
Kinetic and Spectral Resolution of Multiple Nonphotochemical Quenching Components in Arabidopsis...
14
Kinetic and Spectral Resolution of Multiple Nonphotochemical Quenching Components in Arabidopsis Leaves 1[C] Petar H. Lambrev, Manuela Nilkens, Yuliya Miloslavina, Peter Jahns, and Alfred R. Holzwarth* Max-Planck-Institut fu ¨r Bioanorganische Chemie, 45470 Mulheim an der Ruhr, Germany (P.H.L., Y.M., A.R.H.); and Institut fu ¨ r Biochemie der Pflanzen, Heinrich-Heine Universita ¨t Du ¨ sseldorf, 40225 Duesseldorf, Germany (M.N., P.J.) Using novel specially designed instrumentation, fluorescence emission spectra were recorded from Arabidopsis (Arabidopsis thaliana) leaves during the induction period of dark to high-light adaptation in order to follow the spectral changes associated with the formation of nonphotochemical quenching. In addition to an overall decrease of photosystem II fluorescence (quenching) across the entire spectrum, high light induced two specific relative changes in the spectra: (1) a decrease of the main emission band at 682 nm relative to the far-red (750–760 nm) part of the spectrum (DF 682 ); and (2) an increase at 720 to 730 nm (DF 720 ) relative to 750 to 760 nm. The kinetics of the two relative spectral changes and their dependence on various mutants revealed that they do not originate from the same process but rather from at least two independent processes. The DF 720 change is specifically associated with the rapidly reversible energy-dependent quenching. Comparison of the wild-type Arabidopsis with mutants unable to produce or overexpressing the PsbS subunit of photosystem II showed that PsbS was a necessary component for DF 720 . The spectral change DF 682 is induced both by energy-dependent quenching and by PsbS-independent mechanism(s). A third novel quenching process, independent from both PsbS and zeaxanthin, is activated by a high turnover rate of photosystem II. Its induction and relaxation occur on a time scale of a few minutes. Analysis of the spectral inhomogeneity of nonphotochemical quenching allows extraction of mechanistically valuable information from the fluores- cence induction kinetics when registered in a spectrally resolved fashion. One of the most important photoprotective mecha- nisms against high-light (HL) stress in photosynthetic organisms is the nonphotochemical quenching (NPQ) of excitation energy, which is mostly due to thermal deactivation of pigment excited states in the antenna of PSII. There exist a number of literature reviews on the subject (Demmig-Adams and Adams, 1992; Horton et al., 1996; Horton and Ruban, 1999, 2005; Niyogi, 1999, 2000; Mu ¨ ller et al., 2001; Golan et al., 2004; Krause and Jahns, 2004). Chlorophyll (Chl) fluo- rescence, and in particular pulse amplitude-modulated (PAM) fluorometry as introduced by Schreiber et al. (1986), has become by far the dominant technique to measure NPQ in leaves, chloroplasts, and intact mi- croorganisms (Krause and Weis, 1991; Govindjee, 1995; Maxwell and Johnson, 2000; Krause and Jahns, 2003; Schreiber, 2004), more recently often combined with specific NPQ mutant studies (Golan et al., 2004; Kalituho et al., 2006, 2007; Dall’Osto et al., 2007). In this technique, periodic saturating light pulses are applied, superimposed on the continuous actinic irradiation applied to induce NPQ, in order to transiently close the PSII reaction centers (RCs). Since the photochem- istry contribution (photochemical quenching) is thus brought to zero, the method allows us to follow the dynamics of the NPQ development and relaxation by fluorescence in a relatively simple manner (Krause and Jahns, 2003, 2004). Mostly based on its relaxation kinetics, NPQ has been divided technically into the three kinetic compo- nents qE, qT, and qI, for the rapid, middle, and slow phases of relaxation (Horton and Hague, 1988), ini- tially attributed to energy-dependent quenching, state transitions, and photoinhibitory quenching (Quick and Stitt, 1989). The rapidly forming and reversible part of NPQ, qE, is the most thoroughly studied. It is well established that this type of quenching is a finely regulated process in which the main governing factors are the proton gradient across the chloroplast thy- lakoid membrane, DpH (Wraight and Crofts, 1970; Briantais et al., 1979), the xanthophyll cycle (i.e. con- version of violaxanthin to antheraxanthin and zeaxan- thin [Zx]; Demmig et al., 1987; Demmig-Adams, 1990; Demmig-Adams and Adams, 1992), and the action of the PsbS protein (Funk et al., 1995; Li et al., 2000, 2004; Niyogi et al., 2005). The actual molecular mechanism is 1 This work was supported by the Deutsche Forschungsgemein- schaft, Sonderforschungsbereich 663. * Corresponding author; e-mail holzwarth@mpi-muelheim. mpg.de. The author responsible for distribution of materials integral to the findings presented in this article in accordance with the policy described in the Instructions for Authors (www.plantphysiol.org) is: Alfred R. Holzwarth ([email protected]). [C] Some figures in this article are displayed in color online but in black and white in the print edition. www.plantphysiol.org/cgi/doi/10.1104/pp.109.148213 Plant Physiology Ò , March 2010, Vol. 152, pp. 1611–1624, www.plantphysiol.org Ó 2009 American Society of Plant Biologists 1611 www.plant.org on April 26, 2016 - Published by www.plantphysiol.org Downloaded from Copyright © 2010 American Society of Plant Biologists. All rights reserved.
-
Upload
uni-duesseldorf -
Category
Documents
-
view
4 -
download
0
Transcript of Kinetic and Spectral Resolution of Multiple Nonphotochemical Quenching Components in Arabidopsis...

Kinetic and Spectral Resolution of MultipleNonphotochemical Quenching Components inArabidopsis Leaves1[C]
Petar H. Lambrev, Manuela Nilkens, Yuliya Miloslavina, Peter Jahns, and Alfred R. Holzwarth*
Max-Planck-Institut fur Bioanorganische Chemie, 45470 Mulheim an der Ruhr, Germany (P.H.L., Y.M.,A.R.H.); and Institut fur Biochemie der Pflanzen, Heinrich-Heine Universitat Dusseldorf, 40225 Duesseldorf,Germany (M.N., P.J.)
Using novel specially designed instrumentation, fluorescence emission spectra were recorded from Arabidopsis (Arabidopsisthaliana) leaves during the induction period of dark to high-light adaptation in order to follow the spectral changes associatedwith the formation of nonphotochemical quenching. In addition to an overall decrease of photosystem II fluorescence(quenching) across the entire spectrum, high light induced two specific relative changes in the spectra: (1) a decrease of themain emission band at 682 nm relative to the far-red (750–760 nm) part of the spectrum (DF682); and (2) an increase at 720 to 730nm (DF720) relative to 750 to 760 nm. The kinetics of the two relative spectral changes and their dependence on various mutantsrevealed that they do not originate from the same process but rather from at least two independent processes. The DF720 changeis specifically associated with the rapidly reversible energy-dependent quenching. Comparison of the wild-type Arabidopsiswith mutants unable to produce or overexpressing the PsbS subunit of photosystem II showed that PsbS was a necessarycomponent for DF720. The spectral change DF682 is induced both by energy-dependent quenching and by PsbS-independentmechanism(s). A third novel quenching process, independent from both PsbS and zeaxanthin, is activated by a high turnoverrate of photosystem II. Its induction and relaxation occur on a time scale of a few minutes. Analysis of the spectralinhomogeneity of nonphotochemical quenching allows extraction of mechanistically valuable information from the fluores-cence induction kinetics when registered in a spectrally resolved fashion.
One of the most important photoprotective mecha-nisms against high-light (HL) stress in photosyntheticorganisms is the nonphotochemical quenching (NPQ)of excitation energy, which is mostly due to thermaldeactivation of pigment excited states in the antennaof PSII. There exist a number of literature reviewson the subject (Demmig-Adams and Adams, 1992;Horton et al., 1996; Horton and Ruban, 1999, 2005;Niyogi, 1999, 2000; Muller et al., 2001; Golan et al.,2004; Krause and Jahns, 2004). Chlorophyll (Chl) fluo-rescence, and in particular pulse amplitude-modulated(PAM) fluorometry as introduced by Schreiber et al.(1986), has become by far the dominant technique tomeasure NPQ in leaves, chloroplasts, and intact mi-croorganisms (Krause and Weis, 1991; Govindjee,1995; Maxwell and Johnson, 2000; Krause and Jahns,2003; Schreiber, 2004), more recently often combined
with specific NPQ mutant studies (Golan et al., 2004;Kalituho et al., 2006, 2007; Dall’Osto et al., 2007). In thistechnique, periodic saturating light pulses are applied,superimposed on the continuous actinic irradiationapplied to induce NPQ, in order to transiently closethe PSII reaction centers (RCs). Since the photochem-istry contribution (photochemical quenching) is thusbrought to zero, the method allows us to follow thedynamics of the NPQ development and relaxation byfluorescence in a relatively simple manner (Krauseand Jahns, 2003, 2004).
Mostly based on its relaxation kinetics, NPQ hasbeen divided technically into the three kinetic compo-nents qE, qT, and qI, for the rapid, middle, and slowphases of relaxation (Horton and Hague, 1988), ini-tially attributed to energy-dependent quenching, statetransitions, and photoinhibitory quenching (Quickand Stitt, 1989). The rapidly forming and reversiblepart of NPQ, qE, is the most thoroughly studied. It iswell established that this type of quenching is a finelyregulated process in which the main governing factorsare the proton gradient across the chloroplast thy-lakoid membrane, DpH (Wraight and Crofts, 1970;Briantais et al., 1979), the xanthophyll cycle (i.e. con-version of violaxanthin to antheraxanthin and zeaxan-thin [Zx]; Demmig et al., 1987; Demmig-Adams, 1990;Demmig-Adams and Adams, 1992), and the action ofthe PsbS protein (Funk et al., 1995; Li et al., 2000, 2004;Niyogi et al., 2005). The actual molecular mechanism is
1 This work was supported by the Deutsche Forschungsgemein-schaft, Sonderforschungsbereich 663.
* Corresponding author; e-mail [email protected].
