
JBLS: Vol. 5, No. 2, August 2014, published
202
-
Upload
macrothink -
Category
Documents
-
view
0 -
download
0
Transcript of JBLS: Vol. 5, No. 2, August 2014, published


Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
http://jbls.macrothink.org
Editorial Team
Editorial Assistant Guiju Yang, Macrothink Institute, United States
Associate Editors
Weichao Guo, Stowers Institute for Medical Research, United States Dr Lakshmi Chandramohan, Baylor College of Medicine/ Texas Children's Hospital, United States
Editorial BoardAdeline Chia, Taylor's University, Malaysia Ali Akbar Ghanbari, Seed and Plant Improvement Research Institute (SPII), Iran, Islamic Republic of Anand Minajigi, Harvard Medical School, Massachusetts General Hospital, United States Dr. Anirban Mukherjee, The University of Texas at Austin, United States Dr Antonio Andrade-Torres, INBIOTECA, Universidad Veracruzana, Mexico Dr. Bharat Jagannathan, University of California, Berkeley, United States Buxin Chen, University of California, San Diego, United States DR. Chien-I Chang, UMass Medical School, United States Danqi Chen, Pennsylvania state university, United States Dr. Devaveena Dey, Brigham & Women's Hospital Harvard Medical School, United States DR DEVENDRA KUMAR RAI, University of Missouri-Columbia, United States Dr D M Firake, ICAR Research Complex for NEH Region, Meghalaya, India Ewa Moliszewska, Opole University, Poland Fazal Shirazi, MD Anderson Cancer Center,, United States Dr. Fernando Cardona, Institute of Biomedicine of Valencia Spanish National Research Council (C.S.I.C.), Spain Geonyzl Lepiten Alviola, Davao Doctors College, Philippines Gizele Ingrid Gadotti, Oregon State University Universidade Federal de Pelotas, United States Mr. GNANARAJA RAJARETHINAM, University Allahabad, India Mr. GODSWILL NTSOMBOH NTSEFONG, Institute of Agricultural Research for Development (IRAD), Cameroon Dr Gokul Babarao Kale, Department of Zoology, G.S. Science, Arts and Commerce College, India Haiyan Li, University of Texas MD Anderson Cancer Center, United States Dr. Hari Kishore Annavarapu, University of Texas Southwestern Medical Center, United States Dr Helen Evelyn Malone, School of Nursing & Midwifery Trinity College, Dublin 2, Ireland Dr. Hongbo Pang, Sanford-Burnham Medical Research Institute, United States Hong Zhao, The Pennsylvania State University, United States professor hossam el-din Mohamed omar, Assiut University, Egypt Dr. Hui Xiang, Allergan Inc, United States

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
http://jbls.macrothink.org
Ivo Vaz Oliveira, Minho University, Braga, Portugal Dr Jean-Luc Desseyn, Inserm U995 University Lille 2, France Jianfeng Li, University of Pittsburgh, United States Dr. Jinhua Wu, Torrey Pines Institute for Molecular Studies, United States Jittima Charoenpanich, Department of Biochemistry, Faculty of Science, Burapha University Loretta Gratani, Department of Environmental Biology, Sapienza University of Rome Lucélia Cabral, University of Campinas - Unicamp, Brazil Marcos Taveira, University of Porto, Portugal MELTEM SESLİ, College of Tobacco Expertise, Celal Bayar University, Turkey Mohamed Ahmed El-Esawi, Botany Department, Faculty of Science, Tanta University, Egypt Mr Muhammad Ali, COMSATS Institute of Information Technology, Pakistan Dr. Mirza Hasanuzzaman, Department of Agronomy, Faculty of Agriculture, Sher-e-Bangla Agricultural University, Dhaka, Bangladesh Doctor Nayana tara Arunkumar, Kasturba Medical College, India Othman El Mahdy, Cell Biology Department National Research Center Dokki, Egypt, Egypt Mr Palmiro Poltronieri, Institute of Sciences of Food Productions, Italy Pramod K Mishra, University of Texas at San Antonio, United States Prashanth Chandramani Shivalingappa, Iowa State University, United States Dr Preeti Viswanathan, Children's hospital at Montefiore, Albert Einstein College of Medicine Dr. Rajeshwari Valiathan, Wayne State University School of Medicine, United States Ravi Kant, Panjab University, Chandigarh, India Dr Samir AYDI, University of Gabes, Tunisia Dr Sankaranarayanan Srinivasan, Baylor College of Medicine, United States Dr Sarita Prabhakaran, King Faisal Specialist Hospital & Research Centre, Riyadh, KSA, Saudi Arabia Dr. Senthilkumar Sivagurunathan, Boston Children's Hospital/Harvard Medical School, United States Doctor Sergey Alexandrovitch Shukolyukov, Sechenov Institute of Evolutionary Physiology & Biochemistry of the RAS, Russian Federation Srikanth Reddy Medipally, Universiti Putra Malaysia, Malaysia Dr. Subramanian Krishnan, Children's Hospital Los Angeles, University of Southern California., United States Dr. Sumanta Mukherjee, University of Michigan, United States Tiaojiang Xiao, Niddk, National Institute of Health, United States Ujwal S Patil, University of Ne Orleans, United States Uksha Saini, Ohio State University, United States Dr. Waldiney Mello, Centro Universitário Celso Lisboa, Brazil Mr. Weibin Liu, Mayo Clinic College of Medicine, United States Yuan Sun, Stanford University, United States Dr. Zeynep Ahsen Kocer, St Jude Children's Research Hospital, United States Dr Zhen Shen, The Rockefeller University, United States, United States

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
http://jbls.macrothink.org I
Contents
Preventive Effect of Tephrosia Purpurea Against N,N-Diethylnitrosamine Induced Hepatocellular Carcinoma In Swiss Albino Mice
R. Gnanaraja, Veeru Prakash 1-9
Parasitic Burden and Pathologic Effects of Anisakis Sp. (Nematoda : Anisakinae, Anisakidae) and Contracaecum Sp. Larvae (Nematoda : Anisakinae, Anisakidae) on Mugilids from Senegalese Estuaries
Ephigénie Ndew DIONE, Malick DIOUF, Alassane SARR, Jean FALL, Cheikh Tidiane BÂ 10-24
Evaluating the Nutritional Potential of Acacia Sieberiana Seeds (Dc) Growing In North West of Nigeria
Abubakar Salisu, Godwin H. Ogbadu, Paul C. Onyenekwe, Omotoye Olorode, Rebecca W. Ndana, Olayanju Segun
25-36
Effects of Occupational Exposure to Wood Smoke in Tandoor Occupants
Ghulam Nabi, Jeena Urooj, Ayaz Ali Khan, Ghazala Yasmin Zamani, Mussarat Wahid, Saleem Ur Rahman
37-47
The Effects of Long Term Sleep and Exercise Deprivation on Total Serum Testosterone in Male Professional Drivers
Ghulam Nabi, Muhammad Amin, Amjad Khan, Rahim Ullah, Muhammad Shuaib, Ayaz Ali Khan 48-57
Effect of Fetal Sex on Total Levels of Maternal Serum Testosterone
Ghulam Nabi, Tariq Aziz, Muhammad Amin, Ayaz Ali Khan 58-64
Link between Chronic Pesticides Exposure and Reproductive Problems in Male Farmers
Ghulam Nabi, Muhammad Amin, Tabinda Rauf, Khwaja Muhammad Khan, Ayaz Ali Khan 65-76
In Vitro Antibacterial Activity of Two Mosses: Calymperes erosum C. Mull and Bryum coronatum Schwaegr from South-Western Nigeria
Tedela P. O., Adebiyi A. O., Aremu A., David O. M. 77-84
Phytohormones Influence on the In-Vitro Regeneration of Saccharum Officinarum L. from Apical Meristem Culture
Ejiroghene Felix Lawyer, Z. O. Jamaleddine, P. T. Lyam, I. T. Borokini, A. A. Adedeji, A. U. Okere
85-94
The Impact of Wood, Cigarette and Marijuana Smoke on the Reproductive Health of Tandoor Occupants
Ghulam Nabi, Muhammad Amin, Jeena Urooj, Muhammad Kamil, Ayaz Ali Khan 95-105
Reproductive Biology and Bioturbatory Activities of Two Sympatric Species of Fiddler Crabs Uca lactea annulipes and Uca triangularis bengali (Decopada : Ocypodidae) at the East Midnapore Coastal Belt of West Bengal, India
Subhasish Chatterjee, Tapas Kumar Das, Susanta Kumar Chakraborty 106-127
Insects Associated With Tea and Their Identification at Ntri (Shinkiari) Mansehra, Pakistan

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
http://jbls.macrothink.org II
Hamid Ullah, Ibrar Muhammad, Waheed Ullah, Farzana Parveen, Sohail Aslam 128-134
Root Endophytic Fungi: Research Update
Neha Chadha, Manjita Mishra, Ram Prasad, Ajit Varma 135-158
Methanol Extract of Barringtonia asiatica, Kurz. Quite Effective Induce Mortality Of Spodopthera litura, Fabr. Larvae on soybean plants
Meity N Tanor, Abdul Latief Abadi, Bambang Tri Raharjo, Jantje Pelealu 159-164
Environmental Factors and Puberty Onset: An Update
Ghulam Nabi, Muhammad Amin, Rishma Sultan, Muhammad Kamil 165-174
Eco-Diversity of Edible Insects of Nigeria and Its Impact on Food Security
Adeoye Olubusola Temitope, Oyelowo Oyetayo Job, Adebisi-Fagbohungbe Tola Abiodun, Akinyemi Olukayode Dare
175-187
Profile of Catfish (Clarias sp) Oocyte Exposed by Laserpuncture
Dyah Hariani, Marhendra A. P. W, Aulanni Am, Suprayitno E 188-195

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 1
Preventive Effect of Tephrosia Purpurea Against N,N-Diethylnitrosamine Induced Hepatocellular
Carcinoma In Swiss Albino Mice
R. Gnanaraja (Corresponding Author)
Department of Biochemistry and Biochemical Engineering
SHIATS (Formally Allahabad Agricultural Institute Allahabad) U.P India
Tel: 91-896-027-7743 E-mail: [email protected]
Veeru Prakash
Associate Professor, Department of Biochemistry and Biochemical Engineering
SHIATS (Formally Allahabad Agricultural Institute Allahabad) U.P India
Tel: 91-983-995-2882 E-mail: [email protected]
Received: January 21, 2014 Accepted: February 3, 2014
doi:10.5296/jbls.v5i2.5276 URL: http://dx.doi.org/10.5296/jbls.v5i2.5276
Abstract
The aim of the present study is to divulge the preventive nature of Tephrosia purpurea Methanolic Extract (TpME) during DiEthylNitrosAmine (DENA) induced liver cancer in male Swiss albino mice. Anticancer activity of methanolic extract of Tephrosia purpurea was evaluated in DENA induced Swiss albino mice at the doses of 200 and 300 mg/kg body weight orally and both doses were administered for 14 days. After 24 h of the last dose, the mice were sacrificed and antitumor effect of TpME was assessed by evaluating liver serum marker enzyme and haematological parameters. Decrease in Aspartate transaminase, Alanine transaminase, Alkaline phosphatase, Bilirubin and Total protein count were observed in TpME treated animals when compared to DENA treated animals. Haematological profiles such as haemoglobin, red blood cell (RBC) and white blood cell (WBC) count reverted to normal level in TpME treated mice. A comparative histopathology study of liver from test group exhibited almost normal architecture, as compared to DENA treated group. The results demonstrated that Tephrosia purpurea methanolic extract at 300 mg/kg dose exhibits

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 2
significant antitumor activity than 200 mg/kg.
Keywords: Diethylnitrosamine, Tephrosia purpurea, Liver cancer, Haematology and carcinogenesis
1. Introduction
Cancer is a complex genetic disease that is caused primarily by environmental factors. The cancer-causing agents (carcinogens) can be present in food and water, in the air, and in chemicals and sunlight that people are exposed to. The cancer becomes the second major cause of death in the human after cardiovascular disease (Gruddy, 1991). Therefore, there is a continuing search for better control and preventive methods in order to reduce cancer mortality and related side effects. Many investigations are now being carried out to discover naturally occurring compounds, which can suppress or prevent the process of carcinogenesis (Wargovich., et al 1988 and Samaranayake et al., 2000).
Medicinal plants play a significant role in providing primary health care services to rural people. They serve as therapeutic agents as well as important raw materials for the manufacture of traditional and modern medicine. Tephrosia purpurea is a species of flowering plant in the pea family, Fabaceae, that has a pantropical distribution. It is used in the treatment of some major diseases. The antioxidant and anticancer activity of the extracts from medical plants and herbs was associated with their components of phenolic compounds; the major types of phenolic compounds included phenolic acids, flavonoids, tannins, coumarins, lignans, quinones, stilbenes, and curcuminoids (Cai et al., 2004).
In view of this, we have evaluated the preventive effect of TpME against hepatocellular carcinoma in Swiss Albino Mice. The discovery and development of chemotherapeutics that are effective for the treatment and control of liver cancer is urgently needed for present hour.
2. Material and Methods
2.1 Animals
Male Swiss albino mice strain approximately 8 weeks young mice (weighing approximately 22-25g) were used in this study. The healthy animals were procured from United Institute of Pharmacy, Allahabad, Uttar Pradesh India. The animals were housed in spacious polypropylene cages bedded with rice husk. The animal room was well ventilated and maintained under standard experimental conditions (temperature 27±2ºC and 12 hours light / dark cycle) throughout the experimental period. All the animals were fed with standard pellet diet and water ad libitum. The animals were acclimatization to the environment for 7days prior to experimental use. The experiment was carried out according to the guidelines of the Committee for the Purpose of Control and Supervision of Experiments on Animals (CPCSEA), New Delhi, India.
2.2 Plant Materials and Preparation of Plant Extract
The Tephrosia purpurea plant was collected from Yamuna river bank, Allahabad District, in Uttar Pradesh, India. The collected plant materials were washed, sliced and completely dried in

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 3
a hot-air oven at 37°C. The dried materials was ground into make a fine powder and used for extraction. Two hundred grams (200g) of the powered plants were extracted with methanol (80%) using “Soxhlet Apparatus” for 48 hours. A semi solid extract was obtained after complete elimination of alcohol under reduced pressure. The extract was stored in refrigerator until used. The extract contains both polar and non-polar phytocomponents. For experiments 200mg/kg and 300mg/kg body weight of TpME were used. This effective dose was selected based on dose dependent studies of TpME carried out in our laboratory.
2.3 Experimental Design
Body weights of the animals were recorded and they were divided into 6 groups. Each group have 6 animals and the details as follow.
Group 1 (Positive Control)
Normal control mice will be fed with standard diet and served as a control, which received saline.
Group 2 (Negative Control)
Mice induced with hepatocellular carcinoma by providing DENA 175 μl /kg/days for a week through intra peritoneal injection
Group 3
Mice induced with hepatocellular carcinoma by providing DENA 175 μl /kg/day for 7 days through intra peritoneal injection with methanolic extract of Tephrosia purpurea oral gavage daily at the dose of 200 mg/kg for 14 days.
Group 4
Mice induced with hepatocellular carcinoma by providing DENA 175 μl /kg/day for 7 days through intra peritoneal injection with methanolic extract of Tephrosia purpurea oral gavage daily at 300 mg/kg body weight for 14 days.
Group 5
Alone methanolic extract of Tephrosia purpurea oral gavage daily at a different dose of 200 mg/kg body weight for 14 days.
Group 6
Alone methanolic extract of Tephrosia purpurea oral gavage daily at a different dose of 300 mg/kg body weight for 14 days.
2.4 Collection of Samples and Hemato & Biochemical Assay
On completion of the experimental period, animals were anaesthetized with diethyl either (2ml/kg). The blood was collected with and without EDTA as anticoagulant. Blood, plasma for the estimation Hemoglobin (Armour et al., (1965) content, Red blood cell count (Wintrobe et al., (1961) and White blood cells (Dacie and Lewis., (1958) and serum were separated for the

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 4
estimation of various biochemical parameters such as Bilirubin, Asparate amino transaminase (IFCC method., 1986), Alanine amino transaminase (IFCC method., 1986) and Alkaline phosphatase (King and Armstrong., 1934) and Total Protein (Lowry et al., 1951) were determined.
2.5 Histopathology Studies
The liver were excised from the experimental animals of each group after collecting the blood sample and washed with normal saline. Initially, the materials were fixed in 10%neutral formalin solution. Sections were cut using microtome techniques after paraffin embedding. The sections were possessed in alcohol xylene series and were stained with hematoxylin and eosin. The different sections were examined microscopically for the evaluation of histopathological cha nges.
2.6 Statistical Methods
All values are expressed as mean SEM. Statistical significance was determined by two-way analysis of variance (ANOVA) and the difference were considered significant at p <0.05 level.
3. Results
Recently, the use of some herbs has attracted a great deal of attention as one of the alternative cancer therapies from the point of less toxicity and cost benefits. Therefore, an attempt has been made to evaluate the anticancer activity of methanolic extract of Tephrosia purpurea.
3.1 Effect of Methanolic Extract of Tephrosia Purpurea on Hematological Parameters
The hematological parameters of tumor bearing mice were found to be significantly differences compared to the extracts treated groups. In tumor bearing mice it was found that increased in WBC count (15.1±0.21) and decreased in HB content (7.78±0.04) with RBC count (2.88±0.10 106). The methanolic extract treated group at dose of 200 mg/kg and 300 mg/kg body weight restored all the altered hematological parameters to almost near normal (Table 1). All these results suggest the anticancer nature of the extract, the dose of 300 mg/kg body weight was found to be effective potential than 200 mg/kg body weight.
Table 1. Effect of methanolic extract of Tephrosia purpurea on the hematological parameters in Swiss albino mice
Design of Treatment Hemoglobin (gm %) WBC (103 cells/mm3)RBC
(106 cells/mm3)
Normal 13.45 0.09 8.8 0.07 5.23 0.03
Tumor Control DENA (175ul/kg)
7.78 0.04 15.1 0.21 2.88 0.10
DENA (175ul/kg) + TPME 200 mg/kg
9.38 0.21* 13.12 0.15* 3.39 0.17*
DENA (175ul/kg) + TPME 300 mg/kg
11.02 0.04* 10.31 0.08* 4.63 0.06*
TPME 200 mg/kg 13.11 0.06* 8.67 0.10* 5.22 0.02*

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 5
TPME 300 mg/kg 13.17 0.04* 8.73 0.04* 5.29 0.03*
Values are expressed as mean S.E.M., (n = 6)
*P < 0.05 as compared to tumor control group (Two-way ANOVA).
3.2 Effect of Methanolic Extract of Tephrosia Purpurea on Liver Enzyme and Histopathology Studies
The clinical liver parameters of tumor bearing mice were found to be significantly differences compared to the extracts treated groups. In tumor bearing mice it was found that increased in Bilirubin, ALT, AST, ALP and Total Protein. The methanolic extract treated group at dose of 200 mg/kg and 300 mg/kg body weight restored all the altered Clinical liver enzymes are almost near normal (Table 2). All these results suggest the anticancer nature of the extract, the dose of 300 mg/kg body weight was found to be potential activity than 200 mg/kg body weight. Histopathology study of liver in each group of animals given Fig-1.
Table 2. Effect of methanolic extract of Tephrosia purpurea on the Clinical parameters in Swiss albino mice
Treatments Billrubin (mg/dl) ALT (U/L) AST (U/L) ALP (U/L) T. Protein (g/dl)
Normal 0.77 0.02 37.3 0.33 32.67 0.21 74.33 0.49 7.16 0.10
Tumor Control DENA (175ul/kg)
1.93 0.04 100.17 0.70 91.5 0.22 188.33 1.42 9.98 0.18
DENA (175ul/kg) + TPME 200 mg/kg
1.75 0.03* 86.17 0.40* 74.5 0.22* 150 0.51* 9.13 0.09*
DENA (175ul/kg) + TPME 300 mg/kg
1.48 0.03* 62.5 0.22* 59.50 0.22* 135.5 1.78* 8.32 0.06*
TPME 200 mg/kg
0.78 0.02* 37 0.26* 31.67 0.21* 72.17 0.30* 7.17 0.05*
TPME 300 mg/kg
0.80 0.02* 34 0.37* 28.83 0.17* 62.5 0.22* 7.48 0.08*
Values are expressed as mean S.E.M., (n = 6)
*P < 0.05 as compared to tumor control group (Two-way ANOVA).

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 6
Figure 1. photomicrograph of liver (10x) (T1) control group showing normal architecture of liver; T2 animal treated with DENA showing cancer cell developed; (T3) animal treated with DENA+200mg/kg of TPME showing less cancer cell compared to T2; (T4) animal treated with DENA+300mg/kg of TPME showing very less cancer cell compared to cancer control T2; (T5&T6) animal treated with alone 200mg/kg & 300mg/kg of TPME showing normal architecture of liver compared to T1.
4. Discussion
It is well known that liver cancer is one of the most important cancers in the world, resulting in more than 1 million patients and over 260,000 deaths per year (Liu et al., 2006). Therefore, the chemoprevention and treatment of liver cancer is very important. Liver is the major site of metabolism of ingested components; it is highly susceptible to carcinogenic effect. Natural products have long been used to prevent and treat many diseases, including cancer and thus they are good candidates for the development of anti-cancer drugs. The large population use ayurvedic medicine worldwide, there is urgent need for additional, carefully conducted, high quality intensive research to evaluate its efficacy and to develop this discipline to meet ever-new challenges of modern medicine in the field cancer research. Different in vivo and in vitro screening models are available for anticancer activity (Vishal and Thangavel, 2011).
The result of present study showed anticancer effects of TPME leaves against DENA in Swiss albino mice. A significant enhancement of TPME of haemoglobin, red blood cells and decrement of liver marker enzymes and WBC. The proposed study revealed that the fractions

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 7
obtained from TPME plant potent in inhibiting the cancer growth.
Hepatocellular carcinoma is the most frequent primary malignancy of the liver. DENA widely used for studying the mechanism of Hepatocellular carcinoma in animal experimental model (Stelzner, 2006). Tephrosia Purpurea are rich in flavonoid content and the cytotoxic might be due to the presence of flavonoids in the fractions utilized in this study as flavonoids have been shown to possess antimutagenic and antimalignant effect (Hirano et al., 1989; Brown, 1980). The flavonoids have a chemopreventive role in cancer through their effects on signal transduction in cell proliferation and angiogenesis (Osman et al., 2011; Fotis et al., 1997; Weber et al., 1996). Our results suggests that the fractions of extracts obtained in from TPME have good cytotoxic activity against DENA induced liver cancer, which may be attributed to the secondary metabolites and antioxidant of the fractions.
5. Conclusion
The methanolic extracts of Tephrosia purpurea showed good potent anticancer activity against n,n-diethylnitrosamine induced hepatocellular carcinoma in Swiss albino. Further studies are in progress in our laboratory in screening of phytochemicals and antioxidant and investigation of molecular mechanisms and responsible for the anticancer activity of this plant. These results will help in the efficient cancer treatments and that would be more beneficial to our social and economic aspects of world.
Acknowledgement
The authors would like to thank the Sam Higginbottom Institute of Agriculture, Technology and sciences & United Institute of Pharmacy, Allahabad, India for providing the laboratory facilities to carry out the work.
Reference
Bergmeyer, H., Bernt, E., Varley, H., Gowenlock, A. H, & Bell, M. (1980). Practical Clinical Biochemistry, 5th Edn., Willian Heinmann Medical Books Ltd, London; pp 741.
Brown, J. P. (1980). A review of the genetic effect of naturally occurring flavonoids, anthraquinones and related compounds. Mutat Res, 75, 243-247. http://dx.doi.org/10.1016/0165-1110(80)90029-9
Cai, Y., Luo, Q., Sun, M., & Corke, H. (2004). Antioxidant activity and phenolic compounds of 112 traditional Chinese medicinal plants associated with anticancer. Life Sci, 74, 2157–2184. http://dx.doi.org/10.1016/j.lfs.2003.09.047
D’Armour F. E., Blood F. R., & Beldena, D. A. (1965). The manual for laboratory work in mammalian physiology. 3rd ed. Chicago: The University of Chicago Press. p. 4-6.
Dacie, J. V., & Lewis, S. M. (1958). Practical hematology. 2nd ed. London: J and A Churchill; p. 38-48.
Fotis, T., Pepper, M. S., Aktas, E., Breit, S., Rasku, S., & Adlercreutz, H. (1997). Flavonoids, dietary-derived inhibitors of cell proliferation and in vitro angiogenesis. Cancer Res, 57,

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 8
2916-2921.
Gruddy, S. M. (1991). Recent Nutrition Research, implications for foods of the future. Ann Med, 23, 187-193. http://dx.doi.org/10.3109/07853899109148046
Hirano, T. K., Oka, M., & Akiba, M. (1989). Antiproliferative effect of synthetic and naturally occurring flavonoids on tumour cells of human breast carcinoma cell lines, ZR-75-1. Res Commun Chem Pathol Pharmacol, 64, 69-78.
IFCC methods (1986). The measurement of catalytic concentration of enzymes, J. Clin. Chem. Clin Biochem, 24, 481.
IFCC methods for the measurement of catalytic concentration of enzymes, J. Clin. Chem. Clin Biochem, 24, 494-97.
King, E. J., Armstrong, A. R., & Harris, R. I. (1934). Phosphatase in obstructive jaundice. Canad. M. A J. J, 31, 14.
Liu, J. G., Zhao, H J., Liu, Y. J., & Wang, X. L. (2006). Effect of selenium-enriched malt on hepatocarcinogenesis, paraneoplastic syndrome and the hormones regulating blood glucose in rats treated by diethylnitrosamine. Life Sci. 78(20), 2315-2321. http://dx.doi.org/10.1016/j.lfs.2005.09.033
Lowry, O. M., Rosebrough, N. J., Farr, A. L., & Randall, R. J. L. (1951). Protein measurement with folin phenol reagent. J. Biol. Chem, 193, 265-275.
Osman, M. A., Rashid, M. M., Aziz, M. A., Habib, M. R., & Karim, M. R. (2011). Inhibition of Ehrlich ascites carcinoma by Manilkara zapota L. stem bark in Swiss albino mice. Asian Pac J Trop Biomed, 1(6), 448-451. http://dx.doi.org/10.1016/S2221-1691(11)60098-1
Samaranayake, M. P., Wickramasinghe, S. N., Angunawela, P., Jayasekara, S., Iwai, S., & Fukushima, S. (2000). Inhibition of chemically induced liver carcinogenesis in Wistar rats by garlic (Allium sativum). Phytotherapy Research, 14, 1-3. http://dx.doi.org/10.1002/1099-1573(200011)14:7<564::AID-PTR664>3.0.CO;2-Z
Stelzner, F. (2006). (Autoregulatory growth control of a DEN Aomatous polyps and carcinogenesis in the colorectal region. Basics of tumor surgery Part 1). Chirurg, 77, 1048-1055. http://dx.doi.org/10.1007/s00104-006-1258-4
Thirunavukkarasu, C., Premkumar, K., Jagadeeswaran, R., & Sakthisekaran, D. (2005). The inhibitory effect of sodium selenite on Nnitrosodiethylamine- induced and phenobarbital promoted liver tumourigenesis in rats based on the modulation of polyamine levels. Mol. Cell Biochem, 280(1-2), 165-172. http://dx.doi.org/10.1007/s11010-005-8907-7
Thiruvengadam, D., & Mangalamani (2011). Daisy Glory, Induction of Hepatic Antioxidant Enzyme by Chrysin in N- Nitrosodiethylamine induced Hepatocellular carcinoma in Wiser albino rats. Int.J. Res. Pharm. Sci, 2(2), 266-271.
Varley, H., Gowenlock, A. H., & Bell, M. (1980). Practical Clinical Biochemisty, 5th Edn.,William Heinmann Medical Books Ltd, London 1980; 897.

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 9
Vishal, G, & Thangavel S. (2011). Anticancer activity of Tephrosia purpurea and Ficus religiosa using MCF 7 cell lines. Asian Pacific Journal of Tropical Medicine, 54, 526-529.
Wargovich, M. J., Woods, C., & Eng, V. W. S (1988). Chemoprevention of NNitrosomethyl benzyllamine-induced oesophageal cancer in rats by the naturally Cancer thiother, diallyl sulphide. Cancer Res, 48, 6872-6875.
Weber, G., Shen, F., Prajda, N., Yeh, Y. A., & Yang, H. (1996). Increased signal transduction activity and down regulation in human cancer cells. Anticancer Res, 16, 3271-3282.
Wintrobe, M. M., Lee, G. R., Boggs, D. R., Bithel, T. C., Athens, J. W., & Foerester (1961). Clinical Hematology 5th ed. Philadelphia;. p. 326.
Copyright Disclaimer
Copyright reserved by the author(s).
This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 10
Parasitic Burden and Pathologic Effects of Anisakis Sp.
(Nematoda : Anisakinae, Anisakidae) and
Contracaecum Sp. Larvae (Nematoda : Anisakinae,
Anisakidae) on Mugilids from Senegalese Estuaries
Ephigénie Ndew DIONE
Laboratoire d’Ecologie et de Biologie Evolutive, Département de Biologie Animale
Faculté des Sciences et Techniques, Université Cheikh Anta Diop, Dakar, Senegal
Tel: 221-77-456-3138
Malick DIOUF
Laboratoire d’Ecologie et de Biologie Evolutive, Département de Biologie Animale
Faculté des Sciences et Techniques, Université Cheikh Anta Diop, Dakar, Senegal
Institut Universitaire de Pêche et d’Aquaculture (IUPA), Université Cheikh Anta Diop
UCAD II bâtiment pédagogique/Rez de chaussée BP 5005 DAKAR
Tel: 221-33-864-5981; 221-77-501-2780
Alassane SARR
Institut Universitaire de Pêche et d’Aquaculture (IUPA), Université Cheikh Anta Diop
UCAD II bâtiment pédagogique/Rez de chaussée BP 5005 DAKAR,
Tel: 221-77- 656- 8907
Jean FALL (Corresponding author)
Institut Universitaire de Pêche et d’Aquaculture (IUPA), Université Cheikh Anta Diop
UCAD II bâtiment pédagogique/Rez de chaussée BP 5005 DAKAR
Tel: 221-77-956-4339 E-mail: [email protected]
Cheikh Tidiane BÂ

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 11
Laboratoire d’Ecologie et de Biologie Evolutive, Département de Biologie Animale
Faculté des Sciences et Techniques, Université Cheikh Anta Diop, Dakar, Senegal
Tel: 221-77-501-4441
Received: December 7, 2013 Accepted: December 14, 2013
doi:10.5296/jbls.v5i2.5286 URL: http://dx.doi.org/10.5296/jbls.v5i2.5286
Abstract Consumption of mugilids is widespread in Senegal, especially in the estuaries, where there production is of economic importance. The prevalence, mean intensity and pathologic effects of Anisakis sp. and Contracaecum sp. larvae which infected mugilids from Senegalese estuaries were investigated. In 2009, individuals of mugilids belonging to 6 and 5 species were trapped from the estuaries of Saloum, Senegal and Casamance rivers respectively. The prevalence and mean intensity of Anisakis sp. and Contracaecum sp. are higher in Senegal and Saloum estuaries respectively. The increase in the load of Contracaecum sp., related to the season in L. falcipinnis, the season, the size, the weight and/or the sex in M. cephalus from the estuaries of Saloum and Senegal and M. curema from Saloum estuary were due to an accumulation of parasites over time. Liver, kidney, body cavity and mesentery and ovaries are the respective sites of infection of Anisakis sp. and Contracaecum sp. larvae. The immune response observed is the formation of a fibrous capsule surrounding the parasites with melanin granules all around the capsule. The implementation of the capsule and the presence of melanin granules suggest the involvement of lymphocytes, monocytes, neutrophils and macrophages. The lesions caused by Anisakis sp. and Contracaecum sp. larval settlement in target organs may cause the death of the host and/or a reduction in fertility.
Keywords: Anisakidae, Fibrous capsule, Prevalence, Mean intensity, Mugilids, Immune response, Casamance, Saloum, Senegal.
1. Introduction Species of the family Mugilidae commonly known as mugilids are permanent and often abundant coastal, estuarine or lagoon fish. Locally popular, their economic importance is great and they are often the subject of specialized fisheries (Albaret and Legendre, 1985; Albieri et al, 2010; Wang et al, 2010; Durand et al., 2012). Nematode larvae in individual Anisakidae larvae have infected several species of marine, freshwater and brackish water fish including M. cephalus. The presence of larvae Anisakidae in fish may reduce their quality and be dangerous for consumers. Immunological studies of cellular responses induced by these parasites in birds (Nemeth et al 2012), mammals (Cattan et al., 1976; Migaki et al., 1982), human (Chord-Auger et al., 1995; Cuéllar et al., 2010; Daschner et al., 2011; Gonzalez et al., 2010, 2011) , and fish (Munday et al., 2003 ; Longshaw et al., 2004. Murphy et al., 2010; Abowei and Ezekiel, 2011; Buchmann, 2012) have been conducted by many authors around the world. In Africa, histopathological studies in fish from several families have been made but investigations on the effect of Anisakidae on mugilids are a first in Africa, especially in Senegal.
This paper’s main purpose is to determine the parasitic load and study the effect of season, size, weight and sex of six species of mugilids (Liza dumerili, Liza falcipinnis, Liza grandisquamis, Mugil bananensis, Mugil cephalus and Mugil curema) from the Senegalese estuaries on parasitism by larvae of Anisakis sp. and Contracaecum sp. and the histopathological effects

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 12
induced by the establishment of these larvae in the host tissues.
2. Materials and methods 2.1 Sampling procedure Casamance, Saloum and Senegal estuaries constitute the localities chosen within the framework of this study. The estuaries of the Casamance and Saloum rivers are “reverse” estuaries while the estuary of the Senegal river is normal. These ecosystems are characterized by a particular vegetation type, the mangrove swamp. Mangrove of the Casamance and saloum estuaries is composed of six species belonging to three families: Rhizophoraceae, Verbenaceae and Combretaceae. The mangrove of the estuary of the Senegal river is of minor importance in comparison with Casamance and Saloum mangroves. Low to sparse density and height rarely exceeding 5 m, it consists of two species: Rhizophora racemosa and Avicenia africana.
In these estuaries, samplings were done systematically per quarter. Three mugilid samplings were carried out in the dry season and one in the rainy season of 2009. The techniques of fishing used were the beach seines and the gill nets.
2.2 Parasite Prevalence and Mean Intensity The prevalence (P) or parasitism rate is the percentage ratio of the number of infected hosts (N) by a given parasite species on the number of fish examined (H).
P (%) = N / H * 100
The parasite mean intensity (I) refers to the total number of individuals of a parasite species (n) by the number of infected fish (N).
I = n / N
2.3. Effect of weight, size and sex of Mugil cephalus and season on parasitism
We consider intensity or parasite load as a dependent variable or response variable and the weight or size, the season and the sex as variables. Among the explanatory variables, there is a continuous variable which is the weight or the size and two categorical variables or factors, the season and the sex, which are at two levels. A linear regression relationship allows us to study the effect of size, weight and/or sex of each host and the season on the parasite load. The meaning of the data is established at 0.05. R software was used for statistical analysis.
2.4 Histopathological Study Collected first, livers, kidneys and mesentery of host fish underwent three ethanol baths to 95% on the first day, then 3 butanol baths on the second day and then three paraffin baths on the third day before being lumped in "Leuckart bars" placed on a glass plate. Cuts of 5 to 7 microns thick are made two to three days after making the block, with a paraffin microtome Minot kind. They were then plated and bonded to albumin on cleaned slides of ethanol 95%. Drying was done in a paraffin oven at 60 °C. The slides thus prepared were stained with Groat haematoxylin and dyes of Masson Trichome, which helped differentiate tissues.
3. Results 3.1 Prevalence and Mean Intensity Ecological parameters prevalence and mean intensity of Anisakis sp. and/or Contracaecum sp. larvae parasites of L. dumerili, L. grandisquamis, M. bananensis, M. cephalus and M. curema are summarized in the tables 1 to 6 below.

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 13
The prevalence of Anisakis sp. recorded in the estuary of the Senegal river is higher than that found in estuaries of the Saloum and Casamance rivers whatever the host species. It also varies according to the season. Indeed, this ecological parameter is higher in rainy season than in dry season except for M. bananensis originating in the Saloum estuary.
In hosts of the genus Mugil, the mean intensity of Anisakis sp. obtained in Senegal estuary is higher than that recorded in the estuaries of the Casamance and Saloum rivers, with a significant difference in M. bananensis and M. cephalus. In L. grandisquamis, parasite intensity is higher in the Saloum estuary than in the estuaries of the Casamance and Senegal rivers. In host species where the parasite was met throughout the year, the mean intensity is higher in rainy season than in dry season, but a significant difference was only observed in M. cephalus from the estuary Saloum.
The prevalence and mean intensity of Contracaecum sp. identified in the Saloum estuary are generally higher than those recorded in the estuaries of the Casamance and/or Senegal rivers regardless of host species. These ecological parameters vary seasonally. Indeed, in species where the parasite was met throughout the year, the prevalence and mean intensity is higher in dry season than in rainy season except for M. curema native to the Saloum estuary for the prevalence and L. dumerili from the estuary of the Casamance river for the mean intensity. The difference between the values of the mean intensity according to the season is significant in M. cephalus and L. falcipinnis from the Saloum estuary.
Table 1. Prevalence and mean intensity of Contracaecum sp. larvae parasite of L. dumerili
Contracaecum sp. P (%) I
Localities RS DS RS DS Casamance 10 (H=10, N=1) 50 (H=6, N=3) 5 3.33±3.21
Saloum NE 100 (H=60, N=60) NE 50.06±41.25 Senegal 11.76 (H=17, N=2) 54.35 (H=46, N=25) 9.5±10.61 10.25±7.08
RS: rainy season
DS: dry season
NE=host not examined
Table 2. Prevalence and mean intensity of Contracaecum sp. larvae parasite of L. falcipinnis
Contracaecum sp.
P (%) I Localities RS DS RS DS
Casamance NF NF NF NF Saloum 35.85 (H=53, N=19) 81.08 (H=37, N=30) 10.47±17.96 56.52±69.1 Senegal 1.2 (H=25, N=3) 25.81 (H=63, N=16) 2±1.73 4.79±6.68
RS: rainy season; DS: dry season; NF=parasite not found
Table 3. Prevalence and mean intensity of Anisakis sp. and Contracaecum sp. larvae parasite of L. grandisquamis
Anisakis sp. Contracaecum sp. P (%) I P (%) I
Localities RS DS RS DS DS DS
Casamance 6.67 (H=30, 3.37 (H=148, 1±0 1±0 2.01 (H=148, 18±9.54

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 14
N=2) N=5) N=3)
Saloum NE 6.67 (H=30, N=2) NE 1.5±0.71 63.64 (H=33,
N=21) 4.52±3.82
Senegal 23.07 (H=13, N=3) NF 1±0 NF NF NF
RS: rainy season; DS: dry season; NE=host not examined; NF=parasite not found
Table 4. Prevalence and mean intensity of Anisakis sp. and Contracaecum sp. larvae parasite of M. bananensis
Anisakis sp. Contracaecum sp.
P (%) I P (%) I
Localities RS DS RS DS RS DS RS DS
Casamance NE 100
(H=1, N=1)
NE 1±0 NE 100
(H=1, N=1)
NE 26±0
Saloum 60
(H=20, N=12)
81.81 (H=11, N=9)
1.25±0.45 1.33±0.5 52.63
(H=19, N=10)
100 (H=9, N=9)
10.6±14.06 11.89±9.44
Senegal NE 90.47
(H=21, N=19)
NE 2.84±1.74 NE 53.85
(H=13, N=3)
NE 20.8±20.7
RS: rainy season
DS: dry season
NE=host not examined
Table 5. Prevalence and mean intensity of Anisakis sp. and Contracaecum sp. larvae parasite of M. cephalus (Dione et al. 2014)
Anisakis sp. Contracaecum sp.
P (%) I P (%) I
Localities RS DS RS DS RS DS RS DS
Casamance NE NE NE NE NE NE NE NE
Saloum 51.02
(H=49, N=25)
32.5 (H=80, N=26)
1.6±0.71 1.58±0.64 22.44
(H=49, N=11)
51.25 (H=80, N=41)
3.09±1.81 11.71±14.87
Senegal 90
(H=60, N=54)
73.83 (H=107, N=79)
3.09±1.92 2.38±1.5 6.66
(H=60, N=4)
25.23 (H=107, N=27)
3.5±3 5.93±7.23
RS: rainy season
DS: dry season
NE=host not examined
Table 6. Prevalence and mean intensity of Anisakis sp. and Contracaecum sp. larvae parasite of M. curema

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 15
Anisakis sp. Contracaecum sp.
P (%) I P (%) I
Localities RS DS RS DS RS DS RS DS
Casamance NF NF NF NF NF NF NF NF
Saloum 70.59
(H=17, N=12)
37 (H=100, N=37)
1.4±0.97 1.16±0.37 78.57
(H=14, N=11)
65.31 (H=98, N=74)
13.45±10.03 21.75±52.91
Senegal NE 84.62
(H=13, N=11)
NE 1.64±1.03 NE 11.11 (H=9, N=1)
NE 1±0
RS: rainy season
DS: dry season
NE=host not examined
NF=parasite not found
3.2 Effect of Size, Weight and Sex of the Host and Season on the Parasitism For L. dumerili, L. grandisquamis and M. bananensis from the estuaries of the Casamance, Saloum and Senegal rivers, season, weight, height and sex can not explain the variations in the load or parasite intensity of Contracaecum sp. or Anisakis sp larvae. In L. falcipinnis from the Saloum estuary, the variations in the intensity of Contracaecum sp. can not be elucidated by the season, the other above variables are not related to this ecological setting. Individuals operating in the estuary of the Senegal host larvae Contracaecum sp. with a load which changes can not be explained either by season, size, weight or sex.
; R2=13.8%; p=0.008
In M. cephalus, the variations in the parasite load of Anisakis sp. can not be explained either by size or by weight or by sex or season. In contrast to Contracaecum sp., the parasite load is related to the size, weight and season in the Saloum estuary while in the estuary of Senegal, it is correlated to the size, weight and gender. The following models are used to show the linear regression relationship existing between the parasite load and the different explanatory variables.
In individuals infected from the Saloum Delta, the variations in the parasite intensity can be explained by the size or weight and season. Thus, individuals with large size or weight harbor much more Contracecum sp. larvae than those of small size or weight. In addition, the parasite load was higher in the dry season than the rainy season.
; R2=27.66%; p=0.002
; R2=26.78%; p=0.002

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 16
In the estuary of the Senegal river, the variations in the parasite load of Contracaecum sp. larvae can be explained by size, season and sex or weight and sex. Indeed, the number of larvae found in a host increases with the size or weight. It also depends on the season that is to say, is higher in dry season than in rainy season and it is even higher in males than individuals of indeterminate sex.
; R2=71.11%; p=0.000
; R2=85.66%; p=0.000
Anisakis sp. larvae parasites of M. curema from the estuaries of the Saloum and Senegal rivers have a parasitic intensity whose changes can be associated with the season, the size, the weight or the sex of the host. As the larvae Contracaecum sp. parasitizing hosts individuals from the Saloum estuary, they have a parasite load which is related to the above explanatory variables. Indeed, the linear relationship between the environmental parameters and the explanatory variables shows that host individuals which have a size or heavy weight are the most infested. This relationship also shows that the parasitic intensity is higher in dry season than in rainy season and the female individuals harbor more Contracaecum sp. larvae than males.
; R2=12, 67%; p=0,021
; R2=16, 64%; p=0,005
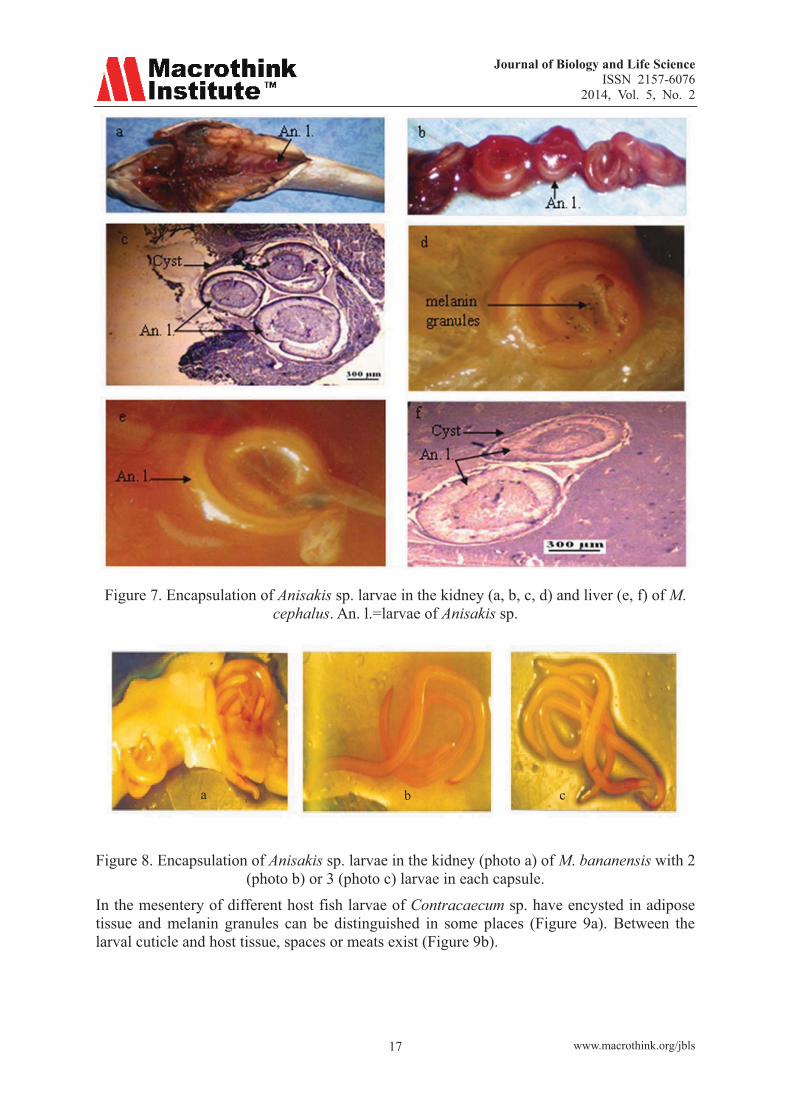
3.2 Histopathological Study The kidney is noisy in its posterior part (Figure 7a) by Anisakis sp. resulting in the formation of capsules in which the worm is wound several times (Figure 7b) to give sectional multiple sections of the worm (Figure 7c). In the liver, the larvae of Anisakis sp. have encysted (Figure 7e) and histological sections also show worms at different levels (Figure 7f). The turn of parasites, liver and kidney tissue was disorganized revealing gaps and black granules (Figure 7d). Two or 3 larvae of Anisakis sp. can be found in a capsule (Figure 8a, b, c).
Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 17
Figure 7. Encapsulation of Anisakis sp. larvae in the kidney (a, b, c, d) and liver (e, f) of M. cephalus. An. l.=larvae of Anisakis sp.
Figure 8. Encapsulation of Anisakis sp. larvae in the kidney (photo a) of M. bananensis with 2 (photo b) or 3 (photo c) larvae in each capsule.
In the mesentery of different host fish larvae of Contracaecum sp. have encysted in adipose tissue and melanin granules can be distinguished in some places (Figure 9a). Between the larval cuticle and host tissue, spaces or meats exist (Figure 9b).
a b c

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 18
melanin granules
Cont. l. Cont. l.
Cyst
meat
a b
Figure 9. Encapsulation of Contracaecum sp. larvae in the mesentery of M. curema. Cont. l.= larvae of Contracaecum sp.
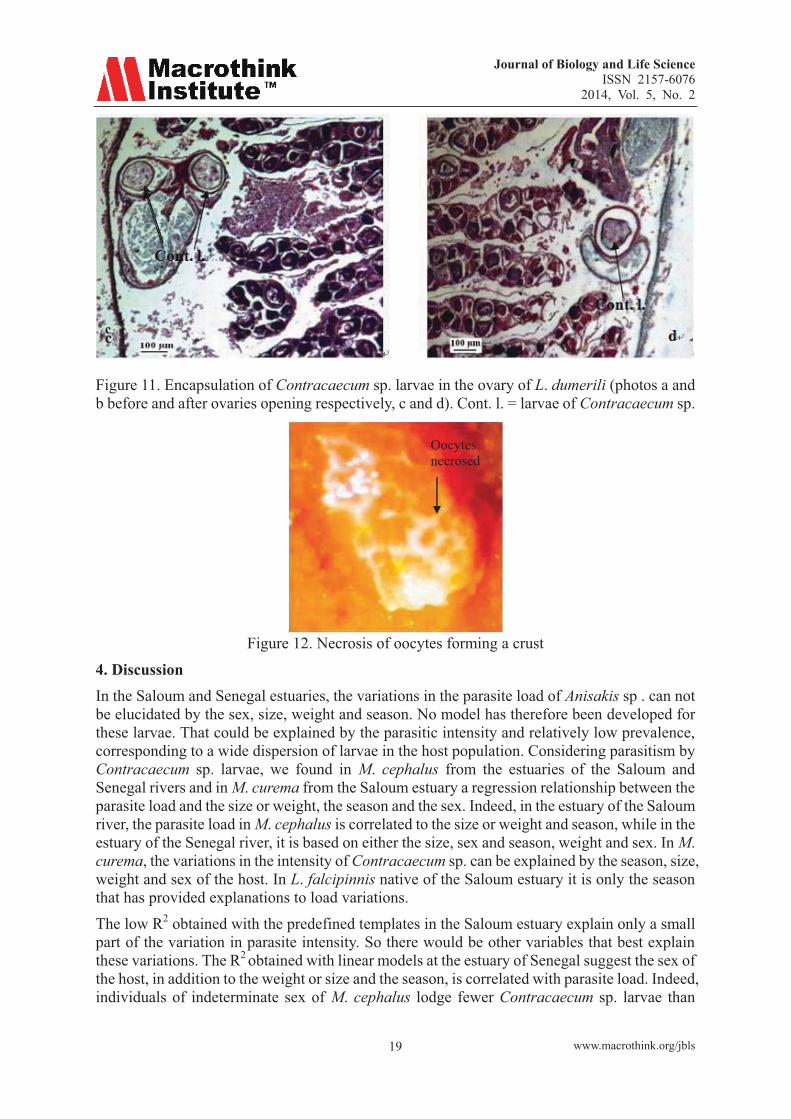
Some larvae of Contracaecum sp. were found encysted on ovarian filaments of L. grandisquamis (Figure 10) or in the ovaries of L. dumerili (Figure 11), L. grandisquamis and M. bananensis. The capsules containing the parasite are peripheral and located in the ovary wall and rely on meats. The stacks of eggs near the cyst suffered disruption to the contact zone. The capsule wall is very thick and we noted some areas of degradation products of oocytes (Figure 9. c, d). This degradation of oocytes can be very deep and result in the formation of a crust (Figure 12).
Figure 10. Fixing Contracaecum sp. larvae on ovarian filaments of L. grandisquamis
Cont. l. Cont. l.
a b
Cont. l.
Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 19
Figure 11. Encapsulation of Contracaecum sp. larvae in the ovary of L. dumerili (photos a and b before and after ovaries opening respectively, c and d). Cont. l. = larvae of Contracaecum sp.
Figure 12. Necrosis of oocytes forming a crust
4. Discussion In the Saloum and Senegal estuaries, the variations in the parasite load of Anisakis sp . can not be elucidated by the sex, size, weight and season. No model has therefore been developed for these larvae. That could be explained by the parasitic intensity and relatively low prevalence, corresponding to a wide dispersion of larvae in the host population. Considering parasitism by Contracaecum sp. larvae, we found in M. cephalus from the estuaries of the Saloum and Senegal rivers and in M. curema from the Saloum estuary a regression relationship between the parasite load and the size or weight, the season and the sex. Indeed, in the estuary of the Saloum river, the parasite load in M. cephalus is correlated to the size or weight and season, while in the estuary of the Senegal river, it is based on either the size, sex and season, weight and sex. In M. curema, the variations in the intensity of Contracaecum sp. can be explained by the season, size, weight and sex of the host. In L. falcipinnis native of the Saloum estuary it is only the season that has provided explanations to load variations.
The low R2 obtained with the predefined templates in the Saloum estuary explain only a small part of the variation in parasite intensity. So there would be other variables that best explain these variations. The R2 obtained with linear models at the estuary of Senegal suggest the sex of the host, in addition to the weight or size and the season, is correlated with parasite load. Indeed, individuals of indeterminate sex of M. cephalus lodge fewer Contracaecum sp. larvae than
Oocytes necrosed
c

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 20
males. Females of M. curema are more frequently infected than males. These results allow us to say that the effect of gender on parasitism is stronger between immature and mature than between mature individuals only. The physico-chemical environmental conditions such as temperature or salinity and the presence of birds, definitive hosts of these nematodes also explain the variations noted in the load of Contracaecum sp.
The increase of the parasite burden with the size or weight of M. cephalus in both estuaries and M. curema from the Saloum estuary means that larvae of Contracaecum sp. have been accumulated, after repeated infestations in fish hosts over time. We confirm this accumulation of parasites over time already mentioned by several authors (Valles -Rios et al., 2000; Bergmann and Motta, 2004; Al- Zubaidy, 2009) particularly in the Senegal estuary where male fish lodge more larvae of Contracaecum sp. than individuals of indeterminate sex. However, some authors (Cremonte and Sardella, 1997, Gonzalez and Acuna, 2000; Sylva and Eiras, 2003; Abattouy et al., 2011; Tantanasi et al., 2012; Tejada, 2013) found no relationship between parasitism by Anisakis spp. and sex of the host fish, but noted an increased risk of infection with the weight, size or season or three variables. Given the importance of fish weight for the consumer, it is necessary to give the following preventive measures: it is safer to consume M. cephalus and M. curema from these estuaries of the Saloum and Senegal rivers with average weight to reduce the risk of contamination because it is the big fish that contain more nematode larvae.
In this study we did not find Anisakidae larvae in the flesh of fish, but in the kidney, the liver, the body cavity, the mesentery and the ovaries. Therefore these larvae were not able to reach the fish. Migration depends on factors related to fish muscles such as wealth even more fat than their preference for high fat tissue was observed (Lymbery et al., 2002 ; Stromnes and Anderson, 2003 ; Abattouy et al., 2011). Many authors (Smith and Wootten, 1975 ; Wootten and Waddell, 1977 ; Smith 1984, Wharton et al., 1999 ; Kino et al., 1993 ; Abollo et al., 2001 ; Martins et al., 2005 ; Mo and al., 2013) supported the idea that Anisakidae larvae perform a post- mortem migration in the fish. In contrast, other authors (Mladineo, 2001; Tejeda, 2010), speak of migration at catching fish.
If the migration of Anisakidae larvae was done after the death of fish, immediate evisceration could prevent. Large firms conduct early fish gutting and nevertheless anisakidose cases continue to be identified in some countries like Japan and the USA. Migration of nematodes for catching fish is plausible. As soon as the fish is trapped, it is in a state of stress, which can lead to birth and signal flow in the body. These stress signals can be picked up by the parasites in particular Anisakidae larvae that will migrate into the fish where they will be safer. That is a good strategy for these larvae to stay alive in the meantime, by chance, to move to another host, such as human. The site of infection in the host fish is an important determinant of the risk of human anisakiasis (Quiazon et al., 2011). The greater presence of larvae of Anisakis sp. in the kidney is a potential risk of contamination because it is usually consumed. Another prophylactic measure would be to remove the kidney during evisceration.
The installation of Anisakis sp. larvae in renal and hepatic serous and/or general cavity of L. grandisquamis, M. bananensis, M. cephalus and M. curema and that of Contracaecum sp. larvae in the mesentery and/or ovaries of L. dumerili, L. falcipinnis, L. grandisquamis, M. bananensis, M. cephalus and M. curema are causing an inflammatory reaction leading to the formation of a fibrous capsule around the parasite. The shell is made of a hard and stratified epithelium and is therefore slightly permeable to nutrients, Berland (1998). Its implementation involves several cells including neutrophils, fibroblasts, lymphocytes and macrophages (Elarifi, 1982; Dezfuli et al., 2007; Murphy et al., 2010). The complete encapsulation of the larvae of

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 21
Anisakis sp. in the liver, kidney and of Contracaecum sp. in the mesentery and/or ovaries of host prevents migration and therefore the infested sites lesion. However, the translucent areas and debris from tissue surrounding the capsule show degradation be it small, of infected organs.
Then, how lysis of host tissues is made while the larvae are not in direct contact with them? According to Larsen (1980), the cephalic tooth of Anisakidae larvae functions as a needle piercing the capsule to permit the entry of host cells. He also states that the cephalic gland secret histolytic enzymes that lead the process of maintaining an opening in the capsule. The lesions of the kidney and liver, essential functional organs could cause a more or less long-term death of the host. The pathogenic action of the larvae of Anisakis sp. was shown by Petter (1969). The experience has been to ingest larvae of Anisakis sp. into a mugilid (M. cephalus), the death of the fish was observed 6 days after ingestion. The larvae of Anisakis sp. are pathogenic for these fish hence the importance of paying attention to these parasites. Given their large size, they cause mechanical compression of tissues causing malfunctions of infected organs.
The consequence of many lesions of the mesentery was slimming host fish seeing that it is an important lipid reserves. These results could be compared with those of Petter et al., (1974) although parasitic nematodes in question and the hosts are different, but also with those of Lymbery et al., (2002 ) in which larvae of Contracaecum sp. found in M. cephalus and Aldrichetta forsteri are causing major morphological changes in the liver. The pathogenic action of larvae of Contracaecum sp. was also studied by Acha and Szyfres, (1987) who reported a decrease in weight of the host and the lipid content of the liver or even the death of young fish when larvae invade the cardiac region. The presence of necrotic plaque in the ovaries can deeply affect the fertility of the host. Indeed, necrosis oocytes lead to decreased fertility of the host and the result in longer term would be a reduction of the fish stock.
5. Conclusion Prevalence and mean intensity of Anisakis sp. and Contracaecum sp. larvae are higer in the estuaries of the Senegal and Saloum rivers respectively. Variations of the load of Anisakis sp. can not be explained by the locality, the season, the size, the weight or the sex of hosts. In contrast, variations of Contracaecum sp. intensity are related to the season, the size, the weight and/or the sex of some hosts. The settlement of the anisakid larvae may induce the death of the host or a reduction in fertility.
Acknowlegment We thank local fishermen from Djilor and St. Louis, Mr Edouard DIATTA and Mr Ousmane (the boatman) from Casamance who helped us a lot to collect fish. We wish to express our sincere thanks to Mr Michel SARR for technical assistance in the implementation of histological sections and coloring, Mr Omar SADIO for his help in statistical analysis and Mr Ferdinand DIATTA from the BSI who kindly corrects the english version of this document.
References Abattouy, N., Valero, A., Benajiba, M. H., Lozano, J., & Martíns-Sánchez, J. (2011). Anisakis simplex s.l. parasitization in mackerel (Scomber japonicus) caught in the North of Morocco – Prevalence and analysis of risk factors. International Journal of Food Microbiology, 150, 136-139. http://dx.doi.org/10.1016/j.ijfoodmicro.2011.07.026
Abollo, E., Gestal, C., & Pascual S. (2001). Anisakis infestation in marine fish and cephalopods from Galician waters: an updated perspective. Parasitology Research, 87, 492-499.

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 22
http://dx.doi.org/10.1007/s004360100389
Abowei, J. F. N., & Ezekiel, E. N. (2011). Trematoda tape worms: infections by larval and other tape worms and nematoda in African fish (a review). International Journal of Animal and veterinary Advances, 3, 352-366. ISSN: 2041-2908
Acha, P. N., & Szyfres, B. (1987). Zoonoses and communicable diseases common to man and animals, 2nd edn. Pan American Health Organisation, Washington, DC.
Al-Zubaidy, A. B. (2009). Prevalence and densities of Contracaecum sp. larvae in Liza abu Heckel 1843 from different Iraqi water bodies. J. K. A. U.: Marine Science, 20, 3-17. http://dx.doi.org/10.4197/Mar.20-1.1
Bergmann, T. G., & Motta, P. J. (2004). Infection by anisakid nematodes Contracaecum spp. in the Mayan cichlid fish ‘Cichlasoma (Nandopsis)’ urophthalmus (Günther 1862). Journal of Parasitology, 90, 405-407. http://dx.doi.org/10.1645/GE-104R
Buchmann, K. (2012). Fish immune responses against endoparasitic nematodes- experimental models. Journal of Fish Diseases, 35, 623-635. http://dx.doi.org/10.1111/jfd.12096
Cattan, P. E., Babero, B. B., & Torres, D. N. (1976). The helminth fauna of Chile: IV Nematodes of general Anisakis Dujardin, 1845 and Phocanema Myers, 1954 in relation with gastric ulcers in a South American sea lion, Otaria byronia. Journal of Wildlife Diseases, 12, 511-515. http://dx.doi.org/10.7589/0090-3558-12.4.511
Chord-Auger, S., Miegeville, M., & Le Pape, P. (1995). L’anisakiase dans la région nantaise: de l’étal du poissonnier au cabinet du médecin. Parasite, 2, 395-400.
Cremonte, F., & Sardella, N. H. (1997). The parasito fauna of Scomber japonicus Houttuyn, 1782 (Pisces: Scombridae) in two zones of the Argentine sea. Fisheries Research, 31, 1-9. http://dx.doi.org/10.1016/S0165-7836(97)00024-6
Cuéllar, C., Rodero, M., & Daschner, A. (2010). Inhibition of cytokine production in gastro-allergic Anisakiasis and Anisakis simplex sensitization-associated chronic urticaria. Parasite Immunology, 32, 528. http://dx.doi.org/10.1111/j.1365-3024.2010.01216.x
Dashner, A., Rodero, M., De Frutos, C., Valls, A., Vega, F., Blanco, C., & Cuéllar, C. (2011). Different serum cytokine levels in chronic vs. acute Anisakis simplex sensitization-associated urticaria. Parasite Immunology, 33, 357-362. http://dx.doi.org/10.1111/j.1365-3024.2011.01282.x
Dezfuli, S., Pivoni, F., Shinn, A. P., Marena, M., & Giari, L. (2007). Histopathology and ultrastructure of Platichthys flesus naturally infected with Anisakis simplex S. L. larvae (Nematoda: Anisakidae). Journal of parasitology, 93, 1416-1423. DOI: http://dx.doi.org/10.1645/GE-1214.1
Dione, E. N., Diouf, M., Fall, J. & Bâ, C. T. (2014). Seasonal and spatial distribution of nematode larvae of the genera Anisakis and Contracaecum (Anisakidae) in two populations of Mugil cephalus (Mugilidae) from Saloum and Senegal rivers. Journal of Biology and Life Science, 5: 41-56.
Elarifi, A. E. (1982). The histopathology of larval anisakid nematode infections in the liver of whiting, Merlangius merlangus (L.), with some observations on blood leucocytes of the fish. Journal of Fish Diseases, 5, 411-419. DOI: 10.1111/j.1365-2761.1982.tb004
Gonzalez, M., Rodriguez-Mahillo, A., & Moneo, I. (2011). Cytokine production and immune modulation by Anisakis simplex. Parasite Immunology, 33, 143.

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 23
http://dx.doi.org/10.1111/j.1365-3024.2010.01254.x
Gonzalez, T., & Acuna, E. (2000). Influence of host size and sex on the endohelminth infracommunities of the Red Rockfish Sebastes capensis off Northern Chile. Journal of Parasitology, 86, 854-857.
Gonzalez–Muñoz, M., Rodriguez-Mahillo, A. I., & Moneo, I. (2010). Different Th1/Th2 responses to Anisakis simplex are related to distinct clinical manifestations in sensitized patients. Parasite Immunology, 32, 67-73. http://dx.doi.org/10.1111/j.1365-3024.2009.01162.x
Kino, H., Watanabe, K., Matsutomo, K., Ueda, M., Sigiura, M., Suzuki, H., Takai, T., Tsuboi, H., Sano, M., Fujiu, H., & Kagel, N. (1993). Occurrence of Anisakis in the western part of Shizuoka Prefecture, with special reference to the prevalence of anisakid infections in sardine, Engraulis japonica. Japanese Journal of Parasitology, 42, 308-312.
Longshaw, M., Green, L., & Feist, S. W. (2004). Histopathology of parasitic infections in greater pipefish, Syngnathus acus L., from an estuary in the UK. Journal of Fish Diseases, 27, 245-248. http://dx.doi.org/10.1111/j.1365-2761.2004.00533.x
Lymbery, A. J., Doupé, R. G., Munshi, M. A., & Wong, T. (2002). Larvae of Contracaecum sp. among inshore fish species of southwestern Australia. Diseases of Aquatic Organisms, 51, 157-159. http://dx.doi.org/10.3354/dao051157
Martins, M. L., Onaka, E. M., & Fenerick Jr, J. (2005). Larval Contracaecum sp. (Nematoda: Anisakidae) in Hoplias malabaricus and Hoplerythrinus unitaeniatus (Osteichthyes: Eruthrinidae) of economic importance in occidental marshlands of Maranhão, Brazil. Veterinary Parasitology, 127: 51-59. http://dx.doi.org/10.1016/j.vetpar.2004.09.026
Mattiucci, S., Paoletti, M., Borrini, F., Palumbo, M., Palmieri, R. M., Gomes, V., Casati, A., & Nascetti, G. (2011). First molecular identification of the zoonotic parasite Anisakis pegreffii (Nematoda: Anisakidae) in a paraffin-embedded granuloma taken from a case of human intestinal anisakiasis in Italy. BMC Infectious Diseases, 11, 82. DOI: http://dx.doi.org/10.1186/1471-2334-11-82
Migaki, G., Heckmann, R. A., & Albert, T. F. (1982). Gastric nodules caused by «Anisakis type» larvae in the bowhead whale (Balaena mysticetus). Journal of Wildlife Diseases, 18, 353-357. http://dx.doi.org/10.7589/0090-3558-18.3.353
Mladineo, I. (2001). Effects of pH values and temperature changes on migration of Anisakis simplex Dujardin, 1845 invasive larvae (Nematoda, Askaridoidea, Anisakidae) in mackerel (Scomber japonicus, L.). Acta Adriatica, 42, 71-75.
Mo, A. T., Gahr, A., Hansen, H., Hoel, E., Oaland, Ø., & Poppe, T. T. (2013). Presence of Anisakis simplex (Rudolphi, 1809 det. Krabbe, 1878) and Hysterothylacium aduncum (Rudolphi 1802) (Nematoda: Anisakidae) in runts of farmed atlantic salmon, Salmo salar L. Journal of Fish Diseases, http://dx.doi.org/10.1111/jfd.12096
Munday, B. L., Sawada, Y., Cribb, T., & Hayward, C. J. (2003). Diseases of tunas, Thunnus spp. Journal of Fish Diseases, 26, 187-206. http://dx.doi.org/10.1046/j.1365-2761.2003.00454.x
Murphy, T. M., Berzano, M., O’keeffe, S. M., Cotter, D. M., McEvoy, S. E., Thomas, K. A., O’Maoile’Idigh, N. P. & Whelan, K. F. (2010). Anisakid larvae in Atlantic salmon (Salmo Salar L.) Grilse and post-smolts: molecular identification and histopathology. Journal of parasitology, 96, 77-82. DOI: http://dx.doi.org/10.1645/GE-2194.1
Nemeth, N. M., D.V.M., Ph.D, Yabsley, M., Ph.D., & Keel, M. K., D.V.M., Ph.D., Dipl.

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 24
A.C.V.P. (2012). Anisakiasis with proventricular perforation in a greater shearwater (Puffinus gravis) off the coast of Georgia, United States. Journal of Zoo and Wildlife Medicine, 43, 412-415. DOI: http://dx.doi.org/10.1638/2011.0193.1
Quiazon, K. M. A., Yoshinaga, T., & Ogawa, K. (2011). Experimental challenge of Anisakis simplex sensu stricto and Anisakis pegreffii (Nematoda: Anisakidae) in rainbow trout and olive flounder. Parasitology International, 60, 126-131. DOI: 10.1016/j/parint.2010.11.007
R Development Core Team. 2009. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0, URL http://www.R-project.org.
Silva, M. E. R., & Eiras, J. C. (2003). Occurrence of Anisakis sp. in the fish off the Portuguese West coast and evaluation of its zoonotic potential. Bulletin of the European Association of Fish Pathologists, 23, 13-17.
Smith, J. W., & Wootten, R. (1975). Experimental studies on the migration of Anisakis sp. larvae (Nematoda: Anisakidae) into the flesh of herring, Clupea harengus L. International Journal for Parasitology, 5, 133-136. http://dx.doi.org/10.1016/0020-7519(75)90019-3
Smith, J. W. (1984). The abundance of Anisakis simplex L3 in the body-cavity and flesh of marine teleosts. International Journal for Parasitology, 14, 491-495. http://dx.doi.org/10.1016/0020-7519(84)90030-4
Strømnes, E., & Andersen, K. (2003). Growth of whaleworm (Anisakis simplex, Nematoda, Ascaridoidea, Anisakidae) third-stage larvae in paratenic fish hosts. Parasitology Research, 89, 335-341. DOI: 10.1007/s00436-002-0756-7
Tantanasi, J., Diakou, A., Tamvakis, A., & Batjakas, I. E. (2012). Anisakis spp. burden in Trachurus trachurus. Helminthologia, 49, 16-20. http://dx.doi.org/10.2478/s11687-012-0003-4
Tejeda, M. (2010). Seguridad alimentariaben pescados: Anisakis. In Seguridad alimentaria e higiene de los alimentos Jiménez Pérez. S (Ed) Instituto Tomás Pascual Sanz para la Nutrición y la Salud y Real Academia de Ciencias Veterinarias de España, Madrid; Spain. pp, 107-127.
Tejeda, M., Karl, H., De Las Heras, C., Vidacek, S., Solas, M. T., & Garcia, M. L. (2013). Does the intensity of Anisakis infection affect the quality of hake muscle ? Journal of aquatic Food Product Technology, ISSN/ISBN, 1547-0636.
Valles-Rios, M. E., Ruiz-Campos, G., & Galaviz-Silva, L. (2000). Prevalencia e intensidad parasitaria en Mugil cephalus (Pisces: Mugilidae), del Rio Colorado, Baja California, Mexico. Revista de biología tropical, 48, 495-501.
Wharton, D. A., Hassal, M. L., & Aalders, O. (1999). Anisakis (Nematoda) in some New Zealand inshore fish. New Zealand Journal of Marine and Freshwater Research, 33, 6436648. http://dx.doi.org/10.1080/00288330.1999.9516907
Wootten, R., & Waddell, I. F. (1977). Studies on the biology of larval nematodes from the musculature of cod and whiting in scottish waters. Journal du Conseil International pour l’Exploration de la Mer, 37, 266-273. http://dx.doi.org/10.1093/icesjms/37.3.266
Copyright Disclaimer Copyright reserved by the author(s).
This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 25
Evaluating the Nutritional Potential of Acacia Sieberiana Seeds (Dc) Growing In North West of
Nigeria
Abubakar Salisu (Corresponding author)
Biotechnology and Genetic Engineering Laboratory, Sheda Science and Technology Complex
(SHESTCO), P.M.B. 186, Garki, FCT Abuja
Tel: 234- 803-969-0544 E-mail: [email protected]
Godwin H. Ogbadu
Biotechnology and Genetic Engineering Laboratory, Sheda Science and Technology Complex
(SHESTCO), P.M.B. 186, Garki, FCT Abuja
Tel: 234-705-777-6677 E-mail: [email protected]
Paul C. Onyenekwe
Biotechnology and Genetic Engineering Laboratory, Sheda Science and Technology Complex
(SHESTCO), P.M.B. 186, Garki, FCT Abuja
Tel: 234-803-634-7293 E-mail: [email protected]
Omotoye Olorode
Department of plant science and Applied Zoology, Olabisi Onabanjo University, Ago-Iwoye,
P.M.B. 2002, Ogun State, Nigeria
Tel: 234-803-226-5244 E-mail: [email protected]
Rebecca W. Ndana
Department of Biological Science, University of Abuja, P.M.B. 117, FCT Abuja.
Tel: 234-703-916-0188 E-mail: [email protected]

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 26
Olayanju Segun
Department of Biological Science, University of Abuja, P.M.B. 117, FCT Abuja
Tel:234-803-947-2307 E-mail: [email protected]
Received: January 1, 2014 Accepted: January 15, 201
doi:10.5296/jbls.v5i2.4860 URL: http://dx.doi.org/10.5296/jbls.v5i2.4860
Abstract
Concerted efforts in search of alternative sources of protein has grown due to dearth of animal protein in developing countries, as food shortage and poverty become more endemic, people increasingly depend on plants rather than animals for proteins in their diets. This work focuses on untapped indigenous wild savannah tree, Acacia sieberiana (seeds) for its nutritional and economical values. The seeds were collected and processed for proximate compositions, phytochemicals and elemental analysis. The high percentage of crude protein content of 49.7% and absence of toxic elements such as Cadmium (Cd), Arsenic (Ar), and very low percentage of Lead (Pb) 0.001ml/g has proven that the Acacia sieberiana seeds are very safe and could serve as an alternative source of protein. The result of the quantitative minerals determination shows the presence of Manganese (3.93mg/g), Calcium (2.02mg/g), and Iron (0.11mg/g). The germination trials of the seeds were also carried out, and the result shows that, soaking, heating the seeds at 600C and even the non treated seeds as well as those planted and mulched gave promising results of the plant’s seeds germination. The statistical package for social science (SPSS) software was used to analysed the significant differences between the treatments at
critical value (p≥0.05).
Keywords: Acacia sieberiana, Elemental analysis, Germination trial, Proximate compositions, Phytochemicals screening.
1. Introduction
Throughout history man has used some 3,000 plant species for food, at least 150 of them have been commercially cultivated to some extent. But over the centuries the tendency has been to concentrate on fewer and fewer crop and a large percentage are underutilized (NAS, 1984) In developing countries, wild plants are exploited as sources of food and other life supporting commodities and thus provide an adequate level of nutrition to the human beings (Aberoumand and Deokule, 2010). These wild plants serve as an indispensable constituent of human diet supplying the body with minerals, vitamins and certain hormone precursors, in addition to protein and energy (Akubugwo et al., 2007).
Acacia sieberiana (English name: White thorn, Hausa: Fara kaya, Yoruba: Aluki sie). A member of the Family Fabaceae – Mimosoideae, is a tree that grows up to 15 m high with

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 27
light-coloured bark and often with a flat crown. The leaves, 10-15 cm long have straight white thorns at their base. The branches and often the leaves are covered with yellow hairs. The flower heads are cream-coloured and spherical. The seeds are contained in straight pods, 8-12 cm long and 2-3 cm broad. The tips of young shoots are intensively browsed particularly toward the end of the dry season. A. sieberiana grows in the savannah and woodland. It occurs with various botanical characteristics in the entire Sahel and other semi-arid regions in Africa (Orwa et al., 2009). The tree possessed some ethnobotanical history; decoction of the root is taken as remedy for stomach-ache. The bark, leaves and gums are used to treat tapeworm, bilharzia, haemorrhage, orchitis, colds, diarrhoea, gonorrhoea, kidney problems, syphilis, ophthalmia, rheumatism and disorders of the circulatory system. It is also used as an astringent. The pods serve as an emollient, and the roots for stomach-ache, acne, tapeworms, urethral problems, oedema and dropsy (Orwa et al., 2009).
This work was aimed at evaluating the seeds of A. sieberiana (DC) which is untapped nutritionally and economically; the proximate composition, phytochemicals, elemental analysis and germination trials were carried out. The work could contribute to educate the indigenous people about the nutritional values of this seeds, and the seeds can also be incorporated among edible and commercial seed plants, which will positively improve the indigenous economy. The research is also being the first attempt to consider the nutritional and economical value of A. sieberiana (DC) seeds.
Figure 1. Acacia sieberiana tree Figure 2. Acacia sieberiana pod and seeds.
Source: BUK, 2013.

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 28
2. Materials and Methods
2.1 Sample Collection
The A. sieberiana seeds were collected from uncultivated land in Bayero University new campus Kano state, Nigeria. It is located in North-West Nigeria, lies between latitude 130N and 110N and longitude 80W and 100E, in the east. The vegetation is the semiarid savannah, the area contained the wild tree plants with nutritional, ethnomedicinal and economical values, such as Acacia nilotica, Parkia biglobosa, Khaya senegalensis, Prosopis africanum, Azadirachta indica, Vitalleria paradoxa, Vitex doniana, Tamarindus indica and Ficus exasperate just to mention but a few (Ibrahim et al., 2012).
2.2 Seed Authentication
The pods containing the seeds were collected between December- January, and brought to the botanical garden of Biological Science Department, University of Abuja, which was identified by Mr. O. Segun and Verified by Professor O. Olorode (Botanist/Taxonomist) before taken to the laboratory for processing.
2.3 Seed Processing
The pods were crushed using mortar and pestle and blown to obtain the seeds .Using laboratory hot plate (Hot Hc 1200, UK, BIBBY) the seeds were boiled for 5hours at 1000C and dehulled, after cooling. The dehulled seeds were oven dried at 60°C for twenty-four (24) h, using (Memmert oven Schutzari 40050-1p 20 Germany) to a constant weight, then cooled and ground to powder (flour), using Mortar and Pestle and stored at 40C prior to use.
2.4 Proximate Analysi
Standard methods of the Association of Official Analytical Chemists (AOAC, 1990) were used to determine the moisture, crude protein, crude lipid, ash content and crude fibre contents of the sample.
Moisture content was determined by weighing 2g of the flour (W1) into preweighed crucible (Wo) and placed into a hot drying oven at 105oC for 3h. The crucible was removed, cooled in desicator and weighed. The process of drying, cooling and weighing were repeated until a constant weight (W2) was obtained. The weight loss due to moisture was obtained by the equation:
Moisture (%) = x 100
Where:
Wo= Weight of the empty crucible (g), W1= weight of the powder sample + empty crucible (g) W2= weight of dried sample + empty crucible (g).
Crude protein (% total nitrogen x 6.25) was determined by the Kjeldahl method, using 2g of the seeds flour;

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 29
Crude lipid was obtained by exhaustively extracting 5g of the sample in a Soxhlet apparatus using petroleum ether (boiling point range 40-60°C) as the extractant.
Ash content was determined by the incineration of 10g sample and placed in a muffle furnace maintained at 550°C for 5h.
Crude fibre was obtained by digesting 2g of sample with H2SO4 and NaOH and incinerating the residue in a muffle furnace maintained at 550°C for 5h.
Carbohydrate value was obtained by (percentage of ash + percentage of Fat + percentage of Protein + percentage of crude fibre.
Each analysis was carried out in triplicate.
2.5 Phytochemical Screening of A. Sieberiana Seed Flour
Sixteen (16) preliminary qualitative phytochemicals screening were conducted, in accordance
with the standard procedure (Harborne, 1974; 1992; Debela, 2002; Sofowora, 1993). Test for alkaloids, steroids, tannins, saponins, glycoside, cardenolides, terpenoid, reducing sugar, flavonoids, cardiac glycosides, balsams, resins, volatile oils, phlobatannins, phenols, and triterpenoids tests are all conducted, on both aqueous and methanolic flour extract.
2.6 Sample Digestion For Elemental Analysis
About 2g of the sample was digested with (Aqua regia) 3ml of HCL to 1ml OF HNO3 until a clear solution was observed; the suspension was then filtered into a 100m volumetric flask and made it up to the mark (100ml) using deionized water. Atomic absorption spectrophotometer (Younglin AAS 8010 Model, Korea) was used for the determination.
2.7 Germination Trials
One hundred seeds were subjected to various treatments for breaking the dormancy of A. sieberiana for its propagation, treatments such as soaking the seeds in concentrated H2SO4, and 50% H2SO4, Concentrated (nitric acid) HNO3 and 50% HNO3 for 1hrs each, Boiling the seeds at 100oC, heating the seeds at 600C, using (water bath), socking the seeds in cold water for 24hrs, mechanical treated and untreated seeds were all planted in polyethene bag containing soil for four weeks germination trials. Mulching the seeds was also attempted.
3. Results and Discussions
The proximate composition results in table 1 shows that Moisture content of A. sieberiana seeds flour was 6.2±0.00% which is low compared to that of Parkia biglobosa 11.21%, reported by Elemo et al., (2011). The low moisture content will afford a long shelf life for A. sieberiana seeds flour. The crude lipid is high 30.6±0.02% compared with that of Parkia biglobosa 15.48%, Elemo et al., (2011) pigeon pea 1.7% and soybeans 23.5% (Paul and Southgate, 1985). This is an indication that A. sieberiana seeds can be grouped as part of plants oil-rich legume seeds. The most interesting value is that of crude protein 49.7±0.03 which is considered very high compared to protein rich foods such as soyabeans 36.00% (Edema et al.,

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 30
2005), pigeon pea 21.0% (Abdelrahman et al., 2010), prosopis africanum 23.6±1.5 (Aremu et al., 2006) and Parkia biglobosa 27.9% (Elemo et al., 2011).
Table 1. Proximate constituents of A. sieberiana seed flour in percentages (%)
S/No. Constituents (%) Seeds flour 1 Moisture content 6.2±0.00
2 Ash Content 5.8±0.04
3 Crude lipids 30.6±0.02
4 Crude fibre 4.9±0.29
5 Carbohydrate 8.2±0.32
6 Crude protein 49.7±0.03
Values are mean ± standard deviation of triplicate.
Phytochemicals screening of aqueous and ethanolic extract of the seeds flour in table 2, revealed the present of alkaloids, which generally exert pharmacological activity particularly in mammals such as humans, many of our most commonly used drugs are alkaloids from natural sources (Robert and Wink 1998). Alkaloids have many pharmacological activities including antihypertensive effect, antiarrhythmic effect, antimalarial activity and cancer action these are few functions of alkaloids illustrated by Cordell (1983).Tannins are also present which serve as natural defense against microbial infection. Tannins have also been reported to exert other physiological effects, such as to accelerate blood clotting, reduce blood pressure, decrease the serum lipid level, produce liver necrosis and modulate immunoresponses (Bele et al., 2012). The seed flour also contains terpenoids, terpenes have biological activities and are also used for the treatment of human diseases. The worldwide sales of terpenes–based pharmaceuticals in 2002 were approximately US $12billion. Among these pharmaceutical, the anticancer drug taxol, and the antimalarial drug artimesinin are two of the most renowned terpenoids based drugs (Lixin and Arnold, 2005). Saponins are also present, a secondary plant metabolite well known for antimicrobial properties. Flavonoids are polyphenols known for antioxidants activity improve blood circulation and relieve tissues damaged by pathogens. Flavonoids play a role in pigmentation and also have anti-inflammatory properties. The seeds flour also contains volatile oil which is a component of plant that offers the strong aroma to attract foraging wildlife, and this oil often provides concentrated medicinal benefits as well. Some components of volatile oil are strongly antiseptic and normally possess anti-inflammatory properties as well. Aromatherapy involves the use of volatile (essential) oil to alter moods and perception as well as to detoxify the body (Kirk, 2009). Cardiac glycoside is also present which is the natural diuretics that directly strengthen the heart by increasing the contraction rate, cardiac glycosides also improve circulation, lower blood pressure and relieve the urinary tract.
Table 2. Phytochemicals screening of aqueous and ethanolic A. sieberiana seeds flour.
S/No. Test Results
1 Alkaloids +

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 31
2 Steroids -
3 Tannins +
4 Saponins +
5 Glycosides +
6 Cardenolides -
7 Terpenoides +
8 Reducing sugar -
9 Flavonoids +
10 Cardiac glycosides +
11 Balsams -
12 Resins -
13 Volatile Oil +
14 Phlobatannins -
15 Phenols -
16
Triterpenoids -
KEY:- (+) Indicate present. (-) Indicate absent
From the table 3 below the concentrations of the essential elements appeared to be low but within safety limit (W. H. O., 1996). These elements support human biochemical processes by serving structural and functional roles as electrolytes (Nelson and Cox, 2008). The values of Calcium (2.02±0.5), Magnesium (3.93±0.03), Cupper (0.19±0.01), and Manganese (0.02±0.00) are considerable, as they play important roles in metabolic activities, manganese (Mn) used extensively in chemotherapy and others are known to help in bone and teeth development (Khan, 1996; Ogugbuaja et. al., 1997). The result of the minerals content of A. sieberiana was also comparable with minerals analysis on Pakia biglobosa seed (Elemo et al., 2011), and Prosopis africanum seed (Aremu, et al., 2006).
The result shows that Cadmium (Cd), and Arsenic (Ar) are not detected and the infinitesimal value of Lead (Pb) 0.001±0.00, indicated that the seed flour contains little or no heavy metals, as heavy metals have been implicated in cancer, liver and kidney problems (Ogugbuaja et. al., 1997). Minerals help maintain acid-base balance, to keep the body pH neutral (Akpanyung, 2005). Minerals help regulate body processes, such as in enzyme systems. They also function in nerve impulse transmission and muscle contraction.
Table 3. Elemental analysis of A. sieberiana processed seeds flour.
S/No. Test Elements Conc. in Mg/g 1 Fe 0.11±0.01
2 Mg 3.93±0.03

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 32
3 Ni 0.045±0.00
4 Ca 2.02±0.5
5 Cu 0.19±0.01
6 Zn 0.15±0.01
7 Cd Nil
8 Pb 0.001±0.00
9 Mn 0.02±0.00
10 Cr 0.02±0.01
11 Ar Nil
Values are mean ± standard deviation of triplicate.
The result of germination trials of A. sieberiana seeds after subjecting each group of 10 seeds to different treatments in Figure 3, shows that out of 100 seeds treated, a total of 50seeds germinated in all. All the seeds (10) treated with cold soaking and mulching germinated, 9 seeds germinated from the group of untreated seeds, 8 seeds germinated from the group that was subjected to boiling at 600c, 7 seeds germinated from the group that was subjected to mechanical treatments, 3,2, and 1 seed(s) germinated from groups treated with 50% HNO3 for (1hr), 50% H2SO4 for (1hr) and boiling at 1000c for (1hr) respectively. Seeds treated with 100% HNO3 for (1hr) and 100% H2SO4 for (1hr) produced no germination. Thus, cold soaking, mulching and natural (no-treatment) prove promising methods of propagating A. sieberiana. The result was in line with the world agroforestry data base (www.worldagroforestry.org/af/treedb/). The data collected from the germination trials were subjected to analysis of variance one way (ANOVA). Means with significant differences were separated by Least Significant Difference (LSD) after Post-hocTest, using SPSS software at
critical value (p ≥ 0.05).
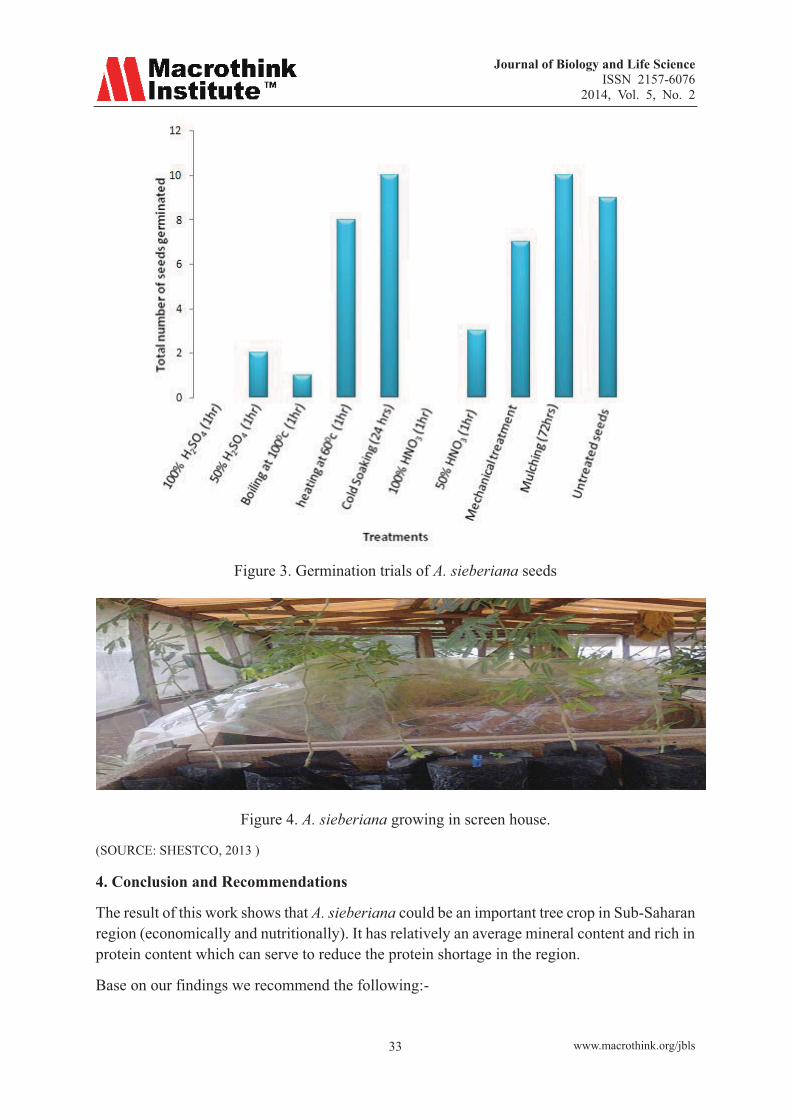
Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 33
Figure 3. Germination trials of A. sieberiana seeds
Figure 4. A. sieberiana growing in screen house.
(SOURCE: SHESTCO, 2013 )
4. Conclusion and Recommendations
The result of this work shows that A. sieberiana could be an important tree crop in Sub-Saharan region (economically and nutritionally). It has relatively an average mineral content and rich in protein content which can serve to reduce the protein shortage in the region.
Base on our findings we recommend the following:-

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 34
There is a growing concern about human destruction of vegetation, it is necessary to collect the germplasm of this untapped nutritive and economically important tree species for conservation before it becomes extinct.
The seeds should be incorporated in agricultural development programmes placing special emphasis on its nutritional value and production potential.
More work should be carried out on its nutritional value i.e. Amino acid profiles, Vitamins, and other nutritive elemental analysis.
Acknowledgement
The authors are grateful to their colleagues in Chemistry Advanced Laboratory and those in Biotechnology Advanced Laboratory (SHESTCO) for their invaluable comments for improving the research work. The authors also warmly acknowledge Mr. Charles Osuji for his help in writing of this manuscript.
References
Abdelrahman Sirelkhatim Mohammed Eltayeb, Ali Osman, & Rea Haron. (2010). The chemical composition of Pigeon pea (Cajanus cajan) seed and functional properties of protein isolate. Pakistan journal of nutrition 9: pp.1069-1073.
Aberoumad A., & S. S. Deokule. (2010). Preliminary studies on proximate and mineral composition of Marchubeh stem (Asparagus officinalis) Vegetable consumed the Behbahan of Iran. World Applied Science. Journal, 9, 127-130.
Akpanyung, E. O. (2005). Proximate and mineral composition of bouillon cubes produced in Nigeria. Pakistan Journal of Nutrition, 4, 327-329. http://dx.doi.org/10.3923/pjn.2005.327.329
Akubugwo, I. E., A. N. Obasi, & S.C. Ginika. (2007). Nutritional potential of the Nutritional potential of the leaves and seeds of black night shade- solanum nigrum L.Var Virginicum from afikpo-Nigeria. Pakistan Journal of Nutrition. 6: pp. 323-326. http://dx.doi.org/10.3923/pjn.2007.323.326
AOAC. (1990), Official Methods ofAnalysis.15th Edition, Association of Official Analytical Chemists Washington DC., Alington, Virginia., USA, ISBN: 0-935584-42-0.
Aremu, M. O., Olonisakin, A., Atolaye, B. O., & Ogbu, C. F., (2006). Some nutritional and functional studies of Prosopis africanum. Electronic Journal of Environment and Agriculture Food Chemistry. 5, 1640-1648.
Bele, A. A., V. M. Jadhav & V. J. Kadam. (2010). Potentials of Tannins: A review. Journal of plants science 9, 209-214.
Cordell, G. A. (1983). Introduction to Alkaloids: A Biogenic Approach, Wiley, New York.
Debela, A. (2002). Manual for photochemical screening of medicinal plants. Ethiopian Health and Nutrition Research Institute, Addis Ababa, Ethopia.pp 35-47.
Edema, Mojibola O. Sanni, Lateef I., & Sanni, Abiodun I. (2005). Evaluation of Maize-

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 35
soybean flour blends for sour maize bread production in Nigeria. African journal of biotechnology, 4, 911-918.
Elemo, Gloria N., Babajide O. Elemo, Oladunmoye, & Ochuko L. Erukainre. (2011). Comprehensive investigation into the nutritional composition of dehulled and defatted African locust bean seed (Pakia biglobosa). African journal of plants science. 5, 291-295.
Harborne J. B. (1992). Photochemical methods. Chapman and Hall publications, London. pp. 7-8.
Harborne, J. B. (1974). Textbook of Phytochemicals method: A guide to modern technique of plant analysis. 2nd edition, Champenan and Hall Limited. pp. 99-103.
Ibrahim, H. A. Ali, G. Y. Halliru, Usaini S., & Abdullahi I. I. (2012). Ethnobotanical Survey of the Wild Edible Food Plants Consumption among Local Communities in Kano State, North-Western, Nigeria. International Journal of Science and Technology, 2(10), 713-717
James, C. S., (1995). Analytical Chemistry of Food. Champman and Hall, London, 64-65. http://dx.doi.org/10.1007/978-1-4615-2165-5
Khan, I. Z. (1996). The role of Chemistry in health disease and ageing. A seminar presentation in the Department of chemistry, University of Maiduguri, Nigeria.
Kirk Patric. (2009). Sixteen Active Plant Components Explained. The Encyclopedia of Medicinal Plants.
Lixin Zhang, Arnold L. Demain. (2005). Natural product; Drug Discovery and Therapeutic Medicine. Humana press Inc., Totowa. Nj. pp.197.
National Academy of Science. (1984). Underexploited Tropical plants with Promising Economic Value. Washington, D.C, Seventh printing, pp, 1-2.
Nelson, D. L., & M. M. Cox, (2008). Lehninger Principles of Biochemistry. 5th Edn. W.H. Freeman and Company. Madison Avenue, New York, pp: 343.
Ogugbuaja, V. O, Akinniyi, J. A., Abdulrahman, F. I., & Ogarawu, V. C., (1997). Elemental Contents of Medicinal plants. A monograph. Department of chemistry faculty of science, University of Maiduguri, Maiduguri, Nigeria.
Orwa C, Mutua A, Kindt R, Jamnadass R, & Simons A. (2009). Agroforestree Database: a tree reference and selection guide, version 4.0 (http://www.worldagroforestry.org/af/treedb/).
Paul, A. A., & Southgate, D. A. T. (1985). McCance and Widdowsen’s The Corporation of Foods Royal Society of Chemistry. London.
Roberts Margaret F., & Michael Wink (1998). Alkaloids/ Biochemistry, Ecology and Medicinal Application. Plenum press. New York and London pp. 5-6.
Sofowora A. (1993). Medicinal plants and Traditional medicine in Africa. Spectrum Books Ltd, Ibadan, Nigeria, pp. 289.

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 36
WHO. (1996). Guidelines for elemental concentration, trace elements in health and human nutrition, pp. 50-68.
Copyright Disclaimer
Copyright reserved by the author(s).
This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 37
Effects of Occupational Exposure to Wood Smoke in Tandoor Occupants
Ghulam Nabi
Student, Department of Zoology, University of Malakand Chakdara
Dir (L), Khyber Pakhtunkhwa, Pakistan
E-mail: [email protected]
Jeena Urooj
Student, Department of Zoology, University of Malakand Chakdara
Dir (L), Khyber Pakhtunkhwa, Pakistan
E-mail:[email protected]
Ayaz Ali Khan (Corresponding Author)
Assistant Professor, Department of Biotechnology, University of Malakand Chakdara
Dir (L), Khyber Pakhtunkhwa, Pakistan
E-mail: [email protected]
Ghazala Yasmin Zamani
Lecturer, Department of Biotechnology, Bacha Khan Unviersity Charsada
Khyber Pakhtunkhwa,Pakistan
E-mail: [email protected]
Mussarat Wahid
M. Phil, Department of Bio Sciences
Comsats Institute of Information Technology, Islamabad
E-mail: [email protected]

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 38
Saleem-Ur-Rahman
Student, Department of Biotechnology, University of Malakand Chakdara
Dir (L), Khyber Pakhtunkhwa, Pakistan
E-mail: [email protected]
Received: August 3, 2013 Accepted: August 17, 2013
doi:10.5296/jbls.v5i2.4079 URL: http://dx.doi.org/10.5296/jbls.v5i2.4079
Abstract
Exposure to wood smoke leads to mucosal irritation and physiological and neurological abnormalities in human.Blood samples along with histories were taken from 100(50Tandoor occupants and 50 controls) male individualsliving in the same locality. SYSMIX KX-21 (Japan) and Shimadzu Double Beam Spectrophotometer 1700 Pharma (Japan) were used for blood profiling and serum biochemistry. In workers, blood cholesterol was 131 ± 4.8 mg/dl, triglyceride was 125 ± 10mg/dl, and serum glutamate pyruvate transaminase level was 68 ± 2.4IU/L and glucose level was 113 ± 2.1 mg/dl. The total red blood cell count was 5.6 ± 0.035mill/cmm, hemoglobin level was15 ± 0.054mg/dl, hematocrit value was49 ± 0.22 mg/dl, mean corpuscular volume was 86 ± 0.22 fl,mean corpuscular hemoglobin was 31 ± 0.12 pg, mean corpuscular hemoglobin concentration was 35 ± 0.12gm/dl, neutrophils were 77 ± 0.50%, lymphocytes were 45 ± 1.0 %, eosinophilswere6.0 ± 0.14%, monocytes were 7.7 ± 0.19 %, total leukocyte count were 11014 ± 115/cmm, and platelets were235864 ± 5491 /cmm. In control group the blood cholesterol, triglyceride, serum glutamate pyruvate transaminase and glucose level was111 ± 2.4mg/dl,97 ± 1.5mg/dl, 28 ± 1.1IU/L and 101 ± 1.2 mg/dl respectively, while thetotal red blood cell count, hemoglobin level, hematocrit value, mean corpuscular volume, mean corpuscular hemoglobin, mean corpuscular hemoglobinconcentration, neutrophils, lymphocytes, eosinophils, monocytes, leukocyte, and platelets were4.3 ± 0.027mil/cmm, 12 ± 0.064 mg/dl, 38 ± 0.11mg/dl, 75 ± 0.30fl, 25 ± 0.37pg, 31 ± 0.11gm/dl, 56 ± 0.82%, 34 ± 1.0, 3.1 ± 0.20%, 4.2 ± 0.12%, 6573 ± 125, and 169162 ± 2454 /cmm respectively.The levels in both groups were statistically compared using t test and the p value was determined.The results showed that there was significant increase in blood biochemical parameters as well as in complete blood count in workers as compared to control and are therefore at more risk to heart, lungs and blood diseases.
Keywords: Tandoor, Wood smoke, Hematology, Blood biochemical parameters
1. Introduction
A Tandoor is a cylindrical clay oven used for cooking and baking in many countries like Pakistan, India, Turkey, Iran, Afghanistan, Burma and Bangladesh (Steven 2011). Burning wood used in Tandoor emits toxic particles in air that contains carbon monoxide (CO),

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 39
Nitrogen Oxides (NOx), Volatile Organic Compounds (VOCs), Dioxin, and Inhalable Particulate Matter (PM) (Minnesota Pollution Control Agency 2012) due to which cardiovascular, coronary artery diseases, cerebro-vascularsdiseases and venous thrombo-embolism occurred in people that are exposed (Fraanchini 2011). According to the recent studies, each year, ten of thousands of premature cardiovascular death occurs, the mechanisms of which remain uncertain (Schwartz 2001). Long time exposure also leads to death from coronary heart diseases (Nemmar et al 2006). Air pollutant when inhale, affects blood pressure (Simkhovich et al 2008). The improved stove intervention was associated with 3.7mm Hg lower systolic blood pressure (SBP) after adjusting for age, body mass index, smoking, second hand tobacco smoke, apparent temperature, season, day of week and time of day (Mccracken 2010). Exposure to wood smoke leads to irritation of the mucous as well as physiological and neurological effects in human (Riddervold et al 2011) and they causes reduced lung function in rural Africans (Fullerton et al2011). The particulate matter in kitchen is associated with chronic obstructive pulmonary diseases and lung cancer in women (Perez et al 2010).In Pakistan 94% of households in rural areas and 58% in urban areas depend on biomass fuels (wood, dung, and agricultural waste) due to incomplete combustion of the biomass fuels, the resulting smoke contains a range of health-worseningsubstances that, at varying concentrations, can causea serious threat to human health. Indoor air pollution accounts for 28,000 deaths a year and 40 million cases of acute respiratory illness. It places a significant economic burden on Pakistan with an annual cost of 1% of GDP (Colbeck et al 2010). The present study was conducted to study the general health status of Tandoor occupants exposed to wood smoke and its effects on blood biochemical parameters of the occupants and hematological parameters.
2. Materials and Methods
Questionnaire
For the present study,a standard questionnaire was formulated for collecting specific information from Tandoor occupants and control group regarding their age, marital status, type of fuel, addiction, health status, exposure time to smoke and other particulars. The study was approved by the ethical board of University of Malakand.
Data collection
The study was approved by the ethical board of University of Malakand. During sampling, 100 (50 workers irrespective to their age and working hours in Tandoor and 50 controls) maleindividuals were selected. A written consent was signed from all individuals before taking interviews.The blood samples from Tandoor occupants and healthy individuals (Control) were collected from (Chakdara, Gulabad and Tazagram), at Dir (Lower), Khyber Pakhtunkhwa, Pakistan, that was made in October, 2011 till April,2012.
Blood sampling from Tandoor occupants
Blood sample (5 ml) was taken from the workers and 3 ml of the blood was transferred to heparinized test tube for blood profiling. Blood sample (2 ml) was transferred to other test tube having no any anticoagulant for isolation of serum that was used for biochemical tests. The

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 40
samples were immediately analyzed and those which were kept pending, transferred to -20°C refrigerator for storage.
Blood sampling from control
The people living all around the area near Tandoor shops were kept as control. A total of 50 people were randomly selected from those areas. Same procedure was adopted for collection of blood and isolation of serum as that of workers.
Serum isolation
Test tubes were kept in slant position. Serum oozed out, and transferred to the other sterilized test tubes. Serum was used for analysis of different chemicals (Cholesterol, Glucose, Serum Glutamate Pyruvate Transaminase (SGPT) and Triglycerides (TG) in blood.
Equipment used
SYSMIX KX-21(Japan) was used for the determination of complete blood profile and Shimadzu Double Beam Spectrophotometer 1700 Pharma (Japan) was used for blood biochemistry.
Statistical analysis
The data obtained was statistically analyzed. Mean, standard deviation, column statistics/row statistics andt test were used for the comparison of both the groups (control and workers) were performed for each parameter using Graphpad Prism, Demo version 05 (www.graphpad.com).
3. Results and Discussions
The present study is based on studying the effects of Tandoor smoke on health status of the occupants. The adverse effects of the toxicants in wood smoke are a common phenomenon in all the developing countries. The awareness about this problem is lacking in Pakistan, particularly in Khyber Pakhtunkhwa. To collect the specific information from workers as well as control, a questionnaire was formulated regarding their age and health status. The information collected was summarized (Table.1).The results of biochemical parameters such as Cholesterol, Triglycerides, Glucose, SGPT and complete blood profile of both workers and control are listed(Table.2).
History of workers and control
In the present study, both workers and control group were divided into different age groups. Control group was mostly normal (Table. 1) while workers, who were frequently exposed to wood smoke, had hypertension, lungs abnormalities, chest pain and physical weakness (Table. 1). These problems were correlated with age and thus more prominent in elder people than young ones.Kodgule and Salvi 2012 stated that increased mortality and morbidity is associated with exposure to biomass smoke. Minnesota Pollution Control Agency2012 have also given same report that irritation of eyes, lungs, throat and sinuses, increases severity of existing lung diseases such as asthma, emphysema, pneumonia and bronchitis, and chronic obstructive lung disease and chronic bronchitis are associated with short term and long term exposure to wood smoke.Kunzliet al2006studied exposure to heavy smoke of wildfires and concluded thatthe nose, eyes, and throat irritations, cough, bronchitis, cold, wheezing, asthma attacks were associated with

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 41
individually reported exposure differences within communities.Rajpandey 1984 found that wood smoke inhalation causes chronic bronchitis in adults; chronic interstitial pneumonitis andfibrosis (Ramage et al 1988), interstitial lung disease, pulmonary arterial hypertension (Sandoval et al 1993), and altered pulmonary immune defense mechanisms (Demarest et al 1979 and Ramage et al 1988).
Table 1. History of workers and control
Age Group (Years)
No. of Workers
Economic status
Marital Status
Type of
Fuel used
Exposure Time
(Months)
Addiction (%)
Use of
Mask
Health status
(Before Job) (%)
Health status (After Job)
(%)
Workers 18-25
20 Poor 15 Unmarried 05 married
Wood 72-90 09 Snuff 00 Cigarettes
No No Specific
06 RTI*, 03 Rhinitis, 08 Eye discharge, 02 Hypertension
26-33 15 Poor 06Unmarried 09 married
Wood 120-140 10 Snuff 01Cigarettes
No 02 GIT* disorders.
05 RTI*, 03 Rhinitis, 02 Eye discharge, 05 Hypertension
34-41 10 Poor 02Unmarried 08 married
Wood 110-120 10 Snuff 00 Cigarettes
No 01 G.B.A*.
10 RTI*, 05Rhinitis, 05 Eye discharge 00 Hypertension
42-49 5 Poor 00 unmarried
05 Married
Wood 140-160 07 Snuff 02 Cigarettes
No 05 Physical weakness
03RTI*, 03Rhinitis, 02 Eye discharge, 01Hypertension, 02 Lumbago, 04 Chest pain
Control 18-25
22 Poor 17 Unmarried 05 Married
No 00 03 Snuff 00Cigarettes
No 00 00
26-33 13 Poor 01 Unmarried 12 Married
No 00 06 Snuff 00 Cigarettes
No 00 00
34-41 8 Poor 00 Unmarried 08 Married
No 00 03 Snuff 00 Cigarettes
No 00 00
42-49 7 Poor 00 Unmarried
No 00 00 Snuff 00
No 00
00

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 42
07 Married Cigarettes
*RTI: Respiratory Tract Infection, *GIT: Gastro Intestinal Tract, *GBA: General Body Aches,
Blood Cholesterol
The mean and standard deviation of workers was high (131 ± 4.8mg/dl) as compared to control (111 ± 2.4 mg/dl) for cholesterol (Table2).Particulate matter coming from fossil fuel can cause atherosclerosis, thrombosis (Nemmar et al 2006),affects blood pressure(Simkhovich et al 2008) and increases risk of coronary events(Bareegard et al 2006). It may be concluded that wood smoke has an ill effect on health by raising the blood cholesterol level.
Blood Triglycerides
Blood triglyceride level in workers was high (125 ± 10 mg/dl) as compared to control (97 ± 1.5mg/dl) (Table 2).The smoking raises triglyceride level (American Heart Association 1983)and raises the Low Density Lipid (Mikhailidi 1998).
Serum Glutamate Pyruvate Transaminase (SGPT)
SGPT level of workers and control was 68 ± 2.4 IU/L and 28 ± 1.1 IU/L (Table 2). Workers have high SGPT level. The serum alanine aminotransferase (ALT) about 1.5 fold the normal limit is caused by the inhalation of smoke having high concentration of Hydrocarbons and Chlorinated vapors (Loh et al 2006). Among HCV seropositive subjects smoking and alcohol consumption significantly elevated ALT level (Wang et al 2002).
Blood Glucose
There was a significant increase in blood glucose level of workers (113 ± 2.1 mg/dl) as compared to control group (101 ± 1.2mg/dl) (Table 2). Same results have also been given byBornemisza and Suciu 1980 after smoke inhalation and blood glucose level raised as compared to control group.
Blood profile
Total Red Blood Cells (TRBC) count
The TRBC in workers was5.6 ± 0.035 mil/cmm and in control was 4.3 ± 0.027 mil/cmm(Table 2). Workers have high TRBC than control. Smoking at high altitude further increases RBC count(Ramirez et al 1991). Roethig et al2010stated that by stopping smoke cigarette, a significant decrease up to 9% in RBC is reported within three days. It has been concluded that wood smoke increases total red blood cell count.
Hemoglobin
The hemoglobin level in workers was 15 ± 0.054mg/dl and in control was 12 ± 0.064mg/dl(Table 2). The smoking increases the hemoglobinlevel both in men and women (Nordenberg et al1990 andBinkin et al 1993).
Hematocrit Value
The Hematocrit level in workers was 49 ± 0.22mg/dl while in control was 38 ± 0.11mg/dl(Table 2).

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 43
The present study is also in correlation with Goldbourt and Medalie1977,who determined that current smokers have high hematocrit level than non smokers.
Mean corpuscular Volume
The MCV in workers was 86 ± 0.22fl and in control was 75 ± 0.30fl(Table 2). Same results have been reported by Kondo et al1993.
Mean Corpuscular Hemoglobin
The MCH for workers was 31 ± 0.12pg and for control, was 25 ± 0.37pg(Table 2).The results are in agreement with Kondo et al1993, who found that in smokers MCH level is high. It has beenconcluded that MCH level is raised when compared to control.
Mean Corpuscular Hemoglobin Concentration
The MCHC was high in workers (35 ± 0.12gm/dl) as compared to control (31 ± 0.11gm/dl)(Table 2). Similar resulthas also been reported by Kondo et al1993,who found that individual chronically exposed to smoke has higher level of MCHC.
Neutrophil
In control, neutrophils count was 56 ± 0.82%, while in workers it was77 ± 0.50%(Table 2).Human exposure to diesel particles for 1 hour at 300 microgram/cubic meter resulted in increased level of peripheral neutrophils(Schwartz 2001).
Eosinophils
For workers, the mean value of eosinophil was 6.0 ± 0.14% and for control was 3.1 ± 0.20%(Table 2).Lal et al1993obtained similar results by exposing rats to wood smoke for 30 and 45 days and found increased number of esosinophils in the blood.
Lymphocytes
There was a significant increase in lymphocyte count of workers (45 ± 1.0 %) as compared to control (34 ± 1.0 %)(Table 2).Lymphocyte percentage is associated with smoking and is therefore higher in smokers (Huang et al 2001 and Rutgerz et al 2000).
Monocytes
The mean count of monocyte for workers was 7.7 ± 0.19% while for control was 4.2 ± 0.12% (Table 2). Present results are in agreement with the finding of Adams et al1997and Huang et al2001. They found that smoke increases monocyte count. Ray et al2006found that biomass smoke exposure causes activation of monocyte.
Total Leukocytes Count
The TLC was high in workers (11014 ± 115/cmm) as compared to control (6573 ± 125/cmm)(Table 2).Thesmoke exposure increase white blood cells count (Yanbaeva et al 2007 and Megan 2003). Kondo et al1993studied the relationship of cigarette smoke and hematological parameters and analyzed that smoking increases TLC.

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 44
Platelets
The mean value of platelets for workers was 235864 ± 5491/cmm and for control were169162 ± 2454 /cmm(Table 2).Milton and Cuyler1956, Reed et al2006,Ray et al2006and Sithu et al2010havestated same results.
Table 2. Comparative analysis of the blood biochemical parameters of the control and Tandoor occupants.
Parameters
Control Workers 95% C.I
P value
Mean ±SD
Mean ±SD Glucose 101 ± 1.2 113 ± 2.1 -16 to -6.7 < 0.0001 Lymphocytes 34 ± 1.0 45 ± 1.0 -14 to -8.5 < 0.0001 Platelets 169162 ± 2454 235864 ± 5491 54749 to 78654 < 0.0001 SGPT 28 ± 1.1 68 ± 2.4 -45 to -34 < 0.0001 Hb% 12 ± 0.064 15 ± 0.054 2.8 to 3.1 < 0.0001 HCT 38 ± 0.11 49 ± 0.22 10 to 11 < 0.0001 Cholesterol 111 ± 2.4 131 ± 4.8 -31 to -9.3 .0003 MCH 25 ± 0.37 31 ± 0.12 5.5 to 7.1 < 0.0001 MCHC 31 ± 0.11 35 ± 0.12 3.2 to 3.8 < 0.0001 MCV 75 ± 0.30 86 ± 0.22 10 to 12 < 0.0001 Neutrophil 56 ± 0.82 77 ± 0.50 -23 to -19 < 0.0001 Eusinophils 3.1 ± 0.20 6.0 ± 0.14 -3.4 to -2.4 < 0.0001 Monocytes 4.2 ± 0.12 7.7 ± 0.19 -3.9 to -3.0 < 0.0001 TG 97 ± 1.5 125 ± 10 -48 to -8.0 0.0066 TLC 6573 ± 125 11014 ± 115 -4778 to -4104 < 0.0001 TRBC 4.3 ± 0.027 5.6 ± 0.035 1.2 to 1.4 < 0.0001
SD= Standard Deviation, C.I= Confidence Interval
4. Conclusion
From the present study, it is concluded that wood smoke has harmful effects on the persistently exposed people. In workers,wood smoke exposure leads to hypertension, lungs infection, dry cough, chest pain and physical weakness. In adults such problems were more severe as compared to young ones. It has been concluded that workers are at more risk to heart and liver diseases because the biochemical parameters such as SGPT, blood cholesterol, blood triglyceride and blood glucose level were found high as compared to control group. Such workers are also at more risk of getting lungs infections/abnormalities as compared to control group.
Recommendations
To check the effects of chronic exposure to wood smoke and its toxicants; a study on large population should be performed, and a work on female Tandoor occupants will also be necessary in the same locality. The printed as well as electronic media should aware the people from the hazardous effects of wood smoke generating during cooking and baking process.

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 45
Limitations
In present study, a small number of population sizeand only the male individuals were considered. The toxicants inindoor environment of work place were not measured due to unavailability of facilities.Toxicants were not measured in blood, it is recommended to check the toxicants in indoor environment as well as in blood to co-relate the results and to boost the validity of the results.
References
Adams, M. R., Jessup, W.,&Celermajer.(1997). Cigarette smoking is associated with increased human Monocyte adhesion to endothelial cells. Journal of American College of Cardiology, 491-497. http://dx.doi.org/10.1016/S0735-1097(96)00537-2
American Heart Association, Triglyceride, (1983). www.americanheart.orq
Barregard, L., Sallste, G., Andersson, L., Almstmansd, A. C., Gustafson, P., Andersson, M., & Olin, A. C. (2006). Experimental exposure to wood-smoke particles in healthy humans: Effects on markers of inflammation - coagulation and lipid peroxidation. Inhalation Toxicology, 845-53. http://dx.doi.org/10.1080/08958370600685798
Binkin.,Nancy, J., Nordenberg., Dale., Yip., & Ray. (1993). The effect of cigarette smoking on hemoglobin level and anemia screening. Journal of American Medical Association, 1556-9.
Bornemisza, P., & Suciu, I. (1980). Effect of cigarette smoking on the blood glucose level in normal and diabetics.Medicine Interne, 353-6.
Colbeck, I., Nasir, Z. A., & Ali, Z. (2010). The state of indoor air quality in Pakistan- a review.Environmental Science and Pollution Research International, 17(6), 1187-96. http://dx.doi.org/10.1007/s11356-010-0293-3
Franchini, M., & Mannucci, P. M. (2011). Thrombogenecity and cardiovascular effects of ambient air pollution. Journal of the American College of Cardiology, 52(9), 719-26.
Fullerton, D. G., Suseno, A., Semple, S., Kalambo, F., Malambo, R., White, S., Jack, S., Calverly, P. M., & Gordon, S. B. (2011). Wood smoke exposure, poverty and impaired lung function in Malawian adults. International Journal of Tuberculosis and Lung Diseases, 15(3), 391-8.
Goldbourt, U., & Jack, H. M. (1977). Characteristics of smokers, non- smokers and ex- smokers among 10,000 adult males in Israel. American Journal of Epidemiology, 75-86.
Huang, Z. S., Chien, K. L., Yang, C. Y., Tsai, K. S., & Wang, C. H. (2001). Peripheral differential leukocyte counts in humans vary with hyperlipidemia, smoking, and body mass index. Lipids, 237-45. http://dx.doi.org/10.1007/s11745-001-0713-9
Kondo, H., Kusaka, Y., & Morimoto, K. (1993). The effects of life style on hematological parameters: Analysis of hematological data in association with smoking habit and age. SangyoIgaku, 98-104. http://dx.doi.org/10.1539/joh1959.35.98
Lal, K., Dutta. K. K., Vachhrajani, K. D., Gupta, G. S., & Srivastava, A. K. (1993). Histomorphological changes in lung of rats following exposure to wood smoke. Indian Journal of

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 46
Experimental Biology, 761-4.
Loh, C. H., Chang, Y. W., Lious, S. H., Chang, J. H., & Chen, I. (2006). Case report: Hexa chloroethane smoke inhalation: A rare cause of hepatic injuries. Enviromental Health Perspectives, 763-5. http://dx.doi.org/10.1289/ehp.8635
Mccraken, J. P., Smith, K. R., Mittleman, M. A., & Schwartz, J. (2010). Chimney stove intervention to reduce long term wood smoke exposure lowers blood pressure among Guatemalan women. Environmental Health Perspectives, 115(7), 996-1001. http://dx.doi.org/10.1289/ehp.9888
Megan, R., & Smith. (2003). Smoking status and differential white cell count in men and women in the EPIC- Norfolk population.Atherosclerosis, 331-337.
Mikhailidis, D. P. (1998). Smoking, Diabetes and Hyperlipidaemia. Journal of the Royal Society of Health, 91-3. http://dx.doi.org/10.1177/146642409811800209
Milton, E. E., & Cuyler, H. (1956). The effects of smoking on packed cell volume, RBC and platelets count. Cannadian Medical Association Journal, 520-53.
Minnesota Pollution Control Agency. (2012). Health Effects of Wood Smoke; Home, Air, Air Quality and Pollutants, General Air Quality, Wood Smoke.http://www.Pca.state.mn.us/indexphp/air/aiqualityandpollutants/generalairquality/woodsmoke/healtheffectswoodmokhtml. accessed onOctober, 2012.
Nemmar, A., Hoylaerts, M. F., & Nemery, B. (2006). Effects of particulate air pollution on hemostasis.Clinics in Occupational and Enviromental Medicine, 865-81.
Nordenberg, D., Ray, Y., & Nancy, J. B. (1990). The effects of cigrate smoking on hemoglobin and anemia screening. Journal of American Medical Association, 1556-1559. http://dx.doi.org/10.1001/jama.1990.03450120068031
Perez, P. R., Schil, M. A., & Riojas, R. H. (2010). Respiratory health effects of indoor air pollution. International Journal of Tuberculosis and Lung Diseases, 14(9), 1079-86.
Ramirez, G., Bittle, P. A., Colice, G. L., Herrera, R., Agosti, S. J., & Foulis, P. R. (1991). The effects of cigarette smoking upon hematological adaptation to moderately high altitude living. Wilderness Medicine, 274-286. http://dx.doi.org/10.1580/0953-9859-2.4.274
Ray, M. R., Mukherjee, S., Roychoudhury, S., Bhattacharya, P., Banerjee, M., Siddique, S., Chakraborty, S., & Lahiri,T. (2006). Platelet activation, upregulation of CD11b/CD18 expression on leukocytes and increase in circulating leukocyte-platelet aggregates in Indian women chronically exposed to biomass smoke. Human and Experimental Toxicology, 25(11), 627-35. http://dx.doi.org/10.1177/0960327106074603
Reed, M. D., Campen, M. J., Gigliotti, A. P., Harrod, K. S., McDonald, J. D.,Seagrave, J. C., Mauderly, J. L., & Seilkop, S. K. (2006). Health effects of subchronic exposure to environmental levels of hardwood smoke.Inhalation Toxicology, 523-39. http://dx.doi.org/10.1080/08958370600685707
Riddervold, I. S., Bonlokke, J. H., Molhave, L., Massling, A., Jenson, B., Gronborg, T. K., Bossi, R.,

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 47
Forchhammer, L., Kiargaard, S. K., &Sigsgaard, T. (2011). Wood smoke in a controlled exposure experiment with human volunteers.Inhalation Toxicology, 23(5), 277-88. http://dx.doi.org/10.3109/08958378.2011.567401
Roethig, H. J., Koval, T., Muhammad, K. R., Jin, Y., Mendes, P., & Unverdorben, M. (2010). Short term effects of reduced exposure to cigarette smoke on White Blood Cells, Platelates and Red Blood Cells in adult cigarette smokers. Regulatory Toxicology and Pharmacology, 333-7. http://dx.doi.org/10.1016/j.yrtph.2010.04.005
Rutgers, S. R., Postma, D. S., Ten, N. H., Kauffman, H. F., Vander, M. T. W., Koeter, G. H., & Timens, W. (2000). Ongoing air way inflammation in patient with Chronic Obstructive Pulmonary Disease (COPD) who do not currently smoke.Thorax, 12-8. http://dx.doi.org/10.1136/thorax.55.1.12
Schwartz, J. (2001). Air pollution and blood markers of cardiovascular risk.Enviromental Health Perspectives, 405-409.
Simkhovich, B. Z., Kleinman, M. T., & Kloner, R. A. (2008). Air pollution and cardiovascular injury epidemiology, toxicology and mechanisms.Journal of the American College of Cardiology, 719-26. http://dx.doi.org/10.1136/thorax.55.1.12
Sithu, S. D., Srivastava, S., Siddiqui, M. A., Vladykovskaya, E., Riggs, D. W., Conklin, D. J., Haberzettl, P., O'Toole, T. E., Bhatnagar, A., & D'Souza, S. E. (2010). Exposure to acrolein by inhalation causes platelet activation. Toxicology and Applied Pharmacology, 100-10. http://dx.doi.org/10.1016/j.taap.2010.07.013
Steven, R. (2011). A Tandoori Oven brings India's heat to the backyard. New York Times, Wikipedia, Accessed on 2nd December, 2012.
Wang, C. S., Wang, S. T., Chang, T. T., Yao, W. J., & Chou, P. (2002). Smoking and alanine aminotransfease level in hepatitis c virus infection: implifications for prevention of hepatitis c virus progression. Archives of International Medicine, 811-5. http://dx.doi.org/10.1001/archinte.162.7.811
Yanbaeva, D. G., Dentener, M. A., Creutzberg, E. C., Wesseling, G., & Wouters, E. F. (2007). Systemic effect of smoking. Chest, 1557-66. http://dx.doi.org/10.1378/chest.06-2179
Copyright Disclaimer
Copyright reserved by the author(s).
This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 48
The Effects of Long Term Sleep and Exercise Deprivation on Total Serum Testosterone in Male
Professional Drivers
Ghulam Nabi (Corresponding author)
Department Animal sciences, Laboratory, Reproductive neuro-endocrinology,
Quaid-i-Azam University, Pakistan
Tel: 92-345-811-2741 E-mail: [email protected]
Muhammad Amin
Department of Zoology, University of Karachi, Pakistan
Tel: 92-315-902-0833 E-mail: [email protected]
Amjad Khan
Department Animal sciences, Quaid-i-Azam University, Pakistan
Tel: 92-3329744605 E-mail:[email protected]
Rahim Ullah
Department Animal sciences, Quaid-i-Azam University, Pakistan
Tel: 92-301-835-054 E-mail: [email protected]
Muhammad Shuaib
Department Animal sciences, Quaid-i-Azam University, Pakistan
Tel: 92-343-977-2779 E-mail: [email protected]
Ayaz Ali khan
Department of Biotechnology, University of Malakand, Chakdara
Dir Lower, Khyber pakhtunkhwa, Pakistan

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 49
Tel: 92-345-5886-0677 E-mail: [email protected]
Received: February 7, 2014 Accepted: February 24, 2014
doi:10.5296/jbls.v5i2.5073 URL: http://dx.doi.org/10.5296/jbls.v5i2.5073
Abstract
The aim of the present study was to find out the effects of long term sleep deprivation and lack of exercise on total serum testosterone level and their consequences in male professional drivers.
This study was conducted at Mingora and Timergara in District Swat and District Dir respectively, Khyber Pakhtunkhwa, Pakistan. Blood samples were taken from 50 sleep deprived sluggish drivers and from 50 normal people of the same locality. A standard format questionnaire was designed regarding their age, marital status, number of children, mating frequency, duration and timing of sleep and habit of exercise. Bio-check (USA) kit was used according to the manufacturer protocol.
The mean and standard deviation of total serum testosterone in control was 650 ± 110 ng/dl (P < 0.0001), and in drivers was 410 ± 33 ng/dl (P < 0.0001). Total serum testosterone level was significantly lower in sleep deprived and sluggish drivers as compared to control group. The ill effects like reduce sexual desire, erectile dysfunction, Absent morning erection, scare pubic hairs, decrease frequency for shaving, muscle weakness, fatigue, depression and poor memory were also reported from the drivers during blood sampling, but no such problems were found in the control group.
Sleep deprivation especially REM and lack of exercise has a negative effect on total serum testosterone level. This low testosterone level in turn produces ill effects both on sexual and general body health.
Keywords: Fatigue, Erection, Pubic.
1. Introduction
In industrialized countries the duration of sleep has been considerably decreased during last decades. Sleep is very crucial for skeletal muscles and other physiologies. Chronic sleep deprivation has serious consequences such as producing muscle atrophy by increasing glucocorticoids and decreasing testosterone, growth hormone and insulin-like growth factor-I. It is also a risk factor for the chronic diseases such as cardiovascular diseases, metabolic syndromes, cancer and hormonal imbalance. However by performing resistance exercises the level of testosterone, growth hormone and insulin-like growth factor-I can be slightly raised (Monico et al., 2013). It has been found that sleep deprived people are in stress condition and this causes hormonal imbalance by secreting more cortisol, thyroid and less Testosterone, dehydroepiandrosterone- sulphate and insulin like growth factor-1 levels. Sleep has a direct

