
Journal of Food Bioactives
96
Volume 15 September 2021 Journal of Food Bioactives An Official Scientific Publication of the International Society for Nutraceuticals and Functional Foods (ISNFF) Editor-in-Chief Fereidoon Shahidi Co-Editor-in-Chief (Honorary) Beiwei Zhu ISNFF & ASSOCIATES Publishing Company, Inc. An Official Journal of the International Union of Food Science and Technology (IUFoST)
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Journal of Food Bioactives

Volume 15 September 2021
Journal of
Food BioactivesAn Official Scientific Publication of the
International Society for Nutraceuticals and Functional Foods (ISNFF)
Editor-in-ChiefFereidoon Shahidi
Co-Editor-in-Chief (Honorary)Beiwei Zhu
ISNFF & ASSOCIATES Publishing Company, Inc.
An Official Journal of the International Union of Food Scienceand Technology (IUFoST)

Journal of Food BioactivesBring together the results of fundamental and applied research on food bioactives, functional food ingredients, nutraceuticals and natu-ral health products that are known to possess or perceived to have health-promoting properties
www.isnff-jfb.com
Editorial Board
Editor-in-Chief
Fereidoon Shahidi (Canada)
Co-editor-in-Chief (Honorary)
Beiwei Zhu (China)
Editorial Board Members
Cesarettin Alasalvar (Turkey)Rotimi Aluko (Canada)Emilio Alvarez (Mexico)Ryszard Amarowicz (Poland)Anna Arnoldi (Italy)Joseph Banoub (Canada)
Colin Barrow (Australia)Adriano Costa de Camargo (Chile)Rong Cao (Tsao) (Canada)Richard FitzGerald (Ireland)Bruce Hamaker (USA)Chi-Tang Ho (USA)Farah Hosseinian (Canada)Charles C. Hu (USA)Amin Ismail (Malaysia)You-Jin Jeon (South Korea)Bo Jiang (China)Hitomi Kumagai (Japan)Shiming Li (USA)Jack Losso (USA)Kazuo Miyashita (Japan)Ganiyu Oboh (Nigeria)
Min-Hsiung Pan (Taiwan)Ronald B. Pegg (USA)Kenji Sato (Japan) Angela Shaw (USA)Young-Joon Surh (South Korea)Francisco Tomas-Barberan (Spain)Petras R. Venskutonis (Lithuania)Chin-Kun Wang (Taiwan)Dao-Ying Wang (China)Mingfu Wang (Hong Kong)Yu Wang (USA)Hanny C. Wijaya (Indonesia)Hang Xiao (USA)Wallace Yokoyama (USA)Hui Zhao (China)Da-Yong Zhou (China)
Instructions for Authors
Journal of Food Bioactives with open access option is an official scientific publication of the International Society for Functional Foods and Nutraceuticals (ISNFF), a not-for-profit Disciplinary Interest Group of the International Union of Food Science and Technology (IUFoST) that was founded in 2007. It publishes original research articles, short communications/research notes, opinion pieces and review articles.
ISSN 2637-8752 Print ISSN 2637-8779 Online
ISNFF & ASSOCIATES Publishing Company, Inc.Connecticut, USA

Copyright: © 2021 International Society for Nutraceuticals and Functional Foods.All rights reserved.
1
Meeting Report J. Food Bioact. 2021;15:1–2
Journal ofFood Bioactives International Society for
Nutraceuticals and Functional Foods
Consumer behaviour and a healthy diet: a challenge or an opportunity?
Fereidoon Shahidi
Functional Foods and Nutraceuticals Laboratory, Department of Biochemistry, Memorial University of Newfoundland, St. John’s, NL, Canada A1B 3X9, Canada. E-mail: [email protected]: 10.31665/JFB.2021.15276Received: September 28, 2021; Revised received & accepted: September 28, 2021Citation: Shahidi, F. (2021). Consumer behaviour and healthy diet: a challenge or an opportunity? J. Food Bioact. 15: 1–2.
Abstract
Healthy food choices by consumers are dictated by several factors but most would like healthy and affordable supply. Of course, traditional foods that are familiar to the consumers and have the appropriate sensory char-acteristics are most desirable to enhance immunity. Cultural background of the consumers often dictates their food habits and availability of local foods, driven by economic factors, are important considerations. Food safety, sustainability and traceability along with regulatory issues are to be considered in order to use the opportunities to address various challenges.
Keywords: Consumer behaviour; Healthy food; Sustainability; Traceability; Functional foods; Food safety; toxins; Health claims; Bioac-tives.
The International Academy of Food Science and Technology (IAFoST) and International Union of Food Science and Technolo-gy (IUFoST) hosted a round-table to discuss the consumer percep-tion, behaviour, food habits, cultural background, gender, socio-economic, food choices and dietary patterns and their influence on a healthy diet. The round Table was co-chaired by Dr. Aman Wirakartakusumah and Dr. Ugugua Charles Aworh. The rappor-teurs were Dr. Chin-Kun Wang and Dr. Jairo Romero.
There were 5 panel members who spoke on different aspects of the importance of consumer behaviour on developments related to a healthy diet. Lifestyle and balanced diet, sustainability, nutri-tion, and functional foods were considered as important aspects. In addition, safety and consumer demands were considered as being components that should help in providing evidence-based and transparent information. The importance of education, cultural behaviour and food literacy in healthy food choices should also be considered. Attention to food loss and waste, local access and availability, role of food science and technology, incentives on healthy diet, and finally value addition and circular economy in food industry were considered important.
The first panel member was Professor Sir Charles Godfray of Oxford University. He discussed the environmental and health is-sues related to livestock and the fact that the largest green house emission, i.e., methane, is due to ruminants and their feed. He
noted that for environmental issues, animal welfare, health, and consumers are factors needed for consideration as well as produc-tion cost and legal aspects.
The second panel member was Dr. Pavinee Chinachoti, chair of Food Innovation and Regulation Network from Thailand. She highlighted gaps in understanding safety and health claim regula-tions and that the food reaching the consumer must be safe and healthy. Product innovators must pay attention to the value chain. In this regard, to innovate, regulation should be recognized along with scientific developments, whilst health claims and functional food innovation roadmap must consider the fact that consumers are interested in transparency, traceability and more advanced food safety considerations that require appropriate technology and regu-lation in the supply chain.
The third speaker was Professor Sebastiano Porretta of the Ministry of Economic Development from Italy. He discussed consumer based new healthy food development. In this, he em-phasized that consumers are demanding clean label products with very few lists of ingredients and no use of unfamiliar chemical names, especially of synthetic compounds (added by the author). Thus, attention needs to be paid to the paradigm shift. Consum-ers are interested in reasons for performance of products. In this regard, functional foods and ingredients from industrial wastes and process stream might lead to consumer preference because these

Journal of Food Bioactives | www.isnff-jfb.com2
Consumer behaviour and a healthy diet Shahidi
co-products often have much higher concentration of bioactive compounds (added by the author). He noted that conjoint analysis provides a best approach. He also discussed segmentation and use of orthogonal functional design. In this regard, he noted the impor-tance of natural, generated or induced compounds, price, sustain-ability, consumer preference, claims and label issues. Although it might be said that the consumer is ready to pay extra for healthy functional foods, in practice they often look for cheap and healthy food options.
The next speaker was Dr. Caroline Smith DeWaal, Global Al-liance for Improved Nutrition. She talked about food safety and consumer demand and new tools for achieving development. She noted that 600 million food borne illnesses occur each year, thus ensuring access to safe and nutritious food is important. These re-sult in 420,000 deaths each year and 125,000 of them are children of less than 5 years. These are due to microbial, helminths, afla-toxin, and other toxins. While 41% of world population is in Asia and Africa, 75% of such deaths is there. In addition, while 9% the population there is from children under 5, 30% of deaths is in this age group. While access to safe and nutritious food is necessary, consumers are a major driving force. Establishing a functional knowledge platform to share information with all is essential so that progress could be made without duplication.
The last speaker was Dr. Petra Klassen Wigger, Global R & D Scientific Advisor for Nutrition and Health of Nestle. She noted that leveraging science and technology to address challenges of sustainable healthy diet was desired as consumers demand food and beverages that are safe and healthful from a nutritional and ingredient standpoint and are respectful of the environment. They also want food and nutrition security for the present and for future generations. In this regard, climate change is putting pressure on food systems and the need for affordable food with consideration of sustainability warrants timely attention. Consumers are also able to recognize healthy foods and distinguish them from less de-sirable choices. However, some problems persist. These are related
to the lack of understanding of how to achieve a balanced diet and nutritional risk when following a special diet, such as vegan food. Lack of knowledge to manage portion and frequency of consump-tion as well as awareness of micronutrients and benefits of fortifi-cation remain to be a challenge for most. Finally, misinterpretation of processing and its effects on food wholesomeness is an issue, especially for the ready-to-eat foods. In addition, understanding the local needs and attention to carbon footprint might be a consid-eration (from the author).
With regard to formulation and re-formulation, positive aspects related to micronutrients, protein, fiber, whole grain, nuts, fruits, vegetables and healthy oils as well as negative features related to sugar, sodium and salt content as well as trans fats (from the author) and energy content must all be considered when address-ing dietary needs. To achieve these goals, a multi-disciplinary ap-proach is needed to ensure science-based innovations. It is also essential to pay attention to the risk of exposure to plant toxins, mycotoxins, co-harvest issues, climate change, risk of adultera-tion, allergens and cross reactivity and cross contamination.
Overall, one might conclude that affordable nutrition, attention to raw material and loss upon harvest, storage and transportation or processing waste, use of innovative technologies, including fermen-tation, as well and use of adequately processed food are essential in addressing consumer’s needs. To achieve desirable sensory charac-ter fermentation and generation of a sour taste, for example, may help in better appreciation of formulations that might otherwise be unfamiliar to the consumers. While there are many challenges, there are also opportunities that we must take advantage of. In this regard, climate change and growing world population require urgent actions to ensure sustainability, healthfulness, and affordable diet. Science based innovation is a key driver and may constitute a shift to a plant-based diet, avoiding food loss and valorize side streams. Therefore, a holistic approach to ensure various aspects of consumer’s diet for better health and wellbeing of ecosystem that supports healthier so-ciety, and the planet, must be considered.

Copyright: © 2021 International Society for Nutraceuticals and Functional Foods.All rights reserved.
3
Perspective J. Food Bioact. 2021;15:3–12
Journal ofFood Bioactives International Society for
Nutraceuticals and Functional Foods
Is the gluten-free and casein-free diet efficacious in the treatment of childhood autism spectrum disorder?
Klaus W. Lange* and Andreas Reissmann
Department of Experimental Psychology, University of Regensburg, Regensburg, Germany*Corresponding author: Klaus W. Lange, Institute of Psychology, University of Regensburg, 93040 Regensburg, Germany. Tel: +49 941 9433815; Fax: +49 941 9434496; E-mail: [email protected]: 10.31665/JFB.2021.15277Received: September 16, 2021; Revised received & accepted: September 28, 2021Citation: Lange, K.W., and Reissmann, A. (2021). Is the gluten-free and casein-free diet efficacious in the treatment of childhood autism spectrum disorder? J. Food Bioact. 15: 3–12.
Abstract
Autism spectrum disorder (ASD) is a set of heterogeneous neurodevelopmental conditions, characterized by ear-ly-onset difficulties in social communication as well as repetitive and unusually restricted behaviors and interests. The treatment of ASD is based primarily on psychoeducational and behavioral interventions. Since the effective-ness of available treatments for ASD is limited, many families search for alternative therapies, such as the gluten-free and casein-free (GFCF) diet. Despite the popularity of the GFCF diet as a supplementary treatment in chil-dren with ASD, several rigorous evaluations have failed to confirm its effectiveness. The majority of the available studies examining the efficacy of the GFCF diet are seriously flawed and allow no firm conclusions. The available evidence regarding the effectiveness of the GFCF diet in the treatment of childhood ASD is very weak and cannot be considered promising. The GFCF diet should be used only if an allergy or intolerance to nutritional gluten or casein has been established. The identification of a hypothetical diet-related ASD phenotype may help in select-ing children who could benefit from a GFCF dietary intervention. An important consideration is that potentially ineffective therapies may imply considerable opportunity costs, with other possibly more effective treatment approaches remaining unutilized.
Keywords: Autism spectrum disorder; Children; Gluten-free and casein-free diet; Opioid excess hypothesis; Treatment.
1. Introduction
Autism spectrum disorder (ASD) is a set of heterogeneous de-velopmental conditions; the core features of ASD are early-onset social communication difficulties and repetitive, stereotypical and restricted sensory-motor behaviors (Kanner, 1943; Levy et al., 2009). While ASD was previously considered a rare and narrowly defined condition of childhood, it is viewed today as a lifelong condition with a spectrum ranging from very mild to severe. The disorder was grouped with neurodevelopmental disorders in the 5th edition of the Diagnostic and Statistical Manual of Mental Disorders (American Psychiatric Association, 2013). Previously distinguished subtypes, such as Asperger’s disorder and pervasive developmental disorder not otherwise specified, are now consoli-
dated under the diagnosis of ASD. Comorbid conditions, such as attention-deficit hyperactivity disorder and anxiety disorders, are common and have been reported in more than 70% of individuals diagnosed with ASD (Gotham et al., 2015; Simonoff et al., 2008). The prevalence of ASD has been estimated to be approximate-ly 1% globally (Elsabbagh et al., 2012) and 1.5% in developed countries (Lyall et al., 2017). Prevalence estimates have risen sig-nificantly over the last two decades. While this rise in prevalence might reflect a concomitant rise in the incidence of ASD, changes in the concepts and diagnostic criteria of ASD have been suggested as alternative explanations (Fombonne, 2009). A wide variety of risk factors for ASD, such as prenatal/perinatal and maternal life-style and dietary factors, have been suggested (Lyall et al., 2014; Mandy and Lai, 2017). Genetic studies have identified risk pat-terns (Gaugler et al., 2014; Tick et al., 2016), and gene-environ-

Journal of Food Bioactives | www.isnff-jfb.com4
Gluten-free and casein-free diet in autism Lange et al.
ment interactions with risk-inducing environmental compounds have been discussed (Herbert, 2010). Neurobiological examina-tions have found early changes of brain development as well as of neuronal reorganization and connectivity in ASD (Bauman and Kemper, 2005; Ecker et al., 2015; Hazlett et al., 2017; Lewis et al, 2014; O’Reilly et al., 2017). However, genetic and neurobiological studies have as yet been unable to provide any clinical benefit or identify reliable biomarkers for routine use (Walsh et al., 2011). There are therefore no biological diagnostic tools available, and the diagnosis of ASD must rely solely on behavioral assessment (American Psychiatric Association, 2013).
There is no curative therapy for ASD. The treatment is pri-marily based on psychoeducational and behavioral interventions (Reichow et al., 2014), with medication used as an adjunct. While some studies using low-intensity interventions involving parent-child interaction have shown significant effects on children’s so-cial behavior and communication (Weitlauf et al., 2014), others have been unable to find any positive effects (Carter et al., 2011a). Meta-analyses of treatment studies using early comprehensive and targeted behavioral interventions (naturalistic developmental be-havioral interventions) (Schreibman et al., 2015) have shown some positive effects on adaptive skills and language skills (Reichow et al., 2012; Weitlauf et al., 2014). However, only one trial was truly randomized, and no effects were found when these approaches were compared to other developmental interventions of equal in-tensity (Weitlauf et al., 2014). While drugs do not directly improve social communication in children with ASD, they may be able to reduce comorbid symptoms. Evidence-based pharmacotherapy of children and adolescents with ASD, using atypical antipsychotics (risperidone, aripripazole), is limited to the treatment of co-occur-ring behaviors, such as agitation, irritability, aggression and other disruptive behaviors (Fung et al., 2016). It is important to note that the reactions of a child’s family to the diagnosis of ASD affects the outcome as much as any treatment (Dykens et al., 2014).
Since the effectiveness of available treatments for ASD is lim-ited, many families search for alternative therapies (Owen-Smith et al., 2015; Perrin et al., 2012). A potential role of nutrition in the etiology of ASD has been suggested. It has been reported that up to a third of parents of children with ASD conceal information on nutritional interventions from the physician responsible for treat-ing their children (Trudeau et al., 2019). Moreover, up to a fifth of preschool children with ASD have been given some form of restriction diet (Rubenstein et al., 2018). In particular, the nutri-tional proteins gluten (from wheat, barley, rye and oats) and casein (from milk and other dairy products) have been hypothesized to be involved in ASD, and special diets free of gluten and casein have been suggested to be of value in the treatment of ASD (Lange et al., 2015). The present short review critically discusses the evi-dence regarding the therapeutic efficacy of the gluten-free and ca-sein-free (GFCF) diet in ASD.
2. Rationale for the GFCF diet in ASD
The rationale for the administration of GFCF diets in people with ASD stems mainly from the effects of opioid peptides, released by the digestion of nutritional gluten and casein. The opioid ex-cess hypothesis, first proposed in 1979, draws parallels between the acute behavioral effects of opiates and the symptoms of ASD and speculates that autism may be “an emotional disturbance aris-ing from an upset in the opiate systems in the brain” (Panksepp, 1979). The opioid excess hypothesis postulates that opioid pep-tides produced through the metabolism of gluten and casein can
pass across an abnormally permeable intestinal membrane and, in consequence, are capable of exerting effects on neurotransmission by binding to opioid receptors (Panksepp, 1979). Opioid receptors may be involved in the regulation of social behavior and possibly also in the pathogenesis of ASD (Genuis and Lobo, 2014). Gluten and casein have similar molecular structures and are metabolized to gluteomorphine and casomorphine, respectively. These peptides have been suggested to accumulate and to enter the blood circula-tion through an elevated permeability of the intestinal membrane (“leaky gut”) in ASD (D’Eufemia et al., 1996). The opioid pep-tides formed during digestion might then permeate the blood-brain barrier and act directly on the brain by binding to opiate receptors and mimicking the effects of opiate drugs (Shattock and Whiteley, 2002). In doing so, they could cause an increase in the activity of the endogenous opioid system, which has been speculated to be linked to the behavioral symptoms of ASD (Panksepp, 1979). However, the opioid excess hypothesis has been criticized, since the available studies have been unable to reveal abnormal opioid levels in the plasma or brain of children with ASD (Lázaro et al., 2016).
Gastrointestinal problems, such as gastrointestinal inflamma-tion, abdominal pain, diarrhea and chronic constipation, are com-monly seen in children with ASD (Buie et al., 2010a; Buie et al., 2010b; Kang et al., 2014). These problems have been found to cor-relate with the severity of ASD (Adams et al., 2011) and may be partly due to digestive enzyme deficiencies, food sensitivities and small bowel inflammation (Kang et al., 2017). The gastrointestinal symptoms in children with ASD indicate a potential link between ASD and celiac disease or gluten sensitivity. Celiac disease is a gastrointestinal disease characterized by a gluten sensitivity, which is responsible for the immunogenic response and mucosal inflam-mation, intestinal villi atrophy and increased intestinal permeabil-ity associated with the disorder (Kelly et al., 2015). Since celiac disease is triggered by the consumption of food containing gluten (Elli et al., 2015), the only treatment currently available for celiac disease is to follow a strict lifelong gluten-free diet (Hill et al., 2016). Today, large numbers of individuals without celiac disease avoid gluten in the belief that a gluten-free diet is associated with health benefits, potentially resulting in the unnecessary consump-tion of gluten-free foods (Gaesser and Angadi, 2015).
A comorbidity between ASD and celiac disease has been sug-gested by several large-scale epidemiological studies (Atladóttir et al., 2009; Butwicka et al., 2017; Lebwohl et al., 2021). Further-more, a predisposition towards autoimmunity has been suggested (Money et al., 1971). In comparison with controls, children with ASD have been reported to display elevated markers of innate and adaptive immune response (Jyonouchi et al., 2001) and, in particu-lar, to have higher concentrations of proinflammatory cytokines following exposure to gluten and casein (Jyonouchi et al., 2002). The mechanism of an elevated immune reactivity to gluten ob-served in a subgroup of children with ASD has been suggested to be distinct from that in celiac disease (Lau et al., 2013). An increase in antigliadin antibody response and its association with gastrointestinal symptoms in these children has been interpreted as an indication of immunological or intestinal permeability abnor-malities (Lau et al., 2013). These findings are the basis of inves-tigations into a link between ASD and food allergies (Jyonouchi, 2009; Li et al., 2021).
The opioid excess hypothesis predicts elevated urinary levels of opioid peptides as potential biomarkers of ASD. As a thera-peutic consequence, diets low in gluten and casein have been hypothesized not only to normalize the urinary peptide levels but also to ameliorate the behavioral symptoms of children with ASD (Reichelt et al., 1991; Shattock and Whiteley, 2002). Gluten-free

Journal of Food Bioactives | www.isnff-jfb.com 5
Lange et al. Gluten-free and casein-free diet in autism
diets involve the dietary exclusion of gluten in grains and various processed food products, while casein-free diets exclude casein in dairy products. Numerous studies have examined the combination of these diets, the GFCF diet, as an alternative intervention in chil-dren with ASD in light of the limited effectiveness of the available therapies.
Studies examining the urinary profiles of children with autistic syndromes have found elevated concentrations of certain peptides (Cade et al., 2000; Israngkun et al., 1986; Knivsberg et al., 1990; Knivsberg et al., 1995; Reichelt et al., 1981; Reichelt et al., 1986; Reichelt et al., 1991). Furthermore, reductions in both these pep-tide concentrations and autistic symptomatology in children adher-ing to a GFCF diet have been reported (Cade et al., 2000; Knivs-berg et al., 1990; Knivsberg et al., 1995), advancing the popularity of the GFCF diet in media reports (Schreck et al., 2013).
In addition to a hypothetical opioid activity of improperly di-gested gluten products, research on pathological interactions be-tween gluten and ASD has focused on inflammation caused by oxi-dative stress and reactivity with anti-gluten antibodies (see Table 1). These hypotheses provide some explanation for the comorbid-ity found between ASD and celiac disease. The intestinal microbi-ome has been suggested to be a possible causative or facilitative agent in mental disorders, including ASD (Doenyas, 2018a; Lange et al., 2020; Li et al., 2017; Vuong and Hsiao, 2017). ASD has been linked to an abnormal gut microbiota composition, with distinct increases or decreases in certain microbial groups being found in the stool of children with ASD (Finegold, 2011; Mulle et al., 2013; O’Mahony et al., 2015; Rosenfeld, 2015; Strati et al., 2017; van de Sande et al., 2014; Wang et al., 2013). However, other studies have found no compositional perturbations of gut microbiota in people
with autism (Son et al., 2015). The changes in gut microbial com-munities are frequently accompanied by strong food preferences and may contribute to the comorbid gastrointestinal symptoms and possibly also to the behavioral problems observed in children with ASD (McElhanon et al., 2014). However, reverse causation, with behavioral changes leading to microbiota perturbations, may also explain the associations between the gut microbiome and ASD. Interactions of probiotics and gluten-casein-free diets with gut microbiota may downregulate gastrointestinal inflammation and intestinal permeability (Doenyas, 2018b).
3. GFCF diet in ASD
A systematic review of the available literature (Reissmann et al., 2020) identified 18 survey studies seeking to explore the popular-ity of complementary and alternative medicine (CAM) treatments among parents of children diagnosed with ASD. Parents reported the use of multiple CAM treatments, particularly dietary treatment forms. Dietary CAM treatments included vitamin or mineral sup-plementation and specific forms of diet (e.g. Feingold diet, GFCF diet, sugar-free diet), of which the GFCF diet was the most com-mon (Green et al., 2006; Salomone et al., 2015; Smith and An-tolovich, 2000). Higher levels of parental education, more severe ASD symptoms, comorbid disorders and younger age of children with ASD have been shown to be associated with the use of CAM treatments (Reissmann et al., 2020). The prevalence, reported by parents, of a previous use of one or more of various CAM treat-ments in their children with ASD was 54–81%, while the current
Table 1. Hypothetical mechanisms of action of the GFCF diet in ASD
Hypothetical mechanism Findings
Excess opioid activity Opioid overactivity leading to decreased social behavior in animal models of ASD (Panksepp, 1979). Possible involvement of opioid receptors in the pathogenesis of ASD (Genuis and Lobo, 2014). Increased (and also decreased) levels of opioid-like peptides, produced through gluten and casein metabolism and entering the blood circulation through a “leaky gut” (D’Eufemia et al., 1996), in the serum, cerebrospinal fluid or urine of individuals with ASD (Pellissier et al., 2018).
Increased autoimmunity Elevated markers of innate and adaptive immune response (cytokines, cytokine tumor necrosis factor-α etc.) in children with ASD (with regression) (Jyonouchi et al., 2001; Money et al., 1971) leading to a hypothetical “autoimmune phenotype” with environmental stimuli triggering an immune response which exacerbates ASD features. Hypothetical knock-on effects of dietary triggers on molecular pathways with high levels of antibodies against gliadin, casein and the digestive enzyme dipeptidyl peptidase IV, which is involved in the processing of gliadin (Vojdani et al., 2003, 2004a, 2004b).
Oxidative stress and inflammation
Impaired antioxidant defense in the cerebellum of individuals with ASD (Gu et al., 2013). Problems in the metabolism of nitrous oxide leading to increased oxidative stress (Frye and Slattery, 2016; Wrońska-Nofer et al., 2012). Increased oxidative stress triggered by gliadin (Monguzzi et al., 2019). Increased oxidative stress demonstrated by elevated markers of oxidative stress in untreated children with celiac disease (Stojiljković et al., 2007). Possible release of pro-inflammatory cytokines contributing to gastrointestinal inflammation (leading to “leaky gut”) due to oxidative stress associated with low glutathione levels (Elder et al., 2006).
Reactivity of antibodies to gluten products
Possible indication of gluten sensitivity in celiac disease and ASD by presence of antibodies against tissue-transglutaminase (Cervio et al., 2007), transglutaminase 6 (Hadjivassiliou et al., 2013; Józefczuk et al., 2018) and gliadin (Cade et al., 2000; De Magistris et al., 2013; Kawashti et al., 2006; Lau et al., 2013; Vojdani et al., 2003, 2004a, 2004b) Potential for antibodies and other irritants to travel along the gut-brain axis indicated by findings of inflammation and abnormal intestinal permeability in ASD (Ashwood et al., 2003; Ashwood et al., 2004; Jyonouchi et al., 2002; Souza et al., 2012; Torrente et al., 2002).
Gut microbiota perturbations
Possible link between abnormal gut microbiota composition and ASD (Finegold, 2011; Mulle et al., 2013; O’Mahony et al., 2015; Rosenfeld, 2015; Wang et al., 2013). Possible downregulation of gastrointestinal inflammation and intestinal permeability by interaction between GFCF diet and gut microbiota (Doenyas, 2018b).

Journal of Food Bioactives | www.isnff-jfb.com6
Gluten-free and casein-free diet in autism Lange et al.
use of at least one CAM treatment was reported by 28–62% of parents (Reissmann et al., 2020). The current use of a GFCF diet was found in 8–32% and previous use in 20–55% of families. Up to 41–69% of parents reported positive effects of the GFCF diet on various aspects of their children’s functioning (Carter et al., 2011b; Christon et al., 2010; Hanson et al., 2007; Hopf et al., 2016; Smith and Antolovich, 2000; Winburn et al., 2014). However, the core features of autistic symptoms may be other than those influ-enced most effectively by the GFCF diet. In a survey study from the United Kingdom, 20–29% of the parents reported a significant improvement in the ASD core dimensions (social interaction, com-munication, repetitive behaviors, restricted interests) (Winburn et al., 2014), 54% of parents reported significant improvements re-garding gastrointestinal symptoms and 42% reported significant positive effects on attention and concentration in their children (Winburn et al., 2014). Significant effects of the GFCF diet on co-morbid problems were also shown in another survey, which found more positive effects of a GFCF diet, as reported by parents, when their children presented with gastrointestinal symptoms or signs of allergy (Pennesi and Klein, 2012). These findings suggest that there may be a subgroup of children with ASD who could benefit from a GFCF diet. However, this should be further investigated and needs to be validated by clinical observations in addition to those of parents.
3.1. Intervention studies using the GFCF diet
First reports of the potential effectiveness of a GFCF diet in ASD were published in the 1990s. For example, a study comprising 15 individuals with autistic syndromes found a reduction in urinary peptides resulting from the metabolism of gluten and casein and an improvement in some behaviors associated with autism in follow-up assessments after one year (Knivsberg et al., 1990) and four years (Knivsberg et al., 1995). Similar behavioral results were re-ported in a follow-up study of a 5-month administration of a glu-ten-free diet in 22 children with autism and associated spectrum disorders, while no significant reductions in urinary peptide levels were found (Whiteley et al., 1999). However, major limitations of these studies included open methodology and a lack of randomiza-tion and blinding. Numerous investigations of variable scientific rigor have since been conducted to determine the effects of a GFCF diet on behavioral changes in children diagnosed with ASD. The effects of a GFCF diet on the symptoms ASD have been assessed in several interventional studies, some of which were reviewed in a Cochrane report (Millward et al., 2008). This report included only two small randomized controlled trials and reported mixed results regarding GFCF dietary effects. A more recent systematic review of the literature included dietary intervention trials and food chal-lenge studies (Reissmann et al., 2020), with the classification of report strength and the judgment of quality indicators performed according to criteria proposed for the evaluation of intervention studies in ASD research (Reichow et al., 2008). The main findings of this systematic review will be presented briefly below.
Several case studies, including anecdotal case reports (Adams and Conn, 1997; Fields and Fields, 1976) and more scientific tri-als (Herbert and Buckley, 2013; Hsu et al., 2009; Knivsberg et al., 1999), have sought to establish a role of the GFCF diet in the relief of symptoms of autism. These studies found some evidence for positive effects of the GFCF diet for at least some of the measures assessed, such as symptoms of autism, cognitive skills and physi-cal development. However, the findings of these reports should be considered as weak evidence at best, since none of them were con-ducted with adequate scientific rigor (see Reissmann et al., 2020).
The effects of GFCF dietary intervention were also examined in several group studies. Of six uncontrolled group trials assess-ing the elimination of both gluten and casein (Cade et al., 2000; Gemmell and Chambliss, 1997; Jyonouchi et al., 2002; Knivsberg et al., 1990; Knivsberg et al., 1995; Lucarelli et al., 1995; Pontino et al., 1998; Whiteley et al., 1999), all but one study (Gemmell and Chambliss, 1997; Pontino et al., 1998) found positive dietary ef-fects on the core symptoms of ASD, cognitive deficits, gastrointes-tinal problems or comorbid symptoms. These trials lacked control procedures and had various other major methodological problems, resulting in only weak scientific evidence, which should be treated with caution (see Reissmann et al., 2020). In regard to five con-trolled group trials (Elder et al., 2006; Ghalichi et al., 2016; John-son et al., 2011; Knivsberg et al., 2002; Knivsberg et al., 2003; Pedersen et al., 2014; Seung et al., 2007; Whiteley et al., 2010), the scientific rigor was adequate in all but one study (Knivsberg et al., 2002; Knivsberg et al., 2003). Two favorable evaluations of GFCF diet effects are at variance with the negative findings of two stud-ies with diet adherence lasting six (Elder et al., 2006; Seung et al., 2007) to 12 weeks (Johnson et al., 2011). These negative results are supported by another small, randomized controlled study of the GFCF diet in young children with ASD (Johnson et al., 2011). A further GFCF diet study in children with ASD, conducted with adequate scientific rigor, failed to show positive effects on ASD symptoms, neurodevelopmental ratings or symptoms of attention and hyperactivity (Pedersen et al., 2014; Whiteley et al., 2010).
Another systematic review based on the findings of six rand-omized controlled trials updated the evidence of the therapeutic effectiveness of the GFCF diet in childhood ASD and concluded that there is little evidence supporting beneficial effects of the diet (Piwowarczyk et al., 2018). A recent systematic review and meta-analysis, based on six relevant randomized controlled trials investigating possible benefits of the GFCF diet in children diag-nosed with ASD found no effects on clinician-reported autism core symptoms, parent-reported functional level or behavioral difficul-ties (Keller et al., 2021). The quality of evidence ranged from low to very low due to serious risk of bias, inconsistency and impreci-sion.
Taken together, the available dietary intervention studies have found highly divergent results and do not therefore allow for clear-cut conclusions regarding the effects of the GFCF diet in children with ASD (see Table 2). It is important to note that the majority of trials conducted without adequate scientific rigor found evidence of beneficial effects of the GFCF diet, while more rigorous scien-tific evaluations were unable to show a consistent pattern of results. Many trials are hampered by various methodological flaws, such as reliance on parental reports as the sole source of information on ASD symptoms as well as a lack of adequate control groups, rater blindness, control for additional treatments or assessment of treatment fidelity. Future studies on the efficacy of the GFCF diet would need to take these aspects into account. In particular, the ratings of ASD symptoms should be performed by uninvolved and blinded clinicians rather than parents.
Given that many positive findings following a GFCF diet in ASD have been reported by studies with longer rather than shorter follow-up periods (see Reissmann et al., 2020), longer follow-up periods involving multiple clinical assessments should be em-ployed. Significant beneficial effects have been found in studies with a duration of 12 months, while studies with interventions lasting 12 weeks showed no significant effects (de Magistris et al., 2013; Elder et al., 2006; Gogou and Kolios, 2017; Gogou and Kolios, 2018; Hyman et al., 2016; Knivsberg et al., 2002; Whiteley et al., 2010). A prolonged administration may be required to reveal benefits of GFCF diets, since inflammation, dysbiosis and altered

Journal of Food Bioactives | www.isnff-jfb.com 7
Lange et al. Gluten-free and casein-free diet in autism
intestinal permeability found in many people with ASD may be normalized only following extended periods of dietary interven-tion.
3.2. Challenge studies with gluten and/or casein
On the basis of research on food allergies, several studies have ex-amined the effects of challenges with gluten and/or casein in chil-dren adhering to some form of GFCF diet. In a case study, acute negative effects were reported in a child following the consump-tion of gluten-/casein-containing foods, particularly wheat prod-ucts (O’Banion et al., 1978). An uncontrolled group study found that a challenge with casein and other food allergens in children with ASD on a casein-free diet was associated with an increase in behavioral symptoms, such as inappropriate emotional responses, motor disturbances as well as disturbed concentration and percep-tion (Lucarelli et al., 1995). Other studies failed to show any clear effects of such dietary challenges (Bird et al., 1977; Hyman et al., 2016; Irvin, 2006; McCarthy and Coleman, 1979; Navarro et al., 2015). The most persuasive evidence stems from a small rand-omized double-blind placebo-controlled challenge study, in which children with ASD who had followed a GFCF diet regimen for two weeks before being randomly assigned to either a gluten/casein challenge or a placebo group (Navarro et al., 2015). The results of this study found no consistent effects of the gluten/casein chal-lenge on gastrointestinal symptoms or behavioral problems, such as inattention, hyperactivity or inattention (Navarro et al., 2015). Taken together, the majority of the available challenge studies were unable to show clear effects of a gluten/casein challenge on ASD symptoms, gastrointestinal symptoms, comorbid behavior problems or cognitive functioning. However, the majority of chal-lenge studies were conducted with inadequate scientific rigor, and the available evidence needs to be viewed with caution.
3.3. Potential harms of the GFCF diet
The available studies exploring potential harms of the GFCF diet in children with ASD have mainly assessed nutritional adequacy or physical development. Four studies have examined possible nu-tritional deficiencies of children with ASD on restriction diets in comparison with children with ASD on unrestricted diets or healthy control children (Arnold et al., 2003; Cornish, 2002; Hyman et al., 2012; Marí-Bauset et al., 2016). Three of these studies estimated nutritional adequacy from dietary records maintained by parents and found no evidence of more nutritional deficiencies following a GFCF diet than in the comparison groups (Cornish, 2002; Hyman et al., 2012; Marí-Bauset et al., 2016). The fourth study sought to estimate deficiencies from plasma-derived concentrations of
amino acids and found deficiencies in several essential amino ac-ids, including the neurotransmitter precursors tyrosine and trypto-phan (Arnold et al., 2003). Two studies of physical development following a GFCF diet examined bone development of children with ASD on a casein-free diet (with a low intake of calcium) with children on unrestricted diets (Hediger et al., 2008; Neumeyer et al., 2013). These studies reported that children with ASD generally displayed a decrease in bone density, with a significantly greater reduction in the children on a casein-free diet (Hediger et al., 2008; Neumeyer et al., 2013). These aspects require further considera-tion in future investigations of GFCF diets.
4. Future directions
The gluten-free diet is a well-established treatment for individuals with heightened sensitivity to gluten and gluten-related disorders, such as celiac disease, allergic reactions (wheat allergy), non-ce-liac gluten sensitivity and other immune-mediated disorders (e.g. gluten ataxia and dermatitis herpetiformis) (Sapone et al., 2012). However, the utility of the gluten-free diet in other conditions is not evidence-based (Jones, 2017).
Studies examining potential benefits of the GFCF diet in chil-dren with ASD need to address numerous methodological problems and use research designs with adequate control procedures (see Ta-ble 3). Other recommendations include an (extended) trial duration of at least 12 months and the use of a broader range of measures, which may provide insight into other relevant aspects, such as moderators and mediators of therapeutic effects as well as potential risks of the GFCF diet. Another important problem to be addressed is the reliance on parents as an information source. Parents as un-blinded providers of treatment may be biased in their perception of dietary outcomes. Thus, future studies should implement clinician-administered rating procedures and use objective observational measures and standardized and ecologically valid assessment in-struments. In order to identify potential responders to GFCF diets, a range of variables, including comorbid gastrointestinal symptoms (Díaz-Atienza et al., 2012), intestinal permeability (Boukthir et al., 2010) as well as gastrointestinal enzymatic and inflammatory activ-ity (Wang et al., 2009), deserve further examination.
Future studies should include children with established distur-bances in the metabolic breakdown of food proteins or those at risk for allergies to gluten or casein. A diet-related phenotype of ASD may be associated with biological aberrations linked to abnormal gut-brain axis functioning (Whiteley, 2015). The identification of such a diet-related ASD phenotype may help in selecting those children who might benefit from a GFCF dietary intervention.
The findings of a recent study suggest that an intervention in-cluding a wider variety of dietary elements may be effective in the
Table 2. Summary of findings of GFCF dietary intervention studies in children with ASD
Highly divergent findings of GFCF diet intervention studies
Methodological flaws of many GFCF intervention studies: strong reliance on (unblinded) parental reports as the sole information source regarding ASD symptoms; frequent lack of control procedures (control groups, measurement baselines, control for additional treatments); frequent lack of assessment of treatment fidelity
Positive effects of GFCF diet adherence mainly found in studies conducted without adequate scientific rigor
No consistent pattern of findings following a GFCF diet in more rigorous scientific evaluations
More positive findings reported by case/group studies with longer follow-up periods
Summary: No clear-cut conclusions possible regarding GFCF diet effects in children with ASD

Journal of Food Bioactives | www.isnff-jfb.com8
Gluten-free and casein-free diet in autism Lange et al.
treatment of childhood ASD (Adams et al., 2018). The randomized, controlled, single-blind 12-month trial of a comprehensive dietary and nutritional intervention in 67 children and adults with ASD aged 3–58 years and 50 non-sibling neurotypical controls exam-ined the effects of a special vitamin/mineral supplement and fur-ther treatments added sequentially, including essential fatty acids, Epsom salt baths, carnitine, digestive enzyme and a GFCF soy-free diet (Adams et al., 2018). In comparison with the non-treat-ment group, the treated group showed significant improvements in nonverbal intellectual ability as well as ASD symptoms and de-velopmental age. According to parental reports, vitamin/mineral supplements, essential fatty acids and the gluten/casein/soy-free diet were the most beneficial (Adams et al., 2018). Furthermore, FODMAP food components (fermentable oligosaccharides, disac-charides, monosaccharides and polyols) have increasingly been in-vestigated due to their possible relation with extraintestinal medi-cal conditions. In particular, diets low in FODMAPs might have beneficial effects on various mental conditions, including ASD (Aranburu et al., 2021). However, this requires further research.
5. Conclusion
Despite the popularity and widespread use of the GFCF diet as a supplementary treatment in children with ASD, several rigorous scientific evaluations have failed to confirm the effectiveness of this approach. While the available dietary studies do not confirm the conceptualization of ASD as a metabolic disorder or the predic-tions of the opioid excess hypothesis, the increasing evidence for associations between gut anomalies and the brain in children with ASD points to a role of immunological factors in the frequently reported gastrointestinal problems. Therefore, the consideration of dietary factors, including gluten and casein, should not be dis-regarded prematurely. Food-derived proteins could play a role in triggering allergic responses, which may influence brain devel-opment and neuronal functions. More scientifically rigorous and methodologically sound investigations are required to examine the importance of dietary factors in the etiology and treatment of ASD.
In summary, the available evidence in support of the effec-tiveness of the GFCF diet in the treatment of childhood ASD is very weak and cannot currently even be considered promising. Potentially ineffective dietary therapies may imply considerable opportunity costs, with other possibly more effective treatment ap-proaches remaining unutilized.
References
Adams, J.B., Audhya, T., Geis, E., Gehn, E., Fimbres, V., Pollard, E.L., Mitch-ell, J., Ingram, J., Hellmers, R., Laake, D., Matthews, J.S., Li, K., Na-viaux, J.C., Naviaux, R.K., Adams, R.L., Coleman, D.M., and Quig, D.W. (2018). Comprehensive nutritional and dietary intervention for au-tism spectrum disorder – A randomized, controlled 12-month trial. Nutrients 10: 369.
Adams, J.B., Johansen, L.J., Powell, L.D., Quig, D., and Rubin, R.A. (2011). Gastrointestinal flora and gastrointestinal status in children with au-tism – comparisons to typical children and correlation with autism severity. BMC Gastroenterol. 11: 22.
Adams, L., and Conn, S. (1997). Nutrition and its relationship to autism. Focus Autism Other Dev. Disabl. 12: 53–58.
American Psychiatric Association. (2013). Diagnostic and statistical manu-al of mental disorders. 5 ed. American Psychiatric Publishing, Wash-ington, DC.
Aranburu, E., Matias, S., Simón, E., Larretxi, I., Martínez, O., Bustamante, M.Á., Del Fernández-Gil, M.P., and Miranda, J. (2021). Gluten and FODMAPs relationship with mental disorders: Systematic review. Nu-trients 13: 1894.
Arnold, G.L., Hyman, S.L., Mooney, R.A., and Kirby, R.S. (2003). Plasma amino acids profiles in children with autism: potential risk of nutri-tional deficiencies. J. Autism Dev. Disord. 33: 449–454.
Ashwood, P., Anthony, A., Pellicer, A.A., Torrente, F., Walker-Smith, J.A., and Wakefield, A.J. (2003). Intestinal lymphocyte populations in chil-dren with regressive autism: Evidence for extensive mucosal immu-nopathology. J. Clin. Immunol. 23: 504–517.
Ashwood, P., Anthony, A., Torrente, F., and Wakefield, A.J. (2004). Spon-taneous mucosal lymphocyte cytokine profiles in children with au-tism and gastrointestinal symptoms: Mucosal immune activation and reduced counter regulatory interleukin-10. J. Clin. Immunol. 24: 664–673.
Atladóttir, H.O., Pedersen, M.G., Thorsen, P., Mortensen, P.B., Deleuran, B., Eaton, W.W., and Parner, E.T. (2009). Association of family history of autoimmune diseases and autism spectrum disorders. Pediatrics 124: 687–694.
Bauman, M.L., and Kemper, T.L. (2005). Neuroanatomic observations of the brain in autism: a review and future directions. Int. J. Dev. Neu-rosci. 23: 183–187.
Bird, B.L., Russo, D.C., and Cataldo, M.F. (1977). Considerations in the analysis and treatment of dietary effects on behavior: a case study. J. Autism Child. Schizophr. 7: 373–382.
Boukthir, S., Matoussi, N., Belhadj, A., Mammou, S., Dlala, S.B., Helayem, M., Rocchiccioli, F., Bouzaidi, S., and Abdennebi, M. (2010). Anoma-lies de la perméabilité intestinale chez l’enfant autiste: [Abnormal intestinal permeability in children with autism]. Tunis Med. 88: 685–686.
Buie, T., Campbell, D.B., Fuchs, G.J., Furuta, G.T., Levy, J., Vandewater, J.,
Table 3. Methodological recommendations regarding GFCF studies in ASD (see also Reissmann et al., 2020)
Randomized controlled trial with adequate control condition and control for attention effects (e.g. nutritionist counseling) in control group
Assessment of additional therapies
Sufficient trial duration (>12 months) with multiple follow-up assessments
Ensuring and maintaining treatment fidelity (diet adherence)
Blinded clinician ratings in addition to parent ratings of behavior
Multi-method/multi-rater assessment using standardized assessment tools: clinical measures, rating scales for parents and teachers, neuropsychological and cognitive tests, ecologically valid behavioral observations in natural settings
Sufficiently large sample size (>30) with control of attrition
Risk measurement regarding nutritional status, physical development, bone density etc
Assessment of potential moderating and mediating variables, such as food allergies, gastrointestinal symptoms and behavioral symptoms (e.g. attention, hyperactivity)

Journal of Food Bioactives | www.isnff-jfb.com 9
Lange et al. Gluten-free and casein-free diet in autism
Whitaker, A.H., Atkins, D., Bauman, M.L., Beaudet, A.L., Carr, E.G., Gershon, M.D., Hyman, S.L., Jirapinyo, P., Jyonouchi, H., Kooros, K., Kushak, R., Levitt, P., Levy, S.E., Lewis, J.D., Murray, K.F., Natowicz, M.R., Sabra, A., Wershil, B.K., Weston, S.C., Zeltzer, L., and Winter, H. (2010a). Evaluation, diagnosis, and treatment of gastrointestinal disorders in individuals with ASDs: a consensus report. Pediatrics 125(Suppl. 1): S1–S18.
Buie, T., Fuchs, G.J., Furuta, G.T., Kooros, K., Levy, J., Lewis, J.D., Wershil, B.K., and Winter, H. (2010b). Recommendations for evaluation and treatment of common gastrointestinal problems in children with ASDs. Pediatrics 125(Suppl. 1): S19–S29.
Butwicka, A., Lichtenstein, P., Frisén, L., Almqvist, C., Larsson, H., and Lud-vigsson, J.F. (2017). Celiac disease is associated with childhood psy-chiatric disorders: A population-based study. J. Pediatr. 184: 87–93.
Cade, R., Privette, M., Fregly, M., Rowland, N., Sun, Z., Zele, V., Wagemak-er, H., and Edelstein, C. (2000). Autism and schizophrenia: intestinal disorders. Nutr. Neurosci. 3: 57–72.
Carter, A.S., Messinger, D.S., Stone, W.L., Celimli, S., Nahmias, A.S., and Yo-der, P. (2011a). A randomized controlled trial of Hanen’s ‘More Than Words’ in toddlers with early autism symptoms. J. Child Psychol. Psy-chiatry 52: 741–752.
Carter, M., Roberts, J., Williams, K., Evans, D., Parmenter, T., Silove, N., Clark, T., and Warren, A. (2011b). Interventions used with an Aus-tralian sample of preschool children with autism spectrum disorders. Res. Autism Spectr. Disord. 5: 1033–1041.
Cervio, E., Volta, U., Verri, M., Boschi, F., Pastoris, O., Granito, A., Barbara, G., Parisi, C., Felicani, C., Tonini, M., and De Giorgio, R. (2007). Sera of patients with celiac disease and neurologic disorders evoke a mitochondrial-dependent apoptosis in vitro. Gastroenterology 133: 195–206.
Christon, L.M., Mackintosh, V.H., and Myers, B.J. (2010). Use of comple-mentary and alternative medicine (CAM) treatments by parents of children with autism spectrum disorders. Res. Autism Spectr. Disord. 4: 249–259.
Cornish, E. (2002). Gluten and casein free diets in autism: a study of the effects on food choice and nutrition. J. Hum. Nutr. Diet. 15: 261–269.
De Magistris, L., Picardi, A., Siniscalco, D., Riccio, M.P., Sapone, A., Cariello, R., Abbadessa, S., Medici, N., Lammers, K.M., Schiraldi, C., Iardino, P., Marotta, R., Tolone, C., Fasano, A., Pascotto, A., and Bravaccio, C. (2013). Antibodies against food antigens in patients with autistic spectrum disorders. BioMed Res. Int. 2013(2013): 729349.
D’Eufemia, P., Celli, M., Finocchiaro, R., Pacifico, L., Viozzi, L., Zaccagnini, M., Cardi, E., and Giardini, O. (1996). Abnormal intestinal permeabil-ity in children with autism. Acta Paediatr. 85: 1076–1079.
Díaz Atienza, F., Serrano Nieto, S., Gonzalez Domenech, P., and García Pablos, C. (2012). Prevalence of feeding disorders, gastrointestinal disorders and recurrent infections in children with autism spectrum disorders (ASD) compared with their healthy siblings. Rev. Psiquiatr. Infant.-Juv. 29: 11–16.
Doenyas, C. (2018a). Gut microbiota, inflammation, and probiotics on neural development in autism spectrum disorder. Neuroscience 374: 271–286.
Doenyas, C. (2018b). Dietary interventions for autism spectrum disor-der: new perspectives from the gut-brain axis. Physiol. Behav. 194: 577–582.
Dykens, E.M., Fisher, M.H., Taylor, J.L., Lambert, W., and Miodrag, N. (2014). Reducing distress in mothers of children with autism and other disabilities: a randomized trial. Pediatrics 134: e454–e463.
Ecker, C., Bookheimer, S.Y., and Murphy, D.G.M. (2015). Neuroimaging in autism spectrum disorder: brain structure and function across the lifespan. Lancet Neurol. 14: 1121–1134.
Elder, J.H., Shankar, M., Shuster, J., Theriaque, D., Burns, S., and Sherrill, L. (2006). The gluten-free, casein-free diet in autism: results of a pre-liminary double blind clinical trial. J. Autism Dev. Disord. 36: 413–420.
Elli, L., Branchi, F., Tomba, C., Villalta, D., Norsa, L., Ferretti, F., Roncoroni, L., and Bardella, M.T. (2015). Diagnosis of gluten related disorders: Celiac disease, wheat allergy and non-celiac gluten sensitivity. World J. Gastroenterol. 21: 7110–7119.
Elsabbagh, M., Divan, G., Koh, Y.-J., Kim, Y.S., Kauchali, S., Marcín, C., Mon-tiel-Nava, C., Patel, V., Paula, C.S., Wang, C., Yasamy, M.T., and Fom-bonne, E. (2012). Global prevalence of autism and other pervasive
developmental disorders. Autism Res. 5: 160–179.Fields, M., and Fields, P. (1976). The relationship between problem behav-
ior and food allergies: one family’s story. J. Autism Child. Schizophr. 6: 75–91.
Finegold, S.M. (2011). Desulfovibrio species are potentially important in regressive autism. Med. Hypotheses 77: 270–274.
Fombonne, E. (2009). Epidemiology of pervasive developmental disor-ders. Pediatr. Res. 65: 591–598.
Frye, R.E., and Slattery, J. (2016). The potential role of nitrous oxide in the etiology of autism spectrum disorder. Transl. Psychiatry 6: e812.
Fung, L.K., Mahajan, R., Nozzolillo, A., Bernal, P., Krasner, A., Jo, B., Coury, D., Whitaker, A., Veenstra-VanderWeele, J., and Hardan, A.Y. (2016). Pharmacologic treatment of severe irritability and problem behav-iors in autism: A systematic review and meta-analysis. Pediatrics 137(Suppl. 2): S124–S135.
Gaesser, G.A., and Angadi, S.S. (2015). Navigating the gluten-free boom. JAAPA 28: 1–7.
Gaugler, T., Klei, L., Sanders, S.J., Bodea, C.A., Goldberg, A.P., Lee, A.B., Mahajan, M., Manaa, D., Pawitan, Y., Reichert, J., Ripke, S., Sandin, S., Sklar, P., Svantesson, O., Reichenberg, A., Hultman, C.M., Devlin, B., Roeder, K., and Buxbaum, J.D. (2014). Most genetic risk for autism resides with common variation. Nat. Genet. 46: 881–885.
Gemmell, M., and Chambliss, C. (1997). Effects of a gluten-free diet on rate of achievement in autistic children in an applied behavioral anal-ysis program: Research Report: Ursinus College (ED406761).
Genuis, S.J., and Lobo, R.A. (2014). Gluten sensitivity presenting as a neu-ropsychiatric disorder. Gastroenterol. Res. Pract. 2014: 293206.
Ghalichi, F., Ghaemmaghami, J., Malek, A., and Ostadrahimi, A. (2016). Ef-fect of gluten free diet on gastrointestinal and behavioral indices for children with autism spectrum disorders: a randomized clinical trial. World J. Pediatr. 12: 436–442.
Gogou, M., and Kolios, G. (2017). The effect of dietary supplements on clinical aspects of autism spectrum disorder: A systematic review of the literature. Brain Dev. 39: 656–664.
Gogou, M., and Kolios, G. (2018). Are therapeutic diets an emerging ad-ditional choice in autism spectrum disorder management? World J. Pediatr. 14: 215–223.
Gotham, K., Brunwasser, S.M., and Lord, C. (2015). Depressive and anxiety symptom trajectories from school age through young adulthood in samples with autism spectrum disorder and developmental delay. J. Am. Acad. Child Adolesc. Psychiatry 54: 369–376.
Green, V.A., Pituch, K.A., Itchon, J., Choi, A., O’Reilly, M., and Sigafoos, J. (2006). Internet survey of treatments used by parents of children with autism. Res. Dev. Disabil. 27: 70–84.
Gu, F., Chauhan, V., and Chauhan, A. (2013). Impaired synthesis and anti-oxidant defense of glutathione in the cerebellum of autistic subjects: Alterations in the activities and protein expression of glutathione-related enzymes. Free Radic. Biol. Med. 65: 488–496.
Hadjivassiliou, M., Aeschlimann, P., Sanders, D.S., Mäki, M., Kaukinen, K., Grünewald, R.A., Bandmann, O., Woodroofe, N., Haddock, G., and Aeschlimann, D.P. (2013). Transglutaminase 6 antibodies in the diag-nosis of gluten ataxia. Neurology. 80: 1740–1745.
Hanson, E., Kalish, L.A., Bunce, E., Curtis, C., McDaniel, S., Ware, J., and Petry, J. (2007). Use of complementary and alternative medicine among children diagnosed with autism spectrum disorder. J. Autism Dev. Disord. 37: 628–636.
Hazlett, H.C., Gu, H., Munsell, B.C., Kim, S.H., Styner, M., Wolff, J.J., Elison, J.T., Swanson, M.R., Zhu, H., Botteron, K.N., Collins, D.L., Constantino, J.N., Dager, S.R., Estes, A.M., Evans, A.C., Fonov, V.S., Gerig, G., Ko-stopoulos, P., McKinstry, R.C., Pandey, J., Paterson, S., Pruett, J.R., Schultz, R.T., Shaw, D.W., Zwaigenbaum, L., and Piven, J. (2017). Early brain development in infants at high risk for autism spectrum disor-der. Nature 542(7641): 348–351.
Hediger, M.L., England, L.J., Molloy, C.A., Yu, K.F., Manning-Courtney, P., and Mills, J.L. (2008). Reduced bone cortical thickness in boys with autism or autism spectrum disorder. J. Autism Dev. Disord. 38: 848–856.
Herbert, M.R. (2010). Contributions of the environment and environmen-tally vulnerable physiology to autism spectrum disorders. Curr. Opin. Neurol. 23: 103–110.
Herbert, M.R., and Buckley, J.A. (2013). Autism and dietary therapy: case

Journal of Food Bioactives | www.isnff-jfb.com10
Gluten-free and casein-free diet in autism Lange et al.
report and review of the literature. J. Child Neurol. 28: 975–982.Hill, I.D., Fasano, A., Guandalini, S., Hoffenberg, E., Levy, J., Reilly, N., and
Verma, R. (2016). NASPGHAN clinical report on the diagnosis and treatment of gluten-related disorders. J Pediatr Gastroenterol Nutr. 63: 156–165.
Hopf, K.P., Madren, E., and Santianni, K.A. (2016). Use and perceived ef-fectiveness of complementary and alternative medicine to treat and manage the symptoms of autism in children: A survey of parents in a community Population. J. Altern. Complement. Med. 22: 25–32.
Hsu, C.-L., Lin, C.-Y., Chen, C.-L., Wang, C.-M., and Wong, M.-K. (2009). The effects of a gluten and casein-free diet in children with autism: a case report. Chang Gung Med. J. 32: 459–465.
Hyman, S.L., Stewart, P.A., Foley, J., Cain, U., Peck, R., Morris, D.D., Wang, H., and Smith, T. (2016). The gluten-free/casein-free diet: A double-blind challenge trial in children with autism. J. Autism Dev. Disord. 46: 205–220.
Hyman, S.L., Stewart, P.A., Schmidt, B., Cain, U., Lemcke, N., Foley, J.T., Peck, R., Clemons, T., Reynolds, A., Johnson, C., Handen, B., James, S.J., Courtney, P.M., Molloy, C., and Ng, P.K. (2012). Nutrient intake from food in children with autism. Pediatrics 130(Suppl. 2): S145–S153.
Irvin, D.S. (2006). Using analog assessment procedures for determining the effects of a gluten-free and casein-free diet on rate of problem behaviors for an adolescent with autism. Behav. Intervent. 21: 281–286.
Israngkun, P.P., Newman, H.A., Patel, S.T., Duruibe, V.A., and Abou-Issa, H. (1986). Potential biochemical markers for infantile autism. Neuro-chem. Pathol. 5: 51–70.
Johnson, C.R., Handen, B.L., Zimmer, M., Sacco, K., and Turner, K. (2011). Effects of gluten free / casein free diet in young children with autism: A pilot study. J. Dev. Phys. Disabil. 23: 213–225.
Jones, A.L. (2017). The gluten-free diet: Fad or necessity? Diabetes Spectr. 30: 118–123.
Józefczuk, J., Konopka, E., Bierła, J.B., Trojanowska, I., Sowińska, A., Czar-necki, R., Sobol, L., Józefczuk, P., Surdy, W., and Cukrowska, B. (2018). The occurrence of antibodies against gluten in children with autism spectrum disorders does not correlate with serological markers of impaired intestinal permeability. J. Med. Food 21: 181–187.
Jyonouchi, H. (2009). Food allergy and autism spectrum disorders: is there a link? Curr. Allergy Asthma Rep. 9: 194–201.
Jyonouchi, H., Sun, S., and Itokazu, N. (2002). Innate immunity associ-ated with inflammatory responses and cytokine production against common dietary proteins in patients with autism spectrum disorder. Neuropsychobiology 46: 76–84.
Jyonouchi, H., Sun, S., and Le, H. (2001). Proinflammatory and regulatory cytokine production associated with innate and adaptive immune responses in children with autism spectrum disorders and develop-mental regression. J. Neuroimmunol. 120: 170–179.
Kang, D.-W., Adams, J.B., Gregory, A.C., Borody, T., Chittick, L., Fasano, A., Khoruts, A., Geis, E., Maldonado, J., McDonough-Means, S., Pollard, E.L., Roux, S., Sadowsky, M.J., Lipson, K.S., Sullivan, M.B., Caporaso, J.G., and Krajmalnik-Brown, R. (2017). Microbiota transfer therapy al-ters gut ecosystem and improves gastrointestinal and autism symp-toms: an open-label study. Microbiome 5: 10.
Kang, V., Wagner, G.C., and Ming, X. (2014). Gastrointestinal dysfunction in children with autism spectrum disorders. Autism Res. 7: 501–506.
Kanner, L. (1943). Autistic disturbances of affective contact. Nervous Child. 2: 217–250.
Kawashti, M.I., Amin, O.R., and Rowehy, N.G. (2006). Possible immuno-logical disorders in autism: concomitant autoimmunity and immune tolerance. Egypt. J. Immunol. 13: 99–104.
Keller, A., Rimestad, M.L., Friis Rohde, J., Holm Petersen, B., Bruun Korfit-sen, C., Tarp, S., Briciet Lauritsen, M., and Händel, M.N. (2021). The effect of a combined gluten- and casein-free diet on children and ad-olescents with autism spectrum disorders: A systematic review and meta-analysis. Nutrients 13: 470.
Kelly, C.P., Bai, J.C., Liu, E., and Leffler, D.A. (2015). Advances in diagnosis and management of celiac disease. Gastroenterology 148: 1175–1186.
Knivsberg, A.M., Reichelt, K.L., Høien, T., and Nødland, M. (2002). A ran-domised, controlled study of dietary intervention in autistic syn-
dromes. Nutr. Neurosci. 5: 251–261.Knivsberg, A.-M., Reichelt, K.-L., Høien, T., and Nødland, M. (2003). Effect
of a dietary intervention on autistic behavior. Focus Autism Other Dev. Disabl. 18: 248–257.
Knivsberg, A.M., Reichelt, K.L., and Nødland, M. (1999). Dietary interven-tion for a seven year old girl with autistic behaviour. Nutr. Neurosci. 2: 435–439.
Knivsberg, A.-M., Reichelt, K.L., Nødland, M., and Høien, T. (1995). Autis-tic syndromes and diet: a follow-up study. Scand. J. Educ. Res. 39: 223–236.
Knivsberg, A.-M., Wiig, K., Lind, G., Nødland, M., and Reichelt, K.L. (1990). Dietary intervention in autistic syndromes. Brain Dysfunction 3: 315–327.
Lange, K.W., Hauser, J., and Reissmann, A. (2015). Gluten-free and casein-free diets in the therapy of autism. Curr. Opin. Clin. Nutr. Metab. Care 18: 572–575.
Lange, K.W., Lange, K.M., Nakamura, Y., and Kanaya, S. (2020). Is there a role of gut microbiota in mental health? J. Food Bioact. 9: 4–9.
Lau, N.M., Green, P.H.R., Taylor, A.K., Hellberg, D., Ajamian, M., Tan, C.Z., Kosofsky, B.E., Higgins, J.J., Rajadhyaksha, A.M., and Alaedini, A. (2013). Markers of celiac disease and gluten sensitivity in children with autism. PLoS One 8: e66155.
Lázaro, C.P., Pondé, M.P., and Rodrigues, L.E. (2016). Opioid peptides and gastrointestinal symptoms in autism spectrum disorders. Braz. J. Psy-chiatry 38: 243–246.
Lebwohl, B., Haggård, L., Emilsson, L., Söderling, J., Roelstraete, B., Butwic-ka, A., Green, P.H.R., and Ludvigsson, J.F. (2021). Psychiatric disorders in patients with a diagnosis of celiac disease during childhood from 1973 to 2016. Clin. Gastroenterol. Hepatol. 19(10): 2093–2101.E13.
Levy, S.E., Mandell, D.S., and Schultz, R.T. (2009). Autism. Lancet 374(9701): 1627–1638.
Lewis, J.D., Evans, A.C., Pruett, J.R., Botteron, K., Zwaigenbaum, L., Estes, A., Gerig, G., Collins, L., Kostopoulos, P., McKinstry, R., Dager, S., Pat-erson, S., Schultz, R.T., Styner, M., Hazlett, H., and Piven, J. (2014). Network inefficiencies in autism spectrum disorder at 24 months. Transl. Psychiatry 4: e388.
Li, Q., Han, Y., Dy, A.B.C., and Hagerman, R.J. (2017). The gut microbiota and autism spectrum disorders. Front. Cell Neurosci. 11: 120.
Li, H., Liu, H., Chen, X., Zhang, J., Tong, G., and Sun, Y. (2021). Association of food hypersensitivity in children with the risk of autism spectrum disorder: a meta-analysis. Eur. J. Pediatr. 180: 999–1008.
Lucarelli, S., Frediani, T., Zingoni, A.M., Ferruzzi, F., Giardini, O., Quintieri, F., Barbato, M., D’Eufemia, P., and Cardi, E. (1995). Food allergy and infantile autism. Panminerva Med. 37: 137–141.
Lyall, K., Ashwood, P., van de Water, J., and Hertz-Picciotto, I. (2014). Ma-ternal immune-mediated conditions, autism spectrum disorders, and developmental delay. J. Autism Dev. Disord. 44: 1546–1555.
Lyall, K., Croen, L., Daniels, J., Fallin, M.D., Ladd-Acosta, C., Lee, B.K., Park, B.Y., Snyder, N.W., Schendel, D., Volk, H., Windham, G.C., and News-chaffer, C. (2017). The changing epidemiology of autism spectrum disorders. Annu. Rev. Public Health 38: 81–102.
Mandy, W., and Lai, M.-C. (2017). Towards sex- and gender-informed au-tism research. Autism 21: 643–645.
Marí-Bauset, S., Llopis-González, A., Zazpe, I., Marí-Sanchis, A., and Suárez-Varela, M.M. (2016). Nutritional impact of a gluten-free ca-sein-free diet in children with autism spectrum disorder. J. Autism Dev. Disord. 46: 673–684.
McCarthy, D.M., and Coleman, M. (1979). Response of intestinal mucosa to gluten challenge in autistic subjects. Lancet 2(8148): 877–878.
McElhanon, B.O., McCracken, C., Karpen, S., and Sharp, W.G. (2014). Gas-trointestinal symptoms in autism spectrum disorder: a meta-analy-sis. Pediatrics 133: 872–883.
Millward, C., Ferriter, M., Calver, S., and Connell-Jones, G. (2008). Gluten- and casein-free diets for autistic spectrum disorder. Cochrane Data-base Syst. Rev. (2): CD003498.
Monguzzi, E., Marabini, L., Elli, L., Vaira, V., Ferrero, S., Ferretti, F., Branchi, F., Gaudioso, G., Scricciolo, A., Lombardo, V., Doneda, L., and Ron-coroni, L. (2019). Gliadin effect on the oxidative balance and DNA damage: An in-vitro, ex-vivo study. Dig. Liver Dis. 51: 47–54.
Money, J., Bobrow, N.A., and Clarke, F.C. (1971). Autism and autoimmune disease: a family study. J. Autism Child. Schizophr. 1: 146–160.

