
Isolation and molecular characterization of the Arabidopsis TPS1 gene, encoding...
32
This article was downloaded by: [McMaster University] On: 01 August 2013, At: 10:10 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Geomicrobiology Journal Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/ugmb20 Isolation and molecular characterization of arsenite tolerant Alishewanella sp. GIDC-5 originated from industrial effluents Raina Jain a , Sanjay Jha a , Hemanta Adhikary b , Prasant Kumar b , Vipul Parekh a , Anamika Jha c , Mahesh K. Mahatma & G. Naresh Kumar b a Department of Plant Molecular Biology and Biotechnology, ASPEE College of Horticulture and Forestry , Navsari Agricultural University , Navsari , Gujarat , 396456 , India b Department of Biochemistry, Faculty of Science , The Maharaja Sayajirao University of Baroda , Vadodara , Gujarat , 390002 , India c Department of Biotechnology, Ashok & Rita Patel Institute of Integrated Study & Research in Biotechnology & Allied Sciences , New Vidyanagar , Gujarat , 388121 , India Accepted author version posted online: 09 Jul 2013. To cite this article: Geomicrobiology Journal (2013): Isolation and molecular characterization of arsenite tolerant Alishewanella sp. GIDC-5 originated from industrial effluents, Geomicrobiology Journal, DOI: 10.1080/01490451.2013.811317 To link to this article: http://dx.doi.org/10.1080/01490451.2013.811317 Disclaimer: This is a version of an unedited manuscript that has been accepted for publication. As a service to authors and researchers we are providing this version of the accepted manuscript (AM). Copyediting, typesetting, and review of the resulting proof will be undertaken on this manuscript before final publication of the Version of Record (VoR). During production and pre-press, errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal relate to this version also. PLEASE SCROLL DOWN FOR ARTICLE Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensors make no representations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content. Any opinions and views expressed in this publication are the opinions and views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and should be independently verified with primary sources of information. Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use of the Content. This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http:// www.tandfonline.com/page/terms-and-conditions
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Isolation and molecular characterization of the Arabidopsis TPS1 gene, encoding...

This article was downloaded by: [McMaster University]On: 01 August 2013, At: 10:10Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Geomicrobiology JournalPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/ugmb20
Isolation and molecular characterization of arsenitetolerant Alishewanella sp. GIDC-5 originated fromindustrial effluentsRaina Jain a , Sanjay Jha a , Hemanta Adhikary b , Prasant Kumar b , Vipul Parekh a , AnamikaJha c , Mahesh K. Mahatma & G. Naresh Kumar ba Department of Plant Molecular Biology and Biotechnology, ASPEE College of Horticultureand Forestry , Navsari Agricultural University , Navsari , Gujarat , 396456 , Indiab Department of Biochemistry, Faculty of Science , The Maharaja Sayajirao University ofBaroda , Vadodara , Gujarat , 390002 , Indiac Department of Biotechnology, Ashok & Rita Patel Institute of Integrated Study & Researchin Biotechnology & Allied Sciences , New Vidyanagar , Gujarat , 388121 , IndiaAccepted author version posted online: 09 Jul 2013.
To cite this article: Geomicrobiology Journal (2013): Isolation and molecular characterization of arsenite tolerantAlishewanella sp. GIDC-5 originated from industrial effluents, Geomicrobiology Journal, DOI: 10.1080/01490451.2013.811317
To link to this article: http://dx.doi.org/10.1080/01490451.2013.811317
Disclaimer: This is a version of an unedited manuscript that has been accepted for publication. As a serviceto authors and researchers we are providing this version of the accepted manuscript (AM). Copyediting,typesetting, and review of the resulting proof will be undertaken on this manuscript before final publication ofthe Version of Record (VoR). During production and pre-press, errors may be discovered which could affect thecontent, and all legal disclaimers that apply to the journal relate to this version also.
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of theContent. Any opinions and views expressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon andshould be independently verified with primary sources of information. Taylor and Francis shall not be liable forany losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use ofthe Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 1
Original Article
Title: Isolation and molecular characterization of arsenite tolerant Alishewanella sp.
GIDC-5 originated from industrial effluents
Authors: Raina Jain1, Sanjay Jha1*, Hemanta Adhikary2, Prasant Kumar2, Vipul Parekh1,
Anamika Jha3, Mahesh K. Mahatma, G. Naresh Kumar2
Institution at which work was performed:
1Department of Plant Molecular Biology and Biotechnology, ASPEE College of Horticulture and
Forestry, Navsari Agricultural University, Navsari, Gujarat-396456, India
2Department of Biochemistry, Faculty of Science, The Maharaja Sayajirao University of Baroda,
Vadodara, Gujarat-390002, India
3Department of Biotechnology, Ashok & Rita Patel Institute of Integrated Study & Research in
Biotechnology & Allied Sciences, New Vidyanagar, Gujarat-388121.India
* Author for correspondence E-mail: [email protected]
Phone: +91-2637-282745, Ext-659
Fax: +91-2637-282145
Abstract
Dow
nloa
ded
by [
McM
aste
r U
nive
rsity
] at
10:
10 0
1 A
ugus
t 201
3