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 50
modulatory influence on the activity of Hypothalmo Pituitary Adrenal (HPA) axis and other hormonal axes. Sleep deprivation is an example of stressor and has serious consequences depending upon its duration (Maggio et al., 2013). Experiment resulted that sleep deprivation affects the pituitary–gonadal and pituitary- thyroidal axis by lowering the level of testosterone, prolactin and raising the level of Thyroid Stimulating Hormone (TSH), Free Tri-iodothyronine (FT3), and Free Thyroxin (FT4) as compared to control group (Jauch et al., 2013). Those fathers who sleep with their children at night and provide care to their children have low morning testosterone level as compared to those fathers who do not sleep at night with their children and do not provide care. This difference in morning testosterone occurs because the co-sleeper fathers have a disturbed sleep because of their children (Gettler, 2012). The drivers were also exposed to marijuana and cigarette smoke. Both of these inhibit secretion of Gonadotropin releasing hormone, thus significantly reducing secretion of testosterone (Murphy et al., 1998; Briggs et al., 1973; Shaarawy et al., 1982). Different types of exercise of moderate-intensity may reduce age-related physiological dysfunction associated with inflammation and oxidative stress by increasing expression and activity of the Nuclear factor erythroid 2-Related Factor (NRF2), a transcriptional regulator of the cellular anti-oxidant system which ultimately leads to high testosterone level in serum (Zhao et al., 2013). It has been concluded that strength training even in aging men slightly increases testosterone resulting in increased muscle function (Kvorning et al., 2013). Even in trained eu-menorrheic women short-term elevations in testosterone can be induced by prolonged aerobic exercise. This increase may result from either increased androgen production and/or decreased degradation rates of the hormone, and are not solely the result of plasma fluid shifts from the exercise (O'Leary et al., 2013). Low testosterone level in the body for long time due to either reason can causes cardiovascular problems, insulin resistance, obesity, osteoporosis, erectile dysfunction, hypoactive sexual desire, reduced nocturnal and morning erection, fatigue and depression (Buvat et al., 2013)
In the present study sleep deprived sluggish drivers and healthy peoples were selected. In such drivers there is lack of qualitative and quantitative sleep either due to poverty or illiteracy. In their short sleep the drivers wake up several times due to noise, brake etc. so in such drivers all the sleep stages are not achieved especially the REM (Rapid Eye Movement) which is crucial for testosterone secretion. Morning blood samples were taken from the sleep deprived and exercise deficient professional drivers and from people who take 8 hours sleep and regular exercise. The objective of the study was to find the effects of sleep deprivation (REM) and lack of exercise on morning serum total testosterone level and theirs resulting ill effects.
2. Materials and Methods
2.1 Questionnaire
For the present study a standard format questionnaire was designed for collecting particular information from the sleep deprived and exercise deficient drivers and from the control people regarding their age, marital status, number of children, mating frequency, duration and timing of sleep and habit of exercise (Table 1).
2.2 Data Collection

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 51
During sampling, 100 (50 sleep deprived and exercise deficient drivers and 50 controls) male Individuals were selected. Before taking interviews and blood sampling a written consent was signed from all the individuals. The morning blood samples from the drivers and control were collected in Mingora and Timergara, at district Swat and Dir (Lower), Khyber Pakhtunkhwa, Pakistan, from 14 August 2013 to 28 August 2013.
2.3 Blood Sampling from Sleep Deprived and Exercise Deficient Drivers
5 ml of the blood was taken from the brachial vein aseptically through a health technician and were immediately transferred into glass Vacutainer tubes (Becton Dickinson) without anticoagulant or preservative. These blood samples were taken from 9:00 AM to 10:00 AM after breakfast. Samples were transferred on dry ice to laboratory for immediate analysis. Serum was obtained after centrifugation at 3200 rpm at 4°C. Serums were stored at -20°C for later analysis. Total serum testosterone was measured using testosterone enzyme immunoassay test kit, Bio-check (USA) according to the manufacturer protocol.
2.4 Blood Sampling from Control
The people living around the bus stations in the same area were taken as control. A total of 50 people were randomly selected from the given areas. For the collection and analysis of blood samples same procedure was adopted as for drivers. The individuals selected were physically normal, have no history of chronic diseases, addiction and were not using any medications.
Statistical analysis
The results obtained were statistically analyzed by using Graphpad Prism, Demo version 05 (www.graphpad.com). The results were represented through mean and standard deviation and the comparison between two groups was performed by using student T-test. P value < 0.05 was considered to be statistically significant.
3. Results
3.1 History of Drivers and Control
Two groups were made, control and drivers. Each group was then divided into 3 age groups ranging from 25 to 55 years. Specific information from both control and drivers were collected via a standard questionnaire from various age groups and are summarized in Table 1.
Table 1. History of drivers and control
Age Group (Years)
Years after marriage
children number (average)
Mating frequency (average/week)
Sleep duration (hours)
Sleep timing
Exercise (average)
Health Problems
Drivers 25-34 (n= 15)
10-15
02
03
3-4
Variable
Absent
05= absent morning erection 10= reduce libido
35-44
10= absent erection, reduce

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 52
(n=20) 15-20
03 02 2-3 Variable Absent libido 10= muscle weakness, erection problem, fatigue
45-55 (n=15)
20-25
03
01
2-3
Variable
Absent
10= absent morning erection, reduce libido, memory loss, low pubic hairs, fatigue 05= depression, poor memory, low libido, reduce shaving frequency.
Control 25-34 (n=15)
10-15
04
05
7-8
Specific
02 hrs.
No specific
35-44 (n=20)
15-20
05
03
6-7
Specific
02 hrs.
02= hypertension
45-55 (n=15)
20-25
06
02
6
Specific
01 hrs.
03= hypertension 01= Diabetes 02= fatigue
SD*= Standard Deviation, C.I*= Confidence Interval
4. Discussion
In the present study we analyzed the ill effects of disturb short sleep and lack of exercise in drivers on testosterone level. Some long traveling buses have two drivers, one sleep while other drive. The sleeping drivers were expose to noise, brake and other irritating factors as a result they awake several times. They also failed to complete all stages of sleep especially REM stage which is either totally absent or very brief. REM sleep is very important for testosterone secretion. Testosterone maintains our general as well as sexual health. Sleep deprivation and sluggishness is a common phenomenon in both industrialized and developing countries and produces ill effects on general and sexual health. In the developing countries like Pakistan, particularly in Khyber Pakhtunkhwa the awareness about this problem is lacking.
Sleep is a very active, dynamic, complex behavior and occupies one-third of human life span. Sleep modulates cardiovascular, endocrine and metabolic system. Over the last several decades the average sleep duration has decreased. Long term sleep deprivation is a risk factor for several outcomes such as; disturb testosterone level, sexual dysfunction, Type-2 diabetes, insulin resistance and weight gain. In young men testosterone level rises during daytime sleep

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 53
just as in night time sleep, but the levels fall upon waking thus confirming that sleep, rather than a circadian rhythm is critical for testosterone regulation. The level of testosterone begins to rise with the onset of sleep and reaches to a peak at REM sleep onset approximately 90 min later. It has also been found that testosterone levels were highest during the first REM sleep episode and lowest when the subjects were awake. Sleep disturbance can cause both lower and higher circulating testosterone level. Men who sleep less than 4 hours and those who sleep 4-6 hours daily had significantly lower androgen concentrations than those who slept over 8 hours (Monica et al., 2011).
The fragmented sleep and short time spent in REM sleep causes a significant decline in serum total testosterone concentration. After awakening testosterone decline exponentially so old peoples who wake up many times have lower testosterone, which leads to erectile dysfunction, ejaculatory problems, diminished libido etc. (Monica et al., 2010; Penev., 2007). Rafael et al., 2001 found that sleep associated rise in testosterone is associated with the appearance of first REM sleep. Individuals with fragmented sleep and partial sleep deprivation having no REM sleep have a significantly low testosterone level. These findings are in agreements with our results because drivers are partially sleep deprived and have nearly no REM sleeps.
Figa et al, 1996 compared the control group with drivers and found that not only salivary testosterone level was reduced but sperm quality (i.e., sperm concentration, sperm morphology, and motility) was also compromised. They considered urban automobile driving a stressful work activity which negatively affects endocrine function and thus is a risk factor for male fertility. Rafael et al, 1999 compared testosterone level in waking, first NREM (Non Rapid Eye Movement) and REM sleep and found that testosterone level were significantly lower when the individuals were awake and were significantly higher during the first REM. Similarly serum testosterones were significantly higher during the first REM as compared to the pre- REM period. Raul et al, 1992 found that sleep related disordered, REM sleep and sleep efficiency has association with testosterone. Decreased sleep efficiency and number of REM episodes while increased in latency to the onset of REM are associated with low androgen level, and this association is independent of age.
Chronic sleep deprivation produces serious health effects by increasing the level of glucocorticoids, cortisol, TSH (Thyroid Stimulating Hormone), Free Tri-iodothyronine (FT3), Free Thyroxin (FT4) and decreasing the level of testosterone, dehydroepiandrosterone-sulphate, growth hormone and insulin like growth factor-1. Such study is an agreement with our findings (Monico et al., 2013; Maggio et al., 2013; Jauch et al., 2013; Gettler, 2012).
In a series of experiments in normal young adult men the interrelations of plasma LH (Luteinizing Hormone), FSH (Follicle Stimulating Hormone), prolactin (PRL), and testosterone during sleep were measured. It was found that during night baseline LH and FSH concentrations did not change, whereas PRL and testosterone over the course of night did show similar patterns of increasing concentration. In summer season due to shorter sleep duration, testosterone concentration was reported to be lowered. But in Ramadan sleep onset is later, sleep duration is reduced and it did not change the 24-h mean concentration of testosterone, but

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 54
did delay the onset of the increase. After 4 days of sleep deprivation in male rats, decreased testosterone levels and increased levels of progesterone, PRL, corticosterone, and catecholamines were observed. During military operations there is prolong psychological and physical stress, sleep deprivation lasts for several days and androgen levels fall by 70-90% (Monica et al., 2008). Long term low testosterone has serious consequences and can causes decreased libido, osteoporosis, depression, obesity, muscle weakness, fatigue, errection problems and disturb secondary sex characters (Ranjan et al., 2010). Same findings were reported during our study.
Children with better sleep quality were associated with better control over aggressive behaviors, while sleep deprived display aggression. Researchers had found that REM deprived rats engaged in more aggressive behavior (Kimberly et al., 2013).
In addition to sleep the drivers were also exposed to cigarette and marijuana smoke. It is well reported that nicotinic cigarette smoke and cocaine both stimulate the mesolimbic dopamine system that elevated the concentration of extracellular dopamine (Kuhar et al., 1991; Corrigall et al., 1992; Di Chiara , 2000; Watkins et al., 2000). This dopamine negatively regulates GnRH (Gonadotropin Releasing Hormone) secretion which causes significant reduction in testosterone level, affecting sexual health (Murphy et al., 1998; Briggs et al., 1973; Shaarawy et al., 1982). Marijuana activates dopamine and opioid which inhibit GnRH secretion, thus negatively affecting our sexual health (Murphy et al., 1998).
Exercise, competitions and resistance training transiently increase testosterone concentration in men and women. Testosterone levels vary before and after competition. Moderate intensity exercise raises the level of serum testosterone even in aging men and eu-menorrheic women (Zhao et al., 2013; O'Leary et al., 2013; Ruth et al., 2012). Testosterone concentration increases significantly after exercise. Therefore coaches may consider low-intensity resistance exercise as a potential training method for athletes who need to maintain muscle mass and strength during the long in-season (Con et al., 2012).
5. Conclusion
In conclusion it was observed that in professional drivers due to increased work load (because of poverty or illiteracy) there is lack of sleep and proper exercise which produce a stress condition in their bodies. This stress may be then responsible for low serum total testosterone level. Low testosterone has ill effects on general and sexual health. Therefore these peoples should take enough qualitative and quantitative sleep and exercise to maintain normal serum testosterone concentration and thus general and sexual health. They are also advised to take a resistance exercise that raises testosterone level.
Limitations
Due to limited facilities the correlation with other hormones was not studied.
References
Briggs, MH. (1973). Cigarette smoking and infertility in men. Medical Journal of Australia. 1, 616-617.

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 55
Buvat, J. Maggi, M. Guay, A., & Torres, L. (2013). Testosterone deficiency in men: Systematic review and standard operating procedures for diagnosis and treatment. Journal of sexual medicine Sex Med. 10, 245-284. http://dx.doi.org/10.1111/j.1743-6109.2012.02783.x
Con, M. Ikeda, T. Homma, T., & Suzuki, Y. (2012). Effects of Low-Intensity Resistance Exercise under Acute Systemic Hypoxia on Hormonal Responses. Journal of Strength and Conditioning Research. 26, 611-617.
Corrigall, WA. Franklin, KB. Coen, KM., & Clarke, PB. (1992). the mesolimbic dopamine system is implicated in the reinforcing effects of nicotine. Psychopharmacology.107, 285–289. http://dx.doi.org/10.1007/BF02245149
Di, CG. (2000) Role of dopamine in the behavioral actions of nicotine related to addiction. European Journal of Pharmacology. 393, 295-314. http://dx.doi.org/10.1016/S0014-2999(00)00122-9
Figa, T. Cini, C. Varricchio, D., & Lenzi, A. (1996). Effects of Prolonged Auto vehicle driving on male reproductive Function: A Study among taxi drivers. American Journal of Industrial Medicine. 30, 750-758. http://dx.doi.org/10.1002/(SICI)1097-0274(199612)30:6<750::AID-AJIM12>3.0.CO;2-1
Gettler, LT. (2012). Does Cosleeping Contribute to Lower Testosterone Levels in Fathers? Evidence from the Philippines. Public library of science. 7, 41559.
Jauch, K. Schmid, SM. Hallschmid, M. Oltmanns, KM., & Schultes, B. (2013). pituitary- gonadal and pituitary- thyroidal axis hormone concentration before and during a hypoglycemic clamp after sleep deprivation in healthy men. Public library of science 8, 54209.
Kimberly, A. Cheryl, M. Shawn, NG. Ryan, PR., & Stacey, DM. (2013). Sleep Deprivation lowers reactive aggression and testosterone in men. Biopsychology. 92, 249-256.
Kuhar, MJ. Ritz, MC., & Boja, JW. (1991). the dopamine hypothesis of the reinforcing properties of cocaine. Trends in Neurosciences. 14, 299-302. http://dx.doi.org/10.1016/0166-2236(91)90141-G
Kvorning, T. Christensen, LL. Madsen, K. Nielsen, JL. Gejl, KD. Brixen, K. (2013). Mechanical muscle function and lean body mass during supervised strength training and testosterone therapy in aging menwith Low-normal testosterone levels. Journal of the American Geriatric Society. 61, 957-62. http://dx.doi.org/10.1111/jgs.12279
Maggio, M. Colizzi, E. Fisichella, A. Valenti, G. Ceresini, G. Dall'Aglio, E. (2013). Stress hormone, Sleep deprivation and cognition in older adults. Maturitas. 76, 22-44. http://dx.doi.org/10.1016/j.maturitas.2013.06.006
Monica, A. Tathiana, A. Renata, M. Helena, H., & Sergio, T. (2011). The association of Testosterone, sleep, and sexual function in men and women. Brain Research. 1416, 80-104. http://dx.doi.org/10.1016/j.brainres.2011.07.060
Monica, L. Sergio, T. (2008). The effects of testosterone on sleep and sleep- disordered

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 56
Breathing in men: Its bidirectional interaction with erectile function. Sleep medicine review. 12, 365-379. http://dx.doi.org/10.1016/j.smrv.2007.12.003
Monica, LA. Rogerio, SS. Lia, RA. Bittencourt, ST. (2010). Prevalence of erectile dysfunction complaints associated with sleep disturbances in Sao Paulo, Brazil: A population-based survey. Sleep Medicine.11, 1019-1024. http://dx.doi.org/10.1016/j.sleep.2009.08.016
Mônico, MN. Antunes, HK. Dattilo, M. Medeiros, A. Souza, HS., & Lee, KS. (2013). Resistance exercise: A non-pharmacological strategy to minimize or reverse sleep deprivation-induced muscle atrophy. Medical Hypotheses. 80, 701-705. http://dx.doi.org/10.1016/j.mehy.2013.02.013
Murphy, LL. Munoz, RM. Adrian, BA. Villanu, MA. (1998). Function of cannabinoid receptors in the neuroendocrine regulation of hormone secretion. Neurobiology. 5, 432-446.
O'Leary, C. B., Lehman, C., Koltun, K., Smith, Ryan. A., Hackney, A. C. (2013). Response of testosterone to prolonged aerobic exercise during different phases of the menstrual cycle. Eureopean Journal of Applied Physiology. 9, 2419-24. http://dx.doi.org/10.1007/s00421-013-2680-1
Penev, P. (2007). Association between sleep and morning testosterone levels in older men. Sleep.30, 427-432.
Rafael, L. Paula, H. Michal, L. Zila, S., & Peretz, L. (1999). Relationship between rapid eye movement sleep and testosterone secretion in normal men. Journal of Andrology. 20, 731-737
Rafael, L. Ziva, Z. Zill, O. Paula, H., & Peretz, L. (2001). Disruption of the Nocturnal Testosterone Rhythm by Sleep Fragmentation in Normal Men. The journal of clinical endocrinology and metabolism.86, 3.
Ranjan, A. Mohan, A., & Shaun, R. (2010). Androgen deficiency in the aging man. Australian Family Physician. 39, 10.
Raul, CS. Daniel, W., & John, M. (1992). Pituitary- Gonadal function during sleep in healthy aging men. Psych neuroendocrinology.17, 6, 599-609.
Ruth, IW. Steven, JS. (2012). Testosterone and sport: Current perspectives. Hormone and behaviour. 61, 147-155. http://dx.doi.org/10.1016/j.yhbeh.2011.09.010
Shaarawy, M. Mahmoud, KZ. (1982). Endocrine profile and semen characteristics in male smokers. Fertility and Sterility. 38, 255-257.
Watkins, SS. Koob, GF., & Markou, A. (2000). neural mechanisms underlying nicotine addiction: acute positive reinforcement and withdrawal. Nicotine and Tobacco Research.. 2, 19-37. http://dx.doi.org/10.1080/14622200050011277
Zhao, X. Bian, Y. Sun, Y. Li, L. Wang, L. Zhao, C. (2013). Effects of moderate exercise over different phases on age- related physiological dysfunctions in testes of SAMP8 mice. Experimental gerontology. 48, 869-80. http://dx.doi.org/10.1016/j.exger.2013.05.063

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 57
Copyright Disclaimer
Copyright reserved by the author(s).
This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 58
Effect of Fetal Sex on Total Levels of Maternal Serum Testosterone
Ghulam Nabi (Corresponding author)
M. Phil Scholar. Department: Animal sciences, Labortary: Reproductive neuro-endocrinology
Quaid-i-Azam University, (Islamabad) Pakistan.
Tel: 92-345-811-2741 E-mail: [email protected]
Tariq Aziz
M. Phil Scholar, Department: Biotechnology
University of Malakand, Chakdara, Dir Lower, Khyber pakhtunkhwa, Pakistan
Tel: 92-345-945-2645 E-mail: [email protected]
Muhammad Amin
M. Phil Scholar, Department of Zoology, University of Karachi, Pakistan
Tel: 92-315-902-0833 E-mail: [email protected]
Ayaz Ali khan
Assistant Professor, Department of Biotechnology
University of Malakand, Chakdara, Dir Lower,Khyber pakhtunkhwa, Pakistan
Tel: 92-345-5886-0677 E-mail: [email protected]
Received: March 4, 2014 Accepted: March 19, 2014
doi:10.5296/jbls.v5i2.5228 URL: http://dx.doi.org/10.5296/jbls.v5i2.5228
Abstract
The aim of this study was to find out the effect of fetal sex on maternal serum total testosterone level and its application for fetal sex determination.

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 59
Forty healthy pregnant (second trimester) females were recruited in the study from rural areas of district Dir lower, Khyber Pakhtunkhwa, Pakistan, having complete antenatal record. Twenty of them were carrying single male fetuses and twenty female fetuses. The inclusions criteria were age (25 to 30), second trimester, absence of serious diseases, availability of antenatal record, no drug addiction and no exposure to pesticides. Blood samples at 5 ml size were collected from each woman, serum was obtained and was assayed by Bio-check (USA) kit according to the manufacturer protocol.
In male fetus group the mean and SD was 169±27.18 ng/dl and in female fetus group the Mean± SD was 166.6±30.47 ng/dl. There was nosignificant difference (P 0.1062) between the two groups at 95% confidence level.
The results suggest that sex of the fetus has no association with maternal serum total testosterone among the study population-and should not be analyzed for sex determination. Further study with bigger sample size of different population groups in different gestational stages is needed to find the fetal effect on maternal serum testosterone because, increased level of testosterone in females can cause aggression, other behavioral changes, acnes and abrupt growth of pubic and axillary hairs.
Keywords: Fetus, Sex, Serum, Testosterone
1. Introduction
In men testosterone is the most important androgen, 95 % of which is synthesized by the testes and the rest by the adrenal cortex. It is very crucial for the spermatogenesis, fetal male sexual differentiation, pubertal development, maintenance of adult secondary sex characters, immunity, bones and muscles etc. (Stephen, 2003).Fetal testes produce testosterone which is important for male sexual differentiation. As soon as the testes differentiate, testosterone production started (8 week of gestation) and reaches to its peak at 10 -15 week of gestation. A decline occurs at the start of third trimester until after birth (Ellinwood et al., 1980).In Rhesus monkey and also in human, fetal testes are active steroidogenically. In human fetusthe formation of testosterone by testes appeared to be independent of gonadotropin control. (Word et al., 1989). The amniotic fluids of both male and female fetuses were analyze for hormone analysisand was found that male fetuses had significantly higher level of Testosterone, while females had significantly higher level of Estrogen but non-significant correlation were found for the different androgenic hormone between levels assessed in amniotic fluid, maternal plasma, and umbilical cord serum.(Rivarolaet al., 1968; Forest et al., 1971; Meulenberg and Hofman, 1991;Corneliekeet al., 2004). But in case of Asian elephant (Elephasmaximus) carrying male calves, mean progesterone concentration in maternal serum is significantly greater than in those carrying female calves and are thus used for gender determination (Connie et al., 2007). Throughout the pregnancy maternal serum total testosterone concentration increases and the sources for this increase are unknownbut could be the maternal cortex and ovarian theca-interstitial cells (Nahid and Sirous, 2013).
In thisstudy we selected forty pregnant women in their second trimester. Twenty were carryingmale fetuses and twentywere carrying female fetuses. The hypothesiswas that fetal sex

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 60
has effect on the total testosterone level in maternal serum, i.e., testosterone is synthesized only in male fetuses.
2. Materials and Methods
2.1 Participants
Healthy pregnant females with complete antenatal records were randomly selected from the rural areas of district Dir (Khyber pakhtunkhwa, Pakistan). Two groups were made. One group containing twenty pregnant female having single male fetuses. Other group also had twenty pregnant female but carryingsingle female fetuses.A written informed consent was signed from all the participants. This study was allowed by local community leader as well as by a registered doctor.The inclusion criteria were age ranges from 25 to 30, second trimester, absence of serious disease, availability of antenatal record, no drug addiction and no direct exposure to pesticides (excluding women working in fields).
2.2 Blood Sampling
A health technician collected 5 ml of the blood from the antecubital vein of the participants aseptically. Blood was collected in Vacutainer tubes containing no additives. Blood were sampled in their home between 9:00AM to 10:00AM after breakfast not before. At room temperature blood was allowed to clot and then centrifuged. The resulting serum samples were then stored at -20°C for later analysis.
2.3 Assay
For the measurement of serum total testosterone,testosterone enzyme immunoassay test kit, Bio-check (supplier, city, USA) was used according to the manufacturer’s instructions.
2.4 Statistics
The software Graph pad Prism, Demo version 6.03 (www.graphpad.com) was used for statistical analysis. Student paired t test was used for the comparison of serum total testosterone in both groups. Differences between paired values were consistent. The results were represented through Mean±SD. A value of p<0.05 was considered statistically significant.
3. Results
In male fetus group the Mean± SD of maternal serum total testosterone was 169±27.18ng/dl and in female fetuses group the Mean±SD was 166.6±30.47ng/dl (Table 1). The non- significant difference (P 0.1062) indicates that maternal serum total testosterone level is independent of fetal sex.
Table1. Total serum Testosterone in Male fetuses group and Female fetuses group
Parameter Male
Fetuses group Mean±SD*
Female Fetuses group
Mean ±SD
95% CI* of the mean difference
P value
Total Serum Testosterone
Level
169±27.18 (ng/dl) 166.6±30.47 (ng/dl) -5.362 to
0.5615 0.1062

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 61
SD*= Standard Deviation, C.I*= Confidence Interval
4. Discussion
In the present study it was found that maternal serum total testosterone level is not significantly (P 0.1062 at 95% CI) affected by the fetal sex. This study is in agreement with the findings of Barboraet al. (2010) ; Steier et al. (2002) ;Salamalekis et al.(2006); Zondek et al.(1977) and Robertson et al., (1980),who compared the serum total testosterone and estradiol level of pregnant women’s carrying male and female fetuses and found non–significant differences, thus excluding their application for fetal sex determination. They found that plasma testosterone increases in women’s throughout pregnancy and this increase is even more significant in preeclamptic pregnancies. A fetal gender difference was not found in the maternal plasma but only in the amniotic fluids.Rodec.(1985) ;Dawood and sexena.(1977) ; Fineganet al.(1989) ; Judd et al.(1976); Nagamaniet al.(1979) and Robinson et al.(1977), analyzed maternal plasma and amniotic fluids for total testosterone. They found that in male fetal amniotic fluids testosterone concentration were higher than female fetuses but this does not predict the fetal sex reliably until pure fetal plasma is used. But still this is not the method of choice for routine fetal sexing.Jefery.(2012), analyzed umbilical blood of male and female neonates and found that total and free testosterone levels were higher in males(P<0.0001) while dehydroepiandrosterone levels were higher in females (P<0.0001).Castracane and Dela.(1990),performed experiment on rats to find the effects of number of male fetuses on maternal serum total testosterone and observed that there was no relationship between maternal serum total testosterone level and the number of male fetuses. This showing that fetal testis does not affect the maternal serum total testosterone level.According toGlass and Klein.(1981)during pregnancy the fetal testes produce testosterone. To find out whether this fetal synthesized testosterone can be used for gender determination by measuring the level of maternal testosterone. They analyzed serum testosterone (total and free) anddihydrotestosterone in pregnant women’s. The age of fetuses ranged from 4 to 20 weeks and found no any significant difference in maternal serum androgen level in women’s having male fetuses and those having female fetuses.However Steieret al.(2002) and Nahid and Sirous.(2013),found that maternal serum testosterone and HCG(Human Chorionic Gonadotropin) were significantly elevated in preeclamptic than normotensive mothers having male fetuses. As compared to normotensive mothers, in preeclamptic mothers bearing female pregnancies maternal serum testosterone were significantly higher. But there was no significant difference in HCG concentration. Again in preeclamptic women’s bearing male pregnancies had higher serum testosterone as compared to preeclamptic women’s carrying female pregnancies but HCG were not significantly different. In uncomplicated pregnancies testosterone level did not differ significantly, however HCG level were higher in mothers carrying female pregnancies than male pregnancies.
Some studies have found that women’s carrying single male fetuses had higher testosterone level as compared to those women’s bearing single female fetuses during gestation. But most of the studies found no any direct relation between maternal serum testosterone and fetal gender. They even analyzed serum testosterone in women’s carrying two fetuses (female-female. Male-male and opposite sex) and observed no any significant difference in the maternal serum

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 62
testosterone of women’scarrying single sex or mixed sex fetuses (Cohen et al., 2005).
5. Conclusion
It is suggested that sex of the fetus has no association with maternal serum total testosterone among the study population-and should not be analyzed for sex determination
Limitations
Due to limited facilities other hormones were not analyzed. The participants were also not ready to givespecific information’s regarding their health.
References
Castracane, VD., & Dela, JL. (1990). The relationship of conceptus number and of fetal sex to maternal serum testosterone concentration in the rat. Proceedings of the society for experimental biology and medicines. 195(1), 109-13.
Cornelieke, VB. Jos, HH. Thijssen, PT. Cohen, K. Stephanie, HM. Van, G., & Jan, K. (2004). Relationships between sex hormones assessed in amniotic fluid, and maternal and umbilical cord serum: What is the best source of information to investigate the effects of fetal hormonal exposure? Hormone and behavior. 46, 663-669. http://dx.doi.org/10.1016/j.yhbeh.2004.06.010
Cohen, BCC. Van, GSH.Buitilar, JK., & Cohen, KPT. (2005). Twin Research and Human Genetics. 8(2), 173-7. http://dx.doi.org/10.1375/twin.8.2.173
Connie, D. Melissa, C., & Thomas, T. (2007). Detection of fetal gender differences in maternal serum progesterone concentrations of Asian elephants (Elephasmaximus).Animal Reproduction Science. 97, 278-283. http://dx.doi.org/10.1016/j.anireprosci.2006.02.008
Dawood, MY. Saxena, BB. (1977). Testosterone and dihydrotestosterone in maternal and cord blood and in amniotic fluid.American Journal of Obstetrics and Gynecology. 129, 37–42.
Ellinwood, RM. Brenner, DL., & Hess, JA. (1980). Testosterone Synthesis in Rhesus Fetal Testes: Comparison between Middle and Late Gestation. Biology of reproduction, 22(4), 955-963. http://dx.doi.org/10.1095/biolreprod22.4.955
Forest, MG. Ances, IG. Tapper, AJ., & Migeon, CJ. (1971). Percentage binding of testosterone, androstenedione and dehydroisoandrosterone in plasma at the time of delivery.Journal of Clinical Endocrinology. 32, 417-425. http://dx.doi.org/10.1210/jcem-32-3-417
Finegan, JA. Bartleman, B., & Wong, PY. (1989). A window for the study of prenatal sex hormone influences on postnatal development.Journal of Genetic Psychology. 150(1), 101-112. http://dx.doi.org/10.1080/00221325.1989.9914580
Glass, AR. Klien, T. (1981).Changes in maternal serum total and free androgen levels in early pregnancy: lack of correlation with fetal sex. American Journal of Obstetrics and Gynecology. 140(6), 656-60.
Judd, HL. Robinson, JD. Young, PE., & Jones, OW. (1976). Amniotic fluid testosterone levels

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 63
in mid pregnancy.Obstetrics and Gynecology. 48(6), 690-692
Jeffrey, A. Keelan, EM., & Martha, H. (2012). Androgen Concentrations in Umbilical Cord Blood and Their Association with Maternal, Fetal and Obstetric Factors.Public library of science. 7(8), 42827.
Meulenberg, PMM. Hofman, JA. (1991). Maternal testosterone and fetal sex.Journal of Steroid Biochemistry and Molecular Biology. 39(1) 51-54. http://dx.doi.org/10.1016/0960-0760(91)90012-T
Nagamani, M. McDonough, PG. Ellegood, JO., & Mahesh, VB. (1979). Maternal and amniotic fluid steroids throughout human pregnancy.American Journal of Obstetrics and Gynecology. 134(6), 674-680.
Nahid, L. Sirous, K. (2103). The Effects of Fetal Gender on Serum Human Chorionic Gonadotropin and Testosterone in Normotensive and Preeclamptic Pregnancies. Journal of Pregnancy. 518591.
Rivarola, MA. Forest, MG., & Migeon, CJ. (1968). Testosterone, androstenedione and dihydroepiandrosterone in plasma during pregnancy and at delivery: concentration and protein binding. Journal of Clinical Endocrinology. 28, 34-40. http://dx.doi.org/10.1210/jcem-28-1-34
Robinson, JD. Judd, HL. Young, PE. Jones, OW., & Yen, SSC. (1977). Amniotic fluid androgens and estrogens in midgestation.Journal of Clinical Endocrinology and Metabolism. 45(4), 755-761. http://dx.doi.org/10.1210/jcem-45-4-755
Robertson, RD. Henniker, AJ. Luttrell, BM., & Saunders, DM. (1980). The prenatal determination of fetal sex: amniotic fluid testosterone as a preliminary screening test. European Journal of Obstetrics and Gynecology and Reproductive Biology. 10, 77-81. http://dx.doi.org/10.1016/0028-2243(80)90085-4
Rode, GD. Rosenberg, DA., & Collins, WB. (1985). Testosterone levels in midtrimester maternal and fetal plasma and amniotic fluid. Prenatal Diagnosis. 5(3), 175-81. http://dx.doi.org/10.1002/pd.1970050303
Salamalekis, E. Bakas, P. Vitoratos, N. Eleptheriadis, M., & Creatsas, G. (2006). Androgen levels in the third trimester of pregnancy in patients with preeclampsia. European Journal of Obstetrics and Gynecology and Reproductive Biology. 126, 16-19. http://dx.doi.org/10.1016/j.ejogrb.2005.07.007
Steier, JA. Ulstein, M., & Myking, OL. (2002). Human chorionic gonadotropin and testosterone in normal and preeclamptic pregnancies in relation to fetal sex.Obstetrics and Gynecology. 100(3), 552-6.
Stephen, J. Winters, B., & Clark, J. (2003). Testosterone Synthesis, Transport, and Metabolism. Contemporary endocrinology. 3-22. http://dx.doi.org/10.1016/S0029-7844(02)02088-4

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 64
Vlková, B. Vávrová, S. Szemes, T. Minárik, G. Turňa, J., & Celec, P. (2010), Testosterone and estradiol in maternal plasma and their relation to fetal sex. Prenatal Diagnosis. 30, 806-807. http://dx.doi.org/10.1002/pd.2535
Word, AN. Friedriek, W.Georoge, JW., & Bruce, RC. (1989). Testosterone Synthesis and AdenylateCyclase Activity in the Early Human Fetal Testis Appear to Be Independent of Human Chorionic Gonadotropin Control. The journal of clinical endocrinology and metabolism .69, 1. http://dx.doi.org/10.1210/jcem-69-1-204
Zondek, T. Mansfield, MD., & Zondek, LH. (1977). Amniotic fluid testosterone and fetal sex determination in the first half of pregnancy. Br J Obstetrics andGynecology. 84, 714-716.
Copyright Disclaimer
Copyright reserved by the author(s).
This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 65
Link between Chronic Pesticides Exposure and Reproductive Problems in Male Farmers
Ghulam Nabi (Corresponding author)
M. Phil Scholar. Animal sciences Department,: Reproductive neuro-endocrinology Lab Quaid-i-Azam University, (Islamabad) Pakistan
Tel: 92-345-811-2741 E-mail: [email protected]
Muhammad Amin
M. Phil Scholar, Department of Zoology, University of Karachi, Pakistan
Tel: 92-315-902-0833 E-mail: [email protected]
Tabinda Rauf
M.Phil scholar, Department of Zoology, University of SBBU
Sheringal, Dir, Khyber Pakhtunkhwa, Pakistan
E-mail: [email protected]
Khwaja Muhammad Khan
B.sc (hon), Department of agriculture, Hazara University Haripur, Pakistan
Tel: 92-346-344-0311 E-mail: [email protected]
Ayaz Ali khan
Assistant Professor, Department of Biotechnology, University of Malakand
Chakdara, Dir Lower, Khyber pakhtunkhwa, Pakistan
Tel: 92-345-5886-0677 E-mail: [email protected]
Received: April 24, 2014 Accepted: May 4, 2014
doi:10.5296/jbls.v5i2.5501 URL: http://dx.doi.org/10.5296/jbls.v5i2.5501

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 66
Abstract
The demand for potent and new pesticides increases day by day because of pest resistant and more food production. The objective of this study was to find out the effects of chronic pesticides exposure on the reproductive health of poor and reckless farmers.A 5 mL blood was drawn from 80 men (40 farmers, 40 controls) and was analyze for serum total testosterone using Biocheck (USA) kit. A detail questionnaire was designed about socio-demographic, occupational activities and clinical characteristics.In farmer group the Mean±SEM of total serum testosterone was 546±10 ng/dl and in control was 664±17 ng/dl. Total serum testosterone was significantly lowered (P< 0.0001) in farmers at 95% confidence interval as compared to control group. Reproductive health problems like infertility, erectile dysfunction, ejaculatory problems, decreased libido, absent morning and nocturnal erection and general health problems like asthma, rheumatoid arthritis, hypertension, Parkinson and diabetes mellitus were more common in farmers.
Pesticides negatively affect the physiology of endocrine system. It reduces the serum total testosterone concentration in farmers significantly.This low testosterone produces negative effects on the reproductive as well as general health.
Keywords: Libido, Infertility, Nocturnal.
1. Introduction
In today’s world pesticides are one of the most important contaminant actively involved in environmental contamination. These chemicals are toxic to vectors of diseases and pest. Pesticides are marketed as herbicides, insecticides and fungicides. The demands for new and potent pesticides are increasing day by day because of pest resistance and more food production to fulfill the need of world growing populations. Only in European Union 140,000 tons of pesticides are used annually for agriculture purpose, such sort of extensive application raised the questions about hazards of pesticides to human health, including reproductive. On one side pesticides helped the human by controlling infectious diseases and increasing the yield of agriculture products. But on the other hand their extensive long term use can harm the health. Pesticides can disturb the physiology of reproductive, endocrine, renal, cardiovascular, respiratory, nervous and immune system causing Alzheimer, multiple sclerosis, diabetes, Parkinson, renal and cardiovascular problems (Abdollahiet al., 2004;Ramazzini, 2009; De Souza et al., 2011;Mostafalou and Abdollahi, 2012).
Most of the pesticides cause reproductive toxicities by interfering with the physiology of endocrine system (Figa-Talamanca et al., 2001; Cocco, 2002; Tiemann, 2008). Pesticides like,endosulfans, dieldrin, DDT and aldrin, the herbicides atrazine and the fungicide vinclozoline are considered as endocrine disruptor chemicals (PAN, 2009).Both male and female reproductive systems are negatively affected by pesticides exposure (Kumar, 2004; Shojaei and Abdollahi, 2012). Chronic pesticides exposure can causes decreased fertility in both sexes, high rate of miscarriage, altered sex ratio and has also antiandrogenic effects (Frazier, 2007).Pesticides exposure causes hormonal imbalance that eventually leads to infertility and sterility. Pesticides like, carbamates, pyrethroids, organophosphatases, Thio-and

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 67
dithiocarbamates, chlorphenoxy acids and chlormethylphosporic acids reduces testosterone concentrations in male after acute exposure during exposure season (Evamarieet al., 1999).A pesticide that changes the physiology of endocrine system by acting as hormone agonists/antagonists or changing endogenous steroidal hormonal level may negatively affect the developmental, biochemical and behavioral functions that are very important for reproductive success (Gerald et al., 1997). Pesticides are endocrine disrupting chemicals (EDCs) that interfere with the synthesis, transport, metabolism and elimination of hormone there by, changing the concentration of natural hormone (Gore, 2010).Up to now a totals of 101 pesticides have listed to be endocrine disruptors (EDs) by the pesticides action network UK (PAN, 2009).
In Pakistan especially in district Dir (lower) the awareness about pesticides hazardous effects is lacking in farmers either due to illiteracy or poverty. These farmers never use masks, gloves, eye glasses and other protective coverings during fumigations. They even never wash their hands after fumigationand take their breakfast or meal in their fields. During fumigation, all the clothes and body of farmers are washed by pesticides. They are also addicted to snuff which during fumigation are contaminated by pesticides. For the sake of high yields with low efforts they uses excessive and multiple pesticides. These farmers are exposed to pesticides from generation to generation. The present study is design to analyze the ill effects of pesticides on the male reproductive health of farmers.
2. Material and methods
2.1 Pesticides Used in Study Population
Most commonly used pesticides are Mancozeb, cymoxanil, metalaxyl, cypermethrin, tebuconazole, nitonpiram, hexythiazox,chlorothalonil, metiram, pyraclostrobin, dimethomorph, levofenoron, pyrimethanil, imidacloprid, thiophanate methyl, bromoxynil, MCPA, pendimethaline, nicosulfuron, atrazine and endosulfon (now banned).
2.2 Inform Consent
Before starting the study a written informed consent was signed by each participant.
2.3 Study Population
We selected the agriculture community of village Tazagram, district Dir lower, Khyber pakhtunkhwa, Pakistan for study. This area was selected because it is surrounded by agriculture fields whose main products include vegetables (tomatoes, onions, spinach, cauliflowers, cabbage, garlic, brinjals, ladyfingers)crops (maize, rice, wheat, tobaccos) as well as fruits (oranges, grapes, apricots, pears). It is situated at 695 meters above sea level. The climate is temperate so, agricultural production is possible all year round.We selected 80 men randomly from the community. They were divided into two groups i.e. control and exposed. Each group contained 40 participants. The exposed group participants were farmers by profession while control group participants were engage in other professions. Their age ranges from 29 to 50 years and were divided into two age groups i.e. 29 -40 and 41-50 respectively. Each participant was interviewed directly regarding his socio-demographic characteristics, alcohol and smoking

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 68
habit, occupational activities and clinical characteristics.
2.4 Inclusion Criteria
Farmers directly exposed to pesticides for at least 20 years.
2.5 Exclusion Criteria
No history of chronic illness, chemotherapy and radiotherapy.
2.6 Blood Sampling
Samples were collected during the heavy spraying season (March to July).5 mL of the morning blood sample was collected from the antecubital vein of each participant by a phlebotomist aseptically. Immediately blood was transferred into Vacutainer tube containing no additives. At room temperature blood was allowed to clot and then centrifuged at 3200 rpm for 15 minutes at 4 C0. The resulting serum samples were then stored at -20°C for later analysis.
2.7 Assay
For the measurement of serum total testosterone, testosterone enzyme immunoassay test kit, Bio-check USA was used according to the manufacturer protocol and procedures.
2.8 Data Analysis
Analysis of data was done by using Graph Pad Prism (www.graphpad.com), version 6.03 (Graph Pad Software Inc., San Diego, CA, USA). Student un-paired t test was used for the comparison of serum total testosterone in both groups. The results were represented through Mean±SEM. A value of p<0.05 was considered statistically significant.
3. Results
3.1 Direct Fumigation Effects
The farmers did not use protective coverings like, masks, gloves, dresses etc. so are directly exposed to pesticides. The effects like, headache, nausea, dizziness, cough, blurring of vision, throat, skin, nose and eye irritation were common in farmers during and after fumigations.
3.2 Selected Characteristics of the Study Population
The characteristics of the study population are presented in Table 1
Table 1. Characteristics of the study population
Age Group (Years)
Exposure time to
pesticides (years)
Fumigation frequency (per week)
Protective clothing
Hand wash
Addiction (%)
General health problems
Reproductive health problems
Control 29-40 (n=20)
yes 05 snuff 6 cigarette
01 DM*
absent

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 69
41-50 (n=20)
yes 11 snuff 6 cigarette
02 HT* absent
Farmer 29-40 (n=20)
≥ 20 yrs. ≥ 7/week absent no 8 snuff 9 cigarette
04 HT
02 A*
01 RA*
02 infertility 04 DL* 03 EP* 04 AMN*
41-50 (n=20)
≥ 20 yrs. ≥ 7/week absent no 13 snuff 5 cigarette
06 HT 03 DM 02 P*
02 infertility 04 DL 05 EP
DL*= Decreased Libido, EP*= Erection Problems, AMN*= Absent Morning and Nocturnal erection.
DM*= Diabetes Mellitus, HT*= Hypertension, A*= Asthma, RA*= Rheumatoid Arthritis, P*= Parkinson,
3.3 Analysis of total serum testosterone concentration in study population
Total serum testosterone concentrations in both groups are summarized in Table 2. A significant reduction (P< 0.0001) in total serum testosterone concentration was found in farmers as compared to control group at 95% confidence interval.
Table 2. Total serum testosterone concentration in study population
Parameters
Control Mean ± SEM
Farmer Mean ± SEM
95% C.I* P value
Total serum testosterone concentration 664 ± 17 (ng/dl) 546 ± 10 (ng/dl) -158 to -78.61 < 0.0001
CI*= Confidence Interval
4. Discussion
Our main finding in this study were low serum total testosterone, reproductive and general health problems, that were more pronounce in farmers as compared to control group. In this community the most common symptoms related to pesticides reported were nausea, headache, dizziness, blurring of vision, throat, skin, nose and eye irritations that were in agreements with other reports link with direct exposure to Organophosphates (OP) and carbamates (CB) pesticides (Catanoet al., 2008).Total serum testosterone and estradiol level decreases in peoples exposed to OP (Padungtodet al., 1998; Straubeet al., 1999). Experimentally this is confirmed that, OP exposure alters the metabolism of testosterone and estradiol (Butler and Murray 1993; Murray and Butler 1995) thus supporting our results.OPaffects the reproductive functions primarily via decreasing brain acetylcholinesterase (AChE) activity and secondarily via gonads. This decrease in AChE leads to increase the concentration of acetylcholine, gamma-aminobutyric acid (GABA), epinephrine, norepinephrine, 5-hydoxytryptamine and dopamine concentration (Glissonet al., 1974; Gupta et al., 1984). Increased level of GABA then inhibit the release of gonadotropin releasing hormone (GnRH) in the median eminence, which is responsible for the release of luteinizing hormone (LH) and follicle stimulating

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 70
hormone (FSH) from the anterior pituitary. These LH and FSH are then involved in steriodogenesis and gametogenesis (Mitsushimaet al., 1994; Terasawa and Fernandez, 2001). Like nicotine and cocaine, pesticides increases dopamine concentration that negatively regulate GnRH secretion suppressing the reproductive functions( Kuharet al., 1991; Corrigallet al., 1992; Murphy et al., 1998; Di Chiara, 2000; Watkins et al., 2000). OP and carbamates alter pituitary adrenal axes (Clement 1985; Kokkaet al., 1987). Acephate andmetamidophos stimulate the secretion of hypothalamic corticotropin-releasing hormone,which in turn stimulate adrenocorticotropic hormone (ACTH) and so cortisol(Spassovaet al., 2000). Receptor of cortisol are located on GnRH neurons, RFRP-3(mammalian ortholog of GnIH) and on gonadotrops. High cortisol level inhibits the secretion of GnRH and so LH and testosterone while positively regulate RFRP-3 secretion, thus suppressing reproductive system (Kirby et al., 2009). OP and CB affect the pituitary-thyroidal axes (Clement, 1985)reducing the level of serum fT4 (free thyroxin) in farmers (Khan et al., 2013) by increasing T4 metabolism through induction of of hepatic UDP-glucuronosyltranferase enzymes (Hill et al., 1989)and to displacement of T4 from plasma binding-proteins, altering hepatic androgen metabolism (Van et al., 1991).The pesticides like imidacloprid (IMI) and acetamiprid (ACE) bind to insect nicotinic acetylcholine receptor in central nervous systems and kill them (Tomizawa and Casida, 2005). But in vivo studies have found that neonicotinoid pesticides including IMI and ACE affect the reproductive organs of mammals causing retardation of testicular development, damage to spermatogenesis, decrease in sperm quality and change of ovary morphology (Kapooret al., 2010; Kapooret al., 2011; Balet al., 2012a ; Balet al., 2012b). OP exposure can result paternal fertility problems. OPisgenotoxic to animal sperm because it is a very potent phosphorylating agent. It binds to DNA and protamines, altering the chromatin structure that results DNA to become more susceptible to induced denaturation in situ (Evensonet al., 2002; Sergerieet al., 2005;Pina-Guzman et al., 2005;Boe-Hansen et al., 2006;). In Chinese pesticides factory workers the effects of OP exposure on their reproductive system was done. It was found that OP exposure increases LH and FSH but decreases serum total testosterone (Padungtodet al.,1998).In adult men OP and CB metabolites reduces the serum level of LH and total testosterone (Meeker et al., 2006).In Djutitsa (West Cameroon) a significant reduction in serum total testosterone and androstenedionewere found in farmers exposed to agro pesticides (Manfoet al., 2010). A lot of wildlife species have been effected by endocrine disrupting chemicals(EDCs) showing alteration in penis/gonopodium development, decreased testosterone level, alters spermatogenesis, impaired reproduction, reduce hatchability and/ or viability of offspring, aberrant sexual behavior and affect sex determination, differentiation of the gonads and timing of sexual maturation(Oberdörster and Cheek, 2001; IPCS, 2002; Stoker et al., 2003;Joblinget al., 2006; Milneset al., 2006; Crain et al., 2007; Orlando and Guillette, 2007;Guillette and Edwards, 2008;Stoker et al.,2008). Exposure toendosulfan (END) and atrazine (ATZ) during the embryonic life of Caiman latirostisaltered histoarchitecture of the testis as well as the balance between proliferation and apoptosis of hatchlings’ testicular cells. In adult male exposed to END, a decreased testosterone level was observed (Florenciaet al., 2009). Impaired testicular functions were also suggested for factory workers exposed to chlordecone (Anderson et al., 1976). Delayed spermatogenesis, decreased sperm counts, altered sperm motility and a higher incidence of intersex were observed in fishes exposed to

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 71
END (Joblinget al., 2002; Aravindakshanet al., 2004; de Montgolfier et al., 2008). Some studies have found high testosterone level (Sarkaret al., 2000) or no significant change in the testosterone level (Rogelio et al., 2005) of model animals as well as human exposed to pesticides. This may be due to the agrochemicals use, severity and/or length of exposure, agricultural practices and protection equipment pattern of use (Rogelio et al., 2005).
5. Conclusion
In summary pesticides exposure negatively affects the endocrine system directly or indirectly, resulting into serious consequences. In district Dir there is a need to educate the farmers about the appropriate use of pesticides, implement a formal worker protection program for agricultural workers and preventive health monitoring for early detection of pesticide exposure.
References
Anderson, JJH. Cohn, WJ. Guzelian, P. Taylor, JR. Griffith, FD. Blanke, FD. Dos Santos, & JG. Blackard G. (1976). Effects of kepone associated toxicity on testicular functions. Abstr.58th Ann. Meet.Endocrine Society, San Francisco, CA.
Abdollahi, M. Ranjbar, A. Shadnia, S. Nikfar, & S. Rezaie, A. (2004). Pesticides and oxidative stress: a review. Medical Science Monitor, 10, 141-147.
Aravindakshan, J. Paquet, V. Gregory, M. Dufresne, J. Fournier, M. Marcogliese, DJ., & Cyr, DG. (2004). Consequences of xenoestrogen exposure on male reproductive function in spottail shiners (Notropishudsonius).Toxicological Sciences, 78, 156-165. http://dx.doi.org/10.1093/toxsci/kfh042
Butler, AM. Murray, M. (1993). Inhibition and inactivation of constitutive cytochrome P450 in rat liver by parathion.Molecular Pharmacology, 43, 902-908.
Boe-Hansen, GB, Fedder, J. Ersbøll, AK., & Christensen, P. (2006). The sperm chromatin structure assay as a diagnostic tool in the human fertility clinic. Human Reproduction. 21, 1576-82. http://dx.doi.org/10.1093/humrep/del019
Bal, R. Naziroglu, M. Turk, G. Yilmaz, & O. Kuloglu, T. (2012)a. Insecticide imidacloprid induces morphological and DNA damage through oxidative toxicity on the reproductive organs of developing male rats. Cell Biochemistry and Function. 30, 492-499.
Bal, R. Turk, G. Tuzcu, M. Yilmaz, O., & Kuloglu, T. (2012)b. Assessment of imidacloprid toxicity on reproductive organ system of adult male rats. Journal of Environmental Sciences and Health B. 47, 434-444. http://dx.doi.org/10.1080/03601234.2012.663311
Clement, JG. (1985). Hormonal consequences of organophosphate poisoning.Fundamental Applied Toxicology. 5, 67-77. http://dx.doi.org/10.1016/0272-0590(85)90115-0
Corrigall, WA. Franklin, KBI. Coen, KM. Clarke, PBS. (1992). The mesolimbic dopamine system is implicated in the reinforcing effects of nicotine. Psychopharmacology. 107, 285-289. http://dx.doi.org/10.1007/BF02245149

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 72
Cocco, P. (2002). On the rumors about the silent spring. Review of the scientific evidence linking occupational and environmental pesticide exposure to endocrine disruption health effects. Cadernos de SaudePublica. 18, 379-402. http://dx.doi.org/10.1590/S0102-311X2002000200003
Crain, A. Eriksen, M. Iguchi, T. Jobling, S. Laufer, H. LeBlanc, GA., & Guillette, LJ. (2007). An ecological assessment of bisphenol-A: evidence from comparative biology. Reproductive Toxicology. 24, 225-239. http://dx.doi.org/10.1016/j.reprotox.2007.05.008
Cataño, HC. Carranza, E.Huamaní, C., & Hernández, AF. (2008). Plasma cholinesterase levels and health symptoms in Peruvian farm workers exposed to organophosphate pesticides.Archives of Environmental Contamination and Toxicology. 55, 153-9. http://dx.doi.org/10.1007/s00244-007-9095-0
Di Chiara, G. (2000). Role of dopamine in the behavioural actions of nicotine related to addiction. European Journal of Pharmacology. 393, 295-314. http://dx.doi.org/10.1016/S0014-2999(00)00122-9
de Montgolfier, B. Fournier, M.Audet, C.Marcogliese, DJ., & Cyr, DG. (2008). Influence of municipal effluents on the expression of connexins in the brook trout (Salvelinusfontinalis) testis.Aquatic Toxicology. 86, 38-48. http://dx.doi.org/10.1016/j.aquatox.2007.09.013
De Souza, A. Medeiros, AR. De Souza, AC. Wink, M.Siqueira, IR. Ferreira, MB. Fernandes, L.Loayza, MP., & Torres, IL. (2011). Evaluation of the impact of exposure to pesticides on the health of the rural population: Vale do Taquari, State of Rio Grande do Sul (Brazil). CienSaude Colet, 16, 3519-3528.
Evamarie, S. Wolfgang, S.Egon, K. Matthias, B.Margitta, JM., & Hans, JR. (1999). Disruption of male sex hormones with regard to pesticides: pathophysiological and regulatory aspects. Toxicology Letters. 107, 225-231. http://dx.doi.org/10.1016/S0378-4274(99)00051-X
Evenson, DP. Larson, KL., & Jost, LK. (2002). Sperm Chromatin Structure Assay: its clinical use for detecting sperm DNA fragmentation in male infertility and comparisons with other techniques. Journal ofAndrology. 23, 25−43.
Figa-Talamanca, I.Traina, ME., & Urbani, E. (2001). Occupational exposures to metals, solvents and pesticides: recent evidence on male reproductive effects and biological markers. Occupational Medicine. 51, 174-188. http://dx.doi.org/10.1093/occmed/51.3.174
Frazier, LM. (2007). Reproductive disorders associated with pesticide exposure. Journal ofAgromedicine. 12, 27-37.
Florencia, R. Marianela, G. Marcelo, AZ. Cora, S., & Milena, D. (2009). Prenatal exposure to pesticides disrupts testicular histoarchitecture and alters testosterone levels in male Caiman latirostris. General and Comparative Endocrinology. 162, 286-292. http://dx.doi.org/10.1016/j.ygcen.2009.03.032
Glisson, SN. Karczmar, AG., & Barnes, L. (1974). Effects of diisopropylphosphorofluoridate on acetylcholine, cholinesterase and catecholamines of several parts of rabbit

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 73
brain.Neuropharmacology. 13, 623-631. http://dx.doi.org/10.1016/0028-3908(74)90052-5
Gupta, M.Bagchi, GK. Gupta, SD.Sasmal, D.Chatterjee, T., & Dey, SN. (1984). Changes of acetylcholine, catecholamines and amino acid in mice brain following treatment with Nuvacron and Furadan.Toxicology. 30, 171-175. http://dx.doi.org/10.1016/0300-483X(84)90127-6
Gerald, AL. Lisa, JB., & Vickie, SW. (1997)Pesticides: multiple mechanisms of demasculinization. Molecular and Cellular Endocrinology. 125, 1-5.
Guillette, LJ., & Edwards, TM. (2008). Environmental influences on fertility: can we learn lessons from studies of wildlife? Fertility and Sterility. 89, 21-24. http://dx.doi.org/10.1016/j.fertnstert.2007.12.019
Gore, AC. (2010). Neuroendocrine targets of endocrine disruptors. Hormones. 9, 16-27. http://dx.doi.org/10.14310/horm.2002.1249
Hill, RN.Erdreich, LS.Paynter, OE. Roberts, PA. Rosenthal, SL., & Wilkinson, CF. (1989). Thyroid follicular cell carcinogenesis.Fundamental and Applied Toxicology. 12, 629-697. http://dx.doi.org/10.1016/0272-0590(89)90001-8
IPCS. (2002). Global Assessment of the State-of-the-Science of Endocrine Disruptors.International Program of Chemical Safety, World Health Organization, Geneva, Switzerland.
Jobling, S. Beresford, N. Nolan, M. Rodgers-Gray, T. Brighty, GC.Sumpter, JP., & Tyler, CR. (2002). Altered sexual maturation and gamete production in wild roach (Rutilusrutilus) living in rivers that receive treated sewage effluents. Biology of Reproduction. 66, 272-281. http://dx.doi.org/10.1095/biolreprod66.2.272
Jobling, S. Williams, R. Johnson, A. Taylor, A. Gross-Sorokin, M. Nolan, M. Tyler, CR. Van, AR. Santos, E., & Brighty, G. (2006). Predicted exposures to steroid estrogens in U.K. Rivers correlate with widespread sexual disruption in wild fish populations. Environmental Health Perspectives. 114, 32-39. http://dx.doi.org/10.1289/ehp.8050
Kokka, N. Clemons, GK., & Lomax, P. (1987). Relationship between the temperature and endocrine changes induced by cholinesterase inhibitors.Pharmacology. 34, 74-79. http://dx.doi.org/10.1159/000138255
Kuhar, MJ. Ritz, MC., & Boja, JW. (1991). The dopamine hypothesis of the reinforcing properties of cocaine. Trends in Neurosciences.14, 299-302. http://dx.doi.org/10.1016/0166-2236(91)90141-G
Kumar, S. (2004). Occupational exposure associated with reproductive dysfunction. Journal of Occupational Health. 46, 1-19. http://dx.doi.org/10.1539/joh.46.1
Kirby, ED. Geraghty, AC. Ubuka, T. Bentley, GE., & Kaufer, D. (2009). Stress increases putative gonadotropin inhibitory hormone and decreases luteinizing hormone in male rats. Proceedings of the National Academy of Sciences USA. 106, 11324-11329.

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 74
Kapoor, U. Srivastava, MK. Bhardwaj, S. Srivastava, LP. (2010). Effect of imidacloprid on antioxidant enzymes and lipid peroxidation in female rats to derive its. No Observed Effect Level (NOEL). Journal of Toxicological Sciences. 35, 577-581. http://dx.doi.org/10.2131/jts.35.577
Kapoor, U. Srivastava, MK. Srivastava, LP. (2011). Toxicological impact of technical imidacloprid on ovarian morphology, hormones and antioxidant enzymes in female rats.Food Chemical Toxicology. 49, 3086-3089. http://dx.doi.org/10.1016/j.fct.2011.09.009
Khan, DA. Ahad, K. Ansari, WM., & Khan, H. (2013). Pesticide exposure and endocrine dysfunction in the cotton crop agricultural workers of southern Punjab, Pakistan. Asia Pacific Journal of Public Health. 25,181-91. http://dx.doi.org/10.1177/1010539511417422
Mitsushima, D. Hei, DL., & Terasawa, E. (1994). GABA is an inhibitory neurotransmitter restricting the release of luteinizing hormone-releasing hormone before the onset of puberty. Proceedings of the National Academy of Sciences USA. 91, 395-399.
Murray, M., & Butler, AM. (1995). Identification of a reversible component in the in vitro inhibition of rat hepatic cytocrome P450 2B1 by parathion.Journal of Pharmacology and Experimental Therapeutics. 272, 639-644.
Murphy, LL. Munoz, RM. Adrian, BA., & Villanu, MA. (1998). Function of cannabinoid receptors in the neuroendocrine regulation of hormone secretion.Neurobiology of Diseases. 5, 432-446. http://dx.doi.org/10.1006/nbdi.1998.0224
Meeker, J. Ryan, L. Barr, D., & Hauser, R. (2006).Exposure to nonpersistent insecticides and male reproductive hormones. Epidemiology. 17, 61-8. http://dx.doi.org/10.1097/01.ede.0000190602.14691.70
Milnes, MR. Bermudez, DS. Bryan, TA. Edwards, TM. Gunderson, MP. Larkin, IL. Moore, BC., & Guillette, JR. (2006). Contaminant-induced feminization and demasculinization of nonmammalian vertebrate males in aquatic environments.Environmental Resistance. 100, 3-17. http://dx.doi.org/10.1016/j.envres.2005.04.002
Manfo, FP. Moundipa, PF., & Déchaud, H. (2010). Effect of agropesticides use on male reproductive function: a study on farmers in Djutitsa (Cameroon). Environmental Toxicology.Epub ahead of print.
Mostafalou, S., & Abdollahi, M. (2012)a. Concerns of environmental persistence of pesticides and human chronic diseases. Clinical and Experimental Pharmacology. 2, 1000-1108.
Oberdörster, E., & Cheek, AO. (2001). Gender benders at the beach: endocrine disruption in marine and estuarine organisms. Environmental Toxicology Chemistry. 20, 23-36. http://dx.doi.org/10.1002/etc.5620200103
Orlando, EF., & Guillette, JR. (2007). Sexual dimorphic responses in wildlife exposed to endocrine disrupting chemicals. Environmental Resistance. 104, 163-173. http://dx.doi.org/10.1016/j.envres.2006.06.002

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 75
Padungtod, C. Lasley, BL.Christiani, DC. Ryan, LM., & Xu, X. (1998).Reproductive hormone profile among pesticide factory workers.Journal of Occupational and Environmental Medicine. 40, 1038-1047. http://dx.doi.org/10.1097/00043764-199812000-00002
Piña-Guzman, B. Solís-Heredia, MJ., & Quintanilla-Vega, B. (2005). Diazinon alters sperm chromatin structure in mice by phosphorylating nuclear protamines. Toxicology Applied Pharmacology.202, 189-98. http://dx.doi.org/10.1016/j.taap.2004.06.028
PAN. (2009). List of Lists: A Catalogue of Lists of Pesticides Identifying Those Associated with Particularly Harmful Health or Environmental Impacts. P. A. Network.
Rogelio, R. Guadalupe, OG. Javier, MM., & Victor, BA. (2005). Pesticide Exposure Alters Follicle-Stimulating Hormone Levels in Mexican Agricultural Workers. Environmental Health Perspectives. 113, 1160-1163. http://dx.doi.org/10.1289/ehp.7374
Ramazzini, C. (2009). Collegium Ramazzini statement on the control of pesticides in the European Union: a call for action to protect human health. American Journal of Industrial Medicine. 52, 176-177. http://dx.doi.org/10.1002/ajim.20654
Straube, E.Straube, W. Kruger, E.Bradatsch, M. Jacob-Meisel, M., & Rose, HJ. (1999). Disruption of male sex hormone with regard to pesticides: pathophysiological and regulatory aspects. Toxicology Letter. 107, 225-231. http://dx.doi.org/10.1016/S0378-4274(99)00051-X
Sarkar, R. Mohanakumar, KP., & Chowdhury, M. (2000).Effects of an organophosphate pesticide, quinalphos, on the hypothalamopituitary- gonadal axis in adult male rats.Journal of Reproduction and Fertility. 118, 29-38.
Spassova, D. White, T., & Singh, AK. (2000). Acute effects of acephate and metamidophos on acetylcholinesterase activity, endocrine system and amino acid concentration in rats.Comparative Biochemistry and Physiology part C Toxicology and Pharmacology. 126, 79-89.
Stoker, C. Rey, F. Rodriguez, HA. Ramos, JG. Sirosky, P.Larriera, A. Luque, EH., & Muñoz-de-Toro, M. (2003). Sex reversal effects on Caiman latirostris exposed to environmentally relevant doses of the xenoestrogenbisphenol.American General and Comparative Endocrinology. 133, 287-296. http://dx.doi.org/10.1016/S0016-6480(03)00199-0
Sergerie, M. Laforest, G. Bujan, L. Bissonnette, F., & Bleau, G. (2005). Sperm DNA fragmentation: threshold value in male fertility. Human Reproduction. 20, 3446-51. http://dx.doi.org/10.1093/humrep/dei231
Stoker, C. Beldoménico, PM. Bosquiazzo, VL. Zayas, MA. Rey, F. Rodríguez, H. Muñoz-de-Toro, M., & Luque, EH. (2008). Developmental exposure to endocrine disruptor chemicals alters follicular dynamics and steroid levels in Caiman latirostris. General and Comparative Endocrinology. 156, 603-612. http://dx.doi.org/10.1016/j.ygcen.2008.02.011
Shojaei, SH., & Abdollahi, M. (2012). Is there a link between human infertilities and exposure to pesticides? Internal Journal of Pharmacology. 8, 708-710.

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 76
Terasawa, E., & Fernandez, DL. (2001). Neurobiological mechanisms of the onset of puberty in primates.Endocrine Review. 22, 111-151.
Tomizawa, M.Casida, JE. (2005). Neonicotinoid insecticide toxicology: mechanisms of selective action. Annual Review of Pharmacology and Toxicology. 45, 247-268. http://dx.doi.org/10.1146/annurev.pharmtox.45.120403.095930
Tiemann, U. (2008). In vivo and in vitro effects of the organochlorine pesticides DDT, TCPM, methoxychlor, and lindane on the female reproductive tract of mammals: a review. Reproductive Toxicology. 25, 316-326. http://dx.doi.org/10.1016/j.reprotox.2008.03.002
Van den, BKJ. Van Raaij, JAGM. Bragt, PC., & Notten, WRF. (1991). Interactions of halogenated industrial chemicals with transthyretin and effects on thyroid hormone levels in vivo.Archive of Toxicology. 65, 15-19.
Watkins, SS.Koob, GF., & Markou, A. (2000). Neural mechanisms underlying nicotine addiction: acute positive reinforcement and withdrawal. Nicotine Tobacco Research. 2, 19-37. http://dx.doi.org/10.1080/14622200050011277
Copyright Disclaimer
Copyright for this article is retained by the author(s), with first publication rights granted to the journal.
This is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 77
In Vitro Antibacterial Activity of Two Mosses: Calymperes Erosum C. Mull and Bryum coronatum
Schwaegr from South-Western Nigeria
Tedela P. O., Adebiyi, A. O., & Aremu, A.
Department of Plant Science
Ekiti State University, P. M. B. 5363, Ado-Ekiti, Nigeria
David, O. M. (Corresponding author)
Department of Microbiology, Ekiti State University, P. M. B. 5363, Ado-Ekiti, Nigeria
E-mail: [email protected]
Received: February 9, 2014 Accepted: February 24, 2014
doi:10.5296/jbls.v5i2.5730 URL: http://dx.doi.org/10.5296/jbls.v5i2.5730
Abstract
Bryophytes are poikilohydric in nature and the oldest known land plant. Their striking resistance to microbial attack suggests their inherent production of antibacterial compounds. The antibacterial activity of acetone, ethanol, methanol and hexane extracts of Calymperes erosum C. Mull and Bryum coronatum Schwaegr were investigated against twenty clinically important bacteria pathogens. Agar dilution method was used to assess the effectiveness of the extracts on the test organisms. The minimum inhibitory concentrations of the extracts of C. erosum were between ˂0.625 and ˃5.0 mg/ml. Klebsiella pneumoniae ATCC 10031, Enterococcus feacalis ATCC 29212, Bacillus pumilis ATCC 14884 and Enterobacter cloaca ATCC 13047 in decreasing order are most sensitive to the extracts while Proteus vulgaris KZN, Staphylococcus aureus OK2 and Shigella sonnei ATCC 29930 were resistant to the extracts. Ethanolic extract was the most effective among the extracts followed by acetone extract. B. coronatum had relatively lower activity. While the mosses screened proved to be promising sources of antimicrobial and biologically active compounds, their toxicity and action mechanism still needed to be investigated.
Keyword: Calymperes erosum, Bryum coronatum, Mosses, Antibacterial, Pathogen, bryophytes

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 78
1. Introduction
Bryophytes represent the second largest group of land green plants after angiosperms and are taxonomically placed between algae and pteridophytes (Asakawa, 2007). The bryophytes group consist of three subgroups: Bryophyta (mosses), Marchantiophyta (liverworts) and Anthocerotophyta (hornworts) (Campbell and Reece, 2002). It is estimated that there are between about 15000 and 25000 bryophyte species known in the world (Goffinet and Shaw, 2009).
Bryophytes have long been considered to be insignificant in the economy of man except for those used in packing, plugging and decoration. However, Saxena and Harrinder (2004) reported that the ecological role of bryophytes in any ecosystem is significant. In China where herbal medicine is extensively and widely accepted, bryophytes have been used as “crude drugs”. Many medicinal bryophytes have been recognized (Madson and Pates, 1952; Wu, 1977; Ding, 1982; Ando, 1983). In particular, the occurrence of antibiotic substances in bryophytes has been documented by scientists (Dulger, 2005; Bodade et al, 2008; Russell, 2010; Elibol et al, 2011; Savaroglu et al, 2011). Although bryophytes normally grow in humid habitats, they are relatively free from microbial attacks and this scarcity of disease indicates that bryophytes are able to elaborate constitutive or inducible small – molecule antimicrobials. In fact, bryophytes have been proven to be a rich source of antibiotics and attempts to find potent, non- toxic broad spectrum antibiotics from the sources have been widely undertaken (Xie and Lou, 2009). However, reports of ethnobotanical researches into this plant group are minimal (Zhu et al, 2006). The reasons for this are the difficulty that researchers have with their identification, the limited amount of the same species available for analyses due to their inconspicuous position in the ecosystem and the difficulty with which analysis can be conducted since it relies on sophisticated methods (Savaroglu, 2011). Though, few studies have successfully investigated the chemistry of bryophytes (Asakawa, 2001; Jockovic et al, 2008), it is generally known that bryophytes possess extremely high amounts of terpenoids, phenolics (flavonoids and bibenzyl derivatives), glycosides, fatty acids as well as some rare aromatic compounds (Jockovic et al, 2008; Sabovljevic et al, 2008).
Recently, the public demand for herbal medicine and the rise of antibiotic-resistant bacteria have motivated scientists to look for new natural sources with potential pharmaceutical capabilities (Cowan, 1999). Zhu et al (2006) suggested that bryophytes are one of the most significant and promising sources of antibiotics and biologically active compounds in nature.
This study analysed the antimicrobial activities of different extracts of Calymperes erosum C. Mull and Bryum coronatum Schwaegr against some bacterial species.
2. Materials and methods
2.1 Plant Materials
Plant materials were collected in March 2012 in growing areas in Ekiti State University, Ado-Ekiti. The species was authenticated at the Herbarium section of the Department of Plant Science, Ekiti State University, Ado-Ekiti, Nigeria. Voucher specimens were prepared and deposited in the Herbarium of the University for reference. Plant samples were air dried for

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 79
four days and pulverized. Powdered plant material (40 g each) was separately extracted in acetone, ethanol and methanol for 48 h with periodic manual shaking after which the extracts were filtered through Whatman No. 1 filter paper. The extracts were evaporated to dryness under reduced pressure at 40°C using a rotary evaporator (Laborota 4000-efficient, Heldolph, Germany). Individual crude extracts was diluted using 5% dimethylsulphoxide to give 50 mg/mL stock solution (Taylor et al., 1995). This was then diluted to the required concentrations for the bioassay.
2.2 Test Organisms
Twenty bacterial strains used in this study were obtained from the Department of Microbiology, Ekiti State University, Ado-Ekiti, Nigeria. Eight Gram positive were used, the isolates include: Bacillus cereus ATCC 10702, Bacillus pumilis ATCC 14884, Bacillus subtilis KZN, Enterococcus feacalis ATCC 29212, Micrococcus luteus, Staphylococcus aureus ATCC 6538, Staphylococcus aureus OK1 and Staphylococcus aureus OK2. The Gram negative organisms used include: Acinetobacter calcoaceticus CSIR, Escherichia coli ATCC 25932, Escherichia coli ATCC 8739, Enterobacter cloaca ATCC 13047, Klebsiella pneumoniae ATCC 10031, Klebsiella pneumoniae KZN Pseudomonas aeruginosa ATCC 19582, Proteus vulgaris KZN, Proteus vulgaris ATCC 6830, Proteus vulgaris CSIR 0030, Serratia mercesscens ATCC 9986 and Shigella sonnei ATCC 29930. Each strain was maintained on Nutrient Agar (Oxoid) plates. The grown cultures were used for preparation of bacterial suspensions in sterile distilled water with densities adjusted to 0.5 McFarland Standard.
2.3 Antibacterial Activity Assay
Antibacterial activity was determined by the methods of Afolayan and Meyer (1997) using Mueller-Hinton agar (Oxoid). Briefly, different concentrations of the extracts were prepared in 5% Dimethyl Sulfoxide (DMSO) while diluent (5% DMSO) was used as control. Molten medium containing the extracts at final concentrations of 0.625, 1.25, 2.50 and 5.00 mg/ml were poured into Petri dishes, swirled gently until the agar began to set, and left over night for solvent to evaporate completely. Agar plates containing 1% of the extracting solvents were used as controls. The test organisms were streaked in radial pattern on the agar plates, incubated under aerobic conditions at 37 oC and examined after 24 h. Each treatment was performed in triplicate and complete suppression of growth at a specific concentration of an extract was required for it to be declared active (Sindambiwe et al., 1999; Mathekga et al., 2000). The lowest concentration that inhibits the growth of the organisms was recorded and considered as the minimum inhibitory concentration (MIC) value.
Table 1. Minimum inhibitory concentrations (MICs) of extracts of C. erosum
Gram Reaction Isolates Extractants
Acetone Ethanol Methanol Hexane
Positive B. cereus ATCC 10702 2.50 ˂0.625 1.25 ˃5.0 B. pumilis ATCC 14884 2.50 ˂0.625 2.50 ˂0.625 B. subtilis KZN 0.625 ˃5.0 5.00 ˃5.0

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 80
Enter. feacalis ATCC 29212 2.50 ˂0.625 1.25 0.625 M. luteus 2.50 5.00 5.00 2.50 St. aureus ATCC 6538 2.50 ˂0.625 ˃5.0 2.50 St. aureus OK1 ˃5.0 1.25 5.00 5.00 St. aureus OK2 2.50 5.00 ˃5.0 5.00
Negative
A. calcoaceticus CSIR 0.625 1.25 ˂0.625 2.50 E. coli ATCC 25932 2.50 ˂0.625 ˃5.0 2.50 E. coli ATCC 8739 1.25 ˂0.625 ˃5.0 2.50 Ent. cloaca ATCC 13047 1.25 ˂0.625 2.50 5.00 K. pneumonia ATCC 10031 1.25 ˂0.625 0.625 ˂0.625 K. pneumonia KZN 2.50 1.25 ˃5.0 ˂0.625 Ps. aeruginosa ATCC 19582 1.25 ˂0.625 2.50 ˂0.625 P. vulgaris KZN ˃5.0 5.00 5.00 2.50 P. vulgaris ATCC 6830 ˂0.625 ˂0.625 5.00 ˃5.0 P. vulgaris CSIR 0030 0.625 2.50 ˃5.0 2.50 Se. mercescens ATCC 9986 2.50 1.25 ˃5.0 5.00 Sh. sonnei ATCC 29930 5.00 5.00 ˃5.0 2.50
Table 2. Minimum inhibitory concentrations (MICs) of extracts of B. coronatum
Gram Reaction Isolates Extractants
Acetone Ethanol Methanol Hexane
Positive
B. cereus ATCC 10702 ˃5.00 ˃5.00 2.50 2.50 B. pumilis ATCC 14884 1.25 ˃5.00 5.00 ˃5.00 B. subtilis KZN ˃5.00 5.00 ˃5.00 ˃5.00 Enter. feacalis ATCC 29212 ˃5.00 ˃5.00 0.625 ˃5.00 M. luteus 5.00 ˃5.00 ˃5.00 ˃5.00 St. aureus ATCC 6538 ˃5.00 ˃5.00 ˃5.00 ˃5.00 S. aureus OK1 ˃5.00 2.50 2.50 0.625 S. aureus OK2 5.00 5.00 ˃5.00 ˃5.00
Negative
A. calcoaceticus CSIR 0.625 0.625 1.25 5.00 E. coli ATCC 25932 ˃5.00 ˃5.00 ˃5.00 ˃5.00 E. coli ATCC 8739 ˃5.00 0.625 2.50 ˃5.00 Ent. cloaca ATCC 13047 ˃5.00 ˃5.00 ˃5.00 0.625 K. pneumonia ATCC 10031 ˃5.00 5.00 ˃5.00 ˃5.00 K. pneumonia KZN ˃5.00 ˃5.00 ˃5.00 ˃5.00 Ps. aeruginosa ATCC 19582 1.25 ˃5.00 5.00 0.625 P. vulgaris KZN 0.625 0.625 ˃5.00 0.625 P. vulgaris ATCC 6830 ˃5.00 ˃5.00 ˃5.00 ˃5.00 P. vulgaris CSIR 0030 ˃5.00 ˃5.00 0.625 ˃5.00 Se. mercesscens ATCC 9986 2.50 ˃5.00 0.625 ˃5.00 Sh. sonnei ATCC 29930 2.50 ˃5.00 5.00 1.25

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 81
3. Results and Discussion
The emerging resistant bacteria pathogens proved to be serious threat to human health globally (Sanchez and Kouznetsov, 2010; Lopez-Pueyo et al., 2011). Treatment with orthodox medicine is ineffective in most cases due to different resistant mechanisms hence the need for cheap, effective and natural medicinal compounds (Basile et al., 1998; Saboljevic et al., 2006). Nearly all bryophytes are known to resist fungi and bacteria attack. They are also not consumed by molluscs, insects and mammals (Basile et al., 1998; Asakawa, 2001). Compared to vascular plants, the use of bryophytes as antibacterial agent is scanty despite their worldwide occurrence. The antibacterial properties of acetone, ethanol, methanol and hexane extracts of C. erosum and B. coronatum were screened against twenty medically important bacteria. The extracts showed varying degrees of inhibitory effects on the test organisms with the control (5% DMSO) has no inhibition on the organisms. Most of the active substances in medicinal plants are in aromatic or saturated organic nature. Some of the substances are reported to be easily extracted by polar and non-polar solvents (Cowan, 1999; Altuner et al., 2011). The MICs ranges were outside the test concentrations. They were between ˂0.625 and ˃5.0 mg/ml. Klebsiella pneumoniae ATCC 10031, Enter. feacalis ATCC 29212, B. pumilis ATCC 14884 and Ent. cloaca ATCC 13047 in decreasing are most sensitive to the extracts of C. erosum while P. vulgaris KZN, S. aureus OK2 and S. sonnei ATCC 29930 were resistant to the extracts. Mosses are a rich source of secondary metabolites with antimicrobial activity (Asakawa, 1981, Sabovljevic et al., 2006; Asakawa, 2007; Mellegard et al., 2009). Different antimicrobial agents have been isolated from the bryophytes and they are used for the treatment of infectious diseases (Dulger et al., 2005; Singt et al., 2006; Dulger et al., 2009; Bukvieki et al., 2012). Ethanolic extract was the most effective among the extracts followed by acetone extract. Ethanol and acetone invariably extract the active compound from the plant at a rate or amount higher than the other extractants. According to Nikolajeva et al. (2012), ethanolic extract has the best activity against pathogenic bacteria. They reported that extracts of seven out of 11 bryophytes were effective in curtailing the growth of Staphylococcus aureus. Khanam et al. (2011) reported ethanolic extracts of Marchantia palmata to have high antibacterial activity. However, Mukhopadhyay et al. (2013) reported low activity of ethanolic extracts of some bryophytes. Methanolic extract had the least effectiveness among the extracts. The extract of non-polar solvent (hexane) had low activity against the isolates though its activity is relatively better than methanol as shown in Table 1. The activity of the extracts of B. coronatum was very poor compared to the C. erosum. The activity of B. coronatum was pronounced against A. calcoaceticus CSIR, P. vulgaris KZN and S. aureus OK1. The action of the extracts may be due to their actions on the cell wall or any other parts of the test organisms as suggested by Olofin et al. (2013). Acetone extract was very active against A. calcoaceticus CSIR and P. vulgaris KZN while the hexane extract was active against S. aureus OK1 (Table 2). Relatively, Gram negative bacteria were more resistant / susceptible to the extracts of C. erosum while the extracts of B. coronatum of had low inhibitory activity on both Gram positive and negative bacteria. The activity of the mosses on the test organisms may be due to the presence of the secondary metabolites present in the mosses (Singh et al., 2007; Chaudhary and Kumar, 2011). The results of the preliminary screening the both organic and inorganic extracts of two bryophytes tested against the test organisms showed different biological activity considering the

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 82
extraction solvents. The active nature and the mechanism of action(s) of the active compounds in the two plants are open to further investigations.
References
Afolayan, A. J., & Meyer, J. M. (1997). The antimicrobial activity of 3, 5, 7-trihydroxyflavone isolated from the shoots of Helichrysum aureonitens. J. Ethnopharmacol. 57, 177-181. http://dx.doi.org/10.1016/S0378-8741(97)00065-2
Altuner, E. M., Ceter, T. P., Bayar, E. N., Aydin, S. N., Arici, F. R., Suleymanolu G. Z., & Edis, E. (2011). Investigation on antimicrobial effects of some moss species collected from Kastamonu Region. Commun. Fac. Sci. Univ. Ank. 23(1-2), 33-43.
Asakawa, Y. (1981). Biologically active substances obtained from bryophytes. The J. Hattori Botanic. Lab..50, 123-142.
Asakawa, Y. (2001). Recent advances in phytochemistry of bryophytes acetogenins, terpenoids and bis(bibenzyl)s from selected Japanese, Taiwanese, New Zealand, Argentinean and European liverworts. Phytochem. 56, 297-312. http://dx.doi.org/10.1016/S0031-9422(00)00454-4
Asakawa, Y. (2007). Biologically active compounds from bryophytes. Pure and Appl. Chem. 79(4), 557-580. http://dx.doi.org/10.1351/pac200779040557
Basile, A., Sorbo, S., Giordano, S., Lavitola, A. & Castaldo, C. R. (1998). Antibacterial activity in Pleurochaete squarrosa extract (Bryophyta). Inter. J. Antimicrob. Agents. 10, 169-172. http://dx.doi.org/10.1016/S0924-8579(98)00028-4
Bodade, R. G., Borkar, P. S., Saiful, A. M., & Khobragade, C. N. (2008). In vitro screening of Bryophytes for antimicrobial activity. J. Med. Plants Res. 7(4), 23-28.
Bukvieki, D., Veljic, M., Sokovic, M., Grujic, S., & Marin, P. D. (2012). Antimicrobial activity of methanol extracts of Abietinella abietina, Neckera crispa, Platyhypnidium riparoides, Cratoneuron filicinum and Campylium protensum mosses. Arch. Biol. Sci., Belgrade. 64(3), 911-916.
Campbell, N. A., & Reece, J. B. (2002). Biology, 6th edition, Pearson Education, Inc., San Francisco, CA
Chaudhary, B. L., & Kumar, P. (2011). Antibacterial activity and preliminary phytochemical screening of epiphytic moss Stereophyllum ligulatum Jaeg. Intern. J. Pharma. Bio. Sci. 2(4), 674-681.
Cowan, M. M. (1999). Plant products as antimicrobial agents. Clin. Microbiol. Rev.12(4), 564-582.
Da Silva, C. M., Mota, F. B. C., Rodrigues, M. D., Alves, O. D., & Maia, V. H. (2013). Crude extracts and semi-fractions from Myracrodruon urundeuva with antibacterial activity against American Type Culture Collection (ATCC) strains of clinical relevance. J. Med. Plants Res. 7(32), 2407-2413. http://dx.doi.org/10.5897/JMPR12.1264

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 83
Dulger, B., Hacioglu, N., & Uyar, G. (2009). Evaluation of antimicrobial activity of some mosses from Turkey. Asian J. Chem. 21, 4093-4096.
Dulger, B., Yayintas, O. T., & A. Gonuz (2005). Antimicrobial activity of some mosses from Turkey. Fitoterapia. 76, 730-732. http://dx.doi.org/10.1016/j.fitote.2005.07.006
Elibol B, Ezer T, Kara R, Yuvali C., & Colak E (2011). Antifungal and antibacterial effects of some acrocarpic mosses. Afr. J. Biotechnol., 10(6), 986-989.
Goffinet B & Shaw AJ (2009). Bryophyte Biology. Cambridge University Press, Cambridge, UK, p.565.
Jockovic, N., Andrade, P. B., Valentao, P., & Sabovljevic, M. (2008). HPLC-DAD of phenolics in bryophytes Lunularia cruciate, Brachytheciastrum velutinum and Kindbergia praelonga. J. Serb. Chem. Soc. 73, 1161-1167. http://dx.doi.org/10.2298/JSC0812161J
Khanam, R., Chaudhary, B. L. Khanam, S., & Kumar, P. (2011). Antibacterial Activity of Marchantia palmata Nees, Asian J. Biochem Pharmaceut. Res. 1(2), 27-36.
Lopez-Pueyo, M. J., Barcenilla-Gaite, F., Amaya-Villar, R., & Garnacho-Montero, J. (2011). Antibiotic multiresistance in critical care units. Med. Intensiva. 35:41-53.
Madsen, G. C., & A. L. Pates. (1952). Occurrence of antimicrobial substances in chlorophyllose plants growing in Florida. Botanical Gazette. 113, 293-300. http://dx.doi.org/10.1086/335721
Mellegard, H., Stalheim, T., Hormazabel, V., Granum, P. E., & Hardy, S. P. (2009). Antibacterial activity of sphagnum acid and other phenolic compounds found in Sphagnum papillasum against food-borne bacteria. Lett. Appl. Microbiol. 49, 85-90. http://dx.doi.org/10.1111/j.1472-765X.2009.02622.x
Mukhopadhyay S. T., Mitra S., Biswas A., Das N., & Poddar-Sarkar, M. (2013). Screening of antimicrobial and antioxidative potential of selected Eastern Himalayan mosses. Eur. J. Med. Plants. 3(3), 422-428.
Nikolajeva, V., Liepina, L., Petrina, Z., Krumina, G., Grube, M., & Muiznieks, I. (2012). Antibacterial activity of extracts from some bryophytes. Adv. Microbiol. 2, 345-353. http://dx.doi.org/10.4236/aim.2012.23042
Olofin, T. A., Akande, A. O., & Oyetayo, V. O. (2013). Assessment of the antimicrobial properties of fractions obtained from bryophytes. J. Microbiol. Antimicrob. 5(5), 50-54. http://dx.doi.org/10.5897/JMA11.055
Russell MD (2010). Antibiotic activity of extracts from some Bryophytes in South Western British Columbia. Med. Std. J. Austr. 2, 9-14.
Saboljevic, A., Sokovic, M., Saboljevic, M., & Grubisic, D. (2006). Antimicrobial activity of Bryum argenteum. Fitoterapia. 77, 144-145. http://dx.doi.org/10.1016/j.fitote.2005.11.002
Sanchez, J. G. B., & Kouznetsov, V. V. (2010). Antimycobacterial susceptibility testing

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 84
methods for natural products research. Braz. J. Microbiol. 41, 270-277. http://dx.doi.org/10.1590/S1517-83822010000200001
Savaroglu F, Filik IC, Oztopcu VP, Kabadere S, IIhan S, & Uyar R (2011). Determination of antimicrobial and antiproliferative activities of the aquatic moss, Fontinalis antipyretica Hedw. Turk. J. Biol. 35, 361-369.
Saxena, K., & Harinder, S. (2004). Uses of bryophytes. Reson., 9(6), 56-65. http://dx.doi.org/10.1007/BF02839221
Singh, M., Rawat, A. K. S., & Govindarajan R. (2007), Antimicrobial activity of some Indian mosses, Fitoterapia. 78, 156-158. http://dx.doi.org/10.1016/j.fitote.2006.10.008
Wu, P.C. (1977). Rhodobryum giganteum (Schwaegr.) par. can be used for curing cardiovascular disease. Acta phytotax. SIN, 15, 93.
Xie, C. F., & Lou, H. X. (2009). Secondary metabolities in bryophytes: An economical aspect. Chem. Biodiversity. 6, 303-312. http://dx.doi.org/10.1002/cbdv.200700450
Zhu, R.L., Wang D., Xu L., Shi R.P, Wang J., & Zheng, M. (2006). Antibacterial activity in extracts of some bryophytes fron China and Mongolia. J. Bot. Lab. 100, 603-615.
Copyright Disclaimer
Copyright for this article is retained by the author(s), with first publication rights granted to the journal.
This is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 85
Phytohormones Influence on the In-Vitro Regeneration of Saccharum Officinarum L. from Apical Meristem
Culture
F. E. Lawyer (Corresponding author)
Biotechnology Unit, National Centre for Genetic Resources and Biotechnology
PMB 5382 Moor plantation, Ibadan, Oyo state, Nigeria
Tel: 234-806-251-7996 E-mail: [email protected]
Z. O. Jamaleddine
Tel: 234-805-856-7113 E-mail: [email protected]
P. T. Lyam
Tel: 234-806-350-2051 email: [email protected]
I. T. Borokini
Tel: 234 806 350 2051 E-mail: [email protected]
A. A. Adedeji
Tel: 234-805-764-6689 E-mail: [email protected]
A. U. Okere
Tel: 234-803-604-2204 E-mail: [email protected]
Received: January 10, 201 Accepted: January 2 , 201
doi:10.5296/jbls.v5i2.4885 URL: http://dx.doi.org/10.5296/jbls.v5i2.4885

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 86
Abstract
Growth regulators especially auxins and cytokinins are critical for plant in-vitro regeneration. The effect of these plant growth regulators on in-vitro propagation of Saccharum officinarum L (Sugarcane) was investigated. In vitro response of two different varieties of sugarcane (NCS 005 and NCS 008) to Plant Growth Regulators was obtained in this study. Formation of buds was obtained on shoot apical meristem when cultured on MS (Murashige and Skoog) medium supplemented with 0.1mg/l BAP (6-Benzylaminopurine). After two weeks of initiation, regenerated meristem was inoculated into MS (Murashige and Skoog) fortified with different concentrations and combination of cytokinins. Shoot multiplication was optimal on 0.5mg/l BAP + 0.25 mg/l Kin(Kinetin) for NCS 005 variety while for NCS 008 variety, no significant (P≥0.05) difference was observed between 1.5mg/l BAP and 1.5mg/l BAP +0.5mg/l Kin. The best root induction for in vitro derived shoots was obtained on 1.0 mg/l NAA (Naphthalene acetic acid) and 2.0 mg/l IBA( Indole butyric acid) for both varieties of sugarcane within ten days of culture transfer. Successfully established plantlets showed excellent growth response when weaned under regulated green house conditions.
Keywords: Plant growth regulators, Murashige and Skoog, 6-Benzylaminopurine, Naphthalene acetic acid, Indole butyric acid, Kinetin
1. Introduction
Saccharum officinarum L (Sugarcane) a perennial herb belonging to the Poaceae family.It is one of the most important sugar crops in the world which accounts for about 70% of the world’s total sugar production (Anonymous, 2005). This distinguishing quality place a high value on the species as a commercial crop especially in Nigeria where it has been highly cultivated. Saccharum officinarum is a polyploid plant that is grown in different parts of the world. Sugarcane importance has increased over time and in recent years becoming an important industrial raw material for sugar industries and other industries producing alcohol, and animal feed. Saccharin officinarum is mainly propagated vegetative by two to three budded sets. This traditional practice results in high production cost and also a large amount of sugarcane involvement in the cultivation process. Considering the importance in the agricultural industry, concerted efforts are being made for its improvement (Desai et al 2004). Tissue culture technique is an alternative method to overcoming these challenges and to produce uniform propagules for the cultivation, (Nickell, 1964, Heinz and Mee 1969). Micro propagation is important for rapid multiplication of elite genotypes/clones and for the quick spread of new varieties (Nickell and Heinz, 1973). Micro propagation is established, not only as means of clonal and rapid propagation it is also more efficient in a viable and successful method for the production of pathogen free planting materials. In recent decades, in vitro shoot tip culture for mass propagation of sugarcane has been reported by (Hendre et al, 1983; Lee, 1987; Nand and Singh,1994.). In vitro regeneration through leaf sheath culture and plant regeneration through callus culture (Nickell, 1977) has also been reported. The in vitro nutrition requirements for Saccharum officinarum vary according to genotypes as well as explants used. In vitro cultivation method has proven to be an efficient regeneration method for mass propagation of this crop and production of uniform and disease free propagules for

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 87
potential and quality production of sugarcane. In order to ensure highest possible yield and quality of field crops, the disease free stock can be extended to the growers and out growers by using this technique (Aamir et al 2008). Present research focuses on the importance of tissue culture technology in Saccharum officinarum improvement and establishment of efficient micro propagation protocols for healthy, disease free and premium quality planting materials in Nigeria.
2. Material and Methods
Two varieties, NCS005 and NCS008 were obtained from National Cereal Research Institute (NCRI), Badeggi, Nigeria. The varieties were established on the field gene bank of the National Centre for Genetic Resources and Biotechnology (NACGRAB), Ibadan. Apical sections, containing 6-8cm long spindle segment with growth tips and young leaf shealths were collected from the two varieties within 2-4 months old. These collected plants materials were taken into the NACGRAB Biotech lab, where they were washed under running tap water with addition of few drops of household liquid detergents for 20-30 minutes. The apical portions were immersed in 70% Ethanol for 5 min followed by disinfection with 50% v/v (volume per volume) commercial bleach for 20mins. These were further rinsed several times with sterile distilled water. Meristems were kept in 0.001%w/v (weight per volume) citric acid solution for 30 minutes before excision. All surface sterilization was done under sterile conditions. Using the left and right leaf shealths remover technique, the surface sterilized spindle segments were aseptically dissected out into 2 cm long meristems with the help of sterile surgical blades and forceps. The apical meristems were immediately inoculated in MS medium (Murashige and Skoog, 1962) containing 3%(w/v) sucrose, B5 vitamins, 0.01g/L ascorbic acid and 0.7%(w/v) agar. MS basal media was supplemented with different concentrations and combinations of Plant Growth Regulators. The pH of the medium was adjusted to 5.8 and autoclaved at 1210C and 15 lb for 15 min (Desai et al 2004). Cultures were maintained in the growth room at an incubation temperature of 23◦C ± 1◦C with 8 to 16 hour photoperiod.
For axillary shoot proliferation, budding meristems were transferred to MS containing increased cytokinin concentrations. Shoot formation and proliferation was observed after 7 - 20 days of shoot initiation. First sub-culturing was done after three weeks and subsequent sub-culturing after every fourth night. All dead shoots were removed at each sub-culture.
For in-vitro rooting, MS medium containing different concentrations and combination of NAA and IBA was used. Successfully rooted in-vitro sugarcane plantlets were acclimatized in a humid environment, prior to transfer to outside conditions. During acclimatization, plantlets were protected from high temperature and irradiance.
All data were subjected to analysis of variance (ANOVA) using SAS software package and the means were separated using Duncan Multiple Range Test at 5% level of significance (Gbadamosi et al 2013).
3. Results
3.1 Shoot Formation from Apical Meristem

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 88
Enlargement, swelling and browning of the sugarcane explants were observed after 8 day of culture in all the media combinations. After 10 – 15 days of culture auxiliary shoot proliferation occurred from the base of explants, these differ from each media due to the varieties.
3.1.1 Multiplication of Shoot
Generally, best results for shoot multiplication were obtained in semi solid medium. The effect of plant growth regulators (PGRs) revealed differences in growth responses for both varieties (see Tables 1 and 2). During secondary proliferation stage, lateral shoots were formed at the base of newly initiated shoots in both varieties. As a result, a dense mass of shoots (15-20) was developed in each culture bottle. Shoot multiplication was maintained by regular transfer to new medium. This was achieved by sub-dividing shoots in clusters containing 4-5 shoots and transferring them into fresh MS medium augmented with different concentrations and combinations of cytokinins. For NCS 005, BAP in combination with KIN provided better results for shoot formation as compared to the use of BAP alone. Among different concentrations used, 0.5 mg/l of BAP with 0.25 mg/l Kinetin conferred the best shoot formation response (Table 1). For NCS 008, the best results for shoot formation (4.0±0.0 ) was observed on BAP at concentration 1.5mg/L, with this being in proximity with shoot formation (3.6±0.33 )observed on 2.0mg/L BAP. However, for NCS 008, there was no significant difference (P≥0.05) observed between results obtained on MS +1.5mg/L and MS +1.5mg/L BAP +0.5mg/L Kin (Table 1).
3.1.2 Rooting of Regenerated Shoots
Root initiation in the successfully established shoots was achieved on MS medium supplemented with different auxin concentrations. The effect of different plant growth regulators on root length in Saccharum officinarum was significant (P≤0.05) (Table2). The best root induction for in vitro derived shoots was obtained on 1.0 mg/l NAA and 2.0 mg/l IBA for both varieties of sugarcane within ten days of culture transfer. An average shoot number of 6.0±0.57 and 10.0±1.67 were produced for NCS 005 and NCS008 respectively (see Table 2). The lowest mean value of 2.00 ± 0.00 cm was obtained in MS media that had 1.00 mg/l NAA and 0.50 mg/l IBA in case of NCS 005 while for NCS 008 MS media that had 0.50 mg/l NAA (Table 2).
Table1. Effect of different plant growth regulators and their concentration on establishment of shoot Variety, NCS 005
Media Concentration mg/l Number
of explants Shoot number/explants Shoot length(cm)
MS+BAP 0.5 20 1.00±0.00e 1.32±0.01d MS+BAP 1.0 20 1.33±0.33de 1.27±0.10d MS+BAP 1.5 20 1.67±0.33de 1.72±0.05d MS+BAP 2.0 20 2.00±0.00cd 1.81±0.10d MS+BAP 2.5 20 2.00±0.00cd 1.56±0.23d
MS+BAP+KIN 0.5+0.25 20 5.00±0.00a 4.56±0.37c

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 89
MS+BAP+KIN 1.0+0.25 20 3.00±0.00b 5.20±0.15c MS+BAP+KIN 1.5+0.50 20 2.66±0.33b 6.09±0.21b MS+BAP+KIN 2.0+0.50 20 2.66±0.66bc 6.66±0.33b MS+BAP+KIN 2.5+0.50 20 2.43±0.21bc 7.70±0.60a
Variety, NCS 008.
Media Concentration mg/l Number of
explants Shoot no/explants Shoot length (cm)
MS+BAP 0.5 20 2.00±0.00c 3.25±0.00ab MS+BAP 1.0 20 2.33±0.33c 3.90±0.00ab MS+BAP 1.5 20 4.00±0.00a 4.60±0.06a MS+BAP 2.0 20 3.60±0.33a 2.25±0.00b MS+BAP 2.5 20 2.66±0.33bc 4.50±1.25a
MS+BAP+KIN 0.5+0.25 20 2.00±0.00c 3.60±0.35ab MS+BAP+KIN 1.0+0.25 20 2.33±0.33c 4.00±0.45ab MS+BAP+KIN 1.5+0.50 20 4.00±0.00a 4.70±0.05a MS+BAP+KIN 2.0+0.50 20 3.33±0.33ab 3.80±0.11ab MS+BAP+KIN 2.5+0.50 20 2.30±0.33c 4.50±1.23a
Values (mean ± standard deviation) in a column followed by same letter are not significantly different (P ≥ 0.05)
Table2. Effect of different auxin and their concentrations on root initiation variety NCS 005.
Media Concentration mg/l Number of
explants Root number/explants Root length (cm)
MS+NAA 0.5 20 2.33±0.33de 1.10±0.01def MS+NAA 1.0 20 2.00±000e 1.76±0.56cd MS+NAA 1.5 20 3.00±0.00d 0.70±0.00f MS+NAA 2.0 20 2.66±0.33de 1.23±0.07def MS+IBA 0.5 20 2.00±0.00e 0.80±0.00f MS+IBA 1.0 20 2.00±0.00e 0.90±0.20ef MS+IBA 1.5 20 3.00±0.00d 1.10±0.00def MS+IBA 2.0 20 2.33±0.33de 1.60±0.40cde
MS+NAA+IBA 1.0+1.0 20 4.00±0.00c 2.10±0.00bc MS+NAA+IBA 1.0+2.0 20 6.00±0.57a 3.50±0.23a MS+NAA+IBA 2.0+1.0 20 5.00±0.00b 3.90±0.00a MS+NAA+IBA 2.0+2.0 20 5.00±0.00b 2.60±0.10b
Variety NCS 008
Media Concentration mg/l Number
of explants Root number/explants Root length (cm)
MS+NAA 0.5 20 2.33±0.33d 0.70±0.00f MS+NAA 1.0 20 2.33±0.33d 2.30±0.23cd

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 90
MS+NAA 1.5 20 3.00±0.00cd 2.30±0.00cd MS+NAA 2.0 20 4.30±0.66bcd 0.80±0.00f MS+IBA 0.5 20 4.00±0.00bcd 0.80±0.00f MS+IBA 1.0 20 5.00±0.00bc 2.00±0.00d MS+IBA 1.5 20 5.00±0.00bc 2.40±0.00cd MS+IBA 2.0 20 4.00±1.00bcd 2.26±0.23d
MS+NAA+IBA 1.0+1.0 20 6.00±0.00b 2.90±0.00b MS+NAA+IBA 1.0+2.0 20 10.00±1.67a 3.50±0.30a MS+NAA+IBA 2.0+1.0 20 5.00±0.00bc 1.50±0.00e MS+NAA+IBA 2.0+2.0 20 5.66±0.66b 2.73±0.17bc
Values (mean ± standard deviation) in a column followed by same letter are not significantly different (P ≥ 0.05)
Figures 1. Photograph showing the tissue cultured explants at different steps of the micropropagation
A B
C D

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 91
Key of Photograph showing the tissue cultured explants at different steps of the micropropagation
A. Apical meristem cultured in regeneration medium
B. Shoot induction of variety NCS 008 with auxiliary shoot after 10 days
C. Sub cultured sugarcane variety NCS 005 shoots after 2 weeks
D. Sugarcane in culture bottles
E. Sugarcane in the screen house.
3.1.3 Hardening and Acclimatization of Plants of in Vitro Raised Plants
The in-vitro plants were hardened and acclimatized in medium containing a mixture of in River Sand, Topsoil, coconut fibers (1:2:2) after three weeks of transplanting in screen house. After 21 days the potted plant were nurtured in poly pots placed under sunlight for further hardening of the plantlets. These seedlings were transfer to the field after another two weeks and a high rate of seedlings were transferred successfully. Hardening was carried out in screen house under natural light conditions.
4. Discussion and Conclusion
In graminaceous species, plant regeneration has been facilitated by the identification of appropriate explant and in vitro culture conditions (Vasil, 1987, Gbadamosi et al 2013). The method of producing large number of identical clones by in-vitro culture is being routinely used for wide range of plant species (Biondi, 1986, Aamir Ali et al 2008). In this study, different concentration and combination of auxin and cytokinins in MS Medium was successfully utilized for shoot organogenesis and multiple shoot regeneration from apical meristems of sugarcane. This concurs with report by Yutaka et al 1998, that combination of phytohormones often determines the course of morphogenesis (shoot organogenesis or embryogenesis). This study also reconfirms the new phase for plant multiplication for the production of efficient, genetically stable clonal germplasm and pathogen free stock (Hussey, (1983); and Kartha (1986). The best shoot formation and proliferation for both varieties was observed on BAP in combination with KIN. Many authors including Shukla, 1994; Geeta,
E

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 92
2001; Dhillon, et al 2002 and Aamil Ali et al 2008 have reported the use of kinetin in combination with BAP for shoot formation in sugarcane
When auxins were used singly, an average of two to three roots was observed in NCS005. Conversely, NCS008 showed a more roubust root formation on MS fortified with IBA only, as compared to MS fortified with NAA. This indicates that IBA is a more preferred auxin regarding effeiciency and yield for invitro root initiation in the NCS008 variety.
A combination of NAA and IBA was however observed to yield the best results for root formation in both varieties. (Anbalagan et al., 2000 and Nadgauda 2002) had earlier reported that a combination of two auxins (NAA and IBA) promotes rooting in sugarcane, also favoured the combination of NAA and IBA promote for rooting. Also the combination of IBA and NAA best for rooting, and this favoured both when he combined NAA and IBA.
Several authors including Yi et al,(2004), Pruski et al.,(2005) and Aamil Ali et al.,(2008) has also reported optimum results in the combination of NAA and IBA for rooting in Phragmites communis and two different varieties of sugarcane.
Following Sreenivasan and Sreenivasan (1992) which observed 90-95% survival of plantlets in poly screen house under shade, the simple method of hardening, adopted in this study facilitated the successful transfer of 900/0 of Saccharum officinarum plants from in vitro to ex vitro conditions.
This study indicates and strongly support the statement by (Aamil Ali et al 2008) that micro propagation is not only feasible but it can be used as the helpful tool for rapid multiplication of disease free, high yielding and premium quality planting material of highly adapted, genetically stable and newly released varieties of sugarcane.
Acknowledgments
The authors wish to acknowledge the National Centre for Genetic Resources and Biotechnology (NACGRAB), NCRI, Badeggi, Nigeria, for the support during the research work, Mr. Ibor Denis of NACGRAB for his effort in raising the sugarcane plantlet in the screen house and the establishment on the field and also to all that supported the research work.
Reference
Aamir A, Shagufta N., Fayyaz A. S., & Javed I. (2008). An efficient protocol for large scale production of sugarcane through micropropagation. Pak. J. Bot., 40(1), 139-149.
Anbalagan, S., A. Kalamani & M. Sakila. (2000). In vitro propagation of sugarcane: nature of callus, direct regeneration, regeneration through callus and morphological variations. Res. On Crops, 1(2), 138-140.
Anonymous, (2005), Crop husbandary for sugarcane mericlones. Sugarcane Breeding
Biondi, (1986) Practical application of in vitro propagation: Present situation and future prospects. Giom. Bot. Ital., 120, 29-42.
Desai N. S., Suprasanna P., & Bapat V. A. (2004). Simple and reproducible protocol for direct

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 93
somatic embryogenesis from cultured immature inflorescence segments of sugarcane (Saccharum spp.).Current science, vol. 87(6), 764-768.
Dhillon, B. D. S., G Raman & S. S. Gosal. (2002). Evaluation of different plant delivery system in micro propagated sugarcane (saccharum officinarum L.). Crop impro. 29(1), 35 - 40
Gbadamosi A. E., & Shaibu B. (2013). Influence of the phytohormones on the in-vitro regeneration in Senna alata (Linn). Academia Journal of Biotechnology, 1(3), 041-045.
Geeta, S., & D. Padmanabhan. (2001). Effect of hormones on direct somatic embryogenesis in sugarcane. Sugar Tech., 3(3), 120-121. http://dx.doi.org/10.1007/BF03014576
Heinz, D. J., & Mee, G. W. P., (1969), Plant differentiation from callus tissue of Saccharum species, Crop Sci., 9, 346-348. http://dx.doi.org/10.2135/cropsci1969.0011183X000900030030x
Hendre, R. R., Iyer, R. S., & Kotwal, M., (1983), Rapid multiplication of sugarcane by tissue culture. Sugarcane, 1, 58.
Hussey, G (1983). Plant Biotechnology. Cambridge University Press. Cambridge, 111-138.
Institute Report (Quarterly), 12, 2.
Jyoti R. Rout & William J. Lucas 1996 Characterization and manipulation of embryogenic response from in-vitro-cultured immature inflorescences of rice (Oryza sativa L.) Springer-Verlag, 198(1), 127-138
Kartha, K. K. (1986). Elimination of viruses in the presence of Antivirus chemicals production and indeang of disease free plants. 219-238.
Lee, T. S. G. (1986). Multiplication of sugarcane by apex culture. Tumalba, 36, 231-235.
Lee, T. S. G., (1987). Micro propagation of sugarcane (Saccharum sp.) Plant Cell Tissue and Organ culture, 10, 47-55. http://dx.doi.org/10.1007/BF00037496
Murashige, T., & Skoog, F., (1962), A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiol. Plant, 15, 473-497. http://dx.doi.org/10.1111/j.1399-3054.1962.tb08052.x
Nadgauda, R. S. (2002.). Need for tissue culture. Tissue culture pilot plant, National Chemical Laboratory, Pure 1-4.
Nand, L, & H. N. Singi., (1994). Rapid colonal multiplication of sugarcane through tissue culture plant .Tissue culture, 4, 1-7.
Nickell, L. G. (1977). Crop improvement in sugarcane: studies using in vitro methods. Crop Science. 17, 717-719. http://dx.doi.org/10.2135/cropsci1977.0011183X001700050011x
Nickell, L. G., & Heinz, D. J., (1973). Potential of cell and tissue culture techniques as aidsin economic plant improvement. In: Genes enzymes and populations. Plenum, N.Y., A. M. SRB Publications, 109-128.

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 94
Nickell, L.G., (1964). Tissue and Cell Culture of sugar-cane: Another research tool. Hawaii Planters Records. 57, 223-229.
Pruski, K., T. Astatkie, & J. Nowak. (2005). Tissue culture propagation of Mngolian cherry (Prunus fruticosa) and Nanking cherry (Prunus tomentosa) Plant Cell Tissues and Organ Culture., 82(2), 207-211. http://dx.doi.org/10.1007/s11240-004-7836-6
Shukla, R. A. Q. Khan & G. K. Gang. (1994). In vitro clonal multiplication of sugarcane; Optimization of media and hardening of plants. India. 89-94.
Sreenivasan, T. V., & Sreenivasan, J. (1992), Micro propagation of sugarcane varieties for Increasing cane yield. Sugar Tech Journal., 18, 61-64.
Vasil, I. K. (1987), Developing cell and tissue culture systems for the improvement of cereals and grass crops. Journal of Plant Physiology. 1987, 128, 192-218. http://dx.doi.org/10.1016/S0176-1617(87)80234-1
Yi, M., Yi. Guo, Yang, Z. Guo & G. Chen. (2004). Adventitious shoot bud formation and plant regeneration from in vitro-cultured stem segments of reed (Phragmites communis Trin.). In vitro cellular and developmental Biology, 40(4), 412-415. http://dx.doi.org/10.1079/IVP2004553
Yutaka,T. Y, Tomohiro, M. Toshikazu & O. Takeshi. (1998). Plant regeneration via shoot organogenesis from cotyledons in two wild Cucumis species, C. figarei and C. metuliferus Japan Agricultural Research Quarterly., 32, 281-286.
Copyright Disclaimer
Copyright for this article is retained by the author(s), with first publication rights granted to the journal.
This is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 95
The Impact of Wood, Cigarette and Marijuana Smoke on the Reproductive Health of Tandoor Occupants
Ghulam Nabi (Corresponding author)
M. Phil Scholar. Department: Animal sciences, Reproductive neuro-endocrinology Lab
Quaid-i-Azam University, (Islamabad) Pakistan
Tel: 92-345-811-2741 E-mail: [email protected]
Muhammad Amin
M. Phil Scholar, Department of Zoology, University of Karachi, Pakistan
Tel: 92-315-902-0833 E-mail: [email protected]
Jeena Urooj
MSc, Department of Zoology, University of Malakand
Chakdara, Dir (L), Khyber Pakhtunkhwa, Pakistan
E-mail: [email protected]
Muhammad Kamil
M. Phil Scholar, Department of Botany, University of Malakand, Pakistan
Tel: 92-343-925-6626 E-mail: [email protected]
Ayaz Ali Khan
Assistant Professor, Department of Biotechnology, University of Malakand
Chakdara, Dir Lower, Khyber pakhtunkhwa, Pakistan
Tel: 92-345-58860677 E-mail: [email protected]
Received: May 21, 2014 Accepted: June 3, 2014
doi:10.5296/jbls.v5i2.5654 URL: http://dx.doi.org/10.5296/jbls.v5i2.5654

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 96
Abstract
The objective of this study was to determine the effects of wood, cigarette and marijuana smoke on the reproductive health of tandoor occupants. A total of 100 male individuals were selected (50 control and 50 tandoor occupants). A standard questionnaire was designed regarding their age, economic status, marital status, fuel type, exposure time (per day), use of mask, addiction and reproductive health. Morning blood samples of 5 mL of the size were taken from all participants. Serums were obtained and analyzed for total serum testosterone concentration. Bio-check (USA) kit was used according to the manufacturer protocol and procedures for testosterone analysis. In control group the mean ± SEM of total serum testosterone was 671.9 ± 20.02 ng/dl where as in tandoor occupants it was 542.7 ± 16.40 ng/dl. There was a significant reduction (P**** < 0.0001) in total serum testosterone concentration in tandoor occupants as compared to control group. Reproductive health problems like, low libido, erection problems, infertility, decreased frequency for shaving and absent morning and nocturnal erection were common in tandoor occupants as compared to control group.
Wood, cigarette and marijuana smoke negatively affects testosterone concentration and lowers it significantly. This reduced testosterone concentration then produces ill effects like low libido, erection problems, infertility and absent morning and nocturnal erection.
Keywords: Libido, Infertility, Nocturnal
1. Introduction
Tandoor is a cylindrical clay oven used for cooking and baking in Bangladesh, Burma, Afghanistan, Iran, Turkey, India and Pakistan. The heat for a Tandoor is generated by a charcoal or wood fire, burning within the Tandoor itself, thus exposing the food to live-fire. Tandoor is used for cooking certain types of food such as, Tandoori chicken, chicken tikka and bread varieties like Tandoori Roti and Naan in the above countries (Ghulam et al., 2014).
The burning woods add toxic particulate matters (PM) into the air. Wood smoke in addition to particulate matter also emits dioxin, volatile organic compounds (VOCs), nitrogen oxides (NOx) and carbon monoxide (CO). VOCs may be either nauseating, noxious, and/or cancer causing. Polycyclic aromatic hydrocarbons (PAHs) include carcinogens such as benzopyrene, is also present in wood smoke. The chemical composition and total amounts of toxicants depend on how the wood is burned. It mostly depends upon the appliances used for wood burning. Lesser amount of toxicant is produced using high energy efficient wood burning appliances. Wood smoke PM contains wood tars, gases, dust, and ashes. The size of the particulate matter in air is lesser than 2.5 micron (Minnesota Pollution Control Agency, 2011).
Wood smoke exposure can cause cardiovascular events, coronary artery diseases, cerebro-vascular diseases, venous thrombo-embolism (Fraanchini, 2011) chronic obstructive pulmonary diseases (COPD) (Riddervold et al., 2011). During cooking PM concentration increases in kitchen, thus causing acute respiratory infection in children and lung cancer in women (Perez et al., 2010).

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 97
Like wood smoke diesel exhaust is also the important cause of air pollution and contains complex mixture of vapor-phase compounds and particulate matters. Smoke particles consist of carbon cores that adsorb many organic compounds including polycyclic aromatic hydrocarbons, heterocyclic organic compounds, quinines, aldehydes, and aliphatic hydrocarbons (Schuetzle et al,. 1981; Schuetzle, 1983; Li et al., 2000). These particles also contain substances having anti-androgenic, anti-estrogenic and estrogenic properties. The sperm production is severely reduced in adult rats due to decreased number of sertoli cells, when exposed to smoke particles during fetal life (Takeda et al., 2004). Smoke particles are discharged into the atmosphere enters our body via skin, lungs or the intestinal tract and affect reproductive functions (Yoshida et al., 1999; Yoshida et al., 2002; Tsukue et al., 2004; Oberdörster et al., 2005a; Oberdörster et al., 2005b ; Yoshida et al., 2006; Ono et al., 2007). In developing mice it has been shown that exposure to diesel exhaust causes degeneration of leydig cells, increase the number of damaged seminiferous tubules, and reduce daily sperm production (DSP) (Yoshida et al., 1999).
Cigarette smoking affects badly adrenal, thyroidal, pituitary, pancreatic, testicular and ovarian functions (Stárka et al., 2005). Cigarette smoke disrupts fertility in both sexes, lowering testosterone level and negatively shift spermiogram parameters in male (Trummer et al., 2002). Cigarette smoking also causes erectile dysfunction in male (Natali et al., 2005; Corona et al., 2005).
In humans as well as in animals models low plasma testosterone, abnormal spermatogenesis and decreased weight of accessory sex organs have been reported in males using marijuana or methadone (Azizi et al., 1973; kolodny et al., 1974; Cicero et al., 1975; Huang et al., 1978). Unlike methadone marijuana acts both directly and indirectly decreasing plasma testosterone levels (Symons et al., 1976; Smith et al., 1978).
Tandoor occupants are exposed to a heavy wood smoke for years. Particulate matter and other gases accumulate in their shops due to lack of proper exhaust system. In addition to wood smoke exposure they are also addicted to cigarette and marijuana. The present study is designed to determine the effects of wood, cigarette and marijuana smoke on testosterone level and their resulting consequences in tandoor occupants.
2. Materials and Methods
2.1 Informed Consent
Before starting the study informed consent was obtained from all the participants. Enough information was given to them about the purpose of the study. They were also assured that the collected information’s will be used only for scientific purposes.
2.2 Study Area
This study was conducted in district Lower Dir, Khyber pakhtunkhwa, Pakistan.
2.3 Questionnaire
A standard questionnaire was designed. Questions about their age, marital status, economic

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 98
conditions, type of fuel used, exposure time to wood smoke, addictions, use of mask and reproductive problems were asked for every participant.
2.4 Participants
A total of 100 male individuals were selected. Two groups were made. Control and exposed. Each group contains 50 participants. Control peoples were selected randomly from the same area. The age of both group participants ranges from 25 to 55 years and was divided into four age groups i.e. 25-34, 35-44, 45-49 and 50-55.
2.5 Blood Sampling
A morning 5 mL blood samples were taken from the brachial vein of all participants aseptically. Blood samples were collected in vacutainer tube containing no preservatives or additives. To prevent contamination of samples from smoke, collection tube and catheter site were covered. All the blood samples were centrifuged at 3200 rpm for 15 minutes at 4 °C. Serums were obtained and for latter analysis were stored at – 20 °C.
2.6 Exclusion Criteria
Participants with a history of tuberculosis, pneumonia, renal problems, hepatic problems, heart problems and individuals using medications were excluded from the study.
2.7 Assay
Testosterone enzyme immunoassay test kit, Bio-check (USA) was used for the determination of total serum testosterone level according to the manufacturer protocol and procedures.
2.8 Statistics
Software graph pad prism, version 6.03 (Graph Pad Software Inc., San Diego, CA, USA) was used for data analysis. All data were presented as mean ± SEM. Unpaired t test was used for the comparison of both groups. P<0.05 indicated the significant difference.
3. Results
3.1 History
History of both control and exposed group are summarized in Table 1.