The author responsible for distribution of materials integral to thefindings presented in this article in accordance with the policydescribed in the Instructions for Authors (www.plantphysiol.org) is:Alfred R. Holzwarth ([email protected]).
[C] Some figures in this article are displayed in color online but inblack and white in the print edition.
www.plantphysiol.org/cgi/doi/10.1104/pp.109.148213
Plant Physiology�, March 2010, Vol. 152, pp. 1611–1624, www.plantphysiol.org � 2009 American Society of Plant Biologists 1611 www.plant.org on April 26, 2016 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2010 American Society of Plant Biologists. All rights reserved.

still unknown, although there is no shortage of hy-potheses and proposed quencher candidates: energytransfer from Chl to Zx in the major light-harvestingcomplex (LHCII; Frank et al., 2000); electron transferfrom a carotenoid to Chl forming a Zx-Chl or lutein-Chl charge-transfer state (Holt et al., 2005; Avensonet al., 2009); direct or indirect quenching by the PsbSprotein (Li et al., 2000; Niyogi et al., 2005); energytransfer from Chl to lutein in LHCII (Horton et al.,1991; Ruban et al., 2007) linked to the aggregation of ora conformational change in LHCII; and last but notleast, a far-red (FR) light-emitting quenched Chl-Chlcharge-transfer state formed by the aggregation ofLHCII (Miloslavina et al., 2008). Quenching in the PSIIRC has also been proposed (Weis and Berry, 1987;Finazzi et al., 2004; Huner et al., 2005; Ivanov et al.,2008) as an additional type of Zx-independent quench-ing. Alternatively, it has been suggested that quench-ing by lutein can complement the Zx-dependentquenching (Niyogi et al., 2001; Li et al. 2009). Johnsonet al. (2009) have recently given support to the notionthat both Zx-dependent and Zx-independent quench-ing originate from the same PsbS-dependent mecha-nism, which is modulated by Zx (Crouchman et al.,2006).
While the rapidly relaxing phase qE is now wellcharacterized in its dependence on the various factors,the much slower qT and qI phases are still controver-sial, and each of them may have contributions frommore than one mechanism. The qI component hasbeen traditionally attributed to photoinhibition of PSII(Somersalo and Krause, 1988), associated with coordi-nated degradation and repair of the photosystem(Powles and Bjorkman, 1982; Kyle, 1987; Krause,1988; Aro et al., 1993; Long et al., 1994; Murata et al.,2007). Lately, though, it is more widely accepted thatunder most conditions the photoinhibition is low andqI, like qE, is a result of thermal deactivation of excitedstates. Different hypotheses have been put forward toaccount for its seeming irreversibility: persistent trans-membrane DpH (Gilmore and Yamamoto, 1992), stableprotonation of proteins (Horton et al., 1994), accumu-lation of inactive PSII reaction centers (Briantais et al.,1992; Schansker and van Rensen, 1999), or stablebinding of Zx to CP29 (Farber et al., 1997). The con-nection of the qT phase with state transitions has beendoubted as well, and in fact it is now thought that thefraction of energy redistributed from PSI to PSII un-der high-light conditions is negligible (Walters andHorton, 1991, 1993) and that the qT must have adifferent origin or that it has erroneously been ascribedas NPQ (Schansker et al., 2006).
Along with the large amount of contradictory evi-dence on the nature and location of the NPQ quench-ing site(s), the question of whether the light-inducedreversible NPQ represents one single mechanism ofdeexcitation located in a single site brought about bythe combined action of PsbS and Zx (Johnson et al.,2009) or whether it comprises several parallel andlargely independent mechanisms acting on different
parts of the PSII antenna has not been finally an-swered. One way to answer this question might be tocarefully examine the spectral properties of NPQ-related fluorescence changes. Quenching in differentlocations of the PSII antenna or with different mech-anisms might give rise to a differential quenching invarious parts of the PSII antenna that might affect thePSII fluorescence spectra in different ways. This ap-pears possible, since the various pigment-proteincomplexes of the photosynthetic apparatus haveslightly different absorption and emission spectra(Holzwarth, 1991; Holzwarth and Roelofs, 1992).However, in the vast majority of modulated Chl fluo-rescence instrumentation, including the most widelyused PAM fluorometer (Schreiber et al., 1986), thesignal is integrated over a broad wavelength range,usually covering the whole range of 710 nm or greater.This integration over the long-wave part of the spec-trum has several undesirable consequences and isassociated with the unnecessary loss of available in-formation. For example, the fluorescence of PSII peaksin the region of 680 to 685 nm, whereas beyond 700nm, the PSII fluorescence intensity drops to less than20% of its peak intensity. In contrast, the fluorescenceof intact PSI complexes is dominant in the regionabove 710 nm (Haehnel et al., 1982; Karukstis andSauer, 1983; Holzwarth et al., 1985; Holzwarth, 1986;Slavov et al., 2008). Thus, the widely used instrumen-tation measures the NPQ parameters in a region withreduced PSII contribution and relatively high PSIcontribution to total fluorescence, despite the factthat NPQ is generally considered to be primarily aPSII phenomenon. Only in a few studies has thefluorescence in the red and the FR region been sepa-rated in order to evaluate the contribution of PSI andits influence on the NPQ parameters (Genty et al.,1990; Peterson et al., 2001). NPQ might also shift thefluorescence properties of the PSII antenna complexesor give rise to entirely new fluorescing components(Miloslavina et al., 2008). This would remain unde-tected if the NPQ fluorescence changes are not re-solved in the spectral domain. It follows from theseconsiderations that a great deal of insight into the NPQmechanisms and locations may be gained if the spec-tral dimension is added to the NPQ fluorescence char-acterization. Among the many advantages of suchan approach, one would then be able to distinguishwhether NPQ simply leads to a uniform decrease ofPSII fluorescence across the emission range or whetherthis decrease is nonuniform, localized in specificpigment protein complexes, and/or whether new fluo-rescing species are actually being produced in theNPQ process.
The HL-induced NPQ effects on the leaf fluores-cence spectra have often been studied also at lowtemperature, where the differentiation between pig-ment sites is better (Krause et al., 1983; Demmig andBjorkman, 1987; Ruban and Horton, 1994). However,the possibility to resolve the kinetics of NPQ develop-ment and relaxation is largely lost when performing
Lambrev et al.
1612 Plant Physiol. Vol. 152, 2010 www.plant.org on April 26, 2016 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2010 American Society of Plant Biologists. All rights reserved.

the measurements at low temperatures. The 77 Kspectra of leaves and thylakoid membranes are char-acterized by three main peaks, F685, F695, and F730,believed to originate predominantly from Chl a inCP47 of PSII, a specific Chl in CP43 of PSII, and PSI,respectively (Satoh and Butler, 1978; van Dorssen et al.,1987; Andrizhiyevskaya et al., 2005; Komura et al.,2007). Fluorescence from the major LHCII peaks at 680nm (Rijgersberg and Amesz, 1978) and from the PSIIreaction center Chls at 683 nm (Roelofs et al., 1993;Andrizhiyevskaya et al., 2005). Low-temperature stud-ies on the effects of HL irradiation are confined to thechanges in the FR-to-red fluorescence ratio, which arethe result of the quenching of PSII fluorescence orenergy redistribution between the photosystems (statetransitions). Ruban and Horton (1994) have shownthat photochemical quenching in Guzmania is maximalat 688 nm, whereas nonphotochemical processesquench preferentially at 683 and 698 nm.In this study, we undertook a detailed investigation
of the NPQ-associated spectral changes in the fluores-cence spectra of Arabidopsis (Arabidopsis thaliana)measured at room temperature (RT) and at 77 K. Itfollows from the above discussion that deeper insightinto the mechanisms of NPQ processes may be gainedby combining the kinetic and the spectral informationof the fluorescence changes occurring in NPQ. For thispurpose, we developed a multiwavelength spectrom-eter with parallel detection, allowing us to follow theentire time-dependent fluorescence spectra of leavesduring the induction and relaxation phases of NPQwith high sensitivity.Specific questions to be addressed in this study are
the following. Are there more than one NPQ processesand NPQ sites? Are these processes occurring in alinked fashion or are they independent? How do theydepend on the various cofactors known to affect NPQ,in particular regarding the roles of PsbS and Zx? Usingthis novel approach of adding the spectral informationto the NPQ fluorescence changes, we discovered spe-cific spectral changes associated with different NPQcomponents. By comparing the effects measured onvarious NPQ mutants of Arabidopsis, it is possible toassign these NPQ components to specific quenchingprocesses. The results provide evidence that the totalNPQ is a combination of several parallel and largelyindependent processes, likely occurring at differentlocations in the photosynthetic apparatus.
RESULTS
77 K Fluorescence Spectra
Fluorescence emission spectra measured at 77 Kfrom dark-adapted wild-type Arabidopsis leaves andfrom leaves irradiated for 30 min with 600 mmolphotons m22 s21 red light (620 nm) are shown inFigure 1. To facilitate detailed comparison of theirshape, all spectra were normalized. Since NPQ is
supposed to alter the properties of PSII rather thanPSI, we wanted to use PSI fluorescence as a referencefor normalization to better visualize the changes inPSII. According to the notion that at shorter wave-lengths the emission originates from PSII and at longerwavelengths from PSI, we normalized the spectra at760 nm, near the FR tail of the spectra, where thecontribution from PSII is minimal and that from PSI ismaximal. More precisely, the spectra were normalizedto the averaged intensity between 755 and 765 nm inorder to further reduce the already low noise in thedata (the signal-to-noise ratio at 760 nm is about 300:1)and thus avoid inaccurate normalization. The 77 Kfluorescence emission spectra of wild-type leavesshow peaks at 683, 691, and 730 nm. The relative ratiobetween them has been reported to be strongly de-pendent on the leaf anatomy, particularly the thicknessand the Chl content, which determine its opticalproperties, and also on the angle of excitation anddetection (Weis, 1985). However, for leaves at similarphysiological states at a fixed orientation, we found
Figure 1. A, 77 K fluorescence emission spectra, normalized at 760 nm,of dark-adapted Arabidopsis wild-type leaves (solid line) and leavespreilluminated for 30 min with red light (photon flux density 600 mmolm22 s21). The curves represent averages from four to six measurementson different leaves. The SE is shown for selected wavelengths. B,Difference spectrum (light minus dark). rel.u., Relative units.
