemosphere 64 (2006) 998–1005
3.3. Effect of initial dye and laccase concentration on the
decolourization of RBBR
The effect of initial dye concentration was performed byincreasing the RBBR concentration from 50 to 400 mg l�1.As shown in Fig. 3a, the laccase from T. trogii decolourizedup to 95% of RBBR when the concentration used is belowor equal to 100 mg l�1. However, only 85% of colourremoval was obtained when the dye concentrations werebetween 200 and 400 mg l�1. The behaviour observed hereis similar to that of laccase oxidation of substrates in whichthe rate of substrate oxidation increased with the substrateconcentration until saturation. Our results are in agreementwith that of Deveci et al. (2004), who demonstrated thatthe dye decolourization rate was optimal by increasingRBBR concentrations up to 100 mg l�1; however, concen-trations above 100 mg l�1 were inhibitory.
The effect of enzyme concentration on RBBR decolouri-zation was studied by increasing laccase activity from0.025 U ml�1 to 0.8 U ml�1. It can be seen in Fig. 3b, that80% of RBBR was decolourized within 15 min of incuba-tion when 0.8 U ml�1 of laccase was used. We can alsonotice that decolourization rate of RBBR increased withenzyme concentrations up to 0.4 U ml�1.
3.4. Effect of pH and temperature on RBBR
decolourization
The effect of pH was studied at pH values between 3 and8. Fig. 4b shows that decolourization of RBBR occurred at
0
20
40
60
80
100
Dec
olo
uri
zati
on
(%
)
0
20
40
60
80
100
0 50 100 150 200 250
Time (min)
Dec
olo
uri
zati
on
(%
)
a
b
Fig. 3. Effect of dye (a) and enzyme (b) concentration on the decolou-rization of RBBR by the laccase from T. trogii. Dye concentrations(mg l�1): 50 (h), 100 (j), 200 (m), 400 (n). Enzyme concentrations(U ml�1): 0.025 (�), 0.05 (n), 0.1 (m), 0.2 (·), 0.4 (h), 0.8 (j).
pH 4, 5, and 6 and was optimal at pH 5. No decolouriza-tion was observed at pH values of 7 or 8. However, slowdecolourization was achieved at pH 3. These results con-firms that the pH optimum of this laccase is substratedependent as purified laccases showed optimum pH values,estimated in 100 mM tartrate buffer, at 2.5 and 3 for theoxidations of DMP (Zouari-Mechichi et al., in press) how-ever it was only stable at neutral pH values.
The effect of temperature was studied by incubating thereaction mixture at temperatures between 30 and 60 �C.The optimal temperature for decolourization was 50 �C(Fig. 4a). The laccase was stable for 24 h at 50 �C but it lost90% of its activity at 60 �C (Zouari-Mechichi et al., inpress).
3.5. Effect of metal ions on RBBR decolourization
In a previous study, it was shown that metal ions did notaffect laccase stability when used at low concentration (1–10 mM) (Zouari-Mechichi et al., in press). Therefore, itwould be of great importance to know the effect of thepresence of such ions in the dye decolourization process.For this, the decolourization of the dye RBBR in presenceof metal ions was studied. Fig. 5a shows that in the pres-ence of Mg2+, Ni2+, Mn2+, Co2+, Mo2+, Se2+, W2+,Cd2+ and Cu2+ the decolourization of RBBR was not orslightly (less than 25% inhibition) affected by the metalions. Whereas in the presence of Fe2+, a severe inhibitionof the decolourization (more than 80% inhibition) was

0
20
40
60
80
100 D
eco
lou
riza
tio
n (
%)
0
20
40
60
80
100
Dec
olo
uri
zati
on
(%
)
0
20
40
60
80
100
0 50 100 150 200 250Time (min)
Dec
olo
uri
zati
on
(%
)
c
b
a
Fig. 5. (a) Decolourization of RBBR in the presence of metal ions: control(�), Mg (h), Se (n), Fe (s), Ni ( ), Mn (d), Co (+), Mo (�), W (�), Cd(m), Cu (j). (b) Effect of HBT on the decolourization of RBBR: control(h), 5 mM (j). (c) Effect of methanol concentration on the decolouriza-tion of RBBR by the laccase from T. trogii: control (j), 10% (h), 20%(m), 30% (n), 40% ( ).