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 99
Table 1. History of control and Tandoor occupants
Age Group (Years)
No. Economic status
Marital Status
Fuel type
Exposure Time (per day)
Addiction (%)
Mask Reproductive health status
Exposed 25-34
15 Poor 03 single 12 married
Wood ≥9 hrs. 02 Snuff 13 C*+ M*
No 05 low libido 06 LSF*
35-44 20 Poor 05 single 15 married
Wood ≥9 hrs. 04 Snuff 16 C*+ M*
No 05 EP* 03 low libido 02 infertility
05 AME*
45-49 10 Poor 00 single 10 married
Wood ≥9 hrs. 02 Snuff 08 C*+ M*
No 03 infertility 04 low libido
03 AME* 50-55 5 Poor 00 single
05 married Wood ≥9 hrs. 01 Snuff
02 C*+ M* No 01 infertility
04 low libido Control 25-34
15 Poor 05 single 10 married
No 00 02 Snuff
No 00
35-44 20 Poor 07 single 13 married
No 00 03 Snuff No 00
45-49 10 Poor 00 single 10 married
No 00 03 Snuff No 01 infertility
50-55 5 Poor 00 single 05 married
No 00 00 Snuff No 01 EP*
C*= Cigarette, M*= Marijuana, LSF*= Low Shaving Frequency, EP*= Erection Problems, AME*= Absent Morning Erection.
3.2 Total Serum Testosterone Analysis in Control and Tandoor Occupants
Testosterone were analyzed both in control and tandoor occupants. Their results are summarized in Table 2. The results showed that testosterone level is significantly lower (P**** < 0.0001) in tandoor occupants as compared to control group.
Table 2 Total serum testosterone concentration in control and Tandoor occupants
CI*= confidence interval
Parameters Control Mean ± SEM
Tandoor occupants Mean ± SEM
95% C.I* P value
Total serum testosterone
671.9 ± 20.02 (ngdl) 542.7 ± 16.40 (ng/dl)
-180.6 to -77.88 < 0.0001
4. Discussion
Tandoor occupants are exposed to wood smoke, automobile emission as well as they also smoke cigarette. Most of them are also addicted to marijuana. The present study focuses on the reproductive health of these peoples.
Leydig cells take up smoke particles and have shown cytotoxicity in cultured cells in vitro (Don Porto Carero et al., 2001; Renwick et al., 2001; Hussain et al., 2005). Particulate matters

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 100
and related organic compounds produce inflammatory responses in lungs, endothelial cells and alveolar macrophages (Hirano et al., 2003). These particulate matters causes repressions of the StAR (Steriodogenic Acute Regulatory protein) expression, thus affects the production of steroid hormone productions from leydig cells. This also causes Leydig cell degeneration, increase the number of damaged seminiferous tubules, and reduce daily sperm production (; Yoshida et al., 1999; Diemer et al., 2003; Murugesan et al., 2007). This has been reported that various factors like AP-1, C/EBPb, GATA4, and DAX-1 are responsible for the regulation of StAR gene expression. The induction of StAR in Leydig cells by nanoparticles may be associated with these factors. (Jana et al., 2008; Silverman et al., 2006; Manna et al., 2004). In rats particulates matters triggers the suppressions of several genes which are crucial for gonadogenesis. This also reduces expression of genes which encode enzymes that are responsible for androgenesis. Diesel particulate matters also contain polycyclic aromatic hydrocarbons that stimulate Ah receptors. It also contains substances that have anti-androgenic, anti-estrogenic and estrogenic activity. In rats exposure to heavy smoke during fetal life, reduces the expression of Ad4BP/SF-1 mRNA that ultimately affect gonadal development. Such rats have low daily sperm production due to insufficient number of sertoli cells (Takeda et al., 2004).
In rats this has been found that penile erection is badly affected by cigarette smoke. Cigarette smoke also reduces plasma testosterone level and the activity of nitric oxide synthase in corpus cavernosum, ultimately leading to erectile dysfunction (Zhan et al., 2009). In an experiment 12 rats were exposed to cigarette smoke generated by a machine and 12 were exposed to room air. Before and after plasma testosterone, Luteinizing Hormone (LH) and Follicle Stimulating Hormone (FSH) were measured by Elisa. Testes were also examined histologically. It was found that plasma testosterone reduces significantly in cigarette smoke exposed rats as compared to control. No significant changes were found for LH and FSH in both groups. However in histological examination of testes, fewer leydig cells and degeneration of the remaining cells were observed (Yardimci et al., 1997). Cigarette smoking reduces testosterone concentration (Alexandersen et al., 1996). Cigarette smoke contains nicotine that like cocaine stimulates mesolimbic dopamine system which increases extracellular dopamine concentration (Corrigall et al., 1992; Di Chiara, 2000; Watkins et al., 2000). This increase in dopamine suppresses reproduction by regulating the secretion of Gonadotropin Releasing Hormone (GnRH) negatively (Murphy et al., 1998; Mello and Mendelson, 2002).
Marijuana and its constituent delta-9-tetrahydrocannabinol (D-9-THC) affect the reproduction negatively by decreasing the level of testosterone (Okey and Truant 1975; Nashas, 1984). There are several evidences which confirm that marujana regulate the estrogen positively but androgen negatively. Different effects of marijuana suggest that different constituent affect the hormonal profile in a different way (Chakravarty et al., 1975; Dixit et al., 1975; Chakravarty et al., 1976; Solomon et al., 1976; Solomon and Cocchia, 1977; Solomon et al., 1977).
5. Conclusions
Chronic exposure to wood smoke, cigarette and marijuana, negatively regulate testosterone level. Low testosterone level then leads to ill effects such as infertility, low libido, erection

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 101
problems, decreased frequency for shaving and absent morning and nocturnal erection. Tandoor occupants are advised to use mask during working hours, improve exhaust system and if possible replaced the wood by gas. Further studies are needed to analyze the effects of individual gases on reproductive system.
References
Alexandersen, P. Haarbo, J., & Christiansen, C. (1996). The relationship of natural androgens to coronary heart disease in males: a review. Atherosclerosis, 125, 1-13. http://dx.doi.org/10.1016/0021-9150(96)05864-9
Azizi, F. Vagenakis, A., & longcope, C. (1973). Decreased serum testosterone concentration in male heroin and methadone addicts. Steroids, 22, 467-72. http://dx.doi.org/10.1016/0039-128X(73)90002-0
Chakravarty, I. Sengupta, D. Bhattacharya, P., & Ghosh, JJ. (1976). Effect of cannabis extract on the uterine monoamine oxidase activity of normal and estradiol treated rats. Biochemical pharmacology. 25, 377-378. http://dx.doi.org/10.1016/0006-2952(76)90335-X
Cicero, TJ. Bell, RD. Wiest, WG., & Allison, JH. (1975). Functions of the male sex organs in heroin and methadone uses, N. Eng. J. Med, 292, 882-87. http://dx.doi.org/10.1056/NEJM197504242921703
Corona, G. Mannucci, E. Petrone, L. Ricca, v. Mansani, R. Cilotti, A. Balercia, G. Chiarini, v. Giommi, R. Forti, G., & Maggi, M. (2005). Psychobiological correlates of smoking in patients with erectile dysfunction. Int j impot res, 17, 527-534. http://dx.doi.org/10.1038/sj.ijir.3901351
Corrigall, WA. Franklin, KBI. Coen, KM., & Clarke, PBS. (1992). The mesolimbic dopamine system is implicated in the reinforcing effects of nicotine. Psychopharmacology, 107, 285-289. http://dx.doi.org/10.1007/BF02245149
Dichara. G. (2000). Role of dopamine in the behavioural actions of nicotine related to addiction. European Journal of pharmacology, 393, 295-314. http://dx.doi.org/10.1016/S0014-2999(00)00122-9
Diemer, T. Allen, JA. Hales, KH., & Hales, DB. (2003). Reactive oxygen disrupts mitochondria in MA-10 tumor Leydig cells and inhibits steroidogenic acuten regulatory (StAR) protein and steroidogenesis. Endocrinology, 144, 2882-2891. http://dx.doi.org/10.1210/en.2002-0090
Dixit, VP. Gupta, CL., & Agarwal, M. (1977). Testicular degeneration and necrosis induced by chronic administration of cannabis extract in dogs. Endokrinologie, 69, 299-305.
Don, PC. Hoet, PH. Verschaeve, L. Schoeters, G., & Nemery, B. (2001). Genotoxic effects of carbon black particles, diesel exhaust particles, and urban air particulates and their extracts on a human alveolar epithelial cell line (A549) and a human monocytic cell line (THP-1). Environmental and Molecular Mutagenesis, 37, 155-163. http://dx.doi.org/10.1002/em.1023

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 102
Franchini, M., & Mannucci, PM. (2011). Thrombogenecity and cardiovascular effects of ambient air pollution. Journal of the American College of Cardiology, 52(9), 719-26.
Ghulam, N. Jeena , U. Ayaz, AK. Ghazala, YZ. Mussarat, W., & Saleem, UR. (2014). Effects of occupational exposure to wood smoke in Tandoor occupants. Journal of Biology and Life Sciences, 5(2) 37-47. http://dx.doi.org/10.5296/jbls.v5i2.4079
Hirano, S. Furuyama, A. Koike, E., & Kobayashi, T. (2003). Oxidative-stress potency of organic extracts of diesel exhaust and urban fine particles in rat heart microvessel endothelial cells. Toxicology, 187, 161-170. http://dx.doi.org/10.1016/S0300-483X(03)00053-2
Huang, HFS. Nahas, GG., & Hembree, WC. (1978). Morphological changes of spermatozoa during marijuana induced depression of human spermatogenesis, Presented at 62nd annual meeting of FASEB, Atlantic city, N.J. april 9-14, abstract, 2761.
Hussain, SM. Hess, KL. Gearhart, JM. Geiss, KT., & Schlager, JJ. (2005). In vitro toxicity of nanoparticles in BRL 3A rat liver cells. Toxicology in Vitro, 19, 975- 983. http://dx.doi.org/10.1016/j.tiv.2005.06.034
Jana, K. Yin, X. Schiffer, RB. Chen, JJ. Pandey, AK. Stocco, DM. Grammas, P., & Wang, X. (2008). Chrysin, a natural flavonoid enhances steroidogenesis and steroidogenic acute regulatory protein gene expression in mouse Leydig cells. Journal of Endocrinology, 197, 315-323. http://dx.doi.org/10.1677/JOE-07-0282
Kolodny, RC. Masters, WH. Kolodner, RM., & Toro, G. (1974). Depression of plasma testosterone levels after chronic intensive marijuana use. N. Eng. J. Med, 290, 872-74. http://dx.doi.org/10.1056/NEJM197404182901602
Li, N. Venkatesan, MI. Miguel, A. Kaplan, R. Gujuluva, C. Alam, J., & Nel, A. (2000). Induction of heme oxygenase-1 expression in macrophages by diesel exhaust particle chemicals and quinones via the antioxidant-responsive element. Journal of Immunology, 165, 3393-3401. http://dx.doi.org/10.4049/jimmunol.165.6.3393
Manna, PR. Eubank, DW., & Stocco, DM. (2004). Assessment of the role of activator protein-1 on transcription of the mouse steroidogenic acute regulatory protein gene. Molecular Endocrinology, 18, 558-573. http://dx.doi.org/10.1210/me.2003-0223
Mello, NK., & Mendelson, JH. (2002). Cocaine, hormones and behavior: clinical and preclinical studies, in Hormones, Brain and Behavior (Pfaff, D.W., Arnold, AP., Etgen, A.M., Fahrbach, S.E., and Rubin, R.T., eds) Academic Press, New York, pp 665-745. http://dx.doi.org/10.1016/B978-012532104-4/50106-2
Minnesota Pollution Control Agency., (2011). Health Effects of Wood Smoke; Home, Air Quality and Pollutants, General Air Quality, Wood Smoke. http://www.Pca.state.mn.us/indexphp/air/aiqualityandpollutants/generalairquality/woodsmoke/healtheffectswoodmokhtml.
Murphy, LL. Munoz, RM. Adrian, BA., & Villanu, MA. (1998). Function of cannabinoid receptors in the neuroendocrine regulation of hormone secretion. Neurobiol. Dis, 5, 432-446.

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 103
http://dx.doi.org/10.1006/nbdi.1998.0224
Murugesan, P. Balaganesh, M. Balasubramanian, K., & Arunakaran, J. (2007). Effects of polychlorinated biphenyl (Aroclor 1254) on steroidogenesis and antioxidant system in cultured adult rat Leydig cells. Journal of Endocrinology, 192, 325-338. http://dx.doi.org/10.1677/joe.1.06874
Nahas, GG. (1984) In: Nahas, G.G. (Ed.), Marihuana in Science and Medicine, Raven Press, New York, pp. 109-246.
Natali, A. Mondaini, N. Lombardi, G. Del, PG., & Rizzo, M. (2005). Heavy smoking is an important risk factor for erectile dysfunction in young men. International journal of impotence research, 17, 227-230. http://dx.doi.org/10.1038/sj.ijir.3901275
Oberdörster, G. Maynard, A. Donaldson, K. Castranova, V. Fitzpatrick, J. Ausman, K. Carter, J. Karn, B. Kreyling, W. Lai, D. Olin, S. Monteiro-Riviere, N. Warheit, D., & Yang, H. (2005)a. Principles for characterizing the potential human health effects from exposure to nano materials: elements of a screening strategy. Particle and Fibre Toxicology, 2, 8. http://dx.doi.org/10.1186/1743-8977-2-8
Oberdörster, G. Oberdörster, E. Oberdörster, J. (2005)b. Nanotoxicology: an emerging discipline evolving from studies of ultrafine particles. Environmental Health Perspectives, 113, 823-839. http://dx.doi.org/10.1289/ehp.7339
Okey, AB., & Truant, GS. (1975). Cannabis demasculinizes rats but is not estrogenic. Life Sciences, 17, 1113-1118. http://dx.doi.org/10.1016/0024-3205(75)90332-X
Ono, N. Oshio, S. Niwata, Y. Yoshida, S. Tsukue, N. Sugawara, I. Takano, H., & Takeda, K. (2007). Prenatal exposure to diesel exhaust impairs mouse spermatogenesis. Inhalation Toxicology, 19, 275-281. http://dx.doi.org/10.1080/08958370601069257
Perez, PR. Schil, MA., & Riojas, RH. (2010). Respiratory health effects of indoor air pollution. International Journal of Tuberculosis and Lung Diseases, 14(9), 1079-86.
Renwick, LC. Donaldson, K., & Clouter, A. (2001). Impairment of alveolar macrophage phagocytosis by ultrafine particles. Toxicology and Applied Pharmacology, 172, 119-127. http://dx.doi.org/10.1006/taap.2001.9128
Riddervold, IS. Bonlokke, JH. Molhave, L. Massling, A. Jenson, B. Gronborg, TK. Bossi, R. Forchhammer, L. Kiargaard, SK., & Sigsgaard, T. (2011). Wood smoke in a controlled exposure experiment with human volunteers. Inhalation Toxicology, 23(5), 277-88. http://dx.doi.org/10.3109/08958378.2011.567401
Schuetzle, D. (1983). Sampling of vehicle emissions for chemical analysis and biological testing. Environmental Health Perspectives, 47, 65-80. http://dx.doi.org/10.3109/08958378.2011.567401
Schuetzle, D. Lee, FS., & Prater, TJ. (1981). The identification of polynuclear aromatic hydrocarbon (PAH) derivatives in mutagenic fractions of diesel particulate extracts.

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 104
International Journal of Environmental Analytical Chemistry, 9, 93-144. http://dx.doi.org/10.1080/03067318108071903
Silverman, E. Yivgi-Ohana, N. Sher, N. Bell, M. Eimerl, S., & Orly, J. (2006). Transcriptional activation of the steroidogenic acute regulatory protein (StAR) gene: GATA-4 and CCAAT/enhancer-binding protein beta confer synergistic responsiveness in hormone-treated rat granulosa and HEK293 cell models. Molecular and Cellular Endocrinology, 252, 92-101. http://dx.doi.org/10.1016/j.mce.2006.03.008
Smith, CG. Besch, NF. Smith, RG., & Besch, PK. (1978). Mechanism for the anti- gonadotropic actions of tetrahydrocanabinol (THC) , Presented at 62nd annual meeting of FASEB, Atlantic city, N.J. april 9-14, abstract, 2687.
Solomon, J. Cocchia, MA. (1977). Is delta-9-tetrahydrocannabinol estrogenic? Science, 195, 905-906.
Solomon, J. Cocchia, MA. Dimartino, R. (1977). Effect of delta-9- tetrahydrocannabinol on uterine and vaginal cytology of ovariectomized rats. Science, 195, 875-877. http://dx.doi.org/10.1126/science.841311
Solomon, J. Cocchia, MA. Gray, R. Shattuck, D., & Vossmer, A. (1976). Uterotrophic effect of delta-9-tetrahydrocannabinol in ovariectomized rats. Science, 192, 559-561. http://dx.doi.org/10.1126/science.1257790
Stárka, L. Hill, M., & Králíková, E. (2005). Smoking and endocrine system. Diabetologie Metabolismus Endokrinologie Výživa, 8, 180-186.
Symons, AM. Telea, JD., & Marks, V. (1976). Effects of tetrahydrocanabinol on the hypothalamic pituitary gonadal system in the maturing male rat. Journal of endocrinology, 68, 43-44.
Takeda, K. Tsukue, N., & Yoshida, S. (2004). Endocrine-disrupting activity of chemicals in diesel exhaust and diesel exhaust particles. Environmental Sciences, 11(1), 33-45.
Trummer, H., & Habermann, H. (2002). The impact of cigarette smoking on human semen parameters and hormones. Human Reproduction, 17, 1554-1559. http://dx.doi.org/10.1093/humrep/17.6.1554
Tsukue, N. Yoshida, S. Sugawara, I., & Takeda, K. (2004). Effect of diesel exhaust on development of fetal reproductive function in ICR female mice. Journal of Health Science, 50, 174-180. http://dx.doi.org/10.1093/humrep/17.6.1554
Virag, R. Bouilly, P., & Frydman, D. (1985). Is impotence an arterial disorder? A study of arterial risk factors in 440 impotent men. Lancet, 1, 181-184. http://dx.doi.org/10.1016/S0140-6736(85)92023-9
Watkins, SS. Koob, GF., & Markou, A. (2000). Neural mechanisms underlying nicotine addiction: acute positive reinforcement and withdrawal. Nicotine Tobacco Research, 2, 19-37. http://dx.doi.org/10.1080/14622200050011277

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 105
Yardimci, S. Atan, A. Delibasi, T. Sunguroglu, K., & Guven, Mc. (1997). Long-term effects of cigarette-smoke exposure on plasma testosterone, luteinizing hormone and follicle-stimulating hormone levels in male rats. British Journal of Urology, 79(1), 66-9. http://dx.doi.org/10.1046/j.1464-410X.1997.28314.x
Yoshida, M. Yoshida, S. Sugawara, I., & Takeda, K. (2002). Maternal exposure to diesel exhaust decreases expression of Steroidogenic Factor-1 and mullerian inhibiting substance in the murine fetus. Journal of Health Science, 48, 317-324. http://dx.doi.org/10.1248/jhs.48.317
Yoshida, S. Ono, N. Tsukue, N. Oshio, S. Umeda, T. Takano, H., & Takeda, K. (2006). In utero exposure to diesel exhaust increased accessory reproductive gland weight and serum testosterone concentration in male mice. Environmental Sciences, 13, 139-147.
Yoshida, S. Sagai, M. Oshio, S. Umeda, T. Ihara, T. Sugamata, M. Sugawara, I., & Takeda, K. (1999). Exposure to diesel exhaust affects the male reproductive system of mice. International Journal of Andrology, 22, 307-315. http://dx.doi.org/10.1046/j.1365-2605.1999.00185.x
Zhan, XS. Wu, TP. Oyuang, H. Liu, XB., & Cai, HL. (2009). Cigarette smoke affects sexual function of male rats. Zhonghua Nan Ke Xue, 15(9), 788-91.
Zimmerman, AM. Zimmerman, S., & Raj, A. (1979). Effects of cannabinoids on spermatogenesis in mice. In: Nahas GG, Paton W, editors. Marihuana: biological effects. New York: Pergamon. p 407.
Copyright Disclaimer
Copyright for this article is retained by the author(s), with first publication rights granted to the journal.
This is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 106
Reproductive Biology and Bioturbatory Activities of Two Sympatric Species of Fiddler Crabs Uca lactea annulipes and Uca triangularis bengali (Decopada : Ocypodidae) at the East Midnapore Coastal Belt of
West Bengal, India
Subhasish Chatterjee
Department of Zoology, Vidyasagar University
Midnapore (West) -721102, West Bengal, India
Tel: 91-943-421-6333 E-mail: [email protected]
Tapas Kumar Das
Department of Zoology, Vidyasagar University
Midnapore (West) -721102, West Bengal, India
Susanta Kumar Chakraborty (Corresponding Author)
Department of Zoology, Vidyasagar University
Midnapore (West) -721102, West Bengal, India
Tel: 91-943-327-0591 E-mail: [email protected]
Received: March 3, 2014 Accepted: March 19, 2014
doi:10.5296/jbls.v5i2.5809 URL: http://dx.doi.org/10.5296/jbls.v5i2.5809
Abstract
Two sympatric species of fiddler crabs viz. Uca lactea annulipes and Uca triangularis bengali belonging to order decapoda, family ocypodidae have been taken into consideration in respect of their habitat preference, sex ratio, fecundity and role of bioturbatory activities in sustaining the coastal-estuarine-ecosystem dynamics at an ecotone (Talsari) located in the Midnapore

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 107
(East) coastal belt of West Bengal, India. A distinct segregation of habitats of these two species was evident from this field study. In general, monthly sex ratios were found to be clearly male biased for both species. Reproductive potential and intensity of the studied fiddler crabs have been measured by quantifying the relative frequency of ovigerous females and their fecundities. These two crabs have been observed to act as excellent bioturbator mediators and their bioturbatory activities were found to alter geochemistry of sediments by way of making semidomes (only male of U. lactea annulipes) and mudballs (both sexes of U. triangularis bengali). These bioturbatory structures used to play some roles in the social as well as sexual interactions of both species. Keywords: Fiddler crabs, Sex ratio, Fecundity, Bioturbation
1. Introduction
In many tropical environments more than one species of Uca coexist in similar habitats. They may have different patterns of microhabitat preferences, so that their ecological niches are not identical (Backwell et al., 2006). In this study, ecological characteristics of two sympatric species of Uca lactea annulipes and Uca triangularis bengali at the Talsari of the East Midnapore coastal belt of West Bengal, India were studied to note how their different ecological and ethological features by prevaling different density dependant and independant parameters.. Fiddler crabs (Ocypodidae, Uca) are a well-known group of small, intertidal brachyuran crabs (Backwell & Christy, 2000). They are widely distributed in nearly all tropical and subtropical regions of the world. Fiddler crabs burrow in intertidal areas and are important consumers of detritus, bacteria, fungi, and benthic microalgae in coastal marsh, mangroves, sand flat, and mudflat habitats (Backwell et al., 2006). Fiddler crabs are intertidal animals that live in mangrove forests, tidal creeks, sandbars, mudflats or, occasionally, stone or boulder beaches. They can occur in huge numbers, with thousands of individuals living in small, adjacent territories. Their burrowing activities aerate the soil and enhance the growth of marsh plants. Their presence also increases the amount of meiofauna in salt marshes (Burford et al., 2001). Fiddler crabs have long been a favourite animal for ecological studies (Christy et al., 2001). They are highly social animals with a rich behavioural repertoire. These crabs are best known for their remarkable sexual dimorphism: males have highly asymmetrical claws (chelipeds), with the major claw greatly enlarged (up to five times in length) relative to both the male’s own minor claw and the female’s two symmetrical minor claws (Rosenberg, 2002; Sweat, 2009). The major cheliped may constitute up to 48% of an adult male’s body mass and is considered a classic example of sexual selection (Pope, 2005; How et al., 2008). The claw is used for mating displays and in combat with other males. Another conspicuous difference is the form of the abdomen. In most male crabs; this is narrow and triangular in form, while females have a broader, rounded abdomen (Guillermo Guerao & Guiomar Rotllant, 2009). This is due to the fact that female crab’s brood fertilized eggs on their pleopods. Several taxonomic and ecological works on fiddler crabs of Hooghly-Matla estuarine complex and other coastal areas of India have been reported (Chakraborty & Choudhury, 1992). Fiddler crabs are diurnally active (Macia et al., 2001; Mokhtari et al., 2007). Surface activity terminates when the tide covers the area and the crabs retreat into their burrows and plug the entrances with sediment. Fiddler crabs, an important faunal component in the food web of the coastal belt play a

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 108
significant positive role in maintaining the steady state of the ecosystem and enhance its biological potentiality through accelerating the decomposition cycle as macro decomposers (Chatterjee & Chakraborty, 2014) influencing the ecosystem function to a large extent and are morphologically, physiologically and behaviorally well adapted to their environment. The burrow is a very important resource for the crabs (Zeil & Layne, 2002). It offers protection from aquatic predators during high tide and from aerial and terrestrial predators during low tide, when the crabs are active on the surface. It provides a safe refuge for moulting animals and for females while incubating their eggs. The burrow protects the crabs from desiccation during their activity on the surface by offering them access to water, which is needed for respiration and feeding. The burrow walls are important sites for nitrification and de-nitrification processes in the sediment. The burrow also serves as central hub from where the crabs venture out on their feeding excursions (Hemmi & Zeil, 2003). Fiddler crabs by their sheer numbers and abundance are a rich food source for birds (Zeil et al., 2006). They form important links between the primary detritus at the base of the food web and consumers of higher trophic levels (Koch & Wolff, 2002). Fiddler crabs being the most abundant and conspicuous invertebrates in many salt marshes (Montague, 1980), ecologically influence the salt marsh significantly by transferring of energy and nutrients within the marsh ecosystem (Daiber, 1982). Uca lactea annulipes and Uca triangularis bengali showed their co-existence with Suaeda maritima and Salicornia brachiat at present study site. Most of fiddler crabs from temperate and tropical estuaries, male to female ratios significantly exceeding unity have been documented by several researchers (Johnson, 2003). Males may be four times more abundant than females in some species (Emmerson, 1994). Generally, natural selection favours a sex ratio of 1:1 parental expenditure on offspring (May, 1983). Although deviant sex ratios are common in marine crustacean, owing to sequential hermaphroditism, sex specific habitat use, and differential growth rate (Johnson, 2003), the trend in Uca is notable in its widespread and consistent bias toward males. Male fiddler crabs are both territorial and promiscuous conditions that should favour female biased sex ratios. Ovigerous females spend prolonged periods underground and, when on the surface, often forage to closer to water sources, creating spatial separation from foraging males (Montague, 1980). Reproduction is the main mechanism that keeps species continuity so that it contributes to regulate the population size. Adaptations on mating systems jointly with environmental conditions are also factors that influence fiddler crab fecundity (Costa et al., 2006). Ovigerous females of two crabs were used to achieve and compare their reproductive pattern. The presence of ovigerous and non ovigerous female provided one index of reproductive periodicity. Fecundity in brachyuran crabs is commonly defined as the number of eggs produced per female per clutch (Reid & Corey, 1991). In brachyurans, body size is the main determinant of fecundity and it does depend on the allometric constraints on yolk storage within the cephalothorax (Lim, 2006).
The term ‘Bioturbation’ originates from ichnology, to describe traces of life in fossil and modern sediments, and has subsequently been adopted in soil and aquatic sciences (Meysman et al., 2006; Chatterjee et al., 2008; Wilkinson et al., 2009). Sedimentary environments are dynamic habitats where the sediments, the fundamental building blocks of the habitats, are continually structured both by the local physical regime. Both in terrestrial and aquatic environment, animal bioturbation results from comparable activities, including burrow and

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 109
mound construction, the lateral ‘ploughing’ of the surface (e.g. by moles or heart urchins), particle ingestion and egestion during foraging (e.g. deposit feeding, geophagy or lithophagy), food caching and prey excavation, wallowing and trampling, and the infilling of abandoned burrow structure (Meysman et al., 2006). In modern ecological point of view, bioturbation is now recognized as an archetypal example of ‘ecosystem engineering’ (Meysman et al., 2006; D’ Andrea & DeWitt, 2009) and act as major modulators of microbial activities and biogeochemical processes in aquatic environments (Mermillod-Blondin 2011).
Seasons are well pronounced in the coastal environment each with four months duration, Pre monsoon (March to June) with least rainfall and high temperature; monsoon (July to October) with high rainfall and moderate temperature; and post monsoon (November to February) with occasional rainfall and low temperature. The present investigation was undertaken to identify the habitat preference, sex ratio, fecundity and different features of bioturbatory activities during different seasons among the different tidal levels viz. LTL (Low Tidal level), MTL (Mid Tidal level), HTL (High Tidal level).
2. Materials and Methods
2.1 Physiography of Study Site
The coastal tract of Midnapore (East) West Bengal, India extends from the junction of longitudinal extension 87o 20/ E to 88o 5/ E and latitudinal extension 21o 30/ N to 22o 2/ N (Chakraborty et al., 2012). The present study was conducted at Talsari, an ecotone located at the confluence of Subarnarekha estuary with Bay of Bengal, between 21°35'48" Northern Latitude and 87°27'17" Eastern Longitude (Figure 1). This site is characterized in having mixed sand and mudflats endowed with degraded mangroves. The Subarnarekha estuarine delta is the westernmost unit of the topographic expression in the present coastal plain. A tract of mangrove swamp is situated around the estuary link of Talsari tidal channel and extensive saltmarsh tract extends on the more saline flats of eastward tidal basin.
Figure 1. Map showing the location of study site

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 110
2.2 Duration of the Study Period
Seasonal dynamics of reproductive parameters in respect of sex ratios (M : F), size classes (carapace width-CW), fecundity and different features of bioturbatory activities of two species of fiddler crabs were investigated in the study site through different months and seasons during March, 2009 to February, 2010.
2.3 Different Attributes of Sex Ratio, Size Class (Carapace Width) and Fecundity
Samples were collected monthly during low tide period on full moon during day time over a period of approximately 1 hour duration and covering an area of about 500 m2 of the study site for the assessment of sex ratio. Fecundity (the number of eggs per female) determines the reproductive potential of a species and the stock size of its population. On each sampling occasion, a total of twenty quadrats 1.0 m 2 (1.0 × 1.0 m) squares were set out at the study site. Ten quadrats were randomly chosen for sampling and each of which was completely excavated with a corer (0.08 m diameter) to a depth of 0.5 m. Crabs were collected by using diving knives or shovel with a catch effort and all ovigerous females after being unearthed were preserved in 70 % ethanol, bagged and stored until further processing. The carapace width (CW) were measured using a vernier caliper (± 0.05 mm accuracy) or, with the aid of a stereomicroscope (CW<10.0mm) respectively. To estimate fecundity, egg bearing females were selected for egg counting. Pleopods were removed from females, placed in Petri dishes filled up with seawater, and eggs were isolated by gradually adding a solution of sodium hypochlorite (7%). Bare pleopods were then separated by gentle stirring in a beaker filled with 200 ml seawater. Three sub samples (1.5 ml) were taken using a pipette, and eggs were counted under a dissecting microscope. The average value obtained was then extrapolated for the whole suspension to estimate the total number of eggs (Litulo, 2004).
2.4 Statistical Analyses
Different statistical analyses were done by following standard book (Zar, 2009) and utilizing ‘STATISTICA’ (STATSOFT, 2001) with the help of a P-4 computer. Quantification of different sexual forms of two fiddler crabs under present study (male-M, ovigerous female-OF and non ovigerous female -NOF) was analyzed following Chi square test (χ2-test, P<0.01) to test the independence of attributes between sex and months of availability using the frequency data of abundance. Such analysis was repeated for each type of species. Many needful pair wise simple correlation coefficients were calculated and tested by t test e.g. carapace width vs. fecundity. Factorial ANOVA technique was used to compare the main effects of seasons and tidal levels of different characteristics of bioturbatory activities. Means of main effect were further subjected to Duncan's test at 5% level of significance to test the homogeneity among respective means for each factor. Significance of all main and interaction effects were also tested by F tests on the results derived from each ANOVA analysis. Significant main effects for factors having more than two levels were subjected to Duncan’s test, where similar alphabets denote homogenous means at 5% level of significance.
3. Results
3.1 Habitat Preference

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 111
The habitats of two sympatric species of brachyuran, U. lactea annulipes and U. triangularis bengali showed their co-existence with the open mud flat endowed occasionally with saline herbs like Suaeda maritima and Salicornia brachiata. Both the species used to prefer a muddy marsh environment where the substrate is relatively free of dense underground networks of plant’s roots,but also stable enough to allow for construction of burrows. They were collected from the study site (Talsari) with maximum abundance at MTL and HTL respectively and it was the dominant species in the middle part of the muddy intertidal belt. Table 1 has revealed soil textural composition of the habitat of each species.
Table 1. Soil texture of the habitat of each species (%)
Texture (%) Pre Monsoon Monsoon Post Monsoon Uca lactea annulipes LTL MTL HTL LTL MTL HTL LTL MTL HTL
Sand 89.6 91.9 91.5 87.2 89.2 95.5 86.8 88.2 82.2 Silt 3.7 3.6 2.8 4.2 4.8 2.6 3.9 3.2 4.4
Clay 6.7 4.5 5.7 8.6 6 1.9 9.3 8.6 13.4 Uca triangularis bengali Sand 91.2 92 91.5 96.6 93.2 95.5 84.6 84.3 82.6
Silt 4.2 3.6 4.2 1.58 3.14 2.8 8.2 5.5 4.6 Clay 4.6 4.4 4.3 1.84 3.62 1.7 7.2 10.2 12.8
3.2 Sex Ratio and Fecundity
A total of 1313 individuals of U. lactea annulipes (752 males, 261 non-ovigerous females and 300 ovigerous females) were recorded during the study period. The number of crabs encountered and the monthly sex ratio of which has been listed in Table 2. In general, monthly sex ratios were found to be slightly male biased, except March, April, May, October, November and December during the study period. Among 300 ovigerous females of this species, 40 were used for fecundity studies (CW: 5.5-14.5), which showed a mean fecundity of 5930±3685 eggs and measured a mean size of CW(9.21± 2.49). Number of eggs of each size class have been mentioned in Table 3. Males (N=752) were more abundant than females (N=561), but overall sex ratio (1:0.75) was significant deviate from the 1:1 ratio and month has significant effect on the allocation of different sexes (χ2-test, P<0.01, 99% Table 4). Fecundity variation relative to carapace width was fitted into a power function given by the equation F= 1512.x -8889 and yielded a correlation coefficient of R2 =0.951. The scatter plot representing this correlation is shown on Figure 2.
Figure 2. Relation between egg number and carapace width of U. lactea annulipes (F=

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 112
fecundity, R2= determination coefficient, X axis=carapace width, Y axis=Egg Number)
A total of 556 of U. triangularis bengali (296 males, 158 non ovigerous females and 102 ovigerous females) were obtained throughout the study period. The number of crabs encountered and the monthly sex ratio of which has been listed in Table 5. In general, monthly sex ratios were a little male biased, except July, October, November, December and January. Among 102 ovigerous females of this species, 38 were used for fecundity study, which bare a mean fecundity of 3768±2616 eggs and measured a mean size of 8.68± 2.22 CW. Number of eggs for each size class have been mentioned in Table 6. Males (N=296) were more plentiful than females (N=260), but overall sex ratio (1:0.88) was significant from the 1:1 ratio and month has significant effect on the distribution of different sexes (χ2-test, P<0.01, 99% Table 7). Fecundity variation relative to carapace width was fitted into a power function given by the equation F=1121.x -6859 and yielded a correlation coefficient of R2 =0.909. The scatter plot representing this correlation is shown on Figure 3. Fecundity variation of both species in respect of carapace width and number of eggs was correlated and the result exposed for U. lactea annulipes (significance of r at 5%=0.312 and significance of r at 1%=0.403) and U. triangularis bengali (significance of rat 5%=0.320 and significance of rat 1%=0.413) (Table 8).
Figure 3. Relation between egg number and carapace width of U. triangularis bengali (F= fecundity, R2= determination coefficient, X axis=carapace width, Y axis=Egg Number)
Table 2. Number of individuals, percentage and sex ratios of Uca lactea annulipes
Months Mal (N) % NOF (N) % OF (N) % Total (N) Sex ratio (M:F) Mar.'08 50 41.32 15 12.40 56 46.28 121 1:1.42 Apr.'08 62 41.33 25 16.67 63 42.00 150 1:1.42 May,'08 42 48.84 32 37.21 12 13.95 86 1:1.05 Jun.'08 72 72.00 28 28.00 0 0 100 1:0.39 Jul.'08 68 79.07 18 20.93 0 0 86 1:0.26
Aug.'08 84 79.25 22 20.75 0 0 106 1:0.26 Sept.'08 95 84.07 18 15.93 0 0 113 1:0.19 Oct.'08 49 37.12 28 21.21 55 41.67 132 1:1.69 Nov.'08 51 47.22 8 7.41 49 45.37 108 1:1.12

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 113
Dec.'08 57 49.14 0 0 59 50.86 116 1:1.04 Jan.'09 66 57.89 44 38.60 4 3.51 114 1:0.73 Feb.'09 56 69.14 23 28.40 2 2.47 81 1:0.45 Total 752 57.27 261 19.88 300 22.85 1313 1:0.75
Mal, males; NOF, non-ovigerous females; OF, ovigerous females; M, males; F, females; N, Sample size.
Table 3. U. lactea annulipes; Mean fecundity (X) and Standard deviation registered for ovigerous females size classes (CW) (N= 40 ; CW= carapace width)
Size Class (mm) X±SD N 5.5-6.5 1818±60 3 6.5-7.5 2230±201 7 7.5-8.5 3487±255 6 8.5-9.5 4582±442 5
9.5-10.5 6187±179 6 10.5-11.5 7481±581 4 11.5-12.5 9730±1395 3 12.5-13.5 12070±1156 3 13.5-14.5 13283±480 3
Table 4. The result of Chi square (χ2-test, P<0.01) analysis of U. lactea annulipes between months and distribution of different sexes
Chi-Square Tests value df Asymp. Sig.(2- sided) Pearson Chi-Square 404.208 a 22 .000 Likelihood Ratio 503.210 22 .000 N of Valid Cases 1313 a. 0 cells (.0%) have expected count. The minimum expected count is 16.10.
Table 5. Number of individuals, percentage and sex ratios of Uca triangularis bengali
Months Mal (N) % NOF (N) % OF (N) % Total (N) Sex ratio (M:F) Mar.'08 45 78.95 12 21.05 0 0 57 1:0.27 Apr.'08 32 69.57 14 30.43 0 0 46 1:0.44 May,'08 28 71.79 11 28.21 0 0 39 1:0.39 Jun.'08 36 52.94 32 47.06 0 0 68 1:0.89 Jul.'08 24 36.36 42 63.64 0 0 66 1:1.75
Aug.'08 28 62.22 17 3.78 0 0 45 1:0.61 Sept.'08 21 65.63 11 34.38 0 0 32 1:0.52 Oct.'08 19 29.69 1 1.56 44 68.75 64 1:2.37 Nov.'08 22 40.74 0 0 32 59.26 54 1:1.45 Dec.'08 14 43.75 0 0 18 56.25 32 1:1.29 Jan.'09 11 33.33 14 42.42 8 24.24 33 1:2 Feb.'09 16 80.00 4 20.00 0 0 20 1:0.25 Total 296 53.24 158 28.42 102 18.35 556 1:0.88
Table 6. U. triangularis bengali; Mean fecundity (X) and Standard deviation registered for ovigerous females size classes (CW) (N= 38 ; CW= carapace width)
Size Class (mm) X±SD N
5.5-6.5 1472±43 3

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 114
6.5-7.5 1651±64 8
7.5-8.5 1961±133 7
8.5-9.5 2674±345 4
9.5-10.5 4065±683 5
10.5-11.5 5105±653 3
11.5-12.5 7512±1173 4
12.5-13.5 8867±440 4
Table 7. The result of Chi square (χ2-test, P<0.01) analysis of U. triangularis bengali between months and distribution of different sexes
Chi-Square Tests value df Asymp. Sig.(2- sided) Pearson Chi-Square 346.523 a 22 .000 Likelihood Ratio 378.551 22 .000 N of Valid Cases 556 a. 1 cells (2.8 %) have expected count less than 5.The minimum expected count is 3.67
Table 8. The result of correlation coefficient analysis in between carapace width and number of eggs
Uca lactea annulipes Uca triangularis bengali
CW (mm) CW (mm)
CW (mm) 1 CW (mm) 1
No of eggs 0.975 No of eggs 0.954
significance of rat 5=0.312 significance of rat 5%= 0.320
significance of rat 1%=0.403 significance of rat 1%= 0.413
3.3 Different Characteristics of Bioturbation
3.3.1 Burrow Depth and Diameter
All burrows of U. lactea annulipes contain air in the upper two-third parts and water only in the lowest one third part. During high tide, the burrow entrances remained totally closed with mud. The maximum depth of the burrow (0.941 m) was noticed at HTL during pre-monsoon and the minimum depth (0.340 m) was recorded at LTL during monsoon. The maximum burrow diameter (0.032 m) was recorded during monsoon at LTL and that of minimum (0.008 m) was found during pre-monsoon at HTL. The maximum depth (0.858 m) of the burrow of U. triangularis bengali was observed at HTL during pre-monsoon and the minimum depth (0.338 m) was recorded at LTL during monsoon. The maximum burrow diameter (0.028 m) was recorded during monsoon at LTL and that of minimum (0.006 m) was observed during post monsoon at HTL (Table 9).
3.3.2 Semidome and Mudball
Semidome is a roof covering a semicircular space i.e. half a dome. Only male U. lactea annulipes could construct semidomes by scrapping up bits of sediments from the surface with their first four walking legs on the side with the major claw, and depositing the sediment

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 115
materials around the burrow entrance displaying triangular shape (Figures 4A and 4B) which tended to become completely invisible during post monsoon at HTL throughout the study period. The maximum height of semidome (0.049 m) was recorded during monsoon at LTL and the minimum height (0.003 m) was documented during pre-monsoon at HTL. U. triangularis bengali also made mudball (Figure 5) which was composed of large wet soil pellets and the diameter of mudballs ranged from 0.013 m at MTL during post-monsoon to 0.001 m at HTL during monsoon (Table 9).
Figure 4. A. Uca lactea annulipes with a semidome, B. Lateral view of a semidome
Figure 5. Uca triangularis bengali with mudball
3.3.3 Seasonal Variation in Quantity of Excavated Mud
In U. lactea annulipes, the weight of the excavated mud varied from 0.063 kg at MTL during pre monsoon to 0.182 kg at LTL during monsoon and the same in U. triangularis bengali varied from to 0.055 kg at MTL during post monsoon to 0.162 kg at LTL during monsoon (Table 9) respectively.
3.3.4 Seasonal Variation in Distance (M) of Placement of Excavated Mud
The distance of placement of mudballs of U. lactea annulipes also varied from 0.057 m at HTL during pre-monsoon to 0.165 m at LTL during monsoon. The mudballs prepared by U. triangularis bengali were thrown to place of a distance of 0.039 m at HTL during post monsoon to 0.086 m at LTL during monsoon (Table 9).
3.3.5 Seasonal Variation in Frequency of Placement of Mudballs / 900 Seconds

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 116
U. lactea annulipes exihibited highest value in respect of frequency of placement of mudballs (38.50 times) at LTL during monsoon and minimum value (1.00 times) at HTL during post monsoon. In U. triangularis bengali, the frequency of placement of mudball was maximum (29.60 times) at LTL during monsoon and minimum (3.90 times) at HTL during post monsoon (Table 9).
Table 9. Seasonal variations in different characteristics of bioturbation
SL. No.
Species Pre monsoon Monsoon Post monsoon LTL MTL HTL LTL MTL HTL LTL MTL HTL
1 Burrow depth (m) U. lactea
annulipes 0.495
0.647
0.941
0.340
0.400
0.465
0.357
0.412
0.615
U. triangularis bengali
0.519
0.652
0.858
0.338
0.406
0.501
0.381
0.457
0.546
2 Burrow diameter (m) U. lactea
annulipes 0.022
0.017
0.008
0.032
0.024
0.018
0.014
0.015
0.012
U. triangularis bengali
0.026
0.018
0.013
0.028
0.025
0.018
0.021
0.018
0.006
3 Height / diameter of bioturbatory structures (m) Semidome
-Height U. lactea annulipes
0.005
0.004
0.003
0.049
0.029
0.025
0.007
0.006
0
Mudball- Diameter U .triangularis bengali
0.008
0.007
0.006
0.003
0.003
0.001
0.009
0.013
0.004
4 Excavated mud (in terms of weight-kg) U. lactea
annulipes 0.071
0.063
0.070
0.182
0.167
0.146
0.094
0.083
0.068
U .triangularis bengali
0.091
0.081
0.073
0.162
0.137
0.122
0.071
0.055
0.056
5 Distance of placement of excavated mudball (m) U. lactea
annulipes 0.086
0.060
0.057
0.165
0.156
0.064
0.105
0.092
0.095
U.triangularis bengali
0.079
0.064
0.049
0.086
0.067
0.064
0.069
0.049
0.039
6 Frequency of mudball placement / 900 s U. lactea
annulipes 20.50 12.30 7.70 38.50 17.80 9.50 13.60 10.80 1.00
U. triangularis bengali
13.70 10.30 5.90 29.60 20.50 11.60 12.50 7.40 3.90

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 117
3.4 Results of ANOVA Analyses and Duncan’s Test on Different forms of Bioturbation
3.4.1 Uca lactea annulipes
3.4.1.1 Anova Analyses
3.4.1.1. A. Seasons Wise
ANOVA analyses showed that main effect of season was found to be significant at 1 % level of significance due to burrow diameter, height of semidome, weight of excavated mud, distance and frequency of placement of excavated mud, and significant at 5 % level of significance on burrow depth (Table 10).
3.4.1.1. B. Tidal Levels Wise
ANOVA analyses displayed that main effect of tidal level was found significant at 1 % level of significance due to burrow depth, burrow diameter, height of semidome, and weight of excavated mud, distance and frequency of placement of excavated mud (Table 10).
3.4.1.1. C. Seasons and Tidal Levels Wise
ANOVA analyses revealed that main effect of season and tidal level exhibited significant at 1 % level of significance due to height of semidome, weight of excavated mud, distance and frequency of placement of excavated mud respectively (Table 10).
3.4.1.2. Results on Duncan’s Test
3.4.1.2. A. Seasonal Effect of Different Characteristics of Bioturabation at Different Seasons
The main effects of seasons when compared by Duncan’s test at 5% level of significance, it was found that the mean of burrow depth displayed significantly higher during pre-monsoon than monsoon and post monsoon; burrow diameter exhibited significantly higher results during monsoon than pre monsoon and post monsoon; height of semidome demonstrated significantly higher values during monsoon than pre monsoon and post monsoon; weight of excavated matters revealed significantly higher results during monsoon than post monsoon than pre monsoon; distance of placement of excavated mud demonstrated significantly higher results during monsoon than post monsoon than pre monsoon and frequency of placement of excavated matters showed higher values during monsoon than pre monsoon than post monsoon, (Table 11) respectively.
3.4.1.2. B. Seasonal Effect of Different Characteristics of Bioturabation at Different Tidal Levels
The main effects of tidal levels when compared by Duncan’s test at 5% level of significance, it was resulted that the mean of burrow depth showed significantly higher values at HTL than LTL and MTL; burrow diameter obtained significantly higher results at LTL than MTL than HTL; height of semidome displayed significantly higher values at LTL than MTL than HTL; weight of excavated mud exposed significantly higher findings at MTL than LTL and HTL; distance of placement and frequency of placement of excavated mud revealed significantly higher values at LTL than MTL than HTL (Table 11) respectively.

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 118
Table 10. The result of ANOVA analysis of different characteristics of bioturabation of U. lactea annulipes
Source Variable SS df MS F Sig. SEASONS Burrow Depth 1712.37 2 856.18 7.34 0.013* Burrow Diameter 3.84 2 1.92 35.10 0.00** Height of semidome 33.54 2 16.77 406.42 00.00** Weight of Excavated mud 33621.50 2 16810.75 8865.70 00.00** Distance of placement of excavated mudball 143.61 2 71.80 86.37 00.00** Frequency of mudball placement 581.37 2 290.68 284.91 00.00** TIDAL LEVELS Burrow Depth 2391.68 2 1195.84 10.25 0.005** Burrow Diameter 2.10 2 1.05 19.20 0.001** Height of semidome 3.00 2 1.50 36.29 00.00** Weight of Excavated mud 1323.20 2 661.60 348.92 00.00** Distance of placement of excavated mudball 59.34 2 29.67 35.69 00.00** Frequency of mudball placement 1052.96 2 526.48 516.03 00.00** SEASONS * TIDAL LEVELS Burrow Depth 241.93 4 60.48 0.52 0.725NS Burrow Diameter 0.55 4 0.14 2.50 0.117NS Height of semidome 2.18 4 0.55 13.22 0.001** Weight of Excavated mud 797.30 4 199.33 105.12 00.00** Distance of placement of excavated mudball 69.86 4 17.46 21.01 00.00** Frequency of mudball placement 217.75 4 54.44 53.36 00.00** Error Burrow Depth 1049.84 9 116.65 Burrow Diameter 0.49 9 0.05 Height of semidome 0.37 9 0.04 Weight of Excavated mud 17.07 9 1.90 Distance of placement of excavated mudball 7.48 9 0.83 Frequency of mudball placement 9.18 9 1.02 Total Burrow Depth 5395.81 17 Burrow Diameter 6.98 17 Height of semidome 39.09 17 Weight of Excavated mud 35759.07 17 Distance of placement of excavated mud 280.29 17 Frequency of mudball placement 1861.25 17
*Significant of 0.05 level ** Significant of 0.01 level NS- Not Significant
Table 11. Duncan's test of different characteristics of bioturabation of U. lactea annulipes between seasons and tidal levels
Seasons Tidal Levels
Burrow Depth
Burrow Diameter
Height of Semidome
Weight of Excavated mud
Distance of placement of excavated mud
Frequency of mudball placement
Pre monsoon HTL 89.00 1.000 0.420 71.250 5.760 8.030 LTL 52.78 1.950 0.605 71.185 8.900 21.765 MTL 64.13 1.550 0.490 64.110 6.145 12.620 Total 68.63a 1.50b 0.51b 68.85c 6.94c 14.14b Monsoon HTL 61.250 1.700 2.570 147.790 7.625 10.390 LTL 37.975 2.975 4.575 184.615 17.660 39.225 MTL 55.875 2.300 3.050 167.410 16.150 18.545 Total 51.70b 2.33a 3.40a 166.61a 13.81a 22.72a Post monsoon HTL 60.000 1.050 0.000 68.495 9.885 1.190 LTL 35.125 1.325 0.795 94.645 9.890 14.495

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 119
MTL 41.588 1.350 0.705 84.480 9.365 11.125 Total 45.57b 1.24b 0.50b 82.54b 9.71b 8.94c Total HTL 70.08x 1.25z 1.00z 95.85z 7.76z 6.54z LTL 41.96y 2.08x 1.99x 116.82x 12.15x 25.16x MTL 53.86y 1.73y 1.42y 105.33y 10.55y 14.10y
3.4.2 Uca triangularis bengali
3.4.2.1 Anova Analyses
3.4.2.1. A. Seasons Wise
ANOVA analyses established that main effect of season was found significant at 1 % level of significance due to burrow diameter, mudball diameter, weight of excavated mud, distance and frequency of placement of excavated mud, and significant at 5% level of significance on burrow depth (Table 12).
3.4.2.1. B. Tidal Levels Wise
ANOVA analyses displayed that main effect of tidal level was found significant at 1 % level of significance due to mudball diameter, distance and frequency of placement of excavated mud, and only burrow diameter revealed significant at 5 % level of significance (Table 12).
3.4.2.1. C. Seasons And Tidal Levels Wise
ANOVA analyses revealed that main effect of season and tidal level was found significant at 1 % level of significance on mudball diameter and frequency of placement of mudball (Table 12).
3.4.2.2. Results on Duncan’s Test
3.4.2.2. A. Seasonal effect of different characteristics of bioturabation at different seasons
The main effects of seasons when compared by Duncan’s test at 5% level of significance, it was established that the mean of burrow depth displayed significantly higher effects during pre monsoon than post monsoon; burrow diameter exhibited significantly higher impact during monsoon than pre monsoon and post monsoon; mudball diameter established significantly higher results during post monsoon than pre monsoon than monsoon; weight of excavated mud exhibited significantly higher role during monsoon than post monsoon and pre monsoon; distance of placement of excavated mud revealed significantly higher values during monsoon than post monsoon and frequency of placement of mudball showed higher results during monsoon than pre monsoon than post monsoon (Table 13) respectively.
3.4.2.2. B. Seasonal effect of different characteristics of bioturabation at different tidal levels
The main effects of tidal levels when compared by Duncan’s test at 5% level of significance, it was found that the mean of burrow depth showed significantly higher impacts at HTL than LTL; burrow diameter exhibited significantly higher results at LTL and MTL than HTL; mudball diameter displayed significantly higher values at MTL than LTL than HTL; weight of excavated mud showed significantly higher findings at LTL and MTL than HTL; distance of

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 120
placement of excavated mud showed significantly higher influence at LTL than MTL and HTL and frequency of placement mudball revealed significantly higher results at LTL than MTL than HTL (Table 13) respectively.
Table 12. The result of ANOVA analysis of different characteristics of bioturabation of U. triangularis bengali
Source Variable SS df MS F Sig. SEASONS Burrow Depth 1886.71 2 943.36 5.019 0.034* Burrow Diameter 3.45 2 1.72 9.856 0.005** Diameter of mudball 1.31 2 0.66 103.572 00.00** Weight of Excavated mud 13788.11 2 6894.05 25.258 00.00** Distance of placement of excavated mud 8.21 2 4.11 4.946 00.00** Frequency of mudball placement 571.66 2 285.83 124.070 00.00** TIDAL LEVELS Burrow Depth 1523.92 2 761.96 4.054 0.056NS Burrow Diameter 2.66 2 1.33 7.595 0.012* Diameter of mudball 0.55 2 0.27 43.225 00.00** Weight of Excavated mud 1658.01 2 829.01 3.037 0.098NS Distance of placement of excavated mud 26.10 2 13.05 15.727 0.001** Frequency of mudball placement 344.58 2 172.29 74.787 00.00** SEASONS * TIDAL LEVELS Burrow Depth 137.70 4 34.42 0.183 0.941NS Burrow Diameter 0.21 4 0.05 0.301 0.87NS Diameter of mudball 0.46 4 0.11 18.023 00.00** Weight of Excavated mud 2484.80 4 621.20 2.276 0.141NS Distance of placement of excavated mud 0.90 4 0.22 0.270 0.89NS Frequency of mudball placement 79.91 4 19.98 8.671 0.004** Error Burrow Depth 1691.60 9 187.96 Burrow Diameter 1.57 9 0.18 Diameter of mudball 0.06 9 0.01 Weight of Excavated mud 2456.54 9 272.95 Distance of placement of excavated mud 7.47 9 0.83 Frequency of mudball placement 20.73 9 2.30 Total Burrow Depth 5239.93 17 Burrow Diameter 7.89 17 Diameter of mudball 2.38 17 Weight of Excavated mud 20387.46 17 Distance of placement of excavated mud 42.68 17 Frequency of mudball placement 1016.88 17
Table 13. Duncan's test of different characteristics of bioturabation of U. triangularis bengali between seasons and tidal levels
Seasons Tidal Levels
Burrow Depth
Burrow Diameter
Diameter of mudball
Weight of Excavated mud
Distance of placement of excavated mud
Frequency of mudball placement
Pre monsoon HTL 86.588 1.250 0.680 74.700 5.600 6.725 LTL 54.525 2.175 0.940 92.335 9.065 15.580 MTL 69.855 1.500 0.820 81.925 6.630 10.825 Total 70.32a 1.64b 0.81b 82.99b 7.10ab 11.04b Monsoon HTL 66.988 1.750 0.220 103.330 6.715 12.945 LTL 48.675 2.700 0.415 151.730 9.195 30.675 MTL 58.350 2.550 0.380 151.540 6.945 21.895

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 121
Total 58.00ab 2.33a 0.34c 135.53a 7.62a 21.84a Post monsoon HTL 53.963 0.750 0.515 77.540 4.815 6.480 LTL 36.725 1.675 0.950 82.030 7.400 12.025 MTL 45.050 1.410 1.460 56.920 5.780 8.465 Total 45.258b 1.28b 0.98a 72.16b 6.00b 8.99c Total HTL 69.18x 1.25y 0.47z 85.19y 5.71y 8.72z LTL 46.64y 2.18x 0.77y 108.70x 8.55x 19.43x MTL 57.75xy 1.82x 0.89x 96.80x 6.45y 13.73y
4. Discussion
The two species of fiddler crabs under present study display adaptation vis-a-vis distribution in different tidal levels of an intertidal belt(Chakraborty& Chowdhury,1992) .U. lactea annulipes lives in large populations on low lying sandy-muddy loamy sediments along the edges of streams which are always covered by even in small amplitude of high tides. Nutrient-rich filamentous algae, detritus and bacteria are deposited by the receding tide each day, giving this species an ample supply of fresh food. U. triangularis bengali inhabits in small patches on higher areas that are not covered by all high tides; their habitat can remain uncovered by tides for several consecutive days in each 14 days semi-lunar tidal cycle. The species therefore has temporally limited access to fresh food which may affect growth rates. The behavioural differences between the two species appear to be strongly related to their different habitats, as found in other fiddler crab species (Backwell et al., 2006). Although the active burrowing activities were observed for both the species, it was found to be less in the habitat for U. triangularis bengali where the sediment was hard. As in other fiddler crab species, these high burrows did not get damaged by incoming tides and remained intact for several days (Klassen & Ens, 1993; Backwell et al., 2006). Furthermore, in the both sexes, the feeding rate of U. lactea annulipes was faster than U. triangularis bengali. Many other differences in between these two species were detected in respect of sizes,biomass and density.U. lactea annulipes was slightly bigger than U. triangularis bengali but they used to live at comparable densities. The sex-ratio of U. lactea annulipes was slight male biased, and the sex ratio is (M: F: 1:0.75.) In U. triangularis bengali, the bias was less pronounced, and the sex ratio is (M: F: 1:0.88.). Month has significant effect on the allocation of different sexes (χ2-test, P<0.01, 99%) of both species. Reproductive intensity in brachyuran can be measured by quantifying the relative frequency of ovigerous females. According to Sastry (1983), the beginning and the duration of the reproductive period are dependent on the occurrence of favorable environmental conditions. The major controlling factors appear to be latitude, temperature, larval food availability, and intertidal zonation (Sastry, 1983). The breeding period becomes protected from cool to warm subtropical localities (Emmerson, 1994). Since most studies have been conducted in temperate region, seasonal breeding is well reported in several the literatures (Thurman II, 1985 for U. subcylindrica in U.S.A./Mexico; Murai et al., 1987 for U. lactea in Japan; Spivak et al., 1991 for U. uruguayensis in Argentina). Yet, the majority of tropical crabs breed continuously, i.e., throughout the year, or have prolonged breeding seasons compared to species at higher latitudes (Sastry, 1983). The reproduction in U. triangularis bengali was remarkably post monsoonal, with a pronounced activity from October to January, and a pause period from

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 122
March to September, when the ovigerous females were not found. Beyond this, the more pronounced reproductive intensity for U. lactea annulipes occurred in the pre monsoon and post monsoon season. For U. lactea annulipes, ovigerous females were registered all the year round, with exception during June, July, August and September. For many fiddler crab species there is a close association between mating and incubation place and fecundity. Salmon (1987) suggested when mating was underground, the female showed high fecundity (like U. pugilator), but when it happened at the surface, a low egg number was produced (like with U. vocans). According to Thurman II (1985), the fecundity (size and number of eggs) for species of the genus Uca from tropical and temperate areas could vary according to environmental conditions. Nevertheless, fiddler crabs, which lived very near the rivers exhibited larger size and produced higher number of eggs; while semi terrestrial or totally terrestrial species have small size and resulted a low number of progenies. Fecundity differences were observed as much between both species as between individuals of the species. These differences were related to many factors: the weather during which females were collected; food availability; multiple spawning (1ª, 2ª or 3ª spawning of reproductive cycle); individual variation on egg production and natural egg losses related to female activity on the surface (Costa and Negreiros-Fransozo, 1996). The ability of U. lactea annulipes producing a higher number of eggs when compared with U. triangularis bengali did not confirm with the larval production of the former species was higher. The probability of mechanical losses, related to feeding activity on the surface was higher U. lactea annulipes than U. triangularis bengali. These points could be clarified only after undertaking study on fertility potential of both species in relation to their abilty of eggs production. There was also a difference between the species in handedness. In U. lactea annulipes, there were equal numbers of left and right handed males, while in U. triangularis bengali was a significant skew to right handed males. While it is generally thought that female fiddler crabs feed faster than males because they have two feeding claws, it is not always true. Of the four studied species, Weis and Weis (2004) found this to hold in only two species. It was observed in the present study that in both U. lactea annulipes and U. triangularis bengali females fed faster than males. There was a very obvious difference in the structure building behaviour of these two species. U. lactea annulipes arranged mud at some distance from the burrow and built semidomes. Many Uca species build structures such as pillars or hoods from moist sand or mud (Shepard, 1954), some of which play roles in attracting females (Shepard, 1954) or defining territory (Zucker, 1981; Weis & Weis, 2004). Mud balls may be merely a by-product of burrow excavation (Klassen & Ens, 1993, but may also function as a signal (Zeil et al., 2006).
Some of the species in soft sediments are responsible for the structure and functioning of their habitats. These organisms, known as ‘bioturbators’, constantly disturb the sediment by burrowing and feeding. Their activities mix the sediment layers and cause substantial resuspension of the sediment and its transport by waves and current. At the same time, bioturbation enhances organic decomposition, nutrient cycling, redistribution of organic materials and oxygenation of sediments, rather as earthworms do on land. These changes greatly affect estuarine and seafloor habitats, with repercussions for the entire soft-sediment ecosystem (Chatterjee et al., 2008). Burford et al., (2001)has indicated that for U. tangeri males, mudballs mark territory and are attractive to females. While we do not know the significance of

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 123
our observations of crabs arranging mud balls and placing them at a distance from burrows and has not tested the functions of semi-domes, mud ball, it is clear that these structures play some role in the social as well as sexual lives of the crabs involved. Future work will illuminate their various roles. The possible function of semidome building behavior is whether it reduces aggression or attracting females. Mudball formed by the male of U. triangularis bengali seems to be functioning as territory marking or female attraction. It seems that above mentioned different unique structures viz. Semidome and mudball of two species have diverse functional roles, but more intensive study is required to understand their functional as well as ecological significance.
Overall, the present study tried to provide a preliminary report on the habitat, sex ratio, fecundity and some unique features of bioturbatory activities of two species of fiddler crab. There were some similarities and some differences in the ecobiology of the two species of fiddler crabs studied here. The similarities were largely due to the strong effect of seasonal cycles on the bioturbation of the crabs. The differences seem to be related, in part, to the slightly different microhabitats used by the two species. The construction of semidome and mudball that probably play some roles in the social as well sexual lives of both U. lactea annulipes and U. triangularis bengali.
Acknowledgements
Authors are thankful to the authority of Vidyasagar University for providing library, laboratory and internet facilities. Research was funded by the Ministry of Environment and Forest, Government of India [Sanctioned No. 3/6/2001 – CSC (M) dated 5th Nov.2001].
References
Backwell, P. R., & Christy, J. H. (2000). Dishonest signalling in a fiddler crab. Proceedings of the Royal Society of London, 267, 719-724. http://dx.doi.org/10.1098/rspb.2000.1062
Backwell, P. R., Jennions M., Wada K., Murai, M., & Christy, J. (2006). Synchronous waving in two species of fiddler crabs. Acta Ethologica, 9, 22-25. http://dx.doi.org/10.1007/s10211-005-0009-8
Burford, F. R. L., McGregor, P. K., & Oliveira, R. F. (2001). Mudballing revisited: further investigations into the construction behaviour of male Uca tangeri. Behaviour, 138, 221-234. http://dx.doi.org/10.1163/15685390151074393
Chakraborty, S. K., & Chaudhury, A. (1992). Ecological studies on the zonation of brachyuran crabs in a virgin mangrove island of Sundarbans, India. Journal of Marine Biological Association of India, 34, 189-194.
Chakraborty, S. K., Chatterjee, S., Dey, M. K., Bhakat, R. K., & Paul, A. K. (2012). Ecological Changes of an Eco-rehabilitated Delta of Hooghly Estuary. Journal of Coastal Environment, 3(1), 9-28.
Chatterjee, S., & Chakraborty, S. K. (2014). Feeding Behaviour and Functional Role of Some Selected Species of Brachyuran Crabs in Nutrient Cycle at Coastal Belt of Midnapore (East),

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 124
West Bengal, India. Journal of Biology and Life Science, 5(1), 106-129. http://dx.doi.org/10.5296/jbls.v5i1.4869
Chatterjee, S., Bhunia, G., & Chakraborty, S. K. (2008). Bioturbation of Brachyuran crabs and its impact on coastal ecosystem of Midnapore district, West Bengal, India. In proceedings of Forth International Conference on Environmental Science and Technology, at Houston, Texas U.S.A. Pp: 133-148.
Christy, J. H., Backwell, P. R. Y., & Goshima, S. (2001). The design and production of a sexual signal: hoods and hood building by male fiddler crabs Uca musica. Behaviour, 138, 1065-1083. http://dx.doi.org/10.1163/156853901753287145
Costa, T. M., & Negreiros-Fransozo, M. L. (1996). Fecundidade de Callinectes danae Smith, 1869 (Crustacea, Decapoda, Portunidae) na região de Ubatuba (SP), Brasilian Archive of Biology and Technology, 39(2), 393-400.
Costa, T. M., Silva, S. M. J., & Negreiros-Fransozo, M. L. (2006). Reproductive pattern comparison of Uca thayeri Rathbun, 1900 and U. uruguayensis Nobili, 1901 (Crustacea, Decapoda, Ocypodidae). Brazilian Archives of Biology and Technology, 49(1), 117-123. http://dx.doi.org/10.1590/S1516-89132006000100014
D’Andrea, A. F., & DeWitt, T. H. (2009). Geochemical ecosystem engineering by the mud shrimp Upogebia pugettensis (Crustacea: Thalassinidae) in Yaquina Bay, Oregon: density-dependent effects on organic matter remineralization and nutrient cycling. Limnology and Oceanography, 54, 1911-1932. http://dx.doi.org/10.4319/lo.2009.54.6.1911
Daiber, F. C. (1952). Animals of the tidal marsh. Van Nostrand Reinhold, New York.pp. 422.
Emmerson W. D. (1994). Seasonal breeding cycles and sex-ratios of 8 species of crabs from Mgazana, a mangrove estuary in Transkei, southern Africa. Journal of Crustacean Biology, 14(3), 568-578. http://dx.doi.org/10.2307/1549002
Guillermo Guerao & Guiomar Rotllant, (2009). Post-larval development and sexual dimorphism of the spider crab Maja brachydactyla (Brachyura: Majidae). Scientia Marina. http://dx.doi.org/10.3989/scimar.2009.73n4797
Hemmi, J. M., & Zeil, J. (2003). Burrow surveillance in fiddler crabs II. The sensory cues. Journal of Experimental Biology, 206(22), 3951-3961. http://dx.doi.org/10.1242/jeb.00636
How, M. J., Hemmi, J. M., Zeil, J., & Peters, R. (2008). Claw waving display changes with receiver distance in fiddler crabs, Uca perplexa. Animal Behaviour, 75(3), 1015-1022. http://dx.doi.org/10.1016/j.anbehav.2007.09.004
Johnson, P. T. J. (2003). Biased sex ratios in fiddler crabs (Brachyura, Ocypodidae): a review and evaluation of the influence of sampling method, size class and sex-specific mortality. Crustaceana, 76(5), 559-580. http://dx.doi.org/10.1163/156854003322316209
Klassen M., & Ens, B. J. (1993). Habitat selection and energetics of the fiddler crab, Uca

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 125
tangeri. Netherlands Journal of Sea Research, 31, 495-502. http://dx.doi.org/10.1016/0077-7579(93)90061-V
Koch, V., & Wolff, M. (2002). Energy budget and ecological role of mangroveepibenthos in the Caeté estuary, North Brazil. Marine Ecology Progress Series, 228, 119-130. http://dx.doi.org/10.3354/meps228119
Lim, S. L. S. (2006). Fiddler crab burrow morphology: How do burrow dimensions and bioturbative activities compare in sympatric populations of Uca vocans (Linnaeus, 1758) and U. annulipes (H. Milne Edwards, 1837)? Crustaceana, 79(5), 525-540. http://dx.doi.org/10.1163/156854006777584241
Litulo, C. (2004). Reproductive aspects of a tropical populationof the fiddler crab Uca annulipes (H. Milne Edwards, 1837) (Brachyura: Ocypodidae) at Costa do Solmangrove, Maputo Bay, southern Mozambique. Hydrobiologia, 525, 167-175. http://dx.doi.org/10.1023/B:HYDR.0000038864.38435.6a
Macia A., Quincardete, I., & Paula, J. (2001). A comparison of alternative methods for estimating population density of the fiddler crab Uca annulipes at Saco Mangrove, Inhaca Island (Mozambique). Hydrobiologia, 449, 213-219. http://dx.doi.org/10.1023/A:1017502817087
May, R. M. (1983). When to be which sex. Nature London, 301, 16-17. http://dx.doi.org/10.1038/301016a0
Mermillod-Blondin, F. (2011). The functional significance of bioturbation and biodeposition on biogeochemical processes at the water - sediment interfacein freshwater and marine ecosystems. Journal of North American Benthological Society, 30, 770-778. http://dx.doi.org/10.1899/10-121.1
Meysman, F. J. R., Middelburg, J. J., & Heip, C. H. R. (2006). Bioturbation: a fresh look at Darwin’s last idea. Trends in Ecology and Evolution, 21, 688-695. http://dx.doi.org/10.1016/j.tree.2006.08.002
Mokhtari, M., Savari A., Rezai H., Kochanian P., & Bitaab, A. (2007). Population ecology of fiddler crab, Uca annulipes (Decapoda: Ocypodidae) in Sirik mangrove estuary, Iran. Estuarine, Coastal and Shelf Science, 76, 273-281. http://dx.doi.org/10.1016/j.ecss.2007.07.010
Montague, C. L. (1980). A natural history of temperate western Atlantic fiddler crabs (Genus Uca) with reference to their impact on the salt marsh. Contribution in Marine Science, 23, 25-55.
Murai, M., Goshima, S., & Henmi, Y. (1987). Analysis of the mating system of the fiddler crab, Uca lactea. Animal Behaviour, 35, 1334-1342. http://dx.doi.org/10.1016/S0003-3472(87)80005-2
Pope, D. S. (2005).Waving in a crowd: fiddler crabs signal in networks. In: McGregor PK, ed. Animal communication networks. Cambridge: Cambridge University Press, 252-276.