Journal of Food Bioactives | www.isnff-jfb.com 11
Lange et al. Gluten-free and casein-free diet in autism
Mulle, J.G., Sharp, W.G., and Cubells, J.F. (2013). The gut microbiome: a new frontier in autism research. Curr. Psychiatry Rep. 15: 337.
Navarro, F., Pearson, D.A., Fatheree, N., Mansour, R., Hashmi, S.S., and Rhoads, J.M. (2015). Are ‘leaky gut’ and behavior associated with gluten and dairy containing diet in children with autism spectrum disorders? Nutr. Neurosci. 18: 177–185.
Neumeyer, A.M., Gates, A., Ferrone, C., Lee, H., and Misra, M. (2013). Bone density in peripubertal boys with autism spectrum disorders. J. Autism Dev. Disord. 43: 1623–1629.
O’Banion, D., Armstrong, B., Cummings, R.A., and Stange, J. (1978). Dis-ruptive behavior: a dietary approach. J. Autism Child. Schizophr. 8: 325–337.
O’Mahony, S.M., Stilling, R.M., Dinan, T.G., and Cryan, J.F. (2015). The mi-crobiome and childhood diseases: Focus on brain-gut axis. Birth De-fects Res. C. 105: 296–313.
O’Reilly, C., Lewis, J.D., and Elsabbagh, M. (2017). Is functional brain con-nectivity atypical in autism? A systematic review of EEG and MEG studies. PLoS One. 12: e0175870.
Owen-Smith, A.A., Bent, S., Lynch, F.L., Coleman, K.J., Yau, V.M., Pearson, K.A., Massolo, M.L., Quinn, V., and Croen, L.A. (2015). Prevalence and predictors of complementary and alternative medicine use in a large insured sample of children with autism spectrum disorders. Res. Au-tism Spectr. Disord. 17: 40–51.
Panksepp, J. (1979). A neurochemical theory of autism. Trends Neurosci. 2: 174–177.
Pedersen, L., Parlar, S., Kvist, K., Whiteley, P., and Shattock, P. (2014). Data mining the ScanBrit study of a gluten- and casein-free dietary inter-vention for children with autism spectrum disorders: Behavioural and psychometric measures of dietary response. Nutr. Neurosci. 17: 207–213.
Pellissier, L.P., Gandía, J., Laboute, T., Becker, J., and Le Merrer, J. (2018). μ opioid receptor, social behaviour and autism spectrum disorder: Reward matters. Br. J. Pharmacol. 175: 2750–2769.
Pennesi, C.M., and Klein, L.C. (2012). Effectiveness of the gluten-free, ca-sein-free diet for children diagnosed with autism spectrum disorder: based on parental report. Nutr. Neurosci. 15: 85–91.
Perrin, J.M., Coury, D.L., Hyman, S.L., Cole, L., Reynolds, A.M., and Clem-ons, T. (2012). Complementary and alternative medicine use in a large pediatric autism sample. Pediatrics 130(Suppl. 2): S77–S82.
Piwowarczyk, A., Horvath, A., Łukasik, J., Pisula, E., and Szajewska, H. (2018). Gluten- and casein-free diet and autism spectrum disorders in children: a systematic review. Eur. J. Nutr. 57: 433–440.
Pontino, J.L., Schaal, K., and Chambliss, C. Effects of a gluten-free diet on rate of achievement in autistic children in an applied behavioural anaylsis program: summary analysis: Research Report: Ursinus Col-lege (ED413689).
Reichelt, K.L., Hole, K., Hamberger, A., Saelid, G., Edminson, P.D., Braes-trup, C.B., Lingjaerde, O., Ledaal, P., and Orbeck, H. (1981). Biologi-cally active peptide-containing fractions in schizophrenia and child-hood autism. Adv. Biochem. Psychopharmacol. 28: 627–643.
Reichelt, K.L., Saelid, G., Lindback, T., and Bøler, J.B. (1986). Childhood au-tism: a complex disorder. Biol. Psychiatry 21: 1279–1290.
Reichelt, K.L., Knivsberg, A.-M., Lind, G., and Nødland, M. (1991). Probable etiology and possible treatment of childhood autism. Brain Dysfunc-tion 4: 308–319.
Reichow, B., Barton, E.E., Boyd, B.A., and Hume, K. (2012). Early intensive behavioral intervention (EIBI) for young children with autism spec-trum disorders (ASD). Cochrane Database Syst. Rev. 10: CD009260.
Reichow, B., Barton, E.E., Boyd, B.A., and Hume, K. (2014). Early intensive behavioral intervention (EIBI) for young children with autism spec-trum disorders (ASD): A systematic review. Campbell Syst. Rev. 10: 1–116.
Reichow, B., Volkmar, F.R., and Cicchetti, D.V. (2008). Development of the evaluative method for evaluating and determining evidence-based practices in autism. J. Autism Dev. Disord. 38: 1311–1319.
Reissmann, A., Hauser, J., Stollberg, E., and Lange, K.W. (2020). Gluten-free and casein-free diets in the management of autism spectrum disor-der: A systematic literature review. Mov. Nutr. Health Dis. 4: 21–38.
Rosenfeld, C.S. (2015). Microbiome disturbances and autism spectrum disorders. Drug Metab. Dispos. 43: 1557–1571.
Rubenstein, E., Schieve, L., Bradley, C., DiGuiseppi, C., Moody, E., Thom-
as, K., and Daniels, J. (2018). The prevalence of gluten free diet use among preschool children with autism spectrum disorder. Autism Res. 11: 185–193.
Salomone, E., Charman, T., McConachie, H., and Warreyn, P. (2015). Prevalence and correlates of use of complementary and alternative medicine in children with autism spectrum disorder in Europe. Eur. J. Pediatr. 174: 1277–1285.
Sapone, A., Bai, J.C., Ciacci, C., Dolinsek, J., Green, P.H.R., Hadjivassiliou, M., Kaukinen, K., Rostami, K., Sanders, D.S., Schumann, M., Ullrich, R., Villalta, D., Volta, U., Catassi, C., and Fasano, A. (2012). Spectrum of gluten-related disorders: consensus on new nomenclature and classification. BMC Med. 10: 13.
Schreck, K.A., Russell, M., and Vargas, L.A. (2013). Autism treatments in print: media’s coverage of scientifically supported and alternative treatments. Behav. Intervent. 28: 299–321.
Schreibman, L., Dawson, G., Stahmer, A.C., Landa, R., Rogers, S.J., McGee, G.G., Kasari, C., Ingersoll, B., Kaiser, A.P., Bruinsma, Y., McNerney, E., Wetherby, A., and Halladay, A. (2015). Naturalistic developmental behavioral interventions: Empirically validated treatments for autism spectrum disorder. J. Autism Dev. Disord. 45: 2411–2428.
Seung, H., Rogalski, Y., Shankar, M., and Elder, J.H. (2007). The gluten-and casein-free diet and autism: Communication outcomes from a pre-liminary double-blind clinical trial. J. Med. Speech Lang. Pathol. 15: 337–345.
Shattock, P., and Whiteley, P. (2002). Biochemical aspects in autism spec-trum disorders: updating the opioid-excess theory and presenting new opportunities for biomedical intervention. Expert Opin. Ther. Targets. 6: 175–183.
Simonoff, E., Pickles, A., Charman, T., Chandler, S., Loucas, T., and Baird, G. (2008). Psychiatric disorders in children with autism spectrum disorders: prevalence, comorbidity, and associated factors in a pop-ulation-derived sample. J. Am. Acad. Child Adolesc. Psychiatry 47: 921–929.
Smith, T., and Antolovich, M. (2000). Parental perceptions of supplemental interventions received by young children with autism in intensive be-havior analytic treatment. Behav. Intervent. 15: 83–97.
Son, J.S., Zheng, L.J., Rowehl, L.M., Tian, X., Zhang, Y., Zhu, W., Litcher-Kelly, L., Gadow, K.D., Gathungu, G., Robertson, C.E., Ir, D., Frank, D.N., and Li, E. (2015). Comparison of fecal microbiota in children with autism spectrum disorders and neurotypical siblings in the Simons Simplex Collection. PLoS One 10: e0137725.
Souza, N.C., Mendonca, J.N., Portari, G.V., Jordao, A.A. Jr, Marchini, J.S., and Chiarello, P.G. (2012). Intestinal permeability and nutritional sta-tus in developmental disorders. Altern. Ther. Health Med. 18: 19–24.
Stojiljković, V., Todoroviċ, A., Radlović, N., Pejić, S., Mladenović, M., Kasapović, J., and Pajović, S.B. (2007). Antioxidant enzymes, glu-tathione and lipid peroxidation in peripheral blood of children af-fected by coeliac disease. Ann. Clin. Biochem. 44: 537–543.
Strati, F., Cavalieri, D., Albanese, D., Claudio De Felice, C., Donati, C., Hayek, J., Jousson, O., Leoncini, S., Renzi, D., Calabrò, A., and De Filippo, C. (2017). New evidences on the altered gut microbiota in autism spec-trum disorders. Microbiome 5: 24.
Tick, B., Bolton, P., Happé, F., Rutter, M., and Rijsdijk, F. (2016). Heritabil-ity of autism spectrum disorders: a meta-analysis of twin studies. J. Child Psychol. Psychiatry 57: 585–595.
Torrente, F., Ashwood, P., Day, R., Machado, N., Furlano, R.I., Anthony, A., Davies, S.E., Wakefield, A.J., Thomson, M.A., Walker-Smith, J.A., and Murch, S.H. (2002). Small intestinal enteropathy with epithelial IgG and complement deposition in children with regressive autism. Mol. Psychiatry 7: 375–382.
Trudeau, M.S., Madden, R.F., Parnell, J.A., Gibbard, W.B., and Shearer, J. (2019). Dietary and supplement-based complementary and alterna-tive medicine use in pediatric autism spectrum disorder. Nutrients 11: 1783.
Van de Sande, M.M., van Buul, V.J., and Brouns, F.J. (2014). Autism and nutrition: the role of the gut-brain axis. Nutr. Res. Rev. 27: 199–214.
Vojdani, A., Pangborn, J.B., Vojdani, E., and Cooper, E.L. (2003). Infections, toxic chemicals and dietary peptides binding to lymphocyte recep-tors and tissue enzymes are major instigators of autoimmunity in autism. Int. J. Immunopathol. Pharmacol. 16: 189–199.
Vojdani, A., Bazargan, M., Vojdani, E., Samadi, J., Nourian, A.A., Eghbalieh,

Journal of Food Bioactives | www.isnff-jfb.com12
Gluten-free and casein-free diet in autism Lange et al.
N., and Cooper, E.L. (2004a). Heat shock protein and gliadin peptide promote development of peptidase antibodies in children with au-tism and patients with autoimmune disease. Clin. Diagn. Lab. Immu-nol. 11: 515–524.
Vojdani, A., O’Bryan, T., Green, J.A., Mccandless, J., Woeller, K.N., Vojdani, E., Nourian, A.A., and Cooper, E.L. (2004b). Immune response to die-tary proteins, gliadin and cerebellar peptides in children with autism. Nutr. Neurosci. 7: 151–161.
Vuong, H.E., and Hsiao, E.Y. (2017). Emerging roles for the gut microbiome in autism spectrum disorder. Biol. Psychiatry 81: 411–423.
Walsh, P., Elsabbagh, M., Bolton, P., and Singh, I. (2011). In search of bio-markers for autism: scientific, social and ethical challenges. Nat. Rev. Neurosci. 12: 603–612.
Wang, L., Angley, M.T., Gerber, J.P., Young, R.L., Abarno, D.V., McKinnon, R.A., and Sorich, M.J. (2009). Is urinary indolyl-3-acryloylglycine a biomarker for autism with gastrointestinal symptoms? Biomarkers 14: 596–603.
Wang, L., Christophersen, C.T., Sorich, M.J., Gerber, J.P., Angley, M.T., and Conlon, M.A. (2013). Increased abundance of Sutterella spp. and Ru-minococcus torques in feces of children with autism spectrum disor-der. Mol. Autism. 4: 42.
Weitlauf, A.S., McPheeters, M.L., Peters, B., Sathe, N., Travis, R., Aiello, R., Williamson, E., Veenstra-VanderWeele, J., Krishnaswami, S., Jerome,
R., and Warren, Z. (2014). Therapies for children with autism spec-trum disorder: Behavioral interventions update. AHRQ Compara-tive Effectiveness Reviews 137. Agency for Healthcare Research and Quality, Rockville (MD).
Whiteley, P. (2015). Nutritional management of (some) autism: a case for gluten- and casein-free diets? Proc. Nutr. Soc. 74: 202–207.
Whiteley, P., Haracopos, D., Knivsberg, A.-M., Reichelt, K.L., Parlar, S., Jacobsen, J., Seim, A., Pedersen, L., Schondel, M., and Shattock, P. (2010). The ScanBrit randomised, controlled, single-blind study of a gluten- and casein-free dietary intervention for children with autism spectrum disorders. Nutr. Neurosci. 13: 87–100.
Whiteley, P., Rodgers, J., Savery, D., and Shattock, P. (1999). A gluten-free diet as an intervention for autism and associated spectrum disor-ders: Preliminary findings. Autism 3: 45–65.
Winburn, E., Charlton, J., McConachie, H., McColl, E., Parr, J., O’Hare, A., Baird, G., Gringras, P., Wilson, D.C., Adamson, A., Adams, S., and Le Couteur, A. (2014). Parents’ and child health professionals’ attitudes towards dietary interventions for children with autism spectrum dis-orders. J. Autism Dev. Disord. 44: 747–757.
Wrońska-Nofer, T., Nofer, J.R., Jajte, J., Dziubałtowska, E., Szymczak, W., Krajewski, W., Wasowicz, W., and Rydzyński, K. (2012). Oxidative DNA damage and oxidative stress in subjects occupationally exposed to nitrous oxide (N2O). Mutat. Res. 731: 58–63.

Copyright: © 2021 International Society for Nutraceuticals and Functional Foods.All rights reserved.
13
Perspective J. Food Bioact. 2021;15:13–20
Journal ofFood Bioactives International Society for
Nutraceuticals and Functional Foods
The use of vitamin D in depression
Klaus W. Lange*, Katharina M. Lange, Yukiko Nakamura and Andreas Reissmann
Department of Experimental Psychology, University of Regensburg, Regensburg, Germany*Corresponding author: Klaus W. Lange, Institute of Psychology, University of Regensburg, 93040 Regensburg, Germany. Tel: +49 941 9433815; Fax: +49 941 9434496; E-mail: [email protected]: 10.31665/JFB.2021.15278Received: September 22, 2021; Revised received & accepted: September 29, 2021Citation: Lange, K.W., Lange, K.M., Nakamura, Y., and Reissmann, A. (2021). The use of vitamin D in depression. J. Food Bioact. 15: 13–20.
Abstract
Vitamin D has increasingly been associated with the pathophysiology of mental illness and has been suggested to have beneficial effects on depression in adults. Epidemiological studies concerning vitamin D and depression have found inconsistent results and many have significant methodological limitations. The available evidence suggests that depressed individuals show reduced vitamin D concentrations compared to controls without depression. De-spite the available findings suggesting that hypovitaminosis D elevates the risk of depressive mood, the evidence of observational and interventional studies is insufficient to establish causality between low vitamin D levels and the occurrence of depression. The question of whether vitamin D sufficiency has protective efficacy against inci-dent depression or recurrence requires future investigation. In order to examine the therapeutic efficacy of vita-min D, further well-designed, large-scale, long-term intervention trials of vitamin D supplementation in people of different age groups with depressive symptoms, diagnosed depression, postpartum depression or other depres-sive disorders are warranted. In short, current evidence cannot definitively establish whether vitamin D deficiency is a risk factor in the development of depression or whether vitamin D is effective in the treatment of depression.
Keywords: Vitamin D; Depression; Mood; Epidemiology; Treatment.
1. Introduction
Depression is globally a highly prevalent mental disorder that can occur across the entire lifespan (Kessler et al., 2007). The 12-month prevalence of major depressive disorder has been estimated to be approximately 6% (Kessler and Bromet, 2013), with a lifetime risk of 15–18% (Bromet et al., 2011). Major depressive disorder has been predicted to become the leading global cause of disease burden by 2030 (Kessler et al., 2003; World Health Organization, 2008). Major depressive disorder is characterized by depressed mood, anhedonia, reduced self-esteem and fatigue (American Psychiatric Association, 2013) and can cause significant func-tional disability and severely limit the psychosocial functioning and quality of life of those affected (Malhi and Mann, 2018). Both depressive disorders and subthreshold depressive symptoms have been found to be associated with considerable disability, health care costs and mortality (Broadhead et al., 1990; Pietrzak et al.,
2013). While a biopsychosocial model of the etiology of depres-sion has been proposed, with biological, psychological, social and environmental factors contributing to the development of depres-sion, the pathophysiological mechanisms underlying depression remain largely unknown (Krishnan and Nestler, 2010).
The treatment of major depressive disorder, with its high rate of recurrence, remains difficult and less than satisfactory (Malhi and Mann, 2018). While pharmacotherapy and/or cognitive be-havior psychotherapy have been demonstrated to be efficacious in the treatment of depression (Cuijpers et al., 2008; Cuijpers et al., 2009), some degree of non-response to antidepressant medications may occur in up to 60% of individuals with major depressive dis-order (Bschor and Baethge, 2010; Fava, 2003). Almost a third of people treated for depression remain refractory to multistep antide-pressant therapies (Rush et al., 2006a; Rush et al., 2006b), and the long-term effectiveness of pharmacotherapy and psychotherapy remains a matter of debate (Casacalenda et al., 2002). Moreover,

Journal of Food Bioactives | www.isnff-jfb.com14
Vitamin D and depression Lange et al.
poor acceptability of treatment (Jorm, 2000) and adverse effects of antidepressant drugs (Anderson et al., 2012; Goethe et al., 2007) may result in discontinuation of treatment. The need for the devel-opment of further medications and novel approaches to the preven-tion and treatment of depression has therefore been emphasized (Insel, 2015; Miller, 2010).
Diet and nutrition have been found not only to be critically in-volved in the promotion of metabolic health (Bennett et al., 2015), but also to play a role in mental health and to be capable of modi-fying mental disorders (Lange, 2018; Lange, 2020a; Marx et al., 2017). The risk of depression appears to be inversely associated with the Mediterranean diet (Lai et al., 2014; Opie et al., 2015; Psaltopoulou et al., 2013), while Western diets have been report-ed to be positively correlated with depression (Lai et al., 2014). Various food bioactives, such as omega-3 polyunsaturated fatty acids (Lange, 2020b; Lange, 2021; Lange et al., 2021a), probiot-ics (Lange et al., 2020) and L-theanine and epigallocatechine gal-late (Lange et al., 2021b) have shown associations with depression risk. Emerging evidence suggests that vitamin D deficiency is also linked to depression, with people showing normal vitamin D levels having a reduced risk of developing depression (e.g. Anglin et al., 2013; Black et al., 2014; Brouwer-Brolsma et al., 2015; Hoogendi-jk et al., 2008; Kerr et al., 2015; von Känel et al., 2015). These findings have led to investigations of food bioactive and micronu-trient supplementation in depression (Lim et al., 2016).
The present perspective critically discusses the evidence re-garding the therapeutic efficacy of vitamin D in depression. A lit-erature search, including publications up until March 2021, was conducted using PubMed and Google Scholar. Search results were screened for relevant articles involving humans and evaluations of vitamin D interventions.
2. Vitamin D and the brain and the rationale for its use in de-pression
Vitamin D comprises a group of structurally related, fat-soluble secosteroids. It is produced in the skin during sun exposure and can also be obtained from dietary intake, including vitamin D sup-plementation (Holick, 2007). The two main forms of vitamin D relevant for humans are vitamin D2 (ergocalciferol) and vitamin D3 (cholecalciferol) (Feldman et al., 2011). Both forms are impor-tant for calcium and phosphorus homeostasis (Holick, 2007) and consequently for the maintenance of cellular and neuromuscular function as well as the regulation of bone metabolism (Brown et al., 1999). Vitamin D obtained from dietary sources stems mainly from fungi and yeasts (vitamin D2) or fatty fish (vitamin D3) (Hos-sein-Nezhad et al., 2013; Jäpelt and Jakobsen, 2013). However, it is difficult to obtain sufficient vitamin D from dietary intake (Max-well, 1994), and the majority of vitamin D3 is produced in the skin from 7-dehydrocholesterol (provitamin D3) following expo-sure to ultraviolet B radiation (Holick, 2017). Two hydroxylation steps are needed to form either 1,25-dihydroxyvitamin D2 (ergo-calcitriol, 1,25(OH)2D2) or 1,25-dihydroxyvitamin D3 (calcitriol, 1,25(OH)2D3), which are the active metabolic products of vitamin D2 and D3, respectively (Vieth, 2004). Vitamin D is converted in the liver to 25-hydroxyvitamin D (25(OH)D), the level of which is commonly used to determine a person’s vitamin D status (Hollis, 1996), and subsequently in the kidneys to its active form 1,25-di-hydroxyvitamin D (1,25(OH)2D) (Wacker and Holick, 2013). Both 1,25(OH)2D2 and 1,25(OH)2D3 are pleiotropic secosteroid hormones regulating the expression of a large number of genes, responsible for cellular proliferation, differentiation, DNA repair
and apoptosis (Hossein-Nezhad et al., 2013), through binding to the vitamin D receptor (Kongsbak et al., 2013; Pike et al., 2010).
Experimental studies have advanced the understanding of the multiple actions of vitamin D in the central nervous system (Kalu-eff and Tuohimaa, 2007). Vitamin D receptors have been found in brain regions involved in the regulation of behavior, including the cerebral cortex, limbic system and cerebellum, and may also stimulate the release of neurotrophins, which are involved in neu-ronal development (Kalueff and Tuohimaa, 2007). Vitamin D has been found to play a critical role in brain development (Eyles et al., 2013; Groves et al., 2014; Ko et al., 2004). A deficiency of the vita-min appears to be a risk factor for cognitive decline and dementia in elderly people (Annweiler et al., 2015). Less is known concern-ing the role of vitamin D in the regulation of affect and mood. Hy-povitaminosis D has been hypothesized to induce negative mood and depression (Berk et al., 2007). Since vitamin D receptors are present on neurons and glial cells in brain areas thought to be in-volved in the pathophysiology of depression, e.g. cingulate cortex and hippocampus (Eyles et al., 2005), and an interaction between vitamin D and glucocorticoids has been found in the hippocampus (Obradovic et al., 2006), the potential role of vitamin D as a thera-peutic agent in depression has been investigated. Vitamin D ap-pears to be involved in the control of serotonin formation, imply-ing a link between vitamin D deficiency and depression (Patrick and Ames, 2015). Vitamin D has been theorized to act as a neu-roactive hormone (Berridge, 2015a; Berridge, 2015b; Berridge, 2017). Furthermore, vitamin D has been postulated to modulate neuronal calcium ion levels and thereby to cause the onset of de-pressive symptoms, with vitamin D deficiency leading to elevated neuronal Ca2+ and increasing depression (Berridge, 2017). In addi-tion, vitamin D may be involved in neuro-immunomodulation and neuroplasticity, both of which have been proposed as mechanisms underlying its effects on mood (Fernandes de Abreu et al., 2009).
Vitamin D deficiency (defined as serum 25(OH)D concentra-tion of less than 50 nmol/L) or insufficiency are common in peo-ple of all age groups, with approximately one billion individuals affected worldwide (Holick, 2007). Several observational studies, including case-control studies, cross-sectional studies and cohort studies, have investigated the relationship between vitamin D and depression. Various epidemiological studies have shown associa-tions between serum 25(OH)D levels and depression, with vitamin D deficiency being associated with depression (Black et al., 2014; Brouwer-Brolsma et al., 2015; Ganji et al., 2010; Hoang et al., 2011; Hoogendijk et al., 2008; Jamilian et al., 2013; Kerr et al., 2015; Kjærgaard et al., 2011; Lee et al., 2011; Milaneschi et al., 2010; Parker et al., 2017; Tolppanen et al., 2012; von Känel et al., 2015). However, other studies have found no association (e.g. Nanri et al., 2009; Pan et al., 2009; Zhao et al., 2010).
A systematic review and meta-analysis summarizing the avail-able evidence from cross-sectional and prospective cohort studies reported a pooled estimate of the adjusted odds ratio of depression in 11 cross-sectional studies (with a total of 43,137 participants) of 0.96 (95% CI = 0.94–0.99, I2 = 63%) for a 10 ng/ml increase in 25(OH)D level (Ju et al., 2013). For five cohort studies compris-ing 12,648 primarily elderly participants, with 2,663 experiencing depression events during follow-up, the pooled adjusted odds ratio of depression was 0.92 (95% CI = 0.87–0.98, I2 = 50%) for a 10 ng/ml increase in 25(OH)D levels (Ju et al., 2013). A further sys-tematic review and meta-analysis pooling the results of previous epidemiological studies reported a positive association between hypovitaminosis D and depression in adults (odds ratio = 1.31; 95% CI, 1.00–1.71) (Anglin et al., 2013). The link between vita-min D concentrations and depression has been suggested to be less straightforward than commonly assumed. The association may

Journal of Food Bioactives | www.isnff-jfb.com 15
Lange et al. Vitamin D and depression
have been overestimated, particularly in older people, since both hypovitaminosis D and depression are highly prevalent in this age group (Rastmanesh et al., 2012) (see also Table 1).
Vitamin D has also been implicated in antenatal depression and postpartum depression. However, this association is complex and the available findings are inconsistent. In a systematic review, a significant association between vitamin D status and both ante-natal and postpartum depression was reported (Aghajafari et al., 2018). The findings of a meta-analysis of nine longitudinal stud-ies suggested that vitamin D deficiency could be a modifiable risk factor of postpartum depression but not of depressive symptoms during pregnancy (Wang et al., 2018).
The correlation between low vitamin D levels and depressive symptoms does not necessarily indicate that hypovitaminosis D plays a causal role in depression, since a wide range of potential confounders, such as age, dietary intake, physical exercise, time spent outdoors, as well as alcohol and tobacco may play a role (Bertone-Johnson, 2009). It is conceivable that low vitamin D lev-els result from depression, since depressed mood and apathy may lead to a decrease in exposure to sunlight or intake of food rich in vitamin D (Rastmanesh et al., 2012). A decrease in vitamin D concentrations may therefore be an antecedent or a consequence of depression, and causality should be examined in prospective stud-ies assessing mood changes over time. A six-year prospective study comprising 954 elderly individuals aged 65 years or older investi-gated the association of baseline vitamin D levels and subsequent
occurrence of depression (Milaneschi et al., 2010). In this study, people with low baseline concentrations of 25(OH)D3 (<50 nM or <20 ng/ml) were reported to show significantly increased depres-sion scores at three-year and six-year follow-up in comparison with those with high baseline levels (Milaneschi et al., 2010). However, this difference may also be explained by different lifestyles.
3. Intervention studies using vitamin D in depression
Intervention studies administering vitamin D have been conducted in people with major depression and seasonal affective disorder as well as women with postpartum depression and elderly individuals with depressive symptoms (see Table 2). A systematic review of six randomized controlled trials (with 1,203 participants including 71 depressed individuals) investigated the efficacy of oral vitamin D administration versus placebo in adults at risk of depression, with symptoms of depression or with a primary diagnosis of de-pression (Li et al., 2014). No significant effects of vitamin D sup-plementation on depression scores or in subgroup or sensitivity analyses were found; the quality of evidence was considered low (Li et al., 2014). A further systematic review and meta-analysis of randomized controlled trials summarized the effects of vitamin D supplementation on depressive symptoms or depression (Shaf-fer et al., 2014). No overall effects of vitamin D supplementation
Table 1. Observational evidence of a relationship between vitamin D and depression
Associations between hypovitaminosis D and depression in adults (Anglin et al., 2013; Ju et al., 2013)
Reduced risk of developing depression in people with higher serum 25(OH)D levels (Jääskeläinen et al., 2015)
Association of depression with vitamin D deficiency in older adults (Okereke and Singh, 2016)
Association of low baseline levels of 25(OH)D3 with increased depression scores at 3-year and 6-year follow-up in elderly adults (Milaneschi et al., 2010)
Association of depression with vitamin D deficiency in young people (Kerr et al., 2015; Polak et al., 2014)
Association of depression with vitamin D deficiency in individuals with heart failure and cancer (Björkhem-Bergman and Bergman, 2016; Johansson et al., 2016)
Association between vitamin D status and antenatal and postpartum depression (Aghajafari et al., 2018)
Caveat: Correlation between low vitamin D levels and depressive symptoms is not necessarily indicative of a causal role of hypovitaminosis D in depression due to potential confounders (age, dietary intake, physical exercise, time spent outdoors etc.)
Table 2. Interventional evidence of a relationship between vitamin D and depression
No significant meta-analytical effect on depression following vitamin D supplementation (Li et al., 2014; Lázaro Tomé et al., 2021)
No overall meta-analytical effect of vitamin D supplementation on depressive symptoms, but moderate, statistically significant effect in people with clinically significant depressive symptoms or depressive disorder (Shaffer et al., 2014)
No significant improvement of depression following vitamin D supplementation in a meta-analysis of studies with individuals with low depression levels and sufficient vitamin D levels at baseline (Gowda et al., 2015)
Significant worsening of depression following vitamin D supplementation in a meta-analysis of “biologically flawed” studies (e.g. vitamin D deficiency not established at baseline and/or vitamin D dose administered insufficient to produce normal vitamin D levels) (Spedding, 2014)
Significant improvement of depression following vitamin D supplementation in a meta-analysis of studies without biological flaws (Spedding, 2014)
Favorable findings following vitamin D supplementation (≥800 I.U. daily) in studies demonstrating a change in vitamin levels (Spedding, 2014)
Dose dependency of effects of importance of vitamin D supplementation (Mozaffari-Khosravi et al., 2013)
Summary: Insufficient evidence in support of therapeutic efficacy of vitamin D supplementation in depression

Journal of Food Bioactives | www.isnff-jfb.com16
Vitamin D and depression Lange et al.
on depressive symptoms were found. However, subgroup analysis showed that vitamin D supplementation had a moderate, statisti-cally significant effect in individuals with clinically significant de-pressive symptoms or depressive disorder and a small, non-signif-icant effect in those without clinically significant depression. The trials included in this review varied in respect of dose, frequency, duration and mode of delivery of vitamin D administration (Shaf-fer et al., 2014). A meta-analysis of nine randomized controlled trials (with a total of 4,923 participants) estimated the weighted mean effect of vitamin D supplementation in decreasing depres-sive symptoms in adult participants diagnosed with depression or depressive symptoms (Gowda et al., 2015). No significant reduc-tion in depression was observed following vitamin D supplementa-tion. However, most of the studies had included people with low depression levels and sufficient concentrations of serum vitamin D at baseline. The vitamin D doses used and intervention durations varied between the studies included in the meta-analytical review (Gowda et al., 2015). A recent systematic review and meta-analy-sis including 10 randomized controlled trials (with a total of 1,393 participants) found no significant effect of oral vitamin D adminis-tration on post-intervention depression scores (Lázaro Tomé et al., 2021). The vitamin D dose administered appears to be important. In a randomized controlled trial conducted in depressed people with initial hypovitaminosis D, a single intramuscular injection of 300,000 I.U. of vitamin D improved symptoms of depression three months following intervention more than a dose of 150,000 I.U. (Mozaffari-Khosravi et al., 2013).
“Biological flaws” in primary studies have been suggested to be a reason why meta-analyses have failed to show efficacy of vita-min D supplementation in depression (Lappe and Heaney, 2012). The concept of biological flaws refers to limitations in study design, which may affect a study’s ability to test the research hypothesis. For example, the hypothesis that the treatment of a vitamin D defi-cit improves depressive symptoms can be tested only if vitamin D deficiency is established at baseline and the vitamin D dose admin-istered is capable of producing sufficient levels of vitamin D dur-ing the trial (Heaney, 2012). Therefore, meta-analytical studies have been conducted according to the presence or absence of biological flaws (Spedding, 2014). While a meta-analysis of the available stud-ies without flaws showed a statistically significant improvement in depression following vitamin D supplementation, the results of stud-ies with biological flaws were mainly inconclusive, with the meta-analysis performed showing a statistically significant worsening in depression following vitamin D administration (Spedding, 2014). Vitamin D supplementation of ≥ 800 I.U. daily demonstrated favora-ble results in the therapy of depression in the studies with a change in vitamin levels, with the effect size being comparable to that of antidepressant medication (Spedding, 2014).
A recent 28-day trial investigating vitamin D3 supplementation in addition to usual treatment in children and adolescents with vi-tamin D deficiency (25(OH)D ≤ 30 nmol/l) and at least mild de-
pression (according to Beck Depression Inventory) failed to show a vitamin D effect on self-rated depression (Libuda et al., 2020).
In summary, the current evidence is insufficient to support the efficacy of vitamin D supplementation in the treatment of depres-sion. Further large-scale randomized controlled trials assessing the efficacy of vitamin D supplementation are required. In particular, future trials should be conducted in people who are both depressed and deficient in vitamin D, with the vitamin D doses supplemented being large enough to achieve sufficient vitamin levels.
4. Future directions
The role of vitamin D in the etiology and treatment of depres-sion is still unclear (see Table 3). Due to various methodological problems and questionable scientific rigor of the available stud-ies, there is no unequivocal evidence of an association between vitamin D status and depressive symptoms or the diagnosis of de-pression. In particular, the relationship reported in several studies between hypovitaminosis D and mood could have been influenced by confounding variables. The association between vitamin D and depression may be indirectly explained by other factors af-fecting both vitamin status and mood. For example, exposure to sunlight influences not only vitamin D synthesis in the skin but also body circadian rhythms (e.g. melatonin), which have been shown to be involved in depression (Boyce and Barriball, 2010). Another covariable possibly playing a role is body mass index, with fat-soluble vitamin D being sequestered in adipose tissue and hypovitaminosis D arising from both high body mass index and depression (Forsythe et al., 2012). Furthermore, physical exercise levels may affect both vitamin D status and mood, since reduced physical activity has been observed in both individuals with de-pression and people with low levels of vitamin D (Annweiler et al., 2011). Recent investigations have attempted to take relevant covariables into account, with prospective longitudinal studies reporting an elevated hazard ratio for depression in people with hypovitaminosis D at baseline (see Anglin et al., 2013). While this suggests that hypovitaminosis D precedes the onset of depression, observational studies are unable to establish causality. In addition, the link between vitamin D and depression could be related to ge-netic predisposition, since gene locus 11p15 harbors susceptibility genes for mood disorders (Huang et al., 2010) and is also involved in vitamin D metabolism (Wang et al., 2010). Experimental and randomized controlled intervention trials are required to shed fur-ther light on the link between vitamin D status and depression.
Major problems of the available treatment studies examining the efficacy of vitamin D in depression include non-representative samples, problems with the generalization of findings, heteroge-neity of the trial participants regarding the severity of depressive symptoms, the methods used to assess depression, the presence of
Table 3. Problems and future directions regarding vitamin D and depression
Unclear causality of correlations between vitamin D status and depressive symptoms
Intervention trials needed in people who are both depressed and deficient in vitamin D
Assessment of depression level and vitamin D status both at trial baseline and follow-up
Investigations needed regarding vitamin D effects in different types of depressive disorders (major depressive disorder, seasonal affective disorder, bipolar disorder, antenatal and postpartum depression, premenstrual dysphoric disorder)
Optimal dosage, duration and mode of delivery of vitamin D supplementation
Summary: Large-scale, randomized controlled trials of vitamin D in people with well-defined diagnoses of depression needed

Journal of Food Bioactives | www.isnff-jfb.com 17
Lange et al. Vitamin D and depression
comorbidities, the dose and duration of supplementation and the levels of 25(OH)D at baseline and follow-up. Polymorphisms of the vitamin D receptor gene modulate the effectiveness of vitamin D in the central nervous system, and this may explain why some individuals with depression respond to vitamin D and others do not (Kuningas et al., 2009).
Future intervention studies need to distinguish between differ-ent types of depressive disorders, such as major depressive dis-order, seasonal affective disorder, bipolar disorder, postpartum depression and premenstrual dysphoric disorder. Intervention trials with biological flaws may demonstrate the methodological limitations of the study design rather than the efficacy of vitamin D supplementation. Therefore, future studies need to include indi-viduals who are both depressed and vitamin D deficient and should examine depression level and vitamin D status both at baseline and follow-up. Studies employing sufficient doses of vitamin D and intervention periods capable of significantly increasing vitamin D levels in well-defined, homogenous risk groups are warranted.
Given a variety of adverse effects of hypovitaminosis D and the general health benefits and low risk of serious adverse effects of vitamin D supplements (Annweiler et al., 2011; Jorde et al., 2013), vitamin D supplementation may be recommended to pro-tect against depression. In particular, sensible and moderate sun exposure, avoiding even mild sunburn (Kennedy et al., 2003), of arms, legs, abdomen and back can help supplement vitamin D and induce the production of beta-endorphin in the skin, which may also improve wellbeing (Zanello et al., 1999). The combination of sunlight exposure and physical exercise may be particularly ben-eficial (Lange, 2018). Since the synthesis of vitamin D in the skin following sunlight exposure is strongly influenced by various fac-tors, such as age, skin pigmentation, season and latitude, dietary supplementation may be needed to obtain beneficial effects. For the treatment of vitamin deficiency, the administration of 50,000 I.U. of vitamins D2 or D3 once a week for 8 weeks has been rec-ommended in order to raise blood concentrations of 25(OH)D to an adequate range of at least 30 ng/ml or preferably 40–60 ng/ml, while a dose of 50,000 I.U. vitamin D2 or vitamin D3 once every other week has been suggested to be effective in maintaining vita-min D sufficiency (Holick et al., 2011).
5. Conclusion
Despite the available findings suggesting that hypovitaminosis D elevates the risk of depressive mood, the evidence of observa-tional and interventional studies is insufficient to establish causal-ity between low vitamin D levels and the occurrence of depression. Whether or not vitamin D sufficiency has protective efficacy against incident depression or recurrence requires future investigation. Further well-designed, large-scale, long-term intervention trials of vitamin D supplementation in people of different age groups with depressive symptoms, diagnosed depression, postpartum depression or other depressive disorders would be of value in examining the therapeutic efficacy of vitamin D. Given the general health benefits and low risk of adverse effects of vitamin D, modest sun exposure and dietary vitamin D enrichment may be recommended in order to supplement vitamin D and improve mood and wellbeing. However, this recommendation requires confirmation by future studies.
References
Aghajafari, F., Letourneau, N., Mahinpey, N., Cosic, N., and Giesbrecht, G.
(2018). Vitamin D deficiency and antenatal and postpartum depres-sion: a systematic review. Nutrients. 10: 478.
American Psychiatric Association. (2013). Diagnostic and Statistical Man-ual of Mental Disorders, 5th ed. American Psychiatric Association, Washington, DC.
Anderson, H.D., Pace, W.D., Libby, A.M., West, D.R., and Valuck, R.J. (2012). Rates of 5 common antidepressant side effects among new adult and adolescent cases of depression: a retrospective US claims study. Clin. Ther. 34: 113–123.
Anglin, R.E.S., Samaan, Z., Walter, S.D., and McDonald, S.D. (2013). Vita-min D deficiency and depression in adults: Systematic review and meta-analysis. Br. J. Psychiatry. 202: 100–107.
Annweiler, C., Dursun, E., Féron, F., Gezen-Ak, D., Kalueff, A.V., Littlejohns, T., Llewellyn, D.J., Millet, P., Scott, T., Tucker, K.L., Yilmazer, S., and Beauchet, O. (2015). ‘Vitamin D and cognition in older adults’: up-dated international recommendations. J. Intern. Med. 277: 45–57.
Annweiler, C., Souberbielle, J.-C., Schott, A.-M., de Decker, L., Berrut, G., and Beauchet, O. (2011). Vitamine D chez la personne âgée les 5 points à retenir. Geriatr. Psychol. Neuropsychiatr. Vieil. 9: 259–267.
Bennett, B.J., Hall, K.D., Hu, F.B., McCartney, A.L., and Roberto, C. (2015). Nutrition and the science of disease prevention: a systems approach to support metabolic health. Ann. N. Y. Acad. Sci. 1352: 1–12.
Berk, M., Sanders, K.M., Pasco, J.A., Jacka, F.N., Williams, L.J., Hayles, A.L., and Dodd, S. (2007). Vitamin D deficiency may play a role in depres-sion. Med. Hypotheses. 69: 1316–1319.
Berridge, M.J. (2015a). Vitamin D cell signalling in health and disease. Bio-chem. Biophys. Res. Commun. 460: 53–71.
Berridge, M.J. (2015b). Vitamin D: a custodian of cell signalling stability in health and disease. Biochem. Soc. Trans. 43: 349–358.
Berridge, M.J. (2017). Vitamin D and depression: cellular and regulatory mechanisms. Pharmacol. Rev. 69: 80–92.
Bertone-Johnson, E.R. (2009). Vitamin D and the occurrence of depres-sion: causal association or circumstantial evidence? Nutr. Rev. 67: 481–492.
Björkhem-Bergman, L., and Bergman, P. (2016). Vitamin D and patients with palliative cancer. BMJ Support. Palliat. Care. 6: 287–291.
Black, L.J., Jacoby, P., Allen, K.L., Trapp, G.S., Hart, P.H., Byrne, S.M., Mori, T.A., Beilin, L.J., and Oddy, W.H. (2014). Low vitamin D levels are as-sociated with symptoms of depression in young adult males. Aust. N. Z. J. Psychiatry. 48: 464–471.
Boyce, P., and Barriball, E. (2010). Circadian rhythms and depression. Aust. Fam. Physician. 39: 307–310.
Broadhead, W.E., Blazer, D.G., George, L.K., and Tse, C.K. (1990). Depres-sion, disability days, and days lost from work in a prospective epide-miologic survey. JAMA 264: 2524–2528.
Bromet, E., Andrade, L.H., Hwang, I., Sampson, N.A., Alonso, J., de Gi-rolamo, G., de Graaf, R., Demyttenaere, K., Hu, C., Iwata, N., Karam, A.N., Kaur, J., Kostyuchenko, S., Lépine, J.P., Levinson, D., Matsch-inger, H., Mora, M.E., Browne, M.O., Posada-Villa, J., Viana, M.C., Williams, D.R., and Kessler, R.C. (2011). Cross-national epidemiology of DSM-IV major depressive episode. BMC Med. 9: 90.
Brouwer-Brolsma, E.M., Dhonukshe-Rutten, R.A.M., van Wijngaarden, J.P., van der Zwaluw, N.L., Sohl, E., In’t Veld, P.H., van Dijk, S.C., Swart, K.M.A., Enneman, A.W., Ham, A.C., van Schoor, N.M., van der Vel-de, N., Uitterlinden, A.G., Lips, P., Feskens, E.J.M., and de Groot, L.C.P.G.M. (2015). Low vitamin D status is associated with more de-pressive symptoms in Dutch older adults. Eur. J. Nutr. 55(4): 1525–1534.
Brown, A.J., Dusso, A., and Slatopolsky, E. (1999). Vitamin D. Am. J. Physiol. 277: F157–F175.
Bschor, T., and Baethge, C. (2010). No evidence for switching the antide-pressant: systematic review and meta-analysis of RCTs of a common therapeutic strategy. Acta Psychiatr. Scand. 121: 174–179.
Casacalenda, N., Perry, J.C., and Looper, K. (2002). Remission in major de-pressive disorder: a comparison of pharmacotherapy, psychothera-py, and control conditions. Am. J. Psychiatry. 159: 1354–1360.
Cuijpers, P., Dekker, J., Hollon, S.D., and Andersson, G. (2009). Adding psy-chotherapy to pharmacotherapy in the treatment of depressive dis-orders in adults: a meta-analysis. J. Clin. Psychiatry. 70: 1219–1229.
Cuijpers, P., van Straten, A., van Oppen, P., and Andersson, G. (2008). Are psychological and pharmacologic interventions equally effective in

Journal of Food Bioactives | www.isnff-jfb.com18
Vitamin D and depression Lange et al.
the treatment of adult depressive disorders? A meta-analysis of com-parative studies. J. Clin. Psychiatry. 69: 1675–85.
Eyles, D.W., Burne, T.H.J., and McGrath, J.J. (2013). Vitamin D, effects on brain development, adult brain function and the links between low levels of vitamin D and neuropsychiatric disease. Front. Neuroendo-crinol. 34: 47–64.
Eyles, D.W., Smith, S., Kinobe, R., Hewison, M., and McGrath, J.J. (2005). Distribution of the vitamin D receptor and 1 alpha-hydroxylase in hu-man brain. J. Chem. Neuroanat. 29: 21–30.
Fava, M. (2003). Diagnosis and definition of treatment-resistant depres-sion. Biol. Psychiatry. 53: 649–659.
Feldman, D., Pike, J.W., and Adams, J.S. (Ed.). (2011). In: Vitamin D. 3rd ed. Elsevier, New York.
Fernandes de Abreu, D.A., Eyles, D., and Féron, F. (2009). Vitamin D, a neuro-immunomodulator: implications for neurodegenerative and autoimmune diseases. Psychoneuroendocrinology. 34(Suppl 1): S265–S277.
Forsythe, L.K., Livingstone, M.B.E., Barnes, M.S., Horigan, G., McSorley, E.M., Bonham, M.P., Magee, P.J., Hill, T.R., Lucey, A.J., Cashman, K.D., Kiely, M., Strain, J.J., and Wallace, J.M.W. (2012). Effect of adiposity on vitamin D status and the 25-hydroxycholecalciferol response to supplementation in healthy young and older Irish adults. Br. J. Nutr. 107: 126–134.
Ganji, V., Milone, C., Cody, M.M., McCarty, F., and Wang, Y.T. (2010). Serum vitamin D concentrations are related to depression in young adult US population: the Third National Health and Nutrition Examination Survey. Int. Arch. Med. 3: 29.
Goethe, J.W., Woolley, S.B., Cardoni, A.A., Woznicki, B.A., and Piez, D.A. (2007). Selective serotonin reuptake inhibitor discontinuation: side effects and other factors that influence medication adherence. J. Clin. Psychopharmacol. 27: 451–458.
Gowda, U., Mutowo, M.P., Smith, B.J., Wluka, A.E., and Renzaho, A.M.N. (2015). Vitamin D supplementation to reduce depression in adults: meta-analysis of randomized controlled trials. Nutrition. 31: 421–429.
Groves, N.J., McGrath, J.J., and Burne, T.H.J. (2014). Vitamin D as a neu-rosteroid affecting the developing and adult brain. Annu. Rev. Nutr. 34: 117–141.
Heaney, R.P. (2012). Vitamin D--baseline status and effective dose. N. Engl. J. Med. 367: 77–78.
Hoang, M.T., Defina, L.F., Willis, B.L., Leonard, D.S., Weiner, M.F., and Brown, E.S. (2011). Association between low serum 25-hydroxyvita-min D and depression in a large sample of healthy adults: the Cooper Center longitudinal study. Mayo Clin. Proc. 86: 1050–1055.
Holick, M.F. (2007). Vitamin D deficiency. N. Engl. J. Med. 357: 266–281.Holick, M.F. (2017). Ultraviolet B radiation: the vitamin D connection. Adv.
Exp. Med. Biol. 996: 137–154.Holick, M.F., Binkley, N.C., Bischoff-Ferrari, H.A., Gordon, C.M., Hanley,
D.A., Heaney, R.P., Murad, M.H., and Weaver, C.M. (2011). Evalua-tion, treatment, and prevention of vitamin D deficiency: an Endo-crine Society clinical practice guideline. J. Clin. Endocrinol. Metab. 96: 1911–1930.
Hollis, B.W. (1996). Assessment of vitamin D nutritional and hormonal sta-tus: what to measure and how to do it. Calcif. Tissue Int. 58: 4–5.
Hoogendijk, W.J.G., Lips, P., Dik, M.G., Deeg, D.J.H., Beekman, A.T.F., and Penninx, B.W.J.H. (2008). Depression is associated with decreased 25-hydroxyvitamin D and increased parathyroid hormone levels in older adults. Arch. Gen. Psychiatry. 65: 508–512.
Hossein-Nezhad, A., and Holick, M.F. (2013). Vitamin D for health: a global perspective. Mayo Clin. Proc. 88: 720–755.
Hossein-Nezhad, A., Spira, A., and Holick, M.F. (2013). Influence of vitamin D status and vitamin D3 supplementation on genome wide expres-sion of white blood cells: a randomized double-blind clinical trial. PLoS One 8: e58725.
Huang, J., Perlis, R.H., Lee, P.H., Rush, A.J., Fava, M., Sachs, G.S., Lieber-man, J., Hamilton, S.P., Sullivan, P., Sklar, P., Purcell, S., and Smoller, J.W. (2010). Cross-disorder genomewide analysis of schizophrenia, bipolar disorder, and depression. Am. J. Psychiatry. 167: 1254–1263.
Insel, T.R. (2015). The NIMH experimental medicine initiative. World Psy-chiatry. 14(2): 151–153.
Jääskeläinen, T., Knekt, P., Suvisaari, J., Männistö, S., Partonen, T., Sääk-
sjärvi, K., Kaartinen, N.E., Kanerva, N., and Lindfors, O. (2015). Higher serum 25-hydroxyvitamin D concentrations are related to a reduced risk of depression. Br. J. Nutr. 113: 1418–1426.
Jamilian, H., Bagherzadeh, K., Nazeri, Z., and Hassanijirdehi, M. (2013). Vitamin D, parathyroid hormone, serum calcium and phosphorus in patients with schizophrenia and major depression. Int. J. Psychiatry Clin. Pract. 17: 30–34.
Jäpelt, R.B., and Jakobsen, J. (2013). Vitamin D in plants: a review of occur-rence, analysis, and biosynthesis. Front. Plant Sci. 4: 136.
Johansson, P., Alehagen, U., van der Wal, M.H., Svensson, E., and Jaarsma, T. (2016). Vitamin D levels and depressive symptoms in patients with chronic heart failure. Int. J. Cardiol. 207: 185–189.
Jorde, R., Strand Hutchinson, M., Kjærgaard, M., Sneve, M., and Grimnes, G. (2013). Supplementation with high doses of vitamin D to subjects without vitamin D deficiency may have negative effects: pooled data from four intervention trials in Tromsø. ISRN Endocrinology. 2013: 348705.
Jorm, A.F. (2000). Mental health literacy. Public knowledge and beliefs about mental disorders. Br. J. Psychiatry. 177: 396–401.
Ju, S.-Y., Lee, Y.-J., and Jeong, S.-N. (2013). Serum 25-hydroxyvitamin D lev-els and the risk of depression: a systematic review and meta-analysis. J. Nutr. Health Aging. 17: 447–455.
Kalueff, A.V., and Tuohimaa, P. (2007). Neurosteroid hormone vitamin D and its utility in clinical nutrition. Curr. Opin. Clin. Nutr. Metab. Care. 10: 12–19.
von Känel, R., Fardad, N., Steurer, N., Horak, N., Hindermann, E., Fischer, F., and Gessler, K. (2015). Vitamin D deficiency and depressive symp-tomatology in psychiatric patients hospitalized with a current de-pressive episode: a factor analytic study. PLoS One 10: e0138550.
Kennedy, C., Bajdik, C.D., Willemze, R., De Gruijl, F.R., and Bouwes Bavinck, J.N. (2003). The influence of painful sunburns and lifetime sun ex-posure on the risk of actinic keratoses, seborrheic warts, melano-cytic nevi, atypical nevi, and skin cancer. J. Invest. Dermatol. 120: 1087–1093.
Kerr, D.C.R., Zava, D.T., Piper, W.T., Saturn, S.R., Frei, B., and Gombart, A.F. (2015). Associations between vitamin D levels and depressive symp-toms in healthy young adult women. Psychiatry Res. 227: 46–51.
Kessler, R.C., and Bromet, E.J. (2013). The epidemiology of depression across cultures. Annu. Rev. Publ. Health 34: 119–138.
Kessler, R.C., Angermeyer, M., Anthony, J.C., de Graaf, R., Demyttenaere, K., Gasquet, I., de Girolamo, G., Gluzman, S., Gureje, O., Haro, J.M., Kawakami, N., Karam, A., Levinson, D., Medina Mora, M.E., Oakley Browne, M.A., Posada-Villa, J., Stein, D.J., Adley Tsang, C.H., Aguilar-Gaxiola, S., Alonso, J., Lee, S., Heeringa, S., Pennell, B.-E., Berglund, P., Gruber, M.J., Petukhova, M., Chatterji, S., and Ustün, T.B. (2007). Lifetime prevalence and age-of-onset distributions of mental disor-ders in the World Health Organization’s World Mental Health Survey Initiative. World Psychiatry. 6: 168–176.
Kessler, R.C., Berglund, P., Demler, O., Jin, R., Koretz, D., Merikangas, K.R., Rush, A.J., Walters, E.E., and Wang, P.S. (2003). The epidemiology of major depressive disorder: results from the National Comorbidity Survey Replication (NCS-R). JAMA 289: 3095–3105.
Kjærgaard, M., Joakimsen, R., and Jorde, R. (2011). Low serum 25-hydrox-yvitamin D levels are associated with depression in an adult Norwe-gian population. Psychiatry Res. 190: 221–225.
Ko, P., Burkert, R., McGrath, J., and Eyles, D. (2004). Maternal vitamin D3 deprivation and the regulation of apoptosis and cell cycle during rat brain development. Brain Res. Dev. Brain Res. 153: 61–68.
Kongsbak, M., Levring, T.B., Geisler, C., and von Essen, M.R. (2013). The vitamin d receptor and T cell function. Front. Immunol. 4: 148.
Krishnan, V., and Nestler, E.J. (2010). Linking molecules to mood: new in-sight into the biology of depression. Am. J. Psychiatry. 167: 1305–1320.
Kuningas, M., Mooijaart, S.P., Jolles, J., Slagboom, P.E., Westendorp, R.G.J., and van Heemst, D. (2009). VDR gene variants associate with cogni-tive function and depressive symptoms in old age. Neurobiol. Aging. 30: 466–473.
Lai, J.S., Hiles, S., Bisquera, A., Hure, A.J., McEvoy, M., and Attia, J. (2014). A systematic review and meta-analysis of dietary patterns and depres-sion in community-dwelling adults. Am. J. Clin. Nutr. 99: 181–197.
Lange, K.W. (2018). Diet, exercise, and mental disorders - public health

Journal of Food Bioactives | www.isnff-jfb.com 19
Lange et al. Vitamin D and depression
challenges of the future. Mov. Nutr. Health Dis. 2: 39–59.Lange, K.W. (2020a). Lipids in the treatment of mental disorders. In:
Shahidi, F. (Ed.). Bailey’s Industrial Oil and Fat Products. Wiley, pp. 371–390.
Lange, K.W. (2020b). Omega-3 fatty acids and mental health. Glob. Health J. 4: 18–30.
Lange, K.W. (2021). Lack of evidence for efficacy of omega-3 fatty acids in depression. Acta Psychiatr. Scand. 144: 415–416.
Lange, K.W., Nakamura, Y., and Lange, K.M. (2020). The use of probiotics in depression. J. Food Bioact. 12: 3–8.
Lange, K.W., Nakamura, Y., Zhao, H., Bai, D., and Wang, H. (2021a). Are omega-3 fatty acids efficacious in the treatment of depression? A re-view. J. Food Bioact. 14: 10–19.
Lange, K.W., Nakamura, Y., Lange, K.M., and Zhao, H. (2021b). Tea and de-pression. Food Sci. Hum. Wellness in press.
Lappe, J.M., and Heaney, R.P. (2012). Why randomized controlled trials of calcium and vitamin D sometimes fail. Dermatoendocrinol. 4: 95–100.
Lee, D.M., Tajar, A., O’Neill, T.W., O’Connor, D.B., Bartfai, G., Boonen, S., Bouillon, R., Casanueva, F.F., Finn, J.D., Forti, G., Giwercman, A., Han, T.S., Huhtaniemi, I.T., Kula, K., Lean, M.E., Punab, M., Silman, A.J., Vanderschueren, D., Wu, F.C., and Pendleton, N. (2011). Lower vitamin D levels are associated with depression among community-dwelling European men. J. Psychopharmacol. 25: 1320–1328.
Li, G., Mbuagbaw, L., Samaan, Z., Falavigna, M., Zhang, S., Adachi, J.D., Cheng, J., Papaioannou, A., and Thabane, L. (2014). Efficacy of vita-min D supplementation in depression in adults: a systematic review. J. Clin. Endocrinol. Metab. 99: 757–767.
Libuda, L., Timmesfeld, N., Antel, J., Hirtz, R., Bauer, J., Führer, D., Zwan-ziger, D., Öztürk, D., Langenbach, G., Hahn, D., Ring, S., Peters, T., Hinney, A., Bühlmeier, J., Hebebrand, J., Grasemann, C., and Föcker, M. (2020). Effect of vitamin D deficiency on depressive symptoms in child and adolescent psychiatric patients: results of a randomized controlled trial. Eur. J. Nutr. 59: 3415–3424.
Lim, S.Y., Kim, E.J., Kim, A., Lee, H.J., Choi, H.J., and Yang, S.J. (2016). Nutri-tional factors affecting mental health. Clin. Nutr. Res. 5(3): 143–152.
Malhi, G.S., and Mann, J.J. (2018). Depression. Lancet. 392(10161): 2299–2312.
Marx, W., Moseley, G., Berk, M., and Jacka, F. (2017). Nutritional psychia-try: the present state of the evidence. Proc. Nutr. Soc. 76: 427–436.
Maxwell, J.D. (1994). Seasonal variation in vitamin D. Proc. Nutr. Soc. 53: 533–543.
Milaneschi, Y., Shardell, M., Corsi, A.M., Vazzana, R., Bandinelli, S., Gural-nik, J.M., and Ferrucci, L. (2010). Serum 25-hydroxyvitamin D and depressive symptoms in older women and men. J. Clin. Endocrinol. Metab. 95: 3225–3233.
Miller, G. (2010). Is pharma running out of brainy ideas? Science 329(5991): 502–504.
Mozaffari-Khosravi, H., Nabizade, L., Yassini-Ardakani, S.M., Hadinedoush-an, H., and Barzegar, K. (2013). The effect of 2 different single in-jections of high dose of vitamin D on improving the depression in depressed patients with vitamin D deficiency: a randomized clinical trial. J. Clin. Psychopharmacol. 33: 378–385.
Nanri, A., Mizoue, T., Matsushita, Y., Poudel-Tandukar, K., Sato, M., Ohta, M., and Mishima, N. (2009). Association between serum 25-hydroxy-vitamin D and depressive symptoms in Japanese: analysis by survey season. Eur. J. Clin. Nutr. 63: 1444–1447.
Obradovic, D., Gronemeyer, H., Lutz, B., and Rein, T. (2006). Cross-talk of vitamin D and glucocorticoids in hippocampal cells. J. Neurochem. 96: 500–509.
Okereke, O.I., and Singh, A. (2016). The role of vitamin D in the prevention of late-life depression. J. Affect. Disord. 198: 1–14.
Opie, R.S., O’Neil, A., Itsiopoulos, C., and Jacka, F.N. (2015). The impact of whole-of-diet interventions on depression and anxiety: a system-atic review of randomised controlled trials. Public Health Nutr. 18: 2074–2093.
Parker, G.B., Brotchie, H., and Graham, R.K. (2017). Vitamin D and depres-sion. J. Affect. Disord. 208: 56–61.
Polak, M.A., Houghton, L.A., Reeder, A.I., Harper, M.J., and Conner, T.S. (2014). Serum 25-hydroxyvitamin D concentrations and depressive symptoms among young adult men and women. Nutrients 6: 4720–
4730.Pan, A., Lu, L., Franco, O.H., Yu, Z., Li, H., and Lin, X. (2009). Association
between depressive symptoms and 25-hydroxyvitamin D in middle-aged and elderly Chinese. J. Affect. Disord. 118: 240–243.
Patrick, R.P., and Ames, B.N. (2015). Vitamin D and the omega-3 fatty acids control serotonin synthesis and action, part 2: relevance for ADHD, bipolar disorder, schizophrenia, and impulsive behavior. FASEB J. 29: 2207–2222.
Pietrzak, R.H., Kinley, J., Afifi, T.O., Enns, M.W., Fawcett, J., and Sareen, J. (2013). Subsyndromal depression in the United States: prevalence, course, and risk for incident psychiatric outcomes. Psychol. Med. 43: 1401–1414.
Pike, J.W., Meyer, M.B., Martowicz, M.L., Bishop, K.A., Lee, S.M., Nerenz, R.D., and Goetsch, P.D. (2010). Emerging regulatory paradigms for control of gene expression by 1,25-dihydroxyvitamin D3. J. Steroid Biochem. Mol. Biol. 121: 130–135.
Psaltopoulou, T., Sergentanis, T.N., Panagiotakos, D.B., Sergentanis, I.N., Kosti, R., and Scarmeas, N. (2013). Mediterranean diet, stroke, cogni-tive impairment, and depression: a meta-analysis. Ann. Neurol. 74: 580–591.
Rastmanesh, R., Beauchet, O., and Annweiler, C. (2012). Vitamin D defi-ciency and depression: causal relationship or artifact? Biofactors 38: 317–319.
Rush, A.J., Trivedi, M.H., Wisniewski, S.R., Nierenberg, A.A., Stewart, J.W., Warden, D., Niederehe, G., Thase, M.E., Lavori, P.W., Lebowitz, B.D., McGrath, P.J., Rosenbaum, J.F., Sackeim, H.A., Kupfer, D.J., Luther, J., and Fava, M. (2006a). Acute and longer-term outcomes in depressed outpatients requiring one or several treatment steps: a STAR*D re-port. Am. J. Psychiatry. 163: 1905–1917.
Rush, A.J., Trivedi, M.H., Wisniewski, S.R., Stewart, J.W., Nierenberg, A.A., Thase, M.E., Ritz, L., Biggs, M.M., Warden, D., Luther, J.F., Shores-Wil-son, K., Niederehe, G., and Fava, M. (2006b). Bupropion-SR, sertra-line, or venlafaxine-XR after failure of SSRIs for depression. N. Engl. J. Med. 354: 1231–1242.
Shaffer, J.A., Edmondson, D., Wasson, L.T., Falzon, L., Homma, K., Ezeokoli, N., Li, P., and Davidson, K.W. (2014). Vitamin D supplementation for depressive symptoms: a systematic review and meta-analysis of ran-domized controlled trials. Psychosom. Med. 76: 190–196.
Spedding, S. (2014). Vitamin D and depression: a systematic review and meta-analysis comparing studies with and without biological flaws. Nutrients 6: 1501–1518.
Tolppanen, A.-M., Fraser, A., Fraser, W.D., and Lawlor, D.A. (2012). Risk factors for variation in 25-hydroxyvitamin D3 and D2 concentrations and vitamin D deficiency in children. J. Clin. Endocrinol. Metab. 97: 1202–1210.
Lázaro Tomé, A., Reig Cebriá, M.J., González-Teruel, A., Carbonell-Asíns, J.A., Cañete Nicolás, C., and Hernández-Viadel, M. (2021). Efficacy of vitamin D in the treatment of depression: a systematic review and meta-analysis. Actas Esp. Psiquiatr. 49: 12–23.
Vieth, R. (2004). Why “Vitamin D” is not a hormone, and not a synonym for 1,25-dihydroxy-vitamin D, its analogs or deltanoids. J. Steroid Bio-chem. Mol. Biol. 89-90: 571–573.
Wacker, M., and Holick, M.F. (2013). Vitamin D - effects on skeletal and extraskeletal health and the need for supplementation. Nutrients 5: 111–148.
Wang, J., Liu, N., Sun, W., Chen, D., Zhao, J., and Zhang, W. (2018). Associa-tion between vitamin D deficiency and antepartum and postpartum depression: a systematic review and meta-analysis of longitudinal studies. Arch. Gynecol. Obstetr. 298: 1045–1059.
Wang, T.J., Zhang, F., Richards, J.B., Kestenbaum, B., van Meurs, J.B., Berry, D., Kiel, D.P., Streeten, E.A., Ohlsson, C., Koller, D.L., Peltonen, L., Cooper, J.D., O’Reilly, P.F., Houston, D.K., Glazer, N.L., Vandenput, L., Peacock, M., Shi, J., Rivadeneira, F., McCarthy, M.I., Anneli, P., de Boer, I.H., Mangino, M., Kato, B., Smyth, D.J., Booth, S.L., Jacques, P.F., Burke, G.L., Goodarzi, M., Cheung, C.-L., Wolf, M., Rice, K., Goltz-man, D., Hidiroglou, N., Ladouceur, M., Wareham, N.J., Hocking, L.J., Hart, D., Arden, N.K., Cooper, C., Malik, S., Fraser, W.D., Hartikainen, A.-L., Zhai, G., Macdonald, H.M., Forouhi, N.G., Loos, R.J.F., Reid, D.M., Hakim, A., Dennison, E., Liu, Y., Power, C., Stevens, H.E., Jaana, L., Vasan, R.S., Soranzo, N., Bojunga, J., Psaty, B.M., Lorentzon, M., Foroud, T., Harris, T.B., Hofman, A., Jansson, J.-O., Cauley, J.A., Uit-

Journal of Food Bioactives | www.isnff-jfb.com20
Vitamin D and depression Lange et al.
terlinden, A.G., Gibson, Q., Järvelin, M.-R., Karasik, D., Siscovick, D.S., Econs, M.J., Kritchevsky, S.B., Florez, J.C., Todd, J.A., Dupuis, J., Hyp-pönen, E., and Spector, T.D. (2010). Common genetic determinants of vitamin D insufficiency: a genome-wide association study. Lancet. 376: 180–188.
World Health Organization. (2008). The Global burden of disease: 2004 update. World Health Organization, Geneva, Switzerland.
Zanello, S.B., Jackson, D.M., and Holick, M.F. (1999). An immunocyto-chemical approach to the study of beta-endorphin production in hu-man keratinocytes using confocal microscopy. Ann. N. Y. Acad. Sci. 885: 85–99.
Zhao, G., Ford, E.S., Li, C., and Balluz, L.S. (2010). No associations between serum concentrations of 25-hydroxyvitamin D and parathyroid hor-mone and depression among US adults. Br. J. Nutr. 104: 1696–1702.

Copyright: © 2021 International Society for Nutraceuticals and Functional Foods.All rights reserved.
21
Review J. Food Bioact. 2021;15:21–28
Journal ofFood Bioactives International Society for
Nutraceuticals and Functional Foods
Phytochemicals: Potential Lead Compounds for COVID-19 Therapeutics
Srishti Kashyapa, Revathy Nadhana and Danny N. Dhanasekarana,b*
aStephenson Cancer Center, The University of Oklahoma Health Sciences Center, Oklahoma City, OK 73104, USAbDepartment of Cell Biology, The University of Oklahoma Health Sciences Center, Oklahoma City, OK 73104, USA*Corresponding author: Danny N. Dhanasekaran, Stephenson Cancer Center, The University of Oklahoma Health Sciences Center, Okla-homa City, OK 73104, USA. E-mail: [email protected]: 10.31665/JFB.2021.15279Received: September 27, 2021; Revised received & accepted: September 29, 2021Citation: Kashyap, S., Nadhan, R., and Dhanasekaran, D.N. (2021). Phytochemicals: Potential Lead Compounds for COVID-19 Therapeu-tics. J. Food Bioact. 15: 21–28.
Abstract
Coronavirus Disease 2019 (COVID-19) is a global pandemic, caused by severe acute respiratory syndrome corona-virus 2 (SARS- CoV-2). The rising number of cases of this highly transmissible infection has pressed for the urgent need to find effective therapeutics. The life cycle of SARS-CoV-2 includes the viral entry, viral replication, viral as-sembly and release. The symptoms associated with viral infection often leads to fatal outcome with pneumonia, myocarditis, acute respiratory distress syndrome, hypercoagulability, and/or multi-organ failure. Recent studies have reported that phytochemicals such as emodin, epigallocatechin gallate, and berberine could, albeit modest-ly, inhibit different stages of SARS-CoV-2 life cycle. The phytochemicals have been shown to disrupt viral infection and replication by blocking viral-surface spike protein binding to entry receptor angiotensin-converting enzyme (ACE2), inhibiting viral membrane fusion with host cells, inhibiting RNA-dependent RNA polymerase involved in viral replication, and/or pathological host- responses in vitro. The focus of this review is to evaluate the effica-cies of these phytochemicals on inhibiting SARS-CoV-2 viral infection, growth, or disease progression as well as to provide a perspective on the potential use of these phytochemicals in the development of novel therapeutics against SARS-CoV-2.
Keywords: SARS-CoV-2; COVID-19; Coronavirus; Phytochemicals; Cytokine Storm; Cytokine Release Syndrome.
1. Introduction
The worldwide outbreak of highly transmissible Coronavirus Disease-2019 (COVID-19) was caused by a zoonotic pathogenic virus, Severe Acute Respiratory Syndrome Coronavirus-2 (SARS-CoV-2). SARS-CoV-2 and its previous counterpart, SARS-CoV, which resulted in a pandemic in 2002, belong to the β-coronavirus lineage B, both of which are similar to the Middle East Respiratory Syndrome Coronavirus (MERS-CoV) that had emerged in 2012 (Zaki et al., 2012). Towards the end of 2019, the emergence of the COVID-19 pandemic posed an unprecedented global health threat, prompting the need to develop effective antiviral therapeutics im-mediately. During the initial stage of the pandemic, repurposing of
the existing drugs was considered to be one of the most tangible options for rapidly developing the effective therapeutics. Numer-ous clinical trials have been completed, but none of those evalu-ated repurposed drugs could give any promising results (Martinez & chemotherapy, 2020). The failure of these repurposed drugs in their clinical trials stresses the need to unleash novel structural motifs for developing effective COVID-19 drugs. A historical overview of drug discovery has shown that natural products could be well optimized for efficacious therapeutic strategies for many diseases including infectious diseases. The uniqueness of the vast array of chemical structures of natural products has been the inspi-ration behind the development of several effective pharmaceuti-cals (Clercq, 2007). In this review, we briefly discuss the potential

Journal of Food Bioactives | www.isnff-jfb.com22
Phytochemicals and COVID-19 Kashyap et al.
of several plant-derived natural products or phytochemicals, which could serve as lead compounds for synthesizing effective nature-inspired therapeutics against COVID-19.
2. COVID-19 virus and its life cycle
SARS-CoV-2 genome has a length of nearly 30,000 nucleotides and is the largest positive-sense genome of the RNA virus fam-ily (Astuti and Ysrafil, 2020). The genetic material of the virus contain a simple structure consisting of a single stranded RNA genome and bound ribonucleoprotein particles that provide struc-tural integrity to RNA in addition to playing a role in viral replica-tion. This RNA-RNP complex is encapsulated in a viral envelope consisting of lipid membrane. The viral envelope is interspersed with envelope and membrane proteins. The outward projecting and the receptor-interacting spike glycoproteins are also embed-ded in the viral envelope (Figure 1). The infection cycle of the vi-rus can be divided into three stages, namely, viral entry, viral RNA release and replication, and viral assembly and release. To enter the host cells, coronaviruses first bind to specific host cell sur-face receptors and then the viral Spike protein is cleaved by host cell-derived proteases to enable fusion of viral and cellular mem-branes (V’kovski et al., 2021). Receptor binding domain (RBD) of SARS-CoV-2 Spike (S) protein specifically recognizes human angiotensin- converting enzyme-2 (ACE-2) as its receptor (Letko et al., 2020; Wan et al., 2020). Proteolytic cleavage of S protein by host cell surface protease, TMPRSS2, leads to the final con-formational changes in the S2 domain of the spike protein needed for virus-host cell membrane fusion. This process is irreversible and it is stringently regulated (Walls et al., 2017). The release of SARS-CoV-2 viral genome in the host cell cytoplasm marks the onset of spatially and temporally regulated complex program of viral gene expression.
Coronavirus particles contain a single, 5′-capped and 3′-poly-adenylated RNA genome which codes for two large overlapping open reading frames in gene 1, as well as a variety of structural and nonstructural proteins at the 3′ end. At the 5′ end, SARS-
CoV-2 genomic RNA comprises of two overlapping reading frames ORF1a and ORF1b (Gorbalenya et al., 2006). The transla-tion of ORF1a and ORF1b from the genomic RNA produces two polyproteins, pp1a and pp1ab, respectively. The pp1a and pp1ab proteins are then cleaved by the viral proteases, papain-like pro-tease (PLPRO) and 3C-like protease (3CLPRO) to release 16 differ-ent non-structural proteins (Nsp). These encoded Nsp primarily comprises of RNA-dependent RNA polymerase (RdRp), helicase, and exonuclease (ExoN), which together forms a viral replication and transcription complex (RTC) (Subissi et al., 2014). The RdRp contains a catalytic subunit, NSP12, helicase (NSP13), and ac-cessory subunits NSP7 and NSP8 (Hillen et al., 2020). The RdRp complex transcribes the viral genome template of negative-sense genes, including progeny genome and subgenomic RNA as inter-mediate products, followed by the transcription of positive sense mRNAs mediated by RdRp. ExoN, exclusive to the β-coronavirus family, conducts RNA proofreading activity, which is critical for maintaining the viral genome integrity. The 3′ end of the viral ge-nome contains the ORFs which encode for viral structural proteins, namely the spike (S), the membrane (M), the envelope (E), and the nucleocapsid (N) proteins, in addition to the accessory proteins. These structural proteins would be incorporated into the endoplas-mic reticulum membrane and forms the membrane encased struc-tures, which further encloses the nucleocapsid and other neces-sary accessory proteins to form the exocytic vesicle. The exocytic vesicle fuses with the plasma membrane and releases the viruses into the extracellular space, which spreads throughout the human body (Murgolo et al., 2021; Romano et al., 2020; Trougakos et al., 2021). This could elicit pathological inflammatory response including cytokine release syndrome and cytokine storm. Eventu-ally, multiple organ failure ensues with the resultant death of the COVID-19 patient.
In spite of the fact that numerous drugs do exist that have clini-cally proven antiviral properties (Indari et al., 2021) and several drugs that show a limited potential to alleviate COVID-19 symp-toms (Lam et al., 2020; Raju et al., 2021), none of them have prov-en to work efficiently to reduce COVID-19 associated mortality. Nevertheless, several in vitro studies have demonstrated the po-tential of several phytochemicals to inhibit COVID-19 infectious
Figure 1. Structure of SARS-CoV-2 Virus. SARS-CoV-2 virus contains a single stranded RNA serves as the genetic material to which ribonucleoproteins that assist in viral structure and replication.are bound. The encapsulating viral envelope is comprised of a lipid layer interspersed with membrane and envelope proteins as well as characteristic receptor-interacting spike glycoprotein.

Journal of Food Bioactives | www.isnff-jfb.com 23
Kashyap et al. Phytochemicals and COVID-19
cycle in humans at various stages (Alam et al., 2021; Behl et al., 2021; Marmitt et al., 2021; Merarchi et al., 2021). These phyto-chemicals have been demonstrated to inhibit viral entry, viral rep-lication, or viral release – in addition to attenuating the host patho-logical responses (Figure 2). However, these studies have fallen short of proving a preventive or therapeutic role for these phyto-chemicals against COVID19 infection in real-time. Nevertheless, they provide strong evidence that they can serve as potential lead molecules for either the therapy or the prevention of SARS-CoV-2 infection as well as other viral infections that might emerge in the future (Swain et al., 2021).
3. Phytochemicals targeting SARS-CoV-2 entry into the host cell
Coronavirus entry into the host cell is one of the most important determinant of viral infectivity (Li, 2016) and is also the major target for human intervention strategies (Du et al., 2009b) . Coro-navirus spike (S) proteins are homotrimeric class I fusion glyco-proteins that comprise of two functionally distinct subunits: S1 and S2. The peripheral S1 subunit contains the receptor-binding domain that binds to the host cell receptor, ACE2 and is criti-cal in determining tropism of infection and pathogenicity (Li et al., 2003; Zhou et al., 2020). The transmembrane S2 subunit that contains the fusion peptide domain and the twin heptad repeat domains mediates viral and cellular membrane fusion upon exten-sive conformational rearrangements following host cell receptor binding (Letko et al., 2020). The S protein is a class I viral fusion membrane glycoprotein similar to HIV glycoprotein 160 (Env), influenza haemagglutinin (HA), and Ebola virus glycoprotein (Du
et al., 2009). In the first step of viral entry, the peripheral amino (S1) subunit of the SARS-CoV-2 spike glycoprotein (S) indepen-dently binds with the membrane metallopeptidase ACE2, and the carboxy (S2) terminus embedded into the viral envelope mediates fusion of viral and cellular membranes (Gallagher and Buchmeier, 2001). Inhibiting the first step of viral infection significantly min-imizes the chance for the virus to propagate, evolve, and eventu-ally acquire drug resistance (Breitinger et al., 2021; Esté, 2003; Horne and Vohl, 2020). Therefore, several phytochemicals have been investigated for their ability to exhibit inhibitory effect on viral entry (Figure 3).
Emodin, an anthraquinone polyphenol obtained from tradi-tional herbs Rhubarb (Rheum officinale) and Japanese knotweed (Polygonum cuspidatum), significantly inhibited the interaction of S protein with ACE2 in a dose-dependent manner, in a cell-free competition assay, with an IC50 of 200 μM (Ho et al., 2007). Emo-din also reduced the infection of ACE2-expressing Vero E6 cells, a non-human primate kidney cell-line, by an S protein-pseudo typed retrovirus (Ho et al., 2007). High-throughput screening of a library containing the extracts from 121 Chinese herbs led to the identification of two candidate low molecular weight phyto-chemicals, namely Tetra-O-galloyl-β- D-glucose (TGG) and lu-teolin (Yi et al., 2004). TGG and luteolin inhibited SARS-CoV infection by preventing viral entry into Vero E6 cells with the EC50 values of 4.5 μM and 10.6 μM, respectively. Frontal affinity chro-matography-mass spectrometry studies revealed that both TGG and luteolin bound with high affinity to SARS-CoV spike protein, suggesting that the inhibition of viral entry could be due to the interference in the interaction between ACE2 and spike protein (Yi et al., 2004). TGG is a polyphenolic compound obtained from In-dian gooseberry (Phyllanthus emblica), and Luteolin is a common
Figure 2. Potential Lead Phytochemicals that can target SARS-CoV-2. Several phytochemicals have been identified to have the potential to disrupt different stages of viral lifecycle or pathological host response following infection. The structures of candidate phytochemicals that have potential to serve as the lead compounds for drug discovery are presented as follows: (a) Phytochemicals targeting viral entry into the host cell; (b) Phytochemicals targeting viral replication; (c) Phytochemicals inhibiting viral release from the host cell; and (d) Phytochemicals that can attenuate host CRS or cytokine storm (See Text for Details).
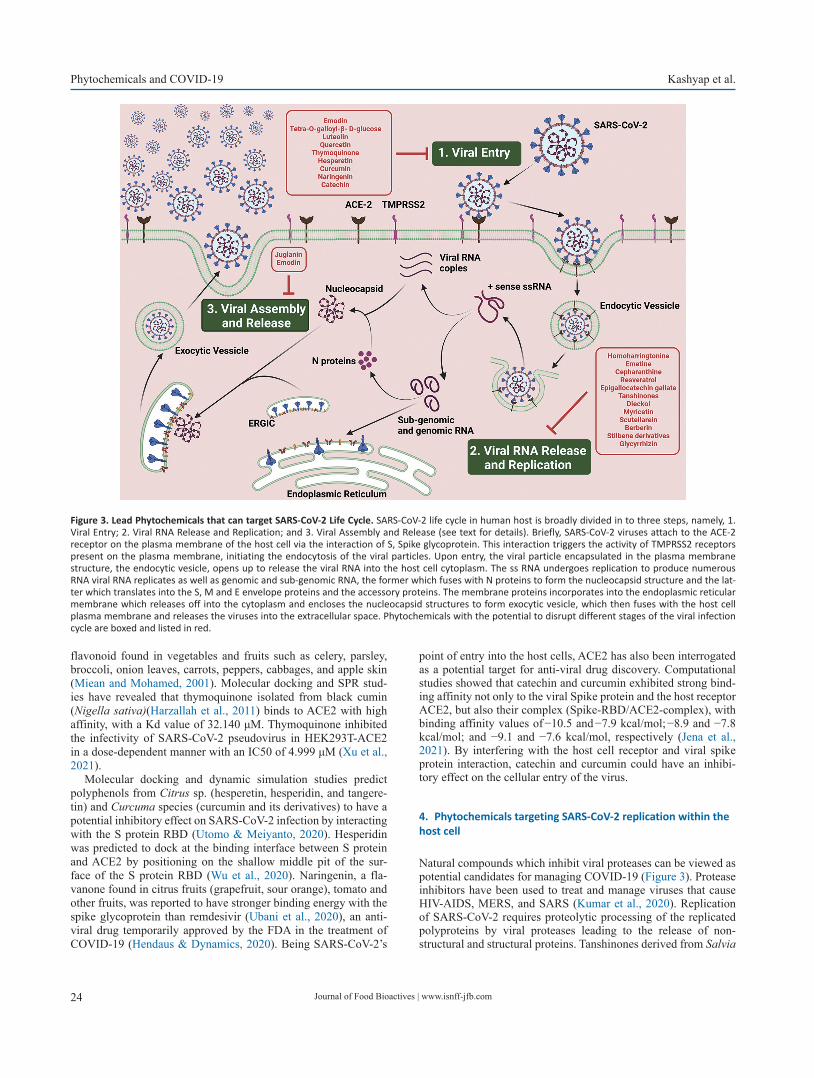
Journal of Food Bioactives | www.isnff-jfb.com24
Phytochemicals and COVID-19 Kashyap et al.
flavonoid found in vegetables and fruits such as celery, parsley, broccoli, onion leaves, carrots, peppers, cabbages, and apple skin (Miean and Mohamed, 2001). Molecular docking and SPR stud-ies have revealed that thymoquinone isolated from black cumin (Nigella sativa)(Harzallah et al., 2011) binds to ACE2 with high affinity, with a Kd value of 32.140 μM. Thymoquinone inhibited the infectivity of SARS-CoV-2 pseudovirus in HEK293T-ACE2 in a dose-dependent manner with an IC50 of 4.999 μM (Xu et al., 2021).
Molecular docking and dynamic simulation studies predict polyphenols from Citrus sp. (hesperetin, hesperidin, and tangere-tin) and Curcuma species (curcumin and its derivatives) to have a potential inhibitory effect on SARS-CoV-2 infection by interacting with the S protein RBD (Utomo & Meiyanto, 2020). Hesperidin was predicted to dock at the binding interface between S protein and ACE2 by positioning on the shallow middle pit of the sur-face of the S protein RBD (Wu et al., 2020). Naringenin, a fla-vanone found in citrus fruits (grapefruit, sour orange), tomato and other fruits, was reported to have stronger binding energy with the spike glycoprotein than remdesivir (Ubani et al., 2020), an anti-viral drug temporarily approved by the FDA in the treatment of COVID-19 (Hendaus & Dynamics, 2020). Being SARS-CoV-2’s
point of entry into the host cells, ACE2 has also been interrogated as a potential target for anti-viral drug discovery. Computational studies showed that catechin and curcumin exhibited strong bind-ing affinity not only to the viral Spike protein and the host receptor ACE2, but also their complex (Spike-RBD/ACE2-complex), with binding affinity values of −10.5 and −7.9 kcal/mol; −8.9 and −7.8 kcal/mol; and −9.1 and −7.6 kcal/mol, respectively (Jena et al., 2021). By interfering with the host cell receptor and viral spike protein interaction, catechin and curcumin could have an inhibi-tory effect on the cellular entry of the virus.
4. Phytochemicals targeting SARS-CoV-2 replication within the host cell
Natural compounds which inhibit viral proteases can be viewed as potential candidates for managing COVID-19 (Figure 3). Protease inhibitors have been used to treat and manage viruses that cause HIV-AIDS, MERS, and SARS (Kumar et al., 2020). Replication of SARS-CoV-2 requires proteolytic processing of the replicated polyproteins by viral proteases leading to the release of non-structural and structural proteins. Tanshinones derived from Salvia
Figure 3. Lead Phytochemicals that can target SARS-CoV-2 Life Cycle. SARS-CoV-2 life cycle in human host is broadly divided in to three steps, namely, 1. Viral Entry; 2. Viral RNA Release and Replication; and 3. Viral Assembly and Release (see text for details). Briefly, SARS-CoV-2 viruses attach to the ACE-2 receptor on the plasma membrane of the host cell via the interaction of S, Spike glycoprotein. This interaction triggers the activity of TMPRSS2 receptors present on the plasma membrane, initiating the endocytosis of the viral particles. Upon entry, the viral particle encapsulated in the plasma membrane structure, the endocytic vesicle, opens up to release the viral RNA into the host cell cytoplasm. The ss RNA undergoes replication to produce numerous RNA viral RNA replicates as well as genomic and sub-genomic RNA, the former which fuses with N proteins to form the nucleocapsid structure and the lat-ter which translates into the S, M and E envelope proteins and the accessory proteins. The membrane proteins incorporates into the endoplasmic reticular membrane which releases off into the cytoplasm and encloses the nucleocapsid structures to form exocytic vesicle, which then fuses with the host cell plasma membrane and releases the viruses into the extracellular space. Phytochemicals with the potential to disrupt different stages of the viral infection cycle are boxed and listed in red.

Journal of Food Bioactives | www.isnff-jfb.com 25
Kashyap et al. Phytochemicals and COVID-19
miltiorrhiza (red sage) moderately inhibit the SAR-CoV 3CLpro and PLpro proteases at IC50 values ranging from 0.8 to 30 μM (Park et al., 2012).
Dieckol, a pholorotannin obtained from Ecklonia cava, is re-ported to exert significant inhibitory activity (IC50= 2.7 μM) on coronavirus 3CLpro. Dieckol which possess two eckol groups linked by a diphenyl ether, also exhibited potent inhibitory activity on the cell-based 3CLprocis-cleavage assay, and its potency was greater than the other phlorotannin derivatives and the natural ref-erence inhibitors (Park et al., 2013). Myricetin, a flavonoid com-monly present in nuts, berries, tea, wine, and Scutellarein, a fla-vone, inhibits the SARS-CoV helicase protein in vitro by affecting its ATPase activity, but not the unwinding activity (Yu et al., 2012).
Recent studies showed that homoharringtonine (HHT) and emetine inhibited SARS-CoV-2 replication in Vero-E6 cells with EC50 at 2.55 and 0.46 μM, respectively (Choy et al., 2020). Lyco-rine, emetine, and cephaeline may inhibit viral protein synthesis by interacting with the host ribosome (Ren et al., 2021). Berberine an alkaloid isolated from plants such as Berberis has been reported to inhibit the replication of SARS-CoV-2 in cultured nasal cell lines (Varghese et al., 2021). Resveratrol, a well-known polyphenol inhibits SARS-CoV-2 viral replication (Pasquereau et al., 2021; ter Ellen et al., 2021; Yang et al., 2021). Glycorhyzin, a triterpene saponin, found in high concentrations in the root of the Glycyr-rhiza glabra (liquorice) plant inhibits viral replication in vitro by inhibiting the CLpro protease (van de Sand et al., 2021). EGCG, a major component in green tea, has also been shown to inhibit viral replication in Vero cells. This inhibition is associated with EGCG’s ability to inhibit Nsp15, a non-structural protein involved in the processing of the viral genome for replication (Hong et al., 2021). In addition to the thymoquinones which has been reported to inhibit the viral entry into the host cells, plumbagin, which is a naphthoquinone, isolated from roots of Plumbago zeylanica has been predicted to induce viral RNA degradation, through its ROS inductive effects, thereby exerting an antiviral therapeutic response (Nadhan et al., 2021).
5. Phytochemicals targeting SARS-CoV-2 viral release from the infected host cell
Phytochemicals belonging to Kaempferol family of flavanol such as Juglanin has been tested for their ability to inhibit the release of assembled mature virions from the host cell into extracellular space to propagate cell-to-cell spread of the infection (Figure 3). The ORF 3a of SARS-CoV codes for a transmembrane protein that forms a homotetrameric ion channel and modulates viral re-lease (Lu et al., 2006). It can be reasoned that the phytochemicals that inhibit the ion channel could exert an inhibitory effect on vi-rus release, and they could serve as lead compounds for the de-velopment of novel therapeutic antiviral agents. Kaempferols are natural flavonols, found in plants such as kale, beans, tea, spinach and broccoli (Baenas et al., 2012). Several kaempferol derivatives were reported to moderately block the 3a channel of Coronavi-rus, hence, inhibiting the viral production and release from the host cells (Schwarz et al., 2014). Kaempferol glycoside deriva-tives were proved to possess more potent inhibitory effect than kaempferol and points towards the significance of sugar residues for the antiviral activity (Schwarz et al., 2014) Juglanin (kaemp-ferol-3-O-a-L-arabinofuranoside) was the most effective glyco-side with an IC50 value of 2.3 μM. Other kaempferol glycosides i.e. tiliroside (kaempferol-3-O-(6-p-coumaroyl)-glucoside) and afzelin (kaempferol-3-O-a-L-rhamnoside) were less potent than
juglanin, but showed similar activity to that of emodin (Schwarz et al., 2014).
6. Phytochemicals targeting cytokine release syndrome/cy-tokine storm
In addition to this array of phytochemicals that target viral entry, replication, and release, there has been increased focus on phyto-chemicals that can have modulatory effect on Cytokine Release Syndrome (CRS) and cytokine storm of the infected patients (Figure 4). CRS and cytokine storm can be broadly described as life-threatening systemic inflammatory syndromes. In CRS and cytokine storm, elevated levels of inflammatory cytokines and excessive activation of immune cells lead to a range of clinical manifestations such as fever, fatigue, disseminated intravascular coagulation (DIC), hypotension, hemostatic imbalance, respira-tory failure (acute respiratory distress syndrome - ARDS), multi-organ failure, and even death, if not treated in time (Mangalmurti and Hunter, 2020). CRS and cytokine storm are primarily asso-ciated with infectious and immune-mediated conditions. Severe pneumonia with elevated cytokine levels such as interferon-γ (IFNγ), Interleukin (IL)- 6, and granulocyte-macrophage colony stimulating factor (GM-CSF) was reported to be a major cause of morbidity in patients infected with highly pathogenic human coronaviruses (SARS-CoV and MERS-CoV) (Channappanavar & Perlman, 2017). In a recent study conducted with 150 patients from Wuhan, China, elevated cytokine levels of IL-6 in patients with COVID-19 was suggested to be the underlying cause of ad-verse clinical outcomes (Ruan et al., 2020). Many of the recent studies have suggested that cytokine storm, composed of an ar-ray of cytokines including IL-1, -2, -6, -7, -8, -10, -12, -17, -18; tumor necrosis factor-α (TNF-α), IFN-γ, granulocyte colony-stimulating factor (G-CSF); GM-CSF, and monocyte chemoat-tractant protein-1 (MCP-1), is the underlying cause of severe COVID-19 immunopathology (Hadjadj et al., 2020; Lucas et al., 2020; McElvaney et al., 2020; Nile et al., 2020; Ronit et al., 2021).
Dehydrozingerone, a phenolic compound found in Zingiber officinale (ginger) and a structural-half analog of curcumin, was shown to exert significant inhibitory effect on LPS induced cy-tokine storm in cultured macrophage cells (Tirunavalli et al., 2021). Many known dietary polyphenols such as curcumin, api-genin, quercetin, cinnamaldehyde, resveratrol, and epigallocat-echin-3-gallate; alone and in combinations, have been reported to have significantly reduced the levels of pro-inflammatory cy-tokines in vitro and in vivo (Giovinazzo and Grieco, 2015; John et al., 2011; Scalbert et al., 2005; Tsao, 2010). Bioactive metabolites Indole-3-carbinol and sulforaphanes present in cruciferous vegeta-bles have also been demonstrated to have a moderate inhibitory ef-fect on LPS-induced inflammatory response in cultured cells (Guo et al., 2012; Jiang et al., 2013). Such phytochemicals can serve as potential lead compounds for the synthesis of effective therapeu-tics, which could reduce the cytokine storm.
7. Perspective and conclusion
COVID-19 has evolved to be a pandemic with global concern since 2020, which brought the world to a standstill. With the novel vaccine strategies, we are slowly emerging out of the abysmal crisis. However, a critical concern is that an effective therapeutic agent and/or a potential preventive strategy is still elusive. This can

Journal of Food Bioactives | www.isnff-jfb.com26
Phytochemicals and COVID-19 Kashyap et al.
be primarily attributed to the enhanced pace of infection and lack of sufficient time for deeper research and validation. During the initial phases of the pandemic, the researchers had come up with the notion of repurposing the existing drugs with antiviral activ-ity, most of which were FDA approved and used against various human diseases. However, most of them, if not all, failed to prove their efficacy in preventing or treating COVID-19 infections in real-world situation. Thus, the need for the hour is to develop nov-el therapeutic agents which can effectively target the COVID-19 infection in humans. The mechanistic role of phytochemicals dis-cussed here needs to be explored further and experimentally vali-dated so that more potent molecules can be developed. It is quite encouraging to note that the limited examples of phytochemicals discussed here provide evidence that they can serve as critical lead compounds for the development of novel therapeutics for treat-ing and preventing COVID-19 infections or similar pathogens that might emerge in the future.
Acknowledgments
The funding supports from the Stephenson Cancer Center and Na-tional Institutes of Health grant (grant no. GM103639) are great-fully acknowledged. Computational services were supported by the National Institute of General Medical Sciences P20 (grant no.
GM103639) and the National Cancer Insti-tute of the National In-stitutes of Health (grant no. P30CA225520).
Conflict of interest
The authors declare that there is no conflict of interest.
References
Alam, S., Sarker, M.M.R., Afrin, S., Richi, F.T., Zhao, C., Zhou, J.-R., and Mo-hamed, I.N. (2021). Traditional Herbal Medicines, Bioactive Metabo-lites, and Plant Products Against COVID-19: Update on Clinical Trials and Mechanism of Actions. Front. Pharmacol. 12: 671498.
Astuti, I., and Ysrafil, (2020). Severe Acute Respiratory Syndrome Coro-navirus 2 (SARS-CoV-2): An overview of viral structure and host re-sponse. Diabetes Metab. Syndr. 14(4): 407–412.
Baenas, N., Moreno, D.A., and García-Viguera, C. (2012). Selecting Sprouts of Brassicaceae for Optimum Phytochemical Composition. J. Agric. Food Chem. 60(45): 11409–11420.
Behl, T., Rocchetti, G., Chadha, S., Zengin, G., Bungau, S., Kumar, A., Me-hta, V., Uddin, M.S., Khullar, G., Setia, D., Arora, S., Sinan, K.I., Ak, G., Putnik, P., Gallo, M., and Montesano, D. (2021). Phytochemicals from Plant Foods as Potential Source of Antiviral Agents: An Overview. Pharmaceuticals 14(4): 381.
Figure 4. Lead Phytochemicals that can Targeting Pathological Host Response. Upon the entry into the human body, SARS-CoV-2 replicates within the hu-man host cells, especially within the alveolar epithelial cells, as the route of entry is via the respiratory tract. Further, it triggers the immune response via activating the immune cells such as T-cell, alveolar macrophages, neutrophils, natural killer cells, dendritic cells, fibroblasts. monocytes and endothelial cells. These activated immune cells potentiates a cascade of signaling events inducing the release of varied cytokines such as IL-1β/2/6/8/10/18, TNF-α, IFN-γ, G-CSF, GM-CSF, CXCL-10 and MCP-1, producing CRS and cytokine storm, which can lead to multiple organ failure, affecting the major human organs includ-ing brain, lungs, liver, heart, intestine and kidney. Phytochemicals that have therapeutic or preventive potential to target pathological CRS and/or cytokine storm are boxed and listed in red.

Journal of Food Bioactives | www.isnff-jfb.com 27
Kashyap et al. Phytochemicals and COVID-19
Breitinger, U., Ali, N.K.M., Sticht, H., and Breitinger, H.-G. (2021). Inhibition of SARS CoV Envelope Protein by Flavonoids and Classical Viroporin Inhibitors. Front. Microbiol. 12: 692423–692423.
Channappanavar, R., and Perlman, S. (2017). Pathogenic human coronavi-rus infections: causes and consequences of cytokine storm and im-munopathology. Semin. Immunopathol. 39(5): 529–539.
Choy, K.-T., Wong, A.Y.-L., Kaewpreedee, P., Sia, S.F., Chen, D., Hui, K.P.Y., Chu, D.K.W., Chan, M.C.W., Cheung, P.P.-H., Huang, X., Peiris, M., and Yen, H.-L. (2020). Remdesivir, lopinavir, emetine, and homohar-ringtonine inhibit SARS-CoV-2 replication in vitro. Antiviral Res. 178: 104786.
Clercq, E.D. (2007). Three decades of antiviral drugs. Nat. Rev. Drug Discov. 6(12): 941–941.
Du, L., He, Y., Zhou, Y., Liu, S., Zheng, B.-J., and Jiang, S. (2009). The spike protein of SARS-CoV — a target for vaccine and therapeutic develop-ment. Nat. Rev. Microbiol. 7(3): 226–236.
Este, J. (2003). Virus Entry as a Target for Anti-HIV Intervention . CMC 10(17): 1617–1632.
Gallagher, T.M., and Buchmeier, M.J. (2001). Coronavirus Spike Proteins in Viral Entry and Pathogenesis. Virology 279(2): 371–374.
Giovinazzo, G., and Grieco, F. (2015). Functional Properties of Grape and Wine Polyphenols. Plant Foods Hum. Nutr. 70(4): 454–462.
Gorbalenya, A.E., Enjuanes, L., Ziebuhr, J., and Snijder, E.J. (2006). Ni-dovirales: Evolving the largest RNA virus genome. Virus Res. 117(1): 17–37.
Guo, S., Qiu, P., Xu, G., Wu, X., Dong, P., Yang, G., Zheng, J., McClements, D.J., and Xiao, H. (2012). Synergistic Anti-inflammatory Effects of Nobiletin and Sulforaphane in Lipopolysaccharide-Stimulated RAW 264.7 Cells. J. Agric. Food Chem. 60(9): 2157–2164.
Hadjadj, J., Yatim, N., Barnabei, L., Corneau, A., Boussier, J., Smith, N., Péré, H., Charbit, B., Bondet, V., Chenevier-Gobeaux, C., Breillat, P., Carlier, N., Gauzit, R., Morbieu, C., Pène, F., Marin, N., Roche, N., Szwebel, T.-A., Merkling, S.H., Treluyer, J.-M., Veyer, D., Mouthon, L., Blanc, C., Tharaux, P.-L., Rozenberg, F., Fischer, A., Duffy, D., Rieux-Laucat, F., Kernéis, S., and Terrier, B. (2020). Impaired type I interferon activ-ity and inflammatory responses in severe COVID-19 patients. Science 369(6504): 718–724.
Harzallah, H.J., Kouidhi, B., Flamini, G., Bakhrouf, A., and Mahjoub, T. (2011). Food Chem. 129(4): 1469–1474.
Hendaus, M.A. (2021). Remdesivir in the treatment of coronavirus dis-ease 2019 (COVID-19): a simplified summary. J. Biomol. Struct. Dyn. 39(10): 3787–3792.
Hillen, H.S., Kokic, G., Farnung, L., Dienemann, C., Tegunov, D., and Cramer, P. (2020). Structure of replicating SARS-CoV-2 polymerase. Nature 584(7819): 154–156.
Ho, T, Wu, S, Chen, J, Li, C, and Hsiang, C (2007). Emodin blocks the SARS coronavirus spike protein and angiotensin-converting enzyme 2 in-teraction. Antiviral Res. 74(2): 92–101.
Hong, S., Seo, S.H., Woo, S.-J., Kwon, Y., Song, M., and Ha, N.-C. (2021). Epi-gallocatechin Gallate Inhibits the Uridylate-Specific Endoribonucle-ase Nsp15 and Efficiently Neutralizes the SARS-CoV-2 Strain. J. Agric. Food Chem. 69(21): 5948–5954.
Horne, J.R., and Vohl, M.-C. (2020). Biological plausibility for interactions between dietary fat, resveratrol, ACE2, and SARS-CoV illness severity. Am. J. Physiol. Endocrinol. Metab. 318(5): E830–E833.
Indari, O., Jakhmola, S., Manivannan, E., and Jha, H.C. (2021). An Update on Antiviral Therapy Against SARS-CoV-2: How Far Have We Come? Front. Pharmacol. 12: 632677.
Jena, A.B., Kanungo, N., Nayak, V., Chainy, G.B.N., and Dandapat, J. (2021). Catechin and curcumin interact with S protein of SARS-CoV2 and ACE2 of human cell membrane: insights from computational studies. Sci. Rep. 11(1): 2043.
Jiang, J., Kang, T.B., Shim, D.W., Oh, N.H., Kim, T.J., and Lee, K.H. (2013). Indole-3-carbinol inhibits LPS-induced inflammatory response by blocking TRIF-dependent signaling pathway in macrophages. Food Chem. Toxicol. 57: 256–261.
John, C.M., Sandrasaigaran, P., Tong, C.K., Adam, A., and Ramasamy, R. (2011). Immunomodulatory activity of polyphenols derived from Cassia auriculata flowers in aged rats. Cell. Immunol. 271(2): 474–479.
Kumar, S., Zhi, K., Mukherji, A., and Gerth, K. (2020). Repurposing Antiviral
Protease Inhibitors Using Extracellular Vesicles for Potential Therapy of COVID-19. Viruses 12(5): 486.
Lam, S., Lombardi, A., and Ouanounou, A. (2020). COVID-19: A review of the proposed pharmacological treatments. Eur. J. Pharmacol. 886: 173451.
Letko, M., Marzi, A., and Munster, V. (2020). Functional assessment of cell entry and receptor usage for SARS-CoV-2 and other lineage B betac-oronaviruses. Nat. Microbiol. 5(4): 562–569.
Li, F. (2016). Structure, Function, and Evolution of Coronavirus Spike Pro-teins. Annu. Rev. Virol. 3(1): 237–261.
Li, W., Moore, M.J., Vasilieva, N., Sui, J., Wong, S.K., Berne, M.A., So-masundaran, M., Sullivan, J.L., Luzuriaga, K., Greenough, T.C., Choe, H., and Farzan, M. (2003). Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 426(6965): 450–454.
Lu, W., Zheng, B.-J., Xu, K., Schwarz, W., Du, L., Wong, C.K.L., Chen, J., Duan, S., Deubel, V., and Sun, B. (2006). Severe acute respiratory syndrome-associated coronavirus 3a protein forms an ion channel and modu-lates virus release. Proc. Natl. Acad. Sci. 103(33): 12540–12545.
Lucas, C., Wong, P., Klein, J., Castro, T.B.R., Silva, J., Sundaram, M., Elling-son, M.K., Mao, T., Oh, J.E., Israelow, B., Takahashi, T., Tokuyama, M., Lu, P., Venkataraman, A., Park, A., Mohanty, S., Wang, H., Wyl-lie, A.L., Vogels, C.B.F., Earnest, R., Lapidus, S., Ott, I.M., Moore, A.J., Muenker, M.C., Fournier, J.B., Campbell, M., Odio, C.D., Casanovas-Massana, A., Yale IMPACT Team, Obaid, A., Lu-Culligan, A., Nelson, A., Brito, A., Nunez, A., Martin, A., Watkins, A., Geng, B., Kalinich, C., Harden, C., Todeasa, C., Jensen, C., Kim, D., McDonald, D., Shepard, D., Courchaine, E., White, E.B., Song, E., Silva, E., Kudo, E., DeIuliis, G., Rahming, H., Park, H.-J., Matos, I., Nouws, J., Valdez, J., Fauver, J., Lim, J., Rose, K.-A., Anastasio, K., Brower, K., Glick, L., Sharma, L., Sewa-nan, L., Knaggs, L., Minasyan, M., Batsu, M., Petrone, M., Kuang, M., Nakahata, M., Campbell, M., Linehan, M., Askenase, M.H., Simonov, M., Smolgovsky, M., Sonnert, N., Naushad, N., Vijayakumar, P., Mar-tinello, R., Datta, R., Handoko, R., Bermejo, S., Prophet, S., Bickerton, S., Velazquez, S., Alpert, T., Rice, T., Khoury-Hanold, W., Peng, X., Yang, Y., Cao, Y., Strong, Y., Herbst, R., Shaw, A.C., Medzhitov, R., Schulz, W.L., Grubaugh, N.D., Dela Cruz, C., Farhadian, S., Ko, A.I., Omer, S.B., and Iwasaki, A. (2020). Longitudinal analyses reveal immunological misfiring in severe COVID-19. Nature 584(7821): 463–469.
Mangalmurti, N., and Hunter, C.A. (2020). Cytokine Storms: Understand-ing COVID-19. Immunity 53(1): 19–25.
Marmitt, D.J., Goettert, M.I., and Rempel, C. (2021). Compounds of plants with activity against SARS-CoV-2 targets. Expert Rev. Clin. Pharmacol. 14(5): 623–633.
Martinez, M.A. (2020). Compounds with Therapeutic Potential against Novel Respiratory 2019 Coronavirus. Antimicrob. Agents Chemother. 64(5): e00399-e00320.
McElvaney, O.J., McEvoy, N.L., McElvaney, O.F., Carroll, T.P., Murphy, M.P., Dunlea, D.M., Ní Choileáin, O., Clarke, J., O’Connor, E., Hogan, G., Ryan, D., Sulaiman, I., Gunaratnam, C., Branagan, P., O’Brien, M.E., Morgan, R.K., Costello, R.W., Hurley, K., Walsh, S., de Barra, E., Mc-Nally, C., McConkey, S., Boland, F., Galvin, S., Kiernan, F., O’Rourke, J., Dwyer, R., Power, M., Geoghegan, P., Larkin, C., O’Leary, R.A., Freeman, J., Gaffney, A., Marsh, B., Curley, G.F., and McElvaney, N.G. (2020). Characterization of the Inflammatory Response to Severe COVID-19 Illness. Am. J. Respir. Crit. Care Med. 202(6): 812–821.
Merarchi, M., Dudha, N., Das, B.C., and Garg, M. (2021). Natural products and phytochemicals as potential anti-SARS-CoV-2 drugs. Phytother. Res.
Miean, K.H., and Mohamed, S. (2001). Flavonoid (Myricetin, Querce-tin, Kaempferol, Luteolin, and Apigenin) Content of Edible Tropical Plants. J. Agric. Food Chem. 49(6): 3106–3112.
Murgolo, N., Therien, A.G., Howell, B., Klein, D., Koeplinger, K., Lieberman, L.A., Adam, G.C., Flynn, J., McKenna, P., Swaminathan, G., Hazuda, D.J., Olsen, D.B., and Hobman, T.C. (2021). SARS-CoV-2 tropism, en-try, replication, and propagation: Considerations for drug discovery and development. PLoS Pathog. 17(2): e1009225.
Nadhan, R., Patra, D., Krishnan, N., Rajan, A., Gopala, S., Ravi, D., and Srinivas, P. (2021). Perspectives on mechanistic implications of ROS inducers for targeting viral infections. Eur. J. Pharmacol. 890: 173621.
Nile, S.H., Nile, A., Qiu, J., Li, L., Jia, X., and Kai, G. (2020). COVID-19: Patho-

Journal of Food Bioactives | www.isnff-jfb.com28
Phytochemicals and COVID-19 Kashyap et al.
genesis, cytokine storm and therapeutic potential of interferons. Cy-tokine Growth Factor Rev. 53: 66–70.
Park, J.-Y., Kim, J.H., Kim, Y.M., Jeong, H.J., Kim, D.W., Park, K.H., Kwon, H.-J., Park, S.-J., Lee, W.S., and Ryu, Y.B. (2012). Tanshinones as selective and slow-binding inhibitors for SARS-CoV cysteine proteases. Bioorg. Med. Chem. 20(19): 5928–5935.
Park, J.-Y., Kim, J.H., Kwon, J.M., Kwon, H.-J., Jeong, H.J., Kim, Y.M., Kim, D., Lee, W.S., and Ryu, Y.B. (2013). Dieckol, a SARS-CoV 3CLpro inhibi-tor, isolated from the edible brown algae Ecklonia cava. Bioorg. Med. Chem. 21(13): 3730–3737.
Pasquereau, S., Nehme, Z., Haidar Ahmad, S., Daouad, F., Van Assche, J., Wallet, C., Schwartz, C., Rohr, O., Morot-Bizot, S., and Herbein, G. (2021). Resveratrol Inhibits HCoV-229E and SARS-CoV-2 Coronavirus Replication In Vitro. Viruses 13(2): 354.
Raju, R., V., P., Biatris, P.S., and J., S.J.U.C. (2021). Therapeutic role of cor-ticosteroids in COVID-19: a systematic review of registered clinical trials. Futur. J. Pharm. Sci 7(1): 67.
Ren, P.-X., Shang, W.-J., Yin, W.-C., Ge, H., Wang, L., Zhang, X.-L., Li, B.-Q., Li, H.-L., Xu, Y.-C., Xu, E.H., Jiang, H.-L., Zhu, L.-L., Zhang, L.-K., and Bai, F. (2021). A multi-targeting drug design strategy for identifying potent anti-SARS-CoV-2 inhibitors. Acta Pharmacol. Sin.
Romano, M., Ruggiero, A., Squeglia, F., Maga, G., and Berisio, R. (2020). A Structural View of SARS-CoV-2 RNA Replication Machinery: RNA Synthesis, Proofreading and Final Capping. Cells 9(5): 1267.
Ronit, A., Berg, R.M.G., Bay, J.T., Haugaard, A.K., Ahlström, M.G., Burgdorf, K.S., Ullum, H., Rørvig, S.B., Tjelle, K., Foss, N.B., Benfield, T., Mar-quart, H.V., and Plovsing, R.R. (2021). Compartmental immunophe-notyping in COVID-19 ARDS: A case series. J. Allergy Clin. Immunol. 147(1): 81–91.
Ruan, Q., Yang, K., Wang, W., Jiang, L., and Song, J. (2020). Clinical predic-tors of mortality due to COVID-19 based on an analysis of data of 150 patients from Wuhan, China. Intensive Care Med. 46(5): 846–848.
Scalbert, A., Manach, C., Morand, C., Rémésy, C., and Jiménez, L. (2005). Dietary Polyphenols and the Prevention of Diseases. Crit. Rev. Food Sci. Nutr. 45(4): 287–306.
Schwarz, S., Sauter, D., Wang, K., Zhang, R., Sun, B., Karioti, A., Bilia, A., Ef-ferth, T., and Schwarz, W. (2014). Kaempferol Derivatives as Antiviral Drugs against the 3a Channel Protein of Coronavirus. Planta. Med. 80(02/03): 177–182.
Subissi, L., Imbert, I., Ferron, F., Collet, A., Coutard, B., Decroly, E., and Canard, B. (2014). SARS-CoV ORF1b-encoded nonstructural proteins 12–16: Replicative enzymes as antiviral targets. Antiviral Res. 101: 122–130.
Swain, S.S., Panda, S.K., and Luyten, W. (2021). Phytochemicals against SARS-CoV as potential drug leads. Biomed. J. 44(1): 74–85.
ter Ellen, B.M., Dinesh Kumar, N., Bouma, E.M., Troost, B., van de Pol, D.P.I., van der Ende-Metselaar, H.H., Apperloo, L., van Gosliga, D., van den Berge, M., Nawijn, M.C., van der Voort, P.H.J., Moser, J., Roden-huis-Zybert, I.A., and Smit, J.M. (2021). Resveratrol and Pterostilbene Inhibit SARS-CoV-2 Replication in Air–Liquid Interface Cultured Hu-man Primary Bronchial Epithelial Cells. Viruses 13(7): 1335.
Tirunavalli, S.K., Gourishetti, K., Kotipalli, R.S.S., Kuncha, M., Kathirvel, M., Kaur, R., Jerald, M.K., Sistla, R., and Andugulapati, S.B. (2021). Dehydrozingerone ameliorates Lipopolysaccharide induced acute respiratory distress syndrome by inhibiting cytokine storm, oxidative stress via modulating the MAPK/NF-κB pathway. Phytomedicine 92: 153729.
Trougakos, I.P., Stamatelopoulos, K., Terpos, E., Tsitsilonis, O.E., Aivalioti, E., Paraskevis, D., Kastritis, E., Pavlakis, G.N., and Dimopoulos, M.A. (2021). Insights to SARS-CoV-2 life cycle, pathophysiology, and ra-tionalized treatments that target COVID-19 clinical complications. J. Biomed. Sci. 28(1): 9.
Tsao, R. (2010). Chemistry and Biochemistry of Dietary Polyphenols. Nutri-ents 2(12): 1231–1246.
Ubani, A., Agwom, F., RuthMorenikeji, O., Nathan, S., Luka, P., Umera, A., Umar, U., Omale, S., Nnadi, N.E., and Aguiyi, J.C. (2021). Molecular Docking Analysis Of Some Phytochemicals On Two SARS-CoV-2 Tar-gets: Potential Lead Compounds Against Two Target Sites of SARS-CoV-2 Obtained from Plants. bioRxiv .
Utomo, R.Y., Ikawati, M., and Meiyanto, E. (2020). Revealing the potency of citrus and galangal constituents to halt SARS-CoV-2 infection. Pre-prints .
V’kovski, P., Kratzel, A., Steiner, S., Stalder, H., and Thiel, V. (2021). Corona-virus biology and replication: implications for SARS-CoV-2. Nat. Rev. Microbiol. 19(3): 155–170.
van de Sand, L., Bormann, M., Alt, M., Schipper, L., Heilingloh, C.S., Stein-mann, E., Todt, D., Dittmer, U., Elsner, C., Witzke, O., and Krawczyk, A. (2021). Glycyrrhizin Effectively Inhibits SARS-CoV-2 Replication by Inhibiting the Viral Main Protease. Viruses 13(4): 609.
Varghese, F., van Woudenbergh, E., Overheul, G., Eleveld, M., Kurver, L., van Heerbeek, N., van Laarhoven, A., Miesen, P., den Hartog, G., de Jonge, M., and van Rij, R. (2021). Berberine and Obatoclax Inhibit SARS-Cov-2 Replication in Primary Human Nasal Epithelial Cells In Vitro. Viruses 13(2): 282.
Walls, A.C., Tortorici, M.A., Snijder, J., Xiong, X., Bosch, B.-J., Rey, F.A., and Veesler, D. (2017). Tectonic conformational changes of a coronavirus spike glycoprotein promote membrane fusion. Proc. Natl. Acad Sci. USA 114(42): 11157–11162.
Wan, Y., Shang, J., Graham, R., Baric, R.S., Li, F., and Gallagher, T. (2020). Receptor Recognition by the Novel Coronavirus from Wuhan: an Analysis Based on Decade-Long Structural Studies of SARS Coronavi-rus. J. Virol. 94(7): e00127-00120.
Wu, C., Liu, Y., Yang, Y., Zhang, P., Zhong, W., Wang, Y., Wang, Q., Xu, Y., Li, M., Li, X., Zheng, M., Chen, L., and Li, H. (2020). Analysis of therapeu-tic targets for SARS-CoV-2 and discovery of potential drugs by com-putational methods. Acta Pharmaceutica Sinica B 10(5): 766–788.
Xu, H., Liu, B., Xiao, Z., Zhou, M., Ge, L., Jia, F., Liu, Y., Jin, H., Zhu, X., Gao, J., Akhtar, J., Xiang, B., Tan, K., and Wang, G. (2021). Computational and Experimental Studies Reveal That Thymoquinone Blocks the Entry of Coronaviruses Into In Vitro Cells. Infect. Dis. Ther. 10(1): 483–494.
Yang, M., Wei, J., Huang, T., Lei, L., Shen, C., Lai, J., Yang, M., Liu, L., Yang, Y., Liu, G., and Liu, Y. (2021). Resveratrol inhibits the replication of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) in cul-tured Vero cells. Phytother. Res. 35(3): 1127–1129.
Yi, L., Li, Z., Yuan, K., Qu, X., Chen, J., Wang, G., Zhang, H., Luo, H., Zhu, L., Jiang, P., Chen, L., Shen, Y., Luo, M., Zuo, G., Hu, J., Duan, D., Nie, Y., Shi, X., Wang, W., Han, Y., Li, T., Liu, Y., Ding, M., Deng, H., and Xu, X. (2004). Small Molecules Blocking the Entry of Severe Acute Respiratory Syndrome Coronavirus into Host Cells. J. Virol. 78(20): 11334–11339.
Yu, M.-S., Lee, J., Lee, J.M., Kim, Y., Chin, Y.-W., Jee, J.-G., Keum, Y.-S., and Jeong, Y.-J. (2012). Identification of myricetin and scutellarein as novel chemical inhibitors of the SARS coronavirus helicase, nsP13. Bioorg. Med. Chem. Lett. 22(12): 4049–4054.
Zaki, A.M., van Boheemen, S., Bestebroer, T.M., Osterhaus, A.D.M.E., and Fouchier, R.A.M. (2012). Isolation of a Novel Coronavirus from a Man with Pneumonia in Saudi Arabia. N. Engl. J. Med. 367(19): 1814–1820.
Zhou, P., Yang, X.-L., Wang, X.-G., Hu, B., Zhang, L., Zhang, W., Si, H.-R., Zhu, Y., Li, B., Huang, C.-L., Chen, H.-D., Chen, J., Luo, Y., Guo, H., Jiang, R.-D., Liu, M.-Q., Chen, Y., Shen, X.-R., Wang, X., Zheng, X.-S., Zhao, K., Chen, Q.-J., Deng, F., Liu, L.-L., Yan, B., Zhan, F.-X., Wang, Y.-Y., Xiao, G.-F., and Shi, Z.-L. (2020). A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 579(7798): 270–273.

Copyright: © 2021 International Society for Nutraceuticals and Functional Foods.All rights reserved.
29
Review J. Food Bioact. 2021;15:29–38
Journal ofFood Bioactives International Society for
Nutraceuticals and Functional Foods
Impact of dietary nutrients (functional foods/nutraceuticals) and mi-cronutrients on COVID-19: a review
Yueching Wonga, Chi-Ho Chanb, Kamesh Venkatakrishnana, Hui-Fang Chiuc, You-Cheng Shend, Oksana Glovinskaiaa, Yi-Chun Hane and Chin-Kun Wanga*
aDepartment of Nutrition, Chung Shan Medical University, 110, Sec. 1, Jianguo North Road, Taichung City, Taiwan, ROCbDepartment of Medical Research, Chung Shan Medical University Hospital, Department of Microbiology and Immunology, Chung Shan Medical University, 110, Sec. 1, Jianguo North Road, Taichung City, Taiwan, ROC
cDepartment of Chinese Medicine, Taichung Hospital Ministry of Health and Well-being, Taichung, Taiwan, ROCdDepartment of Health Industry Technology Management, Chung Shan Medical University, 110, Sec. 1, Jianguo North Road, Taichung City, Taiwan, ROC
eDepartment of nutrition and health science, Fooyin University, Kaohsiung, Taiwan, ROC*Corresponding author: Chin-Kun Wang, Department of Nutrition, Chung Shan Medical University, 110, Sec. 1, Jianguo North Road, Taichung City, Taiwan, ROC. Tel: +886 4 22653397; Fax: +886 4 22654529; E-mail: [email protected]: 10.31665/JFB.2021.15280Received: September 06, 2021; Revised received & accepted: September 29, 2021Citation: Wong, Y., Chan, C.-H., Venkatakrishnan, K., Chiu, H.-F., Shen, Y.-C., Glovinskaia, O., Han, Y.-C., and Wang, C.-K. (2021). Im-pact of dietary nutrients (functional foods/nutraceuticals) and micronutrients on COVID-19: a review. J. Food Bioact. 15: 29–38.
Abstract
The world health organization (WHO) announced that coronavirus 2019 (COVID-19) is a pandemic. Considering the pandemic spread of Covid-19, many researchers have shown immense interest in various functional foods/nutraceuticals and dietary supplements to improve immune function and overall health status and thus lower the risk of COVID-19 infection. Studies have indicated that a balanced diet rich in various nutrients especially micro-nutrients plays a huge role in recovering and preventing COVID-19 related health issues. Improvement of obesity could reduce the risk of COVID-19 infection owing to the less angiotensin-conversion enzyme 2 receptors and in-crease the effect of vaccination. This review signifies the importance of various food bioactives along with dietary supplements (balanced diet) rich in various micronutrients against COVID-19 and its related anomalies. Moreover, this contribution helps non-specialists to understand the importance of various functional foods/nutraceuticals based on the dietary/supplementation recommendations indicated by various popular nutritional or dietary or health organizations. To sum up this contribution, healthy life style and balanced nutrition play important role on immune system. Reducing obesity becomes a critical point on the COVID-19 infection. The most important is to find the potential and effective food bioactives, which might act as supportive or complementary therapy (pro-phylactic) for mitigating the risk and comorbidity associated with COVID-19.
Keywords: Coronavirus 2019 (COVID-19); Functional foods; Nutraceuticals; Dietary supplements; Micronutrients.
1. Introduction
Severe acute respiratory syndrome coronavirus 2 (SARS-CoV2) or coronavirus causes a pneumonia-like respiratory disease, which is commonly called COVID-19 (Coronavirus-2019). Coronavi-rus is a positive sense single standard RNA (ssRNA), which is
enveloped by a spike glycoprotein, which appears like a crown under a 2D-electron microscope, and hence it is generally called a coronavirus (Iddir et al., 2020). Coronavirus is a member of the coronaviridae family and has a close connection with severe acute respiratory syndrome (SARS) and Middle East respiratory syndrome (MERS). Coronavirus structure consists of four major

Journal of Food Bioactives | www.isnff-jfb.com30
Impact of dietary nutrients and micronutrients on COVID-19: a review Wong et al.
proteins including spike glycoprotein, nucleoprotein (attached with RNA), membrane protein, and an envelope protein, which plays a key role in replication (division). SARS-CoV2 can use angiotensin-converting enzyme 2 (ACE2) as a receptor and enter host cells and start to divide and finally cause COVID-19 (Mrit-yunjaya et al., 2020; Wiersinga et al., 2020). Concomitant Obesity and related health issues become the targets of COVID-19 infec-tion and indicator of severe syndrome, and the efficacy of vaccina-tion is suppressed by obesity (Goossens et al., 2020; Bakhshandeh et al., 2021). The novel coronavirus (SARS-CoV2) outbreak from Wuhan, China has inflicted a global emergency health threat to humanity since December 2019. This coronavirus pandemic has impacted the economy and health care system all over the world (Kozlov et al., 2021). This coronavirus spreads from human to hu-man via respiratory and airborne droplets as well as through di-rect contacts. The biggest threat of the coronavirus is that most of the infected patients are asymptomatic and few develop severe symptoms like fever, dry cough, fatigue loss of smell/taste, and headache. If COVID-19 is not treated properly it can result in high mortality and morbidity and also its long-term effects are not clear (Kazemi et al., 2021; Syed, 2020). Especially, elderly people, peo-ple with comorbid conditions (Cardiovascular, diabetes mellitus, cancer, and hypertension), immunosuppressed subjects, and heavy smokers are highly susceptible to Coronavirus infection (Subedi et al., 2021). Recently, WHO has announced four major variants/genera of COVID-19 virus including α, β, γ, δ till date (out of 11 known variants). Out of all these coronavirus variants the δ (delta) is one the deadliest with a high transmissibility rate (Reproductive number) and contributed to high mortality and morbidity globally in recent times (Alizon et al., 2021; Liu et al., 2021).
2. An overview on the pathophysiology of COVID-19
The detailed pathophysiology of COVID-19 is still being explored. However, few major risk factors like oxidative stress, inflamma-tion, and altered immune response are assumed to be the major contributor to COVID-19 and its related health issues (Kozlov et al., 2021; Iddir et al., 2020). In this section, we would like to dis-cuss the connection between oxidative stress, inflammation, and immune response and how they contribute to COVID-19. Once the SARS-CoV2 enters host cells (mostly lung cells) via ACE2 receptor by endocytosis, it starts to replicate and affects the cel-lular homeostasis, resulting in oxidative stress (increased free radi-cal production and reduced antioxidant activity), which inflicts an imbalance between free radicals and antioxidants. These excessive free radicals (reactive oxygen species [ROS]/reactive nitrogen spe-cies [RNS]) could alter various signaling pathways that are respon-sible for various host cellular metabolic processes (Hussain et al., 2016). These excessive ROS/RNS triggers inflammatory response via toll-like receptor (TLR) and nuclear factor-kappa B (NF-κB) signaling pathway by increasing the production of various pro-inflammatory cytokines. The excessive production of inflamma-tory cytokines results in hyperinflammatory condition (cytokines storm) and also alters the release and activation of various immune cells especially monocytes, neutrophils, and macrophages (Wiers-inga et al., 2020; Leisman et al., 2020). These activated immune cells further increase the production of free radicals by suppressing the nuclear factor erythroid related factor 2 (Nrf2) signaling path-way, which is responsible for endogenous antioxidant (cytoprotec-tion) and detoxification related enzymes (Laforge et al., 2020). In addition, elevated cytokines trigger the induction of endothelium HA-Synthase-2 (HAS2) and subsequently increase the production
of hyaluronan (HA), which has a high water binding capacity and eventually results in increased accumulation of lung fluid (sepsis). As mentioned before SARS-CoV2 mostly enters the host system through the respiratory tract and hence lungs cells are highly af-fected by oxidative stress, inflammation, and altered immune re-sponse and result in fibrosis, fluid accumulation, and sepsis (Shi et al., 2020). Hence, most of COVID-19 patients have various lung or respiratory-related problems especially acute respiratory distress syndrome (ARDS), and pneumonia (Kellner et al., 2017). Both ox-idative stress and inflammation are strongly interconnected in that one activates another (vicious cycle). Similarly, oxidative stress and inflammation result in altered immune response (innate and acquired immunity) which plays a dominant role in the pathophys-iology of COVID-19 (Leisman et al., 2020). Furthermore, stud-ies have indicated that elevated ROS production (oxidative burs/stress), cytokine storm (inflammation), immune dysfunction end up in multi-organ failure by altering various normal cellular func-tions like autophagy, apoptosis, necrosis (cell death), cell energy system (mitochondrial dysfunction) along with altered microbiota flora (microbiota dysbiosis) could increase the severity of SARS-CoV2 infection (Yuki et al., 2020; Smith et al., 2020). Because, of the above-mentioned mechanism (pathophysiology), COVID-19 patients start to develop various comorbid conditions which even-tually leads to higher mortality and morbidity. Those individuals who already have comorbid conditions like hypertension, diabetes mellitus, cardiovascular or renal/lung-related disorders are highly vulnerable to high morbidity and mortality (Liu et al., 2020).
3. Modified lifestyle pattern to lower the risk of COVID-19
So far, we have given an overview of COVID-19 and its impacts on humans by explaining various pathophysiological events and their associated factors. We would now like to emphasize the im-portance of a modified lifestyle pattern for reducing the risk of COVID-19 as well as some recommendations for a speedy recov-ery from COVID-19. The common recommendation from various expert includes regular exercise (indoor or outdoor), maintaining social distancing, wearing a mask and washing hands (hygienic lifestyle) as well as limiting smoking and alcohol consumption (Ismail et al., 2021; WHO, 2020). In addition, consumption of healthy foods including regular intake of fruits, vegetables, dietary fibers, nuts, and cereals along with various dietary supplements would lower the risk of COVID-19. Even though various COV-ID-19 vaccines are available in the market, but it does not provide 100% protection against COVID-19, as well as its long-term ad-verse effects are largely unknown (Subedi et al., 2021; Cascella et al., 2021). Moreover, the vaccine can only help to improve or boost the immune response against coronavirus (produce antibodies). However, the individual immune system should be strong enough to produce enough antibodies (plasma/memory cells) to fight against the coronavirus (Ahn et al., 2020). Hence, consumption of a healthy balanced diet would help to boost the host immune system and thereby mitigate the harmful health impacts associated with coronavirus. Along with vaccination, few anti-viral drugs and micronutrients are recommended for treating Covid-19 but follow-ing a healthy lifestyle (healthy food) would be the best choice to help combating this pandemic (Lordan et al., 2021).
One very important issue is obesity control. Obesity and related health issues (hypertension, diabetes and coronary heart diseases) are well known as main reasons of infection and severe syndrome of COVID-19 and vaccination efficacy owing to rich ACE2 receptor (Goossens et al., 2020; Bakhshandeh et al., 2021). ACE2 receptor

Journal of Food Bioactives | www.isnff-jfb.com 31
Wong et al. Impact of dietary nutrients and micronutrients on COVID-19: a review
is a protein on the surface of many cell types. It is an enzyme that generates small proteins, by cutting the larger protein angiotensino-gen, that then go on to regulate functions in the cell. To prevent the infection and avoid severe syndrome of COVID-19 infection, con-comitant obesity control is very critical and effective. Energy intake control and physical activity (healthy lifestyle) could be greatly en-couraged at this time. Even though many strategies until now have been recommended to treat COVOD-19, but the way to help treating this disease is important. Food bioactives are treasures from nature which show great potential to treat COVID-19 problems. Basic studies together with clinical trials are required in the near future. The authors have set up a good model to screen a lot of potential and effective food bioactives on COVID-19 virus, some valuable and exciting results have so far been obtained. Our next step is clinical trial for getting the science-based evidence for them.
4. Dietary guidelines from health/ dietary organization/soci-ety/council to prevent COVID-19
Different nutritional or dietary or health organizations/societies/
councils, including World Health Organization (WHO), Food and Agriculture Organization (FAO) of the UN, and European Food In-formation Council (EFC), Center for Disease Control and Preven-tion (CDC), Dietary Association of Australia (DAA) and United National Children’s Fund (UNICEF) have proposed various health/dietary guidelines and advice pertaining to COVID-19 pandemic. Table 1 epitomizes the summary of health/dietary guidelines and advice indicated by various organizations or councils which most-ly includes the consumption of fruits, vegetables, fish oil, probi-otics, whole grains, cereals, and nuts. These recommended foods could substantially improve the immune system and thereby elicit overall protection from COVID-19 infection and lower the risk of Covid related health issues as well as improve the recovery rate in Covid infected subjects (WHO, 2020; UNICEF, 2020; FAO, 2020; DAA, 2020; de Faria Coelho-Ravagnani et al., 2021). Overall, all major health/ dietary organizations/societies/councils have recom-mended that individuals consume healthy food including fruits and vegetables (rich in fibers, minerals, and vitamins) along with drinking an adequate level of water (hydrated). In addition, avoid-ing smoking and reducing alcohol consumption have been sug-gested (de Faria Coelho-Ravagnani et al., 2021).
Table 1. Summary of dietary/supplementation recommendations and proposed health benefits along with food hygienic guidelines from different nu-tritional or dietary or health organizations/societies/councils
Dietary/supplementation recommendation Proposed health benefits and hy-gienic guidelines (food safety)
Organization/As-sociation/Council
Intake of fresh fruits and vegetables rich in fibers, minerals, and vitamins (Colored fruits and vegetables are highly recommended). Avoid or limit the use of processed or canned food. Lower the consumption of sugar, fat, or salt. Keep hydrated (drink enough water). Do some indoor exercises, yoga, and meditation
Consumption of fruits and vegetables can improve overall health status especially lower oxidative stress and enhance immune system and thus lowering the risk of Covid 19. Wash hands before handling food (before eating or cooking). Wash vegetables/fruits before cooking. Maintain social distancing and wear a mask (outdoor activities)
WHO (2020)
Increase fruits and vegetable consumption. Intake fish oil rich in DHA/EPA. Vitamin and mineral-rich supplements or fruits/vegetables must be consumed. Drink a glass of milk and take only low-fat dairy products are better (especially probiotics). Lower the usage of processed and junk foods. Make cooking and eating fun, makes children happy, and lower stress
Consumption of fruits and vegetables rich in vitamins and minerals could decrease stress and enhance the endurance and immune response (increase antibody production) and thus mitigate the severity of Coronavirus infection. Wash hands before handling food (before eating or cooking). Pregnant women with Covid 19 can still breastfeed. Children should maintain social distancing and wear a mask (outdoor activities)
UNICEF (2020)
Consume a healthy and balanced diet by including vegetables, nuts (healthy fat), whole grains, and fruits. Diversity food consumption and do not avoid any food completely, to balance your system. Limit alcohol consumption. Avoid excessive intake of fats or sugar
Eating healthy and balanced food will significantly improve the immune system (proliferation of T and B cells). Dietary fibers and healthy fat can lower the comorbid conditions associated with Covid 19. Wash all the food materials before cooking. Wash hands regularly
FAO (2020)
Prefer fresh food and try to avoid stored or canned food especially fruits and vegetables. Consume dried nuts like walnut, almond, cashews, peanuts. Intake whole grains/cereals and drink at least a glass of milk. Try to consume herbs and spices with your food. But avoid excessive salt or sugar (carbonated drinks)
Following the dietary plan, it can help maintaining a strong immune function as well as substantially improve your mental health. Sterilize the food before cooking and wash hand with soap for at least 20 seconds
DAA (2020)
Recommended to control the disease by following only hygienic procedure (food safety), no dietary recommendation
Wash your hand regularly for at least 20 seconds. Clean/sanitize the working space and kitchen to avoid food contamination. Refrigerate perishable food and clean than before cooking. Wear a mask as much as possible and maintain social distancing
CDC (2020)

Journal of Food Bioactives | www.isnff-jfb.com32
Impact of dietary nutrients and micronutrients on COVID-19: a review Wong et al.
5. Importance of functional food/nutraceuticals and dietary supplements on COVID-19
Considering this coronavirus pandemic outbreak, people are be-coming highly health-conscious and have started to focus on healthy food. When it comes to healthy food, people mostly re-member functional foods/nutraceuticals, herbs, and dietary supple-ments as they might be the better tool to combat against COVID-19 and its related health issues (Subedi et al., 2021). Both functional foods and nutraceuticals terms are often used interchangeably but there are subtle differences between them. In the case of functional food, they are considered as a traditional or conventional food or food byproduct, which could be fortified or altered to enhance its nutritional value and thus demonstrably enhance its overall bio-logical function and health status (Venkatakrishnan et al., 2019; Shahidi, 2012). However, nutraceuticals are part of the food (bio-active) which can be used in the medicinal form or supplement to improve overall health status (Daliu et al., 2019). Recent studies have indicated that intake of functional foods/nutraceuticals and dietary supplements rich in various micronutrients would lower the risk of COVID-19 infection as well as speeding up the recov-ery in COVID-19 subjects since most viral infections can compro-mise immune function and lower micronutrient levels (especially vitamins/minerals). Moreover, functional foods/nutraceuticals and dietary supplements are well known for their antioxidant, anti-in-flammatory, and immunomodulatory properties. Hence, they can effectively lower oxidative stress and inflammation and thereby positively regulate immune cells (immune booster) which even-tually results in improving overall health status (Subedi et al., 2021; Lordan et al., 2021; Galanakis et al., 2020; Mrityunjaya et al., 2020). Both vitamins and minerals (micronutrients) play a cen-tral role in various cellular activities (metabolic function, immune function, antioxidant, anti-inflammation), structural/body devel-opment (growth), and thereby improve overall health status (Chiu et al., 2021). In this contribution, we havechosen only the popular nutraceuticals/functional food or micronutrients (dietary supple-ments) like probiotics, fish oil, and curcumin, and quercetin which have been reported to show potent anti-viral and anti-respiratory effects. Most of the clinical trials are still ongoing and only a few trials have come up with positive results, thus, it very hard to make any definite conclusions. Hence, we have referred to the previous data of various nutraceuticals/functional foods or micronutrients against viral infection which are related to respiratory disorders like lung sepsis, pneumonia, ARDS, SARS, and MERS, might be also effective against SARS-CoV2 or COVID-19 injection.
6. Functional food/nutraceuticals against COVID-19
6.1. Probiotics (pre and post biotics)
Probiotics are living microorganisms in the gut, which are believed to improve host health status, whereas, prebiotics are defined as non-digestible food acting as a food material that stimulates the growth of host beneficial gut microorganism (probiotics). Simi-larly, postbiotics are fermented by-products (of prebiotics) pro-duced in the gut that help promoting health (Zolkiewicz et al., 2020). The popular probiotics species include Lactobacillus and Bifidobacterium spp., streptococcus (Beneficial bacteria’s) which are commonly used in various commercial food and supplement products to enhance the composition of gut microbiota and hence used as functional foods/nutraceuticals (Chiu et al., 2021). Pro-
and prebiotics also show various biological functions including anti-inflammatory, antioxidant, and immunomodulatory activity through modulating the gut-brain axis (Pandey et al., 2015). The proposed mechanism of pro- and prebiotics against COVID-19 include reduced expression of inflammatory markers by inhibit-ing TLR 2/4, NF-κB, and TNF-α signaling pathway, improving intestinal barrier function, and thus lowering the infiltration of inflammatory and immune cells (Akour, 2020). Various probiotic spp include Lactobacillus and Bifidobacterium which play a key role in clearing the influenza virus by inhibiting the attachment with the receptor (Baud et al., 2020). Likewise, previous studies also indicate that probiotics can inhibit the transmissible gastro-enteritis virus from entering the epithelial cells and thus protect the upper respiratory and gastric tract from viral infection (Wang et al., 2017). Probiotics are reported to modulate immune function (innate immunity) by altering the production of NK and APCs cells (Chong et al., 2019; Mortaz et al., 2013). Clinical trials conducted by Zeng (2016) and Morrow et al. (2010) demonstrated that sup-plementation with prebiotics in respiratory ill subjects on ventila-tion showed less ventilator-associated pneumonia compared with the placebo group. In addition, several reports indicate that pre-/pro- and postbiotics can be beneficial against COVID-19 infection and can lower the severity owing to their potent anti-inflammatory, antioxidant, anti-viral, and immune-modulatory properties via im-proving gut microbiota, which in turn enhance the gut-brain axis and gut-lung axis (Baud et al., 2020; Sundararaman et al., 2020; Akour, 2020). Based on the above data, probiotic/prebiotic supple-mentation may be used as prophylactic and supportive therapeu-tic property (used along with standard drugs) against COVID-19. However, more trials are needed before confirming their definite effect against the coronavirus.
6.2. Fish oil (DHA/EPA)
Fish oil is rich in various polyunsaturated fatty acids (PUFA) which are comprised of fatty acids with multiple double bonds. These include alpha-linolenic acid (ALA), docosahexaenoic acid (DHA) and eicosapentaenoic acid (EPA) which are classified as Omega(ω)-3-fatty acids with an array of pharmaceutical proper-ties including antioxidant and anti-inflammatory activities (Ven-katakrishnan et al., 2019). The proposed mechanism of fish oil against COVID-19, especially DHA and EPA, is by exerting po-tent anti-inflammatory activity via suppressing the production of pro-inflammatory cytokines and Cox-2, and hence can attenuate cytokines Strom process (Szabo et al., 2020). Fish oil also acts as an ACE2 inhibitor and hence acts as an anti-corona viral agent. Adaptive and innate immunity activity of fish oil was demonstrat-ed by altering the production of NK and neutrophil immune cells (Calder, 2007). Fish oil rich in DHA and EPA showed anti-platelet and anti-thrombotic/anti-atherogenic activity and thus used to treat COVID-19 and its related complications (Rogero et al., 2020). A preliminary trial showed that blood samples from 100 COVID-19 patients with a higher DHA/EPA index had a lower risk of death due to Covid-19 (Asher et al., 2021). Even though fish oil was found to be beneficial against COVID-19, but the limited clinical trials make it difficult to recommend fish oil for treating COV-ID-19.
6.3. Curcumin
Curcumin is one of the key active ingredients of turmeric. Tur-meric is commonly used in Asian cuisine especially in India as

Journal of Food Bioactives | www.isnff-jfb.com 33
Wong et al. Impact of dietary nutrients and micronutrients on COVID-19: a review
a flavoring/coloring agent with numerous biological functions, and hence it is considered a popular nutraceutical/functional food ingredient (Chiu et al., 2021). The proposed mechanism of cur-cumin against COVID-19 is by exerting antiviral effects against Zika virus, Influenza virus, Human papillomavirus (HPV), and Human immunodeficiency virus (HIV) through inhibiting viral at-tachment and replication (Chen et al., 2013). In addition, curcumin can suppress ACE2 binding and thus inhibit or control SARS-CoV virus entry (Zahedipour et al., 2020). The available data demon-strate that curcumin can also work effectively against SARS-CoV (coronavirus). Curcumin has been reported to lower antioxidant stress via enhancing antioxidant activity (scavenge free radicals/ anti-lipid peroxidative function) as well as suppressing the produc-tion of inflammatory cytokines via inhibiting NF-κB, STAT, and COX-2 signaling pathways (Rocha et al., 2020). It also stimulates host interferon production and thus trigger innate immunity (Ha-mid and Thakur 2021).
6.4. Quercetin
Quercetin is a flavonoid (flavanol) and is usually present in veg-etables and fruits. Quercetin is available in both aglycone and gly-cosidic form (rutin) and has various biological functions including antioxidant, anti-inflammatory, ant-viral, and immunomodula-tory activities (Venkatakrishnan et al., 2019). The proposed action mechanism of quercetin against COVID-19 is displayed by its strong anti-inflammatory activity and by inhibiting the production of various pro-inflammatory cytokines and inflammatory markers like COX-2 and LOX (Bhasker et al., 2011). It improves antioxi-dant status by upregulating the Nrf2 signaling pathway (Sharma et al., 2020). Previous studies indicate that quercetin can suppress the replication of RNA viruses-like influenza and coronavirus. Quercetin is reported to inhibit SARS CoV2 (spike protein) attach-ment to ACE2 and thus interfering with the endocytosis or entry of SARS CoV2 into cells and thus act as an anti-SARS CoV2 agent (Smith and Smith, 2020). Moreover, a better anti-corona virus ef-fect was found with quercetin, when combined with vitamin C, thus demonstrating their immunomodulatory and anti-viral prop-erties. Furthermore, supplementation of vitamin C with Quercetin for 7 days was shown to improve the recovery rate (reduce hospital stay) in COVID-19 patients (Colunga Biancatelli et al., 2020).
7. Micronutrients against COVID-19
7.1. Vitamins
Vitamins are essential micronutrients, which are classified as wa-ter-soluble (B, C) and lipid-soluble (A, D, E, K). All the vitamins are involved in various biological functions including growth and development. Especially vitamins A, C, D, and E play a critical role in maintaining the immune system along with their antioxi-dant, anti-inflammatory and anti-viral activities which make them a special weapon in combating various upper respiratory illnesses. Hence vitamins A, C, D, and E might also work effectively against SARS- CoV2 or coronavirus (Dehghani-Samani et al., 2020; Jovic et al., 2020).
7.2. Vitamin C (ascorbic acid)
Vitamin C is a water-soluble vitamin that aids to improve immune
function and is also involved in the development of body tissue (collagen formation) and speeds up the wound healing process. In addition, vitamin C has potent antioxidant, anti-inflammatory, and anti-viral properties, which might improve overall health status and thus helps in the management of Covid and its related complica-tions like sepsis and pneumonia (Carr and Rowe, 2020). The possi-ble mechanism of vitamin C against COVID-19 includes modula-tion of the activation of various immune cells (improve immunity) and thereby regulating the production of various inflammatory markers (suppress cytokine storm) and decreasing the histamine levels (Calder, 2020). Vitamin C can increase the differentiation and proliferation of B and T Cells and thus aid in the production of antibodies against SARS- CoV2 (Abobaker et al., 2020). Vitamin C has also been reported to enhance various antioxidant activities via upregulating the Nrf2 signaling pathway (Cheng, 2020) and can reduce the severity of upper respiratory viral infections like ARDS (Hagel et al., 2013). A pilot trial conducted by Zhang et al. (2021) has hinted that administration of vitamin c for 7 days in 56 ill covid subjects resulted in a significant reduction of hospital stay and mortality. Moreover, supplementation of vitamin C with quercetin for 7 days improved the recovery rate (reduce hospital stay) in COVID-19 patients (Colunga Biancatelli et al., 2020). Overall, vitamin C, owing to its potent antioxidant, anti-inflamma-tory, anti-viral, and immune-boosting properties, might help treat-ing COVID-19 patients, however, more evidence is needed before its recommendation for treating COVID-19.
7.3. Vitamin D or 25-hydroxyvitamin D
Vitamin D is a fat-soluble vitamin that plays an important role in several physiological processes, such as bone metabolism, calci-um, and phosphorus absorption, and enhance cellular immunity. Vitamin D is also showing promising results against various upper respiratory disorders or syndrome like ARDS. A mounting num-ber of studies demonstrate that deficiency of vitamin D (hypovi-taminosis D) is strongly associated with an increase in the inci-dences of COVID-19 and its severity (Liu et al., 2021; Pereira et al., 2020; Grant et al., 2020). However, few studies have shown no association between vitamin D and Coronavirus (Jevalikar et al., 2021; Murai et al., 2021). The proposed mechanism against COVID-19 includes intake of vitamin D that could lower inflam-matory cytokines production (cytokines storm) via downregulat-ing the NF-κB signaling pathway. Vitamin D could effectively modulate immune function by altering the activation of various immune cellsthus regulating the production of cytokines as well as increasing the production of antibodies against coronavirus (Mrit-yunjaya et al., 2020; Barlow et al., 2011). In addition, vitamin D is reported to induce cathelicidin (LL-37) and related peptides/pro-tein and thus promoting innate immunity and eventually lowering the replication of SARS-CoV2 and influenza virus (Kazemi et al., 2021; Sharifi et al., 2019). In addition, vitamin D is involved in the regulation of renin-angiotensin-aldosterone system (RAAS) and angiotensin-converting enzyme 2 (ACE2), which are responsible for attachment of SARS CoV2 and COVID-19 infection (Pal et al., 2021). Vitamin D could upregulate the expression Nrf2 signaling pathway and thereby facilitating the balanced mitochondrial func-tion (Mrityunjaya et al., 2020). A meta-analysis conducted by Mar-tineau et al., (2017) found that vitamin D administration reduces the risk of upper respiratory infection. Pal et al. (2021) indicated that vitamin D supplementation (safe) significantly improved vari-ous clinical outcomes like a lower hospital stay, mortality, adverse events, and speedy recovery. A systemic review and meta-analysis conducted by Kazemi et al (2021) also concluded that a significant