Table 1Decolourization of RBBR in the presence of aromatic compounds
Aromatic compounds (1 mM) Effect on decolourization
Catechol �Caffeate +Syringate +Vanilline +Gallate �Protocatechuate �Vanillate +Ferulate �Pyrogallol �3,4-Dihydroxy-phenylacetate �HBT +
(�): Inhibition of RBBR decolourization.(+): No effect on RBBR decolourization.
T. Mechichi et al. / Chemosphere 64 (2006) 998–1005 1003
observed. This could be explained by the instability of thelaccase in the presence of Fe2+ at high concentration.Rodriguez Couto et al. (2005) has demonstrated that thelaccase from T. hirsuta was stable at a concentration of1 mM for 7 d in the presence of all the metal ions testedexcept for Zn2+, CrO2�
4 , Cd2+, Cr2O2�7 , Fe2+, Cu2+ and
especially Hg2+, respectively. When the concentrationwas increased to 10 mM, laccase stability decreased in thepresence of all the metals assayed, in particular againstFe2+.
3.6. Effect of aromatic compounds and HBT on RBBR
decolourization
Dyes contain aromatic compounds in their structurewhich can be released after microbial or chemical attacks
during dye degradation. In addition phenolic compoundswere shown to be efficient laccase mediators (Camareroet al., 2005). The effect of aromatic compounds on thedecolourization of RBBR was tested at increasing con-centrations, namely 0.1, 1 and 10 mM. At a concentrationof 0.1 mM no inhibition or activation on the RBBRdecolourization was detected. It can be seen in Table 1that at a concentration of 1 mM several aromatic com-pounds such as catechol, protocatechuate, caffeate and3,4-dihydroxyphenylacetate showed an inhibition of dyedecolourization. All these compounds possess two hydro-xyl groups in ortho position to each other. This suggeststhat ortho-diphenols are inhibiting for the decolourization,which could be due to the higher specificity of laccase tothese substrates than RBBR. When aromatic compoundswere used at a concentration of 10 mM they all have showninhibition of decolourization.
Laccases are copper-containing enzymes that catalysethe oxidation of electron-rich substrates such as phenols.Laccase alone has a limited effect on bioremediation dueto its specificity for phenolic subunits in lignin. However,recently Camarero et al. (2005) have demonstrated thatphenolic aldehydes, ketones, acids, and esters related tothe three lignin units were among the best mediators,including p-coumaric acid, vanillin, acetovanillone, methylvanillate, and above all, syringaldehyde and aceto-syringone.
The effect of HBT on RBBR decolourization was stud-ied at increasing concentrations from 0.125 to 5 mM. Asshown in Fig. 5b no effect of the laccase mediator wasobserved at HBT concentrations between 0.125 and2.5 mM, however a concentration of 5 mM showed slightinhibition of RBBR decolourization. This could beexplained by the fact that anthraquinone dyes such asRBBR act as enzyme substrates that are directly oxidizedby laccase while decolourization of azo and indigo dyesinvolves some small molecule metabolites like HBT asmediators. On the other hand, the inhibition of the deco-lourization could be due to a toxic effect of the high con-centration of the nitroxide radical resulting from laccaseoxidation of HBT.

0
20
40
60
80
100
120
EDTA SDS CysteineLaccase inhibitors
Inh
ibit
ion
(%
)
0.1 mM
1 mM
10 mM
Fig. 6. Effect of laccase inhibitors on the decolourization of RBBR by thelaccase from T. trogii.
1004 T. Mechichi et al. / Chemosphere 64 (2006) 998–1005
These findings differ from those reported for other lac-cases by Soares et al. (2001b) who demonstrated that apure fungal laccase, obtained from a commercial formula-tion used in the textile industry, did not decolourize RBBR.Decolourization was only observed when a small molecularweight redox mediator (violuric acid or HBT) was addedtogether with the laccase. In contrast higher concentrationsof HBT were inhibitory which could be due to inactivationof laccase by the toxic HBT radical.
3.7. Effect of methanol concentration and laccase
inhibitors on RBBR decolourization
The effect of methanol was tested at concentrationsranging from 10% to 40%. It can be seen in Fig. 5c thatthe presence of methanol in the reaction mixture contain-ing laccase and RBBR had a negative effect on decolouriza-tion. The inhibition of the decolourization increased withmethanol concentration. The effect of organic solvents onlaccase activity has been reported by several authors (Mil-stein et al., 1993; Luterek et al., 1998). In general, both freeand immobilized laccases decrease their activity as the con-centrations of water-miscible organic solvents increase, andin most reported cases, immobilized laccase shows higheractivity than the free enzyme at different organic solventconcentrations.