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 126
http://dx.doi.org/10.1017/CBO9780511610363.016
Reid, D. M., & Corey, S. (1991). Comparative fecundity of decapod crustaceans, II. The fecundity of fifteen species of anomuran and brachyuran crabs. Crustaceana, 61(2), 175-189. http://dx.doi.org/10.1163/156854091X00669
Rosenberg, M. S. (2002). Fiddler crab claw shape variation: a geometric morphometric analysis across the genus Uca. Biological Journal of Linnean Society, 75, 147-162.
Salmon, M. (1987). On the reproductive behavior of the fiddler crab Uca thayeri, with comparisons to U. pugilator and U. vocans: evidence for behavioral convergence. Journal of Crustacean Biology, 7(1), 25-44. http://dx.doi.org/10.2307/1548623
Sastry, A. N. (1983). Ecological aspects of reproduction. In The biology of Crustacea. Environmental adaptations, vol. 8 (ed. F.J. Vernberg and G.W. Vernberg). pp. 179-270. New York: Academic Press.
Shepard, F. P. (1954). Nomenclature based on sand-silt-clay ratios. Journal of Sedimentary Petrology, 24, 151-158.
Spivak, E. D., Gavio, M. A., & Navarro, C. E. (1991). Life history and structure of the word's southernmost Uca population: Uca uruguayensis (Crustacea, Brachyura) in Mar Chiquita lagoon (Argentina). Bulletin of Marine Science, 48(3), 679-688.
Statsoft, (2001). STATISTICA: data analysis software system, version 6.0. Tulsa, StatSoft, Inc.
Sweat, L. H. (2009). "Pachygrapsus transversus". Smithsonian Institution. http://www.sms.si.edu/irLspec /Pachyg transv.htm. Retrieved January 20, 2010.
Thurman II, C. L. (1985). Reproductive biology and population structure of the fiddler crab Uca subcylindrica (Stimpson). Biological Bulletin, 169, 215-229. http://dx.doi.org/10.2307/1541399
Weis J., & Weis P. (2004). Behavior of four species of fiddler crabs, genus Uca, in southeast Sulawesi, Indonesia. Hydrobiologia, 523, 47-58. http://dx.doi.org/10.1023/B:HYDR.0000033093.84155.1d
Wilkinson, M. T., Richards P. J., & Humphreys, G. S. (2009). Breaking ground: pedology, geological, and ecological implications of soil bioturbation. Earth Science Review, 97, 257-272. http://dx.doi.org/10.1016/j.earscirev.2009.09.005
Zar, J. H. (2009). Biostatistical Analysis. 5th ed. Pearson International Edition, Upper Saddle River.
Zeil J., Hemmi, J. M., & Backwell, P. R. (2006). Fiddler crabs. Current Biology, 16, 40-41. http://dx.doi.org/10.1016/j.cub.2006.01.012
Zeil, J., & Layne, J. E. (2002). Path integration in fiddler crabs and its relation to habitat and social life. In: Wiese K (ed.) Crustacean experimental systems in neurobiology. Springer, Berlin, Heidelberg, New York, pp. 227-24. http://dx.doi.org/10.1007/978-3-642-56092-7_13

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 127
Zucker, N. (1981.) The role of hood-building in defining territoriesand limiting combat in fiddler crabs. Animal Behaviour, 29, 387-395. http://dx.doi.org/10.1016/S0003-3472(81)80098-X
Copyright Disclaimer
Copyright for this article is retained by the author(s), with first publication rights granted to the journal.
This is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 128
Insects Associated With Tea and Their Identification at Ntri (Shinkiari) Mansehra, Pakistan
Hamid Ullah (Corresponding author)
M. Phil, Department of Animal sciences, Quiad-i-Azam University, Islamabad,Pakistan
Tel: 92-034-6906-1403 E-mail: [email protected]
Ibrar Muhammad
M. Phil, Department of Animal sciences, Quaid-i-Azam University, Islamabad, Pakistan
Tel: 923078404061 E-mail: [email protected]
Waheed Ullah
M. Phil, Department of Animal sciences, Quaid-i-Azam University, Islamabad, Pakistan
E-mail: [email protected]
Dr. Farzana Perveen (Supervisor)
Assistant Professor, Department of Zoology, SBBU, Sheringal KP, Pakistan
Tel: 92-944-885-529 E-mail: [email protected]
Sohail Aslam
M.Phil Zoology. Scientific Officer in National Tea research institute, Manshera, Pakistan
Tel: 92-333-784-0668 E-mail: [email protected]
Received: March 5, 2014 Accepted: March 20, 2014
doi:10.5296/jbls.v5i2.5233 URL: http://dx.doi.org/10.5296/jbls.v5i2.5233
Abstract
Tea plants are visited by different types of insects including tea pests and pollinator. The objective of the study was to identify tea associated insects in National Tea Research Institute (NTRI), Mansehra, Pakistan during April-June 2012.
A collection of tea associated insects was made during April-June 2012 in Tea Experimental Garden (TEG), NTRI, Mansehra, Pakistan. A total of 128 samples were collected from TEG

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 129
randomly.
During the present research, 10 species were identified. Species collected were brush-footed butterfly, Argynnis hyperbius (Linnaeus, 1763); painted lady, Vanessa cardui (Linnaeus, 1758), Lucerne butterfly, Colias electo (Linnaeus, 1763)( Nymphalidae); small cabbage white butterfly, Pieris rapae (Linnaeus, 1758), white cabbage butterfly, Pieris brassicae (Linnaeus, 1758)( Pieridae); Asiatic honey bee, Apis cerana Fabricius, 1793, Western honey bee, Apis mellifera Linnaeus, 1758(Apidae); seven-spot ladybird Coccinella septempunctata (Linnaeus, 1758)( Coccinellidae); blue marsh hawk Orthetrum glaucum (Brauer, 1865), Crimson-tailed marsh hawk, Orthetrum pruinosum neglectum (Rambur, 1842)( Libellulidae). Different numbers of individuals were collected of each species. Seventeen individuals of P. brassicae, 16 of C. septempunctata, 15 of P. rapae, 14 of C. electo, 13 of V. cardui and A. cerana, 10 of A. mellifera, 9 of O. pruinosum neglectum and 7 of O. glaucum.
It is concluded that there is a diversity to explain tea associated insects in NTRI, Mansehra, Pakistan.
Keywords: Apidae, Coccinellidae, Libellulidae, Mites, Nymphalidae, Pests, Pieridae
1. Introduction
History of tea growing countries shows that after plantation of tea crop within 8-10 yeas some pest/insects come up due to which crop yield affect badly. A number of approaches have been adopted by entomologists to gain knowledge of natural enemies of insects (Takagi, 1973).
The tea plant is attacked by several injurious insects, as the faggot-worm, Enmete Carmerii Linnaeus, 1844 one of the bagworms, whose larva carries a case made up of fragments of twigs and feeds upon leaves of the plant. The tea borer, Zeuzere coffete Nietner, 1861 is the larva of a cossid moth; it bores into the stems of both coffee and tea plants. Three species of bark-slice, Aspidiotus thew Linnaeus 1841; Aspidiotus flaccscens Nietner, 1863 and Aspidiotus transparens Linnaeus, 1832 also occur commonly upon tea, and the first named is one of the most serious enemies of the growing plant, very noticeable at the time of pruning. Three mites feed upon the leaves. One of them a red spider, Tetranychus bimaculatus Harvey, 1891 another five-legged tea mite, Typhlodromus eariatlas Harvey, 1895 closely related to the rust-mite of the orange. It feeds upon the lower leaves. The yellow tea mite, Acarus transluccns Nienter, 1859 feeds upon the buds and produces the condition called 'sulky'.In the world there are, 1031 arthropod species associated with tea; a small number of pests (about 3%) are common throughout the world. As a result of autochthonous and heterochthonous recruitment and the influence of climate, altitude, nature of cultivation, and age of plantation, however, each geographic region may have its own distinctive pest complex (Chen and Chen, 1989).
The flowers are pollinated by insects. Tea is virtually self-sterile and almost entirely cross-pollinated. Supplementary pollination produced more, larger, and heavier capsules, better viability, and a higher grade of seed. Bees are the chief pollinating agents of tea (Bakhtadze, 1932). Syrphid flies, such as hoverflies and drone flies, are important non-selective pollinators. Various midges and thripes are comparatively minor opportunist pollinators. Ants also pollinate some kinds of flowers, but for the most part they are parasites.

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 130
Some Diptera (flies) may be the main pollinators in higher elevations of mountains. Other insect orders are rarely pollinators (e.g.,Hemiptera such as Anthocoridae, Miridae) Jim et al., 2002. Keeping in view the importance of tea associated insects, the present studies were conducted to enlist these insects in National Tea Research Institute (NTRI), Mansehra, Pakistan.
1.1 Study Area
The insects were collected from all Experimental Tea Gardens (ETG) associated with NTRI, District Mansehra of Pakistan.
Mansehra is the North Eastern District of Khyber Pukhtunkhwa and at altitude of 975.36 meters (3200 feet). It is located between 34°–14’ and 35°–11’ north latitudes and 72°–49’ and 74°–08’ east longitude. The Nanga Parbat Mountain is located about 40 kilometers from the north eastern boundary of the district. Climate of the district is warm in summer and cold in winter. Temperature ranges from 2 °C-36 °C in the district (Ali, 2005). Our Objectives were: 1) enlist tea associate insects in NTRI, at district Mansehra, 2) Create awareness regarding tea associated insects and their importance.
2. Material and Methods
The study was conducted during April-June, 2012 in Experimental Tea Garden (ETG), National Tea Research Institute (NTRI), Mansehra, Khyber Pakhtunkhwa, Pakistan and the following strategy were adopted for exploring species diversity and elaborating distribution of the tea associated insects at the study area.
2.1 Collection and Preservation
Insects associated with tea were collected from ETG, NTRI randomly by using aerial nets and naked hands, after killing them in chloroform bottle collected specimens were first placed on white thermo-foil by stretching their wings and antennae and a white paper was placed on their wings of each specimen to keep them smooth. These were pinned properly and leaved for few days in such condition to keep them dry and fixed. On drying these were properly labeled and mounted in the collection boxes. Naphthalene balls were placed in the boxes to keep them safe from the pests.
2.2 Identification and Description
For identification, the specimens were examined under stereoscope. Identification was done up to the specific level with the help of available literature. Help was also taken by already identified specimens placed in National Insects Museum NIM, National Agriculture Research Center NARC, Islamabad and NTRI, Mansehra. Valid names along with synonyms, habitat and distribution were given for the species already recorded from Pakistan. All the identified specimens were deposited in the Zoological Museum, Department of Zoology, Hazara University, Mansehra, Pakistan.
2.3 Morphometry and Photography
Identified specimens were subjected for measurement of their total body length and wingspan

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 131
by using Digital Fractional Caliper (No. 147 UPC 01470, Toronto, Canada). Different number of specimens of each 10 identified species was measured. The specimens were placed one by one on top of a blank white sheet of computer paper for taking their photographs by using digital camera (14.1 mega pixel lenses, Sony Cyber-Shot DSC-W610, Tokyo, Japan). The photography was done of preserved specimens.
2.4 Data Analysis
The data were analyzed by MS Excel by using mean and standard deviation.
3. Results
During the present research, total of 128 samples were collected from Tea Experimental Garden (TEG), randomly and 10 species of 4 orders, Lepidoptera, Hymenoptera, Coleoptera and Odonata were identified. Different number of individuals was collected of each species. Seventeen individuals of P. brassicae, 16 of C. septempunctata, 15 of P. rapae, 14 of C. electo, 13 of V. cardui and A. cerana, 10 of A. mellifera, 9 of O. pruinosum neglectum and 7 of O. glaucum (Table 1).
Table 1. Tea associated insects identified during April-June 2012 in National Tea Research Institute (NTRI), Mansehra, Pakistan.
Order Family Species Number of individuals Lepidoptera Nymphalidae Argynnis hyperbius 14 Vanessa cardui 13 Pieridae Pieris brassicae 17 Pieris rapae 15 Colias electo 14 Hymenoptera Apidae Apis cerana 13 Apis mellifera 10 Coleoptera Coccinellidae Coccinella septempunctata 16 Odonata Libellulidae Orthetrum glaucum 07 Orthetrum pruinosum neglectum 09 Total* 128
*Total number of individuals of species
Table 2. Morphometry of Body length and wing span of different Tea associated insects species collected during April-June 2012 in National Tea Research Institute (NTRI), Mansehra, Pakistan.
SNo Comman name Name of species N* Body Length (M±SD; mm)*
Wing span (M±SD)
01 Brush-footed butterfly Argynnis hyperbius 11 20.64±0.48 75.41±0.59
02 Painted lady Vanessa cardui 09 17.79±0.35 59.85±0.23 03 White cabbage butterfly Pieris brassicae 13 19.57±0.39 69.45±0.38 04 Small cabbage white butterfly Pieris rapae 10 18.50±0.17 58.68±0.29 05 Lucerne butterfly Colias electo 09 15.57±0.35 44.43±0.36 06 Asiatic honey bee Apis cerana 08 11.95±0.22 18.33±0.22 07 Western honey bee Apis mellifera 07 11.69±0.31 18.37±0.24 08 Seven-spot ladybird Coccinella septempunctata 12 8.99±0.17 11.41±0.23 09 Blue marsh hawk Orthetrum glaucum 07 32.69±0.15 52.66±0.17

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 132
10 Crimson-tailed marsh hawk Orthetrum pruinosum neglectum
08 42.27±0.26 70.43±0.22
*Values are represented as mean ± standard deviation. Mean with in a column followed by standard deviation, N*= number of species examined for morphometry.
During the present research, species of four order were identified in which the order Lepidoptera consist of five species greater than the other orders, then order hymenoptera consist of two species, order Odonata consist of two species and the order Coleoptera consist of only one specie which is less than the other orders (Figure 1). All identified species were different population. The most populated species was P. brassicae and less populated is O. glaucum. Different species were different body length and wing span. Argynnis hyperbius have the largest wing span and Coccinella septempunctatathe smallest wing span, similarly Orthetrum pruinosum neglectum have the greatest body length and C. septempunctata have the smallest body length.
Figure 1. Population of insects associated with different insects orders were identified on tea, Camellia sinensis Linneaus, 1753 during April-June 2012 in National Tea Research Institute
(NTRI), Mansehra, Pakistan.
4. Discussion
The present research was conducted in ETG, NTRI during which different butterfly fauna were documented. The study revealed that 5 species in 2 families and 4 genera occur in the area. The families include: Nymphalidae (generaArgynnisand Vanessa are represented); Pieridae (genera Pieris and Colias are represented). Abbas et al., 2002 were conducted a study in six different localities around Skardu to document the butterfly fauna of that region. The study revealed that 16 species in 5 families and 14 genera occur in the area. The families include: Papilionidae (represented only by the genus Parnassius); Pieridae (genera Pieris, Pontia and Colias are represented); Lycaenidae (genera Lycaena, Everes, Aricia, Plebejus, Zizeeria and Zizina are

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 133
represented); Nymphalidae (only two genera, Aglais and Vanessa are represented); and Satyridae (represented by the genera Pararge and Maniola). Genus Pieris, Colias in family Pieridae and Vanessa in Nymphalidae were recorded in both the researches. Most of the genera were not found in the present study. Genus Argynnis was not reported in the previous research but recorded during present study. Differences were found due to climate difference. The previous research study area is harsh than the present research study region.
Only species Coccinella septempunctata Linnaeus, 1758 in family Coccinellidae and order Coleoptera was reported during the present research. Khan et al., 2006 were carried out an extensive survey of predatory Coccinellid beetles (Coleoptera: Coccinellidae) in the Chitral District, Pakistan. Twelve different species belonging to 9 genera of 3 tribes and 2 sub-families were recorded. Two sub-families, Coccinellinae and Chilocorinae were identified. The following 8 species belonged to family Coccinellinae and tribe Coccinellini, Coccinella septempunctata Linnaeus, 1758; Hippodamia variegata Goeze, 1777; Calvia punctata Mulsant, 1853; Adalia bipunctata Linnaeus, 1758; Adalia tetraspilota Hope, 1873; Aiolocaria hexaspilota Hope, 1873; Macroilleis hauseri Mader, 1930 and Oenopia conglobata Linnaeus, 1758. Only one species namely, Halyzia tschitscherini Semenov, 1807 represented tribe Psylloborini of the sub-family Coccinellinae. Three species occurred from sub-family Chilocorinae and tribe Chilocorini, Chilocorus rubidus Hope, 1807; Chilocorus circumdatus Gyllenhal, 1808 and Priscibrumus uropygialis Mulsant, 1853. From the aforementioned species 6 were recorded for the first time from Pakistan, Chilocorus circumdatus Gyllenhyl, 1808; Calvia punctata Mulsant, 1853; A. bipunctata; M. hauseri; P. uropygialis, and Oenopia conglobata Linnaeus, 1758. Coccinella septempunctata was documented from both the research study area. A large number of beetle fauna was not recorded in the present study as documented by previous research. Different host plants and climate are responsible for such huge differences.
Hamid et al., 1992 were conducted a study on insects associated with tea under natural conditions at Experimental Tea Garden, (ETG) Daively and National Tea Research Station, (NTRS) Shinkiari. Six tea varieties i.e., Qi-Men, Jue-Keng, Chuye, Ruopi and Huang-Shun (Chinese type) and High Grown (Sri Lankan) were included in the experiment. The insects observed feeding on tea were surface grasshopper, Acrotylus humbertianus Sauss, 1732; Chrotogonus trachypterus trachypterus Blanchard, 1741; Long horned grasshopper, Tettigonia cantans Fuessly, 1755; short horned grasshopper, Attractomorpha acutipennis Guer, 1777; rice grasshopper, Truxalis grandis Klug, 1722; Acrida exaltata Wlk Jassid, 1733; Amrasca devastans Distant, 1876 and aphid, Toxoptera aurantii Boyer, 1691. There was no similarity between both the researches. The climate and host plant both are the same but still no such species were documented during present study as recorded by previous research. The reason is the use of insecticides and pesticides during the present research work.
5. Conclusions
The present study was conducted pertaining to the biodiversity of tea associated insects in ETG, NTRI, Mansehra, Pakistan. During the present study species of 4 orders were reported. On the whole order Lepidoptera was dominant followed by Hymenoptera, Odonata and Coleoptera. Ruby et al., 2010 conducted studies pertaining to biodiversity of arthropods in the cropland of

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 134
two zones i.e., mixed crop zone (Faisalabad) and Cotton-Wheat zone (Multan) Punjab, Pakistan for a period of one year. The main focus was to collect, identify and compare the species richness and evenness. Sugarcane, Fodder, Wheat and Brassica were sampled round the year showed variations in species composition of their fauna in the two districts representing the two zones. Mixed-crop zone was highly diversified with respect to species and abundance of individuals per species. On the whole order Orthoptera was dominant followed by Araneae, Hemiptera, Coleoptera, Lepidoptera, Hymenoptera, Odonata, Diptera and Thysanoptera, Neuroptera, Prostigmata each represented by single species except Mantodea with two species. All of the 4 orders which were recorded during the present research were also reported by the previous study. Some of the orders which were observed by the previous study were not reported during the present research. Dissimilarities in both the researches were due to the climate and host plant differences.
References
Abbas, M. Rafi, MA. Inayatullah, M. Khan, MR., & Pavulaan, H. (2002). Taxonomy and distribution of butterflies (papilionoidea) of the Skardu region, Pakistan. The Taxanomic Report of the International Lepidoptera Survey. 3(9), 189-196.
Ahmad, N. Hamid, F S. Khan, AR., & Waheed, A. (1992). Insects associated with tea at Mansehra (NWFP) Pakistan. Pakistan Journal of Agricultural Research. 13(4), 222-226.
Ali, I. (2005) Maping and documentation of cultural assests of Mansehra district, united National Scientific and Cultural Organization, Islamabad 1-91.
Bakhtadze, K. (1932). Pollination of tea, Camelia sinensis, in Georgia. Subtropics. 21(12), 63-80.
Chen, Z. Chen, X. (1989). An analysis of the world tea fauna. Journal of Tea Science. 9(15), 13-22.
Jim, G. Mant, F. Schiestl, P. Rod, PP., & Weston, H. (2002). A phylogenetic study of pollinator conservatism among sexually deceptive orchids. Evolution. 56(5), 888-898. http://dx.doi.org/10.1111/j.0014-3820.2002.tb01402.x
Khan, I. Din, S. Khalil, SK., & Rafi, MA. (2006). Survey of predatory Coccinellids (Coleoptera: Coccinellidae) in the Chitral District, Pakistan. Journal of Insect Science. 7(7), 169-174.
Ruby, T. Akhtar, S. Rana, AM., & Hameed, M. (2010). Biodiversity of Foliage Arthropods in the Mixed Crop Zone and Cotton-Wheat Zone in Punjab Province, Pakistan. International Journal of Agriculture and Biology. 12(6), 861–866.
Takagi, k. (1973). Monitoring of Hymenopterous parasite in tea field. National Research Institute of Tea, Japan. 10, 91-131.
Copyright Disclaimer
Copyright for this article is retained by the author(s), with first publication rights granted to the journal.
This is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 135
Root Endophytic Fungi: Research Update
Neha Chadha, Manjita Mishra, Ram Prasad
Amity Institute of Microbial Technology
Amity University of Uttar Pradesh, Sector 125, Noida- 201303, India
Ajit Varma (Corresponding author)
Amity Institute of Microbial Technology
Amity University of Uttar Pradesh, Sector 125, Noida- 201303, India
Tel: 91-981-089-9562 E-mail: [email protected]
Received: May 23, 2014 Accepted: June 5, 2014
doi:10.5296/jbls.v5i2.5960 URL: http://dx.doi.org/10.5296/jbls.v5i2.5960
Abstract
Fungal endophytes are group of microorganisms that reside asymptomatically within the healthy living tissue. Root endophytic fungi like dark septate endophytes (DSE), Piriformospora indica and Trichoderma show mutualistic association with many plant species. The endophytes are known to produce a diverse group of secondary metabolites, extracellular enzymes, antibiotics, antifungal, anticancer agents that can be used as therapeutic drugs, agrochemicals or in industries. Plants associated with fungal endophytes play an important role in the survival of plants in extreme stressed condition. This review focuses on the functions of fungal root endophytes, their relationship with host plant, tolerance to abiotic stress and secondary metabolites production against various pathogens.
Keywords: Fungal Endophytes, Plant Growth Promoting Fungus, Secondary Metabolites, Pirifrmospora indica, Dark Septate Endophyte, Arbuscular Mycorrhizal (AM)
1. Introduction
The term endophyte was coined by the German phytopathologist Heinrich Anton De Bary in 1884, and is used to define fungi or bacteria occurring inside plant tissues without showing disease in the host plant (Wilson 1995). In contrast to others like epiphytes and mycorrhiza, endophytes reside within leaves, bark, stems and roots of the plant whereas mycorrhiza only colonizes the roots of the host plant (Saikkonen et al. 1998). Root endophytes harbor an

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 136
important component of microbial community that is different from above ground plant tissue (Addy et al. 2005), but as common as mycorrhizal fungi that reside below the ground (Weishampel and Bedford 2006). They differ from mycorrhizal fungi in morphology, development and nutrient transfer (Brundrett 2004). Brundrett and Kendrick (1988) stated that endophytic fungi living in the old roots as Glomerocycete can service for more than years even after the collapse of arbuscules. Fungal endophytes are the microorganisms that reside inside the plant tissue that can be roots, leaves or stem. They basically fall in ascomycota and basidiomycota groups (Arnold and Lutzoni 2007; Selosse et al. 2009). Endophytic fungi are plant mutualists and show various benefits to the host plant, for example abiotic stress tolerance (Redman et al. 2002), resistance to pathogens and disease (Benhamou and Garand 2001) and the production of secondary metabolites (Schulz et al. 1995). There are more than one million species of endophytic fungi associated with plants worldwide that can provide a variety of secondary bioactive products such as alkaloid, benzopyranones, flavanoids, phenols, phytochemicals and anticancer agents (Aly et al. 2010; Kharwar et al. 2011; Kusari and Spiteller 2012b). Endophytic fungi promote plant growth through auxin (IAA) (Zou and Tan 1999), phosphate solubilization (Malla et al. 2004; Wakelin et al. 2004) and siderophore production (Costa and Loper 1994).
Fungal root endophytes involve diverse group of fungi. Some specific examples are Dark septate endophytes (DSE), Trichoderma, Piriformospora indica, that are known to be true root endophytes. This review focuses on the functions of fungal root endophytes, their relationship with host plant, and secondary metabolites production against various pathogens.
1.1 Dark Septate Endophyte
Dark septate endophyte (DSE) is a ubiquitous group of ascomycete’s fungi that may function as saprophytes or mutualistic. Kohn and Stasovski (1990) found that these fungi can survive in cold and stressed environment than AM fungi. DSE differ from other group of fungi because of their septate and melanised hyphae (Yu et al. 2001). DSE produces conidia with microsclerotia in plant roots. It has been found that these groups of fungi uses inorganic form of nitrogen that in turn benefits the host plant for nutrient uptake (Upson et al. 2009; Newsham 2011). They are frequently observed in cortex, epidermis as well on root surface. Sieve elements of roots of Atriplex canescens were predominately colonized with the hyaline hyphae and non-pigmented fungal structure (Barrow and Aaltonen 2001). They have been reported from many plants and habitat worldwide. Phialocephala fortinii (Wang and Wilcox 1985) has a broad host and geographical range (Ahlick and Sieber 1996) mostly dominated in Europe and western Canada. Phialocephala dimorphospora, Phialocephala fortinii, Phialocephala finlandia forms association with the root of alpine ericoid. Colonization by this fungus has a unique characteristic; hence they are sometimes termed as ericoid mycorrhizae. Other than ericoid host’s P. fortinii and mycelium radices atrovirens (MRA) also colonizes the roots of Fagus sylvatica, Pinus sylvestris, Phialocephala resinosa, Phialocephala contorta and various alpine perennials (Ahlick and Sieber 1996). Colonization of roots with dark septate endophyte increases the growth of the plant (Read and Haselwandter 1981).
1.2 Trichoderma Species

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 137
Trichoderma species is considered as a major component of soil biodiversity that is mostly associated with plant roots. However, recent studies have revealed that Trichoderma species is not only associated with plant roots but they also persist above ground tissues (Evans et al. 2003; Bailey et al. 2006). Trichoderma species has been widely used as a biocontrol agent against soil borne pathogens (Whipps 2001). Various toxic compounds (e.g., the antibiotics gliotoxin, gliovirin) and extracellular enzymes are released by Trichoderma species to kill fungal pathogen that penetrate and utilize host nutrients (Lorito et al. 1996). Interaction of arbuscular mycorrhiza with Trichoderma shows synergistic effect on plant growth through a wide range mechanism. The use of Trichoderma species as biotrophic decomposer and as endophyte makes it an important tool for agricultural and natural ecosystems (Harman et al. 2004). Some Trichoderma isolates enhances plant growth and reduce damage against nematode damage (Meyer et al. 2001). The genus Trichoderma produces a wide range of metabolites, plant growth regulators (ciclonerodiol), antibiotics (anthraquinone), antifungal (phenolic compounds), antitumor agents and immune modulatory compounds (harzianodiona) that expand the use of this organism commercially (Supothina et al. 2007, Xiao-Yan et al. 2006).
1.3 Piriformospora Species
Piriformospora spp. (Verma et al. 1998) is a root endophytic fungus discovered from Thar Desert of India, belongs to family basidiomycetes (Verma et al. 1999; Pham et al. 2004) that colonize the root cortex of many plant species. Like Arbuscular mycorrhizal fungi, Piriformospora indica and Sebacina vermifera, (Basidiomycota, Sebacinales) has a broad spectrum against soil born fungal pathogens, promotes plant growth and induces resistance against various insects/pests (Varma et al. 2013).
Basiewicz et al. (2012) have described a new species of Piriformospora williamsii and have established its phylogenetic relationship with other members of Sebacinales. Recent study reported 25 Mb genome of P. indica (Zuccaro et al. 2009, 2011). The main features of P. indica genome sequence include 50.68 % GC content, 4.68 % repeat rate, 11,769 protein-coding genes, 5.16 average exons per gene, gene density of 471(number of gene per Mb), 867 secreted proteins, 386 small secreted proteins (SSP), 3,134 unique gene models, 197 unique SSP and 58 tRNA genes. This breakthrough research is the first in-depth genomic study that describes a mutualistic symbiont with a biphasic lifestyle through extensive comparative analysis of the P. indica genome with other Basidiomycota and Ascomycota fungi that have diverse lifestyle strategies identified features typically associated with both, biotrophism and saprotrophism. The tightly controlled expression of the lifestyle- associated gene that sets during the onset of the symbiosis was revealed by microarray analysis that argues for a biphasic root colonisation strategy of P. indica.
This root interacting fungus forms asexual chlamydospores and can easily be grown on various medium (Pham et al. 2004; Prasad et al. 2005). The chlamydospores occur as typical pyriform. P. indica readily colonizes the Arabidopsis thaliana and increases the yield and salt tolerance in barley plant (Oelmüller et al. 2009; Varma et al. 2013). The fungus uses unidentified signaling pathway to protect its host from pathogen and induces systemic resistance (Waller et

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 138
al. 2005; Serfling et al. 2007). Plants infected with P. indica results in higher yield, early flowering and seed production and increase fresh weight (Varma et al. 2013; Prasad et al. 2013). Tolerance to abiotic stress was induced in Arabidopsis thaliana; overall growth and biomass production was achieved in herbaceous mono- and dicots, medicinal plants, and other important crops (Verma et al. 1999).
2. Functions of Fungal Root Endophytes
2.1 Plant Growth Promoting Properties
From the past few decades’ agrochemicals are been used by producers for crop protection, use of this has led to negative impact on crop yield as it increases the resistance of pathogen to antimicrobial agents (Gerhardson 2002). Presently, biological agents are more popular than chemical pesticides; in this concern plant growth promoting activity of root endophytic fungi has the capacity to develop sustainable systems in plant growth (Shoebitz et al. 2009). Plant growth promoting endophytes stimulate growth of plants by solublization of nutrients, production of growth regulating hormones, siderophore, antibiotics, chitinase and cyanide (Pal et al. 2001).
Crop productivity is mainly affected by different stress factors for e.g. high alkaline soil contains high amount of ions that reduces the nitrogen fixation. These adverse conditions reduce the water absorption by plants and induce metabolic changes and decrease the growth rate (Joseph et al. 2007). In such type of soil use of endophytic fungi plays a major role as stress-tolerating organisms. Root endophytic PGPF are those that reside in the roots of the host plants and are beneficial for its host. (Table2,3). PGPF are crop specific and their significance on plant is limited because of variation in climate and soil inconsistency (Khalid et al. 2004, Wu et al. 2005). Plant growth promoting traits of endophytic fungi are phosphate solubilisation; IAA production for e.g. Trichoderma virens produces the auxin related compounds and increases the growth and development of A. thaliana (Cornejo et al. 2009), ammonia and salicylic production, siderophore, HCN production and tolerance to heavy metals. Other factors are also responsible for its effect on PGPF activities i.e., soil type, nutrient, moisture content and organic matter (De Freitas and Germida 1992).
PGPF has been used as biocontrol agent as it cooperatively decreases the pest and pathogens and it is one of the biological and effective approaches to control soil pathogens (Ramamoorthy et al. 2001).
Table 2. Functions of root endophytic fungi on growth of the plant
Fungi Host Effect on plant Reference Cryptosporiopsis spp Larix decidua Increased root length Schulz et al. 2002
Periconia macrospinosa
Brassica compestris Increased root growth Shin et al. 2005
Phialocephala fortinii Rhododendron spp Increased root biomass phosphorus intake
Vohnik et al. 2005

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 139
Piriformospora indica Zea mays, Nicotiana tobaccum, Bacopa monniera, Artemisia annua Spilanthes calva, Withania somnifera
Increased growth and early rooting in tobacco calli Increased overall growth and yield, number of flowers and fruits
Varma et al. 1999; 2012 Rai et al. 2001
Table 3. Functions of root endophytic fungi in host resistance to pathogens Fungi Host Pathogen Effect on host Reference Piriformospora indica
Orzya sativa
Fusarium culmorum, Cochliobolus sativus
Significant improvement in biomass in infected plants
Waller et al. 2005
DSE taxon LtVb3
Brassica campestris
Verticillium longisporum
Symptoms reduced by 90% Narisawa et al. 2004
Phialocephala fortinii
Solanum melongena
Verticillium dahliae Decreased symptoms of pathogen
Narisawa et al. 2004
Fusarium oxysporum
Lycopersicum esculentum
Meliodogyne incognita
Reduced infection through release of antimicrobial compounds by 60%
Hallman and Sikora 1994,1996
Acremonium strictum
Lycopersicum esculentum
Helicoverpa armigera
Decreased development of pupae and larva
Jallow et al. 2004
Acremonium alternatum
Brassica oleracia var. gemmifera
Plutella xlostlla Decreased growth rate of larva
Raps and Vidal 1998
2.2 Mechanism to Control Phytopathogens
2.2.1 Siderophore Production
Iron is a crucial element for microbial growth and mostly it is unavailable to microbes because inorganic iron is extremely insoluble and less in concentration in soil. For iron sequestration many microbes releases the iron binding low molecular weight substances called as siderophores, which has the ability to bind with Fe3+. This bound iron become soluble and transport into the microbial cell and increase its growth (Saharan and Nehra 2011).
2.2.2 Enzyme Production
Hydrolytic enzymes secreted by endophytes can lyse the cell wall of fungal pathogens but not the plant cell wall, and by this it prevents the attack of phytopathogens. Host plants which colonized by endophytes have more vigour because of secretion of phytohormones i.e., cytokines, auxins (Robinson et al. 1998) and help in the absorption of minerals like nitrogen (Lyons et al. 1990) and phosphorus (Malinowski et al. 1999).
2.3 Abiotic Stress Tolerance

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 140
Agriculture is mostly prone to abiotic and biotic stresses, crop yield gets adversely affected as the stress condition increases either due to natural or anthropogenic factors and it may become one of the reasons for productivity limitation. Unfavourable condition like high temperature; elevated CO2, droughts, extreme rainfall, floods, cold waves, global warming etc. cause a negative impact on crops and results in economic losses.
Many genes are involved for adaptation of microbes in stressed environmental conditions (Srivastava et al. 2008). Under extreme environmental condition many microbe posses’ different mechanisms i.e., metabolic process like involvement of enzymatic activities and membrane stability at high temperature and salinity (Madigen 1999). In tropical and sub tropical areas the major problem is high soil temperature for crop production and fungal colonization. Response of all the organisms is by induction of synthesis of specific proteins known as heat shock proteins (HSPs). These HSPs are composed of chaperons (such as GroEL, DnaK, DnaJ, GroES, ClpB, ClpA, ClpX,) and small form of heat shock proteins (HSPs and proteases). The role of chaperons is proper folding and alignment of denatured proteins and proteases that are useful in degradation of irreversible damaged proteins (Munchbach et al. 1999). This has been proved that microbes play a significant role in plant protection by stress management mechanisms; unique properties of stress tolerance, ubiquity, and genetic diversity and now new methods are been employed for their deployment in agriculture. Many reports are also there for AM symbiosis, which improve the plant resistance in drought condition by alteration of plant physiology and metabolic activities, expression of genes (Ruiz-Lozano and Azcon 2000) and dehydration tolerance (Allen and Boosalis1983).
Wu and Xia (2006) demonstrated that induction of Glomus versiforme enhanced the osmotic adjustment in citrus plants under drought condition by increasing the concentration of non-structural carbohydrates, Ca, Mg and K. Under water deficient condition ascorbate and glutathione plays an important role in protection and maintenance of metabolic functions of plants but low concentration of these compounds present in lavender plants shows more drought tolerant. This is because the roots of plant are colonized by Glomus sp. strain (Marulanda et al. 2007). Neto and co workers (2006) reported that A. trifolium plants inoculated with endophytic fungi showed better tolerance to flood by maintenance of osmotic pressure and proline concentration in plant tissues.
One of the examples of endophytic fungus is P. indica that confers the drought tolerance in A. thaliana by priming the expression of stress related genes. In arid and semi arid regions soil salinization is a major problem and it is increasing steadily in various part of world (Giri et al. 2003; Al-Karaki 2006). In natural environmental condition plant are colonized by both external and internal endophytic fungi and they are beneficial fungi helpful in improving plant growth and tolerance under stressed condition (Creus et al. 1998). Most of the AM fungi are incorporated with roots of 80% plants. AMF and endophytic fungi naturally occur in saline areas (Hilderbrandt et al. 2001; Yamato et al. 2008), and the most predominant occurrence of Glomus species i.e., were Glomus intraradices, G. versiform and G. etunicatum in severe saline soil of Tabriz plains having electrical conductivity of 162 dS m-1.
The harmful impact of salinity is not on the host plants but also on endophytic fungi; it can alter

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 141
the colonization capacity, growth of hyphae and germination of spores of fungi. Many reports are there on the effect of salinity on fungi (Jahromi et al. 2008). Percentage of root colonization by fungi decreases as the concentration of NaCl increases in soil (Giri et al. 2007; Sheng et al. 2008) indicating salinity inhibits the fungi colonization and arbuscular mycorrhiza formation (Sheng et al. 2008). Juniper and Abbott (2006) showed that spore germination of fungi is delayed rather than prevented. In synthetic media, the number of spores’ produced by Glomus intraradices decreases as the medium contains the different concentration of salts. This reduction suggests that if salinity continuous there can be decline in colonization percentage by reducing the capacity of inoculums (spores). Abiotic stress results into series of physiological, morphological, molecular and biochemical changes in plants and adversely growth and productivity.
Mainly clavicipitaceous endophytic fungi colonize the tissues and provide the beneficial effects on host plants by increasing the resistance to pathogens and stressed conditions (Kuldau and Bacon 2008). Habitat specific adaptation also supported by F. culmorum, C. protuberate and C. magna endophytes. F. culmorum confers salt but not heat or disease tolerance; C. protuberata confers heat but not disease or salt tolerance; and C. magna confers disease but not heat or salt tolerance (Rodriguez et al. 2008). For e.g. in the geothermal soils of Yellowstone National Park, Dichanthelium lanuginosum plant is heavily colonized by one of the dominant endophyte i.e. Curvularia protuberate. This endophyte confers the heat tolerance to the host plant and when they are separated from one another none of them survive alone when exposed to temperature >38°C (Redman et al. 2002). These endophytes symbiotically tolerates the stress conform the evolutionary dynamics plays differently in various habitats and confer the stress tolerance to plants (Table 4).
Table 4. Some examples of fungal endophytes that conferred abiotic stress tolerance
Fungal endophytes Abiotic stress Host Plants Refrences Neotyphodium spp Drought Festuca pratensis, Perennial Ryegrass,
Festuca arizonica Malinowski et al. 1997 Barker et al. 1997 Morse et al. 2002
Curvularia protuberate
Heat Dichanthelium lanuginosum Redman et al. 2002
Curvularia protuberate (Cp4666D)
Drought Dichanthelium lanuginosum Rodriguez et al. 2008
Curvularia spp Heat/Drought Lycopersicum esculentum Rodriguez and Redman 2008
Fusarium culmorum (Fc18)
Drought Leymus mollis, Oryza sativa, Lycopersicum, esculentum
Rodriguez et al. 2008
Fusarium culmorum (FcRed1)
Salinity Leymus mollis, Oryza sativa, Lycopersicum esculentum, Dichanthelium lanuginosum
Rodriguez et al. 2008
Colletotrichum spp Drought Lycopersicum esculentum Rodriguez et al. 2004 Fusarium spp Alternaria spp
Heat/Drought Lycopersicum esculentum Rodriguez and Redman 2008
Piriformospora indica Salinity Hordeum vulgare Waller et al. 2005
Previous studies documented that stress tolerance by fungal endophytes in host plant plays an important role in the survival of plants in extreme stressed condition (Rodriguez et al. 2004).

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 142
For e.g. Class II type of endophytes confers the heat tolerance to plants in geothermal soils (Redman et al. 2002), extent of colonization by endophytes directly related to the resistance to pathogens of host plant (Arnold et al. 2003) and endophyte can confer drought tolerance to many host plants (Waller et al. 2005). Endophytes colonized plants express different range of adaptations to biotic and abiotic stress condition i.e., drought (West 1994), mineral imbalance (Malinowski et al. 1997) and soil acidity (Malinowski and Belesky 1999).
2.4 Natural Products from Root Endophytes
The endophytic fungi associated with the roots of the plant can produce natural active secondary metabolites that have many industrial and agriculture applications. Endophytes are known to produce various metabolites such as antibacterial, anticancer, antifungal, antiviral and immunosuppressant compounds e.g. paclitaxel, torreyanic acid, etc. which can be used in various field of medicine. Isolation and characterization of endophytic fungi for their use in medicine and industry depends upon the environmental as well as on host endophyte relationship. Since last one decade number of bioactive compounds have been isolated and characterized from root endophytic fungi, which belongs to groups like phenolics, quinones, flavonoids, terpenoids, xanthones and other biological active compound (Tan and Zou 2001; Gunatilaka 2006; Zhang et al. 2006). Various pathways like polyketide, isoprenoid, and amino acid derivation synthesize these secondary metabolites. Strobel et al. (1996) discovered that paclitaxel was produced by Pestalotiopsis microspore, isolated from Taxus wallichiana. Muscodoralbus (Strobel et al. 2001) isolated from Cinnamom zeylanicum produces an antifungal compound that has proved lethal to Aspergillus fumigates and Candida albicans (Woropong et al. 2001). Myco-diesel hydrocarbons from endophyte Gliocladium roseum can be used as an alternative source for bio-diesels (Strobel et al. 2008). Gliocladium catenulatum endophyte isolated from Theobroma cacao is used as a biocontrol agent against Crinipellis perniciosa Witches Broom disease of the Cacao tree (Rubini et al. 2005). Aspergillus fumigates isolated from twigs of the Juniperus communis, produces deoxypodophyllotoxin that has anticancer and antiviral activity. Taxol, a powerful anti cancer drug is produced by an endophyitc fungi Taxus brevifolia Nutt (Strobel et al. 1993). Endophytic fungi Pestalotiopsis microspora has been used for the isolation of antioxidant metabolites such as Pestacin and Isopestacin. Grass-associated endophyes such as Epichlo¨e and Neotyphodium species have been detected with alkaloids.
A number of antifungal products are produced from endophytes. Griseofulvin was first reported from endophyte Abies holophylla that was used as an antifungal against plant pathogen. Griseofulvin and dechloro griseofulvin isolated from Xylaria species have shown high antifungal activity against Magnaporthe grisea, Puccinia recondite and Blumeria graminis f. sp. Hordei (Park et al. 2005). Endophytes are known to produce many antiviral agents. Cytonic acid A and B isolated from Cytonaema sp. act as an inhibitor for human cytomegalovirus (hCMV) protease (Jensen and Roulund 2004).
Kusari et al. (2009) demonstrated one of the examples of horizontal transfer of genes from host to endophyte. Aspergillus fumigatus fresenius was isolated from the twings of Juniperus communis L. Horstmann, which is a producer of deoxypodophyllotoxin that is known for anti

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 143
therapeutically properties like antitumor, antiviral, and anti-inflammatory activities. Pimentel et al. (2011) reported that bioactive compounds produced by endophytes could be use in biotransformation process. Biotransformation of grandisin to tetrahydrofuran by Phomopsis species is a promising method for the production of metabolites by these endophytes. Biotransformation has also been reported from endophytic fungus isolated from Aphelandra tetragona roots (Zikmundová et al. 2002). These endophytic fungi are able to mimic mammalian metabolism through various reactions of biotransformation (Borges et al. 2007).
Table 5. List of secondary metabolites produced by fungal root endophytes
Fungus Source Compound Activity Reference Penicillium expansum
Excoecaria agallocha
Polyphenols, expansols A & B
Cytotoxic Lu et al. 2010
Curvularia lunata
Niphates olemda Cytoskyrins Antibacterial, anticancer agent
Brady et al. 2000, Jadulco et al. 2002
Phoma medicaginis
Medicago sativa, Medi- cago lupulina
Brefeldine A Antibacterial Weber et al. 2004b
Phomopsis spp PSU-D15
Garcinia dulcis Phomoenamide Antimycobacterial activity
Rukachaisirikul et al. 2008
Muscodor albus
Cinnamomum zeylanicum
Colatile organic compounds
Antifungal, antibacterial Ezra et al. 2004
Pestelotiopsis microspora
Taxus wallichiana Paclitaxel Anticancer Strobel et al.1996
Chaetomium acuminata
Edenia gomezpompae
Naphthoquinone spiroketal
Allelochemical activity Macias- Rubalcava et al. 2008
Muscodor vitigenus
Paullinia paullinioides
Naphthalene
Insecticides, antimicrobials, anti helminthics and vermicides
Strobel et al. 2007
3. Host Endophyte Relationship
Several theories have been developed to mark the advantages of root endophyte colonization. Two of the most prominent methods are plant (Newsham 2011) and phytohormones production (Schulz and Boyle 2005). The association between host and endophyte depends upon the environmental condition and ranges from mutualistic, neutral or antagonist (Kogel et al. 2006; Moricca and Ragazzi 2008). The interaction is highly variable and transitory (Bacon and Yates 2006). Although transitory association exists; endophytic fungi are known to show various benefits to the host plant like tolerance to abiotic and biotic stress (Schulz and Boyle 2005; Rodriguez and Redman 2008). Schulz et al. (1999) demonstrated that the host endophyte interaction is a balanced antagonist that refers to a state of equilibrium between fungal virulence and plant defences. Although change can occur through an imbalance in flow of nutrients, change in environmental (Moricca and Ragazzi 2008) and stress condition (Halmschlager et al. 1993). Genetic variation plays an important role in host endophyte

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 144
interactions. The relationship between host genotype and their lifestyle from mutualistic to parasitic has been shown by some fungal isolates (Unterseher and Schnittler 2010).
Stress tolerance to reactive oxygen species (ROS) produced by endophytes protects the host plant from drought and pathogens. ROS produced by endophytes have increased the production of antioxidant such as phenols and/or flavonoids in endophyte-infected plants. ROS denatures the cell membrane of the plant thus involving influx of nutrients that is radially absorbed by fungal hyphae (White and Torres 2010). Host specificity, selectivity and host preference also plays an important role in host endophyte relationship (Cohen 2006).Endophytic fungi interact with the roots of the plants in several ways depending upon the environment and species involved. For e.g. in tree species the interaction between the fungal root endophytes and plants takes place through the formation of mantle and hartig net. Phialocephala finlandica (C. finlandica) develops a hartig net by radial elongation that is confined till the epidermis and the mantle to the roots of yellow birch (Wilcox and Wang 1987 b). It has been reported that roots of oak and pine have been colonized with large number of root endophytic fungi (Reininger and Sieber 2012) as well as with AM fungi (Dickie et al. 2001).
4. Conclusion
Root endophytic fungi are the group of microorganisms that reside inside the plant roots and their association varies from mutualistic to pathogenic. Piriformospora indica has a broad spectrum of resistance against soil-borne pathogens and promotes plant growth. Endophytic fungi are excellent source of bioactive natural metabolites with broad range of functions and structural diversity therefore it can be used as a bio fertilizer. It also plays an important role in conferring resistance to biotic and abiotic stress conditions. The ease of culturing root endophytic fungi can fulfil the future demands in medical, agriculture and pharmaceutical industries.
Acknowledgments
Authors are grateful to Dr. Ashok K. Chauhan, Founder President, Amity University (Noida, India) for his encouragement and providing excellent facilities. Neha Chadha and Manjita Mishra are very thankful to DRDO, ICAR and DBT, Government of India for granting research fellowship.
References
Addy, H. D. Piercey, M. M., & Currah, R. S. (2005). Microfungal endophytes in roots. Canadian Journal of Botany. 83.1-13. http://dx.doi.org/10.1139/b04-171
Ahlich, K., & Sieber, T. N. (1996). The profusion of dark septate endophytic fungi in non-ectomycorrhizal fine roots of forest trees and shrubs. New phytologist. 132, 259-270. http://dx.doi.org/10.1111/j.1469-8137.1996.tb01845.x
Al-Karaki, G. N. (2006). Nursery inoculation of tomato with arbuscular mycorrhizal fungi and subsequent performance under irrigation with saline water. Scientia Horticulturae. 109, 1-7. http://dx.doi.org/10.1016/j.scienta.2006.02.019

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 145
Allen, M. F., & Boosalis, M. G. (1983). Effect of two species of VA mycorrhizal fungi on drought tolerance of winter wheat. New Phytologist. 93, 67-76. http://dx.doi.org/10.1111/j.1469-8137.1983.tb02693.x
Aly, A. H. Debbab, A. Kjer, J., & Proksch, P. (2010). Fungal endophytes from higher plants: a prolific source of phytochemicals and other bioactive natural products. Fungal Diversity. 41, 1-16. http://dx.doi.org/10.1007/s13225-010-0034-4
Ananda, K., & Sridhar, K. R. (2002). Fungal endophytes in submerged roots. In: B. Schulz, C. Boyle, T. Sieber (Eds.), Microbial Root Endophytes (pp. 179-190) Springer Berlin Heidelberg.
Apel, K., & Hirt, H. (2004). Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annual Review of Plant Biology. 55, 373-399. http://dx.doi.org/10.1146/annurev.arplant.55.031903.141701
Arnold, A. E. Mejía, L. C. Kyllo, D. Rojas, E. I. Maynard, Z. Robbins, N., & Herre, E. A. (2003). Fungal endophytes limit pathogen damage in a tropical tree. Proceedings of the National Academy of Sciences, USA. 100, 15649-15654. http://dx.doi.org/10.1073/pnas.2533483100
Arnold, A. E., & Lutzoni, F. (2007). Diversity and host range of foliar fungal endophytes: are tropical trees biodiversity hot spots? Ecology. 88, 541-549. http://dx.doi.org/10.1890/05-1459
Bacon, C. W., & White, J. F. J. (2000). Physiological adaptations in the evolution of endophytism in the Clavicipitaceae. In: C.W. Bacon & J.F.J. White (Eds.), Microbial endophytes (pp. 237-263). New York, NY, USA: Marcel Dekker Inc.
Bacon, C. W., & Yates, I. E. (2006). Endophytic root colonization by Fusarium species: Histology, plant interactions and toxicity. In: B. Schultz, C. Boyle, T. Sieber (Eds.), Microbial root endophytes. Berlin: Springer-Verlag. http://dx.doi.org/10.1007/3-540-33526-9_8
Bailey, B. Bae, H. Strem, M. Roberts, D. Thomas, S. Crozier, J. Samuels, G. Choi, I., & Holmes, K. (2006). Fungal and plant gene expression during the colonization of cacao seedlings by endophytic isolates of four Trichoderma species. Planta. 224(6), 1449-1464. http://dx.doi.org/10.1007/s00425-006-0314-0
Baker, A. C. Rowan, R., & Knowlton, N. (1997). Symbiosis ecology of two Caribbean acroporid corals. In: H.A. Lessios, & I.G. Macintyre (Eds.), Proceedings of the 8th International Coral Reef Symposium (pp. 1295-1300). Smithsonian Tropical Research Institute, Balboa, Panama
Barker, D. J. Hume, D. E., & Quigley, P. E. (1997). Negligible physiological responses to water deficit in endophyte-infected and uninfected perennial ryegrass. In: C.W. Bacon, & N.S. Hill (Eds.), Neotyphodium/Grass Interactions (pp. 137 139). New York: Plenum Press
Barrow, J. R., & Aaltonen, R. E. (2001). A method of evaluating internal colonization of Atriplex canescens (Pursh) Nutt.roots by dark septate fungi and how they are influenced by host physiological activity. Mycorrhiza. 11, 199-205. http://dx.doi.org/10.1007/s005720100111

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 146
Basiewicz, M. Weiß, M. Kogel, K. H. Langen, G. Zorn, H., & Zuccaro, A. (2012). Molecular and phenotypic characterization of Sebacina vermifera strains associated with orchids, and the description of Piriformospora williamsii sp. nov. Fungal Biology. 116, 204-213. http://dx.doi.org/10.1016/j.funbio.2011.11.003
Benhamou, N., & Garand, C. (2001). Cytological analysis of defense related mechanisms induced in pea root tissues in response to colonization by nonpathogenic Fusarium oxysporum F047. Phytopathology. 91, 730-740. http://dx.doi.org/10.1094/PHYTO.2001.91.8.730
Bischoff, J. F., & White, J. F. (2005). Evolutionary development of the Clavicipitaceae. In: J. Dighton, J.F. White, P. Oudemans (Eds.), The fungal community: Its organization and role in the ecosystem (pp. 505-518). Boca Raton, FL, USA: Taylor & Francis.
Borges, K. B. Borges, W. D. S. Pupo, M. T., & Bonato, P. S. (2007). Endophytic fungi as models for the stereo selective biotransformation of thioridazine. Applied Microbiology and Biotechnology. 77, 669- 674. http://dx.doi.org/10.1007/s00253-007-1171-x
Brady, S. F. Wagenaar, M. M. Singh, M. P. Janso, J. E., & Clardy, J. (2000). The cytosporones, new octaketide antibiotics isolated from an endophytic fungus. Organic Letters. 14, 4043-4046. http://dx.doi.org/10.1021/ol006680s
Brundrett, M. (2004). Diversity and classification of mycorrhizal associations. Biological Reviews. 79, 473-495. http://dx.doi.org/10.1017/S1464793103006316
Brundrett, M. C., & Kendrick, W. B. (1988). The mycorrhizal status, root anatomy, and phenology of plants in a sugar maple forest. Canadian Journal of Botany. 66, 1153-1173. http://dx.doi.org/10.1139/b88-166
Campanile, G. Ruscelli, A., & Luisi, N. (2007). Antagonistic activity of endophytic fungi towards Diplodia corticola assessed by in vitro and in planta tests. European Journal of Plant Pathology. 117, 237-246. http://dx.doi.org/10.1007/s10658-006-9089-1
Clay, K. (1988). Fungal endophytes of grasses: a defensive mutualism between plants and fungi. Ecology. 69, 10-16. http://dx.doi.org/10.2307/1943155
Clay, K. (1990). Fungal endophytes of grasses. Annual Review of Ecology and Systematics. 21, 275-297. http://dx.doi.org/10.1146/annurev.es.21.110190.001423
Clay, K., & Schardl, C. L. (2002). Evolutionary origins and ecological consequences of endophyte symbiosis with grasses. American Naturalist. 160, S99-S127. http://dx.doi.org/10.1086/342161
Cohen, S. D. (2006). Host selectivity and genetic variation of Discula umbrinella isolates from two oak species: analyses of intergenic spacer region sequences of ribosomal DNA. Microbial Ecology. 52, 463-469. http://dx.doi.org/10.1007/s00248-006-9073-5
Cornejo, H. A. C. Rodrı´guez, L. M. Penagos, C. C., & Bucio, J. L. (2009). Trichoderma virens, a Plant Beneficial Fungus, Enhances Biomass Production and Promotes Lateral Root Growth through an Auxin-Dependent Mechanism in Arabidopsis. Plant Physiology. 149, 1579-1592.

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 147
http://dx.doi.org/10.1104/pp.108.130369
Costa, J. M., & Loper, J. E. (1994). Characterization of siderophore production by the biological control agent Enterobacter cloacae. Molecular Plant-Microbe Interaction. 7, 440-448. http://dx.doi.org/10.1094/MPMI-7-0440
Creus, C. M. Sueldo, R. J., & Barassi, C. A. (1998). Water relations in Azospirillum-inoculated wheat seedlings under osmotic stress. Canadian Journal of Botany. 76, 238-244. http://dx.doi.org/10.1139/b97-178
Currah, R. S. Sigler, L., & Hambleton, S. (1987). Microbial Endophytes of Orchid Roots. In: B. Schulz, C. Boyle, T. Sieber (Eds.), Microbial Root Endophytes (pp. 153-177). Springer Berlin Heidelberg.
De Bary, H. A. (1884). Vergleichende Morphologie und Biologie der Pilze Mycetozoen und Bacterien.Verlag von Wilhelm Engelmann, Leipzig. Berlin.
De Freitas, J. R., & Germida, J. J. (1992). Growth promotion of winter wheat by fluorescent pseudomonads growth under growth chamber conditions. Soil Biology and Biochemistry. 24, 1127-l 135. http://dx.doi.org/10.5962/bhl.title.42380
Dickie, I. A. Koide, R. T., & Fayish, A. C. (2001). Vesicular-arbuscular mycorrhizal infection of Quercus rubra seedlings. New Phytologist. 151, 257-264. http://dx.doi.org/10.1046/j.1469-8137.2001.00148.x
Dorothy, D. A., & Kandikere, R. S. (2009). Assemblage and diversity of fungal associated with mangrove wild legume Canavalia cathartica. Tropical and Subtropical Agroecosystems. 10, 225-235.
Evans, C. Holmes, K., & Thomas, S. (2003). Endophytes and mycoparasites associated with an indigenous forest tree, Theobroma gileri, in Ecuador and a preliminary assessment of their potential as biocontrol agents of cocoa diseases. Mycological Progress. 2(2), 149-160. http://dx.doi.org/10.1007/s11557-006-0053-4
Ezra, D. Hess, W. M., & Strobel, G. A. (2004). New endophytic isolates of Muscodor albus, a volatile-antibiotic-producing fungus. Microbiology. 150, 4023-4031. http://dx.doi.org/10.1099/mic.0.27334-0
Faeth, S. H. Gardner, D. R. Hayes, C. J. Jani, A. Wittlinger, S. K., & Jones, T. A. (2006). Temporal and spatial variation in alkaloid levels in Achnatherum robustum, a native grass infected with the endophyte eotyphodium. Journal of Chemical Ecology. 32, 307-324. http://dx.doi.org/10.1007/s10886-005-9003-x
Gamboa, M. A., & Bayman, P. (2001). Communities of endophytic fungi in leaves of a tropical timber tree (Guarea guidonia: Meliaceae). Biotropica. 33, 352-360. http://dx.doi.org/10.1111/j.1744-7429.2001.tb00187.x
Gao, K. X., & Mendgen, K. (2006). Seed-transmitted beneficial endophytic Stagonospora sp. can penetrate the walls of the root epidermis, but does not proliferate in the cortex, of

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 148
Phragmites australis. Canadian Journal of Botany. 84, 981-988. http://dx.doi.org/10.1139/b06-056
Gerhardson, B. (2002). Bology substitutes for pesticides. Trends Biotechnology. 20, 338-343. http://dx.doi.org/10.1016/S0167-7799(02)02021-8
Giri, C. Pengra, B. Zhu, Z. Singh, A., & Tieszen, L. (2007). Monitoring mangrove forest dynamics of the Sundarbans in Bangladesh and India using multi-temporal satellite data from 1973-2000. Estuarine, Coastal and Shelf Science. 73, 91-100. http://dx.doi.org/10.1016/j.ecss.2006.12.019
Giri, V. Wilkinson, M., & Gower, D. J. (2003). A new species of Gegeneophis Peters (Amphibia: Gymnophiona: Caeciliidae) from southern Maharashtra, India, with a key to the species of the genus. Zootaxa. 351, 1-10.
Gunatilaka, A. A. L. (2006). Natural products from plant-associated microorganisms: Distribution, structural diversity, bioactivity, and implications of their occurrence. Journal of Natural Products. 69, 509-526. http://dx.doi.org/10.1021/np058128n
Hallmann, J. Sikora, R. A. (1994). Influence of Fusarium oxysporum, a mutualistic fungal endophyte, on Meloidogyne incognita infection of tomato. Journal for Plant Disease and Plant Protection. 101, 475-481.
Hallmann, J., & Sikora, R. A. (1996). Toxicity of fungal endophyte secondary metabolites to plant- parasitic nematodes and soil-borne plant-pathogenic fungi. 102, 155-162.
Halmschlager, E. Butin, H., & Donabauer, E. (1993). Endophytic fungi in leaves and twigs of Quercus petrea. European Journal of Plant Pathology. 23, 51-63. http://dx.doi.org/10.1111/j.1439-0329.1993.tb00805.x
Harman, G. Howell, C. Viterbo, A. Chet, I., & Lorito, M. (2004). Trichoderma species- oportunistic, avirulent plant symbionts. Nature Reviews Microbiology. 2(1), 43 - 56. http://dx.doi.org/10.1038/nrmicro797
Herre, E. A. Bael, S. A. V. Maynard, Z. Robbins, N. Bischoff, J. Arnold, A. E. Rojas, E. Mejía, L. C. Cordero, R. A. Woodward, C., & Kyllo, D. A. (2005). Tropical plants as chimera: some implications of foliar endophytic fungi for the study of host plant defense, physiology, and genetics. In: D.F.R.P. Burslem, M.A. Pinard, S.E. Hartley (Eds.), Biotic interactions in the tropics, Cambridge, UK (pp. 226-240). Cambridge University Press.
Hilderbrandt, U. Janetta, K. Ouziad, F. Renne, B. Nawrath, K. Bothe, H. (2001). Arbuscular mycorrhizal colonization of halophytes in Central European salt marshes. Mycorrhiza. 10. 175-183. http://dx.doi.org/10.1007/s005720000074
Iqbal, S. H. Firdaus-e-Bareen, & Yousaf, N. (1995). Fungal endophytes in submerged roots. In: B. Schulz, C. Boyle, T. Sieber (Eds.), Microbial Root Endophytes (pp. 179-190) Springer Berlin Heidelberg.
Jadulco, R. Brauers, G. Edrada, R. A. Ebel, R. Wray, V. Sudarsono, V., & Proksch, P. (2002).