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 2
An As-hypertolerant Alishewanella sp. GIDC-5 (Accession no. HQ659190) was isolated
from an effluent treatment plant of the industrial area near Sachin, Gujarat (India). In vitro
studies revealed that GIDC-5 can tolerate 18 mM of arsenite [As(III)] and 220 mM of arsenate
[As(V)]. PCR analysis confirmed the presence of arsenite transporters [arsB and ACR3(1)] and
arsenite oxidase gene [aioB]. Specific activities of arsenite oxidase and arsenate reductase,
located on membrane and cytoplasmic fractions respectively, increased in dose dependent
manner with arsenite concentration. Also, specific activities of antioxidant enzymes viz.,
catalase, ascorbate peroxidase, superoxide dismutase and glutathione S-transferase increased in
presence of arsenite. Increased exposure to arsenite changes enzyme activities of the glycolysis,
Krebs and glyoxylate branches dramatically. These results reveal that along with ars operon,
metabolic adaptation and antioxidant activities participate in As(III) tolerance in Alishewanella
sp. GIDC-5.
Key words: Alishewanella, arsenite, ars operon, central carbon metabolism, antioxidant
enzymes.
Abbreviations
As, arsenic; As(III), arsenite; As(V), arsenate; CS, citrate synthase; G6PDH, glucose-6-
phosphate dehydrogenase; GDH, gluconate deydrogenase; ICDH, isocitrate dehydrogenase;
PDH, pyruvate dehydrogenase; α-KGDH, α- ketoglutarate dehydrogenase; MDH, malate
dehydrogenase; ICL, isocitrate lyase; OAA, oxaloacetate; PPC, phosphoenolpyruvate
carboxylase; PYC, pyruvate carboxylase; CAT, catalase; APX, ascorbabate peroxidase; SOD,
superoxide dismutase; GST, glutathione S-transferase; GR, glutathione reductase; TCA,
Tricarboxylic acidIntroduction
Dow
nloa
ded
by [
McM
aste
r U
nive
rsity
] at
10:
10 0
1 A
ugus
t 201
3

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 3
Arsenic, a toxic metalloid of global concern, usually originated geogenically but can be
intensified by human activities (Wang and Mulligan 2006). Consequentially, elevated levels of
arsenic have been reported in soils and groundwater worldwide. The maximum concentration
limit (MCL) recommended for drinking water by the World Health Organization (WHO) is 10
µg l-1. Elevated levels of arsenic in drinking water can seriously impact human health and have
been implicated in human diseases and mortality (Doop et al. 2004). Around 6 million people in
West Bengal and more than 46 million people in Bangladesh are estimated to be at risk from
drinking water with arsenic above 50 µg l-1 (Wang and Mulligan 2006; Kinniburgh and Smedley
2001).
The Sachin region in the south Gujarat, India being the part of “Golden Corridor” has
witnessed rapid industrial development. These industrial areas contain many individual industrial
units, including dye factories, textile, rubber, pesticide and paint manufacturers, pulp and paper
producers, engineering and chemical companies. Effluents of these industries are rich in
persistent organic compounds, heavy metals and metalloids (Bruno 1995, CPCB 1996), and one
which is arsenic with its level up to 150 µg l-1 i.e., about 15 times more than India’s permissible
limits (10 µg l-1). The groundwater arsenic contamination has been increasing day by day. It has
been established that the arsenic concentrations are of geogenic origin. In microcosm-
experiments with sediments from affected aquifers, microbes have been shown to trigger arsenic
release either indirectly by using solid-phase minerals such as iron hydroxides for energy gain or
after changing the aquifer redox conditions by direct reduction of arsenate to arsenite (Akai et al.
2004; Islam et al. 2004). Microorganisms play a major role in the biochemical cycle of arsenic
Dow
nloa
ded
by [
McM
aste
r U
nive
rsity
] at
10:
10 0
1 A
ugus
t 201
3

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 4
and can convert it to different oxidation states with different solubility, mobility and toxicity
(Silver and Phung 2005).
Arsenic (‘As’) occurs in nature in four oxidation states (+5, +3, 0, –3), with pentavalent
arsenate [+5, As(V)], and trivalent arsenite [+3, As(III)] being the most common forms (Jackson
et al. 2003). Arsenate is chemically similar to inorganic phosphate and the potential to substitute
for phosphate in glycolytic and cellular respiration pathways makes it toxic for cells. Arsenate
cannot form the high-energy phosphate bonds that are needed and eventually oxidative
phosphorylation is uncoