
Laboratory-Induced Endolithic Growth in Calcarenites: Biodeteriorating Potential Assessment
14
ENVIRONMENTAL MICROBIOLOGY Laboratory-Induced Endolithic Growth in Calcarenites: Biodeteriorating Potential Assessment A. Z. Miller & M. A. Rogerio-Candelera & L. Laiz & J. Wierzchos & C. Ascaso & M. A. Sequeira Braga & M. Hernández-Mariné & A. Maurício & A. Dionísio & M. F. Macedo & C. Saiz-Jimenez Received: 16 September 2009 / Accepted: 10 February 2010 / Published online: 4 May 2010 # Springer Science+Business Media, LLC 2010 Abstract This study is aimed to assess the formation of photosynthetic biofilms on and within different natural stone materials, and to analyse their biogeophysical and biogeochemical deterioration potential. This was performed by means of artificial colonisation under laboratory conditions during 3 months. Monitoring of microbial development was performed by image analysis and biofilm biomass estimation by chlorophyll extraction technique. Microscopy investigations were carried out to study relationships between microorganisms and the mineral substrata. The model applied in this work corroborated a successful survival strategy inside endolithic microhabitat, using natural phototrophic biofilm cultivation, composed by cyanobacteria and algae, which increased intrinsic porosity by active mineral dissolution. We observed the presence of mineral-like iron derivatives (e.g. maghemite) around the cells and intracellularly and the precipitation of hausmannite, suggesting manganese transformations related to the biomineralisation. Introduction There is a growing interest in the microbial ecology of phototrophic microorganisms on rock surfaces [4, 13, 33, 42]. Photoautotrophic microorganisms are primary producers that include cyanobacteria, green algae and diatoms and commonly inhabit alkaline rather than acid siliceous rocks [2]. Laboratory experiments on stone bioreceptivity have demonstrated that carbonate rocks are extremely suscep- A. Z. Miller Departamento de Conservação e Restauro, Faculdade de Ciências e Tecnologia, Universidade Nova de Lisboa, Monte de Caparica, 2829-516 Caparica, Portugal A. Z. Miller (*) : A. Maurício : A. Dionísio Centro de Petrologia e Geoquímica, Instituto Superior Técnico, Av. Rovisco Pais, 1049-001 Lisbon, Portugal e-mail: [email protected] M. A. Rogerio-Candelera : L. Laiz : C. Saiz-Jimenez Instituto de Recursos Naturales y Agrobiologia, CSIC, Av. Reina Mercedes 10, 41012 Sevilla, Spain J. Wierzchos : C. Ascaso Instituto de Recursos Naturales, Centro de Ciencias Medioambientales, CSIC, Serrano, 115 bis, 28006 Madrid, Spain M. A. Sequeira Braga Centro de Investigação Geológica, Ordenamento e Valorização de Recursos (CIG-R), Universidade do Minho, Campus de Gualtar, 4710-057 Braga, Portugal M. Hernández-Mariné Facultat de Farmacia, Universitat de Barcelona, Av. Joan XXIII, s/n, 08028 Barcelona, Spain M. F. Macedo Vicarte, Faculdade de Ciências e Tecnologia, Universidade Nova de Lisboa, Monte de Caparica, 2829-516 Caparica, Portugal Microb Ecol (2010) 60:55–68 DOI 10.1007/s00248-010-9666-x
Transcript of Laboratory-Induced Endolithic Growth in Calcarenites: Biodeteriorating Potential Assessment

ENVIRONMENTAL MICROBIOLOGY
Laboratory-Induced Endolithic Growth in Calcarenites:Biodeteriorating Potential Assessment
A. Z. Miller & M. A. Rogerio-Candelera & L. Laiz & J. Wierzchos & C. Ascaso &
M. A. Sequeira Braga & M. Hernández-Mariné & A. Maurício & A. Dionísio &
M. F. Macedo & C. Saiz-Jimenez
Received: 16 September 2009 /Accepted: 10 February 2010 /Published online: 4 May 2010# Springer Science+Business Media, LLC 2010
Abstract This study is aimed to assess the formation ofphotosynthetic biofilms on and within different naturalstone materials, and to analyse their biogeophysical andbiogeochemical deterioration potential. This was performedby means of artificial colonisation under laboratoryconditions during 3 months. Monitoring of microbialdevelopment was performed by image analysis and biofilmbiomass estimation by chlorophyll extraction technique.Microscopy investigations were carried out to studyrelationships between microorganisms and the mineralsubstrata. The model applied in this work corroborated asuccessful survival strategy inside endolithic microhabitat,using natural phototrophic biofilm cultivation, composedby cyanobacteria and algae, which increased intrinsicporosity by active mineral dissolution. We observed the
presence of mineral-like iron derivatives (e.g. maghemite)around the cells and intracellularly and the precipitation ofhausmannite, suggesting manganese transformations relatedto the biomineralisation.
Introduction
There is a growing interest in the microbial ecology ofphototrophic microorganisms on rock surfaces [4, 13, 33, 42].Photoautotrophic microorganisms are primary producersthat include cyanobacteria, green algae and diatoms andcommonly inhabit alkaline rather than acid siliceous rocks[2]. Laboratory experiments on stone bioreceptivity havedemonstrated that carbonate rocks are extremely suscep-
A. Z. MillerDepartamento de Conservação e Restauro,Faculdade de Ciências e Tecnologia,Universidade Nova de Lisboa,Monte de Caparica,2829-516 Caparica, Portugal
A. Z. Miller (*) :A. Maurício :A. DionísioCentro de Petrologia e Geoquímica, Instituto Superior Técnico,Av. Rovisco Pais,1049-001 Lisbon, Portugale-mail: [email protected]
M. A. Rogerio-Candelera : L. Laiz : C. Saiz-JimenezInstituto de Recursos Naturales y Agrobiologia, CSIC,Av. Reina Mercedes 10,41012 Sevilla, Spain
J. Wierzchos : C. AscasoInstituto de Recursos Naturales,Centro de Ciencias Medioambientales, CSIC,Serrano, 115 bis,28006 Madrid, Spain
M. A. Sequeira BragaCentro de Investigação Geológica,Ordenamento e Valorização de Recursos (CIG-R),Universidade do Minho,Campus de Gualtar,4710-057 Braga, Portugal
M. Hernández-MarinéFacultat de Farmacia, Universitat de Barcelona,Av. Joan XXIII, s/n,08028 Barcelona, Spain
M. F. MacedoVicarte, Faculdade de Ciências e Tecnologia,Universidade Nova de Lisboa,Monte de Caparica,2829-516 Caparica, Portugal
Microb Ecol (2010) 60:55–68DOI 10.1007/s00248-010-9666-x

tible to colonisation by photoautotrophs [21, 29, 30].These studies provide useful information about microbialbiodiversity, their distribution patterns, and the relationshipsamong the different populations and the mineral substrata. Ithas been stated that mineralogy, porosity, surface roughnessand permeability control rock bioreceptivity [20].
Phototrophic microorganisms develop forming biofilmson the rock surface composed of a mono- or multilayer ofcells embedded in a hydrated extracellular polymericmatrix which hold the cells together and to the surface.It may also contain exoenzymes and inorganic inclusionssuch as clay particles and minerals [47]. In such biofilmsmicroorganisms use structural irregularities of the rockmaterial for their attachment and growth, which isstimulated in the presence of locally enhanced moistureavailability in rough or porous surfaces, cracks andfissures [1].