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 149
New metabolites from sponge- derived fungi Curvularia lunata and Cladosporium herbarum. Journal of Natural Products. 65, 730-733. http://dx.doi.org/10.1021/np010390i
Jahromi, F. Aroca, R. Porcel, R., & Ruiz-Lozano, J. M. (2008). Influence of salinity on the in vitro development of Glomus intraradices and on the in vivo physiological and molecular responses of mycorrhizal lettuce plants. Microbial Ecology. 55, 45-53. http://dx.doi.org/10.1007/s00248-007-9249-7
Jallow, M. F. A. Dugassa-Gobena, D., & Vidal, S. (2004). Indirect interaction between an unspecialized endophytic fungus and a polyphagous moth. Basic and Applied Ecology. 5, 183-191. http://dx.doi.org/10.1078/1439-1791-00224
Jensen, A. M. D., & Roulund, N. (2004). Occurrence of Neotyphodium endophytes in permanent grassland with perennial ryegrass (Lolium perenne) in Denmark. Agriculture, Ecosystems and Environment. 104, 419-427. http://dx.doi.org/10.1016/j.agee.2004.01.044
Joseph, B. Patra, R. R., & Lawrence, R. (2007). Characterization of plant growth promoting rhizobacteria associated with chickpea (Cicer arietinum L.). International Journal of Plant Production. 2, 141-152.
Juniper, S., & Abbott, L. K. (2006). Soil salinity delays germination and limits growth of hyphae from propagules of arbuscular mycorrhizal fungi. Mycorrhizal. 6, 371-379. http://dx.doi.org/10.1007/s00572-006-0046-9
Khalid, A. Arshad, M., & Zahir, Z. A. (2004). Screening plant growth promoting rhizobacteria for improving growth and yield of wheat. Journal of Applied Microbiology. 96(3), 473-480. http://dx.doi.org/10.1046/j.1365-2672.2003.02161.x
Kharwar, R. N. Mishra, A. Gond, S. K. Stierle, A., & Stierle, D. (2011). Anticancer compounds derived from fungal endophytes: their importance and future challenges. Natural Product Reports. 28, 1208-1228. http://dx.doi.org/10.1039/c1np00008j
Kimmons, C. A. Gwinn, K. D., & Bernard, E. C. (1990). Nematode reproduction on endophyte-infected and endophyte-free tall fescue. Plant disease. 74, 757-761. http://dx.doi.org/10.1094/PD-74-0757
Kogel, K. H. Franken, P., & Hückelhoven, R. (2006). Endophyte or parasite— what decides. Current Opinion in Plant Biology. 9, 358-363. http://dx.doi.org/10.1016/j.pbi.2006.05.001
Kohn, L. M., & Stasovski, E. (1990). The mycorrhizal status of plants at Alexandra Fiord, Ellesmere Island, Canada, a high Arctic site. Mycologia, 82, 23-35. http://dx.doi.org/10.2307/3759959
Kuldau, G., & Bacon, C. (2008). Clavicipitaceous endophytes: their ability to enhance resistance of grasses to multiple stresses. Biological Control. 46, 57-71. http://dx.doi.org/10.1016/j.biocontrol.2008.01.023
Kusari, S. Lamshoft, M., & Spiteller, M. (2009). Aspergillus fumigatus fresenius, an endophytic fungus from Juniperus communis L. Horstmann as a novel source of the anticancer

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 150
pro-drug deoxypodophyllotoxin. Applied Microbiology. 107, 1364-5072. http://dx.doi.org/10.1111/j.1365-2672.2009.04285.x
Kusari, S., & Spiteller, M. (2012b). Metabolomics of endophytic fungi producing associated plant secondary metabolites: progress, challenges and opportunities. In: U. Roessner (Ed), (pp. 241-266). Metabolomics, Rijeka, Croatia: InTech.
Lorito, M. Woo, S. L, D’Ambrosio, M. Harman, G. E. Hayes, C. K. Kubicek, C. P., & Scala, F. (1996). Synergistic Interaction between Cell Wall Degrading Enzymes and Membrane Affecting Compounds. Molecular Plant-Microbe Interactions. 9, 206-213. http://dx.doi.org/10.1094/MPMI-9-0206
Lu, Z. Zhu, H. Fu, P. Wang, Y. Zhang, Z. Lin, H. Liu, P. Zhuang, Y. Hong, K., & Zhu, W. (2010). Cytotoxic polyphenols from the marine-derived fungus Penicillium expansum. Journal of Natural Products. 73, 911-914. http://dx.doi.org/10.1021/np100059m
Lyons, P. C. Evans, J. J., & Bacon, C. W. (1990). Effects of the fungal endophyte Acremonium coenophialum on nitrogen accumulation and metabolism in Tall Fescue. Plant Physiology. 92, 726-732. http://dx.doi.org/10.1104/pp.92.3.726
Macías-Rubalcava, M. L. Hernández-Bautista, B. E. Jiménez-Estrada, M. González, M. C. Glenn, A. E. Hanlin, R. T. Hernández-Ortega, S. Saucedo-Garcia, A. Muria-González, J. M. Anaya, A. L. (2008). Naphthoquinone spiroketal with allelochemical activity from the newly discovered endophytic fungus Edenia gomezpompae. Phytochemistry. 69, 1185-1196. http://dx.doi.org/10.1016/j.phytochem.2007.12.006
Madigen, O. (1999). Thermophilic and halophilic extremophiles. Current Opinion in Microbiology. 2, 265-269. http://dx.doi.org/10.1016/S1369-5274(99)80046-0
Malinowski, D. Leuchtmann, A. Schmidt, D. Nsberger, J. (1997). Growth and water status in meadow fescue (Festuca pratensis) is differently affected by its two natural endophytes. Agronomy Journal. 89, 673-678. http://dx.doi.org/10.2134/agronj1997.00021962008900040021x
Malinowski, D. P. Brauer, D. K., & Belesky, D. P. (1999). Neotyphodium coenophialum-endophyte affects root morphology of tall fescue grown under phosphorus deficiency. Journal of Agronomy and Crop Science. 183, 53-60. http://dx.doi.org/10.1046/j.1439-037x.1999.00321.x
Malinowski, D. P., & Belesky, D. P. (1999). Endophyte infection enhances the ability of tall fescue to utilize sparingly available phosphorus. Journal of Plant Nutrition. 22, 835-853. http://dx.doi.org/10.1080/01904169909365675
Malla, R. Prasad, R. Giang, P. H. Pokharel, U. Oelmueller, R. Varma, A. (2004). Characteristic features of symbiotic fungus Piriformospora indica. Endocytobiosis and Cell Research. 15, 579-600.
Marulanda, A. Porcel, R. Barea, J., & Azcón, R. (2007). Drought tolerance and antioxidant activities in lavender plants colonized by native drought-tolerant or drought- sensitive Glomus

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 151
species. Microbial Ecology. 54, 543-552. http://dx.doi.org/10.1007/s00248-007-9237-y
Marvanova, L., & Fisher, P. J. (1991). Fungal endophytes in submerged roots. In: B. Schulz, C. Boyle, T. Sieber (Eds.), Microbial Root Endophytes (pp. 179-190). Springer Berlin Heidelberg.
McKendrick, S. L. Leake, J. R., & Read, D. J. (2000). Microbial Endophytes of Orchid Roots. In: B. Schulz, C. Boyle, T, Sieber (Eds.), Microbial Root Endophytes (pp. 153-177). Springer Berlin Heidelberg.
Meyer, S. L. F. Roberts, D. P. Chitwood, D. J. Carta, L. K. Lumsden, R. D., & Mao, W. (2001). Application of Burkholderia cepacia and Trichoderma virens alone and in combinations against Meloidogyne incognita on bell pepper. Nematropica. 31, 75-86.
Moricca, S., & Ragazzi, A. (2008). Fungal endophytes in Mediterranean oak forests: a lesson from Discula quercina. Phytopathology. 98, 380-386. http://dx.doi.org/10.1094/PHYTO-98-4-0380
Morse, L. J. Day, T. A. Faeth, & S. H. (2002). Effect of Neotyphodium endophyte infection on growth and leaf gas exchange of Arizona fescue under contrasting water availability regimes. Environmental and Experimental Botany. 48, 257-268. http://dx.doi.org/10.1016/S0098-8472(02)00042-4
Munchbach, M. Nocker, A., & Narberhaus, F. (1999). Multiple small heat shock proteins in rhizobia. Journal of Bacteriology. 181, 83-90.
Narisawa, K. Usuki, F., & Hashiba, T. (2004). Control of Verticillium yellows in Chinese cabbage by the dark septate endophytic fungus LtVB3. Phytopathology. 94, 412-418. http://dx.doi.org/10.1094/PHYTO.2004.94.5.412
Neto, D. Carvalho, L. M. Cruz, C., & Martin-Loucao, M. A. (2006). How do mycorrhizas affect C and N relationships in flooded Aster trifolium plants. Plant Soil. 279, 51-63. http://dx.doi.org/10.1007/s11104-005-6333-y
Newsham, K. K. (2011). A meta-analysis of plant responses to dark septate root endophytes. New Phytolologist. 190. 783-793. http://dx.doi.org/10.1111/j.1469-8137.2010.03611.x
Oelmuller, R. Sherameti, I. Tripathi, S., & Varma, A. (2009). Piriformospora indica- a cultivable root endophyte with multiple biotechnological applications. Symbiosis. 49, 1-17. http://dx.doi.org/10.1007/s13199-009-0009-y
Pal, K. K. Tilak, K. V. B. R. Saxena, A. K. Dey, R., & Singh, C. S. (2001). Supression of maize root disesase caused byMacrophomina phaseolina, Fusarium moniliforme and Fusarium graminearum by plant growth prmoting rhzobacteria. Microbiological Research. 156, 209-223. http://dx.doi.org/10.1078/0944-5013-00103
Park, J. H. Choi, G. J. Lee, H. B. Kim, K. M. Jung, H. S. Lee, S. W. Jang, K. S., & Cho, K. Y. (2005). Griseofulvin from Xylaria sp. strain F0010, and endophytic fungus of Abies holophylla and its antifungal activity against plant pathogenic fungi. Journal Microbiology and Biotechnology. 15, 112-117.

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 152
Patterson, C. G. Potter, D. A., & Fanin, F. F. (1991). Feeding deterrence of alkaloids from endophyte-infected grasses to Japanese beetle grubs. Entomologia Experimentalis et Applicata. 61, 285-289. http://dx.doi.org/10.1111/j.1570-7458.1991.tb01561.x
Pham, G. H. Kumari, R. Singh, A. N. Sachdev, M. Prasad, R. Kaldorf, M. Buscot, F. Oelmüller, R. Peśkan, T. Weiss, M. Hampp, R., & Varma, A. (2004). Axenic cultures of Piriformospora indica. In: A. Varma, L. Abbott, D. Werner, R. Hampp (Eds.), Plant Surface Microbiology (pp 593-616). Springer-Verlag, Germany.
Pimentel, M. R. Molina, G. Dionísio, A. P. Junior, M. R. M., & Pastore, G. M. (2011). The Use of Endophytes to Obtain Bioactive Compounds and Their Application in Biotransformation Process. Biotechnology Research International. 1-11. http://dx.doi.org/10.4061/2011/576286
Prasad, R. Kamal, S. Sharma, P. K. Oelmueller, R., & Varma, A. (2013). Root endophyte Piriformospora indica DSM 11827 alters plants morphology, enhances biomass and antioxidant activity of medicinal plant Bacopa monniera. Journal of Basic Microbiology. 53(12), 1016-1024. http://dx.doi.org/10.1002/jobm.201200367
Prasad, R. Pham, G. H. Kumari, R. Singh, A. Yadav, V. Sachdev, M. Peskan, T. Hehl, S. Oelmuller, R. Garg, A. P., & Varma, A. (2005). Sebacinaceae: culturable mycorrhiza-like endosymbiotic fungi and their interaction with non-transformed and transformed roots. In: S. Declerck, D.G. Strullu, A. Fortin (Eds.), In vitro Culture of Mycorrhizas (pp. 291-312). Soil Biology Series, Springer-Verlag, Germany
Rai, M. Acharya, D., & Singh, A. (2001). Positive growth responses of the medicinal plants Spilanthes calva and Withania somnifera to inoculation by Piriformospora indica in a field trial. Mycorrhiza. 11, 123-128. http://dx.doi.org/10.1007/s005720100115
Ramamoorthy, V. Viswanathan, R. Raguchander, T. Prakasam, V., & Samiyappan, R. (2001). Induction of systemic resistance by plant growth promoting rhizobacteria in crop plants against pests and diseases. Crop Protection. 20, 1-11. http://dx.doi.org/10.1016/S0261-2194(00)00056-9
Raps, A., & Vidal, S. (1998). Indirect effects of an unspecialized endophytic fungus on specialized plant-herbivorous insect interactions. Oecologia. 114, 541-547. http://dx.doi.org/10.1007/s004420050478
Read, D. J., & Haselwandter, K. (1981). Observation on the mycorrhizal status of some alpine plant communities. New phytolologist. 88, 341-352. http://dx.doi.org/10.1111/j.1469-8137.1981.tb01729.x
Redman, R. S. Sheehan, K. B. Stout, R. G. Rodriguez, R. J., & Henson, J. M. (2002). Thermotolerance conferred to plant host and fungal endophyte during mutualistic symbiosis. Science. 298 (5598). 1581.
Reininger, V., & Sieber, T. N. (2012). Mycorrhizza reduces adverse effects of dark septate endophyte (DSE) on growth of conifers. Plos One. 7 (8): e42865. http://dx.doi.org/10.1371/journal.pone.0042865

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 153
Rodriguez, R. J. Henson, J. Van Volkenburgh, E. Hoy, M. Wright, L. Beckwith, F. Kim, Y., & Redman, R. S. (2008). Stress tolerance in plants via habitat-adapted symbiosis. International Society of Microbial Ecology. 2, 404-416.
Rodriguez, R. J. Redman, R. S., & Henson, J. M. (2004). The role of fungal symbioses in the adaptation of plants to high stress environments. Mitigation and Adaptation Strategies for Global Change. 9, 261-272. http://dx.doi.org/10.1023/B:MITI.0000029922.31110.97
Rodriguez, R., & Redman, R. (2008). More than 400 million years of evolution and some plants still can’t make it on their own: plant stress tolerance via fungal symbiosis. Journal of Experimental Botany. 59, 1109-1114. http://dx.doi.org/10.1093/jxb/erm342
Rubini, M. R. Silva-Ribeiro, R. T. Pomella, A. W. V. Maki, C. S. Arau ́ Jo, W. L. Dos Santos, D. R., & Azevedo, J. L. (2005). Diversity of endophytic fungal community of cacao (Theobroma cacao L.) and biological control of Crinipellis perniciosa, causal agent of Witches’ Broom Disease. International Journal of Biological Sciences. 1, 24-33. http://dx.doi.org/10.7150/ijbs.1.24
Ruiz-Lozano, J. M., & Azcon, R. (2000). Symbiotic efficiency and infectivity of an autochthonous arbuscular mycorrhizal Glomus sp. from saline soils and Glomus deserticola under salinity. Mycorrhiza. 10, 137-143. http://dx.doi.org/10.1007/s005720000075
Rukachaisirikul, V. Sommart, U. Phongpaichit, S. Sakayaroj, J., & Kirtikara, K. (2008). Metabolites from the endophytic fungus Phomopsis sp. PSU-D15. Phytochemistry. 69(3), 783-787. http://dx.doi.org/10.1016/j.phytochem.2007.09.006
Saharan, B. S., & Nehra, V. (2011). Plant growth promoting Rhizobateria: A critical review. Life science and medicine research. 21, 1-30.
Saikkonen, K. Faeth, S. H. Helander, M., & Sullivan, T. J. (1998). Fungal endophytes: A continuum of interactions with host plants. Annual Review of Ecology and Systematics. 29, 319-343. http://dx.doi.org/10.1146/annurev.ecolsys.29.1.319
Saikkonen, K. Ion, D., & Gyllenberg, M. (2002). The persistence of vertically transmitted fungi in grass metapopulations. Proceedings of the Royal Society B: Biological Sciences. 269, 1397-1403. http://dx.doi.org/10.1098/rspb.2002.2006
Schulz, B. (2006). Mutualistic interactions with fungal root endophytes. In: B. Schulz, C. Boyle, T. Sieber (Eds.), Microbial root endophytes (pp. 261-280). Berlin, Germany: Springer-Verlag. http://dx.doi.org/10.1007/3-540-33526-9_15
Schulz, B. Boyle, C. Draeger, S. Römmert, A. K., & Krohn, K. (2002). Endophytic fungi: a source of biologically active secondary metabolites. Mycological Research. 106, 996-1004. http://dx.doi.org/10.1017/S0953756202006342
Schulz, B. Rommert, A. K. Dammann, U. Aust, H. J., & Strack, D. (1999). The endophyte-host interaction: a balanced antagonism. Mycological Research. 10, 1275-1283. http://dx.doi.org/10.1017/S0953756299008540

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 154
Schulz, B. Sucker, J. Aust, H. J. Krohn, K. Ludewig, K., & Jones, P. G. (1995). Biologically active secondary metabolites of endophytic Pezicula species. Mycological Research. 99, 1007-1015. http://dx.doi.org/10.1016/S0953-7562(09)80766-1
Schulz, B., & Boyle, C. (2005). The endophytic continuum. Mycological Research. 109, 661-686. http://dx.doi.org/10.1017/S095375620500273X
Selosse, M. A. Dubois, M. P., & Alvarez, N. (2009). Do Sebacinales commonly associate with plant roots as endophytes. Mycological Research. 113, 1062-1069. http://dx.doi.org/10.1016/j.mycres.2009.07.004
Serfling, A. Wirsel, S. G. R. Lind, V., & Deising, H. B. (2007). Performance of the biocontrol fungus Piriformospora indica on wheat under greenhouse and field conditions. Phytopathology. 97, 523-531. http://dx.doi.org/10.1094/PHYTO-97-4-0523
Sheng, M. Tang, M. Chan, H. Yang, B. Zhang, F., & Huang, Y. (2008). Influence of arbuscular mycorrhizae on photosynthesis and water status of maize plants under salt stress. Mycorrhiza. 18, 287-296. http://dx.doi.org/10.1007/s00572-008-0180-7
Shin, D. S. Oh, M. N. Yang, H. C., & Oh, K. B. (2005). Biological characterization of periconicins, bioactive secondary metabolites, produced by Periconia sp. OBW-115. Journal of Microbiology and Biotechnology. 15, 216-220.
Shoebitz, M. Ribaudo, C. M. Pardo, M. A. Cantore, M. L. Ciampi, L., & Curá, J. A. (2009). Plant growth promoting properties of a strain of Enterobacter ludwigii isolated from Lolium perenne rhizosphere. Soil Biologyand Biochemistry. 41, 1768-1774. http://dx.doi.org/10.1016/j.soilbio.2007.12.031
Srivastava, S. Yadav, A. Seem, K. Mishra, S. Chaudhary, V., & Srivastava, C. S. (2008). Effect of high temperature on Pseudomonas putida NBRI0987 biofilm formation and expression of stress sigma factor RpoS. Current Microbiology. 56(4), 453-457. http://dx.doi.org/10.1007/s00284-008-9105-0
Strobel, G. A. Dirksie, E. Sears, J., & Markworth, C. (2001). Volatile antimicrobials from Muscodor albus, a novel endophytic fungus. Microbiology. 147, 2943-2950.
Strobel, G. A. Kluck, K. Hess, W. M. Sears, J. Ezra, D., & Vargas, P. N. (2007). Muscodor albus E-6, an endophyte of Guazuma ulmifolia, making volatile antibiotics: isolation, characterization and experimental establishment in the host plant. Microbiology. 153, 2613-2620. http://dx.doi.org/10.1099/mic.0.2007/008912-0
Strobel, G. A. Stierle, A. Stierle, D., & Hess, W. M. (1993). Taxomyces andreanae a proposed new taxon for a bulbilliferous hyphomycete associated with Pacific yew. Mycotaxon. 47, 71-78.
Strobel, G. Knighton, B. Kluck, K. Ren, Y. Livinghouse, T. Griffen, M. Spakowicz, D., & Sears, J. (2008). The production of myco- diesel hydrocarbons and their derivatives by the endophytic fungus Gliocladium roseum. Microbiology. 154, 3319-3328. http://dx.doi.org/10.1099/mic.0.2008/022186-0

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 155
Strobel, G. Yang, X. S. Sears, J. Kramer, R. Sidhu, R. S., & Hess, W. M. (1996). Taxol from Pestalotiopsis microspora, an endophytic fungus of Taxus wallachiana. Microbiology. 142, 435-440. http://dx.doi.org/10.1099/13500872-142-2-435
Supothina, S. Isaka, M., & Wongsa, P. (2007). Optimization of culture conditions for production of the anti-tubercular alkaloid hirsutellone A by Trichoderma gelatinosum BCC 7579. Letters in Applied Microbiology. 44(5), 531- 537. http://dx.doi.org/10.1111/j.1472-765X.2006.02098.x
Tan, R. X., & Zhou, W. X. (2001). Endophytes: a rich source of functional metabolites. Natural Product Reports. 18, 448-459. http://dx.doi.org/10.1039/b100918o
Taylor, D. L. Bruns, T. D. Szaro, T. M., & Hodges, S. A. (2003). Microbial Endophytes of Orchid Roots. In: B. Schulz, C. Boyle, T. Sieber (Eds.), Microbial Root Endophytes (pp. 153-177). Springer Berlin Heidelberg.
Tudzynski, B., & Sharon, A. (2002). Biosynthesis, biological role and application of fungal phyto-hormones. In: H.D. Osiewacz (Ed) The mycota X. Industrial applications. (pp. 183-212). Berlin, Germany: Springer-Verlag. http://dx.doi.org/10.1007/978-3-662-10378-4_9
Unterseher, M., & Schnittler, M. (2010). Species richness analysis and ITS rDNA phylogeny revealed the majority of cultivable foliar endophytes from beech (Fagus sylvatica). Fungal Ecology. 3, 366-378. http://dx.doi.org/10.1016/j.funeco.2010.03.001
Upson, R. Read, D. J., & Newsham, K. K. (2009). Nitrogen form influences the response of Deschampsia antarctica to dark septate root endophytes. Mycorrhiza. 20(1), 1-11. http://dx.doi.org/10.1007/s00572-009-0260-3
Varma, A. Sherameti, I. Tripathi, S., & Prasad, R. (2012). The symbiotic fungus Piriformospora indica: Review. In: B. Hock (Ed.), Fungal Associations, 2nd edn, The Mycota IX (pp. 231-254). Springer-Verlag Berlin Heidelberg.
Varma, A. Verma, S. Sudha, Sahay, N. Butehorn, B., & Franken, P. (1999). Piriformospora indica, a cultivable plant-growth-promoting root endophyte. Applied Environmental Microbiology. 65, 2741-2744.
Verma, S. Varma, A. Rexer, K. H. Hassel, A. Kost, G. Sarbhoy, A. Bisen, P. Butehorn, & B. Franken, P. (1998). Piriformospora indica, gen. et sp. nov., a new root-colonizing fungus. Mycologia. 90, 896-903. http://dx.doi.org/10.2307/3761331
Vohnik, M. Albrechtova, J., & Vosatka, M. (2005). The inoculation with Oidiodendron maius and Phialocephala fortinii alters phosphorus and nitrogen uptake, foliar C: N ratio and root biomass distribution in Rhododendron cv. Azurro. Symbiosis. 40(2), 87-96.
Vu, T. Hauschild, R., & Sikora, R. A. (2006). Fusarium oxysporum endophytes induced systemic resistance against Radopholus similis on banana. Nematology. 8, 847-852. http://dx.doi.org/10.1163/156854106779799259
Vujanovic, V. St-Arnaud, M. Barabe, D., & Thibeautl, G. (2000). Microbial Endophytes of

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 156
Orchid Roots. In: B. Schulz, C. Boyle, T. Sieber (Eds.), Microbial Root Endophytes, (pp. 153-177). Springer Berlin Heidelberg.
Wakelin, S. A. Warren, R. A. Harvey, P. R., & Ryder, M. H. (2004). Phosphate solubilization by Penicillium spp. closely associated with wheat roots. Biology and Fertility of Soils. 40, 36-43. http://dx.doi.org/10.1007/s00374-004-0750-6
Waller, F. Achatz, B. Baltruschat, H. Fodor, J. Becker, K. Fischer, M. Heier, T. Huckelhoven, R. Neumann, C. Von Wettstein, D. Franken, P., & Kogel, K. H. (2005). The endophytic fungus Piriformospora indica reprograms barley to salt-stress tolerance, disease resistance, and higher yield. Proceedings of the National Academy of Sciences, USA. 102(38), 13386-13391. http://dx.doi.org/10.1073/pnas.0504423102
Wang, C. J. K., & Wilcox, H. E. (1985). New species of ectoendomycorrhizal and pseudomycorrhizal fungi: Phialophora finlandia, chloridium paucisporum and Phialocephala fortinii. Mycologia. 77, 951-958. http://dx.doi.org/10.2307/3793308
Wang, W. Vinocur, B., & Altman, A. (2003). Plant responses to drought, salinity and extreme temperatures: toward genetic engineering for stress tolerance. Planta. 218, 1-14. http://dx.doi.org/10.1007/s00425-003-1105-5
Wang, W. X. Barak, T. Vinocur, B. Shoseyov, O., & Altman, A. (2003). Abiotic resistance and chaperones: possible physiological role of SP1, a stable and stabilizing protein from Populus. In: I.K. Vasil (Ed.), Plant biotechnology 2000 and beyond (pp. 439-443). Dordrecht: Kluwer
Weber, R.W. Stenger, E. Meffert, A., & Hahn, M. (2004b). Brefeldin A production by Phoma medicaginis in dead pre- colonized plant tissue: a strategy for habitat conquest. Mycological Research. 108, 662-671. http://dx.doi.org/10.1017/S0953756204000243
Weishampel, P., & Bedford, B. (2006). Wetland dicots and monocots differ in colonization by arbuscular mycorrhizal fungi and dark septateendophytes. Mycorrhiza. 16, 495-502. http://dx.doi.org/10.1007/s00572-006-0064-7
West, C. P. (1994). Physiology and drought tolerance of endophyte infected grasses. In: C.W. Bacon, & J.F. White (Eds.), Biotechnology of endophytic fungi of grasses (pp. 87-99). CRC Press, Boca Raton FL.
Whipps, J. M. (2001). Microbial Interactions and Biocontrol in the rhizosphere. Journal of Experimental Botany. 52, 487-511. http://dx.doi.org/10.1093/jexbot/52.suppl_1.487
White, J. F., & Torres, M. S. (2010). Is plant endophyte-mediated defensive mutualism the result of oxidative stress protection. Physiology Plant. 138, 440-446. http://dx.doi.org/10.1111/j.1399-3054.2009.01332.x
Wilcox, H. E., & Wang, C. J. K. (1987b). Mycorrhizal and pathological associations of dematiaceous fungi in roots of 7-month-old tree seedlings. Candian Journal Forest Research. 17, 884-889. http://dx.doi.org/10.1139/x87-140
Wilson, D. (1995). Endophyte- the evolution of a term and clarification of its use and definition.

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 157
Okios. 73, 274-276. http://dx.doi.org/10.2307/3545919
Woropong, J. Strobel, G. A. Ford, E. J. Li, J. Y. Baird, G., & Hess, W. M. (2001). Muscodor albus anam.nov., an endophyte from Cinnamomum zeylanicum. Mycotaxon. 79, 6 7 - 79 .
Wu, Q. S., & Xia, R. X. (2006). Arbuscular mycorrhizal fungi influence growth, osmotic adjustment and photosynthesis of citrus under well watered and water stress conditions. Journal of Plant Physiology. 163(4), 417-425. http://dx.doi.org/10.1016/j.jplph.2005.04.024
Wu, S. C. Cao, Z. H. Li, Z. G. Cheung, K. C., & Wong, M. H. (2005). Effects of biofertilizer containing N- fixer, P and K solublizers and AM fungi on maize growth: a greenhouse trial. Geoderma. 125, 155-166. http://dx.doi.org/10.1016/j.geoderma.2004.07.003
Xiao-Yan, S. Qing-Tao, S. Shu-Tao, X. Xiu-Lan, C. Cai-Yun, S., & Yu-Zhong, Z. (2006). Broad-spectrum antimicrobial activity and high stability of Trichokonins from Trichoderma koningii SMF2 against plant pathogens. FEMS Microbiology. 260(1), 119-125. http://dx.doi.org/10.1111/j.1574-6968.2006.00316.x
Xing, X. K., & Guo, S. X. (2011). Fungal endophyte communities in four Rhizophoraceae mangrove species on the south coast of China. Ecological Research. 26(2), 403-409. http://dx.doi.org/10.1007/s11284-010-0795-y
Yamato, M. Ikeda, S., & Iwase, K. (2008). Community of arbuscular mycorrhizal fungi in coastal vegetation on Okinawa Island and effect of the isolated fungi on growth of sorghum under salt-treated conditions. Mycorrhiza. 18, 241-249. http://dx.doi.org/10.1007/s00572-008-0177-2
Yu, T. Nassuth, A., & Petersen, R. L. (2001). Characterization of the interaction between the dark septate fungus Phialocephala fortinii and Asparagus officinalis roots. Canadian Journal Microbiology. 47, 741-753. http://dx.doi.org/10.1139/cjm-47-8-741
Zhang, H. W. Song, C. Y., & Tan, R. X. (2006). Biology and chemistry of endophytes. Natural Products Report. 23, 753-771. http://dx.doi.org/10.1039/b609472b
Zikmundová, M. Drandarov, K. Bigler, L. Hesse, M., & Werner, C. (2002). Biotransformation of 2 benzoxazolinone and 2- hydroxy-1,4-benzoxazin-3-one by endophytic fungi isolated from Aphe- landra tetragona. Applied and Environmental Microbiology. 68, 4863-4870. http://dx.doi.org/10.1128/AEM.68.10.4863-4870.2002
Zou, W. X., & Tan, R. X. (1999). Advances in plant science. Beijing: China Higher Education Press. 2. 183-190.
Zuccaro, A. Basiewicz, M. Zurawska, M. Biedenkopf, D., & Kogel, K. H. (2009). Karyotype analysis, genome organization, and stable genetic transformation of the root colonizing fungus Piriformospora indica. Fungal Genetics and Biology. 46, 542-550. http://dx.doi.org/10.1016/j.fgb.2009.03.009
Zuccaro, A. Lahramann, U. Idener, U. Langen, G. Pfiffi, S. Biedenkop, D. Wong, P. Samans, B. Grimm, C. Basiewicz, M. Murat, C. Martin, F., & Kogel, K. H. (2011). Endophytic life

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 158
strategies decoded by genome and transcriptome analyses of the mutualistic root symbiont Piriformospora indica. PLoS Pathogens 7:e1002290. http://dx.doi.org/10.1371/journal.ppat.1002290
Copyright Disclaimer
Copyright for this article is retained by the author(s), with first publication rights granted to the journal.
This is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 159
Methanol Extract of Barringtonia asiatica, Kurz. Quite Effective Induce Mortality of Spodopthera litura, Fabr.
Larvae on Soybean Plants
Meity N Tanor
State University of Manado, Manado, Indonesia
Abdul Latief Abadi, Bambang Tri Raharjo
Plant Pest and Disease Major, Faculty of agriculture, Universitas Brawijaya, Malang, Indonesia
Jantje Pelealu
Faculty of agriculture, Samratulangi University, Manado, Indonesia
Received: May 6, 2014 Accepted: May 20, 2014
doi:10.5296/jbls.v5i2.5587 URL: http://dx.doi.org/10.5296/jbls.v5i2.5587
Abstract
This study aim was to examine the effectiveness of methanol extract derived from keben seed (B. asiatica) on mortality of Spodopthera litura larvae on soybean plant. The instar II larvae of S. litura were treated by the methanol extract with various concentrations i.e. 0.0 ppm (Control), 0.2 ppm, 0.4 ppm, 0.8 ppm and 1.0 ppm using pre-investment and post-investment. The mortality of larvae S. litura was counted after 24-hours of the treatment. The B. asiatica seed extract was quite effective in suppressing the intensity of damage in leaves caused by the attack of S. litura F. on soybean plants. The finding on pre-investment showed that after 24-hour treatment of methanol extract showed to have a significant impact on the mortality of S. litura instar II. The methanol extract dose 0.2 ppm, 0.4 ppm, 0.8 ppm, and 1.0 ppm had all been able to affect larvae mortality of S. litura there are 33.33%, 48.89%, 66.67%, and 88.89% respectively. Moreover, post-investment experiment showed that the methanol extract also induced mortality of S. litura in all doses, i.e 0.2 ppm, 0.4 ppm, 0.8 ppm and 1.0 ppm caused 44.44% (53.33%) ( 55.56%), (71.11%) mortality, respectively.

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 160
Keywords: Methanol extract, B. Asiatica seed, Mortality, Larvae S. litura
1. Introduction
Spodopthera litura F. belongs to Lepidoptera ordo, and is one of the pests that cause serious damage to crops in the tropics and sub-tropics (Haryanti et al., 2006). To control this pest, farmers use chemical insecticides intensively (with high frequency and dose). The situation resulted in adverse impacts such as symptoms of resistance, pest resurgence, death of natural enemies of pests, increased residue on crops produced, pollution on the environment, and increased health risks for users (consumers of the crops) (Haryanti et al., 2006).
Reduction in the use of pesticides could be done by utilizing natural enemies and botanical pesticides for environmentally sustainable form of agriculture (Grace and Yuyun, 2006; Samsudin, 2008). The use of natural insecticides derived from plant extracts has been proven to be safer because they have shorter lifespan residue. After the application, natural insecticides will break down into compounds that may not toxic to humans and to the environment (Desi 2007; Syahputra, 2001).
There are more than 1000 species of plants that can be utilized as natural insecticides, antifeedant substances, repellent substances, acaricides and growth retardants (Susetyo et al., 2008). The major advantage is that natural pesticides can easily degrade. Aided by the natural components, such as sunlight, air and moisture, the degradation process of these natural insecticides happen quickly in nature. Thus, natural pesticides sprayed few days before harvesting time will not leave any residue (Sukrasno, 2003). One of the plants that have the potential to be developed as a natural pesticide is Barringtonia asiatica (L) Kurz or in Indonesia is better known as Bitung (Ecology & Evolutionary Biology Greenhouse (EEBG), 2006). The seeds of the plant can be directly used as a natural pesticide (Kardinan, 2005). The methanol extract of derived from the seed of B. asiatica is toxic to larvae of Crocidolomia pavonana F. (Lepidoptera: Pyralidae) (Dono and Sujana, 2007) and to larvae of Epilachna sp (Herlt et al., 2002). The active compounds found in Bitung include saponins, gallic acid, hydrocyanic acid, and triterpenoids consisting of bartogenat acid, 19-epibartogenat acid, and acid-bartogenat anhidro.
Formulation of B. asiatica seed extract is also safe to apply to plants such as cabbage, broccoli, and cauliflower at the age of five weeks. The best results of B. asiatica seed extract formulation are 30 WP formulation with 100% mortality against C. pavonana larvae on day 5 (Danar and Abdurrohim, 2008). From the afore-presented descriptions, the researchers feel compelled to do this study in order to determine the effectiveness of methanol seed extract (B. asiatica) on mortality of S. litura larvae.
2. Methods
Seed extraction of B. asiatica
The experiment was conducted from February 2012 to January 2013 in the Laboratory of Biology Department, the State University of Manado and the Center for Research and Study of Plants Tomohon in North Sulawesi. B. asiatica seeds were obtained from Malalayang Coastal

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 161
areas of Manado. The seeds were cut into parts, and the cutted-seed was weight until 6350 g and dried. The dried seeds were blended to form of B. asiatica seed flour, as much as 2800 g. The dried sample (450 g) was macerated with methanol for 3x24 hours with occasional stirring, then filtered. The filtrate was concentrated with a rotary evaporator. The total result was 50g crude extract in semi-solid form of a brownish-white color and oily matter.
Treatment of Methanol Extract Derived From B.asiatica Toward S. litura Larvae Instar II
As many as 30 polybags of soybean plants, aged three weeks, were divided into two groups, each group consisted of 15 polybags; one group was for pre-investment treatment, and the other group was for the post-investment treatment. Each treatment, both pre- investment and post-investment, was divided into five dose treatment groups (0.0 ppm, 0.2 ppm, 0.4 ppm, 0.8 ppm and 1.0 ppm of B. asiatica methanol seed extract) with 3 duplicates. Pre-investment treatment was done by evenly spraying methanol extract to soybeans, and 15 minutes later the instar II larvae of S. litura were laid on the leaves of the plant, and subsequent observation was carried out. While the treatment of post-investment was made by giving extra squirt of the methanol extract on the leaves of soybean plants on which the instar II larvae of S. litura were laid out. After 24 hours, the number of edible leaves and mortality rate was observed. Larvae mortality was calculated by using the formula developed by Abbott (1925) in Prijono (1999).
3. Results and Discussion
The Mortality of S. litura Larvae on Treatment Models of Pre-Investment and Post-Investment
The results of the analysis of variance showed that the methanol extract of B. asiatica seeds at various concentration levels significantly affected the mortality of S. litura larvae (Table 1). The results of this study indicate that the highest mortality of S. litura larvae was 88.89% for the pre-investment and 71.11% for post-investment in the treatment level of methanol extract of B. asiatica seeds at the concentration of 1.0 ppm (K4). The higher the concentration of B. asiatica seed extract applied on, the higher the mortality of S. litura larvae was. It is proven that the higher the concentration level of a compound, the more active ingredients it contains, thus the more effective the killing powers it has.
Table 1. The Mortality Rate of S. litura F. Larvae Due to Different Concentration Levels of The Methanol Extract of B. asiatica Seeds on Treatment Models of Pre-Investment and Post-Investment
No
Treatment
Average Mortality (%)
Pre-Investment Post-Investment
1 2 3 4 5
K0 K1 K2 K3 K4
0.67 a 33.33 b 48.89 c 66.67d 88.89e
4.47 a 44.44 b 53.33 c 55.56 d 71.11 e

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 162
Duncan 0.05 1.000 0.138
Note: Figures followed by the same numbers are not significantly different with α 0.05 on BNT. (Duncan)
Furthermore, we analyzed the antifeedant ability of the extract by counting the number of leaves eaten by larvae before dying. The results indicate that the number of leaves eaten by larvae in the treatment group (sprayed with methanol extract of B. asiatica seed) was less than the number of leaves eaten by larvae in the control group (Table 2). It is proven that the extract has antifeedant properties to inhibit the appetite of S. litura larvae to eat the leaves. Appetite reduction is thought to be the cause of death of the observed larvae.
Table 2. Percentage of Leaves Eaten by S. litura Larvae on Day 3 Observation
Treatment
The average number of leaves eaten (%) Pre-Investment Post-Investment
KO K1 K2 K3 K4
58.33 41.67 33.33 19.42 13.89
42.86 38.09 30.95 38.09 33.29
Treatment of B. asiatica methanol extract at a concentration of 0.2 ppm (K1) on pre-investment made instar II larvae of S. litura consume the soybean leaves by 41.67 % on day 3 after treatment, whereas, in treatment 1.0 ppm (K4), the larvae could only eat the soybean leaves by 13.89%. However, the methanol extract could reduce the ability of S. litura larvae in eating soybean leaves compared with the control group (58.33%). These findings are also in line with the research findings Rusdy (2009) which state that the concentration of seed and neem leaf extracts is quite effective in suppressing the intensity of leaf damage caused by the pest of S. litura F. on lettuce plants. The pesticide compounds in the seeds of B. asiatica plant have been known to cause the death of S. litura, and the findings of the present study confirm that the death rate of S. litura depends on the concentration of B. asiatica extract. These results are also in line with the opinion of Yuyun and Grace (2006), in which the analysis of variance in their study show that the different concentration of seed extract and neem leaf extract used against S. litura F. on lettuce plants results in a very noticeable difference on mortality rates. Similarly, the results of the study presented by Makal and Turang (2011) also show that mortality rate of C. binotalis larvae increased with the increasing concentrations of lemongrass extracts applied.
Nursal and Etti (2005) state that steroids are found in various plants and, together with other secondary substances, act as a defense against attacks by insects. Saponins contained in food consumed by insects can decrease the activity of digestive enzymes and absorption of food, while terpenoids and flavonoids are inhibit eating and are toxic for insects. If insects take the active compound and are not resistant, they will experience death; yet, on the other hand, tolerant insects will survive to the next stadia of pupa or imago. For insects that are not resistant to the active compound, before they finally die, they will survive by maximizing the utilization of energy sources in their body. As a consequence, larvae will experience barriers to growth and development. Insecticides derived from B. asiatica seed extract need to be developed, since its active ingredients do not cause phytotoxic symptoms. Active compounds contained in

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 163
methanol extract of B. asiatica seeds are relatively safe on plants due to non-phytotoxic symptoms (Dono and Sujana, 2007).
4. Conclusion
The mortality of S. litura F. larvae is determined by the level of concentration of B. asiatica seed extract tested, the highest mortality percentage was 57.77% obtained at 1.0 ppm for the treatment of pre-investment, and for post-investment the highest mortality was 71.11% at 1.0 ppm treatment. The concentration of B. asiatica seed extract was quite effective in suppressing the intensity of damage in leaves caused by the attack of S. litura F. on soybean plants. Further research is needed to examine the active compounds contained in B. asiatica seed extract that leads to antifeedant effects and mortality in S. litura larvae.
References
Danar, D., & Abdurrohim V. (2008). Selektivitas Formulasi Ekstrak Biji Barringtonia asiatica L. (Lecythidaceae) Terhadap Ulat Krop Kubis Crocidolomia pavonana (F) dan Parasitoid Telur Trichogramma spp. serta Dampaknya Terhadap Bibit Tanaman Brassicaceae. Prosiding Seminar Nasional Pekan Kentang. Lembang 20 s.d 21 Agustus 2008.
Desi, A. (2007). Pemanfaatan biji bengkuang Sebagai Insektisida Alami. http//www. Pkm.dikti. net/pkmi award 2006/pdf/pkmi 06 068.pdf.
Anonymous. (2006). Ecology and Evolutionary Biology Greenhouse: Barringtonia asiatica, Kurz. University of Connecticut. http://florawww.eeb.uconn.edu.
Haryanti, S. M. Suryana & Nurrahmad. (2006). Uji Daya Insektisida Ekstrak Etanol 70 % Biji Buah Mahkota Dewa Terhadap Ulat Grayak (Spodoptera litura Fab.) Instar II. http://www.litbang.depkes.go.id/risbinkes.
Herlt, A. J, L. N. Mander, E. Pongoh, R. J. Rumampuk & P. Tarigan. (2002). Two Major Saponins from Seeds of Barringtonia asiatica: Putative Antifeedants toward Epilachna sp. Larvaee. Journal of Natural Products, 65, 115-120. http://dx.doi.org/10.1021/np000600b
Makal, H. V. G, & Turang, D. A. S. (2011). Pemanfaatan Ekstrak Kasar Batang Serai Untuk Pengendalian Larvae Crosidolomia binotalis Zell pada Tanaman Kubis. Eugenia, Volume 17 No. 1 April 2011
Mordue (Luntz), A. J., M. S. J. Simmonds, S. V. Ley, W. M. Blaney, W. Mordue, M. Nasiruddin & A. J. Nisbet. (1998). Actions of azadirachtin, a plant allelochemical, against insects. Pestic. Sci., 54, 277-284. http://dx.doi.org/10.1002/(SICI)1096-9063(1998110)54:3<277::AID-PS801>3.3.CO;2-9
Nursal & Etti, S. S. (2005). Kandungan Senyawa Ekstrak Lengkuas (Loctuca Indica L.), Toksisitas dan Pengaruh Subletalnya Terhadap Mortalitas Larvae Nyamuk Aedes Aegypti L. http://library. usu. ac.id/download/fmipa/06000449.pdf.
Prijono, D. (1999). Prospek dan Strategi Pemanfaatan Insektisida Alami dalam PHT hal. 1-7

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 164
dalam Bahan Pelatihan Pengembangan dan Pemanfaatan Insektisida Alami. Pusat Kajian Pengendalian Hama Terpadu. Institut Pertanian Bogor.
Rahmat Rukmana, Yuyun Nursal & Etti, S. S. (2005). Kandungan Senyawa Ekstrak Lengkuas (Loctuca Indica L.), Toksisitas dan Pengaruh Subletalnya Terhadap Mortalitas Larvae Nyamuk Aedes Aegypti L. Kanisius, Yogyakarta.
Rahmat Rukmana and Yuyun Yuniarsih Oesman. (2006). Nimba Tanaman Penghasil Pestisida Alami. Kanisius, Yogyakarta.
Rusdy, A. (2009). Efektivitas Ekstrak Nimba Dalam Pengendalian Ulat Grayak (Spodoptera litura F.) Pada Tanaman Selada. J. Floratek, 4, 41-54
Samsudin. (2008). Virus Patogen Serangga: Bio–Insektisida Ramah Lingkungan, http://www.pertaniansehat.or.id.
Sukrasno. (2003). Mimba Tanaman Obat Multi Fungsi, Agromedia Pustaka. Indonesia
Susetyo, T. Ruswandi & Etty Purwanti. (2008). Teknologi Pengendalian Organisme Pengganggu Tumbuhan (OPT) Ramah Lingkungan, Direktorat Perlindungan Tanaman Pangan Jakarta.
Syahputra, E. (2001). Hutan Kalbar Sumber Pestisida Botani: Dulu Kini Dan Kelak, http// tumouto. Net/3 semi. 12 edy saputra. Html.
Copyright Disclaimer
Copyright for this article is retained by the author(s), with first publication rights granted to the journal.
This is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 165
Environmental Factors and Puberty Onset: An Update
Ghulam Nabi (Corresponding author)
M. Phil Scholar. Department: Animal sciences
Reproductive neuro-endocrinology Lab: Quaid-i-Azam University, (Islamabad) Pakistan
Tel: 92-345-811-2741 E-mail: [email protected]
Muhammad Amin
M. Phil Scholar, Department of Zoology, University of Karachi, Pakistan
Tel: 92-315-902-0833 E-mail: [email protected]
Rishma Sultan
M.Phil. Scholar, Department of Zoology
Islamia College University Peshawar, Khyber Pakhtunkhwa, Pakistan
E-mail: [email protected]
Muhammad Kamil
M. Phil Scholar, Department of Botany, University of Malakand, Pakistan
Tel: 92-343-925-6626 E-mail: [email protected].
Received: July 4, 2014 Accepted: July 20, 2014
doi:10.5296/jbls.v5i2.5911 URL: http://dx.doi.org/10.5296/jbls.v5i2.5911
Abstract
Puberty is a process that results from a coordinated series of complex neuroendocrine changes which eventually leads to external and internal physical changes. These changes include attainment of primary and secondary sexual characteristics. Puberty as a temporal process is subdivided into different stages. These pubertal stages occur in a sequential manner but significant variations have been seen among different individuals in the timings of pubertal onset as well as a certain degree of gender difference is also present where puberty is occurring

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 166
earlier in girls than boys. Among many aetiologies which have been hypothesized to be involved, environmental cues are the most ones. This review article describes the recent findings of the influences of the key environmental clues on pubertal timings, emphasizing on the aspects for which the most scientific evidences exist.
Keywords: Puberty, Temporal, Neuroendocrine
1. Introduction
Sexual maturation in humans is physiological variation of pubertal age. Sexual maturation in normal individuals dwelling in the same area is affected by genetics, ethnic, environmental and nutritional factors (Gaudineau et al., 2010). Puberty onset is initiated by activation of hypothalamic GnRH neurons which secrete pulses of gonadotropin-releasing hormone (GnRH). This activation of GnRH pulses is also known as GnRH pulse generator activity and stimulates the secretion of pituitary gonadotropic hormone downstream. Pituitary gonadotropins acts on gonads and activate the process of steroid genesis and subsequent maturation of gametes (Mayer et al., 2010). Average age of puberty onset has been declined over the 20th and early 21st century especially in populations that are characterized by improvements in socioeconomics or social conditions (Boynton-Jarrett and Harville, 2010).
Moreover, many studies showed marked differences in pubertal timing across different ethnic groups. Varieties of genetic and environmental factors have been found to influence timing of puberty. Environmental stressors and supports may accelerate or suppress the HPG axis by influencing the biological stress responses to early adversities (Boynton-Jarrett and Harville, 2010). Environmental conditions that strongly influence the timing of puberty onset include (1) photoperiod (2) nutrition (3) psychosocial factors and (4) endocrine disruptors (Deardroff et al., 2011).
2. Psychosocial Factors
Exposure to a number of psychosocial stressors during pre-pubertal period such as (1) immigration (2) neighborhood environment (3) father absence (4) socioeconomic status (5) childhood social hardships and (6) food insecurity, alter the timing of puberty onset. Dramatic changes in socio-economic context are also involved in altering the timings of puberty onset in children (Tan and Camras, 2013).
2.1 Immigration
Immigration is reviewed as radical socio-cultural alteration for children. In immigration nutritional upshot and quick weight growth following adoption may cause premature puberty onset. Studies found the ubiquity of precocious puberty by the appearance of breast development slightly earlier in adopted Chinese girls as compared to non-adopted American girls (Tan and Camras, 2013).
2.2 Neighbourhood Environment
It also effect puberty onset by either “reduced physical activity or reduced access to healthful foods” or through exposure to chronic and acute stressful familial environment. These factors trigger puberty by influencing hormonal pathways. More recreational outlets in neighbourhood

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 167
delayed timing of puberty onset in study area. Physical activity affects puberty timings by influencing adiposity and body weight (Deardorff et al., 2012).
2.3 Father Absence and Socioeconomic Status
It influences pubertal initiation among adolescents. Recently, studies have associated early onset of puberty to both lower household income and lower parental education. There is strong association between early menarche and high socio-economic status among black girls and vice versa in white girls. However parental education is not significant indicator of early menarche (Braitchwaite et al., 2009). Another studies found that girls with absent biological father from high SES exhibited precocious puberty than those having biological fathers in their homes. In the case of low income families, there is no link between early menarche and father absence (Deardroff et al., 2011; Belachew et al., 2011; James-Todd et al., 2010; Al-Sahab et al., 2010). The possible cause of precocious puberty may possibly be the familial stress due to maternal work load and diminution in maternal affection. Another reason might be the acquaintance to high level of endocrine disrupters (i-e beauty products) or technology (video games, internet) that increases disclosure to light and disturbs sleep-wake cycle which has been linked to early menarche. Another study reveals the relationship between early puberty onset and single parenting as a psychosocial among Mexican-American girls (Jean et al., 2010).
2.4 Childhood Social Hardships
These also play critical role in pubertal timings. (1)Stressful home circumstances (2) family conflicts (3) alterations in family structure (4) attachment relationships (5) child maltreatment (6) parental divorce (7) low parental involvement and (8) marital conflict , all lead to early menarche and precocious sexual maturation through chronic elevation of HPG and HPA axis.(Jean et al., 2010; Boynton-Jarrett et al.,2010) Physical abuse is weakly associated with early menarche while sexual abuse has strong association (Boynton-Jarrett et al., 2010). Overwhelmingly, we can say that children exposed to cumulative aversive condition would have altered timings of puberty. Vigorous physical activity is accompanying with decreased estrogen secretion and increased age at menarche due to disturbance of GnRH pulsatility (Belachew et al., 2011).
2.5 Food Insecurity
Studies found that stress and anxiety about food availability or insufficient access to energy, delays menarche by one year. Malnutrition and increased energy expenditure are also linked with delayed menarche. Age at menarche has declined over last two centuries in most part of the world due to improvement in nutrition and population health (Belachew et al., 2011).
Early family environment covaries with menarcheal age as a function of polymorphic variation in estrogen receptor-α (ESR1)
Studies reported in European-American women that girls reach to puberty at later age that belongs from healthy environment and cohesive families and in contrary those girls who belong from conflicted or divorced families reach to early menarche. This link is regulated by polymorphic variation in the gene encoding estrogen receptor-α (ESR1). Moreover correlated

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 168
variations of family environment positively regulated menarcheal age in those applicants homozygous for minor alleles (rs9304799, rs2234693), but not in women having different genotypes (Manuck et al., 2011). These findings are consistent with hypothesis of Belsky and Pluess, 2009 who found that girls differ genetically in their sensitivity to rearing effects on pubertal maturation.
3. Endocrine Disruptors and Puberty
Recent studies have demonstrated a progressive decrease in age of onset of puberty in children around the world. Perhaps, it is due to complex interaction between genetics, endocrine and environmental factors. The process of industrialization results into a gradual, but significant increase in number and amount of environmental pollutants. Some of these environmental pollutants are natural or synthetic chemicals with alarming effects on the endocrine system. The chemicals that affect the endocrine system are called endocrine disruptors (EDs). Various epidemiological studies on exposure of EDCs show variable tendency towards the puberty onset. In this regard, some EDCs advance puberty, some delay puberty, while others show no association.
To estimate the potential of EDCs which advances the puberty onset, following studies are noteworthy. Prepubertal administration of bisphenol A (BPA) at different doses significantly decreased the body weight from (PND) post natal day 18 to 30. Moreover, bisphenol advances the puberty timing significantly in treated group as compared to control group. BPA induce this effect through its estrogenic activity (Won et al., 2011). In cohort study, exposure of diethylstilbestrol (DES) during the first 15+ week in both low <2500mg and the high (10,000+mg) doses show the strong association with early menarche ≤10 years. While, the odd ratio for prenatal DES exposure in relation to menarche ≤10 years the Odd ratio was 1.41 and for menarche ≤11 years, the OR was 1.16 (Elizabeth et al., 2011). In an experiment of female Long Evans rat, pregnant females were exposed with different doses of ethinyl estradiol and BPA. Exposure of EE2 at different doses accelerated age at vaginal opening as compared to BPA had no effect on puberty onset. Similarly, tributylin chloride (TBTCl) exposure during gestational period significantly accelerated the age at vaginal opening as compared to control group. TBTCl advanced the age of onset of VO in this study, indicative of an estrogenic effect (Jiliang et al., 2012). In another cohort study, data from female participants 12–16 years of age for phthalate and parapens in (NHANES) were analyzed. It was found that 2, 5-dichlorophenol (2,5-DCP) and summed environmental phenols (2,5-DCP and 2,4-DCP) were inversely related with the age of menarche. While, Other exposures (total parabens, bisphenol A, triclosan, benzophenone-3, total phthalates, and 2, 4-DCP) were not significantly associated with age of menarche. This findings suggest an association between 2, 5-DCP, a potential EDC, and earlier age of menarche in the general U.S. population (Buttke et al ., 2012).
Endocrine disruptors may also delay puberty onset such as heavy metals like cadmium and lead is evidenced by following experiments. In a study , three groups of pregnant female rat during gestational to lactational period were exposed to demonstrate the effect of cadmium .It was found that vaginal opening every 24h from postnatal 22 onwards significantly delayed in dose dependent manner. Furthermore, cadmium also reduces binding sites for steroid hormones

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 169
(Jawahar et al., 2011). Similarly, the effect of blood lead levels on puberty onset demonstrated in cohort study of African population in which blood sample were taken to analyze the lead level from 712 girls who had the menarche data. This study found that lead even at low dose (5 μg/dl) blood levels may delay the onset of puberty among adolescent girls. While, higher blood lead levels were associated with significant delays in the onset of puberty (p<0.001) (Nisha et al., 2010). However, in some cases EDCs show no association with puberty onset or their effect depend upon the genotype of individuals. In survey report, the association between age at menarche, and exposure to polyflouroalkyl chemicals during pregnancy was investigated. Serum samples taken from the girls' mothers during pregnancy were analyzed for 8 Polyflouroalkyl among cases and control. All, but one PFC was detectable in most samples. Although, study participants had nearly ubiquitous exposure to most PFCs examined, but PFC exposure did not appear to be associated with altered age at menarche of their offspring (Krista et al., 2011).
Likewise, Sub chronic exposure to 2,000mg /kg and 8,000mg/kg Di(2-ethyl exyl )phthalate after 8 week reduce the maintenance of spermatogenesis in male mice and increase the rate of prenatal death. However, sub chronic administration does not influence vaginal opening (Małgorzata et al ., 2012). In another study, data of adolescent boys, was used association of perinatal dioxin exposure to genetic variability and subsequently their influence on puberty onset (testicular volume>3ml and G2 stage) were investigated. Potential candidates genes were selected for relevance to either the AhR molecular pathway or to pubertal onset. Primary finding was that three SNPs (two from glucocorticoid receptor gene, one from estrogen receptor –alpha) had gene environment interactions robust to correction for multiple comparisons. So, this study suggests that genotype of individual related to pubertal axis may play an important role in EDCs exposure (Olivier et al., 2013).
4. Nutrition
Successful puberty is, of course, important for an animal’s appropriate maturation and subsequent ability to reproduce. The onset of puberty appears to be closely associated with adiposity. For instance, sufficient levels of fat are essential for initiating puberty. The perinatal nutritional environment can permanently influence body weight, potentially leading to changes in puberty onset and reproductive function. Perinatal under- or overfeeding would alter puberty onset. Early life underfeeding in males and overfeeding in female rats significantly affects puberty onset, altering steroid hormone concentrations in males, but this is not related to changes in hypothalamic kisspeptin (Jeremy and Smith, 2012). In mid childhood the effects of nutrition on puberty onset has been addressed variously. Furthermore, both pre- and perinatal exposures have been identified as potential determinants of pubertal timing in a number of studies. There is an association between the magnitudes of dietary factors and the timing of puberty that are in the order of effect sizes observed for endocrine factors; for example, children with a more intensive adrenarchal process (i.e., in the highest group of adrenal androgen excretion) experience onset of breast and genital development 9 months earlier. Similarly, children with the highest intakes of vegetable proteins or animal protein experience at least their growth-related puberty onset up to 7 months later or 7 months earlier, respectively, than children in the lowest groups. Moreover, girls with the highest levels of dietary isoflavone

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 170
intake may experience their onset of breast development and reach their peak height velocity approximately 7–8 months later than girls with the lowest levels of intake (Guo et al., 2012). The adipose derived hormone leptin and gastrointestinal-derived hormone ghrelin communicate information about metabolic status and body weight to the hypothalamus to initiate puberty, so ghrelin and leptin represent metabolic gate for puberty (El-Eshmawy et al., 2010).
Secular trends toward a declining age at puberty onset with correlated changes in body weight have been reported in economically advanced countries. This has been attributed to excess calorie intake along with reduced physical activity in children. However, because the timing of puberty in humans is also influenced by other factors, such as genetic traits, living conditions, geographical location, and environmental chemicals, it is difficult to distinguish the effect of diet and body size from other factors in a human population. Feeding juvenile female Rhesus monkeys with a high-calorie diet results in acceleration of body growth and precocious menarche, the monkeys fed a high calorie diet also had an elevated body mass index (Teraswa et al., 2012). From the above experiment we concluded that nutrition have great impact on the onset of puberty, those individuals which have healthy diet in their juvenile period have reach puberty earlier than those which have poor calories diet or malnutrition .
5. Photoperiod
In case of seasonal breeders, photoperiodic cues have a strong influence on the onset of puberty and first reproduction, helping the animals to adjust their breeding seasons to the most appropriate season. Pregnant females transfer information about photoperiod to developing fetus via blood through melatonin. Females born into decreasing photoperiods delayed maturation by about 30 days (Fritz et al., 2010). In fish, in vivo hypothalamic sGnRH, cGnRH-II, Kiss-1 receptor, pituitary GTHα mRNA, LH-β2 and FSH-β mRNA, Kiss 1 mRNA 17α-hydroxypregnenolone and LH level increases when exposed to green LED for 4 month. Culture cell exposed to green LED only and red LED only shows that green LED significantly increases hypothalamic sGnRH, cGnRH-II, Kiss-1 receptor, Kiss 1 mRNA, GTHα mRNA, LH-β2 and FSH-β level, but no significant change were observed for red LED exposure except GTHα and LH-β2 mRNA level (Hyun et al., 2013). Greater the exposure time to light, early will be the onset of puberty. Studies found that when female Japanese quails were transferred from short days to long days causes significant increase in LH, FSH, 17β-oestradiol and ovary weights while T3, Progesterone and prolactin level decreases but no significant changes were observed in T4 concentrations. The results were opposite when female quails were transferred from long days to short days, still the T4 level was not significantly changed (Henare et al., 2012). According to another studies, photoperiodic changes in kisspeptin level is due to changes in photoperiod driven by the changes in the pattern of melatonin secretion, suggesting melatonin receptor on kisspeptin neurons. This causes increase cellular expression (Kiss1 mRNA and kisspeptin protein) and number of kisspeptin terminal appositions to GnRH neurons (Smith and Clark, 2010). Another similar study, stated that pinealectomy and/or replacement of physiological concentrations and patterns of melatonin in sheep, rhesus monkeys and various hamster species provide convincing evidence that melatonin really is the key hormone regulating the seasonal timing of puberty (Ebling, 2010).

