La Angiotensina ii y la respuesta neurohumoral frente al estrés
10
Disponible en: http://www.redalyc.org/articulo.oa?id=55924202 Red de Revistas Científicas de América Latina, el Caribe, España y Portugal Sistema de Información Científica A. Israel, B. Sosa-Canache, I. Medina La Angiotensina II y la respuesta neurohumoral frente al estrés Archivos Venezolanos de Farmacología y Terapéutica, vol. 24, núm. 2, 2005, p. 0, Sociedad Venezolana de Farmacología Clínica y Terapéutica Venezuela ¿Cómo citar? Fascículo completo Más información del artículo Página de la revista Archivos Venezolanos de Farmacología y Terapéutica, ISSN (Versión impresa): 0798-0264 [email protected], [email protected] Sociedad Venezolana de Farmacología Clínica y Terapéutica Venezuela www.redalyc.org Proyecto académico sin fines de lucro, desarrollado bajo la iniciativa de acceso abierto
-
Upload
independent -
Category
Documents
-
view
6 -
download
0
Transcript of La Angiotensina ii y la respuesta neurohumoral frente al estrés

Disponible en: http://www.redalyc.org/articulo.oa?id=55924202
Red de Revistas Científicas de América Latina, el Caribe, España y Portugal
Sistema de Información Científica
A. Israel, B. Sosa-Canache, I. Medina
La Angiotensina II y la respuesta neurohumoral frente al estrés
Archivos Venezolanos de Farmacología y Terapéutica, vol. 24, núm. 2, 2005, p. 0,
Sociedad Venezolana de Farmacología Clínica y Terapéutica
Venezuela
¿Cómo citar? Fascículo completo Más información del artículo Página de la revista
Archivos Venezolanos de Farmacología y
Terapéutica,
ISSN (Versión impresa): 0798-0264
[email protected], [email protected]
Sociedad Venezolana de Farmacología Clínica y
Terapéutica
Venezuela
www.redalyc.orgProyecto académico sin fines de lucro, desarrollado bajo la iniciativa de acceso abierto

La Angiotensina II y la respuesta neurohumoral frente al estrés
La Angiotensina ii y la respuesta neurohumoral frente al estrés
A Israel1, B Sosa-Canache2 e I Medina2.
1 Facultad de Farmacia, Universidad Central de Venezuela, Caracas.
2 Universidad CentroOccidental "Lisandro Alvarado", Barquisimeto, Estado Lara, Venezuela.
E-mail: [email protected]
RESUMEN
El estímulo eléctrico plantar (EEP) aplicado a ratas conscientes, incrementa la presión arterial media (PAM) y la frecuencia cardíaca (FC). En el presente trabajo evaluamos la hipótesis que la angiotensina II (ANG II) endógena media la respuesta simpática frente al EEP. Nuestros resultados muestran que el bloqueo del receptor AT1 con losartan, valsartan o eprosartan previene la respuesta vasopresora inducida por el EEP. La nefrectomía bilateral (NFX), la cual reduce los niveles circulantes de ANG, suprimió el incremento de la PAM y de las catecolaminas plasmáticas observada durante el EEP. La administración de una dosis subpresora de angiotensina II restauró la respuesta vasopresora inducida por el estrés en las ratas nefrectomizadas. La modulación de los niveles circulantes de angiotensina II alteró la respuesta presora frente al estrés. Así, la reducción de los niveles circulantes de ANG mediante una ingesta elevada de sodio, inhibió la respuesta presora frente al EEP. Por el contrario, el incremento del sistema renina-angiotensina inducido por una dieta baja en sal, facilitó la respuesta presora frente al estrés. Nuestros resultados indican que la angiotensina II endógena soporta la respuesta simpática frente al estrés, a través de la estimulación de los receptores AT1 presinápticos.
Palabras Clave: Angiotensina II, Estrés, Respuesta neurohumoral.
ABSTRACT
Footshocks (FS) increases mean arterial pressure (MAP) and heart rate (HR). We tested the hypothesis that endogenous angiotensin II (ANG) supports sympathetically mediated pressor response to FS. Our results show that AT1 receptor blockade with either losartan, valsartan or eprosartan blunted stress-response to FS. Reduction of circulating ANG induced by bilateral nephrectomy (NPHX), suppressed MAP increase and plasma catecholamines elevation observed after FS. Stress induced-pressor response to FS in nephrectomized rats was restored by the administration of a subpressor dose of synthetic ANG. Reduction of plasma ANG by high sodium diet suppressed, while increased ANG levels induced by low sodium diet facilitated stress-response to FS. Our results indicate that endogenous ANG supports sympathetic mediated stress-response, via stimulation of prejunctional AT1 receptors.