In order to determine the effect of laccase inhibitors onthe decolourization of RBBR, L-cysteine, SDS and EDTAwere tested at concentrations of 0.1, 1 and 10 mM. Fig. 6shows that EDTA does not inhibit the decolourization ofRBBR up to a concentration of 10 mM. While in the pres-ence of SDS decolourization was inhibited at a 10 mM con-centration but not at 0.1 or 1 mM. Cysteine was the mostinhibiting substance used because at 1 mM 70% of inhibi-tion was reached while complete inhibition was achievedwith 10 mM.
The most common laccase inhibitors used are dithio-threitol, thioglycolic acid, cysteine, diethyldithiocarbamicacid, EDTA, sodium fluoride, and sodium azide (Bollagand Leonowicz, 1984; Slomczynski et al., 1995). Theseinhibitors are not laccase specific and their application on
phenoloxidases originates from results obtained with othermetalloenzymes.
4. Conclusion
In view of the results obtained in the present work, it canbe concluded that laccase from T. trogii is a powerful toolfor the decolourization of textile dyes.
Decolourization of RBBR occurred only with culture fil-trates induced with Cu2+, which suggest that laccase is theenzyme responsible for the decolourization of RBBR. Thisresult was confirmed by the use of purified laccase. Thepurified laccase was found to decolourize efficiently thedye at a concentration of 100 mg l�1 in the presence of0.2 U ml�1 of enzyme. In addition, decolourization wasoptimal at pH 5 and at a temperature of 50 �C.
The effect of metal ions was tested at a concentration of10 mM. It was found that among the metal ions tested,Fe2+ was the most inhibiting for the decolourization. Theeffect of HBT was also assessed. At low concentrations itwas neither necessary, nor had any effect on decolouriza-tion of RBBR while at a concentration of 5 mM, HBTinhibited slightly the decolourization. The presence of aro-matic compounds was found to be inhibiting for the deco-lourization at a concentration of 10 mM but not 0.1 mM,while at 1 mM only ortho-diphenols were inhibiting. Prob-ing the effect of methanol it was found that at increasingconcentrations, methanol caused a decrease in the decolou-rization rate of RBBR.
The effect of laccase inhibitors in the decolourization ofRBBR was tested with L-cysteine, SDS and EDTA at con-centrations of 0.1, 1 and 10 mM. It was demonstrated thatL-cysteine was the most inhibiting substrate for the deco-lourization while SDS was only inhibiting at 10 mM con-centration and ETDA was not inhibiting at all testedconcentrations.
Acknowledgments
The authors thank Stephen J. Molyneaux from WoodsHole Oceanographic Institution, USA, for revising theEnglish and Professor Derek Litthauer from the Universityof the Free State, South Africa for his critical review of thepaper. This work was supported by a grant provided by‘‘Contrats Programmes MRSTCD’’ Tunisia.
References
Bajpai, P., 2004. Biological bleaching of chemical pulps. Crit. Rev.Biotechnol. 24, 1–58.
Bollag, J.M., Leonowicz, A., 1984. Comparative studies of extracellularfungal laccases. Appl. Environ. Microbiol. 48, 849–854.
Call, H.P., Mucke, I., 1997. History, overview and applications ofmediated lignolytic systems, especially laccase-mediator-systems (Lig-nozym process). J. Biotechnol. 53, 163–202.
Camarero, S., Ibarra, D., Martınez, M.J., Martınez, A.T., 2005. Lignin-derived compounds as efficient laccase mediators for decolorization of

T. Mechichi et al. / Chemosphere 64 (2006) 998–1005 1005
different types of recalcitrant dyes. Appl. Environ. Microbiol. 71,1775–1784.
Christian, V., Shrivastava, R., Shukla, D., Modi, H., Vya, B.R.M., 2005.Mediator role of veratryl alcohol in the lignin peroxidase-catalyzedoxidative decolorization of Remazol Brilliant Blue R. Enzyme Microb.Technol. 36, 426–431.
Claus, H., 2003. Laccases and their occurrence in prokaryotes. Arch.Microbiol. 179, 145–150.