According to the distribution pattern, the microorganismsthat inhabit the rocks have been classified as epilithic andendolithic [18]. Epilithic microorganisms grow on theexternal surface of the rock, whereas endolithic micro-organisms live in the interior of rocks, penetrating somemillimetres into the porous system. They can survive ininhospitable natural habitats, such as hot deserts and semiaridlands [7] or cold deserts [17], but also in cultural heritageassets such as vertical surfaces of monuments [2, 3, 28, 33,39, 41, 43]. The endolithic microhabitat gives protectionfrom intense solar radiation and desiccation, and it providesmineral nutrients, moisture and growth surfaces [45].
The proliferation of microorganisms under the rocksurface can increase their biodeteriorating potential,contributing to decreasing of grain cohesion to depths upto several millimetres [1, 3, 40]. Field observations andexperiments in the laboratory have demonstrated thesignificant potential for damage by phototrophic organisms[8, 10, 43]. Laboratory-based stone colonisation provides avaluable alternative for natural ecological niches by allowingexperimental manipulation of the microbial ecosystem.
Considering that little if any attempt appear to have beenmade to the study of endolithic growth under laboratoryconditions, this study is aimed to assess the formation ofphotosynthetic biofilms on and within different naturalstone materials, and to analyse their biogeophysical andbiogeochemical deterioration potential.
Materials and Methods
Investigated Stone Materials
Two different calcarenites, comprising natural stoneswidely used in the construction of Southern Europeanmonuments, were acquired from quarries of Cadiz and
Granada provinces (Andalusia, Spain). Fresh cut quarrystone samples of San Cristobal and Escúzar stones werestudied.
San Cristobal lithotype (SC) is an Upper Mioceneyellowish bioclastic calcarenite with an inequigranularheterogeneous texture and a very scarce matrix with lowcementation grade causing a high intergranular porosity.
Escúzar lithotype (ES) is a light-coloured fine-to-medium grained biocalcarenite, very soft and porous fromthe Tortonian age.
The differences in the petrographic and petrophysiccharacteristics of these lithotypes were described in aprevious work [31].
Inoculation and Growth Conditions
Replicate samples of each lithotype (∅ 4.4×2 cm) were cut,using a diamond blade saw and sterilised in autoclave at120°C and 1 atm, for 20 min. The upper surface of thestone samples of each lithotype was inoculated with0.75 ml of a phototrophic community culture, composedmainly by cyanobacteria and algae. The major componentsof the inoculum were Chlorella, Stichococcus, Trebouxiaand Myrmecia among the Chlorophyta and Leptolyngbyaand Pleuroclapsa among the Cyanobacteria. This photo-trophic community was previously collected from Ançãlimestone surfaces of Santa Clara-a-Velha Monastery,Coimbra (Portugal), characterised by molecular techniquesand cultivated in liquid BG11 medium. This multi-speciesculture was used as inoculum for biofilm development onstone samples under laboratory conditions. Further detailsof the characterisation and cultivation procedure of thenatural phototrophic biofilm have been provided by Milleret al. [32].
The samples were incubated for 3 months in a non-commercial incubator system (with overall dimensions of100×60×60 cm) containing 0.5 cm height of sterile waterin the bottom of the chamber. Photosynthesis-inducingfluorescent lamps (Fluora, Osram) were installed below thechamber cover to provide 12 h dark/light (1,200 lx) cyclesfor the growth of cyanobacteria and algae. The experimentwas conducted at room temperature of 20±2°C. Moistureinside the chamber was kept by water circulation in thechamber bottom, fed by a water pump. Sterile distilledwater was periodically added to soak the samples with0.5 cm water height.
Monitoring of Lithotypes Phototrophic Growth
Image Analysis
Image analysis was applied to quantify the covered surfacearea evolution on the stone samples spanning the incuba-
56 A. Z. Miller et al.

tion time. Samples were taken out of the incubator chamberand placed on paper towels to allow excess water to drainout. Subsequently, the samples were placed on millimetricpaper under controlled light to ensure fixed conditions forall photographic records. The photographic recording intriplicates of each lithotype was performed weekly witha digital camera (Kodak EasyShare P850). Since thecolour of the lithotypes masked the possible biofilmgrowth, conventional RGB images obtained with thedigital camera were decorrelated by principal componentanalysis (PCA). The PCA was used to simplify imagesavoiding redundant data present in the different bands ofthe image. This approach allowed choosing the mostappropriate resulting band for the measurements fromthe minority principal components. This analysis strategyhas been previously utilised to improve the visualisationof rock art paintings in highly correlated images [36],and to record separately different elements (of differentnature and composition) present in mural paintings[37]. PCA was performed utilising HyperCube v. 9.5software (US Army Topographic Engineering Centre,Alexandria, VA, USA). Once the geometry of the photo-graphs was digitally rectified with Adobe Photoshop©software, a thresholding algorithm was applied in order toselect the colonised areas, afterwards scaled and measuredusing ImageJ software (National Institutes of Health,Bethesda, MD, USA) following the protocol developedin Rogerio-Candelera et al. [38]. The obtained photo-graphic series, geometrically coherent and comparable,allowed obtaining a series of numerical values related tobiofilm extent.
Chlorophyll Extraction Method
Biofilm growth was also monitored by the increase ofphotosynthetic biomass achieved by chlorophyll extrac-tion method with subsequent UV/Vis spectrophotometryin order to quantify the complete amount of chlorophylla present on and within the stone samples. Every month,three replicate of each lithotype were taken out of theincubator chamber for total chlorophyll a content analysis.Chlorophyll a was extracted by crushing each stonesample into fragments (0.20–0.50 cm3) which were thenadded to 50 ml of dimethyl-sulphoxide and heated to 65°Cfor 1 h, according to the pigment extraction protocol ofVollernweider et al. [44]. The samples were filtered toremove stone particles and absorbancies of the extractswere measured at 664 and 750 nm before and afteracidification with 1 N HCl, in a Perkin Elmer Lambda 35UV/Vis spectrophotometer. The equation of Lorenzen [27]was used to calculate chlorophyll a concentrations.
Linear regression statistics was used to compare thechlorophyll a content data and image analysis data.
Analyses of Endolithic Growth After 3 Monthsof Incubation
Image Analysis
Digital images from sections perpendicular to the inocu-lated stone surfaces were also used in image analysis.RGB images were generated from binocular stereomicroscopy (Zeiss Discover V8 with phototube) andphotographically recorded with a digital camera (CanonPowershot A630). The distribution and penetration ofphotosynthetic biomass in the stone substrata was depictedby means of PCA. This approach allows the detection ofminority elements (of different nature and composition)apparently absent in the RGB digital image but masked bythe redundant data registered in the red, green and bluebands of the image (PC1, PC2 and PC3 bands, respec-tively). As the analysis allows the reduction of the dataset,the redundant information can be securely avoided. Thisdecorrelation allowed choosing the most appropriate PCAband (PC1, PC2 or PC3) which improved the visualisationof the photosynthetic biofilm inside the stone samples.Thus, PC1 plotting more than 95% of the total informationincluded in the three bands of each image, was used asbackground to appreciate the microtopography of thesample, and PC3 band (plotting around 0.5% of the totalinformation of the original image) were used to distin-guish the lesser amounts of information included in theimages, but relevant for the objectives of the work. For abetter visualisation of the decorrelated images reflectingminority principal components, false colours were applied,as human eye can distinguish better colour differencesthan grey tones. As in the case of the quoted above surfacemeasurements of the biofilm extent, PCA and false-colourimages were elaborated using HyperCube 9.5 software.