Key Words: Angiotensin II, Stress, Neurohumoral response.
INTRODUCCIÓN
Existen diversas situaciones excepcionales en las que se producen graves amenazas para la integridad del sujeto que padece estímulos o agentes estresantes muy intensos que pueden afectar de forma considerable la salud del individuo. Ante las situaciones estresantes el ser humano moviliza sus recursos fisiológicos con el fin de responder a éstas con lo que se denomina la "reacción o respuesta al estrés". El estrés constituye un agente nocivo en diversos sistemas del organismo, en algunos casos por su acción directa contribuye a muchas enfermedades que incluyen las cardiovasculares. El estrés y la hipertensión se encuentran muy relacionados, en tanto que incrementos transitorios de la presión arterial que inducen situaciones de estrés pueden convertirse en permanentes al asociarse a factores ambientales, psicológicos o
file:///G|/archivos%202%202005/html/La%20Angiotensin...0respuesta%20neurohumoral%20frente%20al%20estrés.htm (1 of 9)03/02/2006 08:12:29 a.m.

La Angiotensina II y la respuesta neurohumoral frente al estrés
genéticos.
Numerosas factores neurohumorales participan en los efectos nocivos del estrés. Uno de estos factores es la angiotensina II (ANG), considerada en la actualidad como la hormona de estrés por estar implicada en la activación del eje hipotálamo-hipófisis-suprarrenal (Armando y col., 2001). El estrés incrementa la secreción de renina, la producción de ANG circulante y la expresión de receptores para ANG en áreas del cerebro relacionadas con el control central del estrés (Saavedra, 1992). Estudios realizados tanto en modelos animales como en humanos sugieren que la angiotensina II participa en la regulación del sistema nervioso simpático y podría tener un papel en la respuesta cardiovascular inducida por el estrés, al menos en parte regulando el eflujo simpático (Xu y Brooks, 1996). Así, se ha demostrado en ratas descerebradas que el bloqueo del receptor AT1 con losartan resulta en la inhibición de la respuesta cardiovascular a la estimulación simpática de la médula espinal (Wong y col., 1992). En un modelo experimental de estrés agudo inducido por el estímulo eléctrico plantar (EEP), el cual activa el eje simpático-adrenal e incrementa la presión arterial y la frecuencia cardiaca, se demostró que el losartan y el captopril inhiben selectivamente la respuesta vasopresora frente al EEP, lo que indica que esta acción se encuentra mediada por el sistema renina-angiotensina, especificamente a través de la estimulación del receptor AT1 (Cierco e Israel, 1994; Israel y col., 1994; Israel y Sosa, 2002). Efectos similares se observaron en ratas conscientes, donde el tratamiento a largo plazo con candesartan previno la respuesta simpatoadrenal y hormonal así como el incremento de las catecolaminas urinarias inducidas por el estrés agudo (Armando y col., 2001) y en ratas hipertensas en las cuales el SRA se encuentra activado (Seltzer y col., 2004).
La evidencia sugiere, que en condiciones en las que el sistema renina-angiotensina se encuentra activado, como sucede durante el EEP, la vasoconstricción adrenérgica se encuentra marcadamente intensificada por la angiotensina endógena. Aún mas, la vasoconstricción mediada adrenérgicamente podría estar intensificada por la angiotensina endógena en condiciones en las cuales el sistema renina-angiotensina está activado, como por ejemplo durante la estenosis aguda de la arteria renal, durante la ingesta crónica de una dieta hiposódica y en condiciones que conduzcan a una pérdida de sodio (Zimmerman y Kraft, 1979). Por el contrario, en situaciones en las cuales hay una actividad plasmática de la renina baja, como ocurre en ratas con nefrectomía bilateral o con una dieta alta en sodio, la transmisión simpática se encuentra disminuída (Brooks y Osborn, 1995; Feuerstein y col., 1977). Estos resultados sugieren que condiciones que aumenten o disminuyan la actividad del sistema renina-angiotensina afectan la función del sistema nervioso simpático (Xu y Brooks, 1996).
Con el fin de establecer el papel de la angiotensina II endógena y la estimulación del receptor AT1 en la expresión de la respuesta simpática frente al estrés, en el presente estudio se evaluó la respuesta simpática frente al estrés eléctrico plantar en ratas conscientes sujetas a 1) bloqueo de los receptores AT1 con losartan, valsartan o eprosartan, 2) nefrectomía bilateral o, 3) la modulación de los niveles plasmáticos de ANG mediante la aplicación de una dieta alta o baja en sodio.
MATERIAL Y MÉTODOS
Se emplearon ratas Sprague-Dawley, machos, de 180 a 200 g. de peso corporal, procedentes del bioterio de la Facultad de Farmacia, U.C.V., Caracas, mantenidas con Ratarina® y agua ad libitum. Todos los procedimientos fueron aprobados por el Comité de Bioterio de la Facultad de Farmacia, Universidad Central de Venezuela.
El registro de los parámetros cardiovasculares, presión arterial (PA) y frecuencia cardíaca (FC) se realizó en ratas conscientes, por un método no invasivo mediante el uso de un pletismógrafo digital de cola (DIGITAL PRESSURE METER LE 5000 y un PRESSURE CYLINDER LE 5100, LETICA®, Barcelona, España).
Se aplicó un estrés, el estímulo eléctrico plantar, que consiste en someter a los animales en experimentación a descargas eléctricas en la zona plantar y que es un modelo experimental de estrés agudo originado por una descarga simpático-adrenal (Lee y col., 1989).
Las ratas se colocaron individualmente en una jaula de plexiglás con la base enrejada con alambre de cobre conectada a un estimulador (GRASS S48), el cual se graduó para producir un estímulo eléctrico de 2 Hertz, 100 voltios, 5 mseg, 5
file:///G|/archivos%202%202005/html/La%20Angiotensin...0respuesta%20neurohumoral%20frente%20al%20estrés.htm (2 of 9)03/02/2006 08:12:29 a.m.

La Angiotensina II y la respuesta neurohumoral frente al estrés
minutos. Después se determinó la presión arterial y la frecuencia cardíaca, como describimos anteriormente.
Modelos experimentales
Nefrectomía bilateral: La nefrectomía bilateral es un modelo experimental en el cual los niveles plasmáticos de renina se encuentran disminuídos. Las ratas, bajo anestesia con pentobarbital sódico (40 mg/kg) se colocaron en decúbito ventral y se les practicó una incisión subcostal paravertebral izquierda. Al riñón se accedió por su cara avascular posterior y se extrajo mediante disección roma e instrumental, seguidamente se localizó y ligó el hilio renal y posteriormente se seccionó. Se tuvo la precaución de no extraer la glándula suprarrenal.
Dieta alta en sodio: Durante 72 horas las ratas recibieron una solución de cloruro de sodio al 2% en el agua. Esto ocasionó una inhibición del sistema renina-angiotensina.
Dieta hiposódica: Las ratas colocadas en jaulas individuales recibieron una dieta hiposódica (0.02% de sodio, HARLAN TEKLAD, MADISON, WI) durante un período de tres semanas. Durante los dos primeros días de estar recibiendo la dieta hiposódica se les inyectó una dosis diaria de furosemida (1 mg/kg, ip.). Un grupo de ratas fueron anestesiadas con pentobarbital sódico (80 mg/kg) y se colocaron en decúbito dorsal e inmediatamente se les practicó una incisión media longitudinal desde el pubis a la apéndice xifoides, se ubicó la vena cava inferior por delante de la columna vertebral, junto con la aorta abdominal. Seguidamente, se tomaron 4 ml de sangre en una inyectadora que contenía 0.1 ml de EDTA al 5% y se vertieron en un tubo de plástico con 0.2 ml de metabisulfito de sodio al 5%. En un plazo no mayor de 30 minutos las muestras se centrifugaron a 15.000 r.p.m. por 10 minutos a 5°C. Inmediatamente, el plasma fue separado y se procedió a la extracción de las catecolaminas con alúmina o a la congelación a -60°C, hasta que las muestras se procesaron.
Determinación de las catecolaminas: norepinefrina y epinefrina
Para la determinación de las catecolaminas se seguió el método de purificación y adsorción por alúmina de Anton y Sayre (1962). Los niveles sanguíneos de las catecolaminas norepinefrina (NE) y epinefrina (EPI) fueron determinados mediante cromatografía líquida de alta eficiencia de fase reversa y por detección electroquímica. Los niveles sanguíneos de noradrenalina y adrenalina se determinaron en sangre periférica de ratas sometidas a una nefrectomía bilateral y en ratas con una intervención simulada sometidas o no a un estímulo eléctrico plantar. Los niveles plasmáticos de catecolaminas se expresaron en picogramos por mililitros de plasma (pg/ml).