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 171
6. Conclusion
In summary, many studies and evidences suggest that environmental cues play a central role in controlling the timing of pubertal onset through activation of GnRH– gonadotropin axis. These factors range from nutrition and social stress to wide range of endocrine disruptors in our environment. So, it cannot be concluded that any factor, alone alters the timing of puberty onset.
Altered puberty timing causes serious health concerns. Early pubertal onset contributes to both short-term and longer-term negative health outcomes including obesity, reproductive cancers, and cardiovascular disease. Women with early menarche are more susceptible to engross in early sexual activities, smoking, alcohol abuse and maternal conflictions. Further studies are needed to address all the environmental cues that affect the puberty onset and best ways to eliminate exposure to them, in order to improve health conditions in our future generations.
References
Al-Sahab, B. Ardern, CI. Hamadeh, MJ., & Tamin, H. (2010). Age at menarche in Canada: Results from the national longitudinal survey of children and youth. BMC Public Health, 10, 736-744. http://dx.doi.org/10.1186/1471-2458-10-736
Belachew, T. Hadley, C. Lindstrom, D. Getachew, Y. Duchateau, L., & Kolsteren, P. (2011). Food insecurity and age at menarche amond adolescent girls in Jimma Zone Southwest Ethiopia: a longitudinal study. Reproductive Biology and Endocrinology, 9, 125-132. http://dx.doi.org/10.1186/1477-7827-9-125
Belsky, J., & Pluess, M. (2009). Beyond diathesis stress: differential susceptibility to environmental influences. Psychological Bulletin, 135, 885-908. http://dx.doi.org/10.1037/a0017376
Boyton-Jerrett, R., & Harville, EW. (2010). A prospective study of childhood social hardship and age at menarche. Annals of Epidemiology, 22,731-737. http://dx.doi.org/10.1016/j.annepidem.2012.08.005
Braithwaite, D. Moore, DH. Lustig, RH. Epel, ES. Ong, KK. Rehkopf, DH. Wang, MC. Miller, SM., & Hiatt, RA. (2009). Socioeconomic status in relation to early menarche among black and white girls. Cancer Causes Control, 20, 713-720. http://dx.doi.org/10.1007/s10552-008-9284-9
Bryce, C. Ryan, AK. Hotchkiss, KM. Crofton, L., & Earl, GJ. (2011). In Utero and Lactational Exposure to Bisphenol A, In Contrast to Ethinyl Estradiol, Does Not Alter Sexually Dimorphic Behavior, Puberty, Fertility, and Anatomy of Female LE Rats. Toxicological sciences, 114, 133-148.
Danielle, E. Buttke, KS., & Colleen, M. (2012). Exposures to endocrine-Disrupting Chemicals and Age of Menarche in Adolescent Girls in NHANES (2003–2008). Environ Health perspect, 120, 1613-1618. http://dx.doi.org/10.1289/ehp.1104748
Deardorff, J. Fyfe, M. Ekwaru, JP. Kushi, LH. Greenspan, LC., & Yen, IH. (2010). Does

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 172
neighbourhood environment influence girls’ pubertal onset? Findings from a cohort study. BMC Paediatrics, 12, 27-35. http://dx.doi.org/10.1186/1471-2431-12-27
Deardroff, J. Ekwaru, JP. Kush, LH. Ellis, BT. Greenspan, LC. Mirabedi, A. Landaverde, EG, & Hiatt, RA. (2011). Father absence, BMI and pubertal timing in girls: Differential effects by family income and ethnicity. Journal of Adolescent Health, 48, 441-447. http://dx.doi.org/10.1016/j.jadohealth.2010.07.032
Ebling, JP. (2010). Photoperiodic regulation of puberty in seasonal species. Molecular and Cellular Endocrinology, 324, 95-101. http://dx.doi.org/10.1016/j.mce.2010.03.018
Ei, T. Joseph, R. Kurian, KL. Keen, N. Shiel, A. Ricki, J., & Colman, SV. (2012). Body Weight Impact on Puberty: Effects ofHigh-Calorie Diet on Puberty Onset in Female Rhesus Monkeys. Endocrinology, 153, 1696-1705. http://dx.doi.org/10.1210/en.2011-1970
El-Eshmawy, IA. Abdel, A. Amany, K., & El, H. (2010). Association of ghrelin and leptin with reproductive hormones in constitutional delay of growth and puberty. Reproductive Biology and Endocrinology, 8, 153. http://dx.doi.org/10.1186/1477-7827-8-153
Elizabeth, E. Hatcha, RTB. Lauren, A. Wisec, LT. Marianne, H. Julie, R., & Palmerc, W. et al. (2011). Preterm birth, fetal growth, and age at menarche among women exposed prenatally to diethylstilbestrol (DES). Reproductive Toxicology, 31, 151-157. http://dx.doi.org/10.1016/j.reprotox.2010.11.006
Fritz, T. Birte, M. Sylvia, K., & Jana, K. (2010). Puberty in female cavies is affected by photoperiod and social conditions. Physiology of behaviour, 96, 476-80.
Gaudineau, A. Ehlinger, V. Vayssiere, C. Jouret, B. Arnaud, C., & Godeau, E. (2010). Factors associated with early menarche: results from the French Health Behaviour in School-aged Children (HBSC) study. BMC Public Health, 10, 175. http://dx.doi.org/10.1186/1471-2458-10-175
Guo, C. Anette, E. Buyken, L. Shi, N. Karaolis, D. Anja, K. Stefan, A., & Wudy, G. et al. (2012). Beyond overweight: nutrition as an important lifestyle factor influencing timing of puberty. Nutrition Reviews, 7, 133-152.
Henare, SJ. Kikuchi, M. Talbot, RT., & Cockrem, JF. (2012). Changes in plasma gonadotrophins, 17β-oestradiol, progesterone, prolactin, thyroxine and triiodothyronine concentrations in female Japanese quail (Coturnix coturnix japonica) of a heavy body weight line during photo-induced ovarian growth and regression. British Poultry Science, 53, 520-530. http://dx.doi.org/10.1080/00071668.2012.719147
Hyun, SS. Hamid, RH., & Cheol, YC. (2013). The environmental regulation of maturation in goldfish (Carassius auratus): Effects of various LED light spectra. Comparative biochemistry and physiology
James-Todd, T. Tehranifar, P. Rich-Edwards, J. Titievsky, L., & Terry, MB. (2010). The impact of socioeconomic status across early life on age at menarche amonge a racially diverse population of girls. Annals of Epidemiology, 20, 836-842.

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 173
http://dx.doi.org/10.1016/j.annepidem.2010.08.006
Jawahar, B. Samuel, JA., & Stanley, RA. (2011). Gestational Cadmium Exposure-Induced Ovotoxicity Delays Puberty through Oxidative Stress and Impaired Steroid Hormone Levels. Journal of Medical Toxicology, 7, 195-204. http://dx.doi.org/10.1007/s13181-011-0143-9
Jean, RT. Wilkinson, AV. Spitz, MR. Prokhorov, A. Bondy, M., & Formam, MR. (2010). Psychosocial risk and correlates of early menarche in Mexican-American girls. Annals of Epidemiology, 173, 1203-1210. http://dx.doi.org/10.1093/aje/kwq498
Jeremy, T., & Smith, SSJ. (2012). Preweaning Over- and Underfeeding Alters Onset of Puberty in the Rat Without Affecting Kisspeptin. BIOLOGY OF REPRODUCTION, 86(145), 1-8.
Jiliang, S. Xiaoying, H. Fengmei, Z. Quanbin, X. Lihong, A., & Guozhen, L. (2012). Perinatal Exposure to Low Doses of tributyltin Chloride Advances Puberty and Affects Patterns of Estrous Cyclicity in Female Mice. Environmental Toxicology, 27, 662-670. http://dx.doi.org/10.1002/tox.21756
Krista, YC. Mildred, M. Carol, R. Adrianne, H. Antonia, M., & Calafat , KK. Et al. (2011). Exposure to polyfluoroalkyl chemicals during pregnancy is not associated with Offspring age at menarche in a contemporary British cohort. Environment International, 37, 129-131. http://dx.doi.org/10.1016/j.envint.2010.08.007
Levine, JE. (2010). Timing and completion of puberty in female mice depend on estrogen receptor α-signaling in kisspeptin neurons. Proceedings of the national academy of sciences, 107, 22693-22698. http://dx.doi.org/10.1073/pnas.1012406108
Małgorzata, M. Dobrzyńska, EJ. Tyrkiel, ED. Krzysztof, A., & Pachocki, JK. (2012). Two generation reproductive and developmental toxicity following subchronic exposure of pubescent male mice to di(2-ethylhexyl)phthalate. Annals of Agricultures and Environmental Medicines, 19, 31-37.
Manuck, SB. Craif, AE. Flory, JD. Halder, I., & Ferrell, RE. (2011). Reported early family environment covaries with menarcheal age as a function of polymorphic variation in estrogen receptor-α (ESR1). Development and Psychopathology, 23, 69-83. http://dx.doi.org/10.1017/S0954579410000659
Marisa, MF. Erica, AE. (2013). What is in our environment that affects puberty. Reproductive Toxicology.
Mayer, C. Acosta-Martinez, M. Dubois, SL. Wolfe, A. Radovick, S., & Boehm, U. et al. (2010). Lead exposure is associated with a delay in the onset of puberty in South African adolescent females: Findings from the Birth to Twenty cohort. Science of the Total Environment, 408, 4949-4954. http://dx.doi.org/10.1016/j.scitotenv.2010.07.037
Olivier, H. Susan, A. Korrick, PL. Williams, OS. Claude, ELS., & Birnbaum, JS. et al. (2013). Genetic Modification of the Association between Peripubertal Dioxin Exposure and Pubertal Onset in Cohort of Russian Boys. Environmental Health Perspectives, 121, 111-117.

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 174
Pramod, D. Akiko, H. Yasou, N. Takehiro, H. Fumio, S. Kentaro, N., & Kazayoushi, T. (2012). Circulating pituitary and gonadal hormone in spring born thoroughbred fillies and colts from birth to puberty. Journal of reproduction and development, 58, 522-530. http://dx.doi.org/10.1262/jrd.2011-025
Schooling, CM. Hui, LL. Leung, GM., & Lan, TH. (2011). Intergenerational influences on age at onset of puberty: Hong Kong’s children of 1997’ birth cohort. International Journal of Epidemiology, 41, 292-300.
Smith, TJ., & Clark, JL. (2010). Seasonal breeding as a neuroendocrine model for puberty in sheep. Molecular and Cellular Endocrinology, 324, 102-109. http://dx.doi.org/10.1016/j.mce.2010.03.007
Tan, TX., & Camras, LA. (2013). Pre-menarche pubertal development following unique form of immigration: The case girls adopted from China. Journal of Immigrants and Minority Health. http://dx.doi.org/10.1007/s10903-013-9908-3
Terasawa, E. Kurian, JR. Keen, KL. Shiel, NA. Colman, RJ., & Capuano, SV. (2012). Body weight impacts on puberty: effects of a high calorie diet on puberty onset in female rhesus monkeys. Endocrinology, 153, 1696-1705. http://dx.doi.org/10.1210/en.2011-1970
Won, HN. Mi, JP., & Myung, CG. (2011). Effects of early prepubertal exposure to bisphenol A on the onset of puberty, ovarian weights, and estrous cycle in female mice. Clinical and Experimental Reproductive Medicine, 38, 75-81. http://dx.doi.org/10.5653/cerm.2011.38.2.75
Copyright Disclaimer
Copyright for this article is retained by the author(s), with first publication rights granted to the journal.
This is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 175
Eco-Diversity of Edible Insects of Nigeria and Its Impact on Food Security
Adeoye, Olubusola Temitope (Corresponding author)
Department of Forest Conservation and Protection
Forestry Research Institute of Nigeria, Jericho-Hills Ibadan. P.M.B 5054, Oyo State, Nigeria
Tel: 234-803-881-2193 E-mail: [email protected]
Oyelowo, Oyetayo Job
Department of Forest Conservation and Protection
Forestry Research Institute of Nigeria, Jericho-Hills Ibadan. P.M.B 5054, Oyo State, Nigeria
Tel: 234-813-400-7521 E-mail: [email protected]
Adebisi- Fagbohungbe, Tola Abiodun
Department of Forest Conservation and Protection
Forestry Research Institute of Nigeria, Jericho-Hills Ibadan. P.M.B 5054, Oyo State, Nigeria
Tel: 234-806-621-1073 E-mail: [email protected]
Akinyemi, Olukayode Dare
Department of Forest Conservation and Protection
Forestry Research Institute of Nigeria, Jericho-Hills Ibadan. P.M.B 5054, Oyo State, Nigeria
Tel: 234-703-826-2669 E-mail: [email protected]
Received: June 12, 2014 Accepted: June 26, 2014
doi:10.5296/jbls.v5i2.6109 URL: http://dx.doi.org/10.5296/jbls.v5i2.6109
Abstract
Inspite of the strong aversion shown to Entomophagy (consumption of edible insects) due to

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 176
civilization, insects have played an important part in the history of human nutrition in Africa, Australia, Asia and the Americas. Hundreds of species have been used as human food. Globally, about 14 insect orders contain one or more species of edible insects. Africa is one of the leading continents that consume insects as food. In Africa, insects form part of the traditional diets of millions of people and are also used as feed for their farm animals. Nowadays, Entomophagy is a major entomological research interest with focus on its future prospect for food and feed security. Nigeria, the focus of this study is very rich in forest edible insects due to its marked ecological and climatic diversity. Some of the popular edible insects are grasshoppers, caterpillars, beetle grubs and adults, winged termites, bee, wasp and ant brood as well as winged ants, cicadas, and a variety of aquatic insects. Despite their importance as food, biodiversity and conservation efforts have focused mainly on other groups of animals, ignoring the vast world of insects. Therefore, there is a compelling need to collect data on the diversity and conservation of edible insects in Africa, and to make this information available to all interested parties. This study provides information on the list and distribution of these edible insects in Nigeria, their harvesting, processing and preservation techniques, their seasonality, nutritional value and the potential for expanding the market.
Keywords: Edible Insects, Entomophagy, Nutrition, Diversity, Food security
1. Introduction
Entomophagy is the term used to describe the process of eating insects as a food source. It can be divided into two categories: insects used as nutrients source and others as condiments. Insects have played an important part in the history of human nutrition in Africa, Australia, Asia and the Americas. Hundreds of species have been used as human food. Some of the more important groups include grasshoppers, caterpillars, beetle grubs and (sometimes) adults, winged termites (some of which are very large in the tropics), bee, wasp and ant brood (larvae and pupae) as well as winged ants, cicadas, and a variety of aquatic insects. Ordinarily, insects are not used as emergency food to ward off starvation, but are included as a normal part of the diet throughout the year or when seasonally available.
Worldwide, over 1,400 insect species are reportedly eaten as human food. Most are harvested from natural forests. But, while insects account for the greatest amount of biodiversity in forests, they are the least studied of all fauna. Indeed, as many insects cause massive damage and mortality to valuable commercial trees, virtually all insects are considered undesirable pests by many forest managers. Several investigations have shown that, in addition to their fundamental contribution for pollen dissemination, insects may contribute significantly to livelihoods in both rural and urban areas (Stack et al., 2003). Considering Forest insects as food helps in manipulating forest vegetation or harvest practices to increase, maximize, or sustain insect populations. Eating insects puts less strain on the environment because cultivating insects requires forests to be preserved rather than felled.
FAO (1995) notes that insects are also important Non Wood Forest Products that poor people gather, particularly women and children. Insects are a popular food in many cultures all over the world, be it as an occasional delicacy or as a replacement food in times of shortages, droughts, floods or war. Insects and meat play the same role in the human body. As food,

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 177
caterpillars are regulars in the village but meat is a stranger. Insects can be a good source of not only protein, but also vitamins, minerals and fats. Many insects contain abundant stores of lysine, an amino acid deficient in the diets of many people who depend heavily on grain. Further, insects generally have higher food conversion efficiency than most traditional meats and reproduce at a faster rate than beef. Protein production for human consumption would be more effective and consume fewer resources than animal protein. This makes insect meat more ecological than vertebrate meat.
According to Banjo et al. (2006) the consumption of non-toxic insects therefore, should be encouraged. Insects are traditional foods in most cultures, playing an important role in human nutrition and have much nutrient to offer. They can be reared for their high nutritional qualities and sold to the populace that regards them as delicacies. The potential of insects needs to be more seriously considered in food security and poverty alleviation strategies in Sub-Saharan Africa (De Foliart, 1992). Their impact on tree health and on the forest ecosystem in general is also significant enough to yield a wide range of forest management options to address both local food needs and forest ecosystem health and productivity.
Available information suggests that nearly 1700 insect species are reported to be used as human food (Ramos- Elorduy, 2005). While 30 species or more of edible insects are used by indigenous populations in many developing countries as food. DeFoliart (1989, 1990, 2002) reported that scores of species of edible insects are prominent items of commerce in the town and village markets of Africa, tropical and semi- tropical regions of the world.
There is an urgent need to assess insect biodiversity as a whole and the role of ethno-entomophagy in particular in conserving this valuable natural resource and the local traditional knowledge for posterity. This paper therefore provides insights on commonly eaten forest insects of Nigeria, their time of availability, market distribution, nutritional value, together with other evidence from the literature research on ethno-entomophagy in Nigeria with the aim of promoting the contribution of edible forest insects to food security and forest conservation.
1.1 The Study Areas
The survey was carried out in Lagos, Ekiti and Benue State in Nigeria. This area is characterized by a tropical climate and has three distinct vegetation belts, namely: Mangrove, High Forest and Derived Savannah respectively. The States chosen for the study has marked ecological diversity and climatic contrast which makes them to be endowed with abundant edible forest insects and commercial edible insect’s trade are also prominent in the States.
1.1.1 Research Methods
The information presented here was collected by questionnaire, oral interview of edible insect consumers and marketers, reviewing the literature of the works scientists have done on edible insects in Nigeria. Local libraries and the internet were also used as sources of information.
2. Result and Discussion
Nigeria is a country in West Africa with vast ecological and geographic diversity. There are two

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 178
major climatic seasons, namely: Rainy (Wet) and Dry Season. This marked feature makes it to be abundant in Forest and luxuriant green vegetation especially for those in South west and South east Nigeria. This further makes their edible insect population status to be high.
Table 1. Ecological zones and locations of the study sites
Ecological Zones
Location in Nigeria
State Longitude & Latitude
Average Temperature
Land Area Altitude (above sea leve)
Mangrove South West Lagos 6°35′N 3°45′E 22.66°C- 30.72°C
3,474 km2 144m
High Forest South West Ekiti 7° 37' 16N 5° 13' 17E
21oC - 28oC 5887.890km2 455m
Derived Savannah
South East/North East
Benue 7°20’N 8°45’E
21 oC- 32 oC 34,059 km2 104m
Table 1 shows the geographical zones, climate and locations of the states chosen for the study in Nigeria.
2.1 Commonly Eaten Insects in Nigeria
In Nigeria, many insects are edible and can be consumed to combat hunger and malnutrition. Fasoranti and Ajiboye(1993) reported the consumption of 7 edible insect species by the people of Kwara State while Agbidye et al.(2009) reported 4 commercially marketed edible forest insect in Benue State. Most of these insect species are found in the order Isoptera (termites), Lepidoptera (Moths), Orthoptera (Grasshoppers/crickets), Coleoptera (Beetles) and Hymenoptera (Bees). The list of some of them is shown in Table 2.
Table 2. Commonly eaten insects in Nigeria
Scientific name English name
Local name
Order Family Location Consumption stage
Cirina forda Westwood
Pallid Emperor moth
Yoruba: Kanni, Munimuni
Lepidoptera Saturniidae North Central, North East, South-West
Larvae
Bunaea alcinoe Cram
Emperor moth
- Lepidoptera Saturniidae North Central, North-East, North-West
Larvae
Rhynchophorus Phoenics
Palm weevil Yoruba: Ipe, Itun
Coleoptera Curculionidae South-South, South-West
Larvae
Oryctes boas Oryctes monocerus Oliver
Snout beetle Yoruba: Ogongo
Coleoptera Scarabaeidae South-South, South-West
Larvae
Analeptes trifasciata
Rhinocerus beetle
Ibo: Ebe Coleoptera Scarabaeidae South-South, South-West
Larvae
Anaphe venata Caterpillar Yoruba: Ekuku
Lepidoptera Notodontidae South-West Larvae
Heteroligus meles Billberger
Yam beetle - Coleoptera Scarabaeidae South-West South- South South- East
Adults
Zonocerus Grasshopper Yoruba: Orthoptera Pyrgomorphidae North Central, Adults

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 179
Variegatus, Tata Ibo: abuzu, Ukpana
South- South, South- West
Cytacantharis naeruginosus unicolor
Grasshopper Yoruba: Tata Ibo: abuzu, Ukpana
Orthoptera Pyrgomorphidae North Central, South- South, South- West
Adults
Apis mellifera
Honeybee Yoruba: Oyin
Hymenoptera Apidae All the nation All through Eggs, Larvae and Pupae
Macrotermes belliscosus Macrotermes notalensis Haviland
Termites Yoruba: Esunsun Ibo- Aku
Isoptera Termitidae North Central, North-East, South-West
Winged adult, queen
Brachytrupes membranaceus Drury
Cricket Yoruba: Ire
Orthoptera Gryllidae North Central, South-West
Adults
Gryllotalpa Africana P.de.B.
Mole cricket Yoruba: Ire
Orthoptera Gryllidae North Central, South-West
Adults
Nezera viridula L Green Stink Bug
- Hemiptera Pentatomidae North-Central Adults
Source: Field Survey (2013)
Table 2 shows the inventory of commonly eaten edible insects in Nigeria with their locations, their preferred stages of consumption and their common names and scientific names.
2.1.1 Sources and Availability
In Nigeria, insect eaters can be found in all regions of the country (Banjo et al., 2006). In the past, indigenous insect eaters were known to reside mostly in the rural areas of the country. Some kinds of Nigeria edible insects are available all year round and can be collected in forested areas and the wild (Fasoranti and Ajiboye,1993). Although a number of insect species is available throughout the year, some can only be obtained for a short season, dependent either on weather or other natural circumstances (Adeduntan and Bada, 2004; Ashiru, 1988). Insects are often collected and consumed by local people. In the past decade, a large number of insects have become available in Nigeria for domestic consumption (Agbidye et al., 2009; Muyay, 1981). However, available insects have decreased in both quantity and in the number of species due to increased demand and decreased insect habitats (Agbidye et al., 2009). A list of seasons of availability of Nigeria edible insects in local markets is presented in Table 3.
Table 3. Characteristics and seasonal availability of some edible insects in local markets in Nigeria
Scientific name Host Plant Local market Season Available Oryctes boas
Breeds in rotting Vegetation, manure standing palms e.g. coconut, oil and
Bolowou, Opuba, Ajapa, Apata in Ondo, High way road side in Delta,
Most or all of the year

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 180
Raphia palms Rhynchophorus phoenicis
Raphia palm and oil palm
Itokin in Lagos State, south of Okitipupa and the town of Aiyetoro near the coast, Bolowou, Opuba, Ajapa, Apata in Ondo, Highway roadside in Delta, Edo and Bayelsa States
A short period at the near end of the rainy season(September-October)
Cirina forda Vitellaria Paradoxa
North Bank-Benue State, Bisi, Erekesan-Ado-Ekiti, Tede, Saki- Oyo State, kwara State
June to August
Zonocerus Manihot esculenta variegatus
Chromoelena odorata, green plants in general.
Ondo State Osele, Akoko Ondo (November- April).
Mainly dry season
Bunaea alcinoe Parkia biglobosa, Prosopis africana, Khaya senegalensis
Wurukum- Benue State
Macrotermes natalensis
Tree species Gboko main market Mainly the rainy season (May-October)
Brachytrupes membranaceus
Soil and underground tunnel and forest trees nurseries
Emere- Benue State Most or all of the year
Anaphe venata Triplochiton scleroxylon
South-western Nigeria Mainly the rainy season(July-September)
Analeptes trifasciata Oil palm, Raphia palm
Western Nigeria, in Ondo, Delta, Edo and Bayelsa States
A short period at the end of the rainy season (September-October)
Source; Field Survey (2013).
Table 3 shows inventory of the commonly eaten and commercially available edible forest insects at the site of study. This list also shows the main markets where they are sold, their host plant as well as their available season.
2.1.2 Nutritional Quality
Insects and meat play the same role in the human body (Nkouka, 1987). As food, caterpillars are regulars in the village but meat is a stranger. Insects can be a good source of not only protein, but also vitamins, minerals and fats. Many insects contain abundant stores of lysine, an amino acid deficient in the diets of many people who depend heavily on grain (DeFoliart, 1992). Furthermore, insects generally have higher food conversion efficiency than most traditional meats and reproduce at a faster rate than beef. Protein production for human consumption would be more effective and consume fewer resources than animal protein (Capinera, 2004). This makes insect meat more ecological than vertebrate meat.
In Nigeria, many studies have been carried out on the nutritional value of some species of edible insects (Banjo et al., 2006; Ifie and Emeruwa, 2011; Ekop et al., 2010; Ashiru, 1988; Agbidye et al., 2009; Fasoranti and Ajiboye, 1993). These edible insects range from the order

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 181
Orthoptera, Lepidoptera, Coleoptera, Hymenoptera and Isoptera. It is noteworthy that Insects are high in protein energy (Ajayi and Adedire, 2007). In terms of g/100g dry weight, Cirina forda, B. alcinoe, M. natalensis, B. membranaceus have been shown to contain 74.35%, 74.34%, 65.62%, 35.06% of protein respectively (Agbidye,et al., 2009). Proximate analysis in terms of g/100g dry weight in earlier studies also shows M. natalensis, Oryctes boas, Anaphe spp and Rhynchophorus phoenicis having crude fibre content of 7.85%, 3.40%, 1.68 - 3.10% and 2.82 % respectively (Agbidye, et al., 2009; Banjo et al., 2006).
Insects in general are rich in fat, particularly caterpillars, palm weevil larvae and termites. High amounts of Calcium and Phosphorus (61.28mg/100g and 136.4mg/100g) were found in Analeptes trifasciata . Iron, Phosphorus and Vitamin A were 27-29mg/100g, 136mg/100g and 2.56-2.89μg/100g respectively for Macrotermes spp and 85mg/100g Iron for O.monoceros (Banjo et al., 2006; Ifie and Emeruwa, 2011). Malaisse (1997) reported that consumption of 50g dried caterpillars meets the daily human requirements of riboflavin and pantothenic acid as well as 30% of the requirement of niacin. Due to the high nutritional value of insects, flour made from larvae is traditionally used to prepare a pulp given to children to counter malnutrition.
2.1.3 Harvesting, Processing and Consumption Styles
A wide range of insects is consumed at various stages of their life cycles (Chavunduka, 1975). For example, grubs are eaten at both the larval and pupae stages. Crickets, termites and grasshoppers are eaten at the adult stages (Banjo et al., 2006). Because many insects are only seasonally available, they are frequently conserved for later consumption. They can be cooked in various ways and served as side dishes eaten with staple food. Cooked edible insects are sold commercially at roadside food stalls as well as markets in various cities of Nigeria (Agbidye, et al., 2009). Nigerian in the rural areas have used their traditional knowledge for a long time to cook each insect species in a different way; for example deep fried, fried with spices or roasted. The characteristics of local style cooking are shown in Table 4.
Table 4. The Characteristics of Local Style Cooking.
Insect Local name
Harvesting style Processing style Local style of Cooking
Macrotermes spp
Ibo-Aku Yoruba- Esunsun
Since the winged termites are attracted to electric light, they are trapped in a receptacle with water which is placed under or near the light source. In rural areas, they are often caught at the termite mound itself.
The termites wings are broken off.
The wings are broken off fried or roasted, over a fire or hot coals or fried in a pot.
Oryctes boas
Yoruba- Ogongo
By hand picking Collected larvae are washed.
The washed larvae are fried; condiments added include onion, pepper, spices and a little salt.
Rhynchophorus Yoruba- By hand picking from The end of the The washed larvae are fried;

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 182
Phoenicis Ipe Ibo- Ebe
dead raphia or oil palm abdomen, which contains the guts, is removed before the larvae are washed
condiments added include onion, pepper, spices and a little salt.
Cirina forda
Yoruba- kanni, Munimuni
The larvae are either collected from the leaves on the trees or pitfall traps made round the bases of trees with the larvae and descending larvae trapped and collected
The larvae are starved for a day or two to eliminate the contents, and boiled for two hours, then sun-dried on mats.
It is cooked in a stew with vegetables, gut condiments added then include onion, pepper, spices and a little salt
Zonocerus variegates
Yoruba- Tata Ibo- Abuzu, Ukpana
Hand picking and use of sweep net.
Allow the grasshoppers to starve overnight, remove the gut wings, and legs.
Roasting is the method frequently used for cooking. First Season it in spices and then fry in onions, garlic, cayenne, chili pepers or soy sauce may be added.
Bunaea Alcinoe The larvae are starved for a day or two to eliminate the gut contents, and then boiled for two hours, then sun-dried on mats.
It is cooked in a stew with vegetables, condiments added include onion, pepper, spices and a little salt
Brachytrupes membranaceus
Yoruba- Ire Crickets are collected from soil tunnels which they build
The crickets are roasted over a fire or hot coals.
They turn a golden color when roasted. The guts are removed before eating.
Anaphe venata Yoruba- Ekuku
The larvae are starved for a day or two to eliminate the gut contents, and then boiled for two hours, then sun-dried on mats.
It is cooked in a stew with vegetables, condiments added include onion, pepper, spices and a little salt
Source; Field Survey (2013)
Table 4 shows the characteristic ways of harvesting, processing and cooking styles for the insects found during the course of the study.

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 183
Plate 1. Freshly fried Rhynchophorus phoenicis displayed for sale alongside fried snails at Itokin market in Lagos State
Plate 2. Dried Cirina forda larvae for sale at the market in Ekiti State
Plate 3. Dried alate form of Macrotermes natalensis (termites) on sale in Gboko main market, Benue State
Plate 4: Dried Bunaea alcinoe larvae displayed for sale in Wurukum market in Makurdi, Benue State

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 184
Plate5. Adults of Brachytrupes membranaceus (crickets) displayed on sticks for sale at Emere near Okaba in Benue State
3. Potential for Expanding the Market
Insects offer significant advantages in food production, especially when compared with traditional livestock production. There is considerable potential in widening the market for edible insects by incorporating insect protein in supplements, processed foods and animal feeds (Banjo et al., 2006). In addition, their trade, although seasonal, does provide additional income for rural as well as urban people especially women.
Although the market for edible forest insects is lucrative and growing, their collection, processing and marketing is mainly done in the rural areas while some sellers go to rural areas to purchase larvae for resale in cities at high prices (Adeduntan and Bada, 2004; Mbata and Chidumayo, 2003). More attention should be paid by entomologist to the biological potential of edible forest insects, including conservation, forest management, and development of artificial diet for domestication, agriculture, nutrition, and processing aspects.
The Federal as well as State governments should enforce the legislation against indiscriminate bush burning and illegal tree felling to preserve the habitats of these insects. Where necessary, Plantations of host plants should be established to promote large scale production of these edible insects. In instances where insects are traditional foods among a certain group, this fact can serve as a path to commercial development. Relevant commercial Nigerian edible forest insect species include the Rhynchophorus phoenicis (palm grub), Oryctes boas, Cirina forda, Anaphe venata, Bunaea alcinoe.
3.1 Food Safety for Nigeria Edible Forest Insects
Insects are traditional foods in many cultures and play an important role in human nutrition. The insect has been a good nutritional food source, especially for fat and protein. Future attempts to take advantage of this resource should include how to promote Nigeria edible forest insects as a nutritional food source. Information on the toxicity and food safety of insects is needed to satisfy consumer concerns.

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 185
Studies by, Ashiru (1988) show Anaphe larvae among the insects that are an unusually good source of fat, having a calorific value of 6.113 kcal/g. Unfortunately, these larvae have recently come under a shadow for possible involvement as the cause of a seasonal ataxic syndrome. In the absence of strong medical evidence, this insect is still widely consumed by both young and old.
In general, many insects are herbivorous and less problematic than omnivores. But pesticide use can make insects unsuitable for human consumption. Bioaccumulation of herbicides can occur in insects and this may pose a problem since edible plants have been consumed by the insects themselves. For example, in Akoko, Ondo State Nigeria, the grasshopper Cytacanthacris naeruginosus unicolor and Zonocerus variegatus are collected for sale as food, but its suitability as a safe food is controlled by possible contamination with organophosphorus pesticides.
3.2 Future Trends
Nigeria edible forest insects not only serve as a source of food from forest areas of Nigeria, but also reflect rich cultural traditions and a diverse biological resource. As these edible insects are good resources for generating income, there is likely to be an increase in the numbers of Nigeria edible insects collected. However, increases in demand could lead to competition and over exploitation. Forest fragmentation and habitat loss are increasing because of land development in the country. These trends may well lead to decreases in insect diversity as well as in populations of other native fauna and flora. Many Nigeria edible insect species are collected in the wild. The quantity collected and the species found depend on season and location.
Although, several entomologists are showing interest in entomophagy studies, in connection with the local people, studies should be conducted on the development of techniques for mass rearing on a commercial scale for several edible insect species. If such insects are to be used in a sustainable manner, appropriate commercial use depends on people’s awareness of the insects’ habitats and related factors. Furthermore, product development for adding value to edible insects is necessary.
3.3 Constraint
The problems with utilizing insects’ proteins are the lack of social acceptance, nutritional knowledge, information and disbelief about delicacies of these insects that are naturally abundant in our farms and forests. There is need for insect eaters and entomologist to educate the public about the value of edible insects. There is also need for reduction in pesticide use by developing more efficient methods of harvesting pest species that are traditional foods such as grasshoppers.
4. Conclusion
This study further confirms the existence of entomophagy in Nigeria. Considering the economic, nutritional and ecological advantages of this traditional food source, its promotion deserves more attention both from national governments and assistance programmes. To manage insects in the interest of food security more attention should be given to

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 186
environmentally sustainable harvesting methods. They should be made more available throughout the year by developing improved conservation methods or by farming this minilivestock.
In other words, most preferred edible insects, especially those with high nutritional content, can be reared or cultivated in the home gardens with the application of modern tools and techniques and sold to the people, who regard them as delicacies. In the long run, this may serve the twin purpose of insect (natural resource) use as food and conservation.
Finally, there is need for research on industrial scale mass-production of edible insects, for increased recognition of the nutritional and environmental importance of insects by national governments, and for increased involvement of the media and academia in dispelling unfounded cultural biases in the urban dwellers toward insects as food. Collecting these forest edible insects would not only protect their host plants but it could benefit the environment by reducing the need to use pesticides (DeFoliart, 2005).
References
Adeduntan, S. A., & F. A. Bada. (2004). Socio-economic importance of local silkworm Anaphe venata to the rural dwellers in Ondo state, Nigeria. Proceedings of the 35th Annual Conference Entomologia Society Nigeria, Oct. 3-7, The Federal University of Akure, pp: 7-7.
Agbidye, F. S, Ofuya, T. I, & Akindele, S. O. (2009). Marketability and Nutritional Qualities of Some Edible Forest Insects in Benue State, Nigeria. Pakistan Journal of Nutrition, 8, 917-922. http://dx.doi.org/10.3923/pjn.2009.917.922
Ajayi, O. E., & C. O. Adedire (2007). Nutrient characteristics of the subterranean termite, Macrotermes subhyalinus (Rambur) (Isoptera: Termitidae). Nig. J. Entomol., 24, 42-47.
Ashiru, M. O., (1988). The food value of the larvae of Anaphe venata butler (Lepidoptera: Notodontidae). Ecol. Food Nutr., 22, 313-320. http://dx.doi.org/10.1080/03670244.1989.9991080
Banjo, A. D, Lawal,, O. A., & Songonuga,, E. A (2006). The nutritional value of fourteen species of edible insects in southwestern Nigeria. African Journal of Biotechnology, 5(3), 298-301.
Capinera, John L (2004). Encyclopedia of Entomology. Kluwer Academic Publishing.
Chavunduka, D. M (1975). Insects as a source of food to the African. Rhodesia Science News. 9, 217- 220.
De Foliart ,G. R (2002). The Human use of insects is a food resource: A Bibliographic Account in progress. http:// www.food- insects. Com/book.
De Foliart, G. (1992): Insects as human food. Crop Protection, 5(11), 395-399. http://dx.doi.org/10.1016/0261-2194(92)90020-6
DeFoliart, G. R (1989). The Human use of insects as food and animal feed. Bull Ent. Soc. Am., 35, 22-35.

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 187
DeFoliart, G. R (1990). Insects as food in indigenous. Ethno-biology: implications and applications. Proceedings of 1st International Conference on Ethno-biology. Belem, 1, 145-150.
DeFoliart, G. R (2005). Overview of role of edible insects in preserving biodiversity. In: Paoletti MG (ed) Ecological implications of minilivestock. Potential of insects, rodents, frogs and snails. Science Publishers, Inc., Enfield, 123-140
Ekop, E. A, Udoh, A. I., & Akpan, P. E. (2010). Proximate and anti-nutrient compostion of four edible insects in Akwa Ibom State, Nigeria. World Journal of Applied Science and Technology, 2(2), 224-231.
FAO (1995): Non-wood forest products for rural income and sustainable forestry. Non-wood forest products 7. FAO. Rome.
Fasoranti J. O, & Ajiboye D. O (1993). Some Edible Insects of Kwara State, Nigeria. Amer. Entomol. 39(2), 113-116.
Ifie, I., & Emeruwa, C. H. (2011). Nutritional and anti-nutritional charactyeristics of larva of Oryctes monoceros. Agric. Biol. J. N. Am., 2(1), 42-46.
Malaisse, F. (1997). Se nourrir en forêt claire africaine. Approche écologique et nutritionnelle. Les Presses Agronomiques de Gembloux. Gembloux, Belgique.
Mbata, K. J., & E. N. Chidumayo (2003). Traditional value of caterpillars (Insecta: Lepidoptera) among the bisa people of Zambia. Insect Sci. Applied, 23, 341-354.
Muyay T. (1981). Les Insects Comme Aliments de l’Homme. Pubn. Ser. II vol. 69. CEEB A. Bandundu, Zaire pp. 177.
Nkouka E. (1987). Les insectes comestibles dans les sociétés d’Afrique Centrale. Revue Scientifique et Culturelle du CICIBA, Muntu 6, 171-178.
Ramos-Elorduy, J. 2005). Insects: A hopeful food source. In: Ecological implications of minilivestock. Paoletti, M.G. Ed.; Science Pub. Enfield; 263-291.
Stack J, Dorward A, Gondo T, Frost P, Taylor F & Kurebgaseka (2003). Mopane Worm Utilization and Rural Livelihoods in Southern Africa, CIFOR Livelihood Conference, Bonn.
Copyright Disclaimer
Copyright for this article is retained by the author(s), with first publication rights granted to the journal.
This is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 188
Profile of Catfish (Clarias sp) Oocyte Exposed by Laserpuncture
Dyah Hariani (Corresponding author)
Doctoral Program, Faculty of Fisheries and Marine Sciences
University of Brawijaya, Malang-Indonesia
Biology Department, FMIPA, State University Surabaya, Surabaya-Indonesia
E-mail: [email protected]
Marhendra A. P. W
Biology Department, FMIPA, University of Brawijaya, Malang-Indonesia
Aulanni Am
Chemistry Department, FMIPA, University of Brawijaya, Malang-Indonesia
Suprayitno E
Faculty of Fisheries and Marine Sciences, University of Brawijaya, Malang-Indonesia
Received: May 6, 2014 Accepted: May 19, 2014
doi:10.5296/jbls.v5i2.5585 URL: http://dx.doi.org/10.5296/jbls.v5i2.5585
Abstract
An exposure of low-powered laserpuncture at reproductive acupoint of catfish brood stock is known stimulate oocyte maturation. However, the profile of oocyte resulted from laserpuncture exposure has not been known. The present study aimed to identify the profile of catfish (Clarias sp.) oocyte in post-exposure laserpuncture. A total of 48 catfishes with ages of 8-9 months was grouped into two, i.e. fishes were exposed with laserpuncture and without exposure to laserpuncture (control) with 3 replications. Laserpuncture exposure was conducted once a week until the brood stock matured. Profile of oocyte such as, the GSI value, egg diameter, total of oocytes and the gonad maturity stage were examined. The results showed that

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 189
laserpuncture exposure at the catfish reproductive acupoint could trigger the oocyte development without reducing quality of the oocyte indicated by the increase of gonadosomatic index (GSI), oocyte diameter, total of oocytes and gonad maturity stage similar to control. Moreover, laserpuncture exposure accelerated the gonad maturation, three weeks faster than control.
Keywords: Laserpuncture exposure, Oocyte diameter, Total of oocytes, GSI, Gonad maturity stage
1. Introduction
Oocyte development is naturally regulated by gonadotropin hormone (GtH) which is produced by pituitary hypothalamic as a response to environmental signals (Nagahama, 1994; Cornish, 1998; Bhattacharya, 1999; Wijayanti et al., 2009; Ramezani-Fard et al., 2012; Pandey, 2013). Nevertheless, natural oocyte development, in order to reach the oocyte maturation, requires longer time than manipulating the environment by the use of hormone, feed supplementation (Solang and Lamondo, 2009), and low powered laserpuncture exposure (Kusuma et al., 2008).
Laserpuncture exposure on living organisms, such as catfish, with low power (4-5 mW) (Sukarto, 1992) alters the cell membrane potential and stimulates hormone production (Karu, 1988; Koutna et al., 2003; Katona et al., 2004; Gao and Da Xing, 2009; Kusuma et al., 2012b). Moreover, laserpuncture exposure at the reproductive acupoint for 15 seconds increases the gonadotropin hormone (GtH) production (Kusuma et al., 2012b; Kusuma, 2013), which is the regulator for steroidogenesis, oogenesis, and oocyte maturation in catfish (Jalabert, 2008).
Laserpuncture induces maturation of the oocytes, ovulation and spawning (Kusuma and Hariani, 2008; Hariani and Kusuma, 2009; Hariani et al., 2010) and enlarges the GtH dynamics (Kusuma et al., 2012a; Kusuma et al., 2012b; Kusuma, 2013). Although the low power- ed laserpunceture proves the stimulation of GtH production and oocyte maturation, its effect on Gonado Somatic Index (GSI), oocyte diameter, total of oocytes, and gonad maturity stage of female catfish has not yet been studied. Fish gonad development is followed by an increase of diameter and total of oocytes (Lucey, 2009; Konan et al., 2014) and can be determined by GSI value (Poompoung et al., 2012). Therefore, we have examined profile of catfish oocyte after exposed by laserpuncture.
2. Materials and Methods
2.1 Experimental Conditions
This study was carried out in Freshwater Aquaculture Management Unit (UPBAT) Kepanjen; Biochemistry Laboratory, Faculty of Medicine, Brawijaya University; and Clinical Pathology Laboratory of dr. Soetomo Hospital, Surabaya; from January to May 2012. We collected 8-9 month old of F1 catfish brood- stock of cross-breed from female Sangkuriang and male Paiton. A total of 48 females with 760-1300 g of body weight and 48 males with 1140-1759 g of body weight were obtained from one population in UPBAT. All catfish had been acclimated for seven days in 2 x 2 x 1 m3 in tarpaulin ponds, fed in morning and afternoon with 36 % protein (Pokphan 781-3; CP Prima production). After acclimation, the mature gonad male and female catfish (ratio1:1) were transported to spawning pond facilitated with

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 190
kaka-ban for spawning. The following day, after the catfish had been spawned (assuming that they had no mature oocyte), the female catfish were treated by laserpuncture (soft laser Helium-Neon (He-Ne); 5 mW and λ 632.8 nm) for 15 seconds at the reproductive acupoint (2/3 of the ventral body) for once a week until gonads matured (three weeks) (Kusuma et al., 2007).
2.2 GSI, Total Oocyte and Oocyte Diameter Measurement
The catfish were weighed before (W) and after removing their gonad (GW) to obtain GSI value by the formula: GSI = (GW/ (W – GW))*100 (Rocha, 2008). The oocyte diameter was measured by using an ocular micrometer under the light microscope (Olympus CX 4) (Foucher and Beamish, 1980). Total oocytes were counted as follow 0.03 g of oocytes (GS) were accounted, to obtain a number of oocytes (n), and then total oocytes/broodstock were counted based on a formula N = (GW/ GS )*n (Ayidin and Sahin, 2011).
2.3 Gonad Maturity Stage Determination
Gonad maturity stage was determined by Hematoxylin and Eosin staining based on McDowell and Trump (1976) with slight modification. The upper part of the gonad (3 cm long) was fixed by PFA 10% and dehydrated in alcohol series. Then, it was continued by clearing with xylol and embedding in the paraffin block. The preparation was cutted by a microtome (with thickness 4-5μm) and then stained by Hematoxylin and Eosin. Next, it was mounted in Canada balsam and observed under a light microscope (Olympus CX 4) (Santos et al., 2005). The determination of gonad maturity stage refers to Ҫek and Yilmaz (2007).
3. Results and Discussion
This result showed that the laserpuncture accelerated gonad maturation in week four; this was indicated by the brownish red to blackish purple of the pores genitals. The ovary was dominated by oocytes in stage V and VI (Table 1), GSI was 15.52±0.73 (Table 4), oocyte diameter was 1.227±0.031 mm (Table 2), and the total of oocyte was 159885±20577 (Table 3). On the contrary, the control group showed that the gonad was mature in week seven. The ovary was dominated by oocytes in stage V and VI (Table 1), GSI was 16.39±1.23 (Table 4), oocyte diameter was 1.267±0.058 mm (Table 2), and the total of oocytes was 144150±49441 (Table 3). Laserpuncture exposure was confirmed to be able to accelerate the gonad maturity in three weeks faster than that of the control group without reduce the oocyte quality.
Table 1. Total of oocytes development stages after laserpuncture exposed compared to the control group
Week
Oocyte developmental stage (Ҫek and Yilmaz, 2007)
I II III IV V VI
C L C L C L C L C L C L
0 4 ±1 4 ± 1 3 ± 0 3 ± 0 2 ± 0 2 ±0 1 ± 0 1 ±0 1±0 1 ±0 1 ±0 1± 0
1 6 ±1 6 ± 0 3 ± 1 8 ± 1 4 ± 0 6 ±2 2 ± 0 7 ±2 2±0 5 ±1 2± 0
2 4 ±1 6 ±2 5 ± 1 7±1.7 4 ±1 6 ±1 2 ± 0 9±1.7 2±0 6 ±1 2± 0 7 ±2
3 7 ±1 7 ±2 6 ±2.7 7±2.7 4 ±1 6 ±0 8±1.7 6 ±2 3±1.7 8 ±2 2± 1 7 ±1
4 8 ±2 13±2 9 ±1.7 15±2 10±1.7 15±2 6±1.7 13±2.7 4±1 13±2.7 2± 0 10±1.7

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 191
5 10±2.7 - 8 ±1 - 6 ±1.7 - 4±1.7 - 3±1 - 2± 0 -
6 13±1.7 - 10±2 - 12±2.7 - 6±1.7 - 4±1.7 - 3±1 -
7 - - 11±2 - 10±1.7 - 8±1.7 - 6±1.7 -
Notes: C = number of oocytes without laserpuncture exposure (control) L = number of oocyte after laserpuncture exposure = not observed, broodstock already in mature gonad
Table 2. Oocyte diameter after laserpuncture exposed compared to control group:
Week Laserpuncture exposed group (mm) Control group (mm) 0 0.943±0.025 0.943±0.025 1 1.027±0.042 0.97±0.027 2 1.057±0.049 0.993±0.012 3 1.067±0.058 1.057±0.060 4 1.227±0.031* 1.093±0.012 5 - 1.163±0.025 6 - 1.193±0.012 7 - 1.267±0.058 *
Notes: *: oocytes were mature, indicated by brownish yellow and observation was terminated
Table 3. Total oocytes after laserpuncture exposed compared to control group:
Week Laserpuncture exposed group (eggs/ broodstock) Control group (eggs/broodstock) 0 15553±2771 15553±2771 1 45633±1348 29125±7278 2 138693±1205 103368±18903 3 158402±44058 110952±30265 4 159885±20577* 117212±47342 5 - 122254±26947 6 - 127349±17690 7 - 144150±49441*
Notes: *: oocytes were mature, indicated by brownish yellow and observation was terminated
Table 4. GSI after laserpuncture exposed compared to control group
Week Laserpuncture exposed group Control group 0 2.43±0.78 2.43±0.78 1 5.00±0.339 4.19±0.199 2 13.46±0.311 10.98±1.019 3 14.44±3.062* 12.12±2.636 4 15.52±0.728** 1348±3.061 5 - 14.73±1.146 6 - 15.90±4.490* 7 - 16.385±1.237**
Notes: brownish red in porous genitals and small oocytes were separated individually * * : oocytes were mature, indicated by brownish red to blackish purple in porous genitals and large oocytes were separated individually and observation was terminated
The enlargement of oocyte diameter was due to the oocyte cytoplasm filled with yolk mass. Yaron (1995) points out that in vitellogenesis level, both number and size of yolk granule are escalated; thus, it results in a bigger oocyte volume and higher gonad weight.
Laserpuncture accelerated gonad maturation three weeks faster than control (Table 1) and GSI

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 192
enhancement in catfish (Clarias sp) (Table 4). This data corroborates Kusuma et al. (2012b); Kusuma (2013) described that laserpuncture exposure at the reproductive acupoints increased GtH and GSI in catfish. The phenomenon suggested that laserpuncture exposure at the reproductive acupoint be an alternative method to accelerate gonad maturation without reducing the quality of oocytes profile. Therefore, the method is warrant for improving catfish reproduction and can be implemented for aquaculture catfish. Oocyte profile, i.e. oocyte size, total oocyte and GSI has increased in both laserpuncture and without exposure (Table 2,3 and 4). However, the laserpuncture exposure able to stimulate oocyte maturation faster followed by increasing oocyte diameter and GSI value compared to control. It is also suggested by Kusuma (2013) that laserpuncture exposure could significantly raise the gonadotropin hormone production, like GtH I, GtH II and the GSI.
GtH-I and GtH-II stimulate the gonadal gland to produce the steroid hormone namely 17β-estradiol. According to Arukwe and Goksøyr (2003), Berg et al. (2004), Jalabert (2005), and Muhammad et al. (2011), 17β-estradiol stimulates hepatic cell to synthesize vitellogenins, which will be absorbed by the oocyte (Jalabert, 2005; Muhammad et al., 2011). De Vlaming et.al. (1982) also states that mature oocyte has high value of GSI in fish that is ready to spawn. Konan (2014) explained that GSI is required to assess gonad activities and development.
GtH II stimulates the process of oocyte in the final maturation by Maturation Inducing Hormone (MIH) (Nagahama, 1994) resulting upregulation of the Maturation Promoting Factor (MPF). Then, the MPF (Nagahama, 1994; Yaron et al., 2003) stimulates the Germinal Vesicle Break Down (GVBD) so that the mature oocytes will soon be ovulated, and spawning occurs (Clelland and Peng, 2009). Therefore, the data indicated that laserpuncture exposure could accelerate the oocyte developmental stage, and shorten the spawning time (Kusuma, 2013).
4. Conclusion
Laserpuncture-exposed catfish has gonad maturation in week-four indicated by oocyte diameter 1.227±0.031 mm, total of oocytes 159885±20577eggs/broodstock, GSI 15.52 ± 0.73. Laserpuncture accelerated gonad maturation three weeks faster than control of catfish without reducing the oocyte quality.
Acknowledgement
We would like to thank to the Head of Fisheries and Marine Services of East Java Province, who granted permission to conduct this research in UPBAT Kepanjen, and Mr. Wibi Riawan (Biochemistry Laboratory, Faculty of Medicine, Brawijaya University, Malang) for his assistance for this research.
References
Arukwe, A., & Goksøyr, A. (2003). Review. Eggshell and egg yolk proteins in fish: hepatic proteins for the next generation: oogenetic, population, and evolutionary implications of endocrine disruption. Comparative Hepatology, 2(4), 1-21.
Aydin, I., & Sahin, T. (2011). Reproductive performance of turbot (Psetta maxima) in the southeastern Black sea. Tur J. Zool, 35(1), 109-113.

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 193
Berg, B., Modig, C., & Per-Erik, O. (2004). 17β-estradiol-induced vitellogenesis is inhibited by cortisol at the post-transcriptional level in Artic char (Salvelinus alpinus). Reproductive Biology and Endocrinology, 2(62), 1-10.
Bhattacharya, S. (1999). Recent advances in the hormonal regulation of gonadal maturation and spawning in fish. Special Section:Fisheries-Science & Technology. Current Science, 3(3), 342-349. 10 February 1999.
Clelland, E., & Peng, C. (2009). Endocrine/paracrine control of zebrafish ovarian development. Molecular and Cellular Endocrinology, 312, 42-52. http://dx.doi.org/10.1016/j.mce.2009.04.009
Cornish, D. A. (1998). Seasonal steroid hormones profiles in plasma and gonads of the tilapia, Oreochromis mossambicus. Water SA., 24(3), 257-363. July 1998.
De Vlaming, V., Grossman, G., & Chapman, F. (1982). On the use of the gonadosomatic index. Comp. Biochem. Physiol, 73A, 31-39. http://dx.doi.org/10.1016/0300-9629(82)90088-3
Foucher, R. P., & Beamish, R. J. (1980). Production of nonviable oocyte by production of Pacific hake (Merluccius productus). Canadian Journal of Aquatic Science, 37, 41-48. http://dx.doi.org/10.1139/f80-004
Gao, X., & Da Xing. (2009). Review Open Access. Molecular mechanisms of cell proliferation induced by low power laser irradiation. Bio Med Central. Journal of Biomedical Science, 16, 4. http://dx.doi.org/10.1186/1423-0127-16-4
Hariani, D., & Kusuma, P. S. W. (2009). Biostimuli reproduksi ikan lele Dumbo betina dengan penembakan laserpunktur. Jurnal Berkala Penelitian Hayati. PBI Cabang Jawa Timur. Edisi Khusus, 3D, 79-83. 01 December 2009.
Hariani, D., Kusuma, P. S. W., & Widodo, M. S. (2010). Pemberdayaan kelompok pembenih lele untuk peningkatan produksi benih menggunakan laserpunktur sebagai upaya peningkatan pendapatan di Desa Krecek, Kecamatan Pare, Kabupaten Kediri. Jurnal Aksi, 12(2),80-88.
Jalabert, B. (2005). Particularities of reproduction and oogenesis in teleost fish compared to mammals. Reprod. Nutr. Dev, 45, 261-279. http://dx.doi.org/10.1051/rnd:2005019
Jalabert, B. (2008). An overview of 30 years of international research in some selected fields of the Reproductive Physiology of Fish. Cybium, 32(2), 7-13.
Karu, T. I. (1988). Molecular mechanism of the therapeutic effect of low-intensity laser radiation. Laser in the Life Science, 2(1), 53-74.
Katona, E., Katona, G., Doaga, I. O., Seremet, T., Dumitrescu, M., Radesi, S., Matei, R., Horvath, J., Tanos, E., & Katona, L. (2004). Membrane effects of low level infrared laser irradiation, as seen in metabolically intact and impaired human blood cells. Romanian. J. Biophys, 14(1-4), 99-108.
Konan, Y. A., Kone, T., Bamba, M., & Kone, I. (2014). Reproductive strategies of the catfish Clarias buettikoferi (Pisces, Claridae) in the Tanoe-Eby Swamp Forest (South-Eastern Cӧte

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 194
d’Ivoire). World Journal of Fish and Marine Sciences, 6(1), 16-23.
Koutna, M., Janisch, R., & Veselska, R. (2003). Effects of low-power laser irradiation on cell proliferation. Scripta Medica (BRNO), 76(3), 163-172. June 2003.
Kusuma, P. S. W. (2013). Mekanisme pelepasan hormone gonadotropin ikan lele (Clarias sp) setelah dipapar laserpunktur pada titik reproduksi. Disertasi. Pascasarjana Fakultas Perikanan dan Ilmu Kelautan. Universitas Brawijaya, Malang.
Kusuma, P. S. W., & Hariani, D. (2008). Teknologi laserpunktur untuk mempercepat pematangan gonad ikan lele Dumbo. J Penelitian Matematika dan Sains FMIPA-UNESA, 15(2), 124-127. December 2008.
Kusuma, P. S. W., Hariani, D., Mukti, A. T., & Satyantini W. A. (2008). Penyediaan broodstock ikan lele dumbo (Clarias gariepinus) menggunakan teknologi laserpunktur sebagai upaya penyediaan benih skala massal. Jurnal Litbang Provinsi Jawa Tengah, 6(2), 139-146. 2-December 2008.
Kusuma, P. S. W., Hariani, D., Mukti, A. T., & Satyantini, W. A. (2007). Aplikasi teknologi laser untuk peningkatan produksi lele dalam rangka pengembangan ekonomi masyarakat desa di Kabupaten Boyolali Jawa Tengah, LP3K Kabupaten Boyolali, Boyolali.
Kusuma, P. S. W., Marhendra, A. P. M., Aulanni’am., & Marsoedi. (2012b). Mechanism of gonadotropin hormone release in catfish (Clarias sp) upon laserpuncture exposure to reproduction acupoint. International Journal of Basic and Applied Sciences IJBAS-IJENS, 12(6), 177-182. December. 2012.
Kusuma, P. S. W., Marhendra, A. P. W., Aulanni’am., & Marsoedi. (2012a). Mekanisme pelepasan hormon gonadotropin ikan lele (Clarias sp) setelah di induksi laserpunktur pada titik reproduksi. Seminar Nasional Tahunan IX Hasil Penelitian Perikanan dan Kelautan UGM pada Tanggal 14 July 2012.
Lucey, S. M. (2009). Characteristics of fish yolk protein and a method for inducing vitellogenin. Thesis. University of Massachusetts Amherst. September Wildlife and Fisheries Conservation.
McDowell, E. M., & Trump, B. F. (1976). Histologic fixatives suitable for diagnostic light and electron microscopy. Arch. Pathol. Lab. Med. 100:405 in Dykstra, M.J. 1993. A Manual of Applied Techniques for Biological Electron Microscopy, Plenum Press, NY. Laboratory for Advanced Electron and Light Optical Methods College of Veterinary Medicine. 1060 William Moore Drive North Carolina State University. Raleigh, NC 27607.
Muhammad, N. A., Christianus, A., Daud, S. K., Saad, C. R., Harmin, S. A., & Ina-Salwany, M. Y. (2011). Estrogen-Induced vitellogenin in Tor tambroides (Bleeker, 1854): Purification, characterization and elisa development. Journal of Fisheries and Aquatic Science, 6(7), 700-714. http://dx.doi.org/10.3923/jfas.2011.700.714
Nagahama, Y. (1994). Endocrine regulation of gametogenesis in fish. Int. J. Dev. Biol, 38, 217-229.

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
www.macrothink.org/jbls 195
Pandey, A. K. (2013). Dietary and hormonal manipulations for gonadal maturation and seed production of Indian major carps and catfishes. J.Exp.Zool. India, 16(1), 19-37.
Poompoung, P., Poompuang. S., & Kamonrat, W. (2012). Effects of warm temperatures on ovarian development of walking catfish Clarias macrocephalus (Günther, 1864) during post-spawning season. Kasetsart J. (Nat. Sci.), 46, 759-768.
Ramezani-Fard, E., Kamarudin, M. S., & Harmin, S. A. (2012). Endocrine contol of oogenesis in teleosts. Asian Journal of Animal and Veterinary Advances.
Rocha, G. R. A. (2008). Scientific Note. The introduction of the African catfish Clarias gariepinus (Burchell, 1822) into Brazillian inland waters: a growing threat. Neotropical Ichthyology, 6(4), 693-696. http://dx.doi.org/10.1590/S1679-62252008000400020
Santos, R. N., Andrade, C. C., Santos, A. F. G. N., Santos, L. N., & Araújo, F. G. (2005). Hystological analysis of ovarian development of the characiform Oligosarcus hepsetus (Vuvier, 1829) in Brazillian reservoir. Braz. J. Biol, 65(1), 169-177. http://dx.doi.org/10.1590/S1519-69842005000100020
Solang, M., & Lamondo, D. (2009). Peningkatan pertumbuhan dan indeks kematangan gonad ikan nila (Oreochromis niloticus L.) melalui pemotongan sirip ekor. Torani (Jurnal Ilmu Kelautan dan Perikanan), 19(3), 143-149. December 2009.
Sukarto. (1992). Penggunaan laser untuk akupunktur. J. Acupunctur, I, 49-54.
Wijayanti, G. E., Soeminto., & Simanjuntak, S. B. I. (2009). Profil hormon reproduksi dan gametogenesis pada gurame (Osphronemus gouramy Lac) betina. Jurnal Akuakultur Indonesia, 8(1), 77-89.
Yaron, Z,. Gur, G., Melamed, P., Rosenfeld, H., Elizur, A., & Levavi-Sivan, B. (2003). Regulation of fish gonadotropins. Int Rev Cytol, 3(225), 131-85. http://dx.doi.org/10.1016/S0074-7696(05)25004-0
Yaron, Z. (1995). Endocrine control of gametogenesis and spawning exposure in the carp. Aquaculture, 129, 49-73. http://dx.doi.org/10.1016/0044-8486(94)00229-H
Ҫek, Ş., & Yilmaz, E. (2007). Gonad development and sex ratio of Sharptooth Catfish (Clarias gariepinus Burchell, 1822) cultured under laboratory conditions. Turk. J. Zool, 31, 35-46.
Copyright Disclaimer
Copyright for this article is retained by the author(s), with first publication rights granted to the journal.
This is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).

Journal of Biology and Life Science ISSN 2157-6076
2014, Vol. 5, No. 2
http://jbls.macrothink.org 196
Reviewer Acknowledgements
Journal of Biology and Life Science (JBLS) would like to thank the following reviewers for reviewing manuscripts from March 1, 2014, to August 1, 2014. Their comments and suggestions were of great help to the authors in improving the quality of their papers. Many authors, regardless of whether JBLS publishes their work, appreciate the helpful feedback provided by the reviewers. Macrothink Institute appreciates the following reviewers’ rigorous and conscientious efforts for this journal. Each of the reviewers listed below returned at least one review during this period.
Adeline Chia Bharat Jagannathan Devaveena Dey Fazal Shirazi Fernando Cardona Gnanaraja Rajarethinam Godswill Ntsomboh Ntsefong Hong Zhao Hongbo Pang Hossam El-Din Mohamed Omar
Hui Xiang Ivo Vaz Oliveira Jinhua Wu Jittima Charoenpanich Muhammad Ali Palmiro Poltronieri Sarita Prabhakaran Subramanian Krishnan Waldiney Mello Zhen Shen
Guiju Yang
Editorial Assistant
Journal of Biology and Life Science
-------------------------------------------
Macrothink Institute
5348 Vegas Dr.#825
Las Vegas, Nevada 89108
United States
Tel: 1-702-953-1852 ext.510
Fax: 1-702-420-2900
E-mail: [email protected]
URL: http://jbls.macrothink.org





