Kinetic and Spectral Resolution of NPQ Components
Plant Physiol. Vol. 152, 2010 1613 www.plant.org on April 26, 2016 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2010 American Society of Plant Biologists. All rights reserved.

the spectral differences to be insignificant. FollowingHL treatment of the wild-type leaves, the spectrarevealed characteristic and reproducible differences:a decrease of fluorescence in the PSII wavelengthrange (less than 700 nm) and an increase in the FRrange (greater than 710 nm). The light-minus-darkdifference spectra (Fig. 1B) show two negative peaks,at 683 and 691 nm, matching the peaks of the emissionspectra, and a positive band appearing around 727 nm(i.e. somewhat blue shifted to the major PSI emissionpeak; 730 nm). In some cases, a negative differenceband at 715 nm was also observed (only a shoulder inFig. 1B).
Whereas the negative difference bands are undoubt-edly associated with quenching of the PSII antennafluorescence (F685 and F695), the origin of the positive730-nm band is less clear. In order to check whether itsappearance corresponds to the kinetics of NPQ, leaveswere rapidly frozen in the light at different times ofHL irradiation and 77 K fluorescence spectra were re-corded. Furthermore, spectra were taken after keepingpreilluminated leaves for different times in darkness.The resulting time-dependent changes of the ratioF730/F760 are shown in Figure 2. The ratio rose withinthe first several minutes of irradiation toward a max-imum value and declined back after a few minutesof darkness. This is a typical behavior for energy-dependent quenching (qE). However, because thesemeasurements have to be carried out on differentleaves, the level of accuracy is not high enough toquantitatively compare the kinetics of the 77 K fluo-rescence changes at different wavelengths.
RT Fluorescence Spectra
RT measurements allow for a far better quantifica-tion of the relative changes in the fluorescence spectrabefore and after illumination when they are registeredfrom the same leaf without changing its orientation inthe optical path. To exploit this possibility, we con-structed a special dual-light-emitting diode (LED)instrument, based on an Ocean Optics USB2000 CCDspectrometer (see “Materials and Methods”). Since thespectrometer is able to register a full emission spec-trum with high signal-to-noise ratio in less than 10 ms,the instrument can follow fast light-induced spectralchanges with remarkable sensitivity. The dual-LEDmode enables probing of the fluorescence spectra andkinetic development from closed PSII RCs either un-der variable actinic light or in the absence of it. Thisallows us, on the one hand, to correlate the spectralchanges with kinetic components of NPQ and, on theother hand, to also discriminate the processes usingdifferent excitation wavelengths if necessary.
The NPQ kinetics, measured with a single actinic/excitation source, is presented in Figure 3 as the timeand wavelength dependence of the NPQ parameter,calculated as NPQ = F(t0)/F(t) – 1 (Briantais et al., 1979;Bilger and Bjorkman, 1990). Along the time axis, thefigure shows the known kinetics of NPQ formation in
the wild-type Arabidopsis and the mutants npq4 andnpq1. The PsbS-deficient npq4 plants generate muchless NPQ as compared with the wild type, and thekinetics lacks the fast NPQ phase, whereas in npq1,which cannot form Zx, most of the NPQ is formed inthe fast initial phase, attributed to the action of PsbS.Along the wavelength axis, the figure reveals thesignificant nonhomogeneity of the NPQ parameterwith respect to the detection wavelength. A distinctive“valley” in the surface plot between 700 and 730 nm isobserved for wild-type leaves but is lacking in the npq4mutant and is smaller in npq1.
The normalized (to the average intensity at 745–755nm) emission spectra measured in dual-LED modefrom a wild-type leaf in the dark-adapted state, after30 min of irradiation with 600 mmol photons m22 s21
red light and after 5 min of redarkening, are shown inFigure 4. The RT spectra are characterized by a majorPSII band with a single maximum at 685 nm and alower intensity broad band in the 710- to 740-nm
Figure 2. Dependence of the fluorescence ratio F730/F760 on theduration of irradiation with 600 mmol m22 s21 red light (A) or on thetime during redarkening (B) of wild-type Arabidopsis leaves preillumi-nated for 15 min. The parameter is calculated from fluorescenceemission spectra registered at 77 K after rapid freezing of the leaves.Each value represents an average of two to six leaves, and error barsrepresent SE.
Lambrev et al.
1614 Plant Physiol. Vol. 152, 2010 www.plant.org on April 26, 2016 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2010 American Society of Plant Biologists. All rights reserved.

range, which corresponds to the vibrational tails of thetwo photosystems and to the “red” Chls of PSI. Themost pronounced HL-induced effect on the spectralshape is, as expected, the decrease of the main PSIIband. The light-minus-dark difference spectra have anegative peak at 682 nm. In the FR region, the spectraof light-adapted leaves (dashed line in Fig. 4A) show awell-resolved increase above the dark-adapted spec-tra. In the light-minus-dark difference spectra, this isobserved as a positive band with a maximum at 720
nm. Five minutes after switching off the actinic light,the main fluorescence band recovered part of itsintensity (dotted line in Fig. 4A). Interestingly, at thesame time, the observed increase around 720 nmdisappeared completely. This is, to our knowledge,the first indication that the two light-induced effects(i.e. the decrease of F682 and the increase of F720) are notalways matched. The effect is clearly demonstrated bythe difference spectra in Figure 4B. The light-minus-redark difference spectrum (dashed line) shows thatF720 is a fast-relaxing component and the redark-minus-dark spectrum (dotted line) shows the com-plete absence of the F720 difference band after 5 min ofdark, the only difference remaining in the F682 band.
We compared the described light-induced spectralchanges in several Arabidopsis NPQ mutants: npq4,
Figure 3. Three-dimensional plots of the NPQ parameter [NPQ = F(t0)/F(t) – 1] of leaves of Arabidopsis wild-type (A), npq4 (B), and npq1 (C)plants as a function of emission wavelength and irradiation time.Actinic/excitation light was 620 nm, and photon flux density was 1,000mmol m22 s21. Fluorescence spectra were recorded every 2 s using theactinic light as excitation source. [See online article for color version ofthis figure.]
Figure 4. A, RT fluorescence emission spectra, normalized at 750 nm,of wild-type Arabidopsis leaves in the dark-adapted state after 30min ofillumination (actinic light of 620 nm, photon flux density of 600 mmolm22 s21) and after 5 min of redarkening. Additional pulses of blue light(0.2 s, photon flux density of 1,500 mmol m22 s21) were applied every60 s to detect the fluorescence spectra from closed PSII reactioncenters. rel.u., Relative units. B, Relative difference spectra (in per-centage relative to the emission at 750 nm): light minus dark, lightminus redark, and redark minus dark. The plots show results of a singleleaf measurement. Error bars in B represent the SE at 682 and 720 nm fornine measured leaves.
Kinetic and Spectral Resolution of NPQ Components
Plant Physiol. Vol. 152, 2010 1615 www.plant.org on April 26, 2016 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2010 American Society of Plant Biologists. All rights reserved.
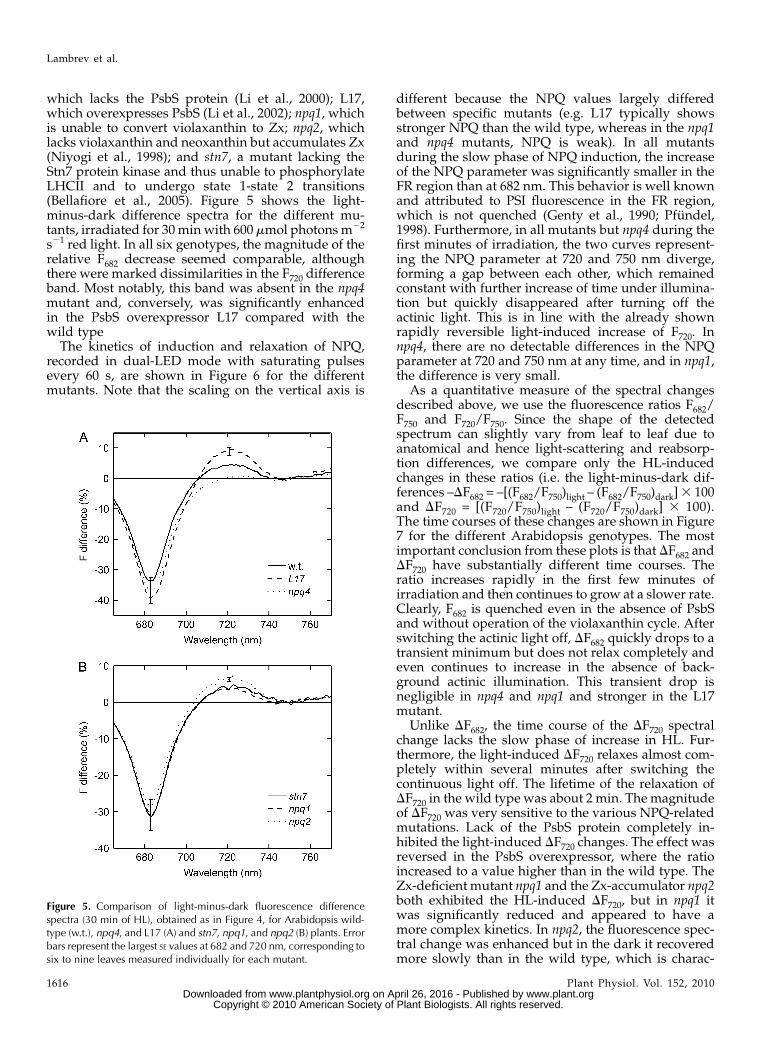
which lacks the PsbS protein (Li et al., 2000); L17,which overexpresses PsbS (Li et al., 2002); npq1, whichis unable to convert violaxanthin to Zx; npq2, whichlacks violaxanthin and neoxanthin but accumulates Zx(Niyogi et al., 1998); and stn7, a mutant lacking theStn7 protein kinase and thus unable to phosphorylateLHCII and to undergo state 1-state 2 transitions(Bellafiore et al., 2005). Figure 5 shows the light-minus-dark difference spectra for the different mu-tants, irradiated for 30minwith 600 mmol photonsm22
s21 red light. In all six genotypes, the magnitude of therelative F682 decrease seemed comparable, althoughthere were marked dissimilarities in the F720 differenceband. Most notably, this band was absent in the npq4mutant and, conversely, was significantly enhancedin the PsbS overexpressor L17 compared with thewild type
The kinetics of induction and relaxation of NPQ,recorded in dual-LED mode with saturating pulsesevery 60 s, are shown in Figure 6 for the differentmutants. Note that the scaling on the vertical axis is
different because the NPQ values largely differedbetween specific mutants (e.g. L17 typically showsstronger NPQ than the wild type, whereas in the npq1and npq4 mutants, NPQ is weak). In all mutantsduring the slow phase of NPQ induction, the increaseof the NPQ parameter was significantly smaller in theFR region than at 682 nm. This behavior is well knownand attributed to PSI fluorescence in the FR region,which is not quenched (Genty et al., 1990; Pfundel,1998). Furthermore, in all mutants but npq4 during thefirst minutes of irradiation, the two curves represent-ing the NPQ parameter at 720 and 750 nm diverge,forming a gap between each other, which remainedconstant with further increase of time under illumina-tion but quickly disappeared after turning off theactinic light. This is in line with the already shownrapidly reversible light-induced increase of F720. Innpq4, there are no detectable differences in the NPQparameter at 720 and 750 nm at any time, and in npq1,the difference is very small.
As a quantitative measure of the spectral changesdescribed above, we use the fluorescence ratios F682/F750 and F720/F750. Since the shape of the detectedspectrum can slightly vary from leaf to leaf due toanatomical and hence light-scattering and reabsorp-tion differences, we compare only the HL-inducedchanges in these ratios (i.e. the light-minus-dark dif-ferences –DF682 = –[(F682/F750)light – (F682/F750)dark]3 100and DF720 = [(F720/F750)light – (F720/F750)dark] 3 100).The time courses of these changes are shown in Figure7 for the different Arabidopsis genotypes. The mostimportant conclusion from these plots is that DF682 andDF720 have substantially different time courses. Theratio increases rapidly in the first few minutes ofirradiation and then continues to grow at a slower rate.Clearly, F682 is quenched even in the absence of PsbSand without operation of the violaxanthin cycle. Afterswitching the actinic light off, DF682 quickly drops to atransient minimum but does not relax completely andeven continues to increase in the absence of back-ground actinic illumination. This transient drop isnegligible in npq4 and npq1 and stronger in the L17mutant.
Unlike DF682, the time course of the DF720 spectralchange lacks the slow phase of increase in HL. Fur-thermore, the light-induced DF720 relaxes almost com-pletely within several minutes after switching thecontinuous light off. The lifetime of the relaxation ofDF720 in the wild type was about 2 min. The magnitudeof DF720 was very sensitive to the various NPQ-relatedmutations. Lack of the PsbS protein completely in-hibited the light-induced DF720 changes. The effect wasreversed in the PsbS overexpressor, where the ratioincreased to a value higher than in the wild type. TheZx-deficient mutant npq1 and the Zx-accumulator npq2both exhibited the HL-induced DF720, but in npq1 itwas significantly reduced and appeared to have amore complex kinetics. In npq2, the fluorescence spec-tral change was enhanced but in the dark it recoveredmore slowly than in the wild type, which is charac-
Figure 5. Comparison of light-minus-dark fluorescence differencespectra (30 min of HL), obtained as in Figure 4, for Arabidopsis wild-type (w.t.), npq4, and L17 (A) and stn7, npq1, and npq2 (B) plants. Errorbars represent the largest SE values at 682 and 720 nm, corresponding tosix to nine leaves measured individually for each mutant.
Lambrev et al.
1616 Plant Physiol. Vol. 152, 2010 www.plant.org on April 26, 2016 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2010 American Society of Plant Biologists. All rights reserved.

teristic for the kinetics of NPQ recovery of this mutant(Niyogi et al., 1998). The absence of the Stn7 kinase didnot result in substantial changes in any of the observedparameters. More importantly, the inability to undergostate transitions was not accompanied by absent orsmaller DF720 change.The values of the parameters DF682 and DF720 ob-
tained after 30 min of irradiation are summarized inTable I for all measured plant genotypes, together withthe technical Stern-Volmer-type parameters NPQ, qE,and qI (Krause and Jahns, 2003), detected at 682 nm.The most striking result is the very strong correlationbetween DF720 and the rapidly forming and relaxingquenching component qE, whose contribution is de-fined as the part of total quenching that relaxes after 10min of darkness. The studied mutants have a largely
different capacity for qE, ranging from 0 (in npq4) toabout double the wild-type value in L17. In all mu-tants, the light-induced DF720 remarkably followed theqE. In contrast, all mutants were able to produce qI (i.e.the part of quenching that did not relax in 5 min), andthe variations in this parameter and in DF682 wererather small. It is important to stress that while qE and(especially) the DF720 fluorescence change can be attrib-uted to a particular (PsbS-dependent) photochemical/physical mechanism of quenching, the qI parametercannot be associated with a single mechanism and ispurely a technical term.
The behavior of the F682/F750 ratio was stronglydependent on the measurement protocol, and partic-ularly the frequency of the saturating pulses. Whensaturating pulses were applied at 3-min intervals (Fig.
Figure 6. Time courses of the light-inducedformation and subsequent dark relaxation ofthe NPQ parameter [NPQ = F(t0)/F(t) – 1] cal-culated at three wavelengths in Arabidopsiswild-type (w.t.) plants and the mutants L17(PsbS overexpressor), npq4 (PsbS less), npq1(Zx deficient), npq2 (Zx accumulator), andstn7 (Stn7 less) irradiated at 600 mmol m22 s21
photon flux density red light for 30 min (whitebars). The fluorescence was recorded every60 s by applying a 0.2-s pulse (460 nm,photon flux density of 1,500 mmol m22 s21).
Kinetic and Spectral Resolution of NPQ Components
Plant Physiol. Vol. 152, 2010 1617 www.plant.org on April 26, 2016 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2010 American Society of Plant Biologists. All rights reserved.

8A), the changes in F682/F750 were significantly smalleras compared with the changes measured with a 1-minpulse interval. The slow phase and particularly theslow increase after switching the actinic light off werebasically abolished. The NPQ parameter and the mag-nitude of DF682 and DF720 are shown in Figure 8B for 1-,3-, and 6-min pulse intervals. The pulse repetition ratehas impact on the NPQ and on the DF682 change, butthe DF720 change is independent from it.
DISCUSSION
Spectral Changes Detected at 77 K
The normalized fluorescence emission spectra ofHL-treated leaves showed the well-established de-
crease of fluorescence in the PSII region comparedwith the PSI region shown by many authors (Krauseet al., 1983; Kyle et al., 1983; McTavish, 1988). Inaddition to this change, a characteristic light-inducedchange, not previously reported, was found in the FRregion: the increase of F730 relative to F760. There aretwo possible explanations to interpret this effect: (1)the negative and the positive relative difference bandshave the same common origin (i.e. quenching of PSIIfluorescence); and (2) there are different mechanismsor sites that are responsible for the different positive/negative light-induced spectral changes. The latterhypothesis brings two further possibilities: (1) therelative increase of F730 is a consequence of the quench-ing of fluorescence components with maxima outsidethe difference band; and (2) F730 is a genuine new
Figure 7. Time-dependent changes in the rel-ative fluorescence differences 2DF682 (whitecircles) and DF720 (black circles) during illu-mination (620 nm, photon flux density of 600mmol m22 s21) and subsequent redarkening ofleaves of Arabidopsis wild-type (w.t.) plantsand the mutants L17 (PsbS overexpressor),npq4 (PsbS less), npq1 (Zx deficient), npq2(Zx accumulator), and stn7 (Stn7 less). Fluo-rescence was excited by saturating pulsesapplied every 60 s. The parameters werecalculated as the light-minus-dark differencein the ratios F682/F750 and F720/F750 (DF720 wasmultiplied by 5 for clarity). The curves repre-sent the results from a single leaf measure-ment. SE values corresponding to six to ninemeasured leaves are shown as error bars.
Lambrev et al.
1618 Plant Physiol. Vol. 152, 2010 www.plant.org on April 26, 2016 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2010 American Society of Plant Biologists. All rights reserved.

fluorescence emission component resulting from theHL-induced formation of a new FR light-emittingexcited-state species. The 77 K fluorescence data alonedid not provide sufficient evidence to distinguishbetween these possibilities. However, the results can-not be explained within the usual concept that at lowtemperature the fluorescence in the FR region origi-nates from PSI alone. The observed increase of F730relative to F760, if regarded as a consequence of PSIIquenching, would only be possible if PSII fluorescencehad a broader vibrational tail extendingmore to the FRthan PSI (i.e. if the ratio F760/F730 of the pure PSIIspectrum were higher than that of the pure PSI spec-trum). A state 1-state 2 transition also cannot explainthe observed effect. Both the quenching of PSII and apotential state transition could explain the negativeF683 and F691 difference band but not the positive F730band demonstrated in the normalized spectra.
Different Mechanisms Are Responsible for theHL-Induced Spectral Changes
The hypothesis that the positive difference band inthe FR region discovered under NPQ conditions has adifferent mechanism of origin than the quenching ofthe main PSII fluorescence band was tested by a seriesof RT experiments in which the time course of light-induced formation and dark relaxation was followedand compared for each fluorescence difference band.The reasoning is that in case these difference bandshave a common origin and location in the PSII an-tenna, they should have the same kinetics and thesame spectral characteristics; conversely, differences inthe kinetics of formation or relaxation would be anindication that the underlying mechanisms or sites ofaction are different for these bands. Although at RT thespectral changes in the FR region are smaller than at 77K, the positive difference band at 720 nm can beresolved clearly and with high precision in our exper-iments, to reveal that it shows drastically differentkinetics of induction and recovery, as compared withthe quenching of PSII detected at 682 nm.The red (682-nm) and FR (720-nm) spectral changes
not only differ in their time course during irradiationand subsequent redarkening but also are affected in a
distinctly different manner bymutations. For example,the presence or absence of PsbS has a strong impact onthe FR change only but no statistically significant effecton the 682-nm changes. The 682-nm changes dependsignificantly on the applied frequency of saturatinglight pulses, yet the 720-nm changes are insensitive to
Table I. NPQ parameters and fluorescence ratios in Arabidopsis leaves
NPQ = Fdark/Flight – 1, qE = Fdark/Flight – Fdark/Fredark, qI = Fdark/Fredark – 1, DF682 = [(F682/F750)light – (F682/F750)dark] 3 100, DF720 = [(F720/F750)light – (F720/F750)dark] 3 100, where Fdark, Flight, and Fredark correspond tothe fluorescence of dark-adapted leaves, after 30 min of HL irradiation, and after 5 min of subsequentredarkening, respectively. Average values 6 SE are shown (n = 6–9).
Genotype NPQ682 qE682 qI682 2DF682 DF720
Wild type 2.9 6 0.1 2.1 6 0.4 0.9 6 0.2 30 6 2 4.4 6 0.2stn7 3.4 6 0.1 2.8 6 0.1 0.6 6 0.0 27 6 3 5.4 6 0.3npq4 1.4 6 0.1 0.0 6 0.0 1.3 6 0.1 29 6 4 0.3 6 0.2L17 6.8 6 0.5 5.6 6 0.5 1.3 6 0.2 35 6 3 10.4 6 0.5npq1 1.5 6 0.1 0.5 6 0.1 1.1 6 0.1 27 6 2 1.9 6 0.3npq2 2.3 6 0.1 1.6 6 0.2 0.8 6 0.2 31 6 5 6.7 6 0.4
Figure 8. A, Time-dependent changes in the relative fluorescencedifference 2DF682 during illumination and redarkening of leaves ofwild-type (w.t.), L17, and npq4 plants. Conditions are as in Figure 7,except that saturating pulses were applied at 3-min intervals. B, NPQparameter at 682 nm, 2DF682, and DF720 obtained after 30 min of HLirradiation of wild-type leaves with additional saturating pulses every 1,3, or 6 min. Error bars represent SE (n = 3–9).
Kinetic and Spectral Resolution of NPQ Components
Plant Physiol. Vol. 152, 2010 1619 www.plant.org on April 26, 2016 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2010 American Society of Plant Biologists. All rights reserved.

it. Therefore, we have to conclude that there are at leasttwo independent processes responsible for the fluo-rescence time changes. One only leads to a relativedecrease of the main PSII fluorescence band at 682 nm.The other quenches the PSII fluorescence as well butconcomitantly develops a specific new fluorescence inthe 720- to 730-nm range.
The FR Fluorescence Change Is Related to qE
We attempted to investigate the particular biologicalprocess underlying the newly found spectroscopicfeature of NPQ, DF720. Although the increase of FR(greater than 700 nm) relative to red fluorescence (680–690 nm) has been commonly interpreted as a result of astate transition (Butler and Kitajima, 1975; Mawsonand Cummins, 1986; McTavish, 1988; Walters andHorton, 1991), this cannot explain the DF720 changefor two reasons: first, the relative increase of the F720 toF750 tail of PSI fluorescence has no obvious explana-tion; and second, the amount of energy redistributionfrom PSII to PSI under HL conditions used in ourexperiments is generally believed to be very low, ifpresent at all (Walters and Horton, 1991). Withoutrelying on any such assumptions, however, we cancompletely exclude a state transition hypothesis, sincein the stn7 mutant of Arabidopsis, which is unable toundergo state transitions (Bellafiore et al., 2005), theDF720 band is unaffected. Therefore, the DF720 spectralchange is definitely not associated with a state 1-state 2transition.
The time course of the HL-induced DF720 spectralchange, which reaches a stationary state in a fewminutes of irradiation and reverses completely withina few minutes in darkness, clearly associates it withthe energy-dependent type of quenching (qE) anddistinguishes it from the so-called “sustained”quenching (qI). The magnitude of the relative differ-ence DF720 is strongly correlated with the value of theqE parameter in all examined mutants. The DF720 bandis thus a spectral marker for the mechanism underly-ing qE, and as such it opens new possibilities to probeqE specifically and independently from other quench-ing components (e.g. photoinhibition or others).
The most striking feature of the HL-induced FRfluorescence change is its complete absence in thePsbS-deficient mutant npq4 and its enhancement in thePsbS overexpressor. This demonstrates the direct roleof the PsbS protein and further confirms the associa-tion of PsbS with qE but not with qI. Since the DF682change was not inhibited by the lack of PsbS (on thecontrary, it seems to be slightly enhanced), it becomesclear that Arabidopsis plants have at least two differ-ent mechanisms for the rapidly inducible NPQ: aPsbS-dependent one and a PsbS-independent one.Furthermore, it follows that the function of PsbS canbe exerted independently from the xanthophyll cycle,since the rapid light-induced DF720 is detected also inthe npq1 and npq2 mutants. However, when Zx cannotbe formed, the amplitude of the spectral change is
significantly smaller, which is consistent with thelower ability of the npq1 mutant to generate both qEand DF720. It must be also noted that double mutantscontaining the npq4 mutation (e.g. the npq1npq4 andthe npq2npq4 mutants) also did not have any detect-able DF720 (data not shown), confirming the com-pulsory role of PsbS for qE and the relation of DF720with qE.
Could the observed FR fluorescence change have adifferent and more trivial origin? The measured fluo-rescence time changes could be in principle influencedby changes in leaf absorbance and reflectance. Butsuch changes would affect the whole fluorescenceintensity and would not primarily give rise to distinctspectral changes of the fluorescence. These effectswould thus be compensated by the normalizationprocedure. Changes in the effective absorption mightalso lead to different reabsorption and thus affect thespectral shape of the fluorescence emission. Clearly,leaves have a strong reabsorption of fluorescence inthe range below 695 nm. However, there is little or noabsorption in the FR region; thus, such artifacts can beexcluded. Another source of error could be light-scattering changes (e.g. associated with DpH-inducedthylakoid swelling). However, light scattering primar-ily affects the measurement of absorbance but has amuch smaller, and almost independent from the opti-cal arrangement, effect on fluorescence. Furthermore,such light-scattering changes could hardly give rise toa distinct spectral change of the fluorescence in arelatively narrow range. More importantly, though, itis extremely unlikely that effects such as DpH-inducedthylakoid swelling and others, that in general mightgive rise to such scattering changes, would follow thewell-known qE dependence on PsbS content, as doesthe observed DF720 fluorescence change. Therefore,while we cannot totally exclude some disturbancesdue to possible optical artifacts, it is extremely unlikelythat they could in fact explain the majority of theobserved DF720 spectral fluorescence change. Rather,the dependence of this change on the different PsbSmutants strongly suggests that it is indeed qE related.
Photophysical Origin of the FR Spectral Change
The actual photophysical origin of the HL-inducedchanges in the FR region of the fluorescence spectrumcannot be determined from our steady-state fluores-cence measurements alone. However, recent ultrafastfluorescence data on isolated LHCII aggregates andintact leaves of Arabidopsis acquired in our laboratory(Miloslavina et al., 2008; Holzwarth et al., 2009) indi-cate the likely origin of this NPQ-related emissioncomponent. Time-resolved fluorescence spectra of HL-treated leaves at RT revealed a new PsbS-dependentantenna fluorescence component not present in thedark-adapted leaves (Holzwarth et al., 2009). Remark-ably, the emission spectrum of this fluorescence decaycomponent showed a strong enhancement in the FRregion, in the range 700 to 730 nm, as compared with
Lambrev et al.
1620 Plant Physiol. Vol. 152, 2010 www.plant.org on April 26, 2016 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2010 American Society of Plant Biologists. All rights reserved.

normal PSII fluorescence. This component was absentin PsbS-deficient plants, and its amplitude was en-hanced in the PsbS overexpressor L17. It can be imag-ined that both the HL-induced FR decay componentand the DF720 change detected here in the steady-statespectra presented originate from the samemechanism.Interestingly, when LHCII forms aggregates, or
higher order oligomers, in vitro, it exhibits enhancedfluorescence in the FR region, particularly strong at lowtemperatures (Ruban and Horton, 1992; Mullineauxet al., 1993; Vasil’ev et al., 1997) but also at RT(Miloslavina et al., 2008). Because oligomerization ofLHCII leads to strong fluorescence quenching (Ideet al., 1987; Ruban and Horton, 1992; Mullineaux et al.,1993), it has actually been proposed as a mechanismfor NPQ in vivo (Horton et al., 1991, 2005; Ruban et al.,2007). An essential finding in this direction is that thespectrum and lifetime of the detached antenna com-ponent measured in intact quenched leaves underNPQ conditions closely match the FR fluorescence ofLHCII aggregates in vitro (Miloslavina et al., 2008).Furthermore, when these LHCII aggregates are cooledto 77 K, a new FR fluorescence located in the range700 to 730 nm appears (Ruban and Horton, 1992;Mullineaux et al., 1993; Miloslavina et al., 2008). Thus,the FR fluorescence at low temperature in LHCIIaggregates also parallels the DF720 band observedhere. We thus suggest that the qE-associated PsbS-dependent FR spectral change has the same origin asthe FR emission of quenched LHCII aggregates.