Todos los resultados se expresaron como la media ± el error estándar del promedio (EEM). La presión arterial se expresó como presión arterial media (PAM). Este parámetro se obtuvo aplicando la siguiente fórmula: PAD + 1/3 (PAS-PAD). La significancia estadística fue determinada usando las prueba: "t" de Student o el análisis de varianza de una vía (ANOVA) y por el rango estadístico de Student-Newman-Keul’s. Se consideraron significativos los valores de p menores de 0.05 (p<0.05 ).
RESULTADOS
Efecto del bloqueo de los receptores at1 sobre
la respuesta vasopresora al eep
El estímulo eléctrico plantar aplicado a ratas conscientes produce un incremento significativo de las presiones media (Figura 1A), sistólica, diastólita y de la frecuencia cardíaca (no se muestran). El bloqueo del receptor AT1 con losartan (10 mg/kg, s.c.), valsartan (10 mg/kg, p.o.) o eprosartan (120 mg, p.o.), inhibió la respuesta presora al EEP (Figura 1A).
Figura 1
file:///G|/archivos%202%202005/html/La%20Angiotensin...0respuesta%20neurohumoral%20frente%20al%20estrés.htm (3 of 9)03/02/2006 08:12:29 a.m.

La Angiotensina II y la respuesta neurohumoral frente al estrés
(A) Respuesta vasopresora al EEP en ratas tratadas con losartan (10 mg/kg, s.c.), valsartan (10 mg/kg, p.o.) o eprosartan (120 mg, p.o.). (B) Respuesta presora al EEP en ratas sujetas a una dieta alta o baja de sal
Efecto de la modulación del sistema renina-angiotensina sobre la respuesta cardiovascular al estímulo eléctrico plantar
I. Efecto del aumento del sistema renina-angiotensina (SRA): La depleción de sodio ocasiona una disminución del volumen de líquido extracelular con el consiguiente aumento del SRA. En las ratas que recibieron la dieta hiposódica se produjo un aumento significativo en la respuesta de la presión arterial media (14.5 ± 1.1 mmHg) frente al EEP (0.2 Hz/100 V/5 mseg/5 min) que fue mayor al observado en ratas con dieta normosódica (7.9 ± 1.1 mmHg). Es decir, la descarga simpático-adrenal producida por el EEP elevó casi al doble los valores de presión arterial media en las ratas depletadas de sodio (p < 0.01), por lo tanto hubo una facilitación de la respuesta simpática al estrés eléctrico plantar (Figura 1B).
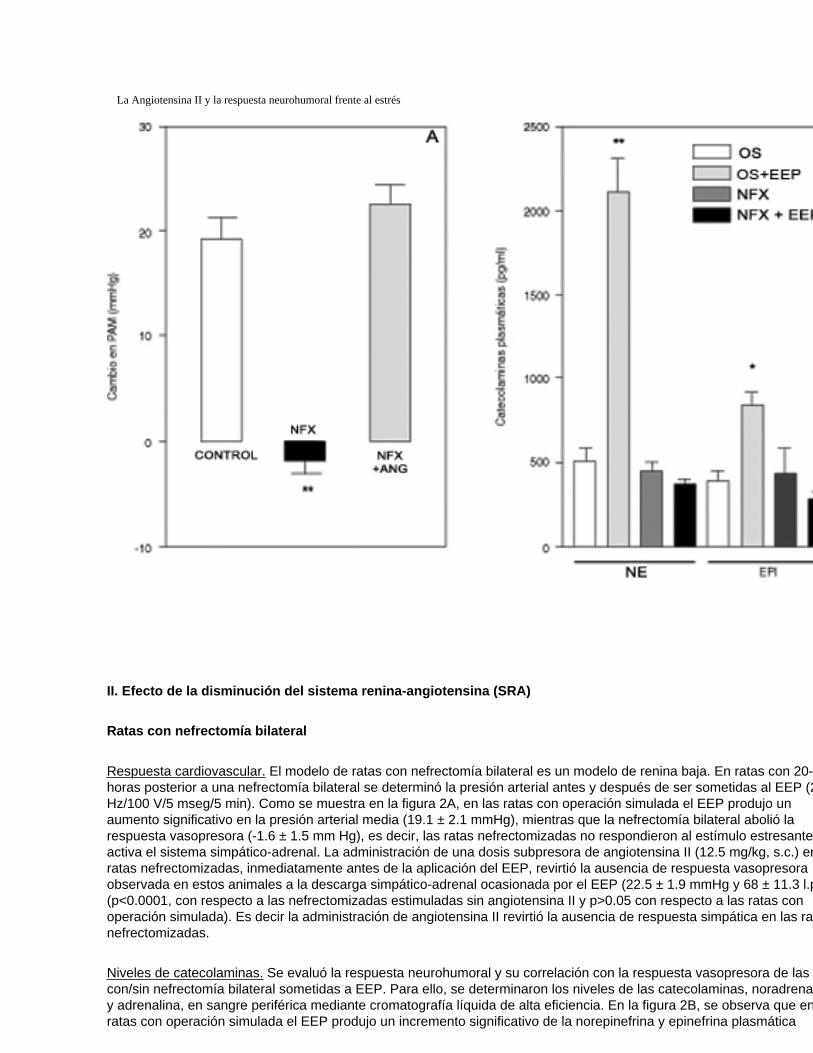
Figura 2
Efecto de la nefrectomía bilateral sobre (A) la respuesta vasopresora (panel izquierda) y (B) catecolaminas plasmáticas (panel derecho) frente al EEP, en ratas
file:///G|/archivos%202%202005/html/La%20Angiotensin...0respuesta%20neurohumoral%20frente%20al%20estrés.htm (4 of 9)03/02/2006 08:12:29 a.m.
La Angiotensina II y la respuesta neurohumoral frente al estrés
II. Efecto de la disminución del sistema renina-angiotensina (SRA)
Ratas con nefrectomía bilateral
Respuesta cardiovascular. El modelo de ratas con nefrectomía bilateral es un modelo de renina baja. En ratas con 20-22 horas posterior a una nefrectomía bilateral se determinó la presión arterial antes y después de ser sometidas al EEP (2 Hz/100 V/5 mseg/5 min). Como se muestra en la figura 2A, en las ratas con operación simulada el EEP produjo un aumento significativo en la presión arterial media (19.1 ± 2.1 mmHg), mientras que la nefrectomía bilateral abolió la respuesta vasopresora (-1.6 ± 1.5 mm Hg), es decir, las ratas nefrectomizadas no respondieron al estímulo estresante que activa el sistema simpático-adrenal. La administración de una dosis subpresora de angiotensina II (12.5 mg/kg, s.c.) en ratas nefrectomizadas, inmediatamente antes de la aplicación del EEP, revirtió la ausencia de respuesta vasopresora observada en estos animales a la descarga simpático-adrenal ocasionada por el EEP (22.5 ± 1.9 mmHg y 68 ± 11.3 l.p.m.) (p<0.0001, con respecto a las nefrectomizadas estimuladas sin angiotensina II y p>0.05 con respecto a las ratas con operación simulada). Es decir la administración de angiotensina II revirtió la ausencia de respuesta simpática en las ratas nefrectomizadas.
Niveles de catecolaminas. Se evaluó la respuesta neurohumoral y su correlación con la respuesta vasopresora de las ratas con/sin nefrectomía bilateral sometidas a EEP. Para ello, se determinaron los niveles de las catecolaminas, noradrenalina y adrenalina, en sangre periférica mediante cromatografía líquida de alta eficiencia. En la figura 2B, se observa que en ratas con operación simulada el EEP produjo un incremento significativo de la norepinefrina y epinefrina plasmática
file:///G|/archivos%202%202005/html/La%20Angiotensin...0respuesta%20neurohumoral%20frente%20al%20estrés.htm (5 of 9)03/02/2006 08:12:29 a.m.

La Angiotensina II y la respuesta neurohumoral frente al estrés
(2110.1 ± 347.7 pg/ml; 833.6 ± 362.7 pg/ml, respectivamente). Por el contrario, en ratas con nefrectomía bilateral se abolió la respuesta simpática ya que no se produjo el incremento de las catecolaminas plasmática frente al EEP. Se determinaron los niveles de las proteínas plasmáticas, a fin de descartar alteraciones en la hemodilución. No se observaron cambios significativos, en los valores de las proteínas plasmáticas, en ninguno de los experimentos donde se usaron ratas nefrectomizadas con respecto a las ratas con operación simulada (no se muestra).
Ratas con sobrecarga de sodio. En la figura 1B, observamos que la sobrecarga de sodio, modelo experimental que origina una reducción de los niveles circulantes de angiotensina II, produce una disminución significativa de la respuesta vasopresora al EEP, es decir una inhibición de la respuesta simpática ante el estrés con respecto a los controles normosódicos (24 ± 3.7 mmHg) (p<0.0001).
DISCUSIÓN
El estrés agudo provocado por el estímulo eléctrico plantar (EEP) causa la activación del eje simpático-adrenal y el consecuente incremento de los niveles plasmáticos de las catecolaminas, noradrenalina y adrenalina, acompañado de un incremento en la presión arterial y en la frecuencia cardíaca (Lee y col., 1989).
Se ha demostrado que varios modelos de estrés incrementan los niveles circulantes de renina y en consecuencia los niveles de angiotensina II (Jindra y Kvetnansky, 1982). La angiotensina II podría contribuir en la respuesta vasopresora al estrés. En efecto, a nivel periférico, la ANG estimula la liberación de catecolaminas en la médula suprarrenal y en el ganglio simpático (Reid, 1972; Wong y col., 1991). A nivel presináptico estimula la liberación e inhibe la recaptación de la noradrenalina y en los terminales postganglionares, la ANG amplifica la vasoconstricción inducida por la estimulación del receptor a1 postsináptico (Dendorfer y col., 1998). Parte de las acciones periféricas de la ANG II parecen estar mediadas centralmente puesto que la angiotensina II actuaría sobre los órganos circunventriculares, ricos en receptores AT1. Se postula que estas acciones facilitatorias podrían estar mediadas por la interacción de la ANG con el receptor AT1.
Con el fin de demostrar la hipótesis de que la ANG es determinante en la respuesta al estrés, se incremento la actividad simpático mediante la aplicación del estímulo eléctrico plantar a ratas conscientes tratadas con diferentes antagonistas no-peptídicos del receptor AT1. Demostramos que el pretratamiento agudo con losartan, valsartan o eprosartan, inhibió la respuesta vasopresora al EEP. Nuestros resultados no coinciden con los reportados por Ohlstein y col. (1997) quienes demostraron que en ratas descerebradas el losartan, valsartan o irbersartan no afectó la respuesta vasopresora a la estimulación eléctrica de la médula espinal. Esta discrepancia podría atribuirse al hecho que estos experimentos fueron realizados en ratas descerebradas que carecen del control central de la presión arterial mientras que nuestro modelo experimental se mantiene intacto al sistema nervioso simpático. Nuestros resultados, aunado al hecho de que el eprosartan es un compuesto que se sabe que actúa preferentemente a nivel de receptores AT1 presinápticos, indica que la interacción entre la ANG y el SNS ocurre principalmente a nivel de los receptores AT1 localizados en los terminales nervioso simpáticos en donde la acción facilitatoria de la ANG endógena se ejerce y que seria bloqueada mediante el bloqueo del receptor AT1 durante la estimulación simpática inducida por el EEP (Cierco y Israel, 1994, Ohlstein y col., 1997).
Si la presencia de angiotensina II es determinante para la manifestación de la respuesta simpática frente al estrés, la modulación del SRA debería alterar dicha respuesta. Nuestros resultados, obtenidos en ratas depletadas de sodio apoyan esta posibilidad. En estas ratas, el EEP produjo la facilitación de la transmisión simpática que se manifestó con un mayor incremento en la presión arterial media y en la frecuencia cardíaca. Estos resultados coinciden con los de Xu y Brooks (1996), quienes demostraron en ratas depletadas de sodio una elevación de los niveles de angiotensina II con respecto a ratas con una dieta normosódica, y tal aumento se correspondió con un incremento en la actividad nerviosa simpática renal y lumbar. Asimismo, en ratas Wistar descerebradas, en las cuales los niveles de angiotensina II se elevaron 20 veces con respecto a los controles, la infusión de angiotensina II produjo una potenciación significativa de la respuesta presora a la estimulación nerviosa simpática. Este efecto es revertido al practicarles a las ratas descerebradas nefrectomía bilateral (Grant y McGrath, 1988). Aunque en este grupo experimental no se midieron los niveles plasmáticos de las catecolaminas ni de la angiotensina II, otros han demostrado un aumento de ambas sustancias en las mismas condiciones experimentales (Friberg y col., 1990; Xu y Brooks, 1996). Además, se ha reportado una elevación de los niveles del ARNm de la enzima tiroxina hidroxilasa a nivel de las glándulas suprarrenales y de los ganglios simpáticos (Kvetnansky y col.,
file:///G|/archivos%202%202005/html/La%20Angiotensin...0respuesta%20neurohumoral%20frente%20al%20estrés.htm (6 of 9)03/02/2006 08:12:29 a.m.

La Angiotensina II y la respuesta neurohumoral frente al estrés
2004). Por el contrario, la sobrecarga de sodio produce disminución de la actividad de la renina plasmática y de los niveles circulantes de angiotensina II. Igualmente produce una disminución del recambio de las catecolaminas, de la resistencia vascular periférica (Krieger y col., 1990) y de los niveles del ARNm de la enzima tiroxina hidroxilasa (Habecker y col., 2003).
Nuestros resultados muestran que la reducción del SRA se acompaña de una disminución de respuesta presora y de la frecuencia cardíaca ante el EEP. Resultados similares se observaron en gatos con sobrecarga de sodio y tratados con mineralocorticoides (Feuerstein y col., 1977). Estos hallazgos parecen indicar que es necesario un nivel adecuado de los diferentes componentes del SRA para que se exprese la respuesta simpática ante el estrés. Podemos postular que se requiere de la integridad del SRA para que la respuesta simpática se manifieste en toda su plenitud en estados de estrés agudo. El bloqueo del receptor AT1 o la sobrecarga de sodio; situaciones en que se reduce el SRA, ocasionan una atenuación marcada de la respuesta de origen simpático.
En apoyo a nuestra hipótesis están nuestros hallazgos en ratas nefrectomizadas. El modelo de nefrectomía bilateral es un modelo de renina baja. Se sabe que la fuente principal de renina circulante es el riñon. La nefrectomía produce una disminución de la actividad de la renina plasmática y de los niveles circulantes de angiotensina II (Grant y McGrath, 1988; Feuerstein y col., 1977). El bloqueo total observado en la respuesta vasopresora y humoral (incremento de las catecolaminas circulantes) frente al EEP en ratas con 20-22 horas de nefrectomía bilateral apoya nuestra hipótesis y nos permiten plantear que el SRA, a través de su péptido activo, la ANG, ejerce un papel "permisivo" para la respuesta simpática ante un estímulo estresor, como el eléctrico plantar.
Si la angiotensina II juega un papel crítico en la manifestación de la respuesta simpática frente a un estrés agudo, la administración exógena de angiotensina II debería revertir la ausencia de respuesta cardiovascular en las ratas nefrectomizadas. Nuestros resultados en ratas con nefrectomía bilateral, a las que se les administró una dosis subpresora de angiotensina II apoyan esta posibilidad. En efecto, la angiotensina II exógena revirtió el efecto de la nefrectomía bilateral, permitiendo observar incrementos en la presión arterial media frente al EEP. Estos hallazgos nos permite afirmar que la ausencia de respuesta simpática en animales nefrectomizados se debe a la falta de integridad del SRA. En apoyo a nuestros resultados están los reportados por Feuerstein y col., 1977, quienes demostraron en gatos, que la nefrectomía bilateral impide la respuesta adrenomedular (liberación de catecolaminas) a la hemorragia. Igualmente, en gatos con sobrecarga de sodio y administración de mineralocorticoides se reportó un bloqueo del aumento de las catecolaminas en sangre posterior a una hemorragia experimental. Asimismo, en pacientes hipertensos con insuficiencia renal crónica se ha demostrado una disminución de las descargas simpáticas luego de la nefrectomía bilateral asociada con disminución de la presión arterial media (Converse y col., 1992).
En conclusión, nuestros resultados indican que para la plena expresión de la respuesta simpática frente al estrés agudo es necesaria la integridad del sistema renina-angiotensina y la estimulación del los receptores AT1 presinápticos.
REFERENCIAS BIBLIOGRÁFICAS
1. Anton AH, Sayre DF. A study of the factors affecting the aluminum oxidetrihydrozyindole procedure for the analysis of catecholamines. J Pharmacol Exp Ther 1962; 138: 360-375.
2. Armando I, Carracza A, Nishimura Y, Hoe K, Barontini M, Terron JA, Falcon-Neri A, Ito T, Juorio AV, Saavedra JM. Peripheral administration of an angiotensin II AT1 receptor antagonist decreases the hypothalamic-pituitary-adrenal response to isolation stress. Endocrinology 2001; 142: 3880-3889.
3. Brooks VL, Osborn JW. Hormonal-sympathetic interactions in long-term regulation of arterial pressure: an hypothesis. Am J Physiol 1995; 268(6 Pt 2): R1343-R1358.
4. Cierco M, Israel A. Role of AT1 receptor in the cardiovascular response to footshock. European J Pharmacol 1994; 251: 103-106.
file:///G|/archivos%202%202005/html/La%20Angiotensin...0respuesta%20neurohumoral%20frente%20al%20estrés.htm (7 of 9)03/02/2006 08:12:29 a.m.

La Angiotensina II y la respuesta neurohumoral frente al estrés
5. Converse RL, Jacobsen TN, Toto RD, Jost CMT, Consentino F, Fouad-Tarazi F, Victor RG. Sympathetic overactivity in patients with chronic renal failure. New Engl J Med 1992; 327: 1912-1918.
6. Dendorfer A, Raasch W, Tempel K, Dominiak P. Interactions between the renin-angiotensin system (RAS) and the sympathetic system. Basic Res Cardiol 1998; 93: 24-29.