Scanning Electron Microscopy
Replicate stone samples of each lithotype were examinedby scanning electron microscopy with back-scatteredelectron imaging (SEM-BSE) and an X-ray energydispersive spectroscopy (EDS), at Centro de CienciasMedioambientales (CSIC—Spain), according to a methoddeveloped by Wierzchos and Ascaso [46]. The inoculatedsamples were investigated as stone fragments coated withgold and as resin-casts. The later consisted of stonefragments perpendicularly cut to the colonised surfacesand fixed with 3.25% glutaraldehyde followed by 1%OsO4. After fixing, the samples were dehydrated in seriesof ethanol solutions, embedded in epoxy resin and fine-polished after polymerisation. The resin-impregnatedcross-sections of colonised stone samples, previouslycarbon coated, were examined using a DMS 940A Zeiss
Endolithic Growth in Calcarenites 57

scanning electron microscope in BSE mode. Chemicalanalyses by EDS were simultaneously conducted. Themicroscope operating conditions were: 0-degree tilt angle,35-degree X-ray take-off angle, 15 kV acceleration potential,25 mm of working distance and 1–5 nA specimen currentrange.
Combined and information supplied by both methods wasused to characterise the distribution pattern of the photosyn-thetic microorganisms on and/or inside stones, and to evaluatetheir interrelations and interactions with the substrata.
Biocalcarenite Mineralogical and MicromorphologicalCharacterisation
X-ray Diffraction
Different analysis were carried out on ES samples (beforeand after inoculation) in order to identify the mineralogicalcomposition, in particular, iron oxides–oxyhydroxidesdetected by SEM-EDS.
The mineralogy was determined by a Philips PW1710(APD-version 3.6 j) diffractometer using CuKα radiation at40 kV and 30 mA. The step size was 0.02° 2θ and thecounting time was 1.25 s. Analytical software, “X’PertGraphics” and “Identify” by Philips, were used. The bulkmineralogy was determined in random powders of ESsamples before and after the colonisation experiment.Regarding the estimations of mineral phases in the bulkrock, the peak-height intensities for diagnostic reflectionswere used: calcite—3.03 Å; quartz—3.34 Å; mica—10 Å;clay minerals—4.43–4.48 Å; siderite—2.79 Å; goethite—4.18 Å; maghemite—2.51 Å; pyrolusite—3.11–3.16 Å; andhausmannite—2.49 Å.
SEM-EDS Analysis
This study was also carried out on the inoculated ES stonefragments. The samples were examined at the CIG-R,Universidade do Minho (Portugal), by optical microscopyand SEM-EDS for chemical analysis, using secondaryX-rays and standard ZAF corrections that allow semi-quantitative microanalyses and characterisation of miner-alogical phases. These analyses were carried out with aLEICA Cambridge S360 microscope, and the sampleswere coated with a high conductance thin film (gold film).
Results
Monitoring of Lithotypes Phototrophic Growth
For the evaluation of the colonisation process during theincubation time span, two different strategies were imple-
mented: (1) the measurement of areas covered by thebiofilm by means of image analysis and (2) the quantifica-tion of photosynthetic biomass by chlorophyll extractionmethod. Image analysis was used as a non-destructivemethod to measure the surface covered area since thechlorophyll extraction method is not suitable for takingsuccessive measurements of photosynthetic biomass on asample.
The distribution pattern of the biofilms on the stonesurfaces was influenced by petrophysical properties of SCand ES lithotypes. During the inoculation procedure, theinoculum (photosynthetic liquid culture) was rapidlyabsorbed by both lithic substrata, penetrating into theirpore system due to large pore size and surface roughness. Inspite of the difficulties for measuring the extent of theepilithic phototrophic biofilm, strongly masked by the highmacroporosity of the lithotypes, the image analysisapproach allowed its quantification. On the surface of theSC samples, the inoculum originally green resulted in abrownish colour biofilm, indicating apparent cessation ofepilithic colonisation after the incubation. Therefore, the totalsurface area covered by the phototrophic biofilm from theseries of triplicate digital photographs showed no significantincrease for SC lithotype over the course of batch incubation.Moreover, a short decrease in the biomass was registered, butthe total covered area was strongly similar, as well as thedistribution of colony sizes (Fig. 1).
The surface covered by the epilithic biofilm on ES sampleswas greater than after inoculation, showing a brownish-greenbiofilm with a moderate growth. This growth was alsoassessed by means of the colony size distribution 1 weekafter the inoculation and after the 3-month colonisationexperiment, as shown in Fig. 2. It is noticeable the epilithicgrowth registered for this lithotype.
The monthly chlorophyll a content, estimated using thechlorophyll extraction method followed by spectrophotome-try, reflected a complete amount of this pigment on andwithin the stone samples. The monthly progress of photo-synthetic growth, expressed as micrograms of chlorophyll aper cubic centimetre, is depicted in Fig. 3. During 3 months,ES presented a steady increase of photosynthetic biomassindicating a successful colonisation. SC photosyntheticbiomass showed stability during the first 2 months ofincubation, experimenting an important increase at the thirdmonth of incubation.
A comparison of the chlorophyll a content and imageanalysis results is presented in Fig. 4. The data for eachmethod are shown as a scatter diagram to illustrate thecorrelation between both monitoring techniques. A fairlygood linear positive experimental correlation was observedfor ES lithotype under the study conditions, whichcompared to SC lithotype, a much lower experimentalcorrelation was obtained.
58 A. Z. Miller et al.

Lithotypes Colonisation by Endolithic Biofilms
Image Analysis
Image analysis by means of pixel value decorrelation wasapplied for the visual enhancement of the presence ofendolithic microorganisms, allowing the quantification ofpenetration depth, and the assessment of the endolithicbiomass distribution into the substrata (Fig. 5). When thestone samples were longitudinally cut after the incubationperiod, some green stains were visible to the naked eye.PCA of optical microscopy images of the cut samplesshowed enhanced stains in both lithotypes, particularly inSC samples. A thin horizontal band with characteristicreflectance differences was observed parallel to the stonesurface (Fig. 5b). These bands and stains, sometimescoinciding with the green stains of the original images,were preliminarily attributed to phototrophic endolithiccolonisation. False-colour images composed of the minorityPC bands helped in distinguishing the extent of theassumed endolithic growth. The detected band ranged from1 to 3 mm in thickness, with a maximum penetration depthof 3 mm. The distribution of the subsurface biofilm wasfairly tabular and strongly parallel to the surface.
Considering ES samples (Fig. 5c, d), the stains weremore discontinuous, in what seems to follow the porous
system of the stone, with penetration depths up to 3 mm.The appearance of this biofilm was more branched andconnected to the outer epilithic biofilm through the poresystem.
Scanning Electron Microscopy
SEM examination of sections perpendicular to the stonesurface enabled studying the distribution of microorganisms,their development on the subsurface of the samples and theirrelationship with the substrata. SEM analyses of SC and ESsamples confirmed they were colonised endolithically byphototrophic microorganisms. The colonisation density,penetration depth and biomass per unit area, varied.