Flash-Induced Quenching Component
Whereas the DF720 band can be ascribed to a specificmechanism of quenching (i.e. the PsbS-dependent qE),this does not hold true for the DF682 band, whichreflects any processes that quench PSII fluorescenceand will depend on both the qE and qI components ofNPQ. The F682/F750 ratio can be then expected todecrease gradually with the slower formation of Zxand not to reverse immediately after turning off theactinic light because of the remaining qI. However, thiscannot explain two important observed effects: (1) theslow changes in the F682/F750 ratio in the npq1 and npq2mutants, in which the Zx content does not change; and(2) the rise of the ratio observed after switching off theactinic light. These changes are obviously induced bythe short saturating flashes only and accumulate witheach flash, provided that the time interval betweenflashes is short. The total irradiation dose from theflashes in our measurements is negligibly small com-pared with the background actinic light: only 0.4% ofthe total number of incident photons at a flash inter-val of 60 s. This rules out the possibility that the fluo-rescence changes are simply an additive effect of themeasuring and actinic light. Therefore, the flash-induced changes reflect a specific process and appearto be triggered by the high turnover rate of PSII duringthe saturating flash. A key observation is that the flash-induced DF682 depends on the flash repetition rate.
This suggests that these flash-induced changes arereversible on a time scale of a few minutes. At shorterintervals between flashes, the time is not sufficient forthese changes to relax and the effects are cumulative.This behavior shows that there is a reversible compo-nent of NPQ, independent from both PsbS and Zx, thatcan be induced by a short pulse of strong light andrelaxes on a time scale of several minutes. This thenrepresents an additional independent mechanism ofNPQ. It might be responsible for the quenching ob-served in mutants having neither PsbS nor xantho-phyll cycle activity, such as the npq1npq4 and npq2npq4double mutants. Based on our data, we cannot tell atpresent what the actual underlying quenching mech-anism might be or whether it takes place in theantenna or the RC of PSII. We hypothesize, however,that this quenching might be located in the RC of PSII,thus relating it to RC quenching that has been pro-posed in the literature (Finazzi et al., 2004; Ivanovet al., 2008).
It must be stressed that the flash-induced quenchingcomponent severely distorts the measured NPQ relax-ation kinetics when measuring flashes are spaced atshort intervals (i.e. shorter than 5 min) and, moreimportantly, that it contributes significantly to qI,interpreted usually as irreversible or slowly reversiblequenching, whereas in fact it is also a quenchingcomponent relaxing on a relatively short time scaleof about 5 to 10min. Thus, in our measurements, by far
Figure 9. A, Schematic representation of the dual-LED fluorescencesetup for registering fluorescence emission spectra of leaves during theformation and relaxation of NPQ. Solid lines represent electricalconnections, and dotted lines represent the optical path. Numbersare as follows: 1, regulated direct current power supply; 2, excitationLED; 3, actinic LED; 4, short-pass filter (630 nm); 5, lens; 6, leaf holder;7, long-pass filter (660 nm); 8, fiber optics; 9, casing; 10, USB2000spectrometer; 11, triggering signal; 12, computer interface. B, Timingdiagram of a typical measurement cycle showing the recording of thebackground fluorescence, turning on of the saturating pulse (SP), andrecording of the fluorescence spectrum.
Kinetic and Spectral Resolution of NPQ Components
Plant Physiol. Vol. 152, 2010 1621 www.plant.org on April 26, 2016 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2010 American Society of Plant Biologists. All rights reserved.

the largest parts of the slow induction and relaxationphases, generally called the qI phase in the literature,are genuine reversible NPQ quenching componentsand are not related to damage or photoinhibition. Infact, our experiments strongly suggest that true photo-inhibition is minor under the conditions applied hereeven for HL irradiation up to 1 h on intact leaves.
CONCLUSION
We have been able with the help of a newly de-signed multiwavelength fluorometer for registeringspectrally resolved fluorescence and NPQ inductionkinetics to dissect and distinguish three contributionsto NPQ. We also discovered spectral features specificto the different components of NPQ. We demonstratethree separate and independent mechanisms or sitesof action of NPQ. The first one is the rapidly inducibleand rapidly relaxing PsbS-dependent qE mechanism,which gives rise to a relative FR fluorescence increasearound 720 nm. This spectral signature of qE suggeststhat a new emitting species is produced together withthe qE quenching induction. The second, slower NPQquenching process occurs essentially independentlyfrom PsbS but can be correlated to the formation of Zx.It does not give rise to new FR-fluorescing species butonly decreases PSII fluorescence uniformly across thespectrum. Finally, a third quenching process, indepen-dent from both PsbS and Zx, was detected that appearsto be reversible in the dark on an approximately 5-mintime scale but is completely unrelated to the mecha-nism of qE and is triggered by a high excitation orturnover rate in PSII. All of these three NPQ mecha-nisms are reversible over the time scales and excitationintensities used in our experiments. Any nonrelaxingquenching contributions appear to be minor.
MATERIALS AND METHODS
Plant Material
Arabidopsis (Arabidopsis thaliana ecotype Columbia 0) wild-type and
mutant plants were grown in soil at a light intensity of 150 mmol photons
m22 s21 and a constant temperature of 20�C under long-day conditions (14 h of
light/10 h of dark). Leaves from 5- to 6-week-old plants were used for all
experiments. The following mutants were used: npq1, defective in the
violaxanthin deepoxidase (Niyogi et al., 1998); npq2 (also termed aba1-6),
defective in the Zx epoxidase (Niyogi et al., 1998); npq4, PsbS deficient (Li
et al., 2000); L17, PsbS overexpressing (Li et al., 2002); and stn7, defective in the
LHCII kinase (Bonardi et al., 2005).
77 K Fluorescence Measurements
In all experiments, plants were dark adapted for at least 1 to 2 h. Prior to
the measurements, leaves were detached, moistened, and placed between two
glass plates, where they were irradiated at RT for certain times using a high-
power red (620-nm) LED (Philips Lumileds Lighting) at an intensity of 600 to
1,000 mmol m22 s21. The leaves were then immediately frozen in liquid
nitrogen and placed in an optical cryostat (Oxford Instruments). Fluorescence
spectra were recorded in a homemade system based on a USB2000 CCD
spectrometer (Ocean Optics) using a blue (465-nm) LED as an excitation
source, filtered with a 550-nm long-pass glass filter.
Dual-LED Fluorescence Spectrometer for
RT Measurements
For measurements of NPQ generation and relaxation at RT, a special dual-
LED fluorescence setup was devised, schematically illustrated in Figure 9A.
The optical setup is assembled in a dark casing. Two high-power LEDs
(Luxeon Star-O; Philips Lumileds Lighting), one red (620 nm) and one blue
(460 nm), are focused with lenses onto the leaf holder. Short-pass 630-nm
filters (Thorlabs) are placed in front of the LEDs to remove long-wavelength
emission. The fluorescence is collected via another lens at an angle of 45�with
the leaf surface. Scattered light is filtered through a custom-made 660-nm
long-pass interference filter (LayrTech). The fluorescence is focused onto a
fiber-optics guide connected to a CCD spectrometer (USB2000; Ocean Optics).
The data-acquisition protocol is programmed inMicrosoft Visual Studio using
an interface library from Ocean Optics. Data analysis is done in MATLAB. In
our experiments, the red LED was used as continuous actinic light for the
induction of NPQ and the blue LED provided periodic short pulses used at the
same time to close the reaction centers and as excitation (measuring) source.
For this purpose, a fast-response, electronically switched direct current power
supply was built, ensuring pulse rise/decay time of less than 50 ms. The
triggering signal for the pulses is supplied by the spectrometer and program-
matically synchronized with the fluorescence detection. Fluorescence spectra
are recorded with each given saturating pulse as depicted in Figure 9B. Before
the pulse, a background spectrum is recorded. The LED is triggered and, after
a predefined delay, the actual pulse-excited fluorescence spectrum is re-
corded, the LED is then switched off, and the background signal is subtracted
by software. This way, the output signal represents only the fluorescence ex-
cited by the saturating pulses but not by the actinic light. After switching off
the actinic light, the relaxation of NPQ can be monitored, provided that a
sufficient time separation is allowed between the measuring pulses to prevent
quenching induction.
The actual measurement conditions used in the reported dual-light exper-
iments were as follows. Blue measuring/saturating pulses of 200 ms duration
and approximate intensity of 1,500 mmol photons m22 s21 were given every
60 s. Fluorescence detection was started 30 ms after the pulse onset. After the
first pulse (to acquire maximum PSII fluorescence in the dark-adapted state), the
red actinic light with intensity of 600 mmol photons m22 s21 was switched on.
The setup can also be used in a single-LED mode, where the same LED is
used as actinic and excitation source. This allows for following faster induc-
tion changes registering a full spectrum every few milliseconds. Thus, in
effect, the described setup provides most of the functionality of both conven-
tional PAM and direct detection fluorometers, although with full spectral
resolution. The time-dependent spectra shown in Figure 3 were acquired in
single-LED mode using red light (1,000 mmol photons m22 s21) and a 2-s
recording interval.
Sequence data from this article can be found in the GenBank/EMBL data
libraries under accession numbers AF134131, AF370251, and AF281655.
Received September 29, 2009; accepted December 16, 2009; published Decem-
ber 23, 2009.
LITERATURE CITED
Andrizhiyevskaya EG, Chojnicka A, Bautista JA, Diner BA, vanGrondelle R,
Dekker JP (2005) Origin of the F685 and F695 fluorescence in photosystem
II. Photosynth Res 84: 173–180
Aro EM, Virgin I, Andersson B (1993) Photoinhibition of photosystem II:
inactivation, protein damage and turnover. Biochim Biophys Acta 1143:
113–134
Avenson TJ, Ahn TK, Niyogi KK, Ballottari M, Bassi R, Fleming GR
(2009) Lutein can act as a switchable charge-transfer quencher in the
CP26 light-harvesting complex. J Biol Chem 284: 2830–2835
Bellafiore S, Barneche F, Peltier G, Rochaix JD (2005) State transitions and
light adaptation require chloroplast thylakoid protein kinase STN7.
Nature 433: 892–895
Bilger W, Bjorkman O (1990) Role of the xanthophyll cycle in photo-
protection elucidated by measurements of light-induced absorbance
changes, fluorescence and photosynthesis in leaves of Hedera canariensis.
Photosynth Res 25: 173–185
Bonardi V, Pesaresi P, Becker T, Schleiff E, Wagner R, Pfannschmidt T,
Lambrev et al.
1622 Plant Physiol. Vol. 152, 2010 www.plant.org on April 26, 2016 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2010 American Society of Plant Biologists. All rights reserved.

Jahns P, Leister D (2005) Photosystem II core phosphorylation and
photosynthetic acclimation require two different protein kinases. Na-
ture 437: 1179–1182
Briantais JM, Ducruet JM, Hodges M, Krause GH (1992) The effects
of low temperature acclimation and photoinhibitory treatments on
photosystem-2 studied by thermoluminescence and fluorescence decay
kinetics. Photosynth Res 31: 1–10
Briantais JM, Vernotte C, Picaud M, Krause GH (1979) A quantitative
study of the slow decline of chlorophyll a fluorescence in isolated
chloroplasts. Biochim Biophys Acta 548: 128–138
Butler WL, Kitajima M (1975) Energy transfer between photosystem II and
photosystem I in chloroplasts. Biochim Biophys Acta 396: 72–85
Crouchman S, Ruban AV, Horton P (2006) PsbS enhances nonphotochem-
ical fluorescence quenching in the absence of zeaxanthin. FEBS Lett 580:
2053–2058
Dall’Osto L, Fiore A, Cazzaniga S, Giuliano G, Bassi R (2007) Different
roles of a- and b-branch xanthophylls in photosystem assembly and
photoprotection. J Biol Chem 282: 35056–35068
Demmig B, Bjorkman O (1987) Comparison of the effect of excessive light
on chlorophyll fluorescence (77K) and photon yield of O2 evolution in
leaves of higher plants. Planta 171: 171–184
Demmig B, Winter K, Kruger A, Czygan FC (1987) Photoinhibition and
zeaxanthin formation in intact leaves: a possible role of the xanthophyll
cycle in the dissipation of excess light energy. Plant Physiol 84: 218–224
Demmig-Adams B (1990) Carotenoids and photoprotection in plants: a role
for the xanthophyll zeaxanthin. Biochim Biophys Acta 1020: 1–24
Demmig-Adams B, Adams WW (1992) Photoprotection and other re-
sponses of plants to high light stress. Annu Rev Plant Physiol Plant Mol
Biol 43: 599–626
Farber A, Young AJ, Ruban AV, Horton P, Jahns P (1997) Dynamics
of xanthophyll-cycle activity in different antenna subcomplexes in
the photosynthetic membranes of higher plants. Plant Physiol 115:
1609–1618
Finazzi G, Johnson GN, Dall’Osto L, Joliot P, Wollman FA, Bassi R (2004)
A zeaxanthin-independent nonphotochemical quenching mechanism
localized in the photosystem II core complex. Proc Natl Acad Sci USA
101: 12375–12380
Frank HA, Bautista JA, Josue JS, Young AJ (2000) Mechanism of non-
photochemical quenching in green plants: energies of the lowest ex-
cited singlet states of violaxanthin and zeaxanthin. Biochemistry 39:
2831–2837
Funk C, Schroder WP, Napiwotzki A, Tjus SE, Renger G, Andersson B
(1995) The PSII-S protein of higher plants: a new type of pigment-
binding protein. Biochemistry 34: 11133–11141
Genty B, Wonders J, Baker NR (1990) Non-photochemical quenching of Fo
in leaves is emission wavelength dependent: consequences for quench-
ing analysis and its interpretation. Photosynth Res 26: 133–139
Gilmore AM, Yamamoto HY (1992) Dark induction of zeaxanthin-
dependent nonphotochemical fluorescence quenching mediated by ATP.
Proc Natl Acad Sci USA 89: 1899–1903
Golan T, Li X-P, Muller-Moule P, Niyogi KK (2004) Using mutants to
understand light stress acclimation in plants. In GC Papageorgiou,
Govindjee, eds, Chlorophyll a Fluorescence: A Signature of Photosyn-
thesis. Springer, Dordrecht, The Netherlands, pp 525–554
Govindjee (1995) Sixty-three years since Kautsky: chlorophyll a fluores-
cence. Aust J Plant Physiol 22: 131–160
Haehnel W, Nairn JA, Reisberg P, Sauer K (1982) Picosecond fluorescence
kinetics and energy transfer in chloroplasts and algae. Biochim Biophys
Acta 680: 161–173
Holt NE, Zigmantas D, Valkunas L, Li XP, Niyogi KK, Fleming GR (2005)
Carotenoid cation formation and the regulation of photosynthetic light
harvesting. Science 307: 433–436
Holzwarth AR (1986) Excited state kinetics of chlorophyll antenna pig-
ments. In LA Staehelin, CJ Arntzen, eds, Encyclopedia of Plant Phys-
iology: Photosynthesis III. New Series, Vol 19. Springer, Berlin, pp
299–309
Holzwarth AR (1991) Excited-state kinetics in chlorophyll systems and its
relationship to the functional organization of the photosystems. In H
Scheer, ed, Chlorophylls. CRC Press, Boca Raton, FL, pp 1125–1151
Holzwarth AR, Miloslavina Y, Nilkens M, Jahns P (2009) Identification of
two quenching sites active in the regulation of photosynthetic light-
harvesting studied by time-resolved fluorescence. Chem Phys Lett 483:
262–267
Holzwarth AR, Roelofs TA (1992) Recent advances in the understanding of
chlorophyll excited state dynamics in thylakoid membranes and iso-
lated reaction centre complexes. J Photochem Photobiol B 15: 45–62
Holzwarth AR, Wendler J, Haehnel W (1985) Time-resolved picosecond
fluorescence spectra of the antenna chlorophylls in Chlorella vulgaris:
resolution of photosystem I fluorescence. Biochim Biophys Acta 807:
155–167
Horton P, Hague A (1988) Studies on the induction of chlorophyll
fluorescence in isolated barley protoplasts. IV. Resolution of non-
photochemical quenching. Biochim Biophys Acta 932: 107–115
Horton P, Ruban AV (1999) Regulation of the structure and function of the
light harvesting complexes of photosystem II by the xanthophyll cycle.
In HA Frank, AJ Young, G Britton, RJ Cogdell, eds, The Photochemistry
of Carotenoids. Kluwer Academic Publishers, Dordrecht, The Nether-
lands, pp 271–291
Horton P, Ruban AV (2005) Molecular design of the photosystem II light-
harvesting antenna: photosynthesis and photoprotection. J Exp Bot 56:
365–373
Horton P, Ruban AV, Rees D, Pascal AA, Noctor G, Young AJ (1991)
Control of the light-harvesting function of chloroplast membranes by
aggregation of the LHCII chlorophyll-protein complex. FEBS Lett
292: 1–4
Horton P, Ruban AV, Walters RG (1994) Regulation of light harvesting in
green plants: indication by nonphotochemical quenching of chlorophyll
fluorescence. Plant Physiol 106: 415–420
Horton P, Ruban AV, Walters RG (1996) Regulation of light harvesting in
green plants. Annu Rev Plant Physiol Plant Mol Biol 47: 655–684
Horton P, Wentworth M, Ruban AV (2005) Control of the light harvesting
function of chloroplast membranes: the LHCII-aggregation model for
non-photochemical quenching. FEBS Lett 579: 4201–4206
Huner NPA, Ivanov AG, Sane PV, Pocock T, Krol M, Balseris A, Rosso D,
Savitch LV, Hurry VM, Oquist G (2005) Photoreaction of photosystem
II: reaction center quenching versus antenna quenching. In B Demmig-
Adams, WW Adams, A Mattoo, eds, Photoprotection, Photoinhibition,
Gene Regulation and Environment. Springer, Dordrecht, The Nether-
lands, pp 155–174
Ide JP, Klug DR, Kuhlbrandt W, Giorgi LB, Porter G (1987) The state of
detergent solubilised light-harvesting chlorophyll- a/b protein complex
as monitored by picosecond time-resolved fluorescence and circular
dichroism. Biochim Biophys Acta 893: 349–364
Ivanov AG, Sane PV, Hurry V, Oquist G, Huner NPA (2008) Photosystem
II reaction centre quenching: mechanisms and physiological role. Photo-
synth Res 98: 565–574
Johnson MP, Perez-Bueno ML, Zia A, Horton P, Ruban AV (2009) The
zeaxanthin-independent and zeaxanthin-dependent qE components of
nonphotochemical quenching involve common conformational changes
within the photosystem II antenna in Arabidopsis. Plant Physiol 149:
1061–1075
Kalituho L, Graßes T, Graf M, Rech J, Jahns P (2006) Characterization of a
nonphotochemical quenching-deficient Arabidopsis mutant possessing
an intact PsbS protein, xanthophyll cycle and lumen acidification. Planta
223: 532–541
Kalituho L, Rech J, Jahns P (2007) The roles of specific xanthophylls in
light utilization. Planta 225: 423–439
Karukstis KK, Sauer K (1983) Fluorescence decay kinetics of chlorophyll in
photosynthetic membranes. J Cell Biochem 23: 131–158
Komura M, Shibata Y, Itoh S (2007) A new fluorescence band F689 in
photosystem II revealed by picosecond analysis at 4–77 K: function of
two terminal energy sinks F689 and F695 in PS II. Biochim Biophys Acta
1757: 1657–1668
Krause GH (1988) Photoinhibiton of photosynthesis: an evaluation of
damaging and protective mechanisms. Physiol Plant 74: 566–574
Krause GH, Briantais JM, Vernotte C (1983) Characterization of chloro-
phyll fluorescence quenching in chloroplasts by fluorescence spectros-
copy at 77 K. I. DpH-dependent quenching. Biochim Biophys Acta 723:
169–175
Krause GH, Jahns P (2003) Pulse amplitude modulated fluorometry and its
application in plant science. In BR Green, WW Parson, eds, Light-
Harvesting Antennas in Photosynthesis. Kluwer Academic Publishers,
Dordrecht, The Netherlands, pp 373–399
Krause GH, Jahns P (2004) Non-photochemical energy dissipation deter-
mined by chlorophyll fluorescence quenching: characterization and
function. In GC Papageorgiou, Govindjee, eds, Chlorophyll a Fluores-
Kinetic and Spectral Resolution of NPQ Components
Plant Physiol. Vol. 152, 2010 1623 www.plant.org on April 26, 2016 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2010 American Society of Plant Biologists. All rights reserved.

cence: A Signature of Photosynthesis. Springer, Dordrecht, The Nether-
lands, pp 463–495
Krause GH, Weis E (1991) Chlorophyll fluorescence and photosynthesis:
the basics. Annu Rev Plant Physiol 42: 313–349
Kyle DJ (1987) The biochemical basis for photoinhibition of photosystem II.
In DJ Kyle, CB Osmond, CJ Arntzen, eds, Photoinhibition. Elsevier,
Amsterdam, pp 197–226
Kyle DJ, Arntzen CJ, Franck F, Inoue Y (1983) Light-induced quenching of
photosystem II fluorescence at 77K. Photochem Photobiol 38: 609–614
Li XP, Bjorkman O, Shih C, Grossman AR, Rosenquist M, Jansson S,
Niyogi KK (2000) A pigment-binding protein essential for regulation of
photosynthetic light harvesting. Nature 403: 391–395
Li XP, Gilmore AM, Caffarri S, Bassi R, Golan T, Kramer D, Niyogi KK
(2004) Regulation of photosynthetic light harvesting involves intra-
thylakoid lumen pH sensing by the PsbS protein. J Biol Chem 279:
22866–22874
Li XP, Muller-Moule P, Gilmore AM, Niyogi KK (2002) PsbS-dependent
enhancement of feedback de-excitation protects photosystem II from
photoinhibition. Proc Natl Acad Sci USA 99: 15222–15227
Li Z, Ahn TK, Avenson TJ, Ballotari M, Cruz JA, Kramer DM, Bassi R,
Fleming GR, Keasling JD, Niyogi KK (2009) Lutein accumulation in the
absence of zeaxanthin restores nonphotochemical quenching in the
Arabidopsis thaliana npq1 mutant. Plant Cell 21: 1798–1812
Long SP, Humphries S, Falkowski PG (1994) Photoinhibition of photo-
synthesis in nature. Plant Physiol Plant Mol Biol 45: 633–662
Mawson BT, Cummins WR (1986) The kinetics of in vivo state transitions
in mesophyll and guard cell chloroplasts monitored by 77 K fluores-
cence emission spectra. Plant Physiol 82: 873–879
Maxwell K, Johnson GN (2000) Chlorophyll fluorescence: a practical
guide. J Exp Bot 51: 659–668
McTavish H (1988) A demonstration of photosynthetic state transitions in
nature. Photosynth Res 17: 247–254
Miloslavina Y, Wehner A, Wientjes E, Reus M, Lambrev P, Garab G, Croce
R, Holzwarth AR (2008) Far-red fluorescence: a direct spectroscopic
marker for LHCII oligomers forming in non photochemical quenching.
FEBS Lett 582: 3625–3631
Muller P, Li X-P, Niyogi KK (2001) Non-photochemical quenching: a
response to excess light energy. Plant Physiol 125: 1558–1566
Mullineaux CW, Pascal AA, Horton P, Holzwarth AR (1993) Excitation
energy quenching in aggregates of the LHC II chlorophyll-protein
complex: a time-resolved fluorescence study. Biochim Biophys Acta
1141: 23–28
Murata N, Takahashi S, Nishiyama Y, Allakhverdiev SI (2007) Photo-
inhibition of photosystem II under environmental stress. Biochim
Biophys Acta 1767: 414–421
Niyogi KK (1999) Photoprotection revisited: genetic and molecular ap-
proaches. Annu Rev Plant Physiol Plant Mol Biol 50: 333–359
Niyogi KK (2000) Safety valves for photosynthesis. Curr Opin Plant Biol 3:
455–460
Niyogi KK, Grossman AR, Bjorkman O (1998) Arabidopsis mutants define
a central role for the xanthophyll cycle in the regulation of photosyn-
thetic energy conversion. Plant Cell 10: 1121–1134
Niyogi KK, Li XP, Rosenberg V, Jung HS (2005) Is PsbS the site of non-
photochemical quenching in photosynthesis? J Exp Bot 56: 375–382
Niyogi KK, Shih C, Soon Chow W, Pogson BJ, DellaPenna D, Bjorkman
O (2001) Photoprotection in a zeaxanthin- and lutein-deficient double
mutant of Arabidopsis. Photosynth Res 67: 139–145
Peterson R, Oja V, Laisk A (2001) Chlorophyll fluorescence at 680 and
730 nm and leaf photosynthesis. Photosynth Res 70: 185–196
Pfundel EE (1998) Estimating the contribution of photosystem I to total leaf
chlorophyll fluorescence. Photosynth Res 56: 185–195
Powles SB, Bjorkman O (1982) Photoinhibition of photosynthesis: effect on
chlorophyll fluorescence at 77K in intact leaves and in chloroplast
membranes of Nerium oleander. Planta 156: 97–107
Quick WP, Stitt M (1989) An examination of factors contributing to non-
photochemical quenching of chlorophyll fluorescence in barley leaves.
Biochim Biophys Acta 977: 287–296
Rijgersberg CP, Amesz J (1978) Changes in light absorbance and chloro-
phyll fluorescence in spinach chloroplasts between 5 and 80K. Biochim
Biophys Acta 502: 152–160
Roelofs TA, Kwa SLS, van Grondelle R, Dekker JP, Holzwarth AR (1993)
Primary processes and structure of the photosystem II reaction center. II.
Low-temperature picosecond fluorescence kinetics of a D1-D2-cyt-b-559
reaction center complex isolated by short Triton exposure. Biochim
Biophys Acta 1143: 147–157
Ruban AV, Berera R, Ilioaia C, van Stokkum IHM, Kennis JTM, Pascal
AA, van Amerongen H, Robert B, Horton P, van Grondelle R (2007)
Identification of a mechanism of photoprotective energy dissipation in
higher plants. Nature 450: 575–578
Ruban AV, Horton P (1992) Mechanism of DpH-dependent dissipation of
absorbed excitation energy by photosynthetic membranes. 1. Spectro-
scopic analysis of isolated light-harvesting complexes. Biochim Biophys
Acta 1102: 30–38
Ruban AV, Horton P (1994) Spectroscopy of non-photochemical and
photochemical quenching of chlorophyll fluorescence in leaves: evi-
dence for a role of the light harvesting complex of photosystem II in the
regulation of energy dissipation. Photosynth Res 40: 181–190
Satoh K, Butler WL (1978) Low temperature spectral properties of sub-
chloroplast fractions purified from spinach. Plant Physiol 61: 373–379
Schansker G, Toth SZ, Strasser RJ (2006) Dark recovery of the Chl a
fluorescence transient (OJIP) after light adaptation: the qT-component of
non-photochemical quenching is related to an activated photosystem I
acceptor side. Biochim Biophys Acta 1757: 787–797
Schansker G, van Rensen JJS (1999) Performance of active photosystem II
centers in photoinhibited pea leaves. Photosynth Res 62: 175–184
Schreiber U (2004) Pulse-amplitude-modulation (PAM) fluorometry and
saturation pulse method: an overview. In GC Papageorgiou, Govindjee,
eds, Chlorophyll a Fluorescence: A Signature of Photosynthesis.
Springer, Dordrecht, The Netherlands, pp 279–319
Schreiber U, Schliwa U, Bilger W (1986) Continuous recording of photo-
chemical and non-photochemical chlorophyll fluorescence quenching
with a new type of modulation fluorometer. Photosynth Res 10: 51–62
Slavov C, Ballottari M, Morosinotto T, Bassi R, Holzwarth AR (2008)
Trap-limited charge separation kinetics in higher plant photosystem I
complexes. Biophys J 94: 3601–3612
Somersalo S, Krause GH (1988) Changes in chlorophyll fluorescence
related to photoinhibition of photosynthesis and cold acclimation of
green plants. In HK Lichtenthaler, ed, Applications of Chlorophyll
Fluorescence. Kluwer Academic Publishers, Dordrecht, The Nether-
lands, pp 157–164
van Dorssen RJ, Breton J, Plijter JJ, Satoh K, van Gorkom HJ, Amesz J
(1987) Spectroscopic properties of the reaction center and of the 47kda
chlorophyll protein of photosystem-II. Biochim Biophys Acta 893:
267–274
Vasil’ev S, Schrotter T, Bergmann A, Irrgang K-D, Eichler H-J, Renger G
(1997) Cryoprotectant-induced quenching of chlorophyll a fluorescence
from light-harvesting complex 2 in vitro: time-resolved fluorescence
and steady state spectroscopic studies. Photosynth Res 33: 553–561
Walters RG, Horton P (1991) Resolution of components of non-
photochemical chlorophyll fluorescence quenching in barley leaves.
Photosynth Res 27: 121–133
Walters RG, Horton P (1993) Theoretical assessment of alternative mech-
anisms for non-photochemical quenching of PS-II fluorescence in barley
leaves. Photosynth Res 36: 119–139
Weis E (1985) Chlorophyll fluorescence at 77 K in intact leaves: character-
ization of a technique to eliminate artifacts related to self-absorption.
Photosynth Res 6: 73–86
Weis E, Berry JA (1987) Quantum efficiency of photosystem II in relation to
energy dependent quenching of chlorophyll fluorescence. Biochim
Biophys Acta 894: 198–208
Wraight CA, Crofts AR (1970) Energy-dependent quenching of chlorophyll
a fluorescence in isolated chloroplasts. Eur J Biochem 17: 319–327
Lambrev et al.
1624 Plant Physiol. Vol. 152, 2010 www.plant.org on April 26, 2016 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2010 American Society of Plant Biologists. All rights reserved.




