Journal of Food Bioactives | www.isnff-jfb.com34
Impact of dietary nutrients and micronutrients on COVID-19: a review Wong et al.
relationship exists between vitamin D and Covid infection, sever-ity, and mortality. Especially vitamin D deficiency (VDD) has a direct association with Coronavirus infection and mortality. Even though vitamin D showed a promising effect against SARS-CoV2, but the dose, mode of administration, duration, and combination with other drugs or nutrients remains largely unknown, and hence further studies are needed before making any recommendations related to Covid patients.
7.4. Vitamin A
Vitamin A exhibits potent anti-inflammatory and immunomodu-latory activities via regulating the cellular and humoral immune response. The possible mechanism underlying vitamin A protec-tion against COVID-19, includes elevated production of T cells, NK cells, and neutrophils (Tanumihardjo et al., 2016). Previously, few researchers indicated that supplementation of vitamin A could significantly reduce the onset of various viral diseases like HIV, HPV as well as decreasing the mortality and morbidity associated with various respiratory illnesses and viral infections (Hamid and Thakur 2021; Villamor and Fawzi, 2005).
7.5. Vitamin E
Vitamin E is a potent antioxidant and plays a crucial role in main-taining immune function especially for the elderly. Vitamin E is also involved in T and B cell proliferation and differentiation as well as improving NK cell and APCs activity and thus regulating immune response (Junaid et al., 2020; Lee and Han, 2018). Vitamin E is a strong anti-inflammatory agent as it suppresses the produc-tion of pro-inflammatory cytokines (Lewis et al., 2019). Hence, it may be used to treat Covid related hyperinflammatory conditions along with its antioxidant and immunomodulatory activities. The evidence of vitamin E against COVID-19 is limited, but it might be used with other essential micronutrients as supportive therapy to enhance immunity. Moreover, vitamin B complex (B6, B9, B12) also helps to improve immune function by enhancing the produc-tion of NK cells (innate immunity), T and B cells proliferation to enhance the antibodies production against coronavirus. However, only limited studies have been conducted on this topic so far and hence the association of vitamin B complex with COVID-19 is weak.
7.6. Minerals
Minerals are essential micronutrients that show a broad spectrum of functions including cellular metabolism, growth and thereby regulating overall health status (Chiu et al., 2021). Few miner-als like zinc (Zn), and selenium (Se), are reported to be effective against COVID-19, but other minerals like magnesium (Mg), cal-cium (Ca), iron (Fe), and copper (Cu) also show mild to moderate effect against COVID-19.
7.7. Zinc (Zn)
Zinc is one of the second most abundant minerals (trace element) used for growth, brain development, bone formation, and wound healing. The proposed mechanism of Zn against COVID-19, in-cludes lowering the production, proliferation, and differentiation of various immune cells like NK cells, APCs, neutrophils and
thus contributing to both innate and acquired immunity (Kumar et al., 2020). Zinc can also regulate the viral entry, fusion/attach-ment (endocytosis), and replication and translation (growth/pro-liferation) of various upper respiratory viruses (Ishida, 2019). Zn is reported to reduce the recombinant human ACE2 activity and may be helpful for treating COVID-19 (Speth et al., 2014). Studies show that chloroquine along with Zn supplementation may show a better anti-corona virus activity by inhibiting the attachment and subsequent multiplication of the SARS-CoV2 virus (Pal et al., 2020; Ishida, 2019), however, this subject is somewhat controver-sial, especially in light of false claims related to hydroxychloro-quine. In addition, Zn is reported to lower the production of pro-in-flammatory cytokines via inhibiting the TLR and NFkB signaling pathways (Hamid and Thakur 2021). A clinical trial, conducted by Prasad et al. (2007) has shown that zinc supplementation in elderly subjects for one year could significantly decrease the incidence of viral infection through lowering of cytokines production and oxi-dative stress. Their data show that low levels of Zn (Zn deficiency) are associated with high susceptibility of viral infection especially COVID-19. The author strongly suggested that all subjects (es-pecially elderly people) maintain the normal Zn levels and in this way lower the risk of Covid infection and its severity.
7.8. Selenium (Se)
Selenium is an essential micronutrient used for various biological functions in the body, especially for immune regulation, anti-in-flammation, and antioxidant properties. Se is usually incorporated as selenoprotein and used by cells for various biological functions especially for lowering oxidative stress (Chiu et al., 2021). The proposed mechanism of Se against COVID-19 includes possible enhancement of glutathione family antioxidants, and thus effec-tively lowering oxidative stress. Se can promote the production and proliferation of NK cells and thus play a major role in innate immunity (Avery and Hoffmann, 2018). Selenium administration can also downregulate various pro-inflammatory cytokines pro-duction (Mrityunjaya et al., 2020). Previous studies have indicated that Se supplementation could significantly lower the inflamma-tory response in respiratory distress syndrome patients (ARDS) as well as lowering the production of various pro-inflammatory cytokines (Mahmoodpoor et al., 2019). The above-mentioned an-tioxidant, anti-inflammatory, and immune-modulatory (innate im-munity) activities of Se recommend itas a good choice for fighting against COVID-19 and its related complications.
Other minor minerals like iron (Fe), and copper (Cu) have also been found to show some positive impact against COVID-19 in-fection and severity. Iron supplementation showed lower inflam-matory cytokines and altered immune cell activation as well as oxidative status (Wu et al., 2019). Similarly, copper intake was linked with antioxidant production and thus lowering oxidative stress in Covid infected subjects. In addition, Cu increased the pro-duction and proliferation of T and B cells, which contributed to antibodies production against the Coronavirus as well as killing several viruses like HIV, poliovirus and could inhibit the coronavi-rus infection (Raha et al., 2020).
7.9. Combination therapy
A few trials have been conducted by combining various dietary supplements (micronutrients) with functional foods/nutraceuticals due to their perceived holistic biological functions. However, the dosage, route of administration, duration, and combination ratio

Journal of Food Bioactives | www.isnff-jfb.com 35
Wong et al. Impact of dietary nutrients and micronutrients on COVID-19: a review
remain to pose the biggest challenge (Colunga Biancatelli et al., 2020; Mittra et al., 2020). Likewise, dietary supplements (micro-nutrients), functional foods/nutraceuticals are combined with other standard anti-viral or anti-inflammatory drugs (synergistic effect). However again, the dose, duration, and combination ratio is an is-sue (Srivastava et al., 2021; Subedi et al, 2021). Figure 1 repre-sents the overview of this review, highlighting the importance of dietary supplements (micronutrients), functional foods/nutraceuti-cals in improving health and immune system for use along with standard drugs and vaccines.
8. Conclusion
This coronavirus pandemic has challenged the entire health system globally and iys economy and has also killed several millions in the last few months. Here, we briefly provided the overall pathophysi-ology and its inter-connection and its subsequent consequences. The importance of hygienic lifestyle, body weight control and food habits to lower the severity of coronavirus infection were iden-tified as important factors in relation to COVID-19. Finally,the importance of functional foods/nutraceuticals and dietary supple-ments (micronutrients) and how they may play a holistic function in improving overall health status in covid patients through sup-pressing oxidative stress, inflammation and positively regulating the immune response was reviewed. Currently limited number of clinical trials are available to support the importance of these vari-ous nutraceuticals/functional foods or dietary supplements against Coronavirus or Covid infection /Severity. Hence, further extensive clinical trials are needed to see if dietary supplements, functional foods/nutraceuticals along standard Covid vaccine (supportive
therapy) would lower the ill effect of COVID-19 and its associated health issues. In addition, whether the dietary supplements, func-tional foods/nutraceuticals could lower the possible adverse effects of Covid vaccine deserves attention. Overall, this review provided some glimpse on the impact of various potential food bioactives against COVID-19 and their therapeutic effects and proposed pos-sible mechanisms of action.
References
Abobaker, A., Alzwi, A., and Alraied, A.H.A. (2020). Overview of the pos-sible role of vitamin C in management of COVID-19. Pharmacol. Rep. 72(6): 1517–1528.
Ahn, D.G., Shin, H.J., Kim, M.H., Lee, S., Kim, H.S., Myoung, J., Kim, B.T., and Kim, S.J. (2020). Current status of epidemiology, diagnosis, therapeu-tics, and vaccines for novel coronavirus disease 2019 (COVID-19). J. Microbiol. Biotechnol. 30(3): 313–324.
Akour, A. (2020). Probiotics and COVID-19: is there any link? Lett. Appl. Microbiol. 71(3): 229–234.
Alizon, S., Haim-Boukobza, S., Foulongne, V., Verdurme, L., Trombert-Paolantoni, S., Lecorche, E., and Sofonea, M.T. (2021). Rapid spread of the SARS-CoV-2 Delta variant in some French regions, June 2021. Euro. Surveill. 26(28): 2100573.
Asher, A., Tintle, N.L., Myers, M., Lockshon, L., Bacareza, H., and Harris, W.S. (2021). Blood omega-3 fatty acids and death from COVID-19: A pilot study. Prostaglandins, Leukotrienes Essent. Fatty Acids 166: 102250.
Avery, J.C., and Hoffmann, P.R. (2018). Selenium, selenoproteins, and im-munity. Nutrients 10(9): 1203.
Bakhshandeh, B., Sorboni, S.G., Javanmard, A.R., Mottaghi, S.S., Mehrabi, M.R., Sorouri, F., Abbasi, A., and Jahanafrooz, Z. (2021). Variants in ACE2; potential influences on virus infection and COVID-19 severity.
Figure 1. Overview of highlighting the importance of dietary supplements (micronutrients), functional foods/nutraceuticals in COVID-19.

Journal of Food Bioactives | www.isnff-jfb.com36
Impact of dietary nutrients and micronutrients on COVID-19: a review Wong et al.
Infect. Genet. Evol. 90: 104773.Barlow, P.G., Svoboda, P., Mackellar, A., Nash, A.A., York, I.A., Pohl, J., and
Donis, R.O. (2011). Antiviral activity and increased host defense against influenza infection elicited by the human cathelicidin LL-37. PLoS ONE 6(10): e25333.
Baud, D., Dimopoulou Agri, V., Gibson, G.R., Reid, G., and Giannoni, E. (2020). Using probiotics to flatten the curve of coronavirus disease COVID-2019 pandemic. Front. Public Health 8: 186.
Bhaskar, V., Yin, J., Mirza, A.M., Phan, D., Vanegas, S., Issafras, H., Michel-son, K., Hunter, J.J., and Kantak, S.S. (2011). Monoclonal antibodies targeting IL-1 beta reduce biomarkers of atherosclerosis in vitro and inhibit atherosclerotic plaque formation in Apolipoprotein E-defi-cient mice. Atherosclerosis 216(2): 313–320.
Calder, P.C. (2007). Immunomodulation by omega-3 fatty acids. Prosta-glandins, Leukotrienes Essent. Fatty Acids 77(5-6): 327–335.
Calder, P.C. (2021). Nutrition, immunity and COVID-19. Eur. J. Clin. Nutr. 75(9): 1309–1318.
Carr, A.C., and Rowe, S. (2020). The emerging role of vitamin C in the pre-vention and treatment of COVID-19. Nutrients 12(11): 3286.
Cascella, M., Rajnik, M., Aleem, A., Dulebohn, S., and Di Napoli, R. (2021). Features, evaluation, and treatment of coronavirus (COVID-19). Stat-Pearls. StatPearls Publishing, Treasure Island (FL).
CDC, Food safety and coronavirus disease 2019 (COVID-19). (2019). Cent-ers for Disease Control and Prevention website. https://www.cdc.gov/foodsafety/newsletter/food-safetyand-Coronavirus.html. Up-dated May 11: 2020. Accessed May 19: 2020.
Chen, T.Y., Chen, D.Y., Wen, H.W., Ou, J.L., Chiou, S.S., Chen, J.M., and Hsu, W.L. (2013). Inhibition of enveloped viruses infectivity by curcumin. PLoS ONE 8(5): e62482.
Cheng, R.Z. (2020). Can early and high intravenous dose of vitamin C pre-vent and treat coronavirus disease 2019 (COVID-19)? Med. Drug Dis-covery 5: 100028.
Chiu, H.F., Venkatakrishnan, K., Golovinskaia, O., and Wang, C.K. (2021). Impact of Micronutrients on Hypertension: Evidence from Clinical Trials with a Special Focus on Meta-Analysis. Nutrients 13(2): 588.
Chong, H.X., Yusoff, N.A.A., Hor, Y.Y., Lew, L.C., Jaafar, M.H., Choi, S.B., and Liong, M.T. (2019). Lactobacillus plantarum DR7 improved upper res-piratory tract infections via enhancing immune and inflammatory parameters: A randomized, double-blind, placebo-controlled study. J. Dairy Sci. 102(6): 4783–4797.
Colunga Biancatelli, R.M.L., Berrill, M., Catravas, J.D., and Marik, P.E. (2020). Quercetin and vitamin C: an experimental, synergistic ther-apy for the prevention and treatment of SARS-CoV-2 related disease (COVID-19). Front. Immunol. 11: 1451.
Colunga Biancatelli, R.M.L., Berrill, M., Catravas, J.D., and Marik, P.E. (2020). Quercetin and vitamin C: an experimental, synergistic ther-apy for the prevention and treatment of SARS-CoV-2 related disease (COVID-19). Front. Immunol. 11: 1451.
DAA. (2020). https://daa.asn.au/smart-eating-for-you/planning-your-pan-try-during-the-Covid-19-pandemic.
Daliu, P., Santini, A., and Novellino, E. (2019). From pharmaceuticals to nu-traceuticals: Bridging disease prevention and management. Expert Rev. Clin. Pharmacol. 12(1): 1–7.
de Faria Coelho-Ravagnani, C., Campos Corgosinho, F., La Flor Ziegler Sanches, F., Marques Maia Prado, C., Laviano, A., and Felipe Mota, J. (2021). Dietary Recommendations During the COVID-19 Pandemic: an Extract. Kompass Nutrition & Dietetics 1: 3–7.
Dehghani-Samani, A., Kamali, M., and Hoseinzadeh-Chahkandak, F. (2020). The Role of vitamins on the prevention and/or treatment of COV-ID-19 infection; A Systematic Review. Mod. Care J. 17(3): e104740.
FAO, Food and Agriculture Organization of the United Nations. (2020). Maintaining a healthy diet during the COVID-19 pandemic. http://www.fao.org/3/ca8380en/CA8380EN.pdf. Published March 27, 2020. Accessed April 12, 2020.
Galanakis, C.M., Aldawoud, T., Rizou, M., Rowan, N.J., and Ibrahim, S.A. (2020). Food ingredients and active compounds against the corona-virus disease (COVID-19) pandemic: A comprehensive review. Foods 9(11): 1701.
Goossens, G.H., Dicker, D., Farpour-Lambert, N.J., Frühbeck, G., Mullerova, D., Woodward, E., and Holm, J.C. (2020). Obesity and COVID-19: A
perspective from the European Association for the study of obesity on immunological perturbations, therapeutic challenges, and oppor-tunities in obesity. Obes. Facts 13: 439–452.
Grant, W.B., Lahore, H., McDonnell, S.L., Baggerly, C.A., French, C.B., Aliano, J.L., and Bhattoa, H.P. (2020). Evidence that vitamin D sup-plementation could reduce risk of influenza and COVID-19 infections and deaths. Nutrients 12(4): 988.
Hagel, A.F., Layritz, C.M., Hagel, W.H., Hagel, H.J., Hagel, E., Dauth, W., and Raithel, M. (2013). Intravenous infusion of ascorbic acid decreases serum histamine concentrations in patients with allergic and non-allergic diseases. Naunyn-Schmiedeberg's Arch. Pharmacol. 386(9): 789–793.
Hamid, A.T., and Thakur, N.S. (2021). Role of functional food components in COVID-19 pandemic: A review. Ann. Phytomed. 10: S240–250.
Hussain, T., Tan, B., Yin, Y., Blachier, F., Tossou, M.C., and Rahu, N. (2016). Oxidative stress and inflammation: what polyphenols can do for us? Oxidative medicine and cellular longevity.
Iddir, M., Brito, A., Dingeo, G., Fernandez Del Campo, S.S., Samouda, H., La Frano, M.R., and Bohn, T. (2020). Strengthening the immune system and reducing inflammation and oxidative stress through diet and nu-trition: considerations during the COVID-19 crisis. Nutrients 12(6): 1562.
Ishida, T. (2019). Review on the role of Zn2+ ions in viral pathogenesis and the effect of Zn2+ ions for host cell-virus growth inhibition. Am. J. Biomed. Sci. Res. 2(1): 28–37.
Ismail, S.A., Saliba, V., Bernal, J.L., Ramsay, M.E., and Ladhani, S.N. (2021). SARS-CoV-2 infection and transmission in educational settings: a pro-spective, cross-sectional analysis of infection clusters and outbreaks in England. Lancet Infect. Disease 21(3): 344–353.
Jevalikar, G., Mithal, A., Singh, A., Sharma, R., Farooqui, K.J., Mahendru, S., and Budhiraja, S. (2021). Lack of association of baseline 25-hydroxy-vitamin D levels with disease severity and mortality in Indian patients hospitalized for COVID-19. Sci. Rep. 11(1): 1–8.
Jovic, T.H., Ali, S.R., Ibrahim, N., Jessop, Z.M., Tarassoli, S.P., Dobbs, T.D., and Whitaker, I.S. (2020). Could vitamins help in the fight against COVID-19? Nutrients 12(9): 2550.
Junaid, K., Ejaz, H., Abdalla, A.E., Abosalif, K.O., Ullah, M.I., Yasmeen, H., and Rehman, A. (2020). Effective immune functions of micronutri-ents against SARS-CoV-2. Nutrients 12(10): 2992.
Kazemi, A., Mohammadi, V., Aghababaee, S.K., Golzarand, M., Clark, C.C., and Babajafari, S. (2021). Association of vitamin D status with SARS-CoV-2 infection or COVID-19 severity: a systematic review and meta-analysis. Adv. Nutr. 12(5): 2040–2044.
Kellner, M., Noonepalle, S., Lu, Q., Srivastava, A., Zemskov, E., and Black, S.M. (2017). ROS signaling in the pathogenesis of Acute Lung Injury (ALI) and Acute Respiratory Distress Syndrome (ARDS). Adv. Exp. Med. Biol. 967: 105–137.
Kozlov, E.M., Ivanova, E., Grechko, A.V., Wu, W.K., Starodubova, A.V., and Orekhov, A.N. (2021). Involvement of oxidative stress and the innate immune system in SARS-CoV-2 infection. Diseases 9(1): 17.
Kumar, A., Kubota, Y., Chernov, M., and Kasuya, H. (2020). Potential role of zinc supplementation in prophylaxis and treatment of COVID-19. Med. Hypotheses. 144: 109848.
Laforge, M., Elbim, C., Frère, C., Hémadi, M., Massaad, C., Nuss, P., and Becker, C. (2020). Tissue damage from neutrophil-induced oxidative stress in COVID-19. Nat. Rev. Immunol. 20(9): 515–516.
Lee, G.Y., and Han, S.N. (2018). The role of vitamin E in immunity. Nutrients 10(11): 1614.
Leisman, D.E., Ronner, L., Pinotti, R., Taylor, M.D., Sinha, P., Calfee, C.S., Hi-rayama, A.V., Mastroiani, F., Turtle, C.J., and Deutschman, C.S. (2020). Cytokine elevation in severe and critical COVID-19: a rapid systematic review, meta-analysis, and comparison with other inflammatory syn-dromes. Lancet Respir. Med. 12: 1233–1244.
Lewis, E.D., Meydani, S.N., and Wu, D. (2019). Regulatory role of vitamin E in the immune system and inflammation. IUBMB Life 71(4): 487–494.
Li, X., Geng, M., Peng, Y., Meng, L., and Lu, S. (2020). Molecular immune pathogenesis and diagnosis of COVID-19. J. Pharm. Anal. 10(2): 102–108.
Liu, H., Chen, S., Liu, M., Nie, H., and Lu, H. (2020). Comorbid chronic dis-eases are strongly correlated with disease severity among COVID-19

Journal of Food Bioactives | www.isnff-jfb.com 37
Wong et al. Impact of dietary nutrients and micronutrients on COVID-19: a review
patients: a systematic review and meta-analysis. Aging Dis. 11(3): 668.
Liu, N., Sun, J., Wang, X., Zhang, T., Zhao, M., and Li, H. (2021). Low vita-min D status is associated with coronavirus disease 2019 outcomes: A systematic review and meta-analysis. Int. J. Infect. Dis. 104: 58–64.
Liu, Y., and Rocklöv, J. (2021). The reproductive number of the Delta variant of SARS-CoV-2 is far higher compared to the ancestral SARS-CoV-2 virus. J. Travel. Med.
Lordan, R., Rando, H.M., and Greene, C.S. (2021). Dietary supplements and nutraceuticals under investigation for COVID-19 prevention and treatment. Msystems 6(3): e00122–21.
Mahmoodpoor, A., Hamishehkar, H., Shadvar, K., Ostadi, Z., Sanaie, S., Sa-ghaleini, S.H., and Nader, N.D. (2019). The effect of intravenous sele-nium on oxidative stress in critically ill patients with acute respiratory distress syndrome. Immunol. Invest. 48(2): 147–159.
Martineau, A.R., Jolliffe, D.A., Hooper, R.L., Greenberg, L., Aloia, J.F., Berg-man, P., and Camargo, C.A. (2017). Vitamin D supplementation to prevent acute respiratory tract infections: systematic review and meta-analysis of individual participant data. BMJ 356: i6583.
Mittra, I., de Souza, R., Bhadade, R., Madke, T., Shankpal, P.D., Joshi, M., and Badwe, R. (2020). Resveratrol and Copper for treatment of se-vere COVID-19: an observational study (RESCU 002). medRxiv.
Morrow, L.E., Kollef, M.H., and Casale, T.B. (2010). Probiotic prophylaxis of ventilator-associated pneumonia: a blinded, randomized, controlled trial. Am. J. Respir. Crit. Care Med. 182(8): 1058–1064.
Mortaz, E., Adcock, I.M., Folkerts, G., Barnes, P.J., Paul Vos, A., and Gars-sen, J. (2013). Probiotics in the management of lung diseases. Media-tors Inflamm. 2013: 751068.
Mrityunjaya, M., Pavithra, V., Neelam, R., Janhavi, P., Halami, P.M., and Ravindra, P.V. (2020). Immune-boosting, antioxidant and anti-inflam-matory food supplements targeting pathogenesis of COVID-19. Front Immunol. 11: 570122.
Murai, I.H., Fernandes, A.L., Sales, L.P., Pinto, A.J., Goessler, K.F., Duran, C.S., and Pereira, R.M. (2021). Effect of a single high dose of vitamin D3 on hospital length of stay in patients with moderate to severe COVID-19: a randomized clinical trial. JAMA 325(11): 1053–1060.
Pal, A., Squitti, R., Picozza, M., Pawar, A., Rongioletti, M., Dutta, A.K., and Prasad, R. (2021). Zinc and COVID-19: basis of current clinical trials. Biol. Trace. Elem. Res. 199: 2882–2892.
Pal, R., Banerjee, M., Bhadada, S.K., Shetty, A.J., Singh, B., and Vyas, A. (2021). Vitamin D supplementation and clinical outcomes in COV-ID-19: a systematic review and meta-analysis. J. Endocrinol. Invest.
Pandey, K.R., Naik, S.R., and Vakil, B.V. (2015). Probiotics, prebiotics and synbiotics-a review. J. Food Sci. Technol 52(12): 7577–7587.
Pereira, M., Dantas Damascena, A., Galvão Azevedo, L.M., de Almeida Ol-iveira, T., and da Mota Santana, J. (2020). Vitamin D deficiency ag-gravates COVID-19: systematic review and meta-analysis. Crit. Rev. Food Sci. Nutr.
Prasad, A.S., Beck, F.W., Bao, B., Fitzgerald, J.T., Snell, D.C., Steinberg, J.D., and Cardozo, L.J. (2007). Zinc supplementation decreases incidence of infections in the elderly: effect of zinc on generation of cytokines and oxidative stress. Am. J. Clin. Nutr. 85(3): 837–844.
Raha, S., Mallick, R., Basak, S., and Duttaroy, A.K. (2020). Is copper benefi-cial for COVID-19 patients? Med. Hypotheses. 142: 109814.
Rocha, F.A.C., and de Assis, M.R. (2020). Curcumin as a potential treat-ment for COVID-19. Phytother. Res. 34(9): 2085–2087.
Rodriguez-Leyva, D., and Pierce, G.N. (2021). The Impact of Nutrition on the COVID-19 Pandemic and the Impact of the COVID-19 Pandemic on Nutrition. Nutrients 13(6): 1752.
Rogero, M.M., Leão, M.D.C., Santana, T.M., de MB Pimentel, M.V., Car-lini, G.C., da Silveira, T.F., and Castro, I.A. (2020). Potential benefits and risks of omega-3 fatty acids supplementation to patients with COVID-19. Free Radic. Biol. Med. 156: 190–199.
Shahidi, F. (2012). Nutraceuticals, functional foods and dietary supple-ments in health and disease. J. Food Drug. Anal. 20(1): 226–30.
Sharifi, A., Vahedi, H., Nedjat, S., Rafiei, H., and Hosseinzadeh-Attar, M.J. (2019). Effect of single-dose injection of vitamin D on immune cy-tokines in ulcerative colitis patients: a randomized placebo-con-trolled trial. Apmis 127(10): 681–687.
Sharma, S.V., Haidar, A., Noyola, J., Tien, J, Rushing, M., Naylor, B.M.,
Chuang, R.J., and Markham, C. (2020). Using a rapid assessment methodology to identify and address immediate needs among low-income households with children during COVID-19. PLoS ONE 15(10): e0240009.
Shi, Y., Wang, Y., Shao, C., Huang, J., Gan, J., Huang, X., Bucci, E, Piacentini, M, Ippolito, G, and Melino, G (2020). COVID-19 infection: the per-spectives on immune responses. Cell Death Differ. 27: 1451–1454.
Singh, P., Tripathi, M.K., Yasir, M., Khare, R., Tripathi, M.K., and Shrivastava, R. (2020). Potential inhibitors for SARS-CoV-2 and functional food components as nutritional supplement for COVID-19: a review. Plant Foods Hum. Nutr. 75: 458–66.
Smith, J.C., Sausville, E.L., Girish, V., Yuan, M.L., Vasudevan, A., John, K.M., and Sheltzer, J.M. (2020). Cigarette smoke exposure and inflamma-tory signaling increase the expression of the SARS-CoV-2 receptor ACE2 in the respiratory tract. Dev. Cell 53(5): 514–529.
Smith, M., and Smith, J.C. (2020). Repurposing therapeutics for COVID-19: Supercomputer-based docking to the SARS-CoV-2 viral spike protein and viral spike protein-human ACE2 interface. ChemRxiv.
Speth, R., Carrera, E., Jean-Baptiste, M., Joachim, A., and Linares, A. (2014). Concentration-dependent effects of zinc on angiotensin-con-verting enzyme-2 activity (1067.4). FASEB J. 28: 1067–4.
Srivastava, S., Furlan, S.N., Jaeger-Ruckstuhl, C.A., Sarvothama, M., Berger, C., Smythe, K.S., Garrison, S.M., Specht, J.M., Lee, S.M., Amezquita, R.A., Voillet, V., Muhunthan, V., Yechan-Gunja, S., Pillai, S.P.S., Rader, C., Houghton, A.M., Pierce, R.H., Gottardo, R., Maloney, D.G., and Riddell, S.R. (2021). Immunogenic Chemotherapy Enhances Recruit-ment of CAR-T Cells to Lung Tumors and Improves Antitumor Effi-cacy when Combined with Checkpoint Blockade. Cancer Cell 39(2): 193–208.e10.
Subedi, L., Tchen, S., Gaire, B.P., Hu, B., and Hu, K. (2021). Adjunctive Nu-traceutical Therapies for COVID-19. Int. J. Mol. Sci. 22: 1963.
Sundararaman, A., Ray, M., Ravindra, P.V., and Halami, P.M. (2020). Role of probiotics to combat viral infections with emphasis on COVID-19. Appl. Microbiol. Biotechnol. 104(19): 8089–8104.
Syed, A. (2020). Coronavirus: a mini-review. Int. J. Curr. Res. Med. Sci 6(1): 8–10.
Szabo, Z., Marosvölgyi, T., Szabó, É., Bai, P., Figler, M., and Verzár, Z. (2020). The potential beneficial effect of EPA and DHA supplementation managing cytokine storm in coronavirus disease. Front. Physiol 11: 752.
Tanumihardjo, S.A., Russell, R.M., Stephensen, C.B., Gannon, B.M., Craft, N.E., Haskell, M.J., and Raiten, D.J. (2016). Biomarkers of Nutrition for Development (BOND)—vitamin A review. J. Nutr. 146(9): 1816S–1848S.
UNICEF. (2020). Easy, affordable and healthy eating tips during the coro-navirus disease (COVID-19) outbreak. United Nations Children’s Fund (UNICEF) website. https://www.unicef.
Venkatakrishnan, K., Chiu, H.F., and Wang, C.K. (2019). Extensive review of popular functional foods and nutraceuticals against obesity and its related complications with a special focus on randomized clinical trials. Food Func. 10(5): 2313–2329.
Venkatakrishnan, K., Chiu, H.F., and Wang, C.K. (2019). Extensive review of popular functional foods and nutraceuticals against obesity and its related complications with a special focus on randomized clinical trials. Food Func. 10(5): 2313–2329.
Venkatakrishnan, K., Chiu, H.F., and Wang, C.K. (2019). Extensive review of popular functional foods and nutraceuticals against obesity and its related complications with a special focus on randomized clinical trials. Food Func. 10(5): 2313–2329.
Venkatakrishnan, K., Chiu, H.F., and Wang, C.K. (2019). Popular functional foods and herbs for the management of type-2-diabetes mellitus: a comprehensive review with special reference to clinical trials and its proposed mechanism. J. Funct. Food 57: 425–438.
Villamor, E., and Fawzi, W.W. (2005). Effects of Vitamin A Supplementation on Immune Responses and Correlation with Clinical Outcomes. Clin. Microbiol. Rev. 18(3): 446–464.
Wang, X., Hu, W., Zhu, L., and Yang, Q. (2017). Bacillus subtilis and surfac-tin inhibit the transmissible gastroenteritis virus from entering the intestinal epithelial cells. Biosci. Rep. 37(2): BSR20170082.
WHO. (2020). Nutrition advice for adults during the COVID-19 outbreak.

Journal of Food Bioactives | www.isnff-jfb.com38
Impact of dietary nutrients and micronutrients on COVID-19: a review Wong et al.
World Health Organization Regional Office for the Eastern Mediter-ranean website. http://www.emro.who.int/nutrition/nutrition-info-cus/nutrition-advice-for-adults-during-theCovid-19-outbreak.html. Published March 27, 2020. Accessed April 12, 2020.
Wiersinga, W.J., Rhodes, A., Cheng, A.C., Peacock, S.J., and Prescott, H.C. (2020). Pathophysiology, transmission, diagnosis, and treatment of coronavirus disease 2019 (COVID-19): a review. JAMA 324(8): 782–793.
Wu, D., Lewis, E.D., Pae, M., and Meydani, S.N. (2019). Nutritional modu-lation of immune function: analysis of evidence, mechanisms, and clinical relevance. Front. Immunol. 9: 3160.
Yuki, K., Fujiogi, M., and Koutsogiannaki, S. (2020). COVID-19 pathophysi-ology: A review. Clin. Immunol. 215: 108427.
Zahedipour, F., Hosseini, S.A., Sathyapalan, T., Majeed, M., Jamialahmadi, T., Al-Rasadi, K., and Sahebkar, A. (2020). Potential effects of curcum-in in the treatment of COVID-19 infection. Phytother. Res. 34(11): 2911–2920.
Zeng, J., Wang, C.T., Zhang, F.S., Qi, F., Wang, S.F., Ma, S., and Wang, Y.P. (2016). Effect of probiotics on the incidence of ventilator-associated pneumonia in critically ill patients: a randomized controlled multi-center trial. Intensive Care. Med. 42(6): 1018–1028.
Zhang, J., Rao, X., Li, Y., Zhu, Y., Liu, F., Guo, G., and Peng, Z. (2021). Pilot trial of high-dose vitamin C in critically ill COVID-19 patients. Ann. Intensive Care 11(1): 1–12.
Zolkiewicz, J., Marzec, A., Ruszczyński, M., and Feleszko, W. (2020). Postbi-otics—a step beyond pre-and probiotics. Nutrients 12(8): 2189.

Copyright: © 2021 International Society for Nutraceuticals and Functional Foods.All rights reserved.
39
Review J. Food Bioact. 2021;15:39–50
Journal ofFood Bioactives International Society for
Nutraceuticals and Functional Foods
Phenolic compounds in cereal grains and effects of processing on their composition and bioactivities: a review
Fereidoon Shahidi*, Renan Danielski and Chiaki Ikeda
Department of Biochemistry, Memorial University of Newfoundland, St. John’s, NL, Canada A1B 3X9*Corresponding author: Department of Biochemistry, Memorial University of Newfoundland, St. John’s, NL, Canada A1B 3X9. E-mail: [email protected]: 10.31665/JFB.2021.15281Received: September 27, 2021; Revised received & accepted: September 29, 2021Citation: Shahidi, F., Danielski, R., and Ikeda, C. (2021). Phenolic compounds in cereal grains and effects of processing on their composi-tion and bioactivities: a review. J. Food Bioact. 15: 39–50.
Abstract
Cereals are a staple food in the diets of many populations globally. Besides their nutritive function in food, they are also rich in various groups of bioactive compounds, especially polyphenols. Wheat, rice, barley, rye, oat, maize, millet, sorghum, and other cereal grains present a great variety of phenolic acids, flavonoids, proanthocyanidins, alkylresorcinols, and lignans, which can be affected in many ways by the post-harvest treatments and further processing of these feedstocks. This review discusses up-to-date studies about the effects of common cereal pro-cessing techniques on their phenolic composition, biological activities, and bioefficiency. Generally, mild thermal and high-pressure treatments enhance cereals’ phenolic composition by releasing the insoluble-bound fraction, which increases their bioaccessibility. On the other hand, processes involving extreme temperature conditions and removal of the grains’ outer layers may drastically reduce the phenolic content. Therefore, it is imperative to optimize the processing conditions of cereals, so their health-promoting benefits are preserved.
Keywords: Polyphenols; Antioxidant activity; Post-harvest treatment; Insoluble-bound; Bioaccessibility.
1. Introduction
Cereal cultivation has been a crucial factor for the development of many countries due to its economic and nutritional importance. As staple foods, they are able to provide a significant portion of energy and dietary needs. Cereals are rich in protein, selected micronutri-ents, and even antinutrients. Maize, rice, wheat, barley, sorghum, millets, oat, and rye are among the most cultivated cereals globally, playing a vital role in the economy of both developed and develop-ing countries (Raheem et al., 2021). However, the importance of cereals extends beyond their essential nutrients. The consumption of whole grains (containing endosperm, germ, and bran) has been associated with a decreased incidence of cardiovascular diseases, some types of cancer, type 2 diabetes, and obesity, among others (Shahidi, 2009). These effects are due to the action of bioactive compounds present in such foods (Shahidi et al., 2019).
The most prominent bioactive classes of compounds found in
cereals include phenolics, carotenoids, phytosterols, beta-glucans, and gamma-oryzanol. In raw cereals, their amount and profile vary according to their location in the grain. Usually, refined cereals possess an underwhelming bioactive composition compared to whole grains due to the removal of germ and bran, where several bioactives are located. The mechanism through which these com-pounds exert their beneficial effects varies according to their type, structure, occurrence in the matrix, and other conditions. Never-theless, a number of bioactive substances have in common their capacity to act as antioxidants, dampening oxidative stress in the body, thus protecting the macromolecules (e.g., lipids, proteins, DNA) against damage that can lead to the development of chronic diseases. This has been deemed one of their most appealing fea-tures, helping the promotion of consumption of bioactive-rich foods and their capacity to serve as ingredients for nutraceutical and functional food development (Naczk and Shahidi, 2006).
The susceptibility of bioactive compounds in cereals is influ-enced by temperature and pH variations, exposure to light and ox-

Journal of Food Bioactives | www.isnff-jfb.com40
Phenolic compounds in cereal grains and effects of processing Shahidi et al.
ygen, and enzymatic reactions. Such factors can compromise their ability to serve as antioxidants and exert their biological activities (Chawda et al., 2017). Moreover, exposure of bioactives to the aforementioned environmental factors is common for cereal grains and cereal products due to post-harvest treatment and storage. At this stage, cereals undergo a series of processing steps designed to clean, sort, and remove inedible portions (primary processing) and can even be transformed into more appealing products (secondary processing), being subjected to more extreme processes such as frying and extrusion (Food and Agriculture Organization, 2021). Therefore, it is imperative to analyze how the major bioactives found in cereals are affected by grain processing.
This review discusses up-to-date information on the transfor-mations of cereal grains upon processing and the effects of com-monly used unit operations on phenolic compounds, one the most relevant groups of bioactive substances encountered in highly pro-duced cereals.
2. Cereal grains: economic and nutritional importance
Cultivation of cereal grains drives the economy of several territo-ries worldwide. The Food and Agriculture Organization (FAO) of the United Nations estimates that around 70% of the world’s farm-land is used to cultivate cereal grains. Approximately 45% of the world’s economically active population is engaged in agriculture-related activities, primarily planting and harvesting cereal grains (Serna-Saldivar, 2016). Data from the 2020/2021 harvest (Statista, 2021) shows that corn (maize) is the most produced cereal in the world, reaching 1.12 billion metric tons, followed by wheat (775.8 million metric tons (Mt)), milled rice (505 Mt), barley (159.74 Mt), sorghum (62.05 Mt), oats (25.53 Mt), and rye (14.3 Mt). China is the largest cereal producer with over 612 Mt harvested in 2018, followed by the United States with 467 Mt, and India with 318 Mt. Most of this production is directed to animal feed and etha-nol manufacture, with human consumption receiving the smallest fraction. According to Food and Agriculture Organization (2020), cereal production is expected to grow by 375 Mt in 2029. This expansion is due to the development of better agricultural practices
and advances in biotechnology.From a nutritional standpoint, the availability of cereals is im-
portant for meeting the global food demand since it is a cost-effec-tive and nutritious food option, especially when considering the population of developing countries (Raheem et al., 2021). Cereals are botanically classified as grasses belonging to the monocot fam-ily Poaceae. According to climate demands, cereals can be either warm-season or cool-season. Warm-season cereals encompass those grown year-round in tropical climates and during the frost-free season in temperate climates. Examples include corn, rice, sorghum, and millet. On the other hand, cool-season cereals are grown in moderate climates and include wheat, barley, oats, and rye (Koehler and Wieser, 2013). Anatomically, cereal grains are composed of bran, germ, and endosperm. The endosperm is the inner layer, making up most of the grain, and it is rich in starch and proteins. The germ is a small nutrient-rich core containing vi-tamins, fatty acids, and bioactive compounds. The bran is the outer layer involving the endosperm, a source of fiber, B vitamins, and trace minerals. The husk is combined with the bran in selected spe-cies, such as oats, barley, and rice (Sidhu et al., 2007).
Chemically, cereal grains are mainly composed of carbohy-drates (56–74%), especially the starch fraction deposited in the endosperm, with fiber (mainly in the bran) representing 2–13% (Koehler and Wieser, 2013). Table 1 shows the chemical composi-tion of some of the most relevant cereal grains grown worldwide.
In general, cereals are high in their starch content which is high in amylopectin (72–75%), while amylose accounts for 25–28%. Besides its nutritional importance in cereals, starch plays a func-tional role when processed and transformed into value-added prod-ucts due to its gelatinization (in pasta, for example) and retrogra-dation properties (in cereal-based baked goods) (Rodehutscord et al., 2016).
As for cereals’ protein composition, a great variability ex-ists depending on the type of cereal. In general, the amino acid glutamine is the predominant one in most cereals, representing 15–31% of all amino acids present. Wheat, rye, and barley pos-sess a significant fraction of proline (12–14%). Leucine (7–14%) and alanine (4–11%) can also be found in high amounts in cereal grains. On the contrary, essential amino acids, such as tryptophan,
Table 1. Chemical composition of the major cereal grains produced worldwide (whole grain)
Wheat Rye Maize Barley Oats Rice Millet
Moisture (g/100g) 12.6 13.6 11.3 12.1 13.1 13.0 12.0
Protein (g/100g) 11.3 9.4 8.8 11.1 10.8 7.7 10.5
Lipids (g/100g) 1.8 1.7 3.8 2.1 7.2 2.2 3.9
Available carbohydrates (g/100g) 59.4 60.3 65.0 62.7 56.2 73.7 68.2
Fiber (g/100g) 13.2 13.1 9.8 9.7 9.8 2.2 3.8
Minerals (g/100g) 1.7 1.9 1.3 2.3 2.9 1.2 1.6
Thiamine (B1) (mg/kg) 4.6 3.7 3.6 4.3 6.7 4.1 4.3
Riboflavin (B2) (mg/kg) 0.9 1.7 2.0 1.8 1.7 0.9 1.1
Nicotinamide (mg/kg) 51.0 18.0 15.0 48.0 24.0 52.0 18.0
Panthothenic acid (mg/kg) 12.0 15.0 6.5 6.8 7.1 17.0 14.0
Vitamin B6 (mg/kg) 2.7 2.3 4.0 5.6 9.6 2.8 5.2
Folic acid (mg/kg) 0.9 1.4 0.3 0.7 0.3 0.2 0.4
Tocols (mg/kg) 41.0 40.0 66.0 22.0 18.0 19.0 40.0
Adapted from Koehler and Wieser (2013).

Journal of Food Bioactives | www.isnff-jfb.com 41
Shahidi et al. Phenolic compounds in cereal grains and effects of processing
methionine, histidine, and lysine, are present in smaller quantities (0.2–1.0, 1.3–2.9, 1.8–2.2, and 1.4–3.3%, respectively). However, with recent advances in genetic engineering, there is a possibility of increasing the levels of essential amino acids, as successfully achieved in the case of high-lysine barley and corn (Yu and Tian, 2018).
The germ is the principal storage site for cereal lipids. Among cereals, oat is a significant source of lipid (6–8%) as opposed to wheat and rye (1.7%). The fatty acid composition of oat lipids is similar to most cereal grains, with a predominance of linoleic acid (39–69%), followed by oleic acid (11–36%) and palmitic acid (18–28%). Although triacylglycerols are the primary lipid class in the germ, a small fraction of phospholipids and glycolipids can also be found in the endosperm (Koehler and Wieser, 2013).
Minerals and vitamins are the minor constituents of cereals. In terms of mineral content (calcium, magnesium, potassium, phosphorus, iron, and sodium), cereals are richer than milk, meat, and vegetables but present lower concentrations when compared to pulses. Nevertheless, as staple foods, cereals are consumed in high quantities; thus, being characterized as significant sources of minerals in the diet. The majority of minerals in cereals are con-centrated in the bran and the germ. Meanwhile, cereals are rich sources of vitamins from the B-group and tocopherols. Similar to minerals, the vitamins are mainly found in the germ, highlighting the importance of consuming whole grains as opposed to refined cereals (Baniwal et al., 2021).
3. Phenolic compounds in cereals
Phenolic compounds are secondary plant metabolites that have the primary role of protecting against oxidative stress, ultraviolet light, and pathogen attack (Shahidi and Naczk, 2003). Structur-ally, phenolics are characterized by the presence of one or more aromatic rings holding one (monophenols) or more (polyphenols) hydroxyl groups (Duodu, 2011). To date, over 8,000 phenolics have been identified in natural sources, being classified into sev-eral subgroups according to their hydroxylation and methylation pattern (Shahidi et al., 2019). Flavonoids, phenolic acids, and tan-nins (proanthocyanidins and hydrolysable tannins) are the most significant phenolic subclasses; however, lignans and alkylresor-cinols are also important phenolic classes in cereals (Cheynier et al., 2013; Shahidi et al., 2019).
Phenolic acids are derived from benzoic and cinnamic acids, being classified into hydroxybenzoic (e.g., p-hydroxybenzoic, pro-tocatechuic, vanillic, syringic, and gallic acids) and hydroxycin-namic acids (e.g., p-coumaric, caffeic, ferulic, and sinapic acids) (Duodu, 2011; Shahidi et al., 2019). Compounds belonging to this subgroup may also be associated with macromolecules in cereal matrices, such as cellulose, hemicellulose, and pectin, acting as the building blocks supporting the cell wall. This fraction is known as insoluble-bound phenolics and can represent up to 60% of all phenolic compounds present in selected grains (Shahidi and Yeo, 2016).
Meanwhile, flavonoids are the largest phenolic class in terms of number of compounds. Their basic structural skeleton is C6-C3-C6, where two benzene rings are connected to a central hetero-cyclic ring through a three-carbon bridge. They can be further broken down into flavanones, flavonols, flavan-3-ols, flavones, isoflavones, flavanonols, and anthocyanins, the latter being water-soluble pigments (Shahidi et al., 2019). Cereal grains are abundant sources of flavonoids. In such materials, they can be present either as insoluble-bound or soluble compounds. The soluble flavonoids
may occur in the free form, or they can be conjugated with poly-saccharides (glycosides) or esterified/etherified to fatty acids (Sha-hidi and Yeo, 2016).
Monomeric flavonoids can form oligomers and polymers, obtaining a higher degree of complexity. These polyphenols are known as proanthocyanidins; polymeric structures composed of (+)-catechin and (−)-epicatechin. Besides flavonoids, phenolic ac-ids can also form polymeric structures known as hydrolysable tan-nins, which can be either esters of gallic (gallotannins) or ellagic (ellagitannins) acids (Shahidi and Yeo, 2016).
Besides phenolic acids, flavonoids, and tannins, other phenolic groups are also considerable in cereals, namely lignans and alkyl-resorcinols, the latter being a class of phenolic lipids. Lignans are dimers with a 2,3-dibenzylbutane structure. They are precursors of phytoestrogen, and can occur in cereal grains such as rye, wheat, barley, and oats (Bhattacharya et al., 2010). Meanwhile, alkylres-orcinols are amphipathic 1,3-dihydroxybenzene derivatives carry-ing an alkyl chain (odd-numbered) at position 5 of the benzene ring. They are present in wheat, rye, barley, millet, and maize (Ross et al., 2003a).
The antioxidant power of phenolic compounds is related to most of their known bioactivities. They can function both as pri-mary (free radical scavenging) or secondary (metal chelation) an-tioxidants. As primary antioxidants, phenolics can donate hydro-gen atoms to free radicals, stabilizing them and interrupting the propagation of oxidation. Then, the compound is transformed into a phenoxyl radical, which gains stability by delocalizing its un-paired electron around the aromatic ring. On the other hand, phe-nolics bearing catechol and galloyl groups can also act as second-ary antioxidants by complexing with transition metals, preventing metal-catalyzed initiation of free radicals and formation of reactive oxygen species (Chen et al., 2020).
According to Naczk and Shahidi (2006), flavonoids and phe-nolic acids in cereals occur as free compounds or conjugated with other molecules, being mainly located in the bran, even though the endosperm and germ also present considerable amounts. Table 2 presents the phenolic composition of major cereal grains.
Zhang et al. (2019) analyzed the pericarp of purple maize and reported a total phenolic content (TPC) of 69.1–229.1 mg of gallic acid equivalents (GAE)/g and total anthocyanin content of 17–93.5 mg of cyanidin-3-glucoside equivalents (C3G)/g, varying accord-ing to the genotype. The anthocyanins cyanidin-3-glucoside, pelargonidin-2-O-glucoside, peonidin-3-O-glucoside, cyanidin-3-(6′-malonylglucoside), pelargonidin-3-(6′-malonylglucoside), and peonidin-3-(6′-malonylglucoside were identified in the sam-ples. Besides, phenolic acids, namely o-coumaric, caffeic, vanillic, protocatechuic, and ferulic acids were also identified, as well as flavonoids rutin, luteolin, quercetin, naringenin, and kaempferol. In terms of bioactivities, the authors demonstrated that quercetin, luteolin, and rutin were the main contributors to the anti-inflam-matory and anti-diabetic actions displayed by the samples. At the same time, vanillic and protocatechuic acids had a major impact in terms of anti-adipogenic potential.
According to Suriano et al. (2021), among pigmented maize, the purple variety is the richest one in terms of total phenolic (4,047 µg GAE/g), total flavonoid (1,998 µg of catechin equivalents (CE)/g), total anthocyanin (780 µg C3G/g), and total proanthocya-nidin (1,381 µg CE/g) contents, followed by blue maize (2,416 µg GAE/g, 1,739 µg CE/g, 216 µg C3G/g, and 1,279 µg CE/g, re-spectively) and red maize (2,373 µg GAE/g, 1,664 µg CE/g, 38.9 µg C3G/g, and 578 µg CE/g, respectively). Conventional yellow maize had the lowest total phenolics (1,359 µg GAE/g) and fla-vonoids (1,162 µg CE/g) and does not contain any anthocyanins.
Pigmented rice is also a remarkable source of anthocyanins.
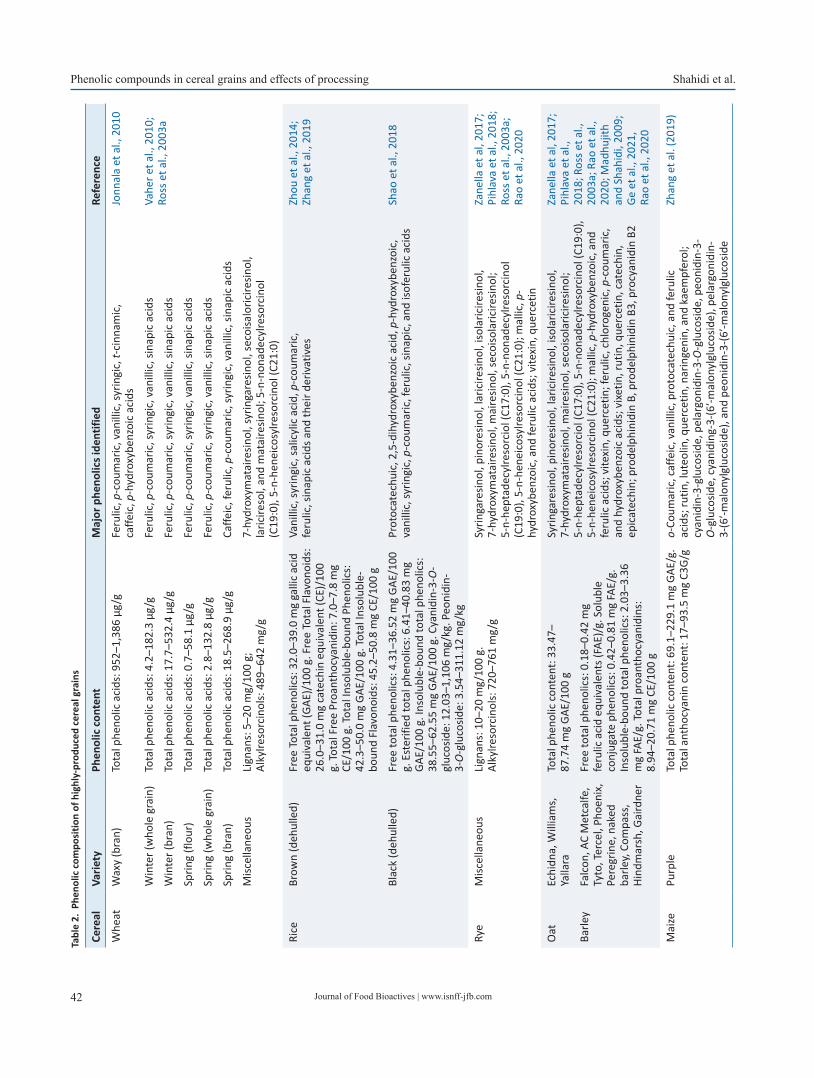
Journal of Food Bioactives | www.isnff-jfb.com42
Phenolic compounds in cereal grains and effects of processing Shahidi et al.Ta
ble
2. P
heno
lic c
ompo
sitio
n of
hig
hly-
prod
uced
cer
eal g
rain
s
Cere
alVa
riety
Phen
olic
con
tent
Maj
or p
heno
lics i
dent
ified
Refe
renc
e
Whe
atW
axy
(bra
n)To
tal p
heno
lic a
cids
: 952
–1,3
86 µ
g/g
Feru
lic, p
-cou
mar
ic, v
anill
ic, s
yrin
gic,
t-ci
nnam
ic,
caffe
ic, p
-hyd
roxy
benz
oic
acid
sJo
nnal
a et
al.,
201
0
Win
ter (
who
le g
rain
)To
tal p
heno
lic a
cids
: 4.2
–182
.3 µ
g/g
Feru
lic, p
-cou
mar
ic, s
yrin
gic,
van
illic
, sin
apic
aci
dsVa
her e
t al.,
201
0;
Ross
et a
l., 2
003a
Win
ter (
bran
)To
tal p
heno
lic a
cids
: 17.
7–53
2.4
µg/g
Feru
lic, p
-cou
mar
ic, s
yrin
gic,
van
illic
, sin
apic
aci
ds
Sprin
g (fl
our)
Tota
l phe
nolic
aci
ds: 0
.7–5
8.1
µg/g
Feru
lic, p
-cou
mar
ic, s
yrin
gic,
van
illic
, sin
apic
aci
ds
Sprin
g (w
hole
gra
in)
Tota
l phe
nolic
aci
ds: 2
.8–1
32.8
µg/
gFe
rulic
, p-c
oum
aric
, syr
ingi
c, v
anill
ic, s
inap
ic a
cids
Sprin
g (b
ran)
Tota
l phe
nolic
aci
ds: 1
8.5–
268.
9 µg
/gCa
ffeic
, fer
ulic
, p-c
oum
aric
, syr
ingi
c, v
anill
ic, s
inap
ic a
cids
Misc
ella
neou
sLi
gnan
s: 5
–20
mg/
100
g;
Alky
lreso
rcin
ols:
489
–642
mg/
g7-
hydr
oxym
atai
resin
ol, s
yrin
gare
sinol
, sec
oisa
loric
iresin
ol,
laric
ireso
l, an
d m
atai
resin
ol; 5
-n-n
onad
ecyl
reso
rcin
ol
(C19
:0),
5-n-
hene
icos
ylre
sorc
inol
(C21
:0)
Rice
Brow
n (d
ehul
led)
Free
Tota
l phe
nolic
s: 3
2.0–
39.0
mg
galli
c ac
id
equi
vale
nt (G
AE)/
100
g. F
ree
Tota
l Fla
vono
ids:
26
.0–3
1.0
mg
cate
chin
equ
ival
ent (
CE)/
100
g. To
tal F
ree
Proa
ntho
cyan
idin
: 7.0
–7.8
mg
CE/1
00 g
. Tot
al In
solu
ble-
boun
d Ph
enol
ics:
42
.3–5
0.0
mg
GAE/
100
g. To
tal I
nsol
uble
-bo
und
Flav
onoi
ds: 4
5.2–
50.8
mg
CE/1
00 g
Vani
llic,
syrin
gic,
salic
ylic
aci
d, p
-cou
mar
ic,
feru
lic, s
inap
ic a
cids
and
thei
r der
ivat
ives
Zhou
et a
l., 2
014;
Zh
ang
et a
l., 2
019
Blac
k (d
ehul
led)
Free
tota
l phe
nolic
s: 4
.31–
36.5
2 m
g GA
E/10
0 g.
Est
erifi
ed to
tal p
heno
lics:
6.4
1–40
.83
mg
GAE/
100
g. In
solu
ble-
boun
d to
tal p
heno
lics:
38
.55–
62.5
5 m
g GA
E/10
0 g.
Cya
nidi
n-3-
O-
gluc
osid
e: 1
2.03
–1,1
06 m
g/kg
. Peo
nidi
n-3-
O-g
luco
side:
3.5
4–31
1.12
mg/
kg
Prot
ocat
echu
ic, 2
,5-d
ihyd
roxy
benz
oic
acid
, p-h
ydro
xybe
nzoi
c,
vani
llic,
syrin
gic,
p-c
oum
aric
, fer
ulic
, sin
apic
, and
isof
erul
ic a
cids
Shao
et a
l., 2
018
Rye
Misc
ella
neou
sLi
gnan
s: 1
0–20
mg/
100
g.
Alky
lreso
rcin
ols:
720
–761
mg/
gSy
ringa
resin
ol, p
inor
esin
ol, l
aric
iresin
ol, i
sola
ricire
sinol
, 7-
hydr
oxym
atai
resin
ol, m
aire
sinol
, sec
oiso
laric
iresin
ol;
5-n-
hept
adec
ylre
sorc
iol (
C17:
0), 5
-n-n
onad
ecyl
reso
rcin
ol
(C19
:0),
5-n-
hene
icos
ylre
sorc
inol
(C21
:0);
mal
lic, p
-hy
drox
yben
zoic
, and
feru
lic a
cids
; vite
xin,
que
rcet
in
Zane
lla e
t al,
2017
; Pi
hlav
a et
al.,
201
8;
Ross
et a
l., 2
003a
; Ra
o et
al.,
202
0
Oat
Echi
dna,
Will
iam
s,
Yalla
raTo
tal p
heno
lic c
onte
nt: 3
3.47
–87
.74
mg
GAE/
100
gSy
ringa
resin
ol, p
inor
esin
ol, l
aric
iresin
ol, i
sola
ricire
sinol
, 7-
hydr
oxym
atai
resin
ol, m
aire
sinol
, sec
oiso
laric
iresin
ol;
5-n-
hept
adec
ylre
sorc
iol (
C17:
0), 5
-n-n
onad
ecyl
reso
rcin
ol (C
19:0
), 5-
n-he
neic
osyl
reso
rcin
ol (C
21:0
); m
allic
, p-h
ydro
xybe
nzoi
c, a
nd
feru
lic a
cids
; vite
xin,
que
rcet
in; f
erul
ic, c
hlor
ogen
ic, p
-cou
mar
ic,
and
hydr
oxyb
enzo
ic a
cids
; vix
etin
, rut
in, q
uerc
etin
, cat
echi
n,
epic
atec
hin;
pro
delp
hini
din
B, p
rode
lphi
nidi
n B3
, pro
cyan
idin
B2
Zane
lla e
t al,
2017
; Pi
hlav
a et
al.,
20
18; R
oss e
t al.,
20
03a;
Rao
et a
l.,
2020
; Mad
hujit
h an
d Sh
ahid
i, 20
09;
Ge e
t al.,
202
1,
Rao
et a
l., 2
020
Barle
yFa
lcon
, AC
Met
calfe
, Ty
to, T
erce
l, Ph
oeni
x,
Pere
grin
e, n
aked
ba
rley,
Com
pass
, Hi
ndm
arsh
, Gai
rdne
r
Free
tota
l phe
nolic
s: 0
.18–
0.42
mg
feru
lic a
cid
equi
vale
nts (
FAE)
/g. S
olub
le
conj
ugat
e ph
enol
ics:
0.4
2–0.
81 m
g FA
E/g.
In
solu
ble-
boun
d to
tal p
heno
lics:
2.0
3–3.
36
mg
FAE/
g. To
tal p
roan
thoc
yani
dins
: 8.
94–2
0.71
mg
CE/1
00 g
Mai
zePu
rple
Tota
l phe
nolic
con
tent
: 69.
1–22
9.1
mg
GAE/
g.
Tota
l ant
hocy
anin
con
tent
: 17–
93.5
mg
C3G/
go-
Coum
aric
, caf
feic
, van
illic
, pro
toca
tech
uic,
and
feru
lic
acid
s; ru
tin, l
uteo
lin, q
uerc
etin
, nar
inge
nin,
and
kae
mpf
erol
; cy
anid
in-3
-glu
cosid
e, p
elar
goni
din-
3-O
-glu
cosid
e, p
eoni
din-
3-O
-glu
cosid
e, c
yani
ding
-3-(6
′-mal
onyl
gluc
osid
e), p
elar
goni
din-
3-(6
′-mal
onyl
gluc
osid
e), a
nd p
eoni
din-
3-(6
′-mal
onyl
gluc
osid
e
Zhan
g et
al.
(201
9)

Journal of Food Bioactives | www.isnff-jfb.com 43
Shahidi et al. Phenolic compounds in cereal grains and effects of processing
Pang et al. (2018) analyzed the free and insoluble-bound phenolic fractions of whole-grain and bran of white, red, and black rice. It is noteworthy that most samples contained around two times more insoluble-bound than free phenolics. This fraction has also been reported as the most prominent one in maize (85%), oats and wheat (75%) (Adom and Liu, 2002). Liyana-Pathirana and Shahidi (2006) demonstrated that hard and soft whole wheat are mainly composed of insoluble-bound phenolics, as opposed to their free and esterified counterparts. This reinforces the role of cereals as one of the main sources of insoluble-bound phenolics in the diet. This fraction has been associated with improved gut health and reduction of the risk of developing colon cancer (Shahidi and Yeo, 2016). According to Pang et al. (2018), rice bran has a phenolic content of almost four times higher than the whole grain, confirm-ing that these compounds are preferably concentrated in the outer layers of cereal grains. Pigmented rice (red and black) is superior to white rice in terms of phenolic content since it has a significant amount of anthocyanins, which is lacking in white rice. Ferulic and p-coumaric acids were among the main phenolic acids identified in both white and pigmented rice. Isoferulic acid was also found in high amounts in white rice, whereas pigmented rice varieties showed a significant content of vanillic acid.
Proanthocyanidins are also relevant in pigmented rice. Shao et al. (2018) investigated the proanthocyanidin composition of brown, red, and black rice genotypes. Among all samples, proanthocyani-dins were only detected in red rice, ranging from 58.13 to 254.8 mg CE/100 g, depending on the genotype. According to the authors, the majority of identified proanthocyanidins were oligomers of 5–8 units (40%), while polymers (over ten units) accounted for 29%.
Proanthocyanidins are also present in barley, mainly in the form of oligomeric prodelphinidins and procyanidins. These compounds are involved in the formation of haze in beer. Besides proanthocya-nidins, barley presents a rich and diverse phenolic profile, includ-ing phenolic acids, their esters and glycosides, anthocyanins, and lignans. The free phenolic fraction of barley is primarily composed of ferulic acid (especially the bran). On the other hand, barleyˋs insoluble-bound phenolic fraction is abundant in p-hydroxyben-zoic acid.
Millet grains play an important role in the diet of several Asian and African populations, besides being used as an ingredient in multigrain and gluten-free cereal products. This cereal has a rich and diverse phenolic composition, especially in regard to its in-soluble-bound fraction. According to Chandrasekara and Shahidi (2011a, b), over 50 phenolic compounds were identified among various whole millet grain varieties. Most phenolic compounds were detected in the insoluble-bound fraction, which accounted for an average TPC of 17.3 µmol ferulic acid equivalent (FAE)/g, followed by free (6.89 µmol FAE/g) etherified (2.05 µmol FAE/g), and esterified (0.99 µmol FAE/g) phenolics. Among the phenolic groups detected, phenolic acids and their derivatives (dehydrod-iferulates and dehydrotriferulates), flavan-3-ols monomers and dimers, flavonols, flavones, flavanonols, and proanthocyanidins were most abundant. The insoluble-bound fraction was mainly composed of hydroxycinnamic acids and their derivatives (e.g., chlorogenic, caffeic, p-coumaric, ferulic acids). Meanwhile, flavo-noids (e.g., catechin, epicatechin, gallocatechin, epigallocatechin, procyanidins B1 and B2) accounted for most of the free phenolics.
Besides its rich phenolic composition, millet has also been shown to present numerous bioactivities. Millet phenolics have demonstrated protection against copper-induced human low-density lipoprotein (LDL) oxidation (Chandrasekara and Sha-hidi, 2012). The accumulation of oxidized LDL can lead to the formation of atheromatous plaques that can eventually cause atherosclerosis and associated cardiovascular diseases. Phenolics Ce
real
Varie
tyPh
enol
ic c
onte
ntM
ajor
phe
nolic
s ide
ntifi
edRe
fere
nce
Mill
etKo
do (w
hole
gra
in)
Free
tota
l phe
nolic
s: 1
6.2
µmol
feru
lic
acid
equ
ival
ent (
FAE)
/g. E
ster
ified
tota
l ph
enol
ics:
2.0
2 µm
ol F
AE/g
. Eth
erifi
ed to
tal
phen
olic
s: 1
.55
µmol
FAE
/g. I
nsol
uble
-bo
und
tota
l phe
nolic
s: 8
1.6
µmol
FAE
/g
Galli
c, p
roto
cate
chui
c, v
anill
ic, s
yrin
gic,
p-h
ydro
xybe
nzoi
c,
p-co
umar
ic, c
hlor
ogen
ic, c
affe
ic, f
erul
ic a
cids
; cat
echi
n,
epic
atec
hin,
myr
icet
in, k
aem
pfer
ol, q
uerc
etin
, lut
eolin
Chan
dras
ekar
a an
d Sh
ahid
i, 20
11a,
b
Foxt
ail (
who
le g
rain
)Fr
ee to
tal p
heno
lics:
4.4
9 µm
ol fe
rulic
ac
id e
quiv
alen
t (FA
E)/g
. Est
erifi
ed to
tal
phen
olic
s: 0
.37
µmol
FAE
/g. E
ther
ified
tota
l ph
enol
ics:
0.3
2 µm
ol F
AE/g
. Ins
olub
le-
boun
d to
tal p
heno
lics:
11.
6 µm
ol F
AE/g
o-Co
umar
ic, c
affe
ic, v
anill
ic, p
roto
cate
chui
c, a
nd fe
rulic
ac
ids;
rutin
, lut
eolin
, que
rcet
in, n
arin
geni
n, a
nd k
aem
pfer
ol;
cyan
idin
-3-g
luco
side,
pel
argo
nidi
n-3-
O-g
luco
side,
peo
nidi
n-3-
O-g
luco
side,
cya
nidi
ng-3
-(6′-m
alon
ylgl
ucos
ide)
, pel
argo
nidi
n-3-
(6′-m
alon
ylgl
ucos
ide)
, and
peo
nidi
n-3-
(6′-m
alon
ylgl
ucos
ide
Zhan
g et
al.
(201
9)
Sorg
hum
Whi
te (w
hole
gra
in)
Free
phe
nolic
aci
ds: 1
23 µ
g/g.
Inso
lubl
e-bo
und
phen
olic
aci
ds: 6
22.9
µg/
gGa
llic,
pro
toca
tech
uic,
p-h
ydro
xybe
nzoi
c, v
anill
ic,
caffe
ic, p
-cou
mar
ic, f
erul
ic, a
nd c
inna
mic
aci
ds; c
atec
hin,
ep
icat
echi
n, p
rocy
anid
in B
1, a
pige
nin,
lute
olin
Dyke
s and
Ro
oney
, 200
6
Galli
c, p
roto
cate
chui
c, p
-hyd
roxy
benz
oic,
van
illic
, ca
ffeic
, p-c
oum
aric
, fer
ulic
, and
cin
nam
ic a
cids
; cat
echi
n,
epic
atec
hin,
pro
cyan
idin
B1,
api
geni
n, lu
teol
in
Dyke
s and
Ro
oney
, 200
6
Tabl
e 2.
Phe
nolic
com
posi
tion
of h
ighl
y-pr
oduc
ed c
erea
l gra
ins -
(con
tinue
d)

Journal of Food Bioactives | www.isnff-jfb.com44
Phenolic compounds in cereal grains and effects of processing Shahidi et al.
from Kodo millet were able to suppress in 40% the formation of conjugated dienes, a product of LDL oxidation. In another study (Chandrasekara and Shahidi, 2011a), millet’s soluble (from whole grain, dehulled grain, and hulls) and insoluble-bound (from whole grain) phenolics exerted protection against free radical-induced supercoiled DNA strand scission in a dose-dependent manner. At a concentration of 0.5 mg/mL, phenolic extracts displayed oxida-tion inhibition varying from 46 to 96% in the presence of hydroxyl radicals and from 31 to 95% in the presence of peroxyl radicals. The attack on the DNA molecule by free radicals can lead to base modification and strand scission, inducing mutagenesis, which could further progress to cancer development. Millet phenolics have been demonstrated to be able to suppress this effect. Besides, phenolics from whole millet grains have also shown inhibition of human colon adenocarcinoma cell proliferation with efficiency ranging from 28 to 100%, which lends support to the anticancer potential of millet phenolics (Chandrasekara and Shahidi, 2011b).
Cereals are the major source of lignan in the Western diet, with wheat and rye having the highest contents of lignan among all cere-als, especially their bran fraction. In wheat, for example, 70–85% of the grain lignan is located in the bran, which accounts for 5–20 mg/100 g on average, depending on the variety. Rye bran possesses a similar content, ranging from 10 to 20 mg/100 g. The most common-ly found lignans in wheat are 7-hydroxymatairesinol, syringaresinol, secoisaloriciresinol, lariciresinol, and matairesinol. Meanwhile, in rye, the major lignans encountered are syringaresinol, pinoresinol, lariciresinol, isolariciresinol, 7-hydroxymatairesinol, mairesinol, secoisolariciresinol (Zanella et al., 2017; Pihlava et al., 2018). Be-sides rye and wheat, other cereals present smaller amounts of lig-nan, namely barley (whole grain flour), buckwheat (whole grain flour), maize (whole grain), and oat (whole grain flour) at average concentrations of 0.28, 0.76, 0.33, and 0.65 mg/100 g, respectively (Rodríguez-García et al., 2019).
Besides their well-documented antioxidant properties, some lignans can also act as regulators of the activity of specific nuclear receptors (e.g., estrogen receptors, peroxisome proliferator acti-vate receptor gamma, liver X receptor alpha) that are involved in glucose and fatty acid metabolism, as well as insulin action. This suggests that they could efficiently be used for selected physiologi-cal functions (Zanella et al., 2017).
In the food matrix, lignans occur as esters, glycosides, and poly-mers. Upon consumption, lignan glycosides undergo deglycosyla-tion, being further converted to glucuronidated, methylated, and sulfated derivatives in the liver. These metabolites can be detected in the urine. However, the non-metabolized fraction of glycosylat-ed lignans is directed to the colon, where they are converted into enterolignans, enterolactone, and enterodiol by the gut microbiota. Evidence suggests that the presence of such metabolites in the gut stimulates the colonization of probiotic microorganisms, decreas-ing the risk of colorectal cancer. However, the exact mechanism for this effect has yet to be clarified (Zanella et al., 2017; Fu et al., 2020).
Among cereal grains, alkylresorcinols (AR) are abundantly present in wheat (489–642 mg/g) and rye (720–761 mg/g). To a lesser extent, they can also be found in barley, millet, and maize (Ross et al., 2003a). In this raw material, common AR homologues include 5-n-pentadecylresorcinol (C15:0), 5-n-heptadecylresorciol (C17:0), 5-n-nonadecylresorcinol (C19:0), 5-n-heneicosylresor-cinol (C21:0), 5-n-tricosylresorcinol (C23:0), and 5-n-pentaco-sylresorcinol (C25:0). Rye is predominantly composed of C17:0 (23–25%), C19:0 (31–32%), and C21:0 (22–25%) homologues, whereas C19:0 (29–35%) and C21:0 (46–51%) represent the ma-jor AR contribution to wheat (Ross et al., 2003a). According to Landberg et al. (2008), ARs are mainly located in the intermediate
layers (inner pericarp, hyaline layer, and testa) of the wheat ker-nel (16.2–16.4 mg/g), followed by the peripheral layers (3.9–4.0 mg/g).
Similar to other phenolic compounds, ARs have also been re-ported to possess antioxidant and antimicrobial activities. Besides, observational and in vitro evidence suggests the anticarcinogenic potential of this group. Reports have pointed out a risk reduction of around 52–66% of distal colon cancer when nanomolar concen-trations of ARs are present in the plasma. In addition, ARs have demonstrated high cytotoxicity toward cancerous cell lines (Kruk et al., 2017; Fu et al., 2018).
ARs have been indicated as possible biomarkers to correlate whole-grain cereals consumption with health benefits due to their high absorption rate in humans. Ross et al. (2003b) administered a high-AR diet based on the consumption of rye bran-enriched bread to ten ileostomy-operated subjects and verified that around 40% of the AR present in the diet were recovered in effluent from the small intestine, while 60% had been absorbed in the small intestine and further metabolized. The authors also suggested that medium-chain AR homologues (C17:0, C19:0) are taken up at a higher fre-quency than their long-chain counterparts (C23:0, C25:0).
4. Effect of processing on cereal phenolics
After harvest and before reaching the consumer’s table, cereals undergo a series of post-harvest processes that can vary according to the type of grain and specific destination. Firstly, the harvested grains are prepared for storage, being subsequently subjected to cleaning, husk removal, and size reduction (in some cases) in the stage known as primary processing. Then, cereals can be commer-cialized as such, or they can be transformed into value-added prod-ucts. This stage is known as secondary processing and is character-ized by the use of primary products (whole grains, flakes, or flour) as ingredients for breakfast cereals, snacks, cereal bars, doughs, batters, and noodles, among others. In secondary processing, vari-ous unit operations (e.g., puffing, flaking, fermentation, baking, extrusion) can be used alone or in combination (Food and Agricul-ture Organization, 2021).
Due to the drastic transformation that cereal grains undergo dur-ing this step, secondary processing is generally the focus of studies interested in the effect of processing on cereals’ bioactive com-pounds. However, evidence points out that even primary process-ing and post-harvest handling of cereals may affect their bioactive content and composition. Table 3 summarizes the possible effects of several cereal processing techniques on their phenolic composi-tion and bioactivities.
4.1. Post-harvest treatment and storage
Cereal harvesting usually begins before the grain is fully ripe and continues until mold and insect damage becomes extensive. The harvest is subsequently subjected to threshing, which is character-ized by removing the grains from the rest of the plant. Winnow-ing follows next, separating the grains from the chaff or straw. A crucial stage in cereal post-harvest treatment is the drying stage, responsible for reducing moisture content to 10–15%. This can happen through sun-drying, a cost-effective technique, or artifi-cial drying in humid locations. Finally, the dried grains are ready to be stored in bulk until further processing. During storage, they should be constantly inspected for spoilage and moisture content. Re-drying may be necessary in some cases (Schmidt et al., 2018).