Considering ES samples, interaction between the inocu-lated phototrophic microorganisms and the substrata wasobserved. It was particularly notable that endolithic micro-organisms actively create small cavities easily penetrating thecalcite crystals (Fig. 6). Figure 7 confirmed the existence ofendolithic microorganisms in the ES samples, as well asepilithic microorganisms on the stone surface (Fig. 7a). Thelikely depths endolithic cells penetrated the stone through theaccessible void spaces. The cells were mainly concentratedin the bottle neck shaped parts of it (200 µm). Beyond thatpart, only a few of them reached approximately 200 µmmore (Fig. 7a). The cells within the epilithic biofilm were
Figure 1 Image analysis results of one representative stone sample ofSan Cristobal lithotype. a Simplified images (PC2) immediately afterinoculation and b after 3 months. Whitish areas comprise the
phototrophic biofilm. c Total colonised surface area immediately afterinoculation (T0) and after 12 weeks (T12). d Colony size distributionafter inoculation and e after 12 weeks
Endolithic Growth in Calcarenites 59
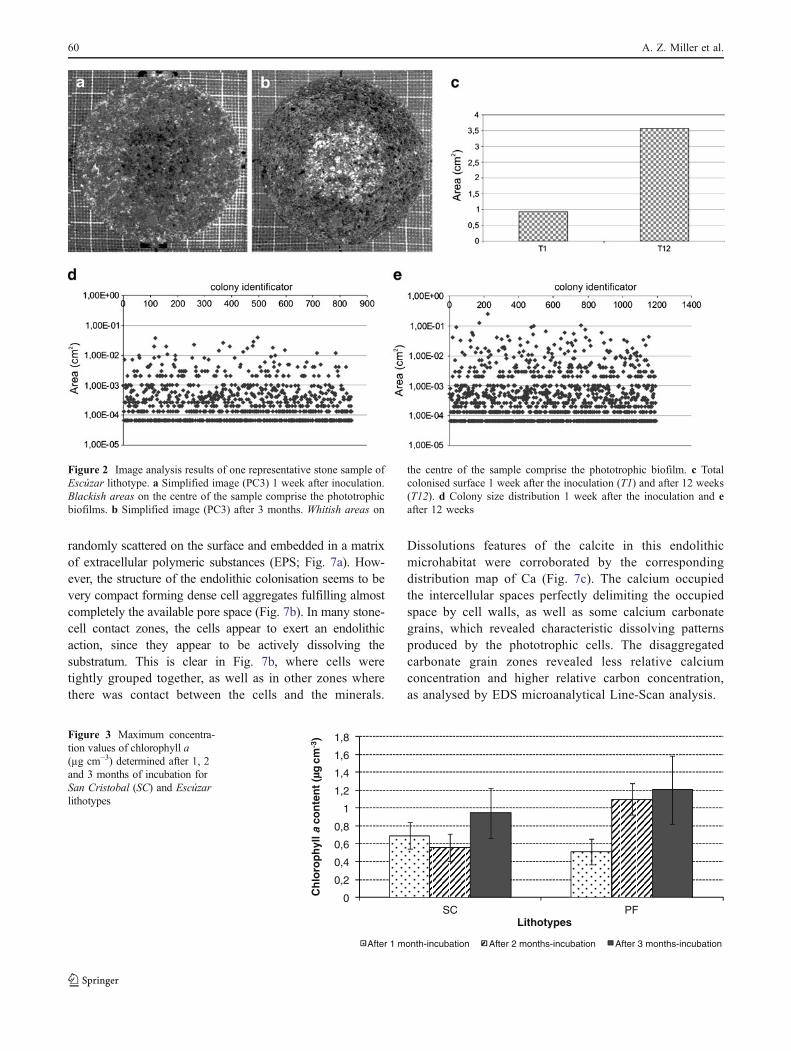
randomly scattered on the surface and embedded in a matrixof extracellular polymeric substances (EPS; Fig. 7a). How-ever, the structure of the endolithic colonisation seems to bevery compact forming dense cell aggregates fulfilling almostcompletely the available pore space (Fig. 7b). In many stone-cell contact zones, the cells appear to exert an endolithicaction, since they appear to be actively dissolving thesubstratum. This is clear in Fig. 7b, where cells weretightly grouped together, as well as in other zones wherethere was contact between the cells and the minerals.
Dissolutions features of the calcite in this endolithicmicrohabitat were corroborated by the correspondingdistribution map of Ca (Fig. 7c). The calcium occupiedthe intercellular spaces perfectly delimiting the occupiedspace by cell walls, as well as some calcium carbonategrains, which revealed characteristic dissolving patternsproduced by the phototrophic cells. The disaggregatedcarbonate grain zones revealed less relative calciumconcentration and higher relative carbon concentration,as analysed by EDS microanalytical Line-Scan analysis.
Figure 2 Image analysis results of one representative stone sample ofEscúzar lithotype. a Simplified image (PC3) 1 week after inoculation.Blackish areas on the centre of the sample comprise the phototrophicbiofilms. b Simplified image (PC3) after 3 months. Whitish areas on
the centre of the sample comprise the phototrophic biofilm. c Totalcolonised surface 1 week after the inoculation (T1) and after 12 weeks(T12). d Colony size distribution 1 week after the inoculation and eafter 12 weeks
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
1,8
SC PF
Ch
loro
ph
yll a
co
nte
nt
( g
-3)
Lithotypes
After 1 month-incubation After 2 months-incubation After 3 months-incubation
cm
Figure 3 Maximum concentra-tion values of chlorophyll a(µg cm−3) determined after 1, 2and 3 months of incubation forSan Cristobal (SC) and Escúzarlithotypes
60 A. Z. Miller et al.

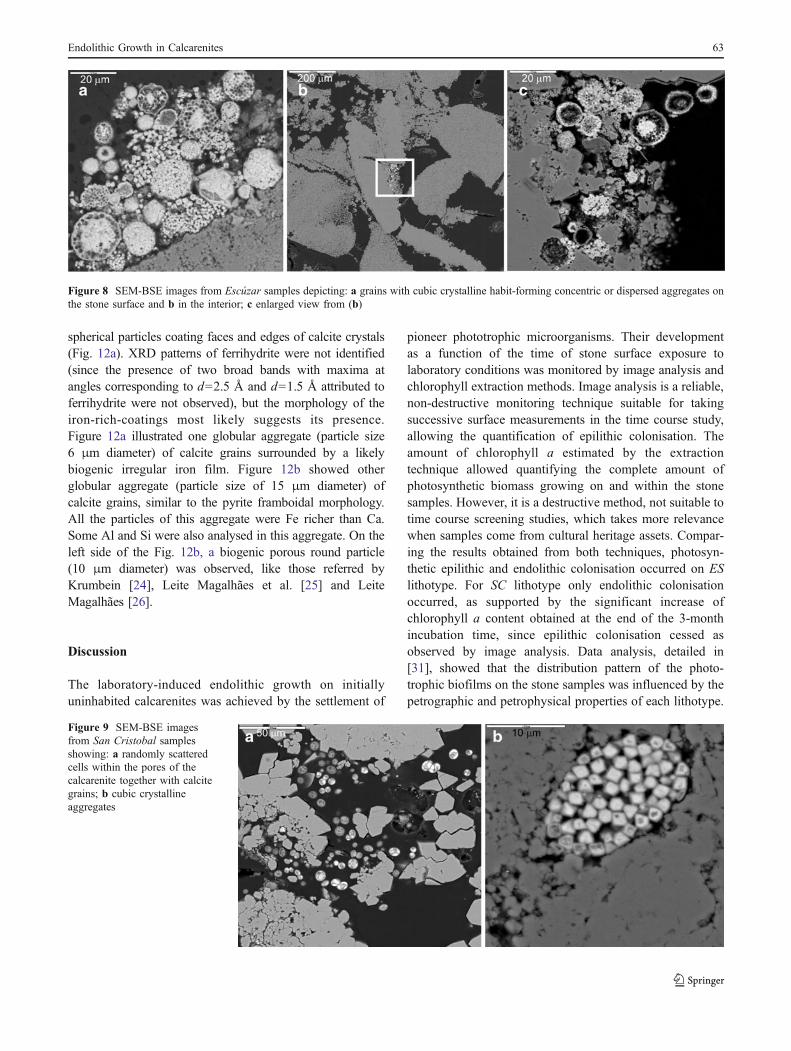
Yet, considering ES samples, SEM-BSE observationsshowed grains with cubic crystalline habit forming concentricor dispersed aggregates (Fig. 8a). These grains were mainlyobserved on the surface of the stone but they were also foundin the interior (at a depth of several millimetres), as shown inFig. 8b, c. The chemical composition of these grains,confirmed by EDS, was the same in each case, indicating theywere deposits of iron oxides–oxyhydroxides. Further details ontheir characterisation are presented in the next section.