7. Feuerstein G, Boonyaviroj P, Gutman Y. Renin-angiotensin mediation of adrenal catecholamine secretion induced by hemorrhage. European J Pharmacol 1977; 44: 131-142.
8. Friberg P, Meredith I, Jennings G, Lambert G, Fazio V, Esle M. Evidence for increased renal norepinephrine overflow during sodium restriction in humans. Hypertension 1990; 16(2): 121-30.
9. Grant TL, McGrath JC. Interactions between angiotensin II, sympathetic nerve-mediated pressor response and cyclo-oxygenase products in pithed rat. Br J Pharmacol 1988; 95: 1220-1228.
10. Habecker BA, Grygielko ET, Huhtala TA, Foote B, Brooks VL. Ganglionic tyrosine hydroxylase and norepinephrine transporter are decreased by increased sodium chloride in vivo and in vitro. Auton Neurosci 2003; 107: 85-98.
11. Israel A, Cierco M, Barbella Y. Role of angiotensin II receptor subtypes, AT1 and AT2, on fluid and electrolite metabolism and on the cardiovascular response to footshock. Progress in Pharmacology and Clinical Pharmacology, Eds. Velasco M and Israel A, Gustav Fischer Verlag-Stuttgard-Jena-N.Y., 1994; 10: 1-12.
12. Israel A, Sosa B. Angiotensin II supports sympathetically mediated vasopressor response to footshock-stress. J. Human Hypertension 2002; 16 (Suppl 1): S84-S88.
13. Jindra AJR, Kvetnansky R. Stress-induced activation of inactive renin. J. Biol. Chem. 1982; 257: 5997-5999.
14. Krieger JE, Liard JF, Cowley AW. Hemodynamics, fluid volume, and hormoral responses to chronic high-salt intake in dogs. Am J Physiol 1990; 259 (Heart Circ Physiol 28): H1629-H1636.
15. Kvetnansky R, Micutkova L, Rychkova N, Kubovcakova L, Mravec B, Filipenko M, Sabban EL, Krizanova O. Quantitative evaluation of catecholamine enzymes gene expression in adrenal medulla and sympathetic Ganglia of stressed rats. Ann N Y Acad Sci. 2004; 1018: 356-369.
16. Lee J, Konarska M, McCarty R. Physiological responses to acute stress in alloxan and streptozotocin diabetic rats. Physiol Behav 1989; 45: 483-489.
17. Ohlstein EH, Brooks DP, Feuerstein GZ, Ruffolo Jr RR. Inhibition of sympathetic outflow by angiotensin II receptor antagonist, eprosartan, but not losartan, valsartan or ibersartan: relationship to differences in prejunctional angiotensin II receptor blockade. Pharmacology 1997; 55: 244-251.
18. Reid IA. Interactions between Ang II, sympathetic nervous system, and baroreceptor reflexes in regulation of blood pressure. Am J Physiol 1992; 262: E763-E778.
19. Saavedra JM. Brain and pituitary angiotensin. Endocrin Rev 1992; 13: 329-380.
20. Seltzer A, Bregonzio C, Armando I, Baiardini G, Saavedra JM. Oral administration of an AT1 receptor antagonist prevents the central effects of angiotensin II in spontaneously hypertensive rats. Brain Research 2004; 1028: 9-18.
file:///G|/archivos%202%202005/html/La%20Angiotensin...0respuesta%20neurohumoral%20frente%20al%20estrés.htm (8 of 9)03/02/2006 08:12:29 a.m.

La Angiotensina II y la respuesta neurohumoral frente al estrés
21. Wong PB, Hart SD, Timmermans PBMWM. Effect of angiotensin II antagonism on canine renal sympathetic nerve function. Hypertension 1991; 17: 1127-1134.
22. Wong PB, Bernard R, Timmermans PBMWM. Effect of blocking angiotensin II receptor subtype on rat sympathetic nerve function. Hypertension 1992; 19: 663-667.
23. Xu l, Brooks VL. ANG II chronically supports renal and lumbar sympathetic activity in sodium-deprived, conscious rats. Am J Physiol 1996; 271 (Heart Circ. Physiol. 40): H2591-H2598.
24. Zimmerman BG, Kraft E. Blockade by saralasin of adrenergic potentiation induced by renin-angiotensin system. J Pharmacol Exp Ther 1979; 210(1): 101-105.
file:///G|/archivos%202%202005/html/La%20Angiotensin...0respuesta%20neurohumoral%20frente%20al%20estrés.htm (9 of 9)03/02/2006 08:12:29 a.m.