Journal of Food Bioactives | www.isnff-jfb.com 45
Shahidi et al. Phenolic compounds in cereal grains and effects of processing
4.1.1. Cultivation and harvesting
Variations regarding the phenolic profile of cereals may take place even before harvesting begins. Climate conditions, agricultural practices, and soil quality play a role in the bioactive composition of such feedstock. This becomes particularly evident when study-ing the phenolic composition of a specific cereal grown in different locations. Emmons and Peterson (2001) reported that oats culti-vated at several locations in Wisconsin presented significant dif-ferences in their phenolic content and profile. Horvat et al. (2020) analyzed the phenolic acid content of wheat, corn, and barley over two crop years. Insoluble-bound phenolic acids (p-hydroxybenzo-ic, vanillic, caffeic, ferulic, syringic, p-coumaric, o-coumaric ac-ids) showed higher contents in the crop year where precipitation and temperature conditions were more extreme, due to a response to abiotic stress toward drastic environmental conditions. A similar outcome was observed for barley and corn, which biosynthesized more insoluble-bound phenolic acids during intense precipitation periods, a response to high water stress.
Additionally, the agronomic management of crops can also af-fect phenolics. According to Rempelos et al. (2018), using mineral NPK fertilizers causes the reduction of phenolic acid and flavonoid contents in wheat. Meanwhile, herbicides, fungicides, and growth regulators do not cause significant variations in the same param-eters. On the other hand, Stumpf et al. (2015) observed that enrich-ing the soil with nitrogen supply in wheat crops positively affects the concentration of free phenolics while reducing the amount of conjugated soluble phenolics and maintaining insoluble-bound phenolics within the same range.
Cultivation system has been found to influence phenolic yield. Fares et al. (2019) cultivated wheat genotypes under organic and traditional systems. Total phenolic content was 10% higher in the wheat grown under the organic system. However, when analyzing the individual contents of free vanillin and ferulic acid, as well as insoluble-bound vanillic, caffeic, syringic, p-coumaric, and ferulic acids, as well as vanillin, the authors did not find significant differ-ences between the organic and the conventional systems.
4.1.2. Drying
Setyaningsih et al. (2016) investigated how sun-drying affects phe-nolics in rice during the post-harvest stage. No major differences
were observed in the content of protocatechuic, vanillic, proto-catechuicaldehyde, p-hydroxybenzoic, p-hydroxybenzaldehyde, ferulic, sinapic, syringic, and p-coumaric acids, and vanillin be-tween the wet and the dried paddy. Ge et al. (2021) studied un-conventional drying techniques (infrared and hot air drying at 60 °C) on the post-harvest processing of naked barley. Samples were dried to 10% moisture. Infrared drying was found to preserve the phenolic compounds better, even leading to higher yields when compared to the original sample. This effect was due to the fact that kinetic energy is converted to thermal energy during infrared drying, releasing bound compounds. Although hot air drying also presented higher phenolic yields than the original sample, infrared drying was deemed more efficient in releasing bound phenolics while avoiding losses.
Wanyo et al. (2014) studied the impact of hot air drying (120 °C/30 min), far-infrared (FIR) radiation, and cellulase treatment on the phenolic content and antioxidant activity of rice bran and husk. Generally, cellulase-aided process and hot-air drying were able to preserve phenolics and their antioxidant activity, measured by 2,2-diphenyl-1-picrylhydrazyl (DPPH) and ferric reducing ability of plasma (FRAP) assays, with little or no significant difference in such parameters in regard to the raw materials. However, FIR man-aged to increase total phenolic and flavonoid contents as well as antioxidant capacity, which is similar to the outcome observed by Ge et al. (2021). Some interesting observations include the effect of cellulase, which significantly increased the content of vanillic acid while drastically reducing ferulic acid concentration.
4.1.3. Storage
Once post-harvest treatment has taken place, cereals can be stored. The storage period and conditions are also crucial for the overall quality of the grains and, especially, their bioactive compounds. Extreme pH, time, temperature, and UV exposure conditions should be avoided, as they may degrade the bioactive content and antioxidant power. Sripum et al. (2016) stored Thai Jasmine par-boiled germinated brown rice for six months under 30 and 40 °C. Neither temperature affected the total phenolic content and antiox-idant activity (measured by DPPH, oxygen free radical scavenging activity, and FRAP), as such parameters kept stable and without major variances during the six-month storage. The rice samples were maintained vacuum-sealed for the whole period, which could
Table 3. Effect of common cereal processing techniques on their phenolic composition
Cereal Process Effect Reference
Barley Infrared and hot air drying (60 °C)
Infrared drying released bound phenolics, increasing the phenolic content in regards to the hot air drying and original samples
Ge et al., 2021
Millet Dehulling Reduction of total phenolic content. Reduction of oxygen radical absorbance capacity and hydroxyl-radical scavenging capacity. Reduction of the concentration of p-coumaric and ferulic acids
Chandrasekara and Shahidi, 2011a, b
Rice Parboiling Release of bound phenolics, increasing the contents of p-coumaric, syringic, protocatechuic, and ferulic acids compared to raw sample
Thammapat et al., 2016
Wheat Steam explosion (215 °C for 120 s)
Increased concentration of soluble phenolic acids Liu et al., 2016
Brown rice, wheat, and oat
Extrusion cooking Enhanced content of insoluble-bound phenolics. Increased bioaccessibility of wheat phenolics
Zeng et al., 2016
Maize Roasting (17 min) Decreased total phenolic and flavonoid contents. Diminished free radical scavenging activity and iron chelating ability
Oboh et al., 2010

Journal of Food Bioactives | www.isnff-jfb.com46
Phenolic compounds in cereal grains and effects of processing Shahidi et al.
have contributed to this outcome. The lack of oxygen avoided the formation of oxidation products, leading to the preservation of phenolics and their antioxidant capacity.
De Oliveira et al. (2017) evaluated three storage temperatures (4, 25, and 40 °C) for sorghum grain and flour; they did not de-tect changes in the contents of 3-deoxyanthocyanins (luteolinidin, 5-methoxyluteolinidin, apigeninidin, and 7-methoxyapigeninidin) and total proanthocyanidins. On the other hand, storage time con-siderably affected sorghum’s phenolics. Up to 60 days of storage, most phenolics in sorghum showed a reduction in their content, stabilizing after this period. After 180 days, 86.7–86.8% of all phe-nolics were retained. The proanthocyanidins were the most affect-ed group, retaining only about 56.6–85.3% after the same period.
4.2. Primary processing
Prior to advanced processing, cereals grains are cleaned and grad-ed based on their size. Depending on the type of cereal, dehulling, which consists of the removal of unpalatable hulls or shells, can be necessary. If grains are used as primary material for flour produc-tion, a milling step will be conducted. This can generally happen in a plate, hammer, or roller mill, according to grain type and pro-duction scale. A common process for rice, parboiling is an optional step, and it involves pre-cooking the grain by soaking and heating it, loosening the hulls, and aiding in grain preservation and cook-ing efficiency. In addition, for white rice production, a polishing stage is required. Although popular in some cultures, nutritionally speaking, white rice is less appealing than its brown counterpart due to the removal of the bran and the germ. These fractions con-centrate the highest proportion of bioactive compounds, including phenolics, while the endosperm is mainly constituted by carbohy-drates (Schmidt et al., 2018; Food and Agriculture Organization, 2021).
4.2.1. Dehulling and parboiling
Dehulling is the process of removing the outer layers of grains, and this is usually carried out as a part of the milling process (Duodu, 2011). This technique reduces the content of phenolic compounds, negatively impacting the antioxidant activity of processed grains (Duodu, 2011). Živković et al. (2021) conducted cold dehulling on germinated buckwheat seeds and analyzed its effect on phenolic profile and antioxidant activity. Regardless of germination time, the dehulled buckwheat showed a sharp decrease in all identified compounds’ content compared to the hulled sample. Some insolu-ble-bound phenolics quantified in the hulled samples could not be detected in their dehulled counterparts, namely orientin, rutin, and hyperin. As expected, antioxidant capacity measured by DPPH de-clined considerably once the seeds were dehulled. The same trend was observed by Dlamini et al. (2007) in sorghum.
Chandrasekara and Shahidi (2011a, b) detected that Pearl millet significantly reduced its total phenolic content, negatively affect-ing the phenolics’ bioactivities (e.g., oxygen radical absorbance capacity (ORAC) and hydroxyl-radical scavenging capacity). Be-sides, two of the major phenolic acids present in millet, ferulic and p-coumaric acid, had their concentrations diminished by 39 and 52%, respectively. A similar study by Madhujith et al. (2006) was carried out in pearled barley fractions. The outermost frac-tions concentrated most phenolic compounds (TPC of 4.16–6.26 mg FAE/g), dropping considerably as the innermost fractions were reached (TPC of 0.17–0.51 mg FAE/g). Additionally, free caffeic and p-coumaric acids were only detected in the outermost fractions
of the grains.Alkylresorcinols are also affected by the dehulling process. Ac-
cording to Giambanelli et al. (2018), whole wheat kernels showed a higher alkylresorcinol content than their hulled counterpart. Af-ter the pearling process, the authors analyzed the waste fractions in order to determine where the phenolic lipids were mostly concen-trated and found that the bran was the main location of akylresor-cinols. This fraction was also the one showing the highest antioxi-dant capacity. Wheat bran was determined to be rich in unsaturated alkylresorcinols homologues, mainly C18:1 and C18:2.
In contrast, parboiling has been reported to improve phenolic concentration considerably. Thammapat et al. (2016) compared two parboiling methods for glutinous rice. The traditional method consisted of soaking the rice in water for 24 h at room temperature, followed by steaming at 100 °C for 1 h and sun-drying for 12–14 h. In the alternative method, soaking was conducted for only 6 h at 60 °C, instead of room temperature. Also, the steaming temperature was 120 °C for 15 min, followed by cabinet-drying at 50 °C for 12–14 h. Both parboiled rice samples exhibited higher content of phenolic acids (traditional – 52.24 µg/g, alternative – 65.39 µg/g) than raw rice (45.94 µg/g), with p-coumaric, syringic, protocat-echuic, and ferulic acids as the compounds showing the sharpest increase. According to the authors, a possible explanation for this effect could be the release of bound phenolics due to the thermal treatment. Similar outcomes have been observed for red rice (Hu et al., 2017) and millet (Bora et al., 2019).
4.3. Secondary processing
Secondary processing of cereals involves operations responsible for further transforming the grains into more palatable and digest-ible forms or using them as ingredients for added-value products. A vast array of techniques can be employed at this stage, such as extrusion, fermentation, high-pressure processing, steam explo-sion, and roasting.
4.3.1. Steam explosion and high-pressure processing
Steam explosion can be used for the pre-treatment of cereals in or-der to release bound phenolics (Baruah et al., 2018; Gong et al., 2012). During this treatment, phenolic compounds are exposed to high-pressure steam (0.69–4.83 MPa) and high temperature rang-ing from 160 to 260 °C for only a few seconds/minutes (Baruah et al., 2018; Rabemanolontsoa and Saka, 2016; Gong et al., 2012). After subjecting the cereal matrix to high pressure, a sudden decline in pressure results in the explosion and releases water, thus lead-ing to the disruption of the cell wall structure (Baruah et al., 2018; Singh et al., 2015). Throughout the process, acetic acid is liber-ated from xylan hemicelluloses, which catalyzes further reactions, such as the hydrolysis of hemicellulose. Therefore, this treatment is called autohydrolysis (Baruah et al., 2018; Rabemanolontsoa and Saka, 2016). The advantages of steam explosion include low en-vironmental impact and low cost as it needs limited chemicals in a simplified process (Baruah et al., 2018; Pielhop et al., 2016).
The efficiency of steam explosion can be affected by steam tem-perature, residence time, sample size, and moisture content (Ba-ruah et al., 2018; Rabemanolontsoa and Saka, 2016). According to Rabemanolontsoa and Saka (2016), lower temperatures and longer residence times are more favorable for extracting bound phenolics. Under drastic temperature conditions, solubilized carbohydrates such as hemicellulose undergo further secondary reactions to form carboxylic acids and soluble polymers (Gong et al., 2012; Sun et

Journal of Food Bioactives | www.isnff-jfb.com 47
Shahidi et al. Phenolic compounds in cereal grains and effects of processing
al., 2005). In addition, reactive precursors cause condensation with lignin, resulting in a lower phenolic yield (Sun et al., 2005). Gong et al. (2012) studied the effect of steam explosion at 220 °C for 10–120 s on phenolic compounds in Tibetan barley bran (dehulled). This study has shown that the steam explosion pre-treatment ef-fectively decomposes carbohydrates in this cereal, which results in a high yield of total free phenolic acids. Total antioxidant capacity was also increased at 220 °C for 120 s (Gong et al., 2012).
Similarly, the amount of soluble phenolic acids in wheat bran and their antioxidant properties were significantly increased after steam explosion at 215 °C for 120 s (Liu et al., 2016). A recent study has shown that steam pressure may also be an important pa-rameter, and milder conditions are more effective for increasing the yield of phenolics, contributing to improved bioactivities. Li et al. (2020) reported that steam explosion enhanced the anti-prolif-erative activity of phenolics in Tartary buckwheat bran on human colon and liver cancer cells. These results highlight the potential of steam explosion as an effective pre-treatment to promote enhanced bioaccessibility of bound phenolics from cereals and other plant foods.
Other technologies have also been investigated for the release of bound phenolics from cereals. Luo et al. (2020) used high-pres-sure boiling on sorghum hull. This method was able to increase the content of free procyanidins and total phenolics by 35.92–58.87 and 6.20–18.70%, respectively, compared to traditional hot water processing. The disruption of bound phenolics caused by the com-bination of high temperature and pressure, along with the inactiva-tion of endogenous polyphenol oxidase, prevented the oxidation and degradation of phenolics.
4.3.2. Extrusion cooking
Extrusion cooking is largely used in the food industry to manu-facture pasta, noodles, and cereal-based snacks. This continuous process uses a combination of high temperature, high pressure, and high shearing to plasticize the raw material so it can be conveyed and forced to flow through a die or specific shape. In terms of phe-nolics, heat-sensitive compounds may be susceptible to degrada-tion and polymerization, decreasing their concentration in the final product. On the other hand, extrusion also causes the disruption of cell walls, breaking the covalent bonds between bound pheno-lics and macromolecules, which results in an increased bioacces-sibility of this phenolic fraction (Wang et al., 2014). Zeng et al. (2016) studied the effect of extrusion on brown rice, wheat, and oat phenolics and their bioaccessibility. All cereal grains had their insoluble-bound phenolic content significantly increased after the extrusion cooking. The bioaccessibility varied according to the grain type. While 11.60% of phenolics from raw wheat were bioac-cessible, extrusion marginally improved it to 12.48%. On the other hand, the bioaccessibility of oat phenolics dropped from 29.83% (raw) to 21.78% (extruded). Brown rice kept the same bioacces-sibility levels before and after extrusion (around 20%). According to the authors, phenolic bioaccessibility was mainly dependent on the cereal type and the way the compounds occur in the matrix since higher phenolic content did not translate into higher bioac-cessibility.
4.3.3. Fermentation, roasting, and popping
Biological treatments, such as fermentation, can also be used to enhance the bioefficiency of cereal phenolics. During this process, enzymes are produced and released, promoting the hydrolysis of
phenolic glycosides, liberating them as free compounds (Wang et al., 2014). Bvochora et al. (2005) reported an increase in the total phenolic content of sorghum during alcoholic fermentation; this brought about higher concentrations of p-hydroxybenzoic acid and p-hydroxybenzyl alcohol. Meanwhile, the levels of p-hydroxyben-zaldehyde decreased.
Total phenolic content of buckwheat, barley, wheat, and rye has been shown to increase after fermentation with Lactobacil-lus rhamnosus (L. rhamnosu) and Saccharomyces cerevisiae (S. cerevisiae) (Đorđević et al., 2010). A recent study on millet has also shown that fermentation with Lactobacilli increased the to-tal phenolic content due to the release of bound polyphenols. The fermentation treatment also enhanced the in vitro effect of millet phenolics on type 2 diabetes markers (Balli et al., 2020).
Roasting is also commonly used in the processing of several cereals, including maize and sorghum. Oboh et al. (2010) roasted yellow and white maize for 17 min and observed a significant de-crease in their total phenolic and flavonoid contents, which was re-flected in the lower free radical scavenging and iron chelating abil-ity of the samples compared to the raw cereal. Irondi et al. (2019) obtained similar results when studying the roasting of red sorghum at 150 and 180 °C for 20 min. Besides reducing the levels of phe-nolic compounds, enzyme inhibitory activities (pancreatic lipase, alpha-amylase, alpha-glucosidase, xanthine oxidase, and angio-tensin 1-converting enzyme) were slightly reduced in comparison to the original sample, with the roasting at 180 °C displaying the most negative effect. These outcomes can be explained by the fact that the high temperature during the roasting process can lead to thermal decomposition and heat-induced oxidation of phenolics.
Popping is used to expand cereals and promote starch gelatini-zation. In this process, grains are subjected to a high-temperature treatment for a short time, which creates superheated vapor inside the grains. The internal pressure causes the endosperm to expand, breaking the outer layer. This process imparts desirable sensory characteristics to cereal-based snacks, and it is commonly used to produce popcorn, popped rice, and popped sorghum, among others (Mishra et al., 2014). Bagchi et al. (2021) analyzed how different processing techniques, including popping, affect phenolic acids, flavonoids, and anthocyanins in black rice. Hydroxybenzoic ac-ids were the predominant phenolic acid class in rice following the popping process, with 75.78 µg/g. Gallic acid was the major phe-nolic acid detected in popped rice, followed by protocatechuic and vanillic acids. Moreover, 83% of all polyphenols quantified in the product were in the free form. In terms of flavonoids, kaempferol and catechin showed high concentrations in popped rice, with 9.19 µg/g and 8.38 µg/g, respectively. Anthocyanins were the main phe-nolic group found in popped rice, with an average of 119.4 µg/g, higher than the content found in beaten and milled rice but lower than the anthocyanin levels in boiled and puffed rice. Among all processes, popping produced the rice product with the highest antioxidant activity, measured by DPPH and FRAP assays. This outcome was due to the fact that the thermal treatment in popping (177 °C for 40–50 s) was not drastic enough to degrade the phe-nolic compounds.
5. Conclusions and final remarks
The economic and nutritional importance of cereal grains is unde-niable. They provide a great portion of the carbohydrate, protein, lipid, vitamins, and minerals consumed in the diet of many popu-lations, helping to fulfill the global food demand. Nevertheless, the consumption of cereals is also advantageous from a bioactive

Journal of Food Bioactives | www.isnff-jfb.com48
Phenolic compounds in cereal grains and effects of processing Shahidi et al.
standpoint, with phenolic compounds assuming the role of provid-ing benefits due to their antioxidant power and related bioactivi-ties.
Notwithstanding, cereals undergo post-harvest treatments, pri-mary, and in some cases, secondary processing before reaching the consumer’s table. Thus, different processing techniques show dis-tinct effects on cereal phenolics. Therefore, such factors should be analyzed in a case-by-case manner. Processes that remove the out-er layers of cereal grains, such as polishing of rice and dehulling of wheat and other cereals, significantly reduce the phenolic content due to the exclusion of fractions that are highly concentrated in these compounds. On the other hand, mild drying techniques and technologies involving the use of high-pressure can enhance the phenolic composition of cereals since they promote the release of the insoluble-bound phenolic fraction, increasing bioaccessibility parameters. Finally, drastic processing, such as extrusion cooking and roasting, may negatively affect cereal phenolics, considerably reducing their biological activities, but may enhance consumer acceptability due to generation of desirable aroma compounds as well as Maillard reaction products that may possess their own ac-tivities.
Therefore, cereal consumption, especially in their whole-grain form, should be stimulated given their large variety of polyphenols, including phenolic acids, flavonoids, tannins, and anthocyanins (pigmented cereals) that are associated with reduced risk of devel-oping chronic diseases, such as colorectal cancer, cardiovascular ailments, and type 2 diabetes. In addition, processing conditions must be carefully analyzed in order to increase the bioefficiency of cereal phenolics and avoid their degradation.
Acknowledgments
We thank the Natural Science and Engineering Research Council (NSERC) of Canada for support in the form of a Discovery Grant to FS.
References
Adom, K.K., and Liu, R.H. (2002). Antioxidant activity of grains. J. Agric. Food Chem. 50(21): 6182–6187.
Bagchi, T.B., Chattopadhyay, K., Sivashankari, M., Roy, S., Kumar, A., Biswas, T., and Pal, S. (2021). Effect of different processing technolo-gies on phenolic acids, flavonoids and other antioxidants content in pigmented rice. J. Cereal Sci. 100: 103263.
Balli, D., Bellumori, M., Pucci, L., Gabriele, M., Longo, V., Paoli, P., Melani, F., Mulinacci, N., and Innocenti, M. (2020). Does fermentation really increase the phenolic content in cereals? A study on millet. Foods 9(3): 303.
Baniwal, P., Mehra, R., Kumar, N., Sharma, S., and Kumar, S. (2021). Cere-als: Functional constituents and its health benefits. Pharma. Innov. 10(3): 01–07.
Baruah, J., Nath, B.K., Sharma, R., Kumar, S., Deka, R.C., Baruah, D.C., and Kalita, E. (2018). Recent Trends in the Pretreatment of Lignocellulosic Biomass for Value-Added Products. Front. Energy Res. 6: 141.
Bhattacharya, A., Sood, P., and Citovsky, V. (2010). The roles of plant phe-nolics in defence and communication during Agrobacterium and Rhizobium infection. Mol. Plant Pathol. 11(5): 705–719.
Bora, P., Ragaee, S., and Marcone, M. (2019). Effect of parboiling on decor-tication yield of millet grains and phenolic acids and in vitro digest-ibility of selected millet products. Food Chem. 274: 718–725.
Bvochora, J.M., Danner, H., Miyafuji, H., Braun, R., and Zvauya, R. (2005). Variation of sorghum phenolic compounds during the preparation of opaque beer. Process Biochem. 40(3-4): 1207–1213.
Chandrasekara, A., and Shahidi, F. (2011a). Determination of antioxidant
activity in free and hydrolyzed fractions of millet grains and charac-terization of their phenolic profiles by HPLC-DAD-ESI-MSn. J. Funct. Foods 3(3): 144–158.
Chandrasekara, A., and Shahidi, F. (2011b). Antiproliferative potential and DNA scission inhibitory activity of phenolics from whole millet grains. J. Funct. Foods 3(3): 159–170.
Chandrasekara, A., and Shahidi, F. (2012). Antioxidant phenolics of mil-let control lipid peroxidation in human LDL cholesterol and food sys-tems. J. Am. Oil Chem. Soc. 89(2): 275–285.
Chawda, P.J., Shi, J., Xue, S., and Young Quek, S. (2017). Co-encapsulation of bioactives for food applications. FQS 1(4): 302–309.
Chen, J., Yang, J., Ma, L., Li, J., Shahzad, N., and Kim, C.K. (2020). Structure-antioxidant activity relationship of methoxy, phenolic hydroxyl, and carboxylic acid groups of phenolic acids. Sci. Rep. 10(1): 1–9.
Cheynier, V., Comte, G., Davies, K.M., Lattanzio, V., and Martens, S. (2013). Plant phenolics: Recent advances on their biosynthesis, genetics, and ecophysiology. Plant Physiol. Biochem. 72: 1–20.
De Oliveira, K.G., Queiroz, V.A.V., de Almeida Carlos, L., de Morais Cardoso, L., Pinheiro-Sant’Ana, H.M., Anunciação, P.C., Menezes, C.B., da Silva, E.C., and Barros, F. (2017). Effect of the storage time and tempera-ture on phenolic compounds of sorghum grain and flour. Food Chem. 216: 390–398.
Dlamini, N.R., Taylor, J.R., and Rooney, L.W. (2007). The effect of sorghum type and processing on the antioxidant properties of African sor-ghum-based foods. Food Chem. 105(4): 1412–1419.
Đorđević, T.M., Šiler-Marinković, S.S., and Dimitrijević-Branković, S.I. (2010). Effect of fermentation on antioxidant properties of some ce-reals and pseudo cereals. Food Chem. 119(3): 957–963.
Duodu, K.G. (2011). Effects of processing on antioxidant phenolics of ce-real and legume grains. Advances in Cereal Science: Implications to Food Processing and Health Promotion. American Chemical Society, pp. 31–54.
Dykes, L., and Rooney, L.W. (2006). Sorghum and millet phenols and anti-oxidants. J. Cereal Sci. 44(3): 236–251.
Emmons, C.L., and Peterson, D.M. (2001). Antioxidant activity and phe-nolic content of oat as affected by cultivar and location. Crop Sci. 41(6): 1676–1681.
Fares, C., Menga, V., Codianni, P., Russo, M., Perrone, D., Suriano, S., Savino, M., and Rascio, A. (2019). Phenolic acids variability and grain quality of organically and conventionally fertilised old wheats under a warm climate. J. Sci. Food Agric. 99(10): 4615–4623.
Food and Agriculture Organization. (2020). OECD-FAO Agricultural out-look 2020-2029. https://www.oecd-ilibrary.org/sites/57d27093-en/index.html?itemId=/content/component/57d27093-en. Accessed 09/10/2021.
Food and Agriculture Organization. (2021). Post-harvesting process. http://www.fao.org/3/au104e/au104e.pdf. Accessed 10 Sep. 2021.
Fu, J., Soroka, D.N., Zhu, Y., and Sang, S. (2018). Induction of apoptosis and cell-cycle arrest in human colon-cancer cells by whole-grain alky-lresorcinols via activation of the p53 pathway. J. Agric. Food Chem. 66(45): 11935–11942.
Fu, J., Zhang, Y., Hu, Y., Zhao, G., Tang, Y., and Zou, L. (2020). Concise re-view: Coarse cereals exert multiple beneficial effects on human health. Food Chem. 325: 126761.
Ge, X., Saleh, A.S., Jing, L., Zhao, K., Su, C., Zhang, B., Zhang, Q., and Li, W. (2021). Germination and drying induced changes in the composition and content of phenolic compounds in naked barley. J. Food Compos. Anal. 95: 103594.
Giambanelli, E., Ferioli, F., and D’Antuono, L.F. (2018). Retention of alkyl-resorcinols, antioxidant activity and fatty acids following traditional hulled wheat processing. J. Cereal Sci. 79: 98–105.
Gong, L., Huang, L., and Zhang, Y. (2012). Effect of Steam Explosion Treat-ment on Barley Bran Phenolic Compounds and Antioxidant Capacity. J. Agric. Food Chem. 60(29): 7177–7184.
Horvat, D., Šimić, G., Drezner, G., Lalić, A., Ledenčan, T., Tucak, M., Plavsic, H., Andric, L., and Zdunić, Z. (2020). Phenolic acid profiles and anti-oxidant activity of major cereal crops. Antioxidants 9(6): 527.
Hu, Z., Tang, X., Liu, J., Zhu, Z., and Shao, Y. (2017). Effect of parboiling on phytochemical content, antioxidant activity and physicochemical properties of germinated red rice. Food Chem. 214: 285–292.
Irondi, E.A., Adegoke, B.M., Effion, E.S., Oyewo, S.O., Alamu, E.O., and Bo-

Journal of Food Bioactives | www.isnff-jfb.com 49
Shahidi et al. Phenolic compounds in cereal grains and effects of processing
ligon, A.A. (2019). Enzymes inhibitory property, antioxidant activity and phenolics profile of raw and roasted red sorghum grains in vitro. Food Sci. Hum. Wellness 8(2): 142–148.
Jonnala, R.S., Irmak, S., MacRitchie, F., and Bean, S.R. (2010). Phenolics in the bran of waxy wheat and triticale lines. J. Cereal Sci. 52(3): 509–515.
Koehler, P., and Wieser, H. (2013). Chemistry of cereal grains. Handbook on sourdough biotechnology. Springer, Boston, MA, pp. 11–45.
Kruk, J., Aboul-Enein, B., Bernstein, J., and Marchlewicz, M. (2017). Die-tary alkylresorcinols and cancer prevention: A systematic review. Eur. Food Res. Technol. 243(10): 1693–1710.
Landberg, R., Kamal-Eldin, A., Salmenkallio-Marttila, M., Rouau, X., and Åman, P. (2008). Localization of alkylresorcinols in wheat, rye and barley kernels. J. Cereal Sci. 48(2): 401–406.
Li, W., Zhang, X., He, X., Li, F., Zhao, J., Yin, R., and Ming, J. (2020). Effects of steam explosion pretreatment on the composition and biological activities of tartary buckwheat bran phenolics. Food Funct. 11(5): 4648–4658.
Liu, J., Zhang, J., Wang, W., and Hou, H. (2021). Effects of microwave treat-ment on the stability and antioxidant capacity of a functional wheat bran. Food Sci. Nutr. 9(5): 2713–2721.
Liu, L., Zhao, M., Liu, X., Zhong, K., Tong, L., Zhou, X. and Zhou, S. (2016). Effect of steam explosion-assisted extraction on phenolic acid pro-files and antioxidant properties of wheat bran. J. Sci. Food and Agr. 96(10): 3484-3491.
Liyana-Pathirana, C.M., and Shahidi, F. (2006). Importance of insoluble-bound phenolics to antioxidant properties of wheat. J. Agric. Food Chem. 54(4): 1256–1264.
Luo, M., Hou, F., Dong, L., Huang, F., Zhang, R., and Su, D. (2020). Compari-son of microwave and high-pressure processing on bound phenolic composition and antioxidant activities of sorghum hull. Int. J. Food Science Technol. 55(9): 3190–3202.
Madhujith, T., and Shahidi, F. (2009). Antioxidant potential of barley as affected by alkaline hydrolysis and release of insoluble-bound phe-nolics. Food Chem. 117(4): 615–620.
Madhujith, T., Izydorczyk, M., and Shahidi, F. (2006). Antioxidant proper-ties of pearled barley fractions. J. Agric. Food Chem. 54(9): 3283–3289.
Mishra, G., Joshi, D.C., and Panda, B.K. (2014). Popping and puffing of ce-real grains: a review. J. Grain Processing Storage 1(2): 34–46.
Naczk, M., and Shahidi, F. (2006). Phenolics in cereals, fruits and vegeta-bles: Occurrence, extraction and analysis. J. Pharm. Biomed. Anal. 41(5): 1523–1542.
Oboh, G., Ademiluyi, A.O., and Akindahunsi, A.A. (2010). The effect of roasting on the nutritional and antioxidant properties of yellow and white maize varieties. Int. J. Food Sci. 45(6): 1236–1242.
Pang, Y., Ahmed, S., Xu, Y., Beta, T., Zhu, Z., Shao, Y., and Bao, J. (2018). Bound phenolic compounds and antioxidant properties of whole grain and bran of white, red and black rice. Food Chem. 240: 212–221.
Pielhop, T., Amgarten, J., Von Rohr, P.R., and Studer, M.H. (2016). Steam explosion pretreatment of softwood: The effect of the explosive decompression on enzymatic digestibility. Biotechnol. Biofuels 9(1): 152–152.
Pihlava, J.M., Hellström, J., Kurtelius, T., and Mattila, P. (2018). Flavonoids, anthocyanins, phenolamides, benzoxazinoids, lignans and alkylresor-cinols in rye (Secale cereale) and some rye products. J. Cereal Sci. 79: 183–192.
Rabemanolontsoa, H., and Saka, S. (2016). Various pretreatments of ligno-cellulosics. Bioresour. Technol. 199: 83–91.
Raheem, D., Dayoub, M., Birech, R., and Nakiyemba, A. (2021). The Contri-bution of Cereal Grains to Food Security and Sustainability in Africa: Potential Application of UAV in Ghana, Nigeria, Uganda, and Namib-ia. Urban Sci. 5(1): 8.
Rao, S., Santhakumar, A.B., Chinkwo, K.A., and Blanchard, C.L. (2020). In-vestigation of phenolic compounds with antioxidant activity in bar-ley and oats affected by variation in growing location. Cereal Chem. 97(4): 772–782.
Rempelos, L., Almuayrifi, A.M., Baranski, M., Tetard-Jones, C., Eyre, M., Shotton, P., Cakmak, I., Ozturk, L., Cooper, J., Volakakis, N., Schmidt, C., Sufar, E., Wang, J., Wilkinson, A., Rosa, E.A.S., Zhao, B., Rose, T.J.,
Leifert, C., and Bilsborrow, P. (2018). Effects of agronomic manage-ment and climate on leaf phenolic profiles, disease severity, and grain yield in organic and conventional wheat production systems. J. Agric. Food Chem. 66(40): 10369–10379.
Rodehutscord, M., Rückert, C., Maurer, H.P., Schenkel, H., Schipprack, W., Bach Knudsen, K.E., Schollenberger, M., Laux, M., Eklund, M., Siegert, W., and Mosenthin, R. (2016). Variation in chemical composition and physical characteristics of cereal grains from different genotypes. Arch. Anim. Nutr. 70(2): 87–107.
Rodríguez-García, C., Sánchez-Quesada, C., Toledo, E., Delgado-Rodríguez, M., and Gaforio, J.J. (2019). Naturally lignan-rich foods: A dietary tool for health promotion? Mol. 24(5): 917.
Ross, A.B., Kamal-Eldin, A., Lundin, E.A., Zhang, J.X., Hallmans, G., and Åman, P. (2003b). Cereal alkylresorcinols are absorbed by humans. J. Nutr. 133(7): 2222–2224.
Ross, A.B., Shepherd, M.J., Schüpphaus, M., Sinclair, V., Alfaro, B., Kamal-Eldin, A., and Åman, P. (2003a). Alkylresorcinols in cereals and cereal products. J. Agric. Food Chem. 51(14): 4111–4118.
Schmidt, M., Zannini, E., and Arendt, E.K. (2018). Recent advances in phys-ical post-harvest treatments for shelf-life extension of cereal crops. Foods 7(4): 45.
Serna-Saldivar, S.O. (2016). Cereal grains: properties, processing, and nu-tritional attributes. CRC press.
Setyaningsih, W., Hidayah, N., Saputro, I.E., Palma, M., and Barroso, C.G. (2016). Profile of phenolic compounds in Indonesian rice (Oryza sa-tiva) varieties throughout post-harvest practices. J. Food Compos. Anal. 54: 55–62.
Shahidi, F. (2009). Nutraceuticals and functional foods: whole versus pro-cessed foods. Trends Food Sci. Technol. 20(9): 376–387.
Shahidi, F., and Naczk, M. (2003). Phenolics in food and nutraceuticals. CRC press.
Shahidi, F., and Yeo, J. (2016). Insoluble-bound phenolics in food. Mol. 21(9): 1216.
Shahidi, F., Varatharajan, V., Oh, W.Y., and Peng, H. (2019). Phenolic com-pounds in agri-food by-products, their bioavailability and health ef-fects. J. Food Bioact. 5(1): 57–119.
Shao, Y., Hu, Z., Yu, Y., Mou, R., Zhu, Z., and Beta, T. (2018). Phenolic acids, anthocyanins, proanthocyanidins, antioxidant activity, minerals and their correlations in non-pigmented, red, and black rice. Food Chem. 239: 733–741.
Sidhu, J.S., Kabir, Y., and Huffman, F.G. (2007). Functional foods from ce-real grains. Int. J. Food Prop. 10(2): 231–244.
Singh, J., Suhag, M., and Dhaka, A. (2015). Augmented digestion of ligno-cellulose by steam explosion, acid and alkaline pretreatment meth-ods: A review. Carbohydr. Polym. 117: 624–631.
Sripum, C., Kukreja, R.K., Charoenkiatkul, S., Kriengsinyos, W., and Sutti-sansanee, U. (2016). The effect of storage conditions on antioxidant activities and total phenolic contents of parboiled germinated brown rice (Khao Dok Mali 105). Int. Food Res. J. 23(4): 1827.
Statista. (2021). Worldwide production of grain in 2020/21, by type. htt-ps://www.statista.com/statistics/263977/world-grain-production-by-type/ Accessed 10 Sep. 2021.
Stumpf, B., Yan, F., and Honermeier, B. (2015). Nitrogen fertilization and maturity influence the phenolic concentration of wheat grain (Triti-cum aestivum). J. Soil Sci. 178(1): 118–125.
Sun, X.F., Xu, F., Sun, R.C., Geng, Z.C., Fowler, P., and Baird, M.S. (2005). Characteristics of degraded hemicellulosic polymers obtained from steam exploded wheat straw. Carbohydr. Polym. 60(1): 15–26.
Suriano, S., Balconi, C., Valoti, P., and Redaelli, R. (2021). Comparison of total polyphenols, profile anthocyanins, color analysis, carotenoids and tocols in pigmented maize. LWT 144: 111257.
Thammapat, P., Meeso, N., and Siriamornpun, S. (2016). Effects of the tra-ditional method and an alternative parboiling process on the fatty acids, vitamin E, γ-oryzanol and phenolic acids of glutinous rice. Food Chem. 194: 230–236.
Vaher, M., Matso, K., Levandi, T., Helmja, K., and Kaljurand, M. (2010). Phe-nolic compounds and the antioxidant activity of the bran, flour and whole grain of different wheat varieties. Procedia Chem. 2(1): 76–82.
Wang, T., He, F., and Chen, G. (2014). Improving bioaccessibility and bio-availability of phenolic compounds in cereal grains through process-ing technologies: A concise review. J. Funct. Foods 7: 101–111.

Journal of Food Bioactives | www.isnff-jfb.com50
Phenolic compounds in cereal grains and effects of processing Shahidi et al.
Wanyo, P., Meeso, N., and Siriamornpun, S. (2014). Effects of different treatments on the antioxidant properties and phenolic compounds of rice bran and rice husk. Food Chem. 157: 457–463.
Yu, S., and Tian, L. (2018). Breeding major cereal grains through the lens of nutrition sensitivity. Mol. Plant 11(1): 23–30.
Zanella, I., Biasiotto, G., Holm, F., and di Lorenzo, D. (2017). Cereal lignans, natural compounds of interest for human health? Nat. Prod. Com-mun. 12(1): 139–146.
Zeng, Z., Hu, X., McClements, D.J., Luo, S., Liu, C., Gong, E., and Huang, K. (2019). Hydrothermal stability of phenolic extracts of brown rice. Food Chem. 271: 114–121.
Zeng, Z., Liu, C., Luo, S., Chen, J., and Gong, E. (2016). The profile and bioaccessibility of phenolic compounds in cereals influenced by im-
proved extrusion cooking treatment. PloS One 11(8): e0161086.Zhang, Q., de Mejia, E.G., Luna-Vital, D., Tao, T., Chandrasekaran, S.,
Chatham, L., Juvik, J., Singh, V., and Kumar, D. (2019). Relationship of phenolic composition of selected purple maize (Zea mays L.) geno-types with their anti-inflammatory, anti-adipogenic and anti-diabetic potential. Food Chem. 289: 739–750.
Zhou, Z., Chen, X., Zhang, M., and Blanchard, C. (2014). Phenolics, flavo-noids, proanthocyanidin and antioxidant activity of brown rice with different pericarp colors following storage. J. Stored Prod. Res. 59: 120–125.
Živković, A., Polak, T., Cigić, B., and Požrl, T. (2021). Germinated Buck-wheat: Effects of Dehulling on Phenolics Profile and Antioxidant Ac-tivity of Buckwheat Seeds. Foods 10(4): 740.

Copyright: © 2021 International Society for Nutraceuticals and Functional Foods.All rights reserved.
51
Review J. Food Bioact. 2021;15:51–62
Journal ofFood Bioactives International Society for
Nutraceuticals and Functional Foods
Potential adverse health effects of dietary lipid oxidation products
Yuanyuan Hua, Guanhua Zhaoa, Min Zhanga, Dayong Zhoua,b,c* and Beiwei Zhua,b,c*
aSchool of Food Science and Technology, Dalian Polytechnic University, Dalian 116034, PR ChinabNational Engineering Research Center of Seafood, Dalian 116034, PR ChinacCollaborative Innovation Center of Seafood Deep Processing, Dalian Polytechnic University, Dalia 116034, PR China*Corresponding author: Dayong Zhou and Beiwei Zhu, School of Food Science and Technology, Dalian Polytechnic University, Dalian 116034, PR China. Tel: +86-0411 86323453 (BZ), +86-0411 86313453 (DZ); E-mail: [email protected] (DZ), [email protected] (BZ)DOI: 10.31665/JFB.2021.15282Received: July 17, 2021; Revised received & accepted: August 11, 2021Abbreviations: Aβ, amyloid β-protein; ACE, angiotensin converting enzyme; ACh, acetylcholine; AD, Alzheimer’s disease; ADAM, A-disintegrin and metalloprotease; ALDH2, aldehyde dehydrogenase-2; ALS, amyotrophic lateral sclerosis; ApoE-/-, apolipoprotein E-defi-cient; APP, amyloid precursor protein; AST, aspartate aminotransferase; ATP, adenosine-triphosphate; BACE-1, beta-secretase 1; BCL-2, B-cell lymphoma 2; BDNF, brain-derived neurotrophic factor; BK, large-conductance calcium-dependent potassium; CAT, catalase; CCK, cholecystokinin; CoA, acyl coenzyme A; COPs, cholesterol oxidation products; CYP, cytochrome P450; dGuo, 2′-deoxyguanosine; DNA, deoxyribonucleic acid; FAS, fatty acid synthase; G6PDH, glucose-6-phosphate dehydrogenase; GPR183, G protein-coupled receptor 183; GPx, glutathione peroxidases; GSH, glutathione; HD, Hungtington’s disease; HHE, 4-hydroxy-2-hexenal; HNE, 4-hydroxy-2-nonenal; KATP, adenosine triphosphate-sensitive potassium; 7-KC, 7-ketocholesterol; LDL, low-density lipoprotein; LOPs, lipid oxidation products; MAPK, mitogen-activated protein kinase; MDA, malondialdehyde; NF-κB, nuclear factor kappa light chain enhancer of activated B cells; NO, nitric oxide; 24-OH, 24-hydroxycholesterol; 27-OH, 27-hydroxycholesterol; PD, Parkinson; PE, phenylephrine; PKC, protein kinase C; PPARα, peroxisome proliferator-activated receptor α; PUFAs, polyunsaturated fatty acids; ROS, reactive oxygen species; SNP, sodium ni-troprusside; SREBP, sterol regulatory element binding protein; t-2-DCA, trans-2-decenal; tt-DDE: trans, trans-2,4-decadienal; tt-2,4-NDE, trans-trans-2,4-nonadienal; TrkA, tropomyosin receptor kinases A; TrkB, tyrosine Kinase receptor B; t-2-UDA, trans-2-undecenal; VLDL, very low-density lipoproteins; ZO-1, occlusion-1Citation: Hu, Y., Zhao, G., Zhang, M., Zhou, D., and Zhu, B. (2021). Potential adverse health effects of dietary lipid oxidation products. J. Food Bioact. 15: 51–62.
Abstract
Lipid oxidation products (LOPs) are widely present in many lipid- containing foods. They usually enter the gastro-intestinal tract from dietary sources and/or are produced in vivo. Part of LOPs were absorbed into the blood and transported into tissues. A growing bulk of evidence suggests that LOPs, mainly reactive aldehydes and oxyster-ols, are potentially involved in the pathogenesis of many chronic diseases, such as atherosclerosis, Alzheimer’s disease, and inflammatory bowel disease. This review summarizes the current knowledge of the adverse effects, cytotoxicity, and the main mechanisms of LOPs involvement in humans and animals.
Keywords: Lipid oxidation products; Aldehyde; Cholesterol oxidation products; Adverse health effects; Toxicity.
1. Introduction
Lipids are the main nutrient components of food and important components of cells in biological systems. However, they are prone to oxidation through various pathways. Lipid oxidation is not only a major reason for the deterioration of food quality, but also has negative impacts on the integrity of the biological system (Shahidi and Zhong, 2010). The lipid oxidation process involves
a series of complex reactions, producing a large number of lipid oxidation products (LOPs), including lipid hydroperoxides, alde-hydes, ketones, alcohols, alcohols, and epoxy compounds, among others (Rodriguez-Amaya and Shahidi, 2021). These oxidative changes in lipids or lipid containing foods result in the forma-tion of peculiar smell, the loss of nutrients and bioactives, mak-ing foods unsuitable for consumption, and even formation of toxic compounds, which potentially risk to human health.
With the diversification of people’s diet, an increasingly large

Journal of Food Bioactives | www.isnff-jfb.com52
Potential adverse health effects Hu et al.
proportion of the human population frequently ingest oxidized/per-oxidized lipids. Oxidized/peroxidized lipids may contain a class of toxic compounds including hydroxyl fatty acids, cyclic mono-mers, dimers, and polymers and polycyclic aromatic hydrocarbons (Rodriguez-Amaya and Shahidi, 2021). Besides, dietary oxidized/peroxidized lipids exist in gastrointestinal tract constantly, and de-compose into a variety of oxygenated and aliphatic fatty acid scis-sion as well as secondary or tertiary lipid oxidation products in the gastric lumen (Vieira, Zhang, and Decker, 2017). Some of them are easily absorbed into the lymph or directly into the blood stream after ingestion (Kanner, 2007). In the past decades, the deleterious effects of regular ingestion of these LOPs on human health have attracted a large amount of high-impacting research interest and focus.
Although a myriad of compounds may be formed during lipid oxidation, the mainly focused research areas are the oxidation prod-ucts of polyunsaturated fatty acids (PUFAs) and cholesterol oxida-tion products (COPs) (Figure 1). For PUFAs, hydroperoxides from PUFA oxidation undergoes β-cleavage and produces various spe-cies of aldehydes. Among these aldehydic end-products, reactive α, β-unsaturated aldehydes, such as 4-hydroxy-2-nonenal (HNE), 4-hydroxy-2-hexenal (HHE), acrolein, malondialdehyde (MDA), and trans, trans-2,4-decadienal (tt-DDE), are the most important secondary oxidation products in terms of both quantity and toxici-
ty (Guillén and Goicoechea, 2008; Rodriguez-Amaya and Shahidi, 2021). Accumulated evidence shows that they are regarded as the second messengers of free radicals and possess potent electrophilic properties that could interfere with many pathophysiological pro-cesses, and exert toxic effects in both human and animal studies (Gęgotek and Skrzydlewska, 2019). For cholesterol, oxidation of cholesterol leads to the formation of a large number of oxidized products, generally known as oxysterols. They are important me-diators of cholesterol-induced effects with detrimental properties. They can enter intracellular membranes and affect the structure and function of membrane-associated proteins (Maldonado-Perei-ra et al., 2018; Testa et al., 2018). Solid evidence indicates that these aldehydes and COPs are implicated in or could aggravate a series of adverse effects on human health, such as cardiovascular diseases, neurodegenerative diseases, mutagenic and carcinogenic properties, gastrointestinal disease, and hepatotoxicity (Table 1). However, very few reviews of this topic have been published.
Taking into account the increasing tendency to enrich foods with LOPs and the increasing incidence of the related diseases, this review summarizes the current knowledge about the studies of LOPs and the related diseases. This evidence might help to en-hance people’s awareness of the health effects of dietary oxidized lipids and provide potential therapeutic directions for reducing the LOPs related diseases.
2. Cardiovascular diseases
2.1. Atherosclerosis
Oxidized lipid and LOPs in foods are absorbed by the intestine, ap-pear in the blood circulation, and can have deleterious cardiovascu-lar effects in humans (Cohn, 2002). Atherosclerosis is a progressive disease characterized by the accumulation of lipid and fibrous com-ponents in the large arteries (Zaguri et al., 2007). Numerous studies from both animals and humans indicate that dietary oxidized lipids, aldehydic LOPs, and COPs have some atherosclerotic effects. Ac-cumulated evidence shows that people who consume LOPs have significant higher oxidized lipids level in chylomicrons and very low density lipoproteins (VLDL), which is positively associated with the occurrence of clinical evidence of atherosclerosis in hu-mans (Ahotupa, 2017; Hajri, 2018; Viktorinova et al., 2016). An-other human study conducted by Menéndez-Carreño et al. (2011) analyzed the relationship between total COPs and different cardio-vascular diseases risk factors in the serum of 88 recruited subjects. The results showed that the serum COPs level of patients with ath-erosclerosis was significantly higher than those of subjects who did not show atherosclerosis. More recent investigation reported that serum 7ß-hydroxycholesterol concentration was the strongest pre-dictor of a rapid progression of carotid atherosclerosis in humans (Maldonado-Pereira, et al. 2018). All these evidences have empha-sized the close relationship between arteriosclerosis and LOPs.
In animal experiments, the COPs have been found to have profound biological effects following the intake of meals rich in oxidized cholesterol, such as increased aortic injury, impaired vascular endothelium, and which also support the role of LOPs in atherosclerosis. Staprans et al. (2000; 2005) found that dietary oxidized cholesterol for respective 7 and 4 months significantly in-creased aortic lesions in apolipoprotein E (ApoE) and low density lipoproteins (LDL) receptor-deficient mice. Meynier et al. (2002) reported that administration of oxysterols in high cholesterol diets for 3 months induced vascular wall thickening, endothelial dam-age and smooth muscle cell proliferation and migration in Golden
Figure 1. Examples of chemical structures of the lipid oxidation products (LOPs).

Journal of Food Bioactives | www.isnff-jfb.com 53
Hu et al. Potential adverse health effectsTa
ble
1. A
brie
f ove
rvie
w o
f ani
mal
stud
ies o
n th
e ro
le o
f die
tary
α, β
-uns
atur
ated
ald
ehyd
es o
r CO
Ps in
rela
ted
dise
ases
and
pat
holo
gica
l cha
nges
Alde
hyde
Expe
rimen
-ta
l mod
elDo
se/
Con-
cent
ratio
nTi
me
Effe
cts
Refe
renc
es
Card
iova
scul
ar d
isea
ses
Athe
rosc
lero
sis
Acro
lein
ApoE
-/- m
ice
2.5
mg/
kg/d
ay8
wee
ksIn
crea
sed
lesio
n fo
rmat
ion
in th
e ao
rtic
val
ve a
nd a
ortic
arc
h,
elev
ated
con
cent
ratio
ns o
f cho
lest
erol
and
pla
tele
t fac
tor
4 le
vels
in th
e pl
asm
a, m
acro
phag
e ac
cum
ulat
ion,
and
the
abun
danc
e of
smal
l and
med
ium
VLD
L pa
rtic
les.
Sriv
asta
va e
t al
., 20
11
Acro
lein
C57B
L/6
mic
e5
mg/
kg/d
ay24
h a
fter
expo
sure
Incr
ease
d pl
asm
a ch
oles
tero
l, tr
iacy
lgly
cero
ls, a
nd V
LDL,
and
de
crea
sed
hepa
tic li
pase
act
ivity
and
hep
atic
lipa
se e
xpre
ssio
n.Co
nklin
et a
l., 2
010
Acro
lein
ApoE
-nul
l mic
e0.
05–1
mg/
kg/d
ay8
wee
ksFe
w h
epat
ic g
enes
wer
e dy
sreg
ulat
ed th
at in
volv
ed
with
lipi
d sy
nthe
sis a
nd tr
affic
king
and
APR
.Co
nklin
et a
l., 2
011
Acro
lein
ApoE
-/- m
ice
3 m
g/kg
/day
1 m
onth
Incr
ease
d se
rum
and
aor
tic c
hole
ster
ol, t
rigly
cerid
es, a
nd li
pid
pero
xide
s;
Incr
ease
d in
trac
ellu
lar o
xida
tive
stre
ss a
nd o
ver-e
xpre
ssio
n of
key
re
gula
tors
of c
ellu
lar l
ipid
bio
synt
hesis
: SRE
BPs,
HM
GCR,
and
DGA
T1.
Rom
et a
l., 2
017
Oxi
dize
d ch
oles
tero
lAp
oE m
ice/
LDL
R-de
ficie
nt m
ice
Diet
con
tain
ing
5% o
xidi
zed
chol
este
rol
4 mon
ths/
7
mon
ths
Incr
ease
d ao
rtic
lesio
ns b
y 38
% a
nd 3
2%, r
espe
ctiv
ely.
Stap
rans
et
al.,
2000
Oxi
dize
d ch
oles
tero
lRa
bbits
Diet
con
tain
ing
0.33
%
chol
este
rol
of w
hich
5%
w
as o
xidi
zed
12 w
eeks
Incr
ease
d fa
tty
stre
ak le
sions
in th
e ao
rtas
by
100%
.St
apra
ns e
t al
., 19
99
Oxy
ster
ols
Gold
en S
yria
n ha
mst
ers
Diet
con
tain
ing
a m
ixtu
re o
f ox
yste
rols
3 m
onth
sIn
duce
d va
scul
ar w
all t
hick
enin
g, e
ndot
helia
l dam
age
and
smoo
th m
uscl
e ce
ll pr
olife
ratio
n an
d m
igra
tion.
Mey
nier
et
al.,
2002
Diab
etes
mel
litus
4-HN
EGa
stro
cnem
ius
mus
cles
isol
ated
fr
om C
D-1
Swiss
mic
e
100
μM\
Decr
ease
d in
sulin
sign
alin
g an
d in
sulin
-indu
ced
gluc
ose
upta
ke in
mus
cle.
Impa
irs
insu
lin a
ctio
n by
indu
cing
oxi
dativ
e st
ress
and
oxi
dativ
e da
mag
e to
pro
tein
s.Pi
llon
et a
l., 2
012
4-HH
EW
istar
rats
10 m
g/kg
\In
duce
d in
sulin
resis
tanc
e, a
ltere
d w
hole
-bod
y in
sulin
sens
itivi
ty
and
decr
ease
d gl
ucos
e in
fusio
n ra
te; a
bolis
hed
insu
lin
indu
ced
phos
phor
ylat
ion
of A
kt in
skel
etal
mus
cle.
Soul
age
et a
l., 2
018
Hype
rten
sion
tt-DD
ESD
rats
500
mg/
kg28
day
sIn
duce
d bl
ood
pres
sure
ele
vatio
n an
d en
doth
elia
l dys
func
tion
by o
xida
tive/
nitr
ativ
e st
ress
and
apo
ptos
is.Hu
et a
l., 2
020
Card
ioto
xici
ty
Acro
lein
ICR
mic
e5
mg/
kg\
Exac
erba
ted
myo
card
ial i
sche
mic
inju
ry a
nd b
lock
ed N
O-
indu
ced
card
iopr
otec
tion
via
disr
uptin
g PK
C sig
nalin
g.W
ang
et a
l., 2
008

Journal of Food Bioactives | www.isnff-jfb.com54
Potential adverse health effects Hu et al.
Alde
hyde
Expe
rimen
-ta
l mod
elDo
se/
Con-
cent
ratio
nTi
me
Effe
cts
Refe
renc
es
Card
iova
scul
ar d
isea
ses
Acro
lein
Spra
gue
Daw
ley
rats
5 m
g/kg
30 d
ays
Indu
ced
card
iac
inju
ry, a
nd d
ysfu
nctio
n in
mito
chon
dria
l bio
ener
getic
s of
hea
rt b
y de
crea
sing
leve
ls of
oxi
dativ
e ph
osph
oryl
atio
n en
zym
es, t
ricar
boxy
lic a
cid
cycl
e en
zym
es a
nd A
TP.
Aydı
n et
al.,
201
8
Neu
rode
gene
rativ
e di
seas
es
4-HN
ESp
ragu
e–Da
wle
y ra
ts3
mL/
kg/d
ay21
day
sIn
duce
d di
srup
tion
of su
rviv
al si
gnal
ing
of fr
onta
l cor
tex
chol
iner
gic
neur
ons.
Zára
te e
t al.,
200
9
Acro
lein
Wist
ar ra
ts2
mg/
kg/d
ay90
day
sIn
duce
d hi
stop
atho
logi
cal c
hang
es o
f bra
in ti
ssue
s an
d ne
urof
ilam
ent a
ccum
ulat
ion.
Selm
anoğ
lu
et a
l., 2
018
Acro
lein
Spra
gue-
Daw
ley
rats
2.5
mg/
kg/d
ay12
wee
ksIn
duce
d co
gniti
ve im
pairm
ent a
nd p
ykno
sis/a
trop
hy o
f hip
poca
mpa
l neu
rons
by
indu
cing
oxi
dativ
e da
mag
e an
d ch
ange
s of A
PP, β
-sec
reta
se, α
-sec
reta
se
and
rece
ptor
, and
act
ivat
ing
astr
ocyt
es, r
egul
atin
g BD
NF/
TrkB
pat
hway
.
Huan
g et
al.,
201
8
Gast
roin
test
inal
dise
ases
4-HN
EW
istar
rats
0.26
μM
\In
duce
d pe
ptic
ulc
ers a
nd re
duce
d th
e GS
H co
nten
t in
the
endo
plas
mic
retic
ulum
.Ja
yara
j et a
l., 1
986
Acro
lein
Wist
ar ra
ts6
mg/
kg70
day
sIn
duce
d er
osio
n, h
yper
plas
ia a
nd h
yper
kera
tosis
in th
e gl
andu
lar
muc
osa
of th
e st
omac
h or
in th
e fo
rest
omac
h.Go
mes
et a
l., 2
002
tt-DD
EF3
44/N
rats
; B6
C3F1
mic
e80
0 m
g/kg
3 m
onth
sIn
duce
d fo
rest
omac
h ul
cera
tion
and
diar
rhea
.Ch
an, 2
011
tt-DD
ESp
ragu
e Da
wle
y ra
ts50
or 1
00 m
g/kg
\In
hibi
t gas
tric
em
ptyi
ng ra
te in
rats
thro
ugh
indu
cing
cho
lecy
stok
inin
secr
etio
n.Hi
ra e
t al.,
201
5
Acro
lein
ApoE
-/- m
ice
3 m
g/kg
/day
1 m
onth
Indu
ced
a m
ajor
shift
in th
e gu
t mic
robi
ota
com
posit
ion
in in
clud
ing
a sig
nific
ant
phyl
um-le
vel c
hang
e w
ith in
crea
sed
Firm
icut
es a
nd d
ecre
ased
Bac
tero
idet
es.
Rom
et a
l., 2
017
Carc
inog
enic
ity
tt-DD
ECD
-1 (I
CR) m
ice
8 or
24
mg/
kg8
wee
ksIn
crea
sed
the
num
ber a
nd ty
pes o
f cel
ls in
the
bron
choa
lveo
lar
lava
ge fl
uid;
Indu
ced
path
olog
ical
cha
nges
, and
infla
mm
ator
y ge
ne m
odul
atio
ns in
the
lung
tiss
ues.
Wan
g et
al.,
201
0
tt-DD
EIC
R m
ice
24 m
g/kg
8 w
eeks
Indu
ced
lung
lesio
ns a
nd c
hang
es o
f am
ino
acid
pro
files
in th
e ur
ine
and
seru
m.
Lin
et a
l., 2
014
MDA
Swiss
mic
e0.
1–10
μg/
g/da
y12
m
onth
sIn
crea
sed
the
tota
l neo
plas
ms a
nd n
eopl
astic
lesio
ns in
the
liver
.Dr
aper
, McG
irr,
and
Hadl
ey, 1
986
Mut
agen
icity
\\
\\
\\
Hepa
toto
xici
ty
4-HN
ESp
ragu
e-Da
wle
y ra
ts0.
5, 2
.5, o
r 12
.5 m
g/kg
28 d
ays
Indu
ced
signi
fican
t alte
ratio
ns in
hep
atot
oxic
ity b
iom
arke
rs, s
uch
as se
rum
alb
umin
, ser
um to
tal b
iliru
bin,
and
AST
act
ivity
.Ka
ng e
t al.,
201
1
Acro
lein
Wist
ar a
lbin
o ra
ts4
mg/
kg7
days
Indu
ced
liver
tiss
ues a
popt
osis,
dila
tatio
ns in
the
perin
ucle
ar sp
ace,
and
cyt
opla
smic
vac
uoliz
atio
n.Gö
kçe
et a
l., 2
020
Tabl
e 1.
A b
rief o
verv
iew
of a
nim
al st
udie
s on
the
role
of d
ieta
ry α
, β-u
nsat
urat
ed a
ldeh
ydes
or C
OPs
in re
late
d di
seas
es a
nd p
atho
logi
cal c
hang
es -
(con
tinue
d)

Journal of Food Bioactives | www.isnff-jfb.com 55
Hu et al. Potential adverse health effects
Syrian hamsters. Besides, Gargiulo et al. (2017) reviewed that ox-ysterols contribute to atherosclerosis by inducing apoptosis, oxida-tive stress, chronic inflammation, foam cell formation, mitochon-drial damage, loss of nitric oxide (NO) bioavailability, as well as enhancing macrophage activity in vitro (Figure 2).
In addition to oxysterols, several previous publications reviewed that aldehydes are involved in the genesis and development of ath-erosclerosis by inducing various key steps, including damage en-dothelial function, modification of LDL, promotion foam cell for-mation, stimulation the adhesion of monocytes and lymphocytes, proinflammatory, redox imbalance, deoxyribonucleic acid (DNA) damage, formation of the monocyte chemoattractant protein-1, and stimulation smooth muscle cells proliferation and migration (Leonarduzzi et al., 2010; Zirak et al., 2019). In this review, the collected evidence suggests that aldehydes, especially acrolein, contribute to the development of atherosclerosis by inducing vas-cular injury, systemic dyslipidemia, and gut microbiota in vivo. According to the results reported by Srivastava et al. (2011), di-etary exposure to acrolein (2.5 mg/kg/day) by gavage for 8 weeks aggravated atherosclerosis in apolipoprotein E-deficient (ApoE-/-) mice, as evidenced by increased lesion formation in the aortic valve and aortic arch, elevated concentrations of cholesterol and platelet factor 4 levels in the plasma, macrophage accumulation, and the abundance of small and medium VLDL particles. Conk-lin and co-workers reported that oral acrolein feeding resulted in increased plasma cholesterol, triacylglycerols, and VLDL, and decreased hepatic lipase activity and hepatic lipase expression in mice (Conklin et al., 2010; Conklin et al., 2011). Rom et al. (2017) found that feeding ApoE-/- mice with acrolein (3 mg/kg/day) for 1 month resulted in a significant dyslipidemia as well as elevations of the prevalence of Ruminococcaceae and Lachnospiraceae, and this altered the gut microbiota composition is positively correlated with enhanced atherosclerosis.
Taken together, these findings indicate that LOPs in the diet are directly related to the level of oxidized lipids in the circulation, which can trigger the occurrence of atherosclerosis involved in a variety of complex ways.
2.2. Diabetes mellitus
Based on the preceding evidence, the process of lipid oxidation and
the resulting LOPs are implicated in the development of diabetes and its complications. Indeed, a study based on the results of hu-man trials in diabetic patients showed that the level of oxidized li-pids in serum chylomicrons increased after people with poor blood glycemic control ingested dietary oxidized fatty acids (Staprans et al., 1999). Besides, oral consumption of a diet contains high oxidized frying oil are demonstrated to induce impairment of glu-cose tolerance and impairment of insulin secretion in rodents (Li et al., 2017; Liao, Shaw, and Chao, 2008). Notably, HNE is con-sidered as a crucial molecule for triggering and sustaining a wide range of biochemical events underlying the development of this processes (Jaganjac et al., 2013). It has been reported that HNE could directly bind to insulin and consequently impair its ability to stimulate glucose uptake in mice (Pillon et al., 2011; Pillon et al., 2012). Following these findings, another by-product of n-3 PUFAs peroxidation, HHE, which has similarities in structure with HNE, was reported to play a causal role in the pathophysiology of type 2 diabetes. The researchers found that circulating levels of HHE in type 2 diabetic humans and diabetic rat models were twice that of non-diabetic counterparts, and it was positively correlated with blood glucose levels. Acute intravenous injection of HHE (10 mg/kg) significantly changed the whole-body insulin sensitivity and reduced the glucose infusion rate in rats (Soulage et al., 2018). Moreover, both HNE and HHE were found to impair insulin ac-tion, insulin signaling, and insulin-induced glucose uptake in mus-cle cells and adipose cells (Cohen et al., 2013; Pillon et al., 2012; Soulage et al., 2018).
For COPs, some evidences show high concentrations of plasma COPs in patients or animals with diabetes mellitus compared with healthy controls (Endo et al., 2008; Ferderbar et al., 2007; Yosh-ioka et al., 2005). However, the question whether dietary COPs can directly lead to the development of diabetes mellitus needs more research to prove.
2.3. Hypertension
As we all know, hypertension is one of the most common chron-ic diseases, and also an important risk factor for cardiovascular disease. Vascular endothelial dysfunction contributes to the oc-currence and development of vascular injury, and it is also an effective independent predictor of hypertension (Endemann and
Figure 2. The effects of COPs on different cell lines that involved in various diseases.

Journal of Food Bioactives | www.isnff-jfb.com56
Potential adverse health effects Hu et al.
Schiffrin, 2004). Evidences from humans and animals indicate that intake of LOPs may be a potential risk factor for the devel-opment of hypertension. A human study conducted by Soriguer et al. (2003) reported that LOPs produced from cooking oil were strongly and positively associated with high blood pressure. In ani-mal experiments, administration of oxidized lipids was reported to induce hypertension by impairing endothelium-dependent and endothelium-independent vasorelaxation, reducing NO bioavail-ability, increasing thromboxane and angiotensin converting en-zyme (ACE), altering the histological structures of blood vessels, and reducing adenosine triphosphate-sensitive potassium (KATP) and large-conductance calcium-dependent potassium channels (BK channels). For example, intake of oxidized oils for 6 months was reported to induce blood pressure elevation, attenuate relaxa-tions in response to acetylcholine (ACh) and sodium nitroprusside (SNP) as well as increase vasoconstriction to phenylephrine (PE) (Leong et al., 2009; Nurul-Iman et al., 2013). Other studies point that ingestion of thermally oxidized oil for 16 weeks induced blood pressure elevation, disrupted elastic lamellae of tunica media and the structures of heart and blood vessels, and simultaneously with increased thromboxane and ACE in rats (Nkanu, Owu, and Osim, 2018; Siti et al., 2017; Siti et al., 2018).
Accumulated evidence reveals that oxidative stress is one of the important reasons leading to the generation and/or maintenance of hypertension, which can directly change the function of vas-cular endothelial or cause changes in vascular tension (Hu et al., 2020). As an inducer of reactive oxygen species (ROS), reactive aldehydes were regarded as an important mediator of oxidized li-pids in inducing blood pressure elevation. Indeed, Conklin et al. (2006) concluded that acrolein generation in the blood vessel wall increased human susceptibility to vasospasm, an event that is en-hanced in hypertension. Recently, it was demonstrated that oral ad-ministration of tt-DDE (500 mg/kg) for 28 day induced blood pres-sure elevation in rats, and this effect was closely associated with tt-DDE induced endothelial dysfunction, oxidative/nitrative stress and apoptosis in both vascular endothelial tissue and endothelial cells (Hu et al., 2020). Besides, other evidences indicate that ex-cess reactive aldehydes contribute to progressive and deleterious changes in hypertensive by increasing cytosolic-free calcium lev-els, altering renal vascular function, and inducing endothelial cell toxicity (Hu et al., 2020; Lee and Park, 2013).
2.4. Cardiotoxicity
In addition to their role in the process of atherosclerosis, diabetes, and hypertension some evidences indicate that LOPs are detrimen-tal to heart and cardiac function, and instigate multiple cellular dis-turbances in cardiomyocytes (Hajri, 2018). Indeed, consumption of oxidized rapeseed oil for 112 days induced transient myocardial lipidosis characterized by accumulation of fat droplets in myo-cardial fibers of rats (Charlton et al., 1975). Rouaki et al. (2013) reported that dietary oxidized sunflower oil induced extensive ne-crosis of heart tissue, increased body weight and LPO levels, and decreased catalase (CAT) and glutathione peroxidases (GPx) ac-tivities in rats. Again, other studies found that dietary heated palm oil diet for 16 weeks significantly increased the myofiber width and area, and induced necrosis in cardiac tissue of rats (Kamisah et al., 2016; Leong et al., 2008).
Specially, reactive aldehydes including acrolein and HNE are consistently demonstrated to be the important mediators that con-tribute to cardiotoxicity. The immunohistochemical examination of myocardial biopsy in patients with cardiomyopathy and/or heart failure showed that HNE protein adduct increased in the heart and
was positively correlated with decreased heart function (Nakamura et al., 2005). Another study reported that HNE can form adducts with cardiac aldehyde dehydrogenase 2 (ALDH2) protein in meta-bolic syndrome and/or type 2 diabetic mice, accompanied by obvi-ous cardiac hypertrophy, cardiac damage, myocardial mitochon-drial dysfunction, and heart weight increase (Mali et al., 2016). For acrolein, Wang et al. (2008) found that acrolein (5 mg/kg) feeding by gavage could exacerbate myocardial ischemic injury and blocks NO-induced cardioprotection via mechanisms that disrupt protein kinase C (PKC) signaling in mice. Aydın et al. (2018) reported that oral administration of acrolein (5 mg/kg) in rats for 30 days induced cardiac injury, with oxidative stress and dysfunction in mitochondrial bioenergetics of heart. On the other hand, reactive aldehydes can induce a wide array of toxic consequences in the cardiomyocytes, including destruction of membrane and cytosolic enzyme activities and actin cytoskeleton, inducing mitochondrial damage, depletion of adenosine-triphosphate (ATP) content, mem-brane swelling and dissolution, alteration of current through K+ channels, imbalance of cellular calcium homeostasis, and inducing oxidative stress, inflammation, and apoptosis (Chapple, Cheng, and Mann, 2013; Gianazza et al., 2019).
For oxysterol, there are significant positive correlations be-tween oxysterol levels and the number of risk factors for coronary heart disease (Samadi et al., 2019). Besides, in a study of Sozen et al. (2018), the data indicated that a high-cholesterol diet caused abnormal heart function by regulating proteins related to lipid metabolism in rabbits. In vitro, oxysterols also exert strong toxic effects on the cardiomyocytes, including increasing inflammatory cytokine, downregulation of B-cell lymphoma 2 (BCL-2) expres-sion, activation of caspases, production of ROS, opening of mito-chondrial permeability transition pore, inducing cell hypertrophy, Ca2+ cycling dysfunctional, DNA fragmentation, and cell death (Cilla et al., 2017; Lee, Nam, and Lee, 2015) (Figure 2). However, more human and animal experiments need to be provided to prove the potential role of COPs intake and cardiotoxicity.
3. Neurodegenerative diseases
Documented evidences have demonstrated that free radical-me-diated oxidative damage is one of the important mechanisms of the pathogenesis of neurodegenerative diseases, including Alz-heimer’s disease (AD), Parkinson (PD), Hungtington’s disease (HD), and Amyotrophic lateral sclerosis (ALS). Since HNE acts both as a signaling molecule and as cytotoxic product, its effect on neurodegenerative diseases has been extensively investigated. Perluigi et al. (2012) pointed that there were elevated levels of HNE and its proteins adducts in relevant brain tissues and body fluids of AD, PD, HD subjects and/or animal models with neuro-degenerative diseases. The formation of protein adducts results in altered energy metabolism and protein homeostasis, mitochondrial dysfunction, as well as reduced antioxidant levels, which provides further support for the physical impairment, cognitive decline, progressive memory loss, and oxidative stress associated with the neurodegenerative disorders (Barrera et al., 2015). For example, Sardar Sinhah et al. (2018) reported that HNE was implicated in the pathogenesis of different PD by modifying α-synuclein, an important protein that linked to synaptic plasticity, neurotransmit-ter release and the maintenance of the synaptic pool. On the other hand, intake of HNE can induce direct adverse effects on neurons and neural cells. Zárate et al. (2009) reported that administration of oxidized sunflower oil containing HNE (3 mL/kg/day) for 21 days induced disruption of survival signaling of frontal cortex choliner-

Journal of Food Bioactives | www.isnff-jfb.com 57
Hu et al. Potential adverse health effects
gic neurons, as evidenced by the reduction of the number of neural cells with positive immunostained tropomyosin receptor kinases A (TrkA) receptor in the frontal cortex of rats.
In addition to HNE, Selmanoğlu et al. (2018) reported that oral exposure to acrolein (2 mg/kg/day) for 90 days induced histo-pathological changes of brain tissues and neurofilament accumula-tion in rats. Huang et al. (2018) found that acrolein (2.5 mg/kg/day) treatment by gavage for 12 weeks induced cognitive impair-ment and pyknosis/atrophy of hippocampal neurons by inducing oxidative damage, activating astrocytes, regulating brain-derived neurotrophic factor (BDNF)/ tyrosine Kinase receptor B (TrkB) pathway, β-secretase 1 (BACE-1) and A-disintegrin and metal-loprotease (ADAM) in hippocampus and cortex, as well as key enzymes involved in amyloid precursor protein (APP) metabolism in rats.
Oxysterols are believed to be the link between altered brain cholesterol metabolism and neurodegenerative diseases. Differ-ent oxysterols are reported to be involved in numerous age-related pathological conditions. Among them, the balance of 24-hydroxy-cholesterol (24-OH) and 27-hydroxycholesterol (27-OH) has been identified as a useful tool for monitoring and/or diagnosing of the progression of neurodegenerative diseases (Poli, Biasi, and Leon-arduzzi, 2013). By analyzing the patient samples, it was found that the increase in plasma 24-OH level and 24-OH/27-OH ratio is related with the development of eventual cognitive impairment (Hughes et al., 2012; Malaguti et al., 2019). Besides, 7-KC ac-cumulation has been correlated to the development of AD. It ex-erts toxic effects on cellular model systems derived from neuronal tissues by multiple mechanisms (Malaguti et al., 2019). Although the direct effect of oral administration of oxysterols on neurode-generative disease has rarely been reported, increasing evidence shows that oxysterols can impair neuronal and muscular develop-ment and induce oxidative stress, inflammation, amyloid β-protein (Aβ) formation and accumulation, and death of neurons in various human neuronal cell lines (Testa et al., 2018) (Figure 2).
4. Gastrointestinal diseases
A growing body of evidence suggests that long-term consumption of oxidized oils may be an important factor in human gastrointesti-nal diseases, and the cause is closely related to the large amount of generated LOPs (Martin Grootveld et al., 2020). Animal toxicol-ogy studies have shown that oxidized oils produced under extreme processing conditions induces severe irritation of gastrointestinal tract and could react with intestinal mucosal proteins and interfere with the absorption process in experimental animals (Esterbauer, 1993; Hajri, 2018). Obembe et al. (2011) found that ingestion of thermally oxidized palm oil may induce distortion in the morphol-ogy of intestinal villi, malabsorption of intestinal juice and glucose absorption in the intestine of rats. Zhou et al. (2016) reported that oral administration of fried oil induced impairment of intestinal mucosa, endocrine tissue destruction and inflammation, and mi-crobiota structure imbalance in rats.
The formation of reactive α, β-unsaturated aldehydes during high fat dietary intake are one of the major contributory factors in aggravation of gastric mucosa and intestinal diseases. They can directly contact intestine lamina, and induce the loss of intestinal epithelial barrier function and abnormal inflammatory response of the intestinal mucosa, and play crucial roles in the pathogenesis of inflammatory bowel disease (IBD) (Rossin et al., 2017; Sottero et al., 2018). Jayaraj et al. (1986) reported that oral administration of HNE to rats at a dose of only 0.26 μM, a concentration similar to
that of healthy human blood plasma induced peptic ulcers in the animals. Gomes et al. (2002) found that acrolein (6 mg/kg) admin-istration by gavage for 70 days in rats induced erosion, hyperplasia and hyperkeratosis in the glandular mucosa of the stomach or in the forestomach. Oral administration of tt-DDE (800 mg/kg) by gavage for 3 months can induce forestomach ulceration and di-arrhea, and inhibit gastric emptying rate in rats through inducing cholecystokinin (CCK) secretion in rats (Chan, 2011; Hira et al., 2015). More recently, Rom et al. (2017) reported that intake of acrolein (3 mg/kg/day) for 1 month induced a major shift in the gut microbiota composition in atherosclerotic ApoE-/- mice, including a significant phylum-level change with increased Firmicutes and decreased Bacteroidetes.
For oxysterols, they are closely related to the occurrence and severity of intestinal inflammation and intestinal endothelial dam-age. Dietary intake of 7-ketocholesterol (7-KC) impaired the integ-rity of intestinal epithelial monolayer barrier by down-regulating tight junction protein zonule occlusion-1 (ZO-1) and reducing the transepithelial electrical resistance (Chalubinski et al., 2014; Cha-lubinski et al., 2013). According to the report presented by Will-inger (2019), high amounts of oxysterols synthesized from diet-derived cholesterols would activate the receptor G protein-coupled receptor 183 (GPR183) and trigger pro-inflammatory immune activity in the intestine. Besides, dietary oxysterols can induce ir-reversible damage of the intestinal epithelial, and can exert a series of toxic effects on human epithelial colonic cells, including mi-tochondrial injury, pro-apoptotic, pro-oxidation, pro-inflammatory activities, thereby impairing the characteristics of the intestinal epithelium (Sottero et al., 2018) (Figure 2).
5. Carcinogenicity
Evidence for a putative role of dietary oxidized oils and LOPs in carcinogenicity has come from both animal and human studies. Stott-Miller et al. (2013) examined the relationship between the intake of deep-fried foods (including French fries, fried chicken, fried fish, etc.) and prostate cancer risk through a food frequen-cy questionnaire survey in 1,549 cases and 1,492 controls, and it was concluded that regular consumption of deep-fried foods is associated with increased prostate risk. Recently, Mhatre et al. (2020) conducted a human case-control study, showing that high consumption of fried mustard oil increased the risk of gallblad-der cancer. On the other hand, the basis of the epidemiological studies suggested that cooking fumes from heated culinary oils are toxic and exposure to human showed to be an increased risk factor of contracting cancer (Chen et al., 2020; Jia et al., 2018). In par-ticular, a positive correlation between the inhaling oil fume from heated oils and high risks of lung cancer (Ma et al., 2020).
Cumulative evidence indicated that lipid oxidation-derived re-active aldehydes may be the main carcinogenic substances in oxi-dized oils that exert carcinogenicity. It has been reported that alde-hydes induce and participate in the occurrence and progression of cancer, and chronic exposure can increase the tumor frequency in animals (Eckl and Bresgen, 2017). For example, tt-DDE, a major aldehyde in cooking oils fumes, was reported to induce lung car-cinogenesis. It has been reported that intratracheally instilled with tt-DDE to mice for 8 weeks significantly increased the number and types of cells in the bronchoalveolar lavage fluid, increased the epithelial hyperplasia and granulomatous nodules at the bron-choalveolar junction (Lin et al., 2014; Wang et al., 2010). Oral ad-ministering MDA as its enol salt over a range of 0.1–10.0 μg/g/day for 12 months in Swiss mice showed a dose dependent increase of

Journal of Food Bioactives | www.isnff-jfb.com58
Potential adverse health effects Hu et al.
total neoplasms and neoplastic lesions in the liver (Draper, Mc-Girr, and Hadley, 1986). Likewise, oral consumption of acrolein induced pulmonary injuries, including airspace enlargement, lung inflammation, lung edema, and even lung cancer (Bein and Lei-kauf, 2011).
The reported biochemical effects of aldehydes are associated with their lipophilicity and high reactivity (Figure 3). They can covalently interact with amino acids cysteine, histidine and lysine, DNA, phospholipids, protein, and sulfhydryl groups (Gęgotek and Skrzydlewska, 2019). Importantly, these aldehyde-induced DNA modification as well as protein adducts play key features in the growth and progression of cancer. Oral administration of HNE by gavage has been found to induce significant increases of 2′-de-oxyguanosine (dGuo) adducts of HNE (HNE-dGp-adducts) and HNE protein adducts in different types of tumors, including liver, lung, renal, colon and forestomach of rats (Barrera, et al., 2015; Zarkovic, Jakovcevic, and Zarkovic, 2017). Hong et al. (2020) an-alyzed a total of 62 urothelial carcinomas patients, and found that acrolein-DNA (Acr-dG) levels were statistically correlated with chronic kidney disease stages in urothelial carcinomas patients. On the other hand, the formation of adducts affects protein function, triggering different mechanism, such as proliferation, cell differ-entiation and apoptosis, enhances resistance to cancer suppressor drugs, and alters cancer progression, among others (Cherkas and Zarkovic, 2018; Martín-Sierra et al., 2019).
The influence of dietary oxysterols on the incidence of can-cer is still a controversial debate. As summarized by Kloudova-Spalenkova et al. (2020), oxysterols not only play important roles in carcinogenesis, proliferation, migration, apoptosis and multiple signaling pathways, but also play many roles in cells in physiologi-cal and pathological conditions thus modulating cancer therapy. Besides, the involvements of oxysterols in hepatocellular carci-noma, pancreatic, colon cancer, and breast cancer have also been reviewed in recent years (Kovač et al.,2019; Nazih et al., 2020). They concluded that the role of oxysterols in carcinogenesis is far from being conclusive.
6. Mutagenicity
To date, some investigations have proven the mutagenic potential of orally administered oxidized oils and LOPs. Hageman et al. (1991) conducted a study to evaluate the mutagenicity of repeated heated oils containing oxidation products of linoleic acid in rats. Results showed that oral consumption the repeatedly heated oils at a level of 10% (by weight) for 4 weeks increased urine mutagenic-ity, and enhanced cell proliferation in the oesophagus of rats. Other studies performed on frying oil obtained by the conventional frying method showed that fatty acid oxidation products and their thermal degradation products produced by deep frying have mutagenic ac-tivity (Chen et al., 2003; Perez, Lopez de Cerain, and Bello, 2002). Generally, severe abusive thermal processing conditions, such as long heating times, high temperature, or overused heated oil, are necessary to produce appreciable mutagenic activity.
For aldehydic LOPs, Wu et al. (2001) reported that four enal compounds identified from the fumes of peanut oil at the dose of 10 μg per plate were mutagenic to Salmonella Typhimurium TA98 and TA100, and the order of mutagenic activity was tt-DDE > trans-trans-2,4-nonadienal (tt-2,4-NDE) > trans-2-decenal (t-2-DCA) > trans-2-undecenal (t-2-UDA). Likewise, Dung et al. (2006) reported that the major alkenal mutagenic compounds (tt-DDE, tt-2,4-NDE, t-2-DCA and t-2-UDA) from heated three cooking oils (soybean oil, sunflower oil, and lard) induced signifi-cant oxidative DNA damage by the 8-hydroxy-20-deoxyguanosine formation in A-549 cells at the concentrations from 50 to 200 μg/mL, which suggested that these aldehydes are generally both mu-tagenic and carcinogenic. Grúz et al. (2017) reported that both HNE and 2-hexenal induced base substitutions in the TA104 and TA100 Ames strains in a dose-dependent manner. Besides, in the review summarized by Eckl and Bresgen (2017), the mutagenici-ties of 2-hexenal, 2-heptenal, 2-octenal and 2-nonenal have been previously evaluated in bacterial systems, and each of these was found to exert significant effects at µmol/L concentrations. The
Figure 3. Summary of the formation of aldehyde adducts and their possible effects on the progression of cancer.

Journal of Food Bioactives | www.isnff-jfb.com 59
Hu et al. Potential adverse health effects
mutagenicity of MDA and acrolein tested in mammalian cell lines in the thioguanine resistance test and in the Ames test with differ-ent strains.
7. Hepatotoxicity
Substantial evidence shows that oxidized oils have hepatotoxic property, and animal studies have shown that they can induce varying degrees of liver damage regardless of short or long ad-ministration periods. These oxidized oils can disturb hepatic redox equilibrium and induce significant changes in the gene expres-sions related to fatty acid composition in animals (Ambreen, Sid-diq, and Hussain, 2020; Ringseis and Eder, 2011). For example, Ringseis and Eder (2011) summarized the phenotypic changes and molecular mechanisms of lipid metabolism observed in oxidized oil feeding studies. They pointed out that oxidized oil played an important role in fatty acid synthesis, cholesterol homeostasis and lipoprotein metabolism. The gene expressions or activities of fatty acid synthase (FAS), peroxisome proliferator-activated receptor α (PPARα), sterol regulatory element binding protein (SREBP), glucose-6-phosphate dehydrogenase (G6PDH), acyl coenzyme A (CoA) oxidase or cytochrome P450 (CYP)4A1 in the liver of rats were regulated. However, whether LOPs are the specific toxic compounds that exert hepatotoxicity remains unclear.
Aldehydes can be absorbed into the circulatory system, subse-quent accumulate and harm various body tissues, particularly the liver (Grootveld et al., 1998). According to the results presented by Kanazawa et al. (1998), radio-labelled hexanal and HNE accu-mulated in the liver of rats with a slight hepatic hypertrophy after administration with radio-labelled hydroperoxides of linoleic acid. Kang and co-authors found that oral daily exposure to HNE (0.5 mg/kg) for 28 days produced hepatotoxicity in rats, as evidenced by significant alterations in serum albumin, serum total bilirubin, and aspartate aminotransferase (AST) activity (Kang et al., 2011). In addition to HNE, Gökçe et al. (2020) reported that consumption of acrolein (4 mg/kg) for 7 days by gavage induced liver tissues apoptosis in rats, as evidenced by chromatin condensation in the nucleus of hepatocytes, dilatations in the perinuclear space, and cytoplasmic vacuolization.
8. Conclusions
Consideration of the popularity of processed and fried foods and the wide-spread consumption of fast foods, the investigation of the effects of dietary LOPs on health is becoming increasingly impor-tant. The collected evidence suggests that the formation of LOPs not only reduces lipid nutritional properties but also represents a risk to human health. Nevertheless, due to the extremely complex processes in the human body, more systematic research about the effects of LOPs on human health are needed, such as the protec-tion of antioxidant enzymes, the interaction of other components in diet, the role that intestinal flora plays in this process, and the effective method of inhibiting lipids oxidation.
Acknowledgments
This work was financially supported by “National Key R&D Program of China (2018YFD0901000)”, “The National Natural Science Foundation of China (31871759)”, “Dalian Science and Technology Innovation Fund Project (2019J11CY005)” and “Cen-
tral Funds Guiding the Local Science and Technology Develop-ment (No. 2020JH6/10500002)”.
References
Ahotupa, M. (2017). Oxidized lipoprotein lipids and atherosclerosis. Free Radic. Res. 51(4): 439–447.
Ambreen, G., Siddiq, A., and Hussain, K. (2020). Association of long-term consumption of repeatedly heated mix vegetable oils in different doses and hepatic toxicity through fat accumulation. Lipids Health Dis. 19(1): 69.
Aydın, B., Atlı Şekeroğlu, Z., and Şekeroğlu, V. (2018). Effects of whey pro-tein and conjugated linoleic acid on acrolein-induced cardiac oxida-tive stress, mitochondrial dysfunction and dyslipidemia in rats. Bi-omed. Pharmacother. 107: 901–907.
Barrera, G., Pizzimenti, S., Ciamporcero, E.S., Daga, M., Ullio, C., Arcaro, A., Cetrangolo, G.P., Ferretti, C., Dianzani, C., Lepore, A., and Gentile, F. (2015). Role of 4-hydroxynonenal-protein adducts in human diseases. Antioxid. Redox Signal. 22(18): 1681–1702.
Bein, K., and Leikauf, G.D. (2011). Acrolein - a pulmonary hazard. Mol. Nutr. Food Res. 55(9): 1342–1360.
Chalubinski, M., Wojdan, K., Gorzelak, P., Borowiec, M., and Broncel, M. (2014). The effect of oxidized cholesterol on barrier functions and IL-10 mRNA expression in human intestinal epithelium co-cultured with dendritic cells in the transwell system. Food Chem. Toxicol. 69: 289–293.
Chalubinski, M., Zemanek, K., Skowron, W., Wojdan, K., Gorzelak, P., and Broncel, M. (2013). The effect of 7-ketocholesterol and 25-hydroxy-cholesterol on the integrity of the human aortic endothelial and intes-tinal epithelial barriers. Inflamm. Res. 62(12): 1015–1023.
Chan, P.C. (2011). NTP toxicity studies of toxicity studies of 2,4-decadienal (CAS No. 25152-84-5) administered by gavage to F344/N Rats and B6C3F1 mice. Toxic. Rep. Ser. (76): 1–94.
Chapple, S.J., Cheng, X., and Mann, G.E. (2013). Effects of 4-hydroxynon-enal on vascular endothelial and smooth muscle cell redox signaling and function in health and disease. Redox Biol. 1: 319–331.
Charlton, K.M., Corner, A.H., Davey, K., Kramer, J.K., Mahadevan, S., and Sauer, F.D. (1975). Cardiac lesions in rats fed rapeseed oils. Can. J. Comp. Med. 39(3): 261–269.
Chen, T., Fang, Y., Chen, H., Chang, C., Huang, H., Chen, Y., Liao, K., Wu, H., Chang, G., Tsai, Y., Wang, C., Chen, Y., Huang, M., Su, W., Yang, P., Chen, C., Hsiao, C., and Hsiung, C. (2020). Impact of cooking oil fume exposure and fume extractor use on lung cancer risk in non-smoking Han Chinese women. Sci. Rep. 10(1): 6774.
Chen, Y.Y., Liu, J.F., Chen, C.M., Chao, P.Y., and Chang, T.J. (2003). A study of the antioxidative and antimutagenic effects of Houttuynia cordata Thunb. using an oxidized frying oil-fed model. J. Nutr. Sci. Vitaminol. 49(5): 327–333.
Cherkas, A., and Zarkovic, N. (2018). 4-Hydroxynonenal in Redox Homeo-stasis of Gastrointestinal Mucosa: Implications for the Stomach in Health and Diseases. Antioxidants (Basel) 7(9): 118.
Cilla, A., Alegría, A., Attanzio, A., Garcia-Llatas, G., Tesoriere, L., and Livrea, M.A. (2017). Dietary phytochemicals in the protection against oxys-terol-induced damage. Chem. Phys. Lipids 207: 192–205.
Cohen, G., Riahi, Y., Sunda, V., Deplano, S., Chatgilialoglu, C., Ferreri, C., Kaiser, N., and Sasson, S. (2013). Signaling properties of 4-hydroxyalk-enals formed by lipid peroxidation in diabetes. Free Radic. Biol. Med. 65: 978–987.
Cohn, J.S. (2002). Oxidized fat in the diet, postprandial lipaemia and cardio-vascular disease. Curr. Opin. Lipidol. 13: 19–24.
Conklin, D.J., Barski, O.A., Lesgards, J.F., Juvan, P., Rezen, T., Rozman, D., Prough, R.A., Vladykovskaya, E., Liu, S., Srivastava, S., and Bhatnagar, A. (2010). Acrolein consumption induces systemic dyslipidemia and lipoprotein modification. Toxicol. Appl. Pharmacol. 243(1): 1–12.
Conklin, D.J., Bhatnagar, A., Cowley, H.R., Johnson, G.H., Wiechmann, R.J., Sayre, L.M., Trent, M.B., and Boor, P.J. (2006). Acrolein generation stimulates hypercontraction in isolated human blood vessels. Toxicol. Appl. Pharmacol. 217(3): 277–288.
Conklin, D.J., Prough, R.A., Juvan, P., Rezen, T., Rozman, D., Haberzettl, P.,

Journal of Food Bioactives | www.isnff-jfb.com60
Potential adverse health effects Hu et al.
Srivastava, S., and Bhatnagar, A. (2011). Acrolein-induced dyslipidem-ia and acute-phase response are independent of HMG-CoA reduc-tase. Mol. Nutr. Food Res. 55(9): 1411–1422.
Draper, H.H., McGirr, L.G., and Hadley, M. (1986). The metabolism of malondialdehyde. Lipids 21(4): 305–307.
Dung, C.H., Wu, S.C., and Yen, G.C. (2006). Genotoxicity and oxidative stress of the mutagenic compounds formed in fumes of heated soybean oil, sunflower oil and lard. Toxicol. In Vitro 20(4): 439–447.
Eckl, P.M., and Bresgen, N. (2017). Genotoxicity of lipid oxidation com-pounds. Free Radic. Biol. Med. 111: 244–252.
Endemann, D.H., and Schiffrin, E.L. (2004). Endothelial dysfunction. J. Am. Soc. Nephrol. 15(8): 1983–1992.
Endo, K., Oyama, T., Saiki, A., Ban, N., Ohira, M., Koide, N., Murano, T., Wa-tanabe, H., Nishii, M., Miura, M., Sekine, K., Miyashita, Y., and Shirai, K. (2008). Determination of serum 7-ketocholesterol concentrations and their relationships with coronary multiple risks in diabetes mel-litus. Diabetes Res. Clin. Pract. 80(1): 63–68.
Esterbauer, H. (1993). Cytotoxicity and genotoxicity of lipid-oxidation prod-ucts. Am. J. Clin. Nutr. 57(5 Suppl): 779S–785S.
Ferderbar, S., Pereira, E.C., Apolinário, E., Bertolami, M.C., Faludi, A., Mon-te, O., Calliari, L.E., Sales, J.E., Gagliardi, A.R., Xavier, H.T., and Abdalla, D.S. (2007). Cholesterol oxides as biomarkers of oxidative stress in type 1 and type 2 diabetes mellitus. Diabetes Metab. Res. Rev. 23(1): 35–42.
Gargiulo, S., Testa, G., Gamba, P., Staurenghi, E., Poli, G., and Leonarduzzi, G. (2017). Oxysterols and 4-hydroxy-2-nonenal contribute to athero-sclerotic plaque destabilization. Free Radic. Biol. Med. 111: 140–150.
Gęgotek, A., and Skrzydlewska, E. (2019). Biological effect of protein modifi-cations by lipid peroxidation products. Chem. Phys. Lipids 221: 46–52.
Gianazza, E., Brioschi, M., Fernandez, A.M., and Banfi, C. (2019). Lipoxida-tion in cardiovascular diseases. Redox Biol. 23: 101119.
Gökçe, A.B., Eren, B., and Sağir, D. (2020). Inhibition of acrolein-induced ap-optosis by the antioxidant selenium. Toxicol. Ind. Health 36(2): 84–92.
Gomes, R., Meek, M.E., and Eggleton, M. (2002). Concise International Chemical Assessment Document 43: Acrolein. Ipcs Concise Interna-tional Chemical Assessment Documents.
Grootveld, M., Atherton, M.D., Sheerin, A.N., Hawkes, J., Blake, D.R., Rich-ens, T.E., Silwood, C.J., Lynch, E., and Claxson, A.W. (1998). In vivo ab-sorption, metabolism, and urinary excretion of alpha,beta-unsaturated aldehydes in experimental animals. Relevance to the development of cardiovascular diseases by the dietary ingestion of thermally stressed polyunsaturate-rich culinary oils. J. Clin. Invest. 101(6): 1210–1218.
Grootveld, M., Percival, B.C., Leenders, J., and Wilson, P.B. (2020). Poten-tial Adverse Public Health Effects Afforded by the Ingestion of Dietary Lipid Oxidation Product Toxins: Significance of Fried Food Sources. Nutrients 12(4): 974.
Grúz, P., Shimizu, M., Sugiyama, K.I., and Honma, M. (2017). Mutagenicity of ω-3 fatty acid peroxidation products in the Ames test. Mutat. Res. 819: 14–19.
Guillén, M.D., and Goicoechea, E. (2008). Toxic Oxygenated α,β-Unsaturated Aldehydes and their Study in Foods: A Review. Crit. Rev. Food Sci. Nutr. 48(2): 119–136.
Hageman, G., Verhagen, H., Schutte, B., and Kleinjans, J. (1991). Biological effects of short-term feeding to rats of repeatedly used deep-frying fats in relation to fat mutagen content. Food Chem. Toxicol. 29(10): 689–698.
Hajri, T. (2018). Effects of oxidized lipids and lipoproteins on cardiac func-tion. Front. Biosci. (Landmark Ed) 23: 1822–1847.
Hira, T., Yahagi, A., Nishimura, S., Sakaino, M., Yamashita, T., and Hara, H. (2015). Diunsaturated Aldehyde, trans,trans-2,4-Decadienal in the In-testinal Lumen Suppresses Gastric Emptying through Serotonin Sign-aling in Rats. J. Agric. Food Chem. 63(37): 8177–8181.
Hong, J.H., Lee, P.A.H., Lu, Y.C., Huang, C.Y., Chen, C.H., Chiang, C.H., Chow, P.M., Jaw, F.S., Wang, C.C., Huang, C.Y., Wang, T.W., Liu, J.H., and Wang, H.T. (2020). Acrolein contributes to urothelial carcinomas in patients with chronic kidney disease. Urol. Oncol. 38(5): 465–475.
Hu, Y., Yin, F., Yu, Z., Peng, Y., Zhao, G., Liu, Z., Zhou, D., Ma, X., Shahidi, F., and Zhu, B. (2020). Trans, trans-2,4-decadienal impairs vascular en-dothelial function by inducing oxidative/nitrative stress and apopto-sis. Redox Biol. 34: 101577.
Huang, Y.J., Zhang, L., Shi, L.Y., Wang, Y.Y., Yang, Y.B., Ke, B., Zhang, T.Y., and
Qin, J. (2018). Caloric restriction ameliorates acrolein-induced neuro-toxicity in rats. Neurotoxicology 65: 44–51.
Hughes, T.M., Kuller, L.H., Lopez, O.L., Becker, J.T., Evans, R.W., Sutton-Tyr-rell, K., and Rosano, C. (2012). Markers of cholesterol metabolism in the brain show stronger associations with cerebrovascular disease than Alzheimer’s disease. J. Alzheimers Dis. 30(1): 53–61.
Jaganjac, M., Tirosh, O., Cohen, G., Sasson, S., and Zarkovic, N. (2013). Re-active aldehydes--second messengers of free radicals in diabetes mel-litus. Free Radic. Res. 47: 39–48.
Jayaraj, A.P., Rees, K.R., Tovey, F.I., and White, J.S. (1986). A molecular basis of peptic ulceration due to diet. Br. J. Exp. Pathol. 67(1): 149–155.
Jia, P.L., Zhang, C., Yu, J.J., Xu, C., Tang, L., and Sun, X. (2018). The risk of lung cancer among cooking adults: a meta-analysis of 23 observational studies. J. Cancer Res. Clin. Oncol. 144(2): 229–240.
Kamisah, Y., Ang, S., Othman, F., Nurul-Iman, B., and Qodriyah, H. (2016). Renoprotective effect of virgin coconut oil in heated palm oil diet-induced hypertensive rats. Appl. Physiol. Nutr. Metab. 41(10): 1033–1038.
Kanazawa, K., and Ashida, H. (1998). Dietary hydroperoxides of linoleic acid decompose to aldehydes in stomach before being absorbed into the body. Biochim. Biophys. Acta. 1393: 349–361.
Kang, S.C., Kim, H.W., Kim, K.B., Kwack, S.J., Ahn, I.Y., Bae, J.Y., Lim, S.K., and Lee, B.M. (2011). Hepatotoxicity and nephrotoxicity produced by 4-hydroxy-2-nonenal (4-HNE) following 4-week oral administration to Sprague-Dawley rats. J. Toxicol. Environ. Health A. 74(12): 779–789.
Kanner, J. (2007). Dietary advanced lipid oxidation endproducts are risk fac-tors to human health. Mol. Nutr. Food Res. 51(9): 1094–1101.
Kloudova-Spalenkova, A., Holy, P., and Soucek, P. (2020). Oxysterols in cancer management: From therapy to biomarkers. Br. J. Pharmacol. 178(16): 3235–3247.
Kovač, U., Skubic, C., Bohinc, L., Rozman, D., and Režen, T. (2019). Oxyster-ols and Gastrointestinal Cancers Around the Clock. Front. Endocrinol. 10: 483.
Lee, D.H., Nam, Y.J., and Lee, C.S. (2015). Apocynin attenuates cholesterol oxidation product-induced programmed cell death by suppressing NF-κB-mediated cell death process in differentiated PC12 cells. Neu-rochem. Int. 89: 28–39.
Lee, S.E., and Park, Y.S. (2013). Role of lipid peroxidation-derived α, β-unsaturated aldehydes in vascular dysfunction. Oxid. Med. Cell Lon-gev. 2013: 629028.
Leonarduzzi, G., Chiarpotto, E., Biasi, F., and Poli, G. (2010). 4-Hydroxynon-enal and cholesterol oxidation products in atherosclerosis. Mol. Nutr. Food Res. 49(11): 1044–1049.
Leong, X.F., Aishah, A., Nor Aini, U., Das, S., and Jaarin, K. (2008). Heated palm oil causes rise in blood pressure and cardiac changes in heart muscle in experimental rats. Arch. Med. Res. 39(6): 567–572.
Leong, X.F., Najib, M.N., Das, S., Mustafa, M.R., and Jaarin, K. (2009). Intake of repeatedly heated palm oil causes elevation in blood pressure with impaired vasorelaxation in rats. Tohoku J. Exp. Med. 219(1): 71–78.
Li, X., Yu, X., Sun, D., Li, J., Wang, Y., Cao, P., and Liu, Y. (2017). Effects of Polar Compounds Generated from the Deep-Frying Process of Palm Oil on Lipid Metabolism and Glucose Tolerance in Kunming Mice. J. Agric. Food Chem. 65(1): 208–215.
Liao, C.H., Shaw, H.M., and Chao, P.M. (2008). Impairment of glucose me-tabolism in mice induced by dietary oxidized frying oil is different from that induced by conjugated linoleic acid. Nutrition 24: 744–752.
Lin, P., Lee, H.L., Cheng, H.I., Chen, C.Y., Tsai, M.H., and Liu, H.J. (2014). Me-tabolomic profiling of mice urine and serum associated with trans-trans 2, 4-decadienal induced lung lesions by liquid chromatography-mass spectrometry. Anal. Bioanal. Chem. 406(17): 4287–4297.
Ma, Y., Deng, L., Ma, P., Wu, Y., Yang, X., Xiao, F., and Deng, Q. (2020). In vivo respiratory toxicology of cooking oil fumes: Evidence, mechanisms and prevention. J. Hazard. Mater. 402: 123455.
Malaguti, M., Cardenia, V., Rodriguez-Estrada, M.T., and Hrelia, S. (2019). Nutraceuticals and physical activity: Their role on oxysterols-mediat-ed neurodegeneration. J. Steroid. Biochem. Mol. Biol. 193: 105430.
Maldonado-Pereira, L., Schweiss, M., Barnaba, C., and Medina-Meza, I.G. (2018). The role of cholesterol oxidation products in food toxicity. Food Chem. Toxicol. 118: 908–939.
Mali, V.R., Pan, G., Deshpande, M., Thandavarayan, R.A., Xu, J., Yang, X.P., and Palaniyandi, S.S. (2016). Cardiac Mitochondrial Respiratory Dys-

Journal of Food Bioactives | www.isnff-jfb.com 61
Hu et al. Potential adverse health effects
function and Tissue Damage in Chronic Hyperglycemia Correlate with Reduced Aldehyde Dehydrogenase-2 Activity. PLoS One 11(10): e0163158.
Martín-Sierra, C., Laranjeira, P., Domingues, M.R., and Paiva, A. (2019). Li-poxidation and cancer immunity. Redox Biol. 23: 101103.
Menéndez-Carreño, M., Varo, N., Mugueta, C., Restituto, P., Ansorena, D., and Astiasarán, I. (2011). Correlation between serum content of the main COPs (cholesterol oxidation products) from autoxidation and cardiovascular risk factors. Nutr. Hosp. 26(1): 144–151.
Meynier, A., Lherminier, J., Demaison-Meloche, J., Ginies, C., Grandgirard, A., and Demaison, L. (2002). Effects of dietary oxysterols on coronary arteries in hyperlipidaemic hamsters. Br. J. Nutr. 87(5): 447–458.
Mhatre, S., Rajaraman, P., Chatterjee, N., and Bray, F. (2020). Mustard oil consumption, cooking method, diet and gallbladder cancer risk in high- and low-risk regions of India. Int. J. Cancer 147(6): 1621–1628.
Nakamura, K., Kusano, K.F., Matsubara, H., Nakamura, Y., Miura, A., Nishii, N., Banba, K., Nagase, S., Miyaji, K., Morita, H., Saito, H., Emori, T., and Ohe, T. (2005). Relationship between oxidative stress and systolic dys-function in patients with hypertrophic cardiomyopathy. J. Card. Fail. 11(2): 117–123.
Nazih, H., and Bard, J. (2020). Cholesterol, Oxysterols and LXRs in Breast Cancer Pathophysiology. Int. J. Mol. Sci. 21: 1356.
Nkanu, E.E., Owu, D.U., and Osim, E.E. (2018). Altered Potassium Ion Chan-nel Function as a Possible Mechanism of Increased Blood Pressure in Rats Fed Thermally Oxidized Palm Oil Diets. J. Diet. Suppl. 15(4): 431–444.
Nurul-Iman, B., Kamisah, Y., Jaarin, K., and Qodriyah, H. (2013). Virgin co-conut oil prevents blood pressure elevation and improves endothelial functions in rats fed with repeatedly heated palm oil. Evidence-based complementary and alternative medicine. Evid. Based Complement. Alternat. Med. 2013: 629329.
Obembe, A.O., Owu, D.U., Okwari, O.O., Antai, A.B., and Osim, E.E. (2011). Intestinal Fluid and Glucose Transport in Wistar Rats following Chronic Consumption of Fresh or Oxidised Palm Oil Diet. ISRN Gastroenterol. 2011: 972838.
Perez, C., Lopez de Cerain, A., and Bello, J. (2002). Modulation of mutagenic activity in meat samples after deep-frying in vegetable oils. Mutagen-esis 17(1): 63–66.
Perluigi, M., Coccia, R., and Butterfield, D.A. (2012). 4-Hydroxy-2-nonenal, a reactive product of lipid peroxidation, and neurodegenerative dis-eases: a toxic combination illuminated by redox proteomics studies. Antioxid. Redox Signal. 17(11): 1590–1609.
Pillon, N.J., Croze, M.L., Vella, R.E., Soulère, L., Lagarde, M., and Soulage, C.O. (2012). The lipid peroxidation by-product 4-hydroxy-2-nonenal (4-HNE) induces insulin resistance in skeletal muscle through both carbonyl and oxidative stress. Endocrinology 153(5): 2099–2111.
Pillon, N.J., Vella, R.E., Souleere, L., Becchi, M., Lagarde, M., and Soulage, C.O. (2011). Structural and functional changes in human insulin in-duced by the lipid peroxidation byproducts 4-hydroxy-2-nonenal and 4-hydroxy-2-hexenal. Chem. Res. Toxicol. 24(5): 752–62.
Poli, G., Biasi, F., and Leonarduzzi, G. (2013). Oxysterols in the pathogenesis of major chronic diseases. Redox Biol. 1(1): 125–130.
Ringseis, R., and Eder, K. (2011). Regulation of genes involved in lipid me-tabolism by dietary oxidized fat. Mol. Nutr. Food Res. 55(1): 109–121.
Rodriguez-Amaya, D.B., and Shahidi, F. (2021). Oxidation of lipids. Chemi-cal Changes during Processing and Storage of Foods. Academic Press, pp. 125–170.
Rom, O., Korach-Rechtman, H., Hayek, T., Danin-Poleg, Y., Bar, H., Kashi, Y., and Aviram, M. (2017). Acrolein increases macrophage atherogenicity in association with gut microbiota remodeling in atherosclerotic mice: protective role for the polyphenol-rich pomegranate juice. Arch. Toxi-col. 91(4): 1709–1725.
Rossin, D., Calfapietra, S., Sottero, B., Poli, G., and Biasi, F. (2017). HNE And Cholesterol Oxidation Products In Colorectal Inflammation And Car-cinogenesis. Free Radic. Biol. Med. 111: 186–195.
Rouaki, F., Mazari, A., Kanane, A., Errahmani, M.B., and Ammouche, A. (2013). Cardiotoxicity induced by dietary oxidized sunflower oil in rats: pro- and antioxidant effects of α-tocopherol. Int. J. Vitam. Nutr. Res. 83(6): 367–376.
Samadi, A., Gurlek, A., Sendur, S.N., Karahan, S., Akbiyik, F., and Lay, I. (2019). Oxysterol species: reliable markers of oxidative stress in dia-
betes mellitus. J. Endocrinol. Invest. 42(1): 7–17.Sardar Sinha, M., Villamil Giraldo, A.M., Öllinger, K., Hallbeck, M., and
Civitelli, L. (2018). Lipid vesicles affect the aggregation of 4-hydroxy-2-nonenal-modified α-synuclein oligomers. Biochim. Biophys. Acta Mol. Basis Dis. 1864(9 Pt B): 3060–3068.
Selmanoğlu, G., Mülayimçelik Özgün, G., and Karacaoğlu, E. (2018). Acr-olein-mediated neurotoxicity in growing Wistar male rats. Pestic. Bio-chem. Physiol. 149: 37–43.
Shahidi, F., and Zhong, Y. (2010). Lipid oxidation and improving the oxida-tive stability. Chem. Soc. Rev. 39(11): 4067–79.
Siti, H., Kamisah, Y., Nur Iliyani, M., Mohamed, S., and Jaarin, K. (2017). Citrus leaf extract reduces blood pressure and vascular damage in repeatedly heated palm oil diet-Induced hypertensive rats. Biomed. Pharmacother. 87: 451–460.
Siti, H.N., Kamisah, Y., Mohamed, S., and Jaarin, K. (2018). Effects of cit-rus leaf extract on aortic vascular reactivity in hypertensive rats fed repeatedly heated vegetable oil. Appl. Physiol. Nutr. Metab. 44(4): 373–380.
Soriguer, F., Rojo-Martínez, G., Dobarganes, M.C., García Almeida, J.M., Esteva, I., Beltrán, M., Ruiz De Adana, M.S., Tinahones, F., Gómez-Zu-maquero, J.M., García-Fuentes, E., and González-Romero, S. (2003). Hypertension is related to the degradation of dietary frying oils. Am. J. Clin. Nutr. 78(6): 1092–1097.
Sottero, B., Rossin, D., Poli, G., and Biasi, F. (2018). Lipid Oxidation Products in the Pathogenesis of Inflammation-related Gut Diseases. Curr. Med. Chem. 25(11): 1311–1326.
Soulage, C.O., Sardón Puig, L., Soulère, L., Zarrouki, B., Guichardant, M., Lagarde, M., and Pillon, N.J. (2018). Skeletal muscle insulin resistance is induced by 4-hydroxy-2-hexenal, a by-product of n-3 fatty acid per-oxidation. Diabetologia 61(3): 688–699.
Sozen, E., Yazgan, B., and Sahin, A. (2018). High Cholesterol Diet-Induced Changes in Oxysterol and Scavenger Receptor Levels in Heart Tissue. Oxid. Med. Cell Longev. 2018: 8520746.
Srivastava, S., Sithu, S.D., Vladykovskaya, E., Haberzettl, P., Hoetker, D.J., Siddiqui, M.A., Conklin, D.J., D’Souza, S.E., and Bhatnagar, A. (2011). Oral exposure to acrolein exacerbates atherosclerosis in apoE-null mice. Atherosclerosis 215(2): 301–308.
Staprans, I., Hardman, D.A., Pan, X.M., and Feingold, K.R. (1999). Effect of oxidized lipids in the diet on oxidized lipid levels in postprandial serum chylomicrons of diabetic patients. Diabetes Care 22(2): 300–306.
Staprans, I., Pan, X.M., Rapp, J.H., and Feingold, K.R. (2005). The role of di-etary oxidized cholesterol and oxidized fatty acids in the development of atherosclerosis. Mol. Nutr. Food Res. 49(11): 1075–1082.
Staprans, I., Pan, X.M., Rapp, J.H., Grunfeld, C., and Feingold, K.R. (2000). Oxidized cholesterol in the diet accelerates the development of ath-erosclerosis in LDL receptor- and apolipoprotein E-deficient mice. Ar-terioscler. Thromb. Vasc. Biol. 20(3): 708–714.
Stott-Miller, M., Neuhouser, M.L., and Stanford, J.L. (2013). Consumption of deep-fried foods and risk of prostate cancer. Prostate 73(9): 960–969.
Testa, G., Rossin, D., Poli, G., Biasi, F., and Leonarduzzi, G. (2018). Implica-tion of oxysterols in chronic inflammatory human diseases. Biochimie 153: 220–231.
Vieira, S.A., Zhang, G., and Decker, E.A. (2017). Biological Implications of Lipid Oxidation Products. J. Am. Oil Chem. Soc. 94(3): 339–351.
Viktorinova, A., Svitekova, K., Stecova, A., and Krizko, M. (2016). Relation-ship between selected oxidative stress markers and lipid risk factors for cardiovascular disease in middle-aged adults and its possible clini-cal relevance. Clin. Biochem. 49(12): 868–872.
Wang, C.K., Chang, L.W., Chang, H., Yang, C.H., Tsai, M.H., Tsai, H.T., and Lin, P. (2010). Pulmonary changes induced by trans,trans-2,4-decadienal, a component of cooking oil fumes. Eur. Respir. J. 35(3): 667–675.
Wang, G.W., Guo, Y., Vondriska, T.M., Zhang, J., Zhang, S., Tsai, L.L., Zong, N.C., Bolli, R., Bhatnagar, A., and Prabhu, S.D. (2008). Acrolein con-sumption exacerbates myocardial ischemic injury and blocks nitric oxide-induced PKCε signaling and cardioprotection. J. Mol. Cell. Car-diol. 44(6): 1016–1022.
Willinger, T. (2019). Oxysterols in intestinal immunity and inflammation. J. Intern. Med. 285(4): 367–380.
Wu, S.C., Yen, G.C., and Sheu, F. (2001). Mutagenicity and identification of mutagenic compounds of fumes obtained from heating peanut oil. J. Food Prot. 64(2): 240–245.

Journal of Food Bioactives | www.isnff-jfb.com62
Potential adverse health effects Hu et al.
Yoshioka, N., Adachi, J., Ueno, Y., and Yoshida, K. (2005). Oxysterols in-crease in diabetic rats. Free Radic. Res. 39(3): 299–304.
Zaguri, R., Verbovetski, I., Atallah, M., Trahtemberg, U., Krispin, A., Nahari, E., Leitersdorf, E., and Mevorach, D. (2007). ‘Danger’ effect of low-density lipoprotein (LDL) and oxidized LDL on human immature den-dritic cells. Clin. Exp. Immunol. 149(3): 543–552.
Zárate, J., Goicoechea, E., Pascual, J., Echevarría, E., and Guillén, M.D. (2009). A study of the toxic effect of oxidized sunflower oil containing 4-hydroperoxy-2-nonenal and 4-hydroxy-2-nonenal on cortical TrkA receptor expression in rats. Nutr. Neurosci. 12(6): 249–259.
Zarkovic, K., Jakovcevic, A., and Zarkovic, N. (2017). Contribution of the HNE-immunohistochemistry to modern pathological concepts of ma-jor human diseases. Free Radic. Biol. Med. 111: 110–126.
Zhou, Z., Wang, Y., Jiang, Y., Diao, Y., Strappe, P., Prenzler, P., Ayton, J., and Blanchard, C. (2016). Deep-fried oil consumption in rats impairs glyc-erolipid metabolism, gut histology and microbiota structure. Lipids Health Dis. 15(1): 86.
Zirak, M.R., Mehri, S., Karimani, A., Zeinali, M., Hayes, A.W., and Karimi, G. (2019). Mechanisms behind the atherothrombotic effects of acrolein, a review. Food Chem. Toxicol. 129: 38–53.

Copyright: © 2021 International Society for Nutraceuticals and Functional Foods.All rights reserved.
63
Review J. Food Bioact. 2021;15:63–73
Journal ofFood Bioactives International Society for
Nutraceuticals and Functional Foods
Functional foods beyond nutrition: therapeutic interventions to combat COVID-19 and other viral diseases
Samudra P. Banika, Rituparna Banik Ghoshb, Bernard W. Downsc, Sanjoy Chakrabortyd, Manashi Bagchie, Tandra R. Chakrabortyf and Debasis Bagchic,e,g*
aDepartment of Microbiology, Maulana Azad College. Kolkata, IndiabDepartment of Physiology, Bhairab Ganguly College, Kolkata, IndiacVictory Nutrition International, Inc., Dept of R&D, Bonita Springs, FL, USAdDepartment of Biological Sciences, New York City College of Technology/CUNY, Brooklyn, NY, USAeDepartment of Biology, Adelphi University, New York City, NY, USAfDr. Herbs LLC, Dept of R&D, Concord, CAgDepartment of Pharmaceutical Sciences, Texas Southern University, Houston, TX, USA*Corresponding author: Debasis Bagchi, Department of Pharmaceutical Sciences, Texas Southern University, Houston, TX, USA. E-mail: [email protected]: 10.31665/JFB.2021.15283Received: September 26, 2021; Revised received & accepted: September 30, 2021Citation: Banik, S.P., Ghosh, R.B, Downs, B.W., Chakraborty, S., Bagchi, M., Chakraborty, T.R., and Bagchi, D. (2021). Functional foods beyond nutrition: therapeutic interventions to combat COVID-19 and other viral diseases. J. Food Bioact. 15: 63–73.
Abstract
SARS-COV2 has emerged as the second biggest threat to humanity of all times, after the Spanish Flu massacre. Due to its brisk rate of mutation, especially that of the host receptor docking Spike protein, a safe immunization is still questioned by some, and protection or recovery has been dependent largely on immune system compe-tence and overall bio-constitutional strength. In order to ensure optimal performance of the body’s immune cells, proper nutrition fortified with functional food ingredients is indispensable. Especially in a scenario where targeted therapeutics against life threatening pandemics like SARS-CoV2 are yet to be invented, formulation of dietary supplements rich in antioxidants, vitamins, and minerals is the need of the hour to promote greater immuno-competence and a heightened immune response; decrease the overall metabolic burden; and restore homeostasis in an infected individual. This review highlights our current understanding and advances made in the regimen of functional foods designed to combat viral diseases together with the shortcomings involved therein.
Keywords: SARS-CoV2; Antivirals; Functional food; Nutraceuticals; Healthy immune system.
1. Introduction
Viral outbreaks had been the cause of serious concern for human health and well-being since time immemorial. The Spanish flu or the Great Influenza Epidemic of 1918 caused by the H1N1 influ-enza virus was one such major documented catastrophe that in-fected about one third’s of the world population and took away the lives of at least 50 million people. Since then, the COVID-19 pandemic, has emerged as the biggest threat to human life and civi-lization with more than 220 million people affected worldwide up
to the date of this writing. The pandemic, caused by an enveloped (+) single stranded and non-segmented RNA virus belonging to Coronaviridae family, has currently a global death toll exceeding 4.5 million. This has been the third and most significant attack unleashed on humankind from the betacoronaviridae family fol-lowing the outbreaks of SARS-CoV-1 in 2003 and MERS-CoV in 2012. The other four human coronaviruses, which resulted in mild symptoms, were HKU1, NL63, OC43 and 229E (Cui et al., 2019). In order to combat these recurrent viral infections, aside from de-veloping natural immunity from getting COVID-19, the best pos-sible way is to prime our immune system with the specific antiviral

Journal of Food Bioactives | www.isnff-jfb.com64
Functional foods beyond nutrition Banik et al.
antibodies is through immunization. However, the development of a vaccine against SARS-CoV-2 has been a challenging task due to incomplete understanding of the life cycle, epidemiology, and specific immune responses directed against the virus (Ahmed et al., 2020). Moreover, emerging evidence indicates that there is a high probability that the efficacy of vaccines will decline over time due to rapid changes in what is believed to be caused by the ge-netic makeup of the virus. Therefore, although getting immunized is critical in maximizing protection against the viral infection, vac-cination cannot safeguard a person completely from acquiring the viral disease. Under these circumstances, boosting our own innate immune response forms the foundation to sufficiently bolster com-petent resistance to viral infection. In order to possess a healthy immune system, maintenance of a lifestyle that promotes an aero-bic metabolism with optimal nutrition is of paramount importance.
Viruses enter our cells through specific entry points utilizing de-fined molecular targets. Therefore, in addition to consumption of a balanced diet fortified with adequate minerals and vitamins, it is also essential at the same time to identify nutraceuticals and functional foods that can be administered in addition to the staple diet. These are supposed to act as therapeutic interventions to help blocking vi-ral entry points and augment the immune response (Figure 1).
2. Identifying potential check points of viral infections for therapeutic intervention
Viruses enter host cells via several mechanisms including endo-cytosis, macrocytosis, pinocytosis, and phagocytosis. They can also fuse at the plasma membrane and spread within the host via cell-to-cell fusion or syncytia. Generally, alpha- and beta-corona-viruses employ angiotensin-converting enzyme 2 (ACE2) recep-tors for infecting lung epithelial cells (Li et al., 2003). ACE2 is a transmembrane carboxypeptidase of the Renin-Angiotensin Sys-tem implicated in vasodilation following hypertension and related ailments (Tikellis and Thomas, 2012), which acts as a receptor for
the viral Spike protein (Shang et al., 2020). However, exceptions to this general route have also been reported frequently. Camel alpha-coronaviruses, human alpha-coronavirus 229E (along with porcine delta-coronavirus) are known to infect host cells by docking onto corresponding Aminopeptidase N receptors (Li et al., 2020). Ad-ditionally, canine respiratory beta-coronavirus is known to employ Human Leukocyte Antigen Class 1 (HLA-1) as receptor for host cell entry (Szczepanski et al., 2019).The coronaviruses are made up of four structural proteins, namely, the spike (S), membrane (M),envelop (E) and nucleocapsid (N) proteins. The spike protein of SARS-CoV-2 is the specific docking ligand of ACE2, which consists of two domains; a pleated S1 domain or Receptor Binding Domain, and a helical S2 domain which enables the virus to fuse with host cell membrane. In SARS-CoV-2 and a few other corona-viruses, there is a stretch of amino acids (PRRARS/V) between the S1 docking and S2 fusion domain which serves as a recognition site for the host protease furin. Cleavage by furin occurs as a natural defence mechanism of the host, but acts in the favour of the virus by exposing the access of the S2 fusion domain to the host membrane for fusion and entry. This also explains why infection by SARS-CoV-2 occurs selectively for the ciliated epithelial cells of the lungs where there is an abundance of furin proteases. The S gene corre-sponding to the spike protein is one of the hypervariable spots of the viral genome (Wen et al., 2020), which enables it to quickly change and successfully use a number of zoonotic carriers as natural res-ervoirs and intermediate hosts before infecting humans. This also explains the significantly faster rate of mutation of SARS-COV2 and appearance of its variants. In addition to ACE-2, the S1 domain of the spike glycoprotein is also known to interact with the human CD26 (also known as Dipeptidyl Peptidase IV or DPP4), a multi-functional transmembrane type II glycoprotein, expressed consti-tutively on several cell types including epithelial cells, endothelial cells, and fibroblasts, as well as many immune cells, including T, B, and natural killer lymphocytes, and macrophages. There is also a soluble counterpart of the molecule in plasma and its enhanced expression has been implicated in down regulating inflammatory response (Busso et al., 2005, Sargiacomo et al., 2020).
Figure 1. Potential targets of therapeutic interventions to arrest viral progression: As shown in numbered red circles 1. Therapeutics to sequester and deplete DPP4, a potent receptor of SARS-coronavirus 2. Drugs targeted against NOX2-depenedent NADH oxidase complex to prevent superoxide mediated oxidation of TLR7 and subsequent inhibition in secretion of IFN 3. Therapeutics to modulate the expression of gelsolin and hence regulate the density de-pendent phenotype of macrophages to minimize host damage arising out of immune response.

Journal of Food Bioactives | www.isnff-jfb.com 65
Banik et al. Functional foods beyond nutrition
CD26 is often hijacked by viruses during infection and subsequent occupancy of the cellular machinery, and therefore forms a potential target molecule for designing anti-viral therapeutics (Vankadari and Wilce, 2020). After successfully mediating membrane fusion, the viral RNA in the pulmonary alveolar epithelial cells is endocytosed and released into the cytosol by lysis of the endosomal membrane with subsequent intracellular release of viral contents. Host immune response is initiated at this point as trapped single stranded viral RNA primes the Toll Like Receptor 7 (TLR7) on the endosomal membrane thereby signalling the release of Type 1 interferon (Die-bold et al., 2004). Thereafter, the virus synthesizes a negative strand RNA from the pre-existing single-strand positive RNA. The nega-tive strand RNA serves to produce new strands of positive RNAs which are deployed in translation. Assembly of new virion particles takes place in the cellular endoplasmic reticulum with the viral N and M proteins from where they are exocytosed into the extracellular space via Golgi derived vesicles. The new viral particles can then initiate a fresh round of infection of the adjacent epithelial cells or infect other individuals through burst of respiratory droplets.
Coronaviruses and a wide group of other RNA viruses includ-ing Sendai virus, HIV, rhinovirus, human meta pneumonia virus, respiratory syncytial virus, human para influenza virus, Dengue virus, etc., have adopted a clever strategy to immediately sup-press the release of interferon. These viruses induce production of superoxides in the endosomal compartment of human alveolar macrophages by NOX2-dependent NADPH oxidase complexes (Sindona et al., 2021). The generated hydrogen peroxide oxidises the Cys98residue on TLR7 that permanently blocks transmission of signal for type 1 interferon production. This was demonstrated in both mutant macrophages deficient in NOX2 activity, and in cell lines with a targeted NOX2 inhibitor (gp91ds-TAT) where the level of type 1 interferon was markedly higher in response to RNA virus infection. Therefore, novel NOX2 inhibitors can act as effec-tive therapeutic agents to arrest viral progression.
Cardinal signs of COVID-19 infection are initiated from this phase onwards when the virus migrates to the upper respiratory tract and fever accompanied with dry cough begins. The immune response also heightens at this phase with the release of C-X-C motif chemokine ligand 10 (CXCL-10) and interferons (IFN-β and IFN-λ) from the virus-infected cells (Tang et al., 2005), chiefly the lung pneumocytes. Cytokines attract and recruit a battery of im-mune cells including a large number of macrophages, neutrophils, CD4+ helper T cells, and CD8+ cytotoxic T cells. The immune response is further augmented by the large scale release of pro-inflammatory cytokines including interleukins (IL-1, IL-6, IL-8, IL-120 and IL-12), tumour necrosis factor-α (TNF-α), IFN-λ, and IFN-β, CXCL-10, monocyte chemo-attractant protein-1 (MCP-1), and macrophage inflammatory protein-1α (MIP-1α). The events collectively lead to the elicitation of the characteristic “cytokine storm” and many cells die out of lung injury. In more extreme cases, fluid starts accumulating inside the lung cells and there is loss of both type 1 and type 2 pneumocytes; if left untreated, these complex events can gain pathological momentum and morph into the development of Acute Respiratory Distress Syndrome (Xu et al., 2020), the most significant cause behind COVID-19 inflicted death. A recent significant finding has indicated that the mac-rophage population at the site of inflammation is regulated by a quorum sensing phenomenon, which limits the extent of inflam-matory response and minimizes host damage as a consequence of cytokine storm. The secreted form of the actin severing protein gelsolin, has been identified as the key auto-inducer of the quorum sensing phenomenon. Gelsolin can therefore be another potential target for developing new generation therapeutics for uncontrolled immune proliferation (Sharma et al., 2021).
3. Role of diet in establishing a healthy gut microbiome and an optimally functioning immune system
It has been unequivocally demonstrated that optimal nutrition is vital for keeping ourselves healthy. Nutritional deficits can ad-versely affect the immune system and hence curb the potential to fight microbial infections (Chandra, 1996). Specific components of nutrients are implicated in activation of immune functions through modulation of gene expression and signalling molecules (Valdés-Ramos et al., 2010). In addition to other numerous ben-efits, dietary ingredients can also influence immune responses through their effects on gut microbiota composition (Singh et al. 2017); the evolution and development of our immune system owes a lot to our healthy microbiome and therefore these two have been intricately associated with each other since the birth of a child. This is best portrayed by regulatory T (Treg) cells, a subset of TH cells, which mediate immune tolerance to self-antigens as well as to the antigenic determinants of commensals colonizing the human microbiome (Cebula et al., 2013).
Babies feeding on breast milk develop a solid foundation of their immune response compared to those consuming formula milk. Apart from its nutritive constituents, mother’s milk contains plenty of bioactive components including lysozyme, sIgA, alpha-lactalbumin, lactoferrin, free oligosaccharides, complex lipids, and glycoconjugates, which play a significant role in the maturation of newborn immunological responses (Lazar et al., 2018). A major glycoprotein of milk, k-casein, is proteolytically cleaved to gener-ate glycomacropeptides, which prevents the growth of pathogens on gut epithelial cells (Gordon et al., 2012). Mother’s milk also contains lactoferricin, a potent antimicrobial agent that combats pathogen colonization. Several key interacting factors between malnutrition and diseases have been delineated and therefore many detrimental effects seen in under-nourished children can be controlled by prompt nutritional supplementation (Schuetz et al., 2019).
The Mediterranean diet has been one of the most sought af-ter food habits globally due to its proven effects in maintaining a healthy lifestyle and achieving wellness (Trichopoulou et al., 2003). It is rich in vegetables, fruits, legumes, nuts, beans, cere-als, grains, fish, and unsaturated fats, and generally includes a bare minimum of meat and dairy foods. Due to the presence of high levels of polyphenols, foods comprising a typical Mediterranean diet exhibit pleiotropic health-augmenting effects, especially with regards to alleviation of cardiovascular troubles and in the treat-ment of type-2 diabetes. Consumption of a plant and fish rich diet also benefits immensely the health of our gut microflora due to the presence of gut friendly dietary fibers and unsaturated fatty acids that have often been found to reverse dysbiosis caused by frenzied lifestyle. For example, strict adherence to a Mediterranean diet pattern resulted in a significant decrease in obesity of individuals habitually fed Western diets (Nagpal et al., 2019).
In spite of consumption of naturally nutritious diets fortified with all proximate and protective principles of food that can sup-posedly augment our immune system and enable us to stay strong and healthy against the menace of infectious diseases, over recent years, outbreak of deadly microbial pathogens have jeopardized human health. Consumption of nutritive foods alone have been found wanting in many cases to safeguard us. This has necessitated the introduction of functional foods and nutraceuticals as a figura-tive ‘shot in the arm’ for boosting our immune response against the onslaught of the deadly viruses and other pathogens.