Polished sections of SC samples examined by SEM-BSEshowed randomly scattered cells within the pores of the SCbioclastic calcarenite together with calcite grains (Fig. 9a).Figure 9b showed cubic crystalline aggregates, similar tothose detected in the ES samples.
Biocalcarenite Mineralogical and MicromorphologicalCharacteristics
X-ray Diffraction
The bulk mineralogy of the non-inoculated biocalcarenitesample comprised calcite (99%) and quartz (1%). K-feldspar,muscovite, biotite, granada, siderite, pyrolusite, clay mineralsand goethite were the associated minerals that occur investigial amounts (<1%). In the bulk colonised sample, calcite(99%), quartz (1%) and both goethite and pyrolusite (<1%)were the identified minerals. This last sample was separatedby hand-picking for some mineral phases which were brown-black in colour, with sub-metallic lustre and with very goodcleavage. These mineral phases were analysed from theinterior of a stone sample fissure showing yellow pigmentzones, visible to the naked eye and also reddish dots. The bulkmineralogical composition of this concentrated sample was:calcite (86%), quartz (3%), mica and clay minerals (<1%),maghemite (1%) and hausmannite (10%).
Taking into account that poorly crystalline hydrous ironoxides (goethite), anhydrous oxides of iron (maghemite)and manganese minerals usually give even poorer diffrac-tion patterns with fewer broader weaker reflections [9], theinterpretation of the obtained XRD patterns led us toconsider that pyrolusite (MnO2) was identified by itsdiagnostic line at 3.16 Å, followed by the reflections at2.42, 2.21, 2.13, 1.98 and 1.56 Å. Other reflections werecoincident with those of calcite. The d values of about 4.93,
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
0 0.5 1 1.5 2 2.5 3 3.5 4
Area (cm2)
Ch
loro
ph
yll a
co
nte
nt
(µg
cm
-3)
ES SC
Figure 4 Comparison of chlorophyll a contents and biofilm-coveredsurface area evolution on the San Cristobal (SC) and Escúzar (ES)lithotypes during 3 months under laboratory conditions
Figure 5 Original micrographs and false-colour images elaborated from the minority principal components of: a, b San Cristobal lithotype and c,d Escúzar lithotype. Colour key: light blue cavities covered by endolithic biofilms, red rock surface. Scale bar 1 mm
Endolithic Growth in Calcarenites 61

2.77, 2.49, 2.46, 1.576 and 1.544 Å resemble those ofhausmannite (Mn2+Mn2
3+O4). Goethite (α-FeOOH) wasidentified by its peaks at 4.17–4.18 Å and 2.69–2.70 Å. Thebehaviour of the reflections indicated the presence of onedisordered goethite. Maghemite (γ-Fe2O3) was identified
by its more intense reflections at 2.519 and 1.474 Å, as wellas by smaller and broader reflections at 2.95, 2.086 and1.604 Å.
SEM-EDS Analyses
The biocalcarenite morphological characterisation wascarried out on the ES colonised stone fragments. Theselected fragments for XRD were the same for SEM-EDSanalyses to enable comparing stone substratum mineralogyand the poorly crystalline mineral phases identified byXRD, in particular iron oxides–oxyhydroxides and manga-nese minerals. The difficulties rose in identifying Mnoxides from XRD could be exceeded combining bothvisual identification (brown-black in colour mineral phases)and SEM-EDS analyses.
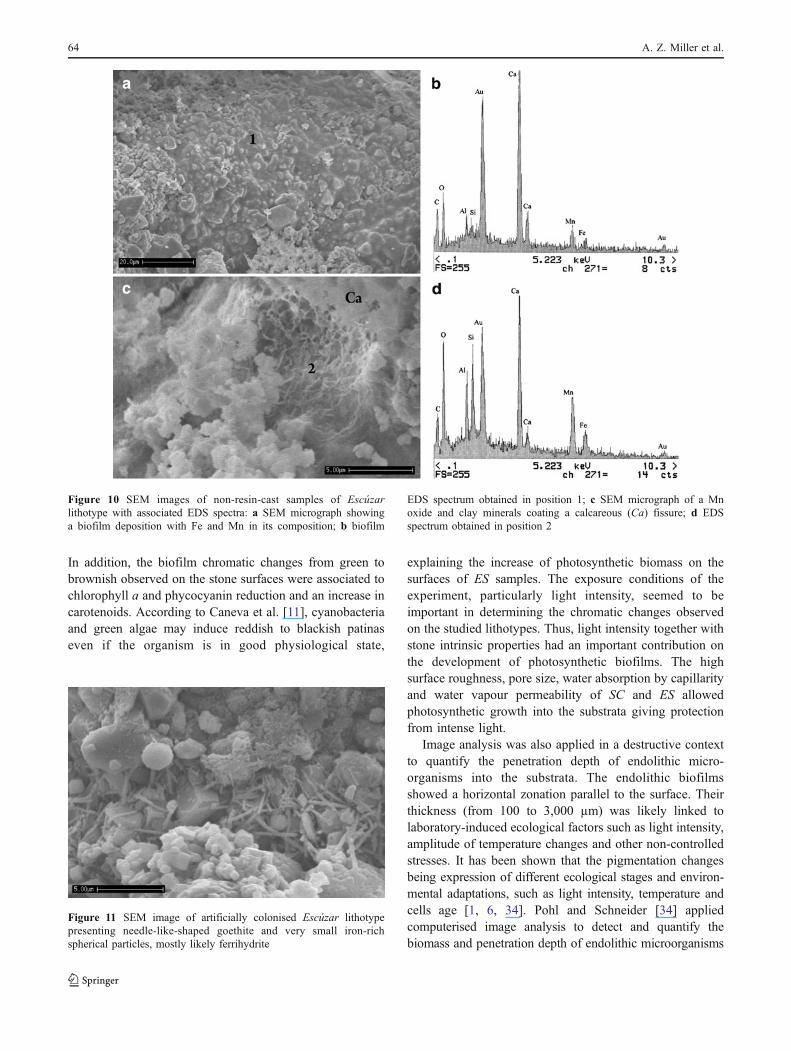
Figure 10a, b showed the biofilm deposition in anendolithic microenvironment of the biocalcarenite with Feand Mn reniform coats associated with kaolin minerals. InFig. 10a it was also clear that the presence of microbialcells adhered to the lithic substratum and embedded in amatrix of EPS. Figure 10c, d depicted clumps of Mn oxideand clay minerals (smectite) coating a massive calcareousfissure.
SEM-EDS study confirmed the goethite occurrence owingto the presence of needle-like shapes (Fig. 11). Goethite,identified by XRD as very poorly crystalline phase, wasassociated with ferrihydrite, which appeared as very small
Figure 6 SEM-BSE image from Escúzar lithotype depicting aninteraction result between the inoculated phototrophic microorganismsand the substratum. Particularly noticeable is the endolithic activitycreating small cavities in the calcite crystals
Figure 7 SEM-BSE images of phototrophic microorganisms in theEscúzar samples illustrating: a epilithic microorganisms randomlyscattered on the stone surface and the likely depths endolithic cellspenetrating the stone through the accessible void spaces; b detail from
(a) showing compact cell aggregates of the endolithic colonisation; cdistribution map of Ca showing the dissolutions features of the calcitein the endolithic microhabitat analysed by EDS microanalytical Line-Scan analysis
62 A. Z. Miller et al.
spherical particles coating faces and edges of calcite crystals(Fig. 12a). XRD patterns of ferrihydrite were not identified(since the presence of two broad bands with maxima atangles corresponding to d=2.5 Å and d=1.5 Å attributed toferrihydrite were not observed), but the morphology of theiron-rich-coatings most likely suggests its presence.Figure 12a illustrated one globular aggregate (particle size6 μm diameter) of calcite grains surrounded by a likelybiogenic irregular iron film. Figure 12b showed otherglobular aggregate (particle size of 15 μm diameter) ofcalcite grains, similar to the pyrite framboidal morphology.All the particles of this aggregate were Fe richer than Ca.Some Al and Si were also analysed in this aggregate. On theleft side of the Fig. 12b, a biogenic porous round particle(10 μm diameter) was observed, like those referred byKrumbein [24], Leite Magalhães et al. [25] and LeiteMagalhães [26].
Discussion
The laboratory-induced endolithic growth on initiallyuninhabited calcarenites was achieved by the settlement of
pioneer phototrophic microorganisms. Their developmentas a function of the time of stone surface exposure tolaboratory conditions was monitored by image analysis andchlorophyll extraction methods. Image analysis is a reliable,non-destructive monitoring technique suitable for takingsuccessive surface measurements in the time course study,allowing the quantification of epilithic colonisation. Theamount of chlorophyll a estimated by the extractiontechnique allowed quantifying the complete amount ofphotosynthetic biomass growing on and within the stonesamples. However, it is a destructive method, not suitable totime course screening studies, which takes more relevancewhen samples come from cultural heritage assets. Compar-ing the results obtained from both techniques, photosyn-thetic epilithic and endolithic colonisation occurred on ESlithotype. For SC lithotype only endolithic colonisationoccurred, as supported by the significant increase ofchlorophyll a content obtained at the end of the 3-monthincubation time, since epilithic colonisation cessed asobserved by image analysis. Data analysis, detailed in[31], showed that the distribution pattern of the photo-trophic biofilms on the stone samples was influenced by thepetrographic and petrophysical properties of each lithotype.
Figure 8 SEM-BSE images from Escúzar samples depicting: a grains with cubic crystalline habit-forming concentric or dispersed aggregates onthe stone surface and b in the interior; c enlarged view from (b)
Figure 9 SEM-BSE imagesfrom San Cristobal samplesshowing: a randomly scatteredcells within the pores of thecalcarenite together with calcitegrains; b cubic crystallineaggregates
Endolithic Growth in Calcarenites 63
In addition, the biofilm chromatic changes from green tobrownish observed on the stone surfaces were associated tochlorophyll a and phycocyanin reduction and an increase incarotenoids. According to Caneva et al. [11], cyanobacteriaand green algae may induce reddish to blackish patinaseven if the organism is in good physiological state,
explaining the increase of photosynthetic biomass on thesurfaces of ES samples. The exposure conditions of theexperiment, particularly light intensity, seemed to beimportant in determining the chromatic changes observedon the studied lithotypes. Thus, light intensity together withstone intrinsic properties had an important contribution onthe development of photosynthetic biofilms. The highsurface roughness, pore size, water absorption by capillarityand water vapour permeability of SC and ES allowedphotosynthetic growth into the substrata giving protectionfrom intense light.
Image analysis was also applied in a destructive contextto quantify the penetration depth of endolithic micro-organisms into the substrata. The endolithic biofilmsshowed a horizontal zonation parallel to the surface. Theirthickness (from 100 to 3,000 µm) was likely linked tolaboratory-induced ecological factors such as light intensity,amplitude of temperature changes and other non-controlledstresses. It has been shown that the pigmentation changesbeing expression of different ecological stages and environ-mental adaptations, such as light intensity, temperature andcells age [1, 6, 34]. Pohl and Schneider [34] appliedcomputerised image analysis to detect and quantify thebiomass and penetration depth of endolithic microorganisms
Figure 11 SEM image of artificially colonised Escúzar lithotypepresenting needle-like-shaped goethite and very small iron-richspherical particles, mostly likely ferrihydrite
Figure 10 SEM images of non-resin-cast samples of Escúzarlithotype with associated EDS spectra: a SEM micrograph showinga biofilm deposition with Fe and Mn in its composition; b biofilm
EDS spectrum obtained in position 1; c SEM micrograph of a Mnoxide and clay minerals coating a calcareous (Ca) fissure; d EDSspectrum obtained in position 2
64 A. Z. Miller et al.

into carbonate rock, showing that endolithic biofilms fromprotected sites (humid and shaded) showed a residualsubstratum thickness of only a few micrometres, whereasfrom intensely isolated dry sites retreated to depths of 150–250 µm below to the rock surfaces. Therefore, it can beconsidered that light intensity inside the chamber was alimiting factor influencing the behaviour and distributionpattern of the microbial communities.
The potential for damage to the stone substrata byphototrophic microorganisms was demonstrated in thisexperimentation. Optical microscopy, SEM-BSE and EDSand X-ray diffraction procedures revealed the close associ-ation between microorganisms and rock alteration processesclearly reflecting the biological contribution to thesedeterioration phenomena. The cyanobacteria and algae andthe diffusion of their excreted products into the poroussystem led to biomechanical and biochemical alterations.De los Ríos et al. [12] referred biogeomechanical andbiogeochemical processes associated with epilithic andendolithic microorganisms detected at the convent of SantaCruz la Real (Segovia, Spain). Due to the inoculationprocedure applied in this study, it was possible to assumethe cells were dragged into the rock (a purely physicalprocess) and thus, they cannot be considered as endolithicactive-colonisers but rather, they were passively installedthere by the percolating inoculum. However, the observedsmall cavities actively created by endolithic microorgan-isms in the calcite crystals from ES samples revealed theinteraction between the microorganisms and the substratum.It indicates that as a result of cells activity the stone wasdissolved due to the biochemical induced activity, decreasingpH conditions. The presence of EPS might increase thelimestones biodeteriorating potential, contributing to bio-chemical activities and disaggregation of rock grains. Kaplanet al. [22] described the chelating properties of Chlorellaexopolysaccharides, which were very important in the way
these organisms deteriorate stone. The stone surface can losecohesion due to contraction and expansion of these biofilmsbecause EPS incorporate large amounts of water into itsstructure ensuring the maintenance of moisture by balancingchanges in humidity and temperature [40].
In the context of the mineralogical and micromorpho-logical characterisation of the ES lithotype brown-blackmineral phases, we consider that the cubic aggregatesobserved in Figs. 8a and 9b, are maghemite (γ-Fe2O3) andhausmannite (Mn2+Mn2
3+O4), which are usually of crystalcube morphology and square shaped, respectively. Sinceseveral manganese and iron minerals are essential forbacterial cell metabolism and are often associated withclays and iron oxides in soils and other environments[9, 14, 15, 23], concomitant precipitation of Fe–Mn oxidesmust be considered in this biogenic microenvironment.However, manganese minerals and hydrous and anhydrousiron oxides are usually poorly crystalline and often showfewer reflections [9], as occurred in this study. According toBrown [9], some difficulties arise in recognising these poorlycrystalline hydrous iron oxides (goethite, ferrihydrite),anhydrous iron oxides (maghemite) and manganese mineralsbecause they often show fewer reflections or they occur insmall amounts in the samples. The XRD patterns of someMn oxides and hydroxides obtained in this study weresimilar and suggested that the materials were structurallyrelated. The scarcity of the manganese minerals and theabsence of peaks could hide the presence of mineralmixtures. Nevertheless, the diffuse nature of the XRDpatterns of Fe–Mn oxides–oxyhydroxides and the coinci-dence of the diagnostic reflections with those of otherminerals could be performed in more detail, following someconcentrate pre-treatment [14].
Previous investigations have shown that in spite of theimportance of Mn oxides in the environment, the molecularmechanisms, rates, intermediates and products of Mn oxide
Figure 12 SEM images of Escúzar lithotype showing: a calcite crystals coated by iron oxyhydroxides and a globular aggregate of calcite grainssurrounded by an iron film; b one globular aggregate of calcite grains with iron-rich coating and other porous round particle, likely biogeneous
Endolithic Growth in Calcarenites 65

biomineralisation are poorly understood. Even the relation-ship between biotic and abiotic Mn oxidation cycles is notwell documented [5]. However, in a recent work carried outby Fischer et al. [16] about a biologically mediated mineralreaction, bioreduction and dissolution of birnessite, a layeredMn3+, 4+ oxide and the concomitant precipitation ofrhodocrosite (Mn2+CO3) and hausmannite (Mn2+Mn2
3+O4)was observed, using both synchrotron and conventionalX-ray sources.
In our study, mineralogical data showed the presenceof pyrolusite (MnO2) in the non-inoculated stone, whereashausmannite (Mn2+Mn2
3+O4) was the Mn oxide identifiedby XRD in the colonised stone sample. The divalentmanganese form is soluble and the more stable tetravalentform, usually represented by the dioxide MnO2, isinsoluble. Thus, the availability of Mn from MnO2 andthe microbial activity alter the Eh/pH of the microenvi-ronment and indirectly modify the valence of manganese.This process is not necessarily cell associated and mayoccur at considerable distances from the cell [23].According to this author it is not surprising that manga-nese transformations occur as an indirect consequence ofmicrobial growth and metabolism. Probably, the precipi-tation of hausmannite (Mn II, III) was conceivable in suchparticular endolithic environment. Figure 10 is one of theevidences of the biomineralisation: (1) presence of abiofilm deposition with Fe and Mn in its composition(Fig. 10a,b); (2) cyanobacteria remains on a fissure,yellow-brown in colour, coated by Mn oxide and clayminerals (Fig. 10c, d).
XRD and SEM-EDS studies allowed the identification ofgoethite and maghemite. In addition to these oxides–oxyhydroxides, a poorly crystalline natural hydrated ironoxide, which is analogous to the so-called “colloidal ferrichydroxide”, named ferrihydrite [9], may be responsible tothe observed coatings depicted in Fig. 12. This authorreferred that materials commonly formed by bacterialoxidation of ferrous solutions, give similar XRD patternsto ferrihydrite. According to Robert [35], some bacteriause both oxidation–reduction reaction energy and carbonfor the soluble Fe transportation. Successive fixation ofFe onto the exopolysaccharides excreted by the micro-bial community creates a capsule involving the cells andforming ferrihydrite, which can develop to iron oxideafter cells death. Other relevant reactions governed bybacteria concern iron and manganese reduction oroxidation [19].
In conclusion, this paper shows, for the first time, anartificially induced biogenic deterioration of calcarenitescaused by endolithic microorganisms, and associatedbiomineralisation. The model applied in this workcorroborated a successful survival strategy inside endo-lithic microhabitat, using natural phototrophic biofilm
cultivation, composed by cyanobacteria and algae, whichincreased intrinsic porosity by active mineral dissolution.
These findings helped to explain: (1) the presence ofmineral-like iron derivatives (e.g. maghemite) around thecells and intracellular; (2) locally enhanced moistureavailability by the occurrence of clay minerals (kaoliniteand smectite) stimulating microbial growth; (3) the role ofMn oxides, in particular by the precipitation of hausmannite,manganese (II, III) suggesting manganese transformationsrelated to the biomineralisation.
With a view towards the future conservation of stonemonuments, a reliable characterisation of the microorgan-isms dwelling on the stone surfaces, as well as theassessment of the relationships between microorganismsand the mineral substrata are a prerequisite. Three mainrecommendations with respect to prevention and treatmentof microbial colonisation can be pointed out: (a) specialattention should be taking when using San Cristobal andEscúzar lithotypes, with high surface roughness, openporosity and water absorption by capillarity, in theconstruction of new buildings without previous hydropho-bic treatments; (b) in the case of materials already placed ina structure, the exact identification of the colonisingmicroflora, as well as, the assessment of the biodeteriora-tion damage are of great importance since it will allow thedesign of accurate intervention; (c) the control anderadication of phototrophic biofilms on the stone surfacesare strongly recommended. When using biocides, great careshould be taken. As an alternative to biocides, hydrophobictreatments can be applied to prevent water from cominginto the stone by the use of organic or silico-organicproducts. It must be said that such products should meet aspecific list of chemical, physical and aesthetic require-ments. Therefore, the benefit of a multidisciplinary evalu-ation with the complementary cooperation of conservatorsand scientists in the diagnosis and handling of thebiodeterioration effects are necessary to the conservationof stone monuments.
Acknowledgements This work was supported by the Ministério daCiência, Tecnologia e Ensino Superior, Portugal, with a doctoral grant(SFRH/BD/21481/2005) and partially financed by the CEPGIST FCTsubproject DECASTONE. The projects TCP CSD2007-00058 and2007PT0041 are acknowledged. We also thank F. Pinto (CCMA,CSIC) for technical assistance and CA and JW thanks for support bygrant CGL2007-62875/BOS from the Ministry of Science andInnovation of Spain and by PIE-631A from CSIC, Spain.
References
1. Alakomi HL, Arrien N, Gorbushina AA, Krumbein WE, MaxwellI, McCullagh C, Robertson P, Ross N, Saarela M, Valero J,Vendrell M, Young ME (2004) Inhibitors of biofilm damage onmineral materials (Biodam). In: Kwiatkowski D, Löfvendahl R
66 A. Z. Miller et al.

(eds) Proceedings of the 10th International Congress on Deteri-oration and Conservation of Stone. ICOMOS, Stockholm, pp399–406
2. Ariño X, Hernandez-Marine M, Saiz-Jimenez C (1997) Colonizationof Roman tombs by calcifying cyanobacteria. Phycologia 36:366–373
3. Ascaso C, Wierzchos J, Castello R (1998) Study of the biogenicweathering of calcareous litharenite stones caused by lichen andendolithic microorganisms. Int Biodeterior Biodegrad 42:29–38
4. Ascaso C, Wierzchos J, Souza-Egipsy V, De los Ríos A, DelgadoRodrigues J (2002) In situ evaluation of the biodeterioratingaction of microorganisms and the effects of biocides on carbonaterock of the Jeronimos Monastery (Lisbon). Int BiodeteriorBiodegrad 49:1–12
5. Bargar JR, Tebo BM, Pecher KH, Chiu V, Villinski JE, Tonner BP(2000) Kinetics and products on Mn oxide biomineralization byspores of the marine Bacillus sp. strain-1. Symposia paperspresented before the division of Environment Chemistry AmericanChemical Society 40(2):491–493
6. BartoliniM, Ricci S, Del Signore G (2004) Release of photosyntheticpigments from epilithic biocenosis alter biocida treatments. In:Kwiatkowski D, Löfvendahl R (eds) Proceedings of the 10thInternational Congress on Deterioration and Conservation of Stone.ICOMOS, Stockholm, pp 519–526
7. Bell RA (1993) Cryptoendolithic algae of hot semiarid lands anddeserts. J Phycol 29:133–139
8. Bellinzoni AM, Caneva G, Ricci S (2003) Ecological trends intravertine colonisation by pioneer algae and plant communities.Int Biodeterior Biodegrad 51:203–210
9. Brown G (1980) Associated minerals. In: Brindley GW, Brown G(eds) Crystal structures of clay minerals and their X-rayidentification, vol 5. Mineralogical Society, London, pp 361–410
10. Büdel B, Weber B, Kühl M, Pfanz H, Sültemeyer D, Wessels D(2004) Reshaping of sandstone surfaces by cryptoendolithiccyanobacteria: bioalkalization causes chemical weathering in aridlandscapes. Geobiology 3:261–268
11. Caneva G, Nugari MP, Salvadori O (1991) Biology in theconservation of work of art. ICCROM, Rome, p 182
12. De Los Ríos A, Galván V, Ascaso C (2004) In situ microscopicaldiagnosis of biodeterioration processes at the convent of Santa Cruzla Real, Segovia, Spain. Int Biodeterior Biodegrad 54:113–120
13. De Los Ríos A, Ascaso C (2005) Contributions of in situmicroscopy to the current understanding of stone biodeterioration.Int Microbiol 8:181–188
14. Dixon J (1977) Minerals in soil environments. Soil ScienceSociety of America, Wisconsin
15. Dorn R (1998) Rock coatings. Developments in Earth SurfaceProcesses 6. Elsevier, Amsterdam
16. Fischer TB, Heaney PJ, Jang JH, Ross DE, Brantley SL, Post JE,Tien M (2008) Continuous time-resolved X-ray diffraction of thebiocatalysed reduction of Mn oxide. American Mineralogist 93(11-12):1929–1932
17. Friedmann EI (1982) Endolithic microorganisms in the AntarcticCold Desert. Science 215:1045–1053
18. Golubic S, Friedmann I, Schneider J (1981) The lithobionticecological niche, with special reference to microorganisms. JSediment Petrol 51:475–478
19. Gonzalez I, Laiz L, Hermosin B, Caballero B, Incerti C,Saiz-Jimenez C (1999) Bacteria isolated from rock art paintings:the case of Atlanterra shelter (south Spain). J Microbiol Methods36:123–127
20. Guillitte O (1995) Bioreceptivity: a new concept for buildingecology studies. Sci Total Environ 167:215–220
21. Guillitte O, Dreesen R (1995) Laboratory chamber studies andpetrographical analysis as bioreceptivity assessment tools ofbuilding materials. Sci Total Environ 167:365–374
22. Kaplan D, Christiaen D, Arad S (1987) Chelating properties ofextracellular polysaccharides from Chlorella spp. Appl EnvironMicrobiol 53:2953–2956
23. Krumbein WE (1983) Microbial geochemistry. Blackwell Scientific,Oxford
24. Krumbein WE (1992) Colour changes of building stones and theirdirect and indirect biological causes. Proceedings of 7th Interna-tional congress on Deterioration and Conservation of Stone,Lisboa, pp 443–452
25. Leite Magalhães S, Sequeira Braga MA, Ascaso C (1998) Pátinasbiogénicas numa fachada granítica do edifício do Largo do Paço(Braga). Actas do V Congresso Nacional de Geologia, Tomo 84,Fas. 2, Lisboa, pp F-194-F-197
26. Leite Magalhães S (2000) Biodeterioração de um monumento dacidade de Braga. Estudo microbiológico da pedra granítica. MScEthesis, Universidade do Minho, Braga, Portugal
27. Lorenzen CJ (1967) Determination of chlorophyll and pheo-pigments:spectrophotometric equations. Limnol Oceanogr 12:343–346
28. McNamara CJ, Perry TD IV, Bearce KA, Hernandez-Duque G,Mitchell R (2006) Epilithic and endolithic bacterial communities inlimestone from a Maya archaeological site. Microb Ecol 51:51–64
29. Miller A, Dionísio A, Macedo MF (2006) Primary bioreceptivity:a comparative study of different Portuguese lithotypes. IntBiodeterior Biodegrad 57:136–142
30. Miller AZ, Laiz L, Gonzalez JM, Dionisio A, Macedo MF,Saiz-Jimenez C (2008) Reproducing stone monumentphotosynthetic-based colonization under laboratory conditions.Sci Total Environ 405:278–285
31. Miller AZ, Leal N, Laiz L, Rogerio-Candelera MA, Gonzalez JM,Silva RJC, Dionísio A, Macedo MF, Saiz-Jimenez C (2010)Primary bioreceptivity of limestones used in Southern Europemonuments. In: Smith BJ, Gomez-Heras M, Viles HA, Cassar J(eds) Limestone in the built environment: Present day challengesfor the preservation of the past. Geological Society of London,London, Special Publications 331:79–92
32. Miller AZ, Laiz L, Dionísio A, Macedo MF, Saiz-Jimenez C (2009)Growth of phototrophic biofilms from limestone monuments underlaboratory conditions. Int Biodeterior Biodegrad 63:860–867
33. Ortega-Calvo JJ, Hernandez-Marine M, Saiz-Jimenez C (1991)Biodeterioration of building materials by cyanobacteria and algae.Int Biodeterior 28:165–185
34. Pohl W, Schneider J (2002) Impact of endolithic biofilms oncarbonate rock surfaces. Geological Society, London, SpecialPublication, pp 177–194
35. Robert M (1993) Rôle du facteur biologique dans la dégradationdes roches et des monuments. In: Vincente Hernández MA,Molina Ballesteros E, Rives Arnau V (eds) Actas del workshop:Alteración de granitos y rocas afines, empleados como materialesde contrucción. CSIC, Madrid, pp 103–115
36. Rogerio-Candelera MA (2008) Una propuesta no invasiva para ladocumentación integral del arte rupestre. MScE thesis, Universidadde Sevilla, Spain
37. Rogerio-Candelera MA, Laiz L, Saiz-Jimenez C (2008) Una experi-encia de laboratorio para la separación de cubiertas en la documentaciónde pinturas rupestres y murales afectadas por biodeterioro. In: Saiz-Jimenez C, Rogerio-Candelera MA (eds) Novena Reunión de la RedTemática del CSIC de Patrimonio Histórico y Cultural “Avancesrecientes en la investigación sobre Patrimonio”. CSIC, Sevilla, pp 71–72
38. Rogerio-Candelera MA, Laiz L, González JM, Saiz-Jimenez C(2008) Monitorización del crecimiento microbiano en una tumbaromana mediante técnicas de teledetección. In: Rovira S, Garcia-Heras M, Gener M, Montero I (eds) Actas del VII CongresoIbérico de Arqueometría, CSIC, Madrid, pp 593–600
39. Saiz-Jimenez C, Garcia-Rowe J, Garcia del Cura MA, Ortega-CalvoJJ, Roekens E, Van Grieten R (1990) Endolithic cyanobacteria inMaastricht limestone. Sci Total Environ 94:209–220
Endolithic Growth in Calcarenites 67

40. Saiz-Jimenez C (1999) Biogeochemistry of weathering processesin monuments. Geomicrobiol J 16:27–37
41. Salvadori O (2000) Characterization of endolithic communities ofstone monuments and natural outcrops. In: Ciferri O, Tiano P,Mastromei G (eds) Of microbes and art—the role of microbialcommunities in the degradation and protection of culturalheritage. Kluwer Academic, New York, pp 89–101
42. Tiano P, Accolla P, Tomaselli L (1995) Phototrophic biodeteriogenson lithoid surfaces: an ecological study. Microb Ecol 29:299–309
43. Tomaselli L, Lamenti G, Bosco M, Tiano P (2000) Biodiversity ofphotosynthetic micro-organisms dwelling on stone monuments.Int Biodeterior Biodegrad 46:251–258
44. Vollenweider RA, Talling JF, Westlake DF (1974) A manual onmethods for measuring primary production in aquatic environments,2nd edn. Blackwell, Oxford
45. Walker JJ, Spear JR, Pace NR (2005) Geobiology of a microbialendolithic community in the Yellowstone geothermal environment.Nature 434:1011–1014
46. Wierzchos J, Ascaso C (1994) Application of backscatteredelectron imaging to the study of the lichen-rock interface. Journalof Microsc-Oxford 175:54–59
47. Wilderer PA, Characklis WG (1989) Structure and function ofbiofilms. In: Characklis WG, Wildener PA (eds) Structure andfunction of biofilms. Wiley, Chichester, pp 5–17
68 A. Z. Miller et al.




