Deveci, T., Unyayar, A., Mazmanci, M.A., 2004. Production of RemazolBrilliant Blue R decolourising oxygenase from the culture filtrate ofFunalia trogii ATCC 200800. J. Mol. Catal. B: Enzym. 30, 25–32.
Dhouib, A., Hamza, M., Zouari, H., Mechichi, T., H’midi, R., Labat, M.,Martinez, M.J., Sayadi, S., 2005. Screening for ligninolytic enzymeproduction by diverse fungi from Tunisia W. J. Microbiol. Biotechnol.21, 1514–1523.
Eichlerova, I., Homolka, L., Lisa, L., Nerud, F., 2005. Orange G andRemazol Brilliant Blue R decolorization by white rot fungi Dichomitus
squalens, Ischnoderma resinosum and Pleurotus calyptratus. Chemo-sphere 60, 398–404.
Levin, L., Forchiassin, F., Ramos, A.M., 2002. Copper induction oflignin-modifying enzymes in the white-rot fungus Trametes trogii.Mycologia 94, 377–383.
Luterek, J., Gianfreda, L., Wojtas-Wasilewska, M., Cho, N.S., Rogalski,J., Jaszek, M., Malarczyk, E.B., Staszczak, M., Fink-Boots, M.,Leonowicz, A., 1998. Activity of free and immobilized extracellularCerreno unicolor laccase in water miscible organic solvents. Holzfors-chung 52, 589–595.
Mazmanci, M.A., Unyayar, A., 2005. Decolourisation of Reactive Black 5by Funalia trogii immobilised on Luffa cylindrica sponge. ProcessBiochem. 40, 337–342.
Milstein, O., Huttermann, A., Majcherczyk, A., Schulze, K., Fruend, R.,Luedermann, H.D., 1993. Transformation of lignin-related com-pounds with laccase in organic solvents. J. Biotechnol. 30, 37–47.
Minussi, R.C., Pastore, G.M., Duran, N., 2002. Potential applicationsof laccase in the food industry. Trends Food Sci. Technol. 13, 205–216.
Palmieri, G., Cennamo, G., Sannia, G., 2005. Remazol Brilliant Blue Rdecolourisation by the fungus Pleurotus ostreatus and its oxidativeenzymatic system. Enzyme Microb. Technol. 36, 17–24.
Peralta-Zamora, P., Pereira, C.M., Tiburtius, E.R.L., Moraes, S.G., Rosa,M.A., Minussi, R.C., Duran, N., 2003. Decolorization of reactive dyesby immobilized laccase. Appl. Catal. B: Environ. 42, 131–144.
Rodriguez Couto, S., Sanroman, M., Gubitz, G.M., 2005. Influence ofredox mediators and metal ions on synthetic acid dye decolourizationby crude laccase from Trametes hirsuta. Chemosphere 58, 417–422.
Slomczynski, D., Nakas, J.P., Tanenbaum, W., 1995. Production andcharacterization of laccase from Botrytis cinerea 61-34. Appl. Environ.Microbiol. 61, 907–912.
Soares, G.M.B., Costa-Ferreira, M., de Amorim, M.T.P., 2001a. Decol-orization of an anthraquinone-type dye using a laccase formulation.Bioresour. Technol. 79, 171–177.
Soares, G.M., de Amorim, M.T., Costa-Ferreira, M., 2001b. Use oflaccase together with redox mediators to decolourize Remazol BrilliantBlue R. J. Biotechnol. 89, 123–129.
Solomon, E.I., Sundaram, U.M., Machonkin, T.E., 1996. Multicopperoxidases and oxygenases. Chem. Rev. 96, 2563–2605.
Wesenberg, D., Kyriakides, I., Agathos, S.N., 2003. White-rot fungi andtheir enzymes for the treatment of industrial dye effluents. Biotechnol.Adv. 22, 161–187.
Yesilada, O., Asma, D., Cing, S., 2003. Decolourization of textile dyes byfungal pellets. Process Biochem. 38, 933–938.
Zouari-Mechichi, H., Mechichi, T., Dhouib, A., Sayadi, S., Martınez,A.T., Martınez, M.J., in press. Purification and characterizationof laccases from a Trametes trogii strain isolated in Tunisia. Decol-orization of textile dyes by the purified enzyme. Enzyme Microb.Technol.




