Journal of Food Bioactives | www.isnff-jfb.com66
Functional foods beyond nutrition Banik et al.
4. Nutraceuticals and functional foods as indispensable boost ups of the immune system
Natural compounds and edible food ingredients contain many bio-active components that are capable of providing health benefits beyond the traditional nutritional value of food. These components have been ascribed the term “functional foods” or the dietary items that in addition to providing nutrients and energy, benefit certain tar-geted functions of the body (Nicoletti, 2012), possess the potential to boost immune response (López-Varela et al., 2002), and also re-duce the risk of diseases (Martirosyan and Singh, 2015). The term is sometimes used synonymously with “nutraceuticals”, which are rather extracted bioactive components of functional foods that can be converted for use in edible pills or capsules, or as fortified foods, dietary supplements, or herbal products marketed commercially.
Over last few decades, a large number of natural reservoirs of functional food components have been discovered including spices like black pepper, turmeric, ginger and garlic, fruits like pomegranate and elderberry, and other edible medicinal plants such as liquorice, which are reservoirs of immunomodulatory and anti-viral agents (Yang et al., 2020). For example, Bromelain, a cysteine protease, obtained from the pineapple stem (Ananas comosus) has tradition-ally been known for the management of trauma (Jain et al. 2012). In addition, bromelain has also been implicated in the inhibition of cyclooxygenase and hydrolysis of bradykinin, thus alleviating COV-ID-19 symptoms.
In spite of these sporadic but significant successes, scientists are yet to get confirmatory evidence regarding the efficacy of many more food plants in human systems in addition to complete elimi-nation of any toxic side effects. The important biomolecules which are found in abundance in plant extracts are as given in the follow-ing sub-sections.
4.1. Polyphenols
These are plant secondary metabolites providing a range of protec-tive functions against ultraviolet radiations, infectious diseases, and oxidative stress. Most of them have planar and fused phenolic rings with hydroxyl groups as substituents. They can be classified into phenolic acids, flavonoids, polyphenolic amides, and other polyphe-nol compounds (including stilbenes or lignans) (Annunziata et al., 2020). Polyphenols are proven inhibitors of inflammation and pre-vent expression of pro-inflammatory genes (Yahfoufi et al., 2018). Polyphenols have been found to be effective against a broad range of viruses including Epstein-Barr virus (Yiu et al., 2010), enterovi-rus 71 (Zhang et al., 2015), herpes simplex virus (HSV) (Faith et al., 2006), influenza virus (Lin et al., 2015), and other virus-causing respiratory tract-related infections (Zang et al., 2011). Most of the polyphenols exert their activity especially as anti-virals in a syner-gistic manner. For example, a crude extract prepared from Sambucas nigra was able to inhibit viral replication successfully in a Vero cell line and led to alteration of viral structure (Chen et al., 2014).
4.2. Molecular mechanism of action of polyphenols
Quite a few mechanisms to explain the antiviral properties of these compounds have been proposed. For example, polyphenols from Broussonetia papyrifera were found to inhibit the proteases of MERS/SARS-COV2 thereby effectively blocking the spread of the virus. Another commonly occurring polyphenol is trans-resveratrol, or 3,5,4′-trihydroxy-trans-stilbene, first isolated as a bioactive com-
ponent from the roots of Polygonum cuspidatum (Lin et al., 2015) (common name is Chinese knotweed) and subsequently discovered to occur in abundance in various types of grapes, berries, nuts, etc. This polyphenol is popular as a powerful antioxidant and prevents development of inflammatory responses. It has been found to be up-regulated under exposure to UV radiation, ozone, and in response to fungal infections (Pannu and Bhatnagar, 2019).In addition, res-veratrol has been demonstrated to be a potent inhibitor of upper res-piratory tract viral infections. Resveratrol directly interferes with the replication of influenza virus by blocking the nuclear to cytoplasmic translocation of Ribonucleoprotein particles (Palamara et al., 2005). In case of infection by coronaviruses, resveratrol has been found to significantly increase the survival rate of virus infected cells in the body by reducing their frequency of apoptosis (Lin et al., 2017). The antiviral properties of resveratrol are augmented by its strong immune modulating capabilities. Resveratrol alters cytokine and chemokine levels to down-regulate ROS-mediated inflammatory responses and leads to suppression of the NF-κB mediated pathway of activation of proinflammatory response (Meng et al., 2020). In addition, resveratrol administration has also been observed to result in upregulation of the IFNβ gene for arrest of viral replication (Lin et al., 2015). However, in spite of so many health augmenting roles of resveratrol, its therapeutic application has been restricted due to poor solubility and low bioavailability (Shaito et al., 2020).
4.3. The basis of antioxidative potential of green tea
Consumption of green tea had become popular globally over the last few decades owing to its proven role as a strong antioxidant. Green tea is made from the leaves of Camellia sinensis and the ac-tive components responsible for its antioxidative property are cat-echins. Catechins are also plant secondary metabolites belonging to the class of hydroxyflavans that have a flavan-3-ol skeleton and its substituted derivatives. Catechins comprise about 10% of the dry weight of green tea leaves and include (−)-epigallocatechin gallate (EGCG), (−)-epigallocatechin (EGC), (−)-epicatechin gallate (ECG) and (−)-epicatechin (EC) as the major constituents with EGCG ac-counting for about 50% of the total amount of catechins in green tea (Reygaert, 2018). EGCG has been reported to possess many dif-ferent pharmacotherapeutic properties including antioxidative, anti-bacterial, and antiviral roles (Song et al., 2005). EGCG serves as a potent inhibitor of HIV replication (Fassina et al., 2002) as well as an agglutinating agent of influenza viruses (Nakayama et al., 1993). Antiviral activity of catechins are believed to be due to their inhibi-tion of host-virus membrane fusion by prevention of acidification of the endosomal compartment (Imanishi et al., 2002).
4.4. Probiotics
The role of intestinal microflora in maintaining the overall health of the individual has been established beyond arguments. However, much like our own organs, the consortium of gut microbes are also susceptible to occasional dysbiosis resulting in altered functionali-ties with undesirable effects. Probiotics are live microbes that can supplement and replenish the ailing gut flora and restore homeo-stasis. Gut commensals connect and communicate with two major organs of the human body to ensure maintenance of physiological homeostasis; with the brain via the gut-brain axis (Carabotti et al., 2015) and with the lung via the gut-lung axis (Dang and Marsland, 2019). Much like the resident gut microbes, probiotics also can mod-ulate many host immune responses (Frei et al., 2015) and thereby curb the infectivity of a lot of viral pathogens, especially those in-

Journal of Food Bioactives | www.isnff-jfb.com 67
Banik et al. Functional foods beyond nutrition
volved in respiratory tract infections (Hao et al., 2015). Probiotics have already been recommended as a mitigation strategy for a weak-ened immune system following COVID-19 infection (Mirzaei et al., 2021) where gut microfloral dysbiosis with significant reductions in Lactobacillus and Bifidobacterium have been seen (Xu et al. 2020). They have been associated with increased expression of mucous glycoproteins to strengthen the epithelial barrier against viral inva-sion. Probiotics are also implicated in resurrection of tight junction complexes following virus inflicted damage through increased ex-pression and redistribution of zonula occluden (ZO-2) proteins and protein kinase C (PKC) (Yi et al., 2018).
4.5. Molecular mechanism of the action of probiotics
Although the exact mechanism of probiotic based prophylaxis of viral infections still remains to be delineated, these microbial strains are believed to exert their immunomodulatory effects via interac-tion of the bacterial constituents like peptidoglycan, lipoteichoic acid, and nucleic acid with their cognate Toll Like Receptors as well as muramyl dipeptide, which binds to the Nod-like receptors (Pimentel-Nunes et al., 2010) and subsequent modulation of Treg cells,TH17 cells, and type3 innate lymphoid cells (Kanauchi et al., 2018). Probiotic strains also boost type I interferon levels for en-hanced viral infection alarm and increase the number and activity of the immune arsenal comprised of NK cells, T cells, and levels of circulating antibodies. These are also implicated in modulating the levels of proinflammatory and immunoregulatory cytokines lead-ing to enhanced viral clearance with minimal bystander damage to lung cells. Collectively, the probiotics restrict viral replication by a) enhancement of the intestinal epithelial barrier b) attaching to and sequestering viruses c) by secreting antiviral components such as bacteriocins and hydrogen peroxide d) by induction of basal level nitric oxide (NO) generation by host cells e) by activation of macrophages and dendritic cells f) by inducing CD8+ T cells into Cytotoxic T Lymphocytes and g) by inducing differentiation of B cells by TH2 cells (Tapiovaara et al., 2016). Although rare incidents of probiotic-associated bacteremia and fungaemia have been reported in a few immunocompromised patients (Bertelli et al., 2015), probiotics are generally safe (Dermyshi et al., 2017).
4.6. Prebiotics
To keep our gut microbiota happy and rejuvenated, they have to be fed with specific nutrients that are not absorbable by our gut, but tailormade for these microbes. The gut microbiota thrives on certain plant derived dietary fibers, which human enzymes are in-capable of digesting. These dietary fibers are collectively known as prebiotics. Numerous common fruits and vegetables including apples, artichokes, bananas, berries, legumes, tomatoes, asparagus, etc. are enriched in prebiotic fibres including fructans, oligosac-charides, arabinooligosaccharides, isomaltooligosaccharides, xy-looligosaccharides, resistant starch, lactosucrose, lactobionic acid, galactomannan, psyllium, polyphenols, and polyunsaturated fatty acids, among others (Guarino et al., 2020). In addition to providing essential nutrients to the gut microbes, the prebiotics also benefit our body in a lot of other ways including facilitating absorption of calcium (Scholz-Ahrens et al., 2007), regulating the glycemic index (Connolly et al., 2012), boosting digestion (Cummings et al., 2001), and keeping the epithelial cell lining healthy (Carlson et al., 2018). New insights have also suggested the beneficial role of prebiotics in management of chronic gut disorders such as Irritable Bowel Syndrome (Wilson et al., 2019).
4.7. Antioxidant vitamins
Micronutrients, comprising vitamins and minerals, are named so due to the fact that requirements by our body are in relatively smaller or trace amounts. However, they are as important as the primary nutri-ents and their deficiency leads to impairment of many key metabolic reactions and biochemical pathways as well as malfunctioning of the host immune response (Opara, 2002). They have a significant role to play in maintaining the functionality of both innate and adaptive immune responses and micronutrient homeostasis is integral to the sustenance of a healthy immune system (Wintergerst et al., 2007). Some vitamins like A, C, E, flavonoids, and carotenoids are read-ily obtainable from the diet and therefore can function relentlessly in neutralizing oxidative free radicals. A loss in the balance of these elements can make a person susceptible to infection and diseases, and supplementation, on the other hand, can augment the recovery process, especially from viral infections (Jayawardena et al., 2020).
4.8. Vitamin D as a model vitamin with a functional food tag
A multidimensional vitamin in terms of therapeutic potential is Vi-tamin D, also referred to as “calciferol”. It is a fat-soluble vitamin existing in different forms that is naturally present in a few foods, but mostly produced endogenously by lower layers of skin epider-mis by a chemical reaction driven by the reaction of photons of sunlight with cholesterol (Holick et al., 1980). Hence, the major forms found in human circulation are Vit D3 (cholecalciferol) and D2 (ergocalciferol) (Bikle, 2014). Vitamin D obtained from food sources or synthesized in presence of sunlight is not biologically active. It is activated by two successive enzymatic hydroxylation events occurring respectively in the kidney and the liver, to yield 1,25-dihydroxyvitamin D (Bikle et al., 2002). Vitamin D is in-volved regulating many physiological processes inside the body including reabsorption of calcium and magnesium, which are also involved as pH buffers, mediating bone and extra-skeletal metabo-lism, regulating cardiovascular homeostasis, and most significant-ly modulating immunological responses of our bodies (Christakos et al., 2016). Vitamin D is also implicated in a marked reduction in pro-inflammatory cytokines thus decreasing the probability of a cytokine storm (Martineau et al., 2017) thereby reducing the need for prolonged inflammatory events. A recommended dose of vita-min D3 ranges from 2,000 to 10,000 IU/day.
4.9. Molecular basis of action of Vitamin D
Vitamin D enters inside the cell and exerts its effect by binding to its cognate nuclear receptor VDR. VDR exists as a heterodimer with an isoform of the retinoid X receptor (RXR). Ligand triggered con-formational change of VDR–RXR heterodimers induces dissocia-tion of co-repressor proteins such as nuclear receptor co-repressor (NCoR) and binding of the VDR-RXR heterodimer to the vitamin D response elements (VDRE) on the promoter regions of antimicrobial peptides (AMPs). These include human cathelicidin (human cation-ic antimicrobial protein; hCAP18) and human b-defensin-2 (hBD-2) (Beard et al., 2011). AMPs are localized on mucosal and epithelial layers and constitute one of the primary defenses against pathogens. On the other hand, epidermal expression of 1,25(OH)2D3 leads to expansion of CD4+ and CD25+ regulatory T cells (Loser and Beis-sert, 2009), and suppresses T cell proliferation. This is implicated in moderation of heightened immune response after the viral infection subsides to minimize host damage.

Journal of Food Bioactives | www.isnff-jfb.com68
Functional foods beyond nutrition Banik et al.
4.10. Role of micronutrients
Micronutrients also play an integral role in the alleviation of pathogen inflicted damage and restoration of homeostasis during recovery after viral replication. However several factors actually regulate the efficacy with which they can augment the recovery process; these include the kind of pathogen; the administered dose of the micronutrient; as well as the age, genetic background, and nutritional status and lifestyle of the patient, i.e. epigenetic factors (Lange, 2021). Immunosenescence or immune dysfunction during aging also profoundly affects the ability of the body to respond to nutrient functions (Yaqoob, 2017). The pharmacotherapeutic suc-cess of micronutrients is dependent upon a synergistic response of a wide range of components rather than administration of indi-vidual nutrients in isolation.
Zn deficiency has been implicated with reduced antibody pro-duction, and impaired activity of the innate immune system with a simultaneous decline in neutrophil population and secretion of cy-tokines by monocytes (Ibs and Rink, 2003). Suboptimal levels of this element have also been found to be associated with suppressed production of thymic hormones resulting in significant decrease in the number of peripheral and thymic T cells. Thymulin is a nona-peptide hormone responsible for inducing differentiation of T cells (Bach and Dardenne, 1989). Its active structure requires zinc and hence zinc deficiency also affects T cell proliferation.
4.11. Antiviral properties of Zinc and the mechanism involved
Zinc has also demonstrated antiviral properties, which is probably executed through metallothioneins (MT), a family of low molecu-lar weight, cysteine-rich proteins that mediates sequestration and release of Zn2+and are also involved in oxidative response, apo-ptosis, and immune responses (Subramanian and Deepe, 2017). Metallothioneins may either act directly by sequestering Zn2+ away from the viral metalloproteins or indirectly by inducing In-terferon gamma signalling. Zinc has also sometimes been used in conjunction with standard antiviral therapy where it has synergis-tically inhibited the folding and/or activity of viral enzymes and resulted in inhibition of viral replication. For example, the element has been shown to inhibit the template binding of RNA Depend-ent RNA Polymerase; arrest the replication of coronaviruses (Te Velthuis et al., 2010); inhibit the protease mediating viral poly-protein cleavage in Encephalomyocarditis virus (Butterworth and Korant, 1974); and inhibit RNA polymerase in Hepatitis C virus (Ferrari et al., 1999), DNA polymerase in Herpes Simplex virus (Fridlender et al., 1978), and many more. Zinc supplementation seems to have prospect in the prophylaxis and treatment of COV-ID-19 (Rahman and Idid, 2021). Accordingly, therapeutic trials of free zinc in the forms of creams, lozenges, and supplements had been initiated sporadically but most of them are yet to get approval for commercial applications.
4.12. Importance of selenium as an antiviral micronutrient
Alongside zinc, selenium is also an important micronutrient im-plicated in maintenance of immune health (Rayman, 2000) and therefore effective in antiviral therapy. Selenium supplementation has been known for its immune-supporting effect, which includes enhancement of activated T cells (cytotoxic lymphocytes) and pro-liferation of NK-cell activity (Kiremidjian-Schumacher and Roy, 1998). The element has also been found to be effective as an im-
munity booster in combination with certain vitamins such as D and E (Delesderrier et al., 2019). When administered in conjunction with phytoncides (present in onions), selenium results in increased T lymphocyte proliferation (Ru-duan et al., 1992) In addition, se-lenium can also prevent the formation of thrombosis in the blood vessels. Blood coagulation disorders leading to the formation of tiny clots constitute one of the significant reasons for death of patients with COVID-19 (Fogarty et al, 2020). Therefore, use of sodium selenite may reduce the formation of blood clots and in-crease the chances of patient’s survival. Selenium is a co-factor in glutathione peroxidase and can be reduced from the tetravalent selenite to the divalent cation (Se2+) thus acting as a powerful oxi-dant. This oxidizing capacity of selenite is exploited in its antiviral activity as selenite readily oxidizes the sulfhydryl groups in the active site of the viral protein disulphide isomerase (PDI) cross-linking them to inactive disulphide. This effectively stops the viral spike protein from engaging in exchange reactions with disulphide groups of cell membrane proteins and thus prevents viral entry into the healthy cell cytoplasm (Kieliszek and Lipinski, 2020).
4.13. Spices as functional foods
Plants are rich sources of flavonoids, antioxidants, polyphenols, coumarins, and steroids that help in boosting the immune system (Messina et al., 2020, Somerville et al., 2016). Spices can be pro-duced from any part of the plant, for example the leaf (e.g., bay leaf), buds (clove), bark (cinnamon), root (ginger), berries (grains of pepper), seeds (cumin), or even the stigma of the flower (saf-fron). Allicin, capsaicin, curcumin, gingerol, mustard oil, piperine, and quercetin glucosides are the major classes of spices implicated in augmentation of human health (García-Casal et al., 2016).
There have been proven immune benefits associated with many spices. Carnosol and carnosic acid are the two chief components of rosemary, which have demonstrated anti-HIV activity (Aruoma et al., 1996). Apigenin, a commonly occurring flavonoid present in many spices including thyme, sage, oregano, and rosemary, also have been associated with inhibition of transcription of HIV (Critchfield et al., 1996). Similarly, hydroalcoholic extracts of hys-sop had been shown to inhibit HIV activity (Bedoya et al., 2002). Many studies have also demonstrated the antiviral activity of cur-cumin (Joe et al., 2004, Lin et al., 2005), especially in Herpes Sim-plex Virus. Curcumin is a yellow polyphenolic curcuminoid ob-tained from Curcuma longa (turmeric) that possess a wide range of bioactivities including antioxidant, antiapoptotic, and antifibrotic properties with concomitant downregulation of Toll-like receptors, NF-κB, inflammatory cytokines and chemokines, and bradykinin. Curcumin has been implicated in blocking cellular entry and rep-lication of SARS-CoV-2 and in the repair of pneumocytes, renal cells, cardiomyocytes, and hematopoietic stem cells in the wake of COVID-19 (Soni, Mehta et al. 2020).
Essential oils are also an important class of compounds present in spices with high contents of terpenes, monoterpenes, and ses-quiterpenes; compounds that mediate several important physiolog-ical functions in our body. The antiviral effect of essential oils, has been reported by many. Eugenol, the principal component of clove bud (Eugenia caryophyllus) was found to possess strong antiviral activity against the types 1 and 2 herpes simplex viruses (HSV-1 and HSV-2) (Benencia and Courreges, 2000). Several nutraceuti-cal formulations based on beneficial spices have also been report-ed to augment health and well-being. For example, consumption of a milk based drink containing turmeric powder (Golden milk) once or twice a day has been claimed to act as an immune booster, largely by virtue of the active component, curcumin. Curcumin is

Journal of Food Bioactives | www.isnff-jfb.com 69
Banik et al. Functional foods beyond nutrition
responsible for suppression of anti-inflammatory cytokines and therefore has the potential to arrest a cytokine storm during a coro-naviral infection. (Sordillo and Helson, 2015).
4.14. Molecular basis of the immune augmenting role of spices
The active ingredients of spices exert their effects by acting as potent anti-oxidants. Their action is mediated mostly by direct or indirect activation of the transcription factor Nuclear factor-erythroid factor 2-related factor 2 (Bousquet et al., 2020). Nrf2 is the most potent antioxidant in humans and can minimize oxidative stress arising from both the Renin-Angiotensin II axis as well from the unfolded protein response of endoplasmic reticulum (Bousquet et al., 2020). Upregulation of Nrf2 signalling curbs the secretion of proinflammatory cytokines like IL-6 and also chemokines, and simultaneously downregulates the activation of nuclear factor-kap-pa b (NFĸB). Natural compounds derived from plants, vegetables, fungi, and micronutrients, or physical exercise can activate Nrf2 (Jiménez-Osorio et al., 2015). The different mortality rates inflict-ed by COVID-19 in different regions of the world may be partly explained by different culinary habits with differential contents of Nrf2 in the spices used for cooking.
Apart from Nrf-2, another important player in this molecular cross-talk is transient receptor potential (TRP). The TRP vanilloid 1 (TRPV1) and ankyrin 1 (TRPA1) belong to the TRP superfam-ily of structurally related, nonselective cation channels. They are often co-located in different cell types including sensory neurons, vascular smooth muscle cells, and many immune cells including monocytes and lymphocytes as well as in keratinocytes and epithe-lial cells, and are implicated in numerous physiological functions (Talavera et al., 2020). These cationic channels are highly sensitive to oxidative damage. Active principles of spices play a major role in preserving the functionality of these molecules and hence en-sure maintenance of physiological homeostasis, especially during intrusion by viral and bacterial pathogens.
4.15. Fatty acids as essential components of functional foods
The polyunsaturated fatty acids (PUFA) that cannot be synthesized by us, but yet are indispensable are termed ‘essential fatty acids’ (EFA). EFAs can be categorized into two classes, the omega-3 (ω-3) fatty acids, which have the last carbon–carbon double bond in the ω-3 position i.e. the third bond from the methyl end of the fatty acid. Omega-6 (ω-6) fatty acids possess a double bond in the ω-6 position i.e. the sixth bond from the methyl end. α-Linolenic acid (ALA), eicosapentaenoic acid (EPA), and docosahexaenoic acid (DHA) are the three chief omega-3 (ω-3) fatty acids that have been given the designation of functional foods for regulating many physiological processes.Among the omega-6 fatty acids, Linoleic acid,gamma-linolenic acid, dihomo-gamma-linolenic acid, and arachidonic acid are especially worth mentioning with regards to their various functionalities inside the body.
Alpha-linolenic acid is the most readily available ω-3 fatty acid in nature. Its most accessible source are the green leafy vegeta-bles. In addition, alpha-linolenic acid is also found in abundance in flax seed (Hunter, 1990). Fish and fish oil are the most abundant sources of EPA whereas DHA is obtained mostly from fish oil and red brown algae. Safflower, sunflower, and corn are rich sources of linoleic acid. Gamma-linolenic acid is obtained from evening primrose oil, black currant seed oil, and borage oil. Dihomo-gam-ma-linolenic acid is synthesized from gamma-linolenic acid, and arachidonic acid is present chiefly in meat and dairy sources.
4.16. Physiological importance of EFAs and their antiviral roles
Most of the EFAs are incorporated into the phospholipid bilayer and therefore are responsible for the structural integrity of the cell. ω-3 enriched diet is implicated in prevention of cardiovascular dis-eases (Das, 2000). In fact, the ratio of ω-3 to ω-6 fatty acids plays a significant role in maintaining normal physiological conditions and minimizing development of inflammatory response. Dietary LA is implicated in inflammatory response. However, in the body, LA is desaturated to GLA which is anti-inflammatory. Very recent-ly, EFAs were also demonstrated to inhibit coronaviral pathogen-esis by blocking the binding of viral spike protein to ACE2 recep-tor. Linolenic acid, eicosapentaenoic acid (EPA), and linoleic acid had the highest binding affinity to spikeprotein and hence arrested COVID-19 infection with highest efficiency (Goc et al., 2021).
4.17. The herbal nutraceuticals
The term nutraceuticals is an amalgamation of the two words “nutri-tion” and “pharmaceutical” originally coined in the year 1989 by Stephen Defelice, founder and chairman of the Foundation for In-novation in Medicine (FIM) Cranford, New Jersey (Brower, 1998). Since then, the quest for the extraordinary potential of herbs has flourished and we have come to know about the therapeutic poten-tials of traditional herbs consumed both as medicines and as culinary delights. Not surprisingly, many such herbs have shown promise to act as effective remedies for both prevention and cure from viral infections, including those caused by SARS-CoV2. The active com-ponent present in many of these herbs have been identified and their antiviral potential have been well demonstrated (Table 1). An early study had indicated the anti-coronaviral activity of concanavalin A (conA), a phytagglutinin found in jack beans (Canavalia ensi-formis). Since then, many oriental herbs have been discovered with strong antiviral activity. Some of these are Lycorisradiata, Artemisia annua, Pyrrosia lingua, Linderaaggregataetc. which inhibited the viral replication by more than 50%. Houttuynia cordata, an edible herb popularly known as fish mint or rainbow plant, arrested SARS-CoV2 replication by blocking the action of two key viral proteins, chymotrypsin-like protease (3CLpro) and RdRp (Abiri et al., 2021). It also led to increased levels of CD4+ and CD8+ T cells to strength-en the immune system for evading the viral attack (Figure 2).
Herbs of Indian origin belonging to the traditional Ayurveda has been protecting us a plethora of ailments and infections since ages. The treasure trove of Ayurveda has plants from the family polygonaceae inhibiting viral entry into the host (Schwarz et al., 2011), edible herbs like Glycyrrhiza glabra inhibiting viral rep-lication (Cinatl et al., 2003), and plants like Sambucus ebulus arresting viral envelope formation (Choudhary et al., 2020). The AYUSH system maintains a database of these medicinal plants for preserving a systematic knowledge of the diverse repertoire of Indian medicinal plants for effective lifestyle management during the COVID-19 pandemic through lifestyle modification, dietary management as well as prophylactic interventions.
5. Concluding remarks: the future of functional food and nutra-ceuticals as effective antivirals as compared to traditional medi-cines
Restoration of physiological homeostasis by functional foods and nutraceuticals is mostly achieved via their anti-inflammatory and antioxidative potential. This to an extent also involves downregu-
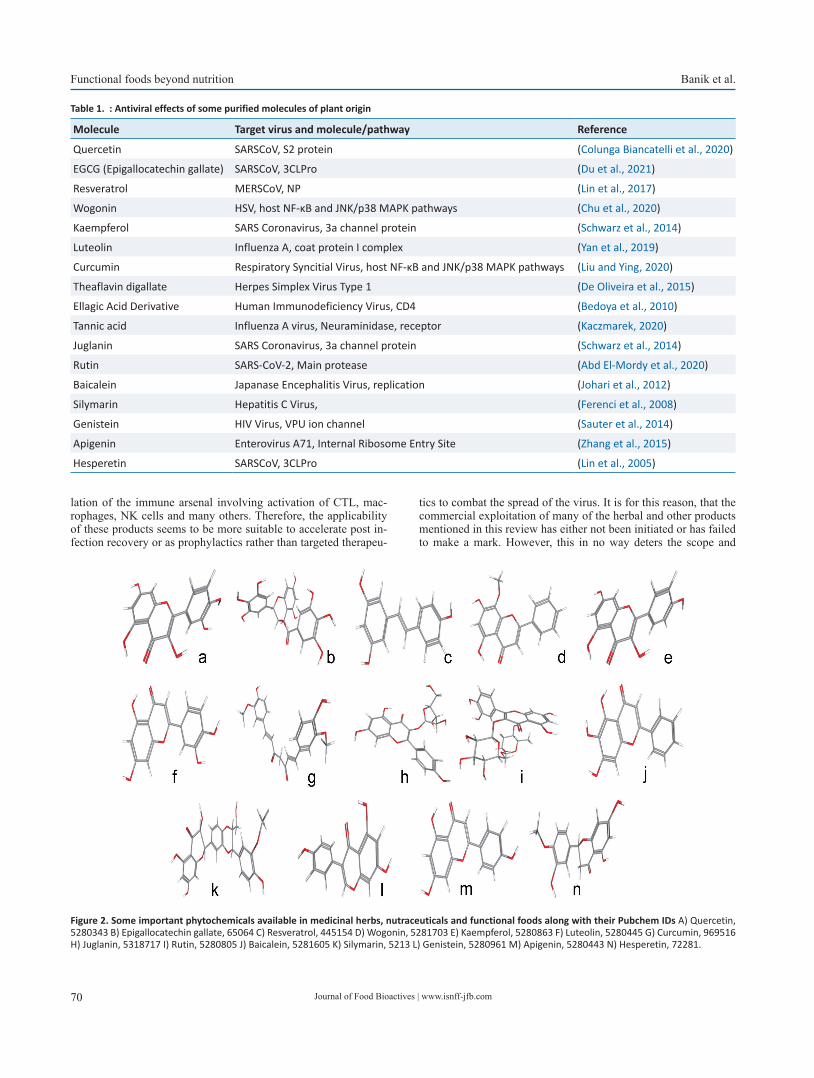
Journal of Food Bioactives | www.isnff-jfb.com70
Functional foods beyond nutrition Banik et al.
lation of the immune arsenal involving activation of CTL, mac-rophages, NK cells and many others. Therefore, the applicability of these products seems to be more suitable to accelerate post in-fection recovery or as prophylactics rather than targeted therapeu-
tics to combat the spread of the virus. It is for this reason, that the commercial exploitation of many of the herbal and other products mentioned in this review has either not been initiated or has failed to make a mark. However, this in no way deters the scope and
Figure 2. Some important phytochemicals available in medicinal herbs, nutraceuticals and functional foods along with their Pubchem IDs A) Quercetin, 5280343 B) Epigallocatechin gallate, 65064 C) Resveratrol, 445154 D) Wogonin, 5281703 E) Kaempferol, 5280863 F) Luteolin, 5280445 G) Curcumin, 969516 H) Juglanin, 5318717 I) Rutin, 5280805 J) Baicalein, 5281605 K) Silymarin, 5213 L) Genistein, 5280961 M) Apigenin, 5280443 N) Hesperetin, 72281.
Table 1. : Antiviral effects of some purified molecules of plant origin
Molecule Target virus and molecule/pathway Reference
Quercetin SARSCoV, S2 protein (Colunga Biancatelli et al., 2020)
EGCG (Epigallocatechin gallate) SARSCoV, 3CLPro (Du et al., 2021)
Resveratrol MERSCoV, NP (Lin et al., 2017)
Wogonin HSV, host NF-κB and JNK/p38 MAPK pathways (Chu et al., 2020)
Kaempferol SARS Coronavirus, 3a channel protein (Schwarz et al., 2014)
Luteolin Influenza A, coat protein I complex (Yan et al., 2019)
Curcumin Respiratory Syncitial Virus, host NF-κB and JNK/p38 MAPK pathways (Liu and Ying, 2020)
Theaflavin digallate Herpes Simplex Virus Type 1 (De Oliveira et al., 2015)
Ellagic Acid Derivative Human Immunodeficiency Virus, CD4 (Bedoya et al., 2010)
Tannic acid Influenza A virus, Neuraminidase, receptor (Kaczmarek, 2020)
Juglanin SARS Coronavirus, 3a channel protein (Schwarz et al., 2014)
Rutin SARS-CoV-2, Main protease (Abd El-Mordy et al., 2020)
Baicalein Japanase Encephalitis Virus, replication (Johari et al., 2012)
Silymarin Hepatitis C Virus, (Ferenci et al., 2008)
Genistein HIV Virus, VPU ion channel (Sauter et al., 2014)
Apigenin Enterovirus A71, Internal Ribosome Entry Site (Zhang et al., 2015)
Hesperetin SARSCoV, 3CLPro (Lin et al., 2005)

Journal of Food Bioactives | www.isnff-jfb.com 71
Banik et al. Functional foods beyond nutrition
applicability of functional foods since their future is bright and ever-expanding with so many unexplored components. Addition-ally, these compounds have proven to be much safer as compared to the corresponding synthetic molecules making them especially more popular for paediatric use. Therefore, if a dual impetus of funding and systematic research is put into this field, the future of functional foods and nutraceuticals will shine brighter than syn-thetic drug research.
References
Abd El-Mordy, F.M., El-Hamouly, M.M., Ibrahim, M.T., Abd El-Rheem, G., Aly, O.M., Abd El-Kader, A.M., Youssif, K.A., and Abdelmohsen, U.R. (2020). Inhibition of SARS-CoV-2 main protease by phenolic com-pounds from Manilkara hexandra (Roxb.) Dubard assisted by metabo-lite profiling and in silico virtual screening. RSC Adv. 10: 32148–3215.
Abiri, R., Abdul-Hamid, H., Sytar, O., Abiri, R., Bezerra De Almeida, E., Shar-ma, S.K., Bulgakov, V.P., Arroo, R.R., and Malik, S. (2021). A Brief Over-view of Potential Treatments for Viral Diseases Using Natural Plant Compounds: The Case of SARS-Cov. Molecules 26: 3868.
Ahmed, S.F., Quadeer, A.A., and Mckay, M.R. (2020). Preliminary identi-fication of potential vaccine targets for the COVID-19 coronavirus (SARS-CoV-2) based on SARS-CoV immunological studies. Viruses 12: 254.
Annunziata, G., Jiménez-García, M., Capó, X., Moranta, D., Arnone, A., Tenore, G., Sureda, A., and Tejada, S. (2020). Microencapsulation as a tool to counteract the typical low bioavailability of polyphenols in the management of diabetes. Food Chem. Toxicol. 139: 111248.
Aruoma, O., Spencer, J., Rossi, R., Aeschbach, R., Khan, A., Mahmood, N., Munoz, A., Murcia, A., Butler, J., and Halliwell, B. (1996). An evalua-tion of the antioxidant and antiviral action of extracts of rosemary and Provencal herbs. Food Chem. Toxicol. 34: 449–456.
Bach, J.-F., and Dardenne, M. (1989). Thymulin, a zinc-dependent hor-mone. Med. Oncol. Tumor. Pharmacother. 6: 25–29.
Beard, J.A., Bearden, A., and Striker, R. (2011). Vitamin D and the anti-viral state. J. Clin. Virol. 50: 194–200.
Bedoya, L., Abad, M., Sánchez-Palomino, S., Alcami, J., and Bermejo, P. (2010). Ellagitannins from Tuberaria lignosa as entry inhibitors of HIV. Phytomedicine 17: 69–74.
Bedoya, L., Palomino, S.S., Abad, M., Bermejo, P., and Alcami, J. (2002). Screening of selected plant extracts for in vitro inhibitory activity on human immunodeficiency virus. Phytother. Res. 16: 550–554.
Benencia, F., and Courreges, M. (2000). In vitro and in vivo activity of eugenol on human herpesvirus. Phytother. Res. 14: 495–500.
Bertelli, C., Pillonel, T., Torregrossa, A., Prod’hom, G., Fischer, C.J., Greub, G., and Giannoni, E. (2015). Bifidobacterium longum bacteremia in preterm infants receiving probiotics. Clin. Infect. Dis. 60: 924–927.
Bikle, D.D. (2014). Vitamin D metabolism, mechanism of action, and clini-cal applications. Chem. Biol. 21: 319–329.
Bikle, D.D., Ng, D., Oda, Y., Hanley, K., Feingold, K., and Xie, Z. (2002). The vitamin D response element of the involucrin gene mediates its regulation by 1, 25-dihydroxyvitamin D3. J. Invest. Dermatol. 119: 1109–1113.
Bousquet, J., Cristol, J.P., Czarlewski, W., Anto, J.M., Martineau, A., Haahte-la, T., Fonseca, S.C., Iaccarino, G., Blain, H., and Fiocchi, A. (2020). Nrf2-interacting nutrients and COVID-19: time for research to de-velop adaptation strategies. Clin. Transl. Allergy 10: 58.
Brower, V. (1998). Nutraceuticals: poised for a healthy slice of the health-care market? Nat. Biotechnol. 16: 728–731.
Busso, N., Wagtmann, N., Herling, C., Chobaz-Péclat, V., Bischof-Delaloye, A., So, A., and Grouzmann, E. (2005). Circulating CD26 is negatively associated with inflammation in human and experimental arthritis. Am. J. Pathol. 166: 433–442.
Butterworth, B.E., and Korant, B.D. (1974). Characterization of the large picornaviral polypeptides produced in the presence of zinc ion. J Vi-rol. 14: 282–291.
Carabotti, M., Scirocco, A., Maselli, M.A., and Severi, C. (2015). The gut-brain axis: interactions between enteric microbiota, central and en-
teric nervous systems. Ann. Gastroenterol. 28: 203.Carlson, A.L., Xia, K., Azcarate-Peril, M.A., Goldman, B.D., Ahn, M., Styner,
M.A., Thompson, A.L., Geng, X., Gilmore, J.H., and Knickmeyer, R.C. (2018). Infant gut microbiome associated with cognitive develop-ment. Biol. Psychiatry 83: 148–159.
Cebula, A., Seweryn, M., Rempala, G.A., Pabla, S.S., Mcindoe, R.A., Den-ning, T.L., Bry, L., Kraj, P., Kisielow, P., and Ignatowicz, L. (2013). Thy-mus-derived regulatory T cells contribute to tolerance to commensal microbiota. Nature 497: 258–262.
Chandra, R.K. (1996). Nutrition, immunity and infection: from basic knowl-edge of dietary manipulation of immune responses to practical ap-plication of ameliorating suffering and improving survival. Proc. Natl. Acad. Sci. USA 93: 14304–14307.
Chen, C., Zuckerman, D.M., Brantley, S., Sharpe, M., Childress, K., Hoiczyk, E., and Pendleton, A.R. (2014). Sambucus nigra extracts inhibit infec-tious bronchitis virus at an early point during replication. BMC Vet. Res. 10: 1–12.
Choudhary, N., Lodha, M., and Baranwal, V. (2020). The role of enzymatic activities of antiviral proteins from plants for action against plant pathogens. 3 Biotech 10: 505.
Christakos, S., Dhawan, P., Verstuyf, A., Verlinden, L., and Carmeliet, G. (2016). Vitamin D: metabolism, molecular mechanism of action, and pleiotropic effects. Physiol. Rev. 96: 365–408.
Chu, Y., Lv, X., Zhang, L., Fu, X., Song, S., Su, A., Chen, D., Xu, L., Wang, Y., and Wu, Z. (2020). Wogonin inhibits in vitro herpes simplex virus type 1 and 2 infection by modulating cellular NF-κB and MAPK path-ways. BMC Microbiol. 20: 1–11.
Cinatl, J., Morgenstern, B., Bauer, G., Chandra, P., Rabenau, H., and Doerr, H. (2003). Glycyrrhizin, an active component of liquorice roots, and replication of SARS-associated coronavirus. Lancet 361: 2045–2046.
Colunga Biancatelli, R.M.L., Berrill, M., Catravas, J.D., and Marik, P.E. (2020). Quercetin and vitamin C: an experimental, synergistic ther-apy for the prevention and treatment of SARS-CoV-2 related disease (COVID-19). Front. Immunol. 11: 1451.
Connolly, M., Tuohy, K., and Lovegrove, J. (2012). Wholegrain oat-based cereals have prebiotic potential and low glycaemic index. Br. J. Nutr. 108: 2198–2206.
Critchfield, J.W., Butera, S.T., and Folks, T.M. (1996). Inhibition of HIV ac-tivation in latently infected cells by flavonoid compounds. AIDS Res. Hum. Retroviruses 12: 39–46.
Cui, J., Li, F., and Shi, Z.-L. (2019). Origin and evolution of pathogenic coro-naviruses. Nat. Rev. Microbiol. 17: 181–192.
Cummings, J.H., Macfarlane, G.T., and Englyst, H.N. (2001). Prebiotic diges-tion and fermentation. Am. J. Clin. Nutr. 73: 415s–420s.
Dang, A.T., and Marsland, B.J. (2019). Microbes, metabolites, and the gut–lung axis. Mucosal Immunol. 12: 843–850.
Das, U.N. (2000). Beneficial effect (s) of n-3 fatty acids in cardiovascular diseases: but, why and how? Prostaglandins Leukot. Essent. Fatty Acids 63: 351–362.
De Oliveira, A., Prince, D., Lo, C.-Y., Lee, L.H., and Chu, T.-C. (015). Antiviral activity of theaflavin digallate against herpes simplex virus type 1. Antiviral Res. 118: 56–67.
Delesderrier, E., Cople-Rodrigues, C.S., Omena, J., Kneip Fleury, M., Bar-bosa Brito, F., Costa Bacelo, A., Correa Koury, J., and Citelli, M. (2019). Selenium status and hemolysis in sickle cell disease patients. Nutri-ents 11: 2211.
Dermyshi, E., Wang, Y., Yan, C., Hong, W., Qiu, G., Gong, X., and Zhang, T. (2017). The “golden age” of probiotics: a systematic review and meta-analysis of randomized and observational studies in preterm infants. Neonatology 112: 9–23.
Diebold, S.S., Kaisho, T., Hemmi, H., Akira, S., and E Sousa, C.R. (2004). In-nate antiviral responses by means of TLR7-mediated recognition of single-stranded RNA. Science 303: 1529–1531.
Du, A., Zheng, R., Disoma, C., Li, S., Chen, Z., Li, S., Liu, P., Zhou, Y., Shen, Y., and Liu, S. (2021). Epigallocatechin-3-gallate, an active ingredient of Traditional Chinese Medicines, inhibits the 3CLpro activity of SARS-CoV-2. Int. J. Biol. Macromol. 176: 1–12.
Faith, S.A., Sweet, T.J., Bailey, E., Booth, T., and Docherty, J.J. (006). Resver-atrol suppresses nuclear factor-κB in herpes simplex virus infected cells. Antiviral Res. 72: 242–251.
Fassina, G., Buffa, A., Benelli, R., Varnier, O.E., Noonan, D.M., and Albini, A.

Journal of Food Bioactives | www.isnff-jfb.com72
Functional foods beyond nutrition Banik et al.
(2002). Polyphenolic antioxidant (–)-epigallocatechin-3-gallate from green tea as a candidate anti-HIV agent. Aids 16: 939–941.
Ferenci, P., Scherzer, T.M., Kerschner, H., Rutter, K., Beinhardt, S., Hofer, H., Schöniger–Hekele, M., Holzmann, H., and Steindl–Munda, P. (2008). Silibinin is a potent antiviral agent in patients with chronic hepatitis C not responding to pegylated interferon/ribavirin therapy. Gastroen-terology 135: 1561–1567.
Ferrari, E., Wright-Minogue, J., Fang, J.W., Baroudy, B.M., Lau, J.Y., and Hong, Z. (1999). Characterization of soluble hepatitis C virus RNA-dependent RNA polymerase expressed in Escherichia coli. J Virol. 73: 1649–1654.
Fogarty, H., Townsend, L., Cheallaigh, C.N., Bergin, C., Martin-Loeches, I., Browne, P., Bacon, C.L., Gaule, R., Gillett, A., Byrne, M., Ryan, K., O’Connell, N., O'Sullivan, J.M., Conlon, N., and O'Donnell, J.S. (2020). COVID-19 coagulopathy in caucasian patients. Br. J. Haematol. 189: 1044–1049.
Frei, R., Akdis, M., and O’mahony, L. (2015). Prebiotics, probiotics, synbiot-ics, and the immune system: experimental data and clinical evidence. Curr. Opin. Gastroenterol. 31: 153–158.
Fridlender, B., Chejanovsky, N., and Becker, Y. (1978). Selective inhibition of herpes simplex virus type 1 DNA polymerase by zinc ions. Virology 84: 551–554.
García-Casal, M.N., Peña-Rosas, J.P., and Malavé, H.G. (2016). Sauces, spices, and condiments: definitions, potential benefits, consumption patterns, and global markets. Ann. N. Y. Acad. Sci. 1379: 3–16.
Goc, A., Niedzwiecki, A., and Rath, M. (2021). Polyunsaturated ω-3 fatty acids inhibit ACE2-controlled SARS-CoV-2 binding and cellular entry. Sci. Rep. 11: 1–12.
Gordon, J.I., Dewey, K.G., Mills, D.A., and Medzhitov, R.M. (2012). The human gut microbiota and undernutrition. Sci. Transl. Med. 4: 137ps12–137ps12.
Guarino, M.P.L., Altomare, A., Emerenziani, S., Di Rosa, C., Ribolsi, M., Bal-estrieri, P., Iovino, P., Rocchi, G., and Cicala, M. (2020). Mechanisms of action of prebiotics and their effects on gastro-intestinal disorders in adults. Nutrients 12: 1037.
Hao, Q., Dong, B.R., and Wu, T. (2015). Probiotics for preventing acute upper respiratory tract infections. Cochrane Database Syst. Rev. (1): CD006895.
Holick, M.F., Maclaughlin, J., Clark, M., Holick, S., Potts, J., Anderson, R., Blank, I., Parrish, J., and Elias, P. (1980). Photosynthesis of previtamin D3 in human skin and the physiologic consequences. Science 210: 203–205.
Hunter, J.E. (1990). n-3 fatty acids from vegetable oils. Am. J. Clin. Nutr. 51: 809–814.
Ibs, K.-H., and Rink, L. (2003). Zinc-altered immune function. J. Nutr. 133: 1452S–1456S.
Imanishi, N., Tuji, Y., Katada, Y., Maruhashi, M., Konosu, S., Mantani, N., Terasawa, K., and Ochiai, H. (2002). Additional inhibitory effect of tea extract on the growth of influenza A and B viruses in MDCK cells. Microbiol Immunol. 46: 491–494.
Jayawardena, R., Sooriyaarachchi, P., Chourdakis, M., Jeewandara, C., and Ranasinghe, P. (2020). Enhancing immunity in viral infections, with special emphasis on COVID-19: A review. Diabetes Metab. Syndr. 14: 367–382.
Jiménez-Osorio, A.S., Gonzalez-Reyes, S., and Pedraza-Chaverri, J. (2015). Natural Nrf2 activators in diabetes. Clin. Chim. Acta 448: 182–192.
Joe, B., Vijaykumar, M., and Lokesh, B. (2004). Biological properties of cur-cumin-cellular and molecular mechanisms of action. Crit. Rev. Food Sci. Nutr. 44: 97–111.
Johari, J., Kianmehr, A., Mustafa, M.R., Abubakar, S., and Zandi, K. (2012). Antiviral activity of baicalein and quercetin against the Japanese en-cephalitis virus. Int. J. Mol. Sci. 13: 16785–16795.
Kaczmarek, B. (2020). Tannic acid with antiviral and antibacterial activity as a promising component of biomaterials—A minireview. Materials 13: 3224.
Kanauchi, O., Andoh, A., Abubakar, S., and Yamamoto, N. (2018). Probiot-ics and paraprobiotics in viral infection: clinical application and ef-fects on the innate and acquired immune systems. Curr. Pharm. Des. 24: 710–717.
Kieliszek, M., and Lipinski, B. (2020). Selenium supplementation in the prevention of coronavirus infections (COVID-19). Med. Hypotheses.
143: 109878.Kiremidjian-Schumacher, L., and Roy, M. (1998). Selenium and immune
function. Zeitschrift fur Ernahrungswissenschaft 37: 50–56.Lange, K.W. (2021). Food science and COVID-19. Food Sci. Hum. Wellness
10: 1–5.Lazar, V., Ditu, L.-M., Pircalabioru, G.G., Gheorghe, I., Curutiu, C., Holban,
A.M., Picu, A., Petcu, L., and Chifiriuc, M.C. (2018). Aspects of gut microbiota and immune system interactions in infectious diseases, immunopathology, and cancer. Front. Immunol. 9: 1830.
Li, W., Moore, M.J., Vasilieva, N., Sui, J., Wong, S.K., Berne, M.A., So-masundaran, M., Sullivan, J.L., Luzuriaga, K., and Greenough, T.C. (2003). Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 426: 450–454.
Li, Y., Dong, J., Xiao, H., Zhang, S., Wang, B., Cui, M., and Fan, S. (2020). Gut commensal derived-valeric acid protects against radiation injuries. Gut Microbes 11: 789–806.
Lin, C.-J., Lin, H.-J., Chen, T.-H., Hsu, Y.-A., Liu, C.-S., Hwang, G.-Y., and Wan, L. (2015). Polygonum cuspidatum and its active components inhibit replication of the influenza virus through toll-like receptor 9-induced interferon beta expression. PLoS One 10: e0117602.
Lin, C.-W., Tsai, F.-J., Tsai, C.-H., Lai, C.-C., Wan, L., Ho, T.-Y., Hsieh, C.-C., and Chao, P.-D.L. (2005). Anti-SARS coronavirus 3C-like protease ef-fects of Isatis indigotica root and plant-derived phenolic compounds. Antiviral Res. 68: 36–42.
Lin, S.-C., Ho, C.-T., Chuo, W.-H., Li, S., Wang, T.T., and Lin, C.-C. (2017). Ef-fective inhibition of MERS-CoV infection by resveratrol. BMC Infect. Dis. 17: 1–10.
Liu, Z., and Ying, Y. (2020). The inhibitory effect of curcumin on virus-in-duced cytokine storm and its potential use in the associated severe pneumonia. Front. Cell Dev. Biol. 8: 479.
López-Varela, S., González-Gross, M., and Marcos, A. (2002). Functional foods and the immune system: a review. Eur. J. Clin. Nutr. 56: S29–S33.
Loser, K., and Beissert, S. (2009). Regulation of cutaneous immunity by the environment: an important role for UV irradiation and vitamin D. Int. Immunopharmacol. 9: 587–589.
Martineau, A.R., Jolliffe, D.A., Hooper, R.L., Greenberg, L., Aloia, J.F., Berg-man, P., Dubnov-Raz, G., Esposito, S., Ganmaa, D., and Ginde, A.A. (2017). Vitamin D supplementation to prevent acute respiratory tract infections: systematic review and meta-analysis of individual partici-pant data. BMJ 356: i6583.
Martirosyan, D.M., and Singh, J. (2015). A new definition of functional food by FFC: what makes a new definition unique? Funct. Foods Health Dis. 5: 209–223.
Meng, X., Zhou, J., Zhao, C.-N., Gan, R.-Y., and Li, H.-B. (2020). Health ben-efits and molecular mechanisms of resveratrol: A narrative review. Foods 9: 340.
Messina, G., Polito, R., Monda, V., Cipolloni, L., Di Nunno, N., Di Mizio, G., Murabito, P., Carotenuto, M., Messina, A., and Pisanelli, D. (2020). Functional role of dietary intervention to improve the outcome of COVID-19: a hypothesis of work. Int. J. Mol. Sci. 21: 3104.
Mirzaei, R., Attar, A., Papizadeh, S., Jeda, A.S., Hosseini-Fard, S.R., Jamasbi, E., Kazemi, S., Amerkani, S., Talei, G.R., and Moradi, P. (2021). The emerging role of probiotics as a mitigation strategy against coronavi-rus disease 2019 (COVID-19). Arch. Virol. 166: 1819–1840.
Nagpal, R., Shively, C.A., Register, T.C., Craft, S., and Yadav, H. (2019). Gut microbiome-Mediterranean diet interactions in improving host health. F1000Research 8: 699.
Nakayama, M., Suzuki, K., Toda, M., Okubo, S., Hara, Y., and Shimamura, T. (1993). Inhibition of the infectivity of influenza virus by tea polyphe-nols. Antiviral Res. 21: 289–299.
Nicoletti, M. (2012). Nutraceuticals and botanicals: overview and perspec-tives. Int. J. Food Sci. Nutr. 63: 2–6.
Opara, E.C. (2002). Oxidative stress, micronutrients, diabetes mellitus and its complications. J. R. Soc. Promot. Health. 122: 28–34.
Palamara, A.T., Nencioni, L., Aquilano, K., De Chiara, G., Hernandez, L., Coz-zolino, F., Ciriolo, M.R., and Garaci, E. (2005). Inhibition of influenza A virus replication by resveratrol. J. Infect. Dis. 191: 1719–1729.
Pannu, N., and Bhatnagar, A. (2019). Resveratrol: From enhanced biosyn-thesis and bioavailability to multitargeting chronic diseases. Biomed. Pharmacother. 109: 2237–2251.

Journal of Food Bioactives | www.isnff-jfb.com 73
Banik et al. Functional foods beyond nutrition
Pavan, R., Jain, S., and Kumar, A. (2012). Properties and therapeutic ap-plication of bromelain: a review. Biotechnol. Res. Int. 2012: 976203.
Pimentel-Nunes, P., Soares, J.B., Roncon-Albuquerque, R., Dinis-Ribeiro, M., and Leite-Moreira, A.F. (2010). Toll-like receptors as therapeu-tic targets in gastrointestinal diseases. Expert Opin. Ther. Targets 14: 347–368.
Rahman, M.T., and Idid, S.Z. (2021). Can Zn be a critical element in COV-ID-19 treatment? Biol. Trace Elem. Res. 199: 550–558.
Rayman, M.P. (2000). The importance of selenium to human health. Lan-cet 356: 233–241.
Reygaert, W.C. (2018). Green tea catechins: Their use in treating and pre-venting infectious diseases. BioMed Res. Int. 2018: 9105261.
Ru-Duan, W., Chang-Sen, W., Zuo-Hua, F., and Yi, L. (1992). Investigation on the effect of selenium on T lymphocyte proliferation and its mecha-nisms. J. Tongji Med. Univ. 12: 33–38.
Sargiacomo, C., Sotgia, F., and Lisanti, M.P. (2020). COVID-19 and chrono-logical aging: senolytics and other anti-aging drugs for the treatment or prevention of corona virus infection? Aging (Albany NY) 12: 6511.
Sauter, D., Schwarz, S., Wang, K., Zhang, R., Sun, B., and Schwarz, W. (2014). Genistein as antiviral drug against HIV ion channel. Planta Med. 80: 682–687.
Scholz-Ahrens, K.E., Ade, P., Marten, B., Weber, P., Timm, W., Aςil, Y., Gluer, C.-C., and Schrezenmeir, J.R. (2007). Prebiotics, probiotics, and syn-biotics affect mineral absorption, bone mineral content, and bone structure. J. Nutr. 137: 838S–846S.
Schuetz, P., Fehr, R., Baechli, V., Geiser, M., Deiss, M., Gomes, F., Kutz, A., Tribolet, P., Bregenzer, T., and Braun, N. (2019). Individualised nutri-tional support in medical inpatients at nutritional risk: a randomised clinical trial. Lancet 393: 2312–2321.
Schwarz, S., Sauter, D., Wang, K., Zhang, R., Sun, B., Karioti, A., Bilia, A.R., Efferth, T., and Schwarz, W. (2014). Kaempferol derivatives as antivi-ral drugs against the 3a channel protein of coronavirus. Planta Med. 80: 177–182.
Schwarz, S., Wang, K., Yu, W., Sun, B., and Schwarz, W. (2011). Emodin inhibits current through SARS-associated coronavirus 3a protein. An-tiviral Res. 90: 64–69.
Shaito, A., Posadino, A.M., Younes, N., Hasan, H., Halabi, S., Alhababi, D., Al-Mohannadi, A., Abdel-Rahman, W.M., Eid, A.H., and Nasrallah, G.K. (2020). Potential adverse effects of resveratrol: A literature re-view. Int. J. Mol. Sci. 21: 2084.
Shang, J., Wan, Y., Liu, C., Yount, B., Gully, K., Yang, Y., Auerbach, A., Peng, G., Baric, R., and Li, F. (2020). Structure of mouse coronavirus spike protein complexed with receptor reveals mechanism for viral entry. PLoS Pathog. 16: e1008392.
Sharma, R.K., Goswami, B., Mandal, S.D., Guha, A., Willard, B., and Ray, P.S. (2021). Quorum Sensing by Gelsolin Regulates Programmed Cell Death 4 Expression and a Density-Dependent Phenotype in Mac-rophages. J. Immunol. 207: 1250–1264.
Sindona, C., Schepici, G., Contestabile, V., Bramanti, P., and Mazzon, E. (2021). NOX2 Activation in COVID-19: Possible Implications for Neu-rodegenerative Diseases. Medicina (Kaunas) 57: 604.
Somerville, V.S., Braakhuis, A.J., and Hopkins, W.G. (2016). Effect of flavo-noids on upper respiratory tract infections and immune function: a systematic review and meta-analysis. Adv. Nutr. 7: 488–497.
Song, J.-M., Lee, K.-H., and Seong, B.-L. (2005). Antiviral effect of catechins in green tea on influenza virus. Antiviral Res. 68: 66–74.
Soni, V.K., Mehta, A., Ratre, Y.K., Tiwari, A.K., Amit, A., Singh, R.P., Sonkar, S.C., Chaturvedi, N., Shukla, D., and Vishvakarma, N.K. (2020). Cur-cumin, a traditional spice component, can hold the promise against COVID-19? Eur. J. Pharmacol. 886: 173551.
Sordillo, P.P., and Helson, L. (2015). Curcumin suppression of cytokine re-lease and cytokine storm. A potential therapy for patients with Ebola and other severe viral infections. In Vivo 29: 1–4.
Subramanian, V.K., and Deepe, G.S. (2017). Metallothioneins: emerging modulators in immunity and infection. Int. J. Mol. Sci. 18: 2197.
Szczepanski, A., Owczarek, K., Bzowska, M., Gula, K., Drebot, I., Ochman, M., Maksym, B., Rajfur, Z., Mitchell, J.A., and Pyrc, K. (2019). Canine respiratory coronavirus, bovine coronavirus, and human coronavirus OC43: receptors and attachment factors. Viruses 11: 328.
Talavera, K., Startek, J.B., Alvarez-Collazo, J., Boonen, B., Alpizar, Y.A.,
Sanchez, A., Naert, R., and Nilius, B. (2020). Mammalian transient re-ceptor potential TRPA1 channels: from structure to disease. Physiol. Rev. 100: 725–803.
Tang, N.L.-S., Chan, P.K.-S., Wong, C.-K., To, K.-F., Wu, A.K.-L., Sung, Y.-M., Hui, D.S.-C., Sung, J.J.-Y., and Lam, C.W.-K. (2005). Early enhanced expression of interferon-inducible protein-10 (CXCL-10) and other chemokines predicts adverse outcome in severe acute respiratory syndrome. Clin. Chem. 51: 2333–2340.
Tapiovaara, L., Pitkäranta, A.I., and Korpela, R. (2016). Probiotics and up-per respiratory tract - a review. Pediatr. Infect. Dis. 1: 19.
Te Velthuis, A.J., Van Den Worm, S.H., Sims, A.C., Baric, R.S., Snijder, E.J., and Van Hemert, M.J. (2010). Zn2+ inhibits coronavirus and arterivi-rus RNA polymerase activity in vitro and zinc ionophores block the replication of these viruses in cell culture. PLoS Pathog. 6: e1001176.
Tikellis, C., and Thomas, M. (2012). Angiotensin-converting enzyme 2 (ACE2) is a key modulator of the renin angiotensin system in health and disease. Int. J. Pept. 2012: 256294.
Trichopoulou, A., Costacou, T., Bamia, C., and Trichopoulos, D. (2003). Ad-herence to a Mediterranean diet and survival in a Greek population. N. Engl. J. Med. 348: 2599–2608.
Valdés-Ramos, R., Martínez-Carrillo, B.E., Aranda-González, I.I., Guadar-rama, A.L., Pardo-Morales, R.V., Tlatempa, P., and Jarillo-Luna, R.A. (2010). Diet, exercise and gut mucosal immunity. Proc. Nutr. Soc. 69: 644–650.
Vankadari, N., and Wilce, J.A. (2020). Emerging COVID-19 coronavirus: gly-can shield and structure prediction of spike glycoprotein and its in-teraction with human CD26. Emerging Microbes Infect. 9: 601–604.
Wen, F., Yu, H., Guo, J., Li, Y., Luo, K., and Huang, S. (2020). Identification of the hyper-variable genomic hotspot for the novel coronavirus SARS-CoV-2. J. Infect. 80: 671.
Wilson, B., Rossi, M., Dimidi, E., and Whelan, K. (2019). Prebiotics in irrita-ble bowel syndrome and other functional bowel disorders in adults: a systematic review and meta-analysis of randomized controlled tri-als. Am. J. Clin. Nutr. 109: 1098–1111.
Wintergerst, E.S., Maggini, S., and Hornig, D.H. (2007). Contribution of se-lected vitamins and trace elements to immune function. Ann. Nutr. Metab. 51: 301–323.
Xu, K., Cai, H., Shen, Y., Ni, Q., Chen, Y., Hu, S., Li, J., Wang, H., Yu, L., and Huang, H. (2020). Management of COVID-19: the Zhejiang experi-ence. J. Zhejiang Univ. Sci. 49: 147–157.
Yahfoufi, N., Alsadi, N., Jambi, M., and Matar, C. (2018). The immunomod-ulatory and anti-inflammatory role of polyphenols. Nutrients 10: 1618.
Yan, H., Ma, L., Wang, H., Wu, S., Huang, H., Gu, Z., Jiang, J., and Li, Y. (2019). Luteolin decreases the yield of influenza A virus in vitro by interfering with the coat protein I complex expression. J. Nat. Med. 73: 487–496.
Yang, F., Zhang, Y., Tariq, A., Jiang, X., Ahmed, Z., Zhihao, Z., Idrees, M., Azizullah, A., Adnan, M., and Bussmann, R.W. (2020). Food as medi-cine: A possible preventive measure against coronavirus disease (COVID-19). Phytother. Res. 34: 3124–3136.
Yaqoob, P. (2017). Ageing alters the impact of nutrition on immune func-tion. Proc. Nutr. Soc. 76: 347–351.
Yi, H., Wang, L., Xiong, Y., Wang, Z., Qiu, Y., Wen, X., Jiang, Z., Yang, X., and Ma, X. (2018). Lactobacillus reuteri LR1 improved expression of genes of tight junction proteins via the MLCK pathway in IPEC-1 cells during infection with enterotoxigenic Escherichia coli K88. Mediators Inflamm. 2018: 6434910.
Yiu, C.-Y., Chen, S.-Y., Chang, L.-K., Chiu, Y.-F., and Lin, T.-P. (2010). Inhibitory effects of resveratrol on the Epstein-Barr virus lytic cycle. Molecules 15: 7115–7124.
Zang, N., Xie, X., Deng, Y., Wu, S., Wang, L., Peng, C., Li, S., Ni, K., Luo, Y., and Liu, E. (2011). Resveratrol-mediated gamma interferon reduc-tion prevents airway inflammation and airway hyperresponsiveness in respiratory syncytial virus-infected immunocompromised mice. J Virol. 85: 13061–13068.
Zhang, L., Li, Y., Gu, Z., Wang, Y., Shi, M., Ji, Y., Sun, J., Xu, X., Zhang, L., and Jiang, J. (2015). Resveratrol inhibits enterovirus 71 replication and pro-inflammatory cytokine secretion in rhabdosarcoma cells through blocking IKKs/NF-κB signaling pathway. PLoS One 10: e0116879.

Copyright: © 2021 International Society for Nutraceuticals and Functional Foods.All rights reserved.
74
Original Research J. Food Bioact. 2021;15:74–79
Journal ofFood Bioactives International Society for
Nutraceuticals and Functional Foods
Redistribution of nutritionally important starch fractions beneficial to diabetic population by using oats as a functional ingredient in a
wheat-based food product – Jereesh
Faiyaz Ahmed
Department of Clinical Nutrition, College of Applied Health Sciences in Ar Rass, Qassim University, Al Qassim Region 51921, Saudi Arabia. Tel: +966163010496; E-mail: [email protected]: 10.31665/JFB.2021.15284Received: July 10, 2021; Revised received & accepted: September 28, 2021Citation: Ahmed, F. (2021). Redistribution of nutritionally important starch fractions beneficial to diabetic population by using oats as a functional ingredient in a wheat-based food product – Jereesh. J. Food Bioact. 15: 74–79.
Abstract
Oats being important sources of β-glucan were incorporated at 10, 20 and 30% level in a wheat-based traditional Arabian breakfast cereal called Jereesh and evaluated for sensory acceptability. Sensory analysis reflected that the products with 10 and 20% oats incorporation were acceptable in terms of color, appearance, taste, aroma, mouthfeel, after taste and overall quality as indicated by the sensory scores which revealed no substantial differ-ences between control jereesh (CJ) and oats incorporated jereesh (OJ). However, jereesh with 30% oats incorpo-ration was very sticky (viscous) and scored low in sensory evaluation. Thus, nutritionally important starch frac-tions were measured in jereesh with 20% oats incorporation using a controlled in vitro multi-enzymatic method. Results indicated that, addition of oats at 20% level resulted in a substantial decrease (p<0.05) in total starch (TS), rapidly available glucose (RAG), and resistant starch content (RS), as well as a significant increase (p<0.05) in slow-ly digestible starch (SDS) compared to the control jereesh. The starch digestibility index remained unchanged. The findings of the study suggest that incorporation of oats helps in value addition of jereesh in terms of slow digest-ibility with increased soluble dietary fiber content that might lower glycemic index.
Keywords: Jereesh; Oats; Sensory analysis; Starch digestibility; Starch fractions.
1. Introduction
Cereal-based foods containing starch as the principal component form a major source of energy worldwide particularly in traditional diets. Nutritionally, distribution of starch fractions namely, rapidly digestible starch (RDS, hydrolyzed within 20 min), slowly digest-ible starch (SDS, hydrolyzed after 20 min but before 120 min), and resistant starch (RS, remain unhydrolyzed even after 120 min) which determine digestibility of starchy foods is of particular im-portance in populations with high prevalence of metabolic diseases (Englyst et al., 1992; Samarakoon, 2020).
Jereesh is one such wheat-based traditional food product with higher gelatinization of starch due to the sheer amount of water used in its preparation (Al-Faris, 2017). Traditionally, jereesh is made
with whole grains and chicken, wherein a single serving of 150 g provides 16% of the RDA for fiber. As a low glycemic index food (GI: 52), it is believed that jereesh intake can make a substantial contribution to the improvement of Saudi diets. However, in recent times consumption of jereesh is reduced and processed grains have now replaced whole grains diminishing the low GI advantage of jereesh (Nasib, 2003; Al-Mssallem, 2014). Several studies on ce-real starches have shown that starch digestibility is affected by both internal and extrinsic parameters, and the degree of gelatinization, which increases with water and cooking time, is one such factor that has a significant impact on starch digestibility because of the altered susceptibility of cereal starches to enzymatic breakdown (Englyst et al., 1992; Samarakoon, 2020; Samarakoon et al., 2020). Studies have shown that, the inclusion of certain food components or modi-fications in processing conditions might result in redistribution of

Journal of Food Bioactives | www.isnff-jfb.com 75
Ahmed Starch fractions in oats incorporated jereesh
starch fractions, opening the door to the production of tailored food products for different health conditions (Giri et al., 2017).
Oats are an example of a functional ingredient that has considera-ble potential for use in the formulation of functional foods. Their use, however, is restricted to porridges and other plain breakfast cereals (Butt et al., 2008). More recently, interest in using oats in food prod-uct development has increased, not only because of the presence of high amounts of β-glucans, which have a lipid lowering/plasma glu-cose stabilizing effect, but also because of the unique gelatinization characteristics of its starch, which behaves like a waxy starch due to high shear susceptibility (Peterson, 2004). In comparison to other cereal starches, the high viscosity starch gel thus generated form oats is less prone to retrogradation upon cooling, producing an elastic, sticky, and less rigid gel (Wang and White, 1994). These properties make oats starch appropriate for use in the formulation of functional foods. It is worth noting that adding oats into food products not only improves soluble dietary fiber content, which has lipid-lowering and plasma glucose-stabilizing effects, but also provides minor phyto-chemicals, which have health benefits including improved gut health (Harasym and Olędzki, 2018). Addition of oats will also substan-tially reduce gluten content of wheat-based food products (Thomp-son, 2003). Furthermore, limited studies have reported nutritionally important starch fractions in food preparations, particularly in tradi-tional food products; hence, modifying food formulations by substi-tuting the basic ingredients with oats is a viable approach to modify-ing starch digestibility. In view of this, the present study investigated the dietary acceptability of oats incorporated jereesh and studied the effect of oats incorporation on different starch fractions of nutritional significance using an in vitro multi-enzyme assay.
2. Materials and methods
2.1. Materials
All food ingredients were purchased from Nesto hypermarket, Ar Rass, Saudi Arabia. Amyloglucosidase (Sigma-Aldrich, St. Lou-is, USA), pancreatin (Sigma-Aldrich, St. Louis, USA), invertase (Sigma-Aldrich, St. Louis, USA), glucose oxidase-peroxidase rea-gent kit (Randox Laboratories, Crumlin, UK) were used. All of the other chemicals were of the highest analytical purity.
2.2. Preparation of jereesh
Crushed wheat (250 g) soaked in water for 4 hours was used as main ingredient. First seasoning was prepared for which oil (30 mL) was heated in a pan placed on low heat (<150 °C). To this chopped onion (40 g), finely chopped garlic (5 g), were added and sautéed till on-ions turned golden brown. Then tomato paste (30 mL), grated carrot (30 g), dried lemon powder (2.5 g), black pepper (5 g) and salt (5 g) were added and sauteed for 2–3 min following with addition of water (250 mL) and soaked crushed wheat. Mixed well and allowed to cook 10 min on medium heat (160–190 °C) till wheat became soft. To prepare oats incorporated jereesh (OJ), crushed wheat was replaced with whole oats at 10, 20 and 30% levels based on prelimi-nary studies incorporating oats in different traditional Indian food preparations of high glycemic index (Giri et al., 2017).
2.3. Sensory analysis
Quantitative descriptive analysis was used to assess the sensory
acceptability of oats incorporated jereesh on a scale of 0–10 cm. The one end of the scale marked as low represented the recognition threshold, while the other end of the scale marked as high indi-cated the saturation threshold (Stone et al., 2008). The developed score card listed appearance, color, taste, after taste, mouthfeel and overall quality as attributes of the product. Sensory analysis was carried out by 25 graduate student panelists having completed food analysis course and familiar with the product attributes and sen-sory evaluation techniques. Sensory analysis was carried out in air-conditioned sensory booths (18 ± 2 °C) lit with white fluorescent light and maintained at RH 60 ± 5% using ultrasonic humidifier (Buerer LB88, Ulm, Germany).
The food samples were coded with three-digit random codes to avoid bias and served hot (50–60 °C) in small cups with disposable spoons. Along with the samples, packaged drinking water was giv-en as palate cleanser. Panelists were asked to draw a vertical line on the scale and write the code number to indicate the perceived strength of each attribute specified on the score card. The scores for each attribute for each sample were tabulated, reflecting the panelists’ decision. Finally, a mean value was calculated for each sample attribute, reflecting the panel’s opinion on the perceived sensory quality and dietary acceptability. A visual representation of sensory quality in the form of sensory profile was plotted us-ing radar plot in OriginPro 2018 software (OriginLab Corporation, MA, USA).
2.4. Measurement starch fractions of nutritional significance and starch digestibility index
Figure 1 depicts a summary of the methodological approach em-ployed. Total starch (TS) and nutritionally important starch frac-tions namely, rapidly digestible starch (RDS), resistant starch (RS) and rapidly available glucose (RAG) were measured by the meth-od of Englyst et al. (1992). Incubation with pancreatin, invertase and amyloglucosidase at 37 °C in 50 mL polypropylene capped tubes positioned in a shaking water bath was used to determine different starch fractions in freshly prepared food samples. Glass beads and guar gum were added to the tubes to simulate intesti-nal digestive conditions by disrupting food particles and increas-ing viscosity of incubation mixture, respectively. Glucose content of the reaction mixture was measured at 20 min (G20) to reflect rapidly available glucose RAG and at 120 min (G120) to measure slowly digestible starch (SDS). The tubes were then placed in boil-ing water to gelatinize starch and treated with 7M KOH at 0 °C and the glucose content was measured to estimate total glucose (TG). After 120 minutes of incubation, resistant starch was characterized as the starch that remained unhydrolyzed. The amount of free glu-cose (FG) in the sample was measured by using acetate buffer and putting the tubes in a boiling water bath for 30 min. Simultaneous experiments with a standard dextrose were performed in a similar manner. To account for the glucose present in enzyme solutions, a blank tube containing buffer, glass beads, and guar gum was used. The glucose levels in all of the samples were measured using a glucose oxidase – peroxidase diagnostic package.
From the values of G20, G120, FG, and TG, the following values for TS, RDS, SDS, and RS were determined.
20
120 20
120
Total Starc s (TS) (TG FG) 0.9Rapidly Digestible Starch (RDS) (G FG) 0.9Slowly Digestible Starch (SDS) (G G ) 0.9Resistant Starch (RS) TS (RDS SDS) 0.9 or (TG-G ) 0.9
= - ´= - ´
= - ´
= - - ´ ´
Starch digestibility index and rapidly available glucose were
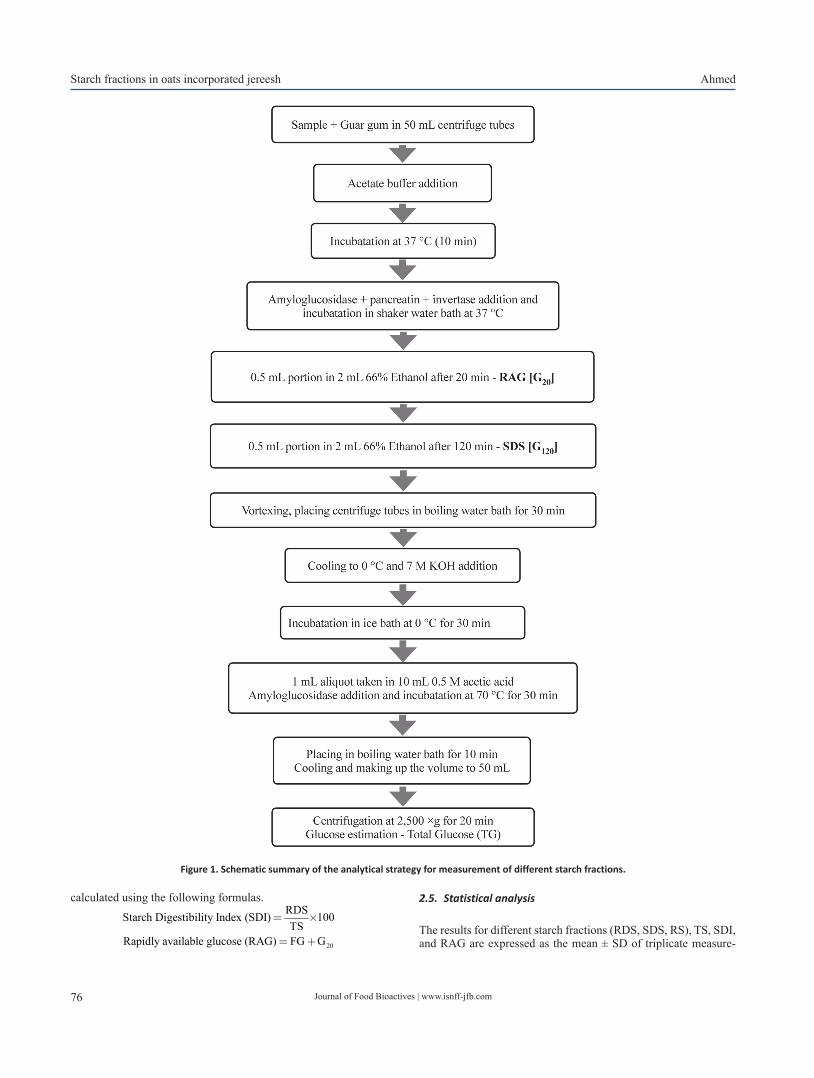
Journal of Food Bioactives | www.isnff-jfb.com76
Starch fractions in oats incorporated jereesh Ahmed
calculated using the following formulas.
20
RDSStarch Digestibility Index (SDI) 100TS
Rapidly available glucose (RAG) FG G
= ´
= +
2.5. Statistical analysis
The results for different starch fractions (RDS, SDS, RS), TS, SDI, and RAG are expressed as the mean ± SD of triplicate measure-
Figure 1. Schematic summary of the analytical strategy for measurement of different starch fractions.
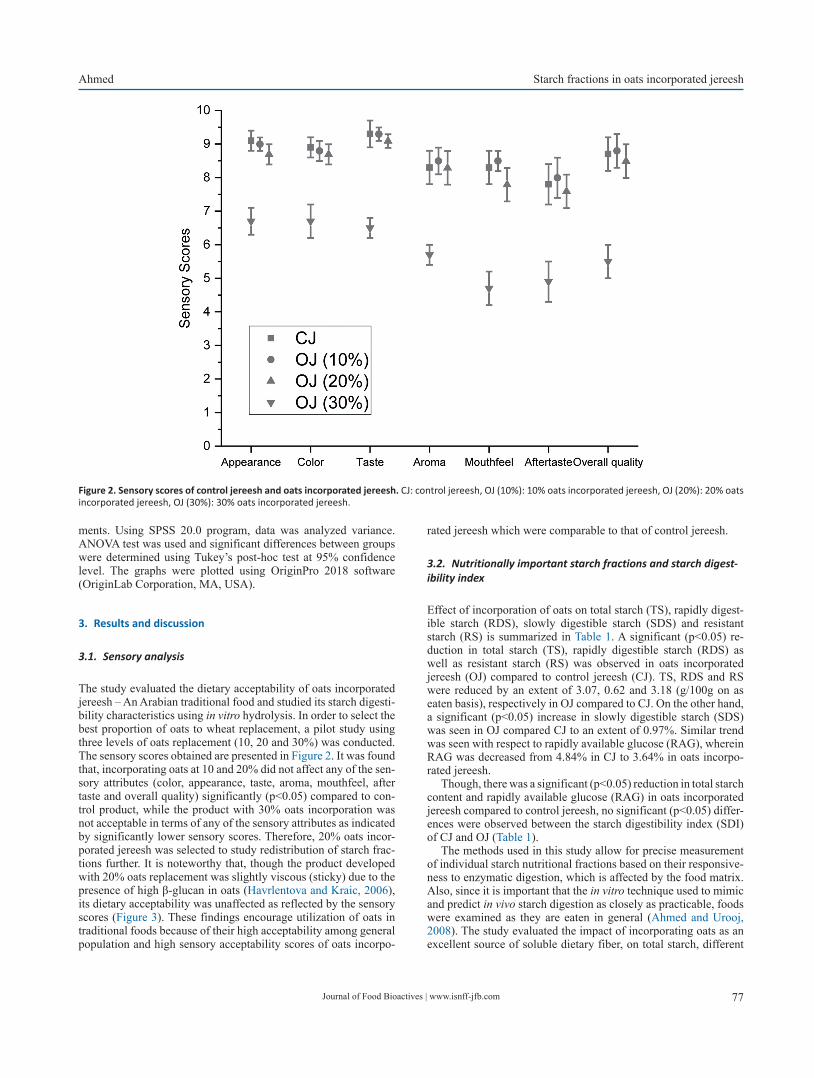
Journal of Food Bioactives | www.isnff-jfb.com 77
Ahmed Starch fractions in oats incorporated jereesh
ments. Using SPSS 20.0 program, data was analyzed variance. ANOVA test was used and significant differences between groups were determined using Tukey’s post-hoc test at 95% confidence level. The graphs were plotted using OriginPro 2018 software (OriginLab Corporation, MA, USA).
3. Results and discussion
3.1. Sensory analysis
The study evaluated the dietary acceptability of oats incorporated jereesh – An Arabian traditional food and studied its starch digesti-bility characteristics using in vitro hydrolysis. In order to select the best proportion of oats to wheat replacement, a pilot study using three levels of oats replacement (10, 20 and 30%) was conducted. The sensory scores obtained are presented in Figure 2. It was found that, incorporating oats at 10 and 20% did not affect any of the sen-sory attributes (color, appearance, taste, aroma, mouthfeel, after taste and overall quality) significantly (p<0.05) compared to con-trol product, while the product with 30% oats incorporation was not acceptable in terms of any of the sensory attributes as indicated by significantly lower sensory scores. Therefore, 20% oats incor-porated jereesh was selected to study redistribution of starch frac-tions further. It is noteworthy that, though the product developed with 20% oats replacement was slightly viscous (sticky) due to the presence of high β-glucan in oats (Havrlentova and Kraic, 2006), its dietary acceptability was unaffected as reflected by the sensory scores (Figure 3). These findings encourage utilization of oats in traditional foods because of their high acceptability among general population and high sensory acceptability scores of oats incorpo-
rated jereesh which were comparable to that of control jereesh.
3.2. Nutritionally important starch fractions and starch digest-ibility index
Effect of incorporation of oats on total starch (TS), rapidly digest-ible starch (RDS), slowly digestible starch (SDS) and resistant starch (RS) is summarized in Table 1. A significant (p<0.05) re-duction in total starch (TS), rapidly digestible starch (RDS) as well as resistant starch (RS) was observed in oats incorporated jereesh (OJ) compared to control jereesh (CJ). TS, RDS and RS were reduced by an extent of 3.07, 0.62 and 3.18 (g/100g on as eaten basis), respectively in OJ compared to CJ. On the other hand, a significant (p<0.05) increase in slowly digestible starch (SDS) was seen in OJ compared CJ to an extent of 0.97%. Similar trend was seen with respect to rapidly available glucose (RAG), wherein RAG was decreased from 4.84% in CJ to 3.64% in oats incorpo-rated jereesh.
Though, there was a significant (p<0.05) reduction in total starch content and rapidly available glucose (RAG) in oats incorporated jereesh compared to control jereesh, no significant (p<0.05) differ-ences were observed between the starch digestibility index (SDI) of CJ and OJ (Table 1).
The methods used in this study allow for precise measurement of individual starch nutritional fractions based on their responsive-ness to enzymatic digestion, which is affected by the food matrix. Also, since it is important that the in vitro technique used to mimic and predict in vivo starch digestion as closely as practicable, foods were examined as they are eaten in general (Ahmed and Urooj, 2008). The study evaluated the impact of incorporating oats as an excellent source of soluble dietary fiber, on total starch, different
Figure 2. Sensory scores of control jereesh and oats incorporated jereesh. CJ: control jereesh, OJ (10%): 10% oats incorporated jereesh, OJ (20%): 20% oats incorporated jereesh, OJ (30%): 30% oats incorporated jereesh.

Journal of Food Bioactives | www.isnff-jfb.com78
Starch fractions in oats incorporated jereesh Ahmed
starch nutritional fractions and its digestibility in a wheat-based breakfast cereal – jereesh. Despite the fact that the developed prod-uct was viscous, its acceptability was high, and the observed rise in product’s viscosity can be attributed to higher β-glucan content of oats (Havrlentova and Kraic, 2006).
As expected, incorporation oats in jereesh significantly de-creased total starch content, rapidly digestible starch, resistant starch and rapidly available glucose but starch digestibility index remained unchanged. The decrease in total starch content is pro-portionate to starch content of the ingredients used which can be attributable to the replacement of wheat starch with β-glucan pre-sent in oats. Similar findings have been reported earlier by Bren-nan et al. (2008) wherein, incorporation of dietary fiber source in extruded breakfast cereal products reduced led to a significant reduction in total starch content. This observation is also in good agreement with an earlier study wherein, incorporation of oats led
to a significant reduction in the total starch content of some tra-ditional Indian foods – idli, dosa, upma, and chapatti (Giri et al., 2017).
The involvement of several factors in food preparations, the most important of which are degree of gelatinization, cooking procedures, food source, and macronutrient composition, regu-lates the in vitro rate of starch digestion (Urooj and Puttraj, 1999). RAG is the sum of RDS and free sugars that are present in the food and considered useful in predicting the glycemic responses in vivo (Englyst et al., 1992; Englyst and Hudson, 1996). Therefore, a starch digestibility index of 18.54% observed in OJ aided with low RDS and RAG could be beneficial in maintaining glucose homeo-stasis amongst obese and glucose intolerant population.
When foods are exposed to conditions that decrease accessibil-ity of starch molecules to enzymatic activity either by creating a physical barrier across the enzyme or by inhibiting the enzyme, the rate of starch digestibility decreases (Tester et al., 2004). It is noteworthy that incorporation oat flours in different foods prod-ucts have been reported to inhibit starch digestibility, resulting in lower GI in foods (Kim and White, 2012). The results obtained in our study further substantiate the earlier findings and support the use of oats as functional ingredients in the development of novel foods products with reduced starch digestibility which is expected in various medical conditions such as diabetes, obesity, polycystic ovarian syndrome and metabolic syndrome. The findings indicate that, in food formulations, soluble fiber can influence the rate of starch digestibility to varying extent depending on several other factors such as food matrix, viscosity afforded by the soluble fibers and the amount of water used (Jenkins, 2007; Remya et al., 2017).
4. Conclusion
This study demonstrated the dietary acceptability of oats incorpo-
Figure 3. Sensory profile of control jereesh and oats incorporated jereesh. CJ: control jereesh, OJ: oats incorporated jereesh.
Table 1. Nutritionally important starch fractions in control and oats in-corporated jereesh (Mean ± SD g/100g)
Component* CJ OJ
Dry matter 28 28
TS 16.64b ± 0.57 13.57a ± 0.24
RDS 3.38a ± 0.21 2.52b ± 0.23
SDS 0.46a ± 0.09 1.43b ± 0.29
RS 12.80b ± 0.40 9.62a ± 0.33
RAG 4.84b ± 0.17 3.64a ± 0.28
SDI 20.30a ± 0.95 18.54a ± 1.39
*Mean values carrying different superscript letters a and b in rows differ significantly (p ≤ 0.05). CJ, control jereesh; OJ, oats incorporated jereesh; TS, total starch; RDS, rapidly digestible starch; SDS, slowly digestible starch; RS, resistant starch; RAG, rap-idly available glucose; SDI, starch digestibility index.

Journal of Food Bioactives | www.isnff-jfb.com 79
Ahmed Starch fractions in oats incorporated jereesh
rated jereesh; a traditional Arabic food. The observed decrease in RAG and RDS with associated increase in SDS levels might lower glycemic index of the developed product that could be beneficial for populations with impaired glucose tolerance, obesity and dia-betes. The findings of this study may help in the development of tailored functional foods with altered starch digestibility character-istics. However, further research is warranted to study the effect of oats incorporation in traditional food products on glycemic index in real life situations.
Acknowledgments
None to declare.
Conflict of interest
The author declares no conflict of interest.
References
Ahmed, F., and Urooj, A. (2008). In vitro starch digestibility and nutrition-ally important starch fractions in processed roots and tubers. Starch/Stärke 60: 493–499.
Al-Faris, N.A. (2017). Nutritional Evaluation of Selected Traditional Foods Commonly Consumed in Saudi Arabia. J. Food Nutr. Res. 5: 168–175.
Al-Mssallem, M.Q. (2014). The Association between the glycaemic index of some traditional saudi foods and the prevalence of diabetes in Saudi Arabia: A review article. J. Diabetes Metab. 5: 452.
Brennan, M.A., Monro, J.A., and Brennan, C.S. (2008). Effect of inclusion of soluble and insoluble fibres into extruded breakfast cereal products made with reverse screw configuration. Int. J. Food Sci. Technol. 43: 2278–2288.
Butt, M.S., Tahir-Nadeem, M., Khan, M.K.I., Shabir, R. and Butt, M.S. (2008). Oat: unique among the cereals. Eur J Nutr. 47: 68–79.
Englyst, H.N., and Hudson, G.J. (1996). The classification and measure-ment of dietary carbohydrates. Food Chem. 57: 15–21.
Englyst, H.N., Kingman, S.M., and Cummings, J.H. (1992). Classification and measurement of nutritionally important starch fractions. Eur. J.
Clin. Nutr. 46: S33–S50.Giri, S., Banerji, A., Lele, S.S., and Ananthanarayan, L. (2017). Starch di-
gestibility and glycaemic index of selected Indian traditional foods: Effects of added ingredients. Int. J. Food Prop. 20: S290–S305.
Harasym, J., and Olędzki, R. (2018). The mutual correlation of glucose, starch, and beta-glucan release during microwave heating and an-tioxidant activity of oat water extracts. Food Biopro. Technol. 11: 874–884.
Havrlentova, M., and Kraic, J. (2006). Content of β-D-glucan in cereal grains. J. Food Nutr. Res. 45: 97–103.
Jenkins, D.J.A. (2007). The Glycemic Index: Looking Back 25 Years. Cereal Foods World 52: 50–53.
Kim, H.J., and White, P.J. (2012). In vitro digestion rate and estimated gly-cemic index of oat flours from typical and high β-glucan oat lines. J. Agric. Food Chem. 60: 5237–5242.
Nasib, O.M. (2003). Effect of chemical component and methods of prepa-ration of some traditional Saudi Arabian foods on glycemic index. Thesis. King Saud University, College of Food & Agricultural Sciences, Saudi Arabia.
Peterson, D.M. (2004). Oat- multifunctional grain. In: 7th international oat conference, Helsinki, Finland. Agri. Food Res. Rep. 51: 21–26.
Remya, R., Jyothi, A.N., and Sreekumar, J. (2017). Comparative study of RS4 type resistant starches derived from cassava and potato starches via octenyl succinylation. Starch-Stärke. 69: 1600264.
Samarakoon, E.R.J. (2020). Impact of physical modifications on starch nu-tritional fractions: rapidly digestible starch, slowly digestible starch and resistant starch. J. Food Bioact. 12: 106–121.
Samarakoon, E.R.J., Waduge, R., Liu, Q., Shahidi, F., and Banoub, J.H. (2020). Impact of annealing on the hierarchical structure and physic-ochemical properties of waxy starches of different botanical origins. Food Chem. 303: 125344.
Stone, H., Sidel, J., Oliver, S., Woolsey, A., and Singleton, R.C. (2008). Sen-sory evaluation by quantitative descriptive analysis. Descriptive sen-sory analysis in practice. Food and Nutrition Press, Inc.
Tester, R.F., Karkalas, J., and Qi, X. (2004). Starch structure and digestibility Enzyme-Substrate relationship. World Poultry Sci. J. 60: 186–195.
Thompson, M.S. (2003). Oats and the gluten-free diet. J. Am. Diet. Assoc. 103: 376–379.
Urooj, A., and Puttraj, S. (1999). Digestibility index and factors affecting rate of starch digestion in vitro in conventional food preparation. Nahrung 43: 42–47.
Wang, L.Z., and White, P.J. (1994). Structure and physicochemical proper-ties of starches from oats with different lipid contents. Cereal Chem. 71: 443–450.

Copyright: © 2021 International Society for Nutraceuticals and Functional Foods.All rights reserved.
80
Original Research J. Food Bioact. 2021;15:80–89
Journal ofFood Bioactives International Society for
Nutraceuticals and Functional Foods
Optimized preparation of quassinoid-rich Eurycoma longifolia Jack phospholipid complex and its effects on oral bioavailability
Wu-Yan Guo#, Shu-Yan Wang#, Jian Ren# and Bo Zhang*
Tianjin Key Laboratory of Early Druggability Evaluation of Innovative Drugs, Tianjin International Joint Academy of Biomedicine, Tian-jin, 300457, China
*Corresponding author: Bo Zhang, Tian Jin International Joint Academy of Biomedicine, No.220 Dongting Road, the Tianjin Economic-Technological Development Area (TEDA), Tianjin, China. Tel: +86 022 65378890; Fax: 86-22-65378036; E-mail: [email protected]: 10.31665/JFB.2021.15285Received: June 24, 2021; Revised received & accepted: September 30, 2021Citation: Guo, W.-Y., Wang, S.-Y., Ren, J., and Zhang, B. (2021). Optimized preparation of quassinoids rich Eurycoma longifolia Jack, TAF2-phospholipid complex and its effects on oral bioavailability. J. Food Bioact. 15: 80–89.
Abstract
Eurycoma longifolia Jack is a plant of the genus Eurycoma in Simaroubaceae. Eurycomanone is one of the main quassinoids of E. longifolia. Unfortunately, these quassinoids exhibit low bioavailability. In the present study, a phospholipid complex (TAF2-PC) was prepared to improve the oral bioavailability of TAF2 (Tongkat Ali Fraction 2). The complexation rate of eurycomanone and phospholipid was used as the index of success in the phospho-lipid complexes. The effects of reaction solvent, reaction temperature, drug concentration, drug-lipid ratio and reaction time on the complexation rate were investigated. Response Surface-Box-Behnken Design Method was performed to optimize the preparation technology and the pharmacokinetics of phospholipid complex of TAF2 were studied in rats. Our results showed that the optimum preparation condition of TAF2-phospholipid complex were as follows: reaction solvent, anhydrous ethanol; reaction time, 1h; the ratio of TAF2 to phospholipid, 2.28; reaction temperature, 40.85 °C; TAF2 concentration, 32.66 mg/ml; and complexation rate, 95.04 ± 0.4% (n=3). The results of pharmacokinetic study showed that the relative bioavailability was increased to 209.20% in the TAF2-phospholipid complex treated rats. Thus, this phospholipid-complex-technique shows potential for enhanc-ing oral bioavailability of poorly absorbed quassinoid-rich E. longifolia extract, TAF2.
Keywords: Eurycoma longifolia; Quassinoids; Eurycomanone; Phospholipid complex; bioavailability.
1. Introduction
Eurycoma longifolia Jack (known as tongkat ali), one of the most popular traditional herbal medicine, is a flowering plant of the family Simaroubaceae, native to Indonesia, Malaysia, Vietnam and also Cambodia, Myanmar, Laos and Thailand. Decoctions of E. longifolia leaves are used for relieving itches, while its fruits are used to treat dysentery. Its bark is used mainly as an insect repel-lent, while the taproots are used to treat hypertension, and the root bark is used for the treatment of diarrhea and fever. The use of Eu-rycoma longifolia Jack as food dates back to the colonial period of 1,700 years ago. Indigenous people in Malaysia are used to taking tongkat ali decoction to treat intermittent fever, and Malay women
also take tongkat ali decoction as a tonic after childbirth. Mostly, the roots extract of E. longifolia are used as indigenous medicine for sexual dysfunction, aging, malaria, cancer, diabetes, anxiety, pain, constipation, exercise recovery, fever, leukemia, osteoporo-sis, stress, syphilis and swollen glands, as well as an aphrodisiac, antibiotic, appetite stimulant and health supplement (Chan et al., 1989; Darise et al., 1983; Fiaschetti et al., 2011; Hussein et al., 2007; Katsunori et al., 2009; Kuo et al., 2004; Morré et al., 1998).
E.longifolia is a rich source of various classes of bioactive com-pounds, which includes quassinoids, β-carboline alkaloids, canthin-6-one alkaloids, squalene derivatives, biphenyl neolignan, and bio-active steroids (Ang et al., 2000; Katsunori et al., 2009; Kuo et al., 2004; Shah et al., 2012; Tran et al., 2014; Zakaria et al., 2009). A

Journal of Food Bioactives | www.isnff-jfb.com 81
Guo et al. Optimized preparation of Eurycoma longifolia Jack phospholipid complex
study by Lin et al. (2013) reported that over 50 quassinoids were present in E.longifolia. while eurycomanone 1 showed the most abundant content (Ling et al., 2013). As the main bioactive compo-nent of E.longifolia, the quassinoids have been reported to be phar-macological active against various diseases including malaria, anxi-ety, aging, osteoporosis treatment and improving sperm motility.
The bioavailability of the constituent eurycomanone was pharma-cologically investigated in animal research. Low et al. demonstrated that following intravenous injection, eurycomanone was detected in the plasma, declining to zero within 8 h (Low et al., 2011). Following oral administration, Cmax and Tmax values were 0.33 ± 0.03 μg/mL and 4.40 ± 0.98 h, respectively. The plasma concentration was lower following oral administration versus intravenous administration, even at a much higher oral dose (five times the dose). It suggested that eury-comanone is poorly bioavailable (10.5%) when given orally. Following oral administration of Tongkat Ali Fraction 2 (TAF2), 13α(21)-epoxy-eurycomanone had a higher Cmax than eurycomanone (1.61 ± 0.41 μg/mL vs. 0.53 ± 0.10 μg/mL) (Chan et al., 2008; Leng et al., 2015; Low et al., 2011). The experimental results demonstrated that the absolute bioavailability was also higher due to increased membrane permeabil-ity. Moreover, eurycomanol and 13α,21-dihydroeurycomanone were not detected in plasma (Low et al., 2011). The distribution, excretion and half life of eurycomanone have been reported in many studies. In animal study, the volume of distribution (Vd) of eurycomanone was relatively high (0.68 ± 0.30 L/kg), suggesting that it is well distributed in the extravascular fluids (Low et al., 2005). Following intravenous injection, the mean elimination rate constant (ke) and clearance (CL) for eurycomanone were 0.88 ± 0.19 h−1 and 0.39 ± 0.08 L/h/kg, re-spectively and 13α(21)-epoxyeurycomanone had a longer biological half-life than eurycomanone (0.75 ± 0.25 h vs. 0.35 ± 0.04 h), due to the elimination rate constant (Low et al., 2005).
E.longifolia has a remarkable pharmacological activity. How-ever, its poor oral absorption, fast metabolic rate and low oral bio-availability lead to its reduced efficacy. This drawback could be figured out by performing drug-phospholipid complex strategy, as they utilize mechanism which is similar to the absorption mecha-nism of nutritional constituents form gastro intestinal tract. The phospholipid complex enhanced the lipid solubility of hydrophilic plant components, enhancing their ability to cross phospholipid bi-layer biofilms. The active components of herbal extracts from de-struction by digestive enzymes and intestinal bacteria are protected by phospholipid complex (Jia et al., 2010).
A traditional Malaysian herb, E.longifolia, contains a large num-ber of active ingredients. Most of these active substances have good water solubility and poor fat solubility, leading to a small amount of these active ingredients that can be effectively absorbed by the hu-man body. Thus, in present study, we attempt to prepare novel quassi-noids rich fractions (Tongkat Ali Fraction 2, TAF2) – phospholipid complex using the solvent evaporation method to improve the lipo-solubility of active substance of E.longifolia and thereby to promote its oral bioavailability. The effects of reaction solvent, reaction tem-perature, drug concentration, drug-lipid ratio and reaction time on the complexation rate were investigated. Box-Behnken design-response surface method was performed to optimize TAF2-phospholipid com-plex (PC) preparation. The characteristics of TAF2-PC were deter-mined and the pharmacokinetics of TAF2-PC was compared with free TAF2 as control after the oral administration in rats.
2. Materials and methods
2.1. Materials
Root of Eurycoma longifolia (Guangzheng Biological Technology
Co. Ltd. Kunming, China); Eurycomanone (mass fraction >95%; Chengdu alfa Biological Technology Co. Ltd. Chengdu, China); DI-AION HP-20 resin (Mitsubishi Chemical Holdings. Tokyo, Japan); Methyl alcohol (chromatographically pure; Tianjin heowns Biologi-cal Technology Co. Ltd. Tianjin, China); Ethyl alcohol (analytically pure; China National Pharmaceutical Group Corporation. Beijing, China); Acetonitrile (chromatographically pure; Merck KGaA, Darmstadt, Germany); Ultra-pure water was generated using a Mil-lipore Milli-Q Gradient System (Millipore Corp. Billerica, MA, USA); Egg yolk lecithin (Beijing Nuoqi Yasheng Biotechnology Co. Ltd. Beijing, China); Tetrahydrofuran, Acetone, Cyclohexane, Dichloromethane (Tianjin Chemical Reagent Supply and Marketing Co. Ltd. Tianjin, China).
2.2. Preparation and fractionation of Eurycoma longifolia extract
The granule roots of E. longifolia of 100 g were dissolved with 5 times volume of solvent (10% ethyl alcohol) and put in a heating jacket and heat for 4 hours/day, for 5 times consecutively. All of the extracts were collected, concentrated and dried under decom-pression, the crude extract was obtained and stored at low tem-perature (4 °C). The crude extract redissolved in a small volume of ethyl alcohol, was then passed through a resin column of Diaion® HP 20 and eluted by gradient mixtures of increasing ratio of ethyl alcohol in water (0:100–100:0) to afford four fractions (TAF1, TAF2, TAF3 and TAF4) (Low et al., 2011), concentrated, dried under decompression, and store at −20 °C.
2.3. Preparation of phospholipid complex
2.3.1. Separation solvent selection
Selection of solvent is one of the critical parameters while prepar-ing drug and phospholipid complex. The solvent should be select-ed based on the solubility of both drug and phospholipid. It must have the ability to dissolve phospholipid complex and phospho-lipid but not the drug or to dissolve drugs and phospholipids but not the phospholipid complex. Selecting the proper solvent is very important to calculate the complexation rate (CR) (Yu et al., 2016). Briefly, the TAF2 and TAF2-PC complexes were placed in white vials with sufficient solvents as follow methyl alcohol, acetone, cyclohexane and 1, 4-dioxane, respectively. Vortex over a period of time to observe the dissolution.
2.3.2. Preparation technique of TAF2-phospholipid complex (PC)
Herein, single factor experiment (Zhou et al., 2012)was used to in-vestigate the influence of several major parameters of the complexa-tion rate of TAF2-PC, such as reaction solvent, reaction tempera-ture, reaction drug concentration, drug-lipid ratio and reaction time. Then, the Box-Behnken Design (BBD) coupled with response sur-face methodology (RSM) with 3 factors and 3 levels was employed to determine the best preparation process of TAF2-PC.
2.3.3. Reactive solvents screening
The complex was initially prepared with TAF2 and egg yolk leci-thin at a mass ratio of 1:2, the concentration of TAF2 was 20 mg/mL, the reaction temperature was 40°C, and the reaction time was

Journal of Food Bioactives | www.isnff-jfb.com82
Optimized preparation of Eurycoma longifolia Jack phospholipid complex Guo et al.
1 h. The effects of methyl alcohol, ethyl alcohol, tetrahydrofuran and dichloromethane on the complexation rate were compared.
2.3.4. Reaction temperature screening
The complex was prepared with TAF2 and egg yolk lecithin at a mass ratio of 1:2, the concentration of TAF2 was 20 mg/mL, the reaction time was 1 h, the reaction solvent was ethyl alcohol. The effects of different reaction temperatures (30, 40, 50, 60°C) on the complexation rate were compared.
2.3.5. Reaction TAF2 concentration screening
The complex was prepared with TAF2 and egg yolk lecithin at a mass ratio of 1:2, the reaction temperature was 40°C, the reaction time was 1 h, and the reaction solvent was ethyl alcohol. The ef-fects of different TAF2 concentrations (20, 40, 60, 80 mg/mL) on the complexation rate were compared.
2.3.6. TAF2-phospholipid ratio screening
A 20 mg/mL TAF2 concentration, the reaction temperature was 40°C, 1h reaction time, and anhydrous ethyl alcohol as reaction solvent were set and the effects of mass ratios of different TAF2 and phospholipids (1:1, 1:2, 1:3, 1:4) on the complexation rate were compared.
2.3.7. Reaction time screening
The complex was prepared with TAF2 and egg yolk lecithin at a mass ratio of 1:2, the concentration of TAF2 was 20 mg/mL, the reaction temperature was 40°C, and ethyl alcohol as reaction sol-vent. The effects of reaction time (1, 2, 3, 4 h) on the complexation rate were compared.
2.3.8. The Box-Behnken Design (BBD) – response surface meth-odology (RSM)
After examining the influence of each single factor on the experi-ment results, the three factors with the most significant influence were selected. The Box-Behnken design (BBD) coupled with re-sponse surface methodology (RSM) with 3 factors and 3 levels was performed to optimize the preparation process of TAF2-PC.
2.3.9. Preparation of TAF2-PC
TAF2 and PC were combined by solvent evaporation method(Xiao et al., 2006). Briefly, a certain amount of TAF2 and egg yolk leci-thin were dissolved in anhydrous ethyl alcohol. The mixture was stirred gently in a water bath for definite time. The solution was evaporated in a rotary evaporator under vacuum, the ethyl alcohol was removed and dried in vacuum for 24 h to obtain the TAF2-phospholipid complex, and store at 4°C.
2.4. TAF2-PC complexation rate
The complexation rates of TAF2-PC was determined as follows.
Briefly, approximately 1 g of TAF2-PC was dissolved in 15 mL of the separation solvent and vortex violently. The residue and uncompounded TAF2 were removed by 0.45µm nylon membrane filtration. Concentrated TAF2-PC was obtained by evaporating filtrate. The filtrated precipitate was dissolved and the amount of uncombined TAF2 was determined by high performance liquid chromatography/HPLC system (Waters-e2695, Waters). The com-plexation rates (CR) (Yu et al., 2016) of TAF2-PC was determined using the following equation:
(%) 100%total uncomplexed
total
W WCR
W-
= ´
where “Wtotal” is the initial quantity of TAF2 and “Wuncomplexed” is the quantity of uncombined TAF2
2.5. Characterization of TAF2-phospholipid complex
There are several characterization instruments available for char-acterizing drug-phospholipid complexes(Semalty et al., 2012). Morphological evaluation of TAF2 and TAF2-PC was performed by scanning electron microscope, SEM (JSM-6390LV, Jeol Ltd, Tokyo, Japan). In brief, a small piece of double-sided adhesive tape was fixed to an aluminum stub. Powders were sprinkled into a dispersed layer on the stub. Prior to examination, the samples were sputter coated with gold–palladium using an ion sputter (JFC1600, Jeol Ltd, Tokyo, Japan).
To analyze the thermal behavior of the obtained complex, dif-ferential scanning calorimeter (DSC) curves were recorded by a DSC instrument (DSC3, Mettler-Toledo International Inc., Swit-zerland). The accurately weighed samples including TAF2, egg yolk lecithin, the physical mixture and TAF2-PC were sealed into aluminium plate separately. The investigation was carried out over the temperature range 25°Cto 200°Cat a continuous heating rate of 10°C/min in 0.2 mL/min atmospheric nitrogen.
Infrared (IR) spectroscopy is the primary tool to predict any interactions between the drug and phospholipid. IR spectro-scopic analyses of TAF2, phospholipid, and TAF2-PC were per-formed using FTIR-850 Fourier transform-Infrared spectrometer (Tianjin Port East Company). In brief, approximately 1–2 mg of TAF2, lecithin, the physical mixture and TAF2-PC were weighed and placed in a mortar, ground and mixed with KBr pellet at a mass ratio of 1:100, then put into the mold and pressed into tab-lets for the test. Spectra were recorded in the range 7,800–350 cm−1.
2.6. In vivo pharmacokinetic study of TAF2-PC
Male Sprague–Dawley (SD) rats (220 ± 20 g) were obtained from Animal Center of the Chinese Academy of Science After passing the quarantine by GLP (Good Laboratory Practice) and feeding for 5 days, the experiment was carried out. During the feeding period, the rats were given standard rodent feed and freely drinking water. The experimental design was approved by the Animal Ethics Com-mittee of Tianjin International Joint Institute of Biomedical Sci-ences. The rats were fasted 24h with free access to water. Twelve animals were divided randomly into two groups (n = 6). TAF2 or TAF2-PC (equivalent to 200 mg/kg of TAF2) was dissolved in 2 mL distilled water and orally administered with a gavage needle. Blood samples of 0.5mL were collected from oculi chorioideae venous of the rats at 0, 0.083, 0.25, 0.5, 0.75, 1, 2, 4, 6, 8 and 10 h after oral administration, respectively. Heparin coated vials containing blood samples were then centrifuged at 8,000 g for 10

Journal of Food Bioactives | www.isnff-jfb.com 83
Guo et al. Optimized preparation of Eurycoma longifolia Jack phospholipid complex
min. A 100 µL of the plasma sample were deproteinized by adding 5 µL of 70% perchloric acid. The mixture was mixed for 2 min on a vortex and then centrifuged at 8,000 g for 10 min. The clear su-pernatant layer was collected, and 20 µL were injected for analysis in the HLPC system(Xiao et al., 2006) to analyze the concentration of eurycomanone.
The maximum concentration (Cmax), time to reach maximum concentration (Tmax), area under the concentration-time curve (AUC0–∞), and elimination half-life (Thalf) were obtained from the plasma concentration–time curve and analyzed by the software Kinetica® 5.0 (Thermo Scientific, American). The results were ex-pressed as mean ± SD. The relative bioavailability (F) of TAF2-PC relative to TAF2 was calculated by the following formula:
2 2rel
2 2
(%) 100%TAF PC TAF
TAF TAF PC
AUC DFAUC D
-
-
= ´ ´
2.7. Statistical analysis
All of the experiments were repeated six times. The statistical analysis was carried out by one-way analysis of variance (ANO-VA), followed by LSD’s multiple comparison test using SPSS 25.0 (SPSS Inc., Chicago, IL, USA)software. Student’s t-test was used to analyze the differences between two groups in pharmacokinetic study. Data were expressed as mean ± standard deviation (SD). Statistical significance was declared at P<0.05.
3. Results and discussion
3.1. Screening of solvent for TAF2-PC preparation
Table 1 shows the dissolution of TAF2 and TAF2-PC in different solvents. Both methyl alcohol and 1, 4-dioxane have good solubil-ity for TAF2 and TAF2-PC complex and neither of them can be dissolved in acetone. Cyclohexane dissolves TAF2-PC complex well, but does not dissolve TAF2. Selection of solvent is one of the critical parameters while preparing drug and phospholipid com-plex. The solvent must have the ability to dissolve phospholipid complex and phospholipid but not the drug. Therefore, we select cyclohexane as the separation solvent instead of methyl alcohol, 1, 4-dioxane and acetone.
3.2. The preparation of TAF2-PC
Phospholipid complexes can be prepared by various methods, such as solvent evaporation, supercritical fluid processing, mechani-cal dispersion, and anhydrous co-solvent lyophilization(Xiao et al., 2006)(Lian and Wang, 2006)Of these, solvent evaporation is relatively mature and easy to perform. In present study, TAF2−PC was prepared using a solvent evaporation method, and the optimal complexation parameters such as reaction solvent, TAF2 to phos-pholipid ratio, temperature, and time were determined by single factor test.
A number of solvents were examined as the complexation me-dium to formulate TAF2-PC. This study showed that the methyl al-cohol has the highest complexation rate (88.0 ± 0.42), followed by ethyl alcohol, dichloromethane, and tetrahydrofuran (85.3 ± 1.50, 75.3 ± 2.36, and 64.6 ± 2.41), respectively (Table 2). The complex-ation rate of both methyl alcohol and ethyl alcohol are significantly higher than dichloromethane and tetrahydrofuran,respectively. The ethanol was finally chosen as the reaction solvent, due to the toxicity of methanol that may not fully removed from the product, which result in the excessive solvent residue.
Different reaction temperatures were applied to obtain the op-timum complexation rate. The results showed that the relatively higher complexation rate were achieved at 40 to 50°C compared with other temperatures, as indicated in table 3. The relatively lower temperature of 40°C was determined as the optimized tem-perature of present study, due to the well know trend that the low temperature the less chances to produce the impurities and degra-dation products.
The drug concentration of fixed drug carrier ratio is another important factor that may significantly affect the complexation rate of the final product. The result of present study suggested that the loading concentration of 40 mg/mL have reaches the saturated level. The complexation rates were not improved by increasing the drug loading anymore (Table 4).
The drug carrier ratio is also necessary to be evaluated, as the entire volume of single dose, the cost of the carrier, and the com-plexation rate need to be collectively balanced. The results in table 5 suggest that the Drug-phospholipid ratio of 1:2 and 1:3 result in very significant higher complexation rate compared with other ratios. However, the appearance of the complex showed that the
Table 1. The solubility of TAF2 and TAF2-PC in different solvents
Solvents TAF2-PC TAF2
Methyl alcohol easily soluble easily soluble
Acetone sparingly soluble sparingly soluble
Cyclohexane easily soluble insoluble
1, 4-dioxane easily soluble easily soluble
Table 2. The effect of reaction solvent on complexation rate
No. Reactive solvents Complexation rate (%) P-value
1 Methyl alcohol 88.0 ± 0.42 <0.05
2 Ethyl alcohol 85.3 ± 1.50 <0.05
3 Tetrahydrofuran 64.6 ± 2.41 –
4 Dichloromethane 75.3 ± 2.36 –
Table 3. The effect of reaction temperature on complexation rate
No. Reaction tem-perature (°C)
Complexa-tion rate (%) P-value
1 30 75.2 ± 2.40 –
2 40 85.0 ± 1.57 <0.05
3 50 83.2 ± 2.14 <0.05
4 60 74.3 ± 3.24 –
Table 4. The effect of reaction drug concentration on complexation rate
No. Reaction drug concentration(mg/mL)
Complexa-tion rate (%) P-value
1 20 85.0 ± 1.57
2 40 92.3 ± 1.10 <0.05
3 60 88.5 ± 2.04
4 80 82.1 ± 0.72
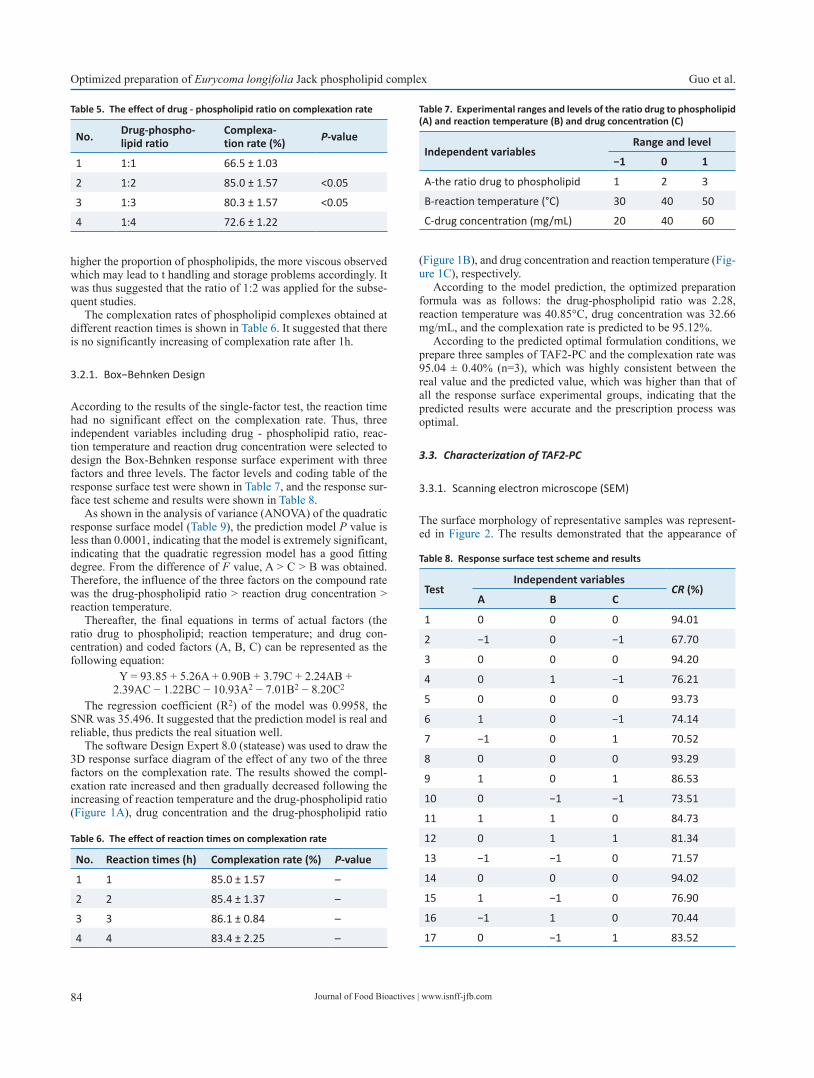
Journal of Food Bioactives | www.isnff-jfb.com84
Optimized preparation of Eurycoma longifolia Jack phospholipid complex Guo et al.
higher the proportion of phospholipids, the more viscous observed which may lead to t handling and storage problems accordingly. It was thus suggested that the ratio of 1:2 was applied for the subse-quent studies.
The complexation rates of phospholipid complexes obtained at different reaction times is shown in Table 6. It suggested that there is no significantly increasing of complexation rate after 1h.
3.2.1. Box−Behnken Design
According to the results of the single-factor test, the reaction time had no significant effect on the complexation rate. Thus, three independent variables including drug - phospholipid ratio, reac-tion temperature and reaction drug concentration were selected to design the Box-Behnken response surface experiment with three factors and three levels. The factor levels and coding table of the response surface test were shown in Table 7, and the response sur-face test scheme and results were shown in Table 8.
As shown in the analysis of variance (ANOVA) of the quadratic response surface model (Table 9), the prediction model P value is less than 0.0001, indicating that the model is extremely significant, indicating that the quadratic regression model has a good fitting degree. From the difference of F value, A > C > B was obtained. Therefore, the influence of the three factors on the compound rate was the drug-phospholipid ratio > reaction drug concentration > reaction temperature.
Thereafter, the final equations in terms of actual factors (the ratio drug to phospholipid; reaction temperature; and drug con-centration) and coded factors (A, B, C) can be represented as the following equation:
Y = 93.85 + 5.26A + 0.90B + 3.79C + 2.24AB + 2.39AC − 1.22BC − 10.93A2 − 7.01B2 − 8.20C2
The regression coefficient (R2) of the model was 0.9958, the SNR was 35.496. It suggested that the prediction model is real and reliable, thus predicts the real situation well.
The software Design Expert 8.0 (statease) was used to draw the 3D response surface diagram of the effect of any two of the three factors on the complexation rate. The results showed the compl-exation rate increased and then gradually decreased following the increasing of reaction temperature and the drug-phospholipid ratio (Figure 1A), drug concentration and the drug-phospholipid ratio
(Figure 1B), and drug concentration and reaction temperature (Fig-ure 1C), respectively.
According to the model prediction, the optimized preparation formula was as follows: the drug-phospholipid ratio was 2.28, reaction temperature was 40.85°C, drug concentration was 32.66 mg/mL, and the complexation rate is predicted to be 95.12%.
According to the predicted optimal formulation conditions, we prepare three samples of TAF2-PC and the complexation rate was 95.04 ± 0.40% (n=3), which was highly consistent between the real value and the predicted value, which was higher than that of all the response surface experimental groups, indicating that the predicted results were accurate and the prescription process was optimal.
3.3. Characterization of TAF2-PC
3.3.1. Scanning electron microscope (SEM)
The surface morphology of representative samples was represent-ed in Figure 2. The results demonstrated that the appearance of
Table 5. The effect of drug - phospholipid ratio on complexation rate
No. Drug-phospho-lipid ratio
Complexa-tion rate (%) P-value
1 1:1 66.5 ± 1.03
2 1:2 85.0 ± 1.57 <0.05
3 1:3 80.3 ± 1.57 <0.05
4 1:4 72.6 ± 1.22
Table 6. The effect of reaction times on complexation rate
No. Reaction times (h) Complexation rate (%) P-value
1 1 85.0 ± 1.57 –
2 2 85.4 ± 1.37 –
3 3 86.1 ± 0.84 –
4 4 83.4 ± 2.25 –
Table 7. Experimental ranges and levels of the ratio drug to phospholipid (A) and reaction temperature (B) and drug concentration (C)
Independent variablesRange and level
−1 0 1
A-the ratio drug to phospholipid 1 2 3
B-reaction temperature (°C) 30 40 50
C-drug concentration (mg/mL) 20 40 60
Table 8. Response surface test scheme and results
TestIndependent variables
CR (%)A B C
1 0 0 0 94.01
2 −1 0 −1 67.70
3 0 0 0 94.20
4 0 1 −1 76.21
5 0 0 0 93.73
6 1 0 −1 74.14
7 −1 0 1 70.52
8 0 0 0 93.29
9 1 0 1 86.53
10 0 −1 −1 73.51
11 1 1 0 84.73
12 0 1 1 81.34
13 −1 −1 0 71.57
14 0 0 0 94.02
15 1 −1 0 76.90
16 −1 1 0 70.44
17 0 −1 1 83.52
Journal of Food Bioactives | www.isnff-jfb.com 85
Guo et al. Optimized preparation of Eurycoma longifolia Jack phospholipid complex
free TAF2 particles is irregular, which is composed of small crys-talline particles and polydisperse aggregates (Figure 2A). When TAF2 form a complex with phospholipids, it showed significantly change in morphology and shape. They form granules of different sizes with the absence of crystallinity of TAF2 (Figure 2B). It sug-gested that the TAF2 molecules may form amorphous mixture with phospholipids, the molecular surface is covered by phospholipid molecules, losing the original crystal structure.
3.3.2. Differential scanning calorimeter (DSC)
DSC was performed to analyze the interactions between multiple components of a drug preparation. TAF2 showed endothermic peak group between 99.8°Cand 127.8°C (Figure 3A), this is related to TAF2 is a mixture of a variety of quassinoids compounds. The phospholipids had a melting point peak at 153.3°C (Figure 3B). In the physical mixture of TAF2 and phospholipids, an endothermic peak was observed approximately 130.5°C(Figure 3C), which was close to the phase transition point of TAF2 at 127.8°C. It was sug-gested that following by increasing temperature, the phospholipids melt, the drug dissolves in the phospholipids and partially forms phospholipid complexes, which making the slightly change of endothermic peak. Thermal analysis diagram of TAF2-PC shows wide endothermic peak at 65.2°C(Figure 3D). The original peaks of TAF2 and phospholipid disappeared from the thermal spectra of TAF2-PC and the temperature of phase transition was lower than that of TAF2 and phospholipid, the decrease in melting point and enthalpy may be due to the decreased crystallinity of the drug (De-vendra et al., 2012).The non-polar part of TAF2 binds to lecithin by hydrogen bonding or van der Waals interaction, which makes TAF2 highly dispersed among the molecules of lecithin, reduces the phase transition temperature of TAF2 and there are no TAF2 drug crystals in the phospholipid complex. Therefore, it is assumed that TAF2 forms a phospholipid complex with lecithin.
3.3.3. Fourier transform infrared spectroscopy (FTIR)
The FTIR spectrum of TAF2 revealed a broad peak at 3,419 cm−1
(Figure 4A), representing the -OH group on the ring structure, and the C-H stretching signal at 2,900 cm−1. The absorption peak near 1,729 cm−1 represents the C=O stretching signal of fatty acid ester, the C=C stretching signal at 1,670 cm−1, and the C-O stretching sig-nal at 1,055 cm−1. These signal peaks are mostly consistent with the structure of diterpenoid rings of quassinoids compounds.
FTIR spectra of lecithin (Figure 4B) showed the characteristic C-H stretching signal of long-chain fatty acids at 2,924 cm−1 and 2,854 cm−1, respectively. The absorption peak of C=O of fatty acid ester at 1,738 cm−1, the absorption peak of P=O at 1,233 cm−1, the absorption peak of P-O-C at 1,087 cm−1 and the absorption peak of -N+(CH3)3 at 970 cm−1 were observed in the spectrum.
The FTIR spectra of the mixture of TAF2 and lecithin (Figure 4C) are essentially superposition of the main absorption peaks of TAF2 and lecithin.
In the FTIR spectrum of TAF2-PC (Figure 4D), the -OH absorp-tion peak of TAF2 changed from 3,419 cm−1 to 3,391 cm−1, the ab-sorption peak of C=C changed from 1,670 cm−1 to 1,678 cm−1, and the absorption peaks at 1,512 cm−1 and 1,153 cm−1 disappeared. The P=O characteristic peak of phospholipids changed from 1,233 cm−1 to 1,228 cm−1, and the C-H stretching signal peak of phospholipids did not change. These results suggested that the phospholipid com-plex is not a simple physical mixture between quassinoids and phos-pholipids, but is bound by intermolecular interaction. Phospholipid complexes may be formed by the interaction between the polar ends of phospholipids and the hydroxyl groups and the carbon-carbon double bonds of the quassinoids.
3.4. Pharmacokinetic studies of TAF2-PC in SD rats
The plasma concentration-time curves of eurycomanone which obtained after single oral administration of TAF2 or TAF2-PC in rats were shown in Figure 5 and the main pharmacokinetic param-eters of eurycomanone were summarized in Table 10. The mean value of the Cmax of eurycomanone in the TAF2-PC group was higher than that in the TAF2 group, 0.50 ± 0.13µg/mL vs 0.27 ± 0.16µg/mL, respectively. Time period to attain the maximum plas-ma concentration (Tmax) of the TAF2-PC group was 0.46 ± 0.29 h,
Table 9. Analysis of variance of quadratic response surface model
Source sum of squares df mean square F-value P-value (prob > F)
Model 1,495.72 9 166.19 185.22 < 0.0001
A-the ratio drug to phospholipid 221.24 1 221.24 246.57 < 0.0001
B-reaction temperature (°C) 6.52 1 6.52 7.26 0.0309
C-drug concentration (mg/mL) 115.14 1 115.14 128.33 < 0.0001
AB 20.07 1 20.07 22.37 0.0021
AC 22.9 1 22.9 25.52 0.0015
BC 5.95 1 5.95 6.64 0.0367
A2 503.13 1 503.13 560.74 < 0.0001
B2 206.83 1 206.83 230.52 < 0.0001
C2 282.86 1 282.86 315.25 < 0.0001
Residual 6.28 7 0.9
Lack of fit 5.78 3 1.93 15.25 0.0118
Pure error 0.5 4 0.13
Total 1,502 16
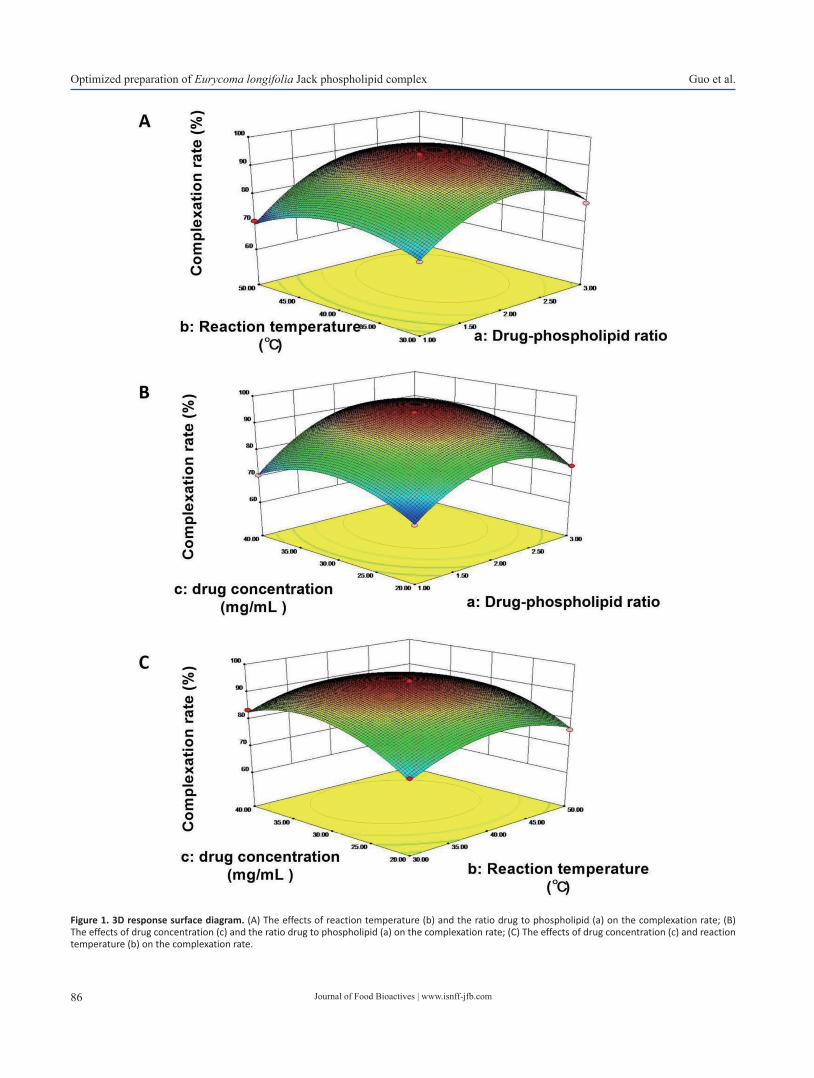
Journal of Food Bioactives | www.isnff-jfb.com86
Optimized preparation of Eurycoma longifolia Jack phospholipid complex Guo et al.
Figure 1. 3D response surface diagram. (A) The effects of reaction temperature (b) and the ratio drug to phospholipid (a) on the complexation rate; (B) The effects of drug concentration (c) and the ratio drug to phospholipid (a) on the complexation rate; (C) The effects of drug concentration (c) and reaction temperature (b) on the complexation rate.

Journal of Food Bioactives | www.isnff-jfb.com 87
Guo et al. Optimized preparation of Eurycoma longifolia Jack phospholipid complex
whereas that of the TAF2 group was 0.32 ± 0.26 h. Tmax was delayed in the TAF2-PC group compared with the TAF2 group, which may be due to the continuous release of the drug from the phospholipid complex or the delayed uptake of the drug by intestinal cells and M cells (Yu et al., 2016). Passive transport was the main absorption mode of eurycomanone. Due to its poor lipid solubility, it is difficult to penetrate the phospholipid bilayer of gastrointestinal cells, result-ing in poor absorption. The phospholipid complex of eurycomanone ameliorated the lipid solubility of TAF2, enhanced the affinity with the cell membrane, thus the membrane absorption of TAF2 was im-proved, and the blood drug concentration in rats was increased. The
improvement of oral bioavailability is the key factor to determine the further application of new drugs in clinical trials. The relative bioavailability of TAF2-PC to TAF2 was 209.20%, suggesting that TAF2-PC improved the absorption of TAF2 and significantly in-creased the relative bioavailability (P < 0.05).
4. Conclusion
The bioavailability is generally determined by the solubility and permeability of the active compounds, except that the P-glycopro-
Figure 2. Scanning electron microscope (SEM) micrographs of (A) TAF2; (B) TAF2-PC.
Figure 3. Differential scanning calorimeter (DSC) pattern of TAF2 (A), lecithin (B), A physical mixture of TAF2 and lecithin (C), and TAF2-PC (D).
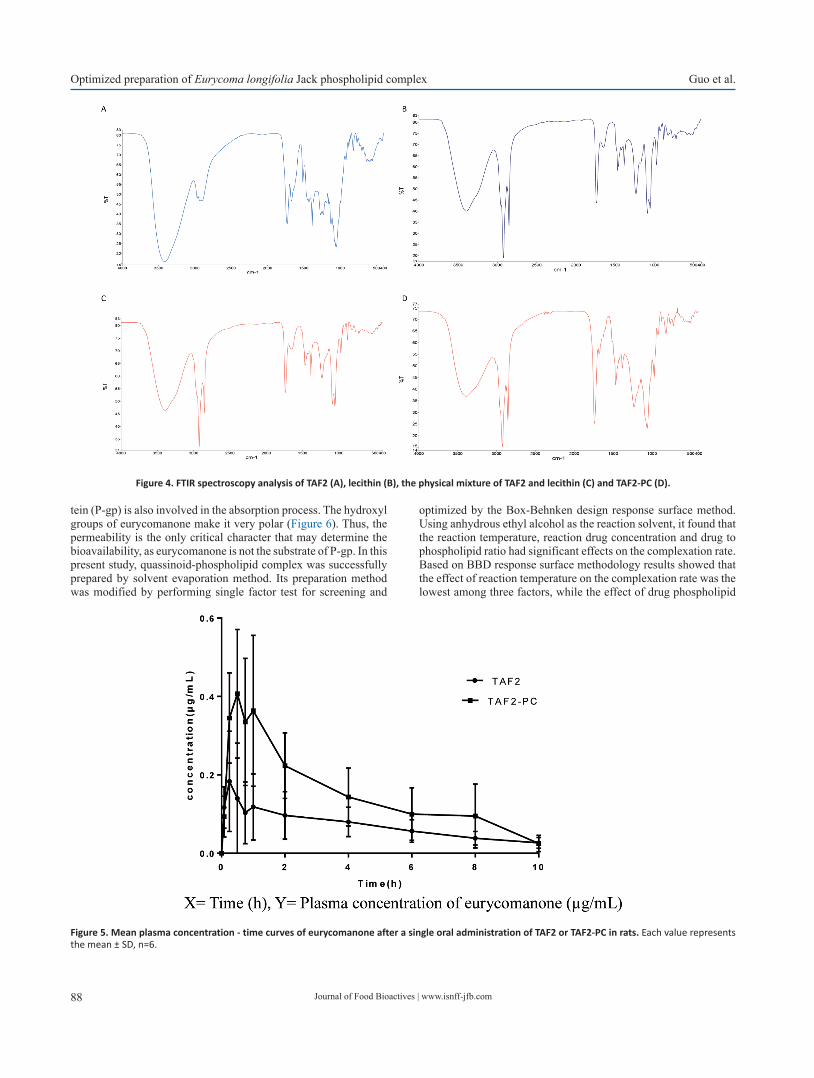
Journal of Food Bioactives | www.isnff-jfb.com88
Optimized preparation of Eurycoma longifolia Jack phospholipid complex Guo et al.
tein (P-gp) is also involved in the absorption process. The hydroxyl groups of eurycomanone make it very polar (Figure 6). Thus, the permeability is the only critical character that may determine the bioavailability, as eurycomanone is not the substrate of P-gp. In this present study, quassinoid-phospholipid complex was successfully prepared by solvent evaporation method. Its preparation method was modified by performing single factor test for screening and
optimized by the Box-Behnken design response surface method. Using anhydrous ethyl alcohol as the reaction solvent, it found that the reaction temperature, reaction drug concentration and drug to phospholipid ratio had significant effects on the complexation rate. Based on BBD response surface methodology results showed that the effect of reaction temperature on the complexation rate was the lowest among three factors, while the effect of drug phospholipid
Figure 4. FTIR spectroscopy analysis of TAF2 (A), lecithin (B), the physical mixture of TAF2 and lecithin (C) and TAF2-PC (D).
Figure 5. Mean plasma concentration - time curves of eurycomanone after a single oral administration of TAF2 or TAF2-PC in rats. Each value represents the mean ± SD, n=6.

Journal of Food Bioactives | www.isnff-jfb.com 89
Guo et al. Optimized preparation of Eurycoma longifolia Jack phospholipid complex
ratio and reaction drug concentration were ranked first and sec-ond, respectively. The optimal preparation process was predicted as follows: drug-phospholipid ratio of 2.28, reaction temperature of 40.85°C, drug concentration of 32.66 mg/mL. The predicted complexation rate was 95.12%. These prediction complexation rate results which was proven to be accurate by the prescription verification experiment. SEM results showed change in morpholo-gy and shape of TAF2-phospolopid complex, which confirmed by the absence of crystallinity of TAF2. The results of DSC showed that the original peaks of TAF2 and lecithin disappeared from the thermal spectrum of TAF2-PC, and the phase transition tem-perature was lower than that of TAF2 and lecithin. The complex demonstrated amorphous characteristics, indicating that TAF2 and lecithin formed phospholipid complex. IR spectra revealed that the phospholipid complex was formed by intermolecular binding of quassinoids and phospholipid molecules, rather than simple mix-ture between them. In summary, it can be confirmed that TAF2 and phospholipids form phospholipid complex. The pharmacokinetics of TAF2-PC and bioavailability of TAF2 were studied in male SD rats. The results showed that the phospholipid complex of eury-comanone significantly enhanced the oral absorption of TAF2, and the relative bioavailability was significantly increased (P < 0.05).
In summary, we successfully prepared the quassinoids-phos-pholipid complex and in vivo study have shown that the develop TAF2-PC significantly improved the oral absorption and bioavail-ability of TAF2 which to overcome the limited absorption of TAF2.
References
Ang, H.H., Hitotsuyanagi, Y., and Takeya, K. (2000). EurycolactonesA–
C, novel quassinoids from Eurycoma longifolia. Tetrahedron Lett. 41(35): 6849–6853.
Chan, K.L., Lee, S.P., Sam, T.W., and Han, B.H. (1989). A quassinoid glyco-side from the roots of Eurycoma longifolia. Phytochemistry 28(10): 2857–2859.
Chan, K.L., Low, B.S., and Ho, D.S.S. (2008). Polar organic extract of eury-coma longifolia Patent.
Darise, M., Kohda, H., Mizutani, K., and Tanaka, O. (1983). Revision of configuration of the 12-hydroxyl group of eurycomanone and eu-rycomanol, quassinoids from eurycoma longifolia. Phytochemistry 22(6): 1514.
Singh, D., Rawat, M.S.M., Semalty, A., and Semalty, M. (2012). Emodin–phospholipid complex. J. Therm. Anal. Calorim. 108: 289–298.
Fiaschetti, G., Grotzer, M.A., Shalaby, T., Castelletti, D., and Arcaro, A. (2011). Quassinoids: From traditional drugs to new cancer therapeu-tics. Curr. Med. Chem. 18(3): 316–328.
Hussein, S., Rusli, I., and Kiong, L. (2007). A summary of reported chemical constituents and medicinal uses of Eurycoma longifolia. J. Trop. Med. Plants 8(10): 103–110.
Jia, D.S., Zhao, J.L., Shi, F., and Jia, X.B. (2010). Preparation of icaritin phy-tosomes and their solid dispersions. Chin. Tradit. Herb. Drugs 41(9): 1449–1453.
Miyake, K., Tezuka, Y., Awale, S., Li, F., and Kadota, S. (2009). Quassinoids fromEurycoma longifolia. J. Nat. Prod. 72(12): 2135–2140.
Kuo, P.C., Damu, A.G., Lee, K.H., and Wu, T.S. (2004). Cytotoxic and anti-malarial constituents from the roots of Eurycoma longifolia. Bioorg. Med. Chem. 12(3): 537–544.
Leng, T.K., Chan, K.L., Sazaly, A.B., Seng, L.B., Ma, H.Q., Fong, W.P., and Davis, K.R. (2015). The in Vitro and in Vivo Anti-Cancer Activities of a Standardized Quassinoids Composition from Eurycoma longifolia on LNCaP Human Prostate Cancer Cells. Plos One 10(3): e121752.
Lian, J., and Wang, B.C. (2006). Research progress in process evaluation standard and application of drug phospholipid complex. Lett. Bio-technol. 017(005): 830–833.
Low, B.S., Ng, B.H., Choy, W.P., Yuen, K.H., and Chan, K.L. (2005). Bioavaila-bility and pharmacokinetic studies of eurycomanone from Eurycoma longifolia. Planta. Med. 71(9): 803–807.
Low, B.S., Teh, C.H., Yuen, K.H., and Chan, K.L. (2011). Physico-chemical ef-fects of the major quassinoids in a standardized Eurycoma longifolia extract (Fr 2) on the bioavailability and pharmacokinetic properties, and their implications for oral antimalarial activity. Nat. Prod. Com-mun. 6(3): 337–341.
Morré, D.J., Grieco, P.A., and Morré, D.M. (1998). Mode of action of the anticancer quassinoids--inhibition of the plasma membrane NADH oxidase. Life Sci. 63(7): 595–604.
Semalty, A., Semalty, M., Singh, D., and Rawat, M. (2012). Phyto-phospho-lipid complex of catechin in value added herbal drug delivery. J. Incl. Phenom. Macro. 73(1-4): 377–386.
Shah, G.R., Chaudhari, M.V., Patankar, S.B., Pensalwar, S.V., Sabale, V.P., and Sonawane, N.A. (2012). Evaluation of a multi-herb supplement for erectile dysfunction: A randomized double-blind, placebo-con-trolled study. BMC Complementary Altern. Med. 12(1): 155.
Tran, T., Malainer, C., Schwaiger, S., Atanasov, A.G., and Stuppner, H. (2014). NF-κB inhibitors from Eurycoma longifolia. J. Nat. Prod. 77(3): 483–488.
Xiao, Y., Song, Y., Chen, Z., and Ping, Q. (2006). The preparation of silybin-phospholipid complex and the study on its pharmacokinetics in rats. Int. J. Pharmaceut. 307(1): 77–82.
Yu, F., Li, Y., Chen, Q., He, Y., Wang, H., Yang, L., and Xue, M. (2016). Monodisperse microparticles loaded with the self-assembled ber-berine-phospholipid complex-based phytosomes for improving oral bioavailability and enhancing hypoglycemic efficiency. Eur. J. Pharm. Biopharm. 103: 136–148.
Zakaria, Y., Rahmat, A., Pihie, A.H., Abdullah, N.R., and Houghton, P.J. (2009). Eurycomanone induce apoptosis in HepG2 cells via up-regu-lation of p53. Cancer Cell Int. 9(1): 16.
Zhou, C., Xia, X., Liu, Y., and Lin, L. (2012). The preparation of a complex of insulin-phospholipids and their interaction mechanism. J Pept Sci. 18(9): 541–548.
Table 10. Pharmacokinetic parameters after oral administration of TAF2 or TAF2-PC in rats
Parameters TAF2 TAF2-PC
Cmax(μg/mL) 0.27 ± 0.16 0.50 ± 0.13
Tmax(h) 0.32 ± 0.26 0.46 ± 0.29
AUC0–∞((μg/mL)·h) 0.87 ± 0.22 1.82 ± 1.08
Data are presented as mean ± standard deviation (SD), n = 10.
Figure 6. Chemical structure of Eurycomanone.

90
Meeting Report
1
Consumer behaviour and healthy diet: a challenge or an opportunity?Fereidoon Shahidi
Healthy food choices by consumers are dictated by several factors but most would like healthy and affordable supply.
Perspectives
3
Is the gluten-free and casein-free diet efficacious in the treatment of childhood autism spectrum disorder?Klaus W. Lange and Andreas Reissmann
Autism spectrum disorder (ASD) is a set of heterogeneous neurodevelopmental conditions, characterized by early-onset difficulties in social communication as well as repetitive and unusually restricted behaviors and interests.
13
The use of vitamin D in depressionKlaus W. Lange, Katharina M. Lange, Yukiko Nakamura and Andreas Reissmann
Vitamin D has increasingly been associated with the pathophysiology of mental illness and has been suggested to have beneficial effects on depression in adults.
Reviews
21
Phytochemicals: Potential Lead Compounds for COVID-19 TherapeuticsSrishti Kashyap, Revathy Nadhan and Danny N. Dhanasekaran
Journal of Food Bioactives
The Journal of Food Bioactives (JFB), a publication of the International Society for Nutraceuticals and Functional Foods (ISNFF), aims to bring together the results of fundamental and applied research on food bioactives, functional food ingredients, nutraceuticals and natural health products that are known to possess or perceived to have health-promoting properties.

91
Coronavirus Disease 2019 (COVID-19) is a global pandemic, caused by severe acute respiratory syndrome coronavirus 2 (SARS- CoV-2).
29
Impact of dietary nutrients (functional foods/nutraceuticals) and micronutrients on COVID-19: a reviewYueching Wong, Chi-Ho Chan, Kamesh Venkatakrishnan, Hui-Fang Chiu, You-Cheng Shen, Oksana Glovinskaia, Yi-Chun Han and Chin-Kun Wang
The world health organization (WHO) announced that coronavirus 2019 (COVID-19) is a pandemic.
39
Phenolic compounds in cereal grains and effects of processing on their composition and bioac-tivities: a reviewFereidoon Shahidi, Renan Danielski and Chiaki Ikeda
Cereals are a staple food in the diets of many populations globally.
51
Potential adverse health ef-fects of dietary lipid oxidation productsYuanyuan Hu, Guanhua Zhao, Min Zhang, Dayong Zhou and Beiwei Zhu
Lipid oxidation products (LOPs) are widely present in many lipid- con-taining foods.

92
63
Functional foods beyond nutri-tion: therapeutic interventions to combat COVID-19 and other viral diseasesSamudra P. Banik, Rituparna Banik Ghosh, Bernard W. Downs, Sanjoy Chakraborty, Manashi Bagchi, Tandra R. Chakraborty and Debasis Bagchi
SARS-COV2 has emerged as the second biggest threat to humanity of all times, after the Spanish Flu massacre.
Original Research
74
Redistribution of nutritionally important starch fractions ben-eficial to diabetic population by using oats as a functional in-gredient in a wheat-based food product – JereeshFaiyaz Ahmed
Oats being important sources of β-glucan were incorporated at 10, 20 and 30% level in a wheat-based traditional Arabian breakfast cereal called Jereesh and evaluated for sensory acceptability.
80
Optimized preparation of quassi-noid-rich Eurycoma longifolia Jack phospholipid complex and its effects on oral bioavailabilityWu-Yan Guo, Shu-Yan Wang, Jian Ren and Bo Zhang
Eurycoma longifolia Jack is a plant of the genus Eurycoma in Simarou-baceae. Eurycomanone is one of the main quassinoids of E. longifolia.


Table of Contents
Journal of Food Bioactives
Meeting ReportConsumer behaviour and healthy diet: a challenge or an opportunity?
Fereidoon Shahidi . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1
PerspectivesIs the gluten-free and casein-free diet efficacious in the treatment of childhood autism spectrum disorder?
Klaus W . Lange and Andreas Reissmann . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3
The use of vitamin D in depressionKlaus W . Lange, Katharina M . Lange, Yukiko Nakamura and Andreas Reissmann . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13
ReviewsPhytochemicals: Potential Lead Compounds for COVID-19 Therapeutics
Srishti Kashyap, Revathy Nadhan and Danny N . Dhanasekaran . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21
Impact of dietary nutrients (functional foods/nutraceuticals) and micronutrients on COVID-19: a reviewYueching Wong, Chi-Ho Chan, Kamesh Venkatakrishnan, Hui-Fang Chiu, You-Cheng Shen, Oksana Glovinskaia, Yi-Chun Han and Chin-Kun Wang . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 29
Phenolic compounds in cereal grains and effects of processing on their composition and bioactivities: a review
Fereidoon Shahidi, Renan Danielski and Chiaki Ikeda . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 39
Potential adverse health effects of dietary lipid oxidation productsYuanyuan Hu, Guanhua Zhao, Min Zhang, Dayong Zhou and Beiwei Zhu . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 51
Functional foods beyond nutrition: therapeutic interventions to combat COVID-19 and other viral diseasesSamudra P . Banik, Rituparna Banik Ghosh, Bernard W . Downs, Sanjoy Chakraborty, Manashi Bagchi, Tandra R . Chakraborty and Debasis Bagchi . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 63
Original ResearchRedistribution of nutritionally important starch fractions beneficial to diabetic population by using oats as a functional ingredient in a wheat-based food product – Jereesh
Faiyaz Ahmed . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 74
Optimized preparation of quassinoid-rich Eurycoma longifolia Jack phospholipid complex and its effects on oral bioavailability
Wu-Yan Guo, Shu-Yan Wang, Jian Ren and Bo Zhang . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 80




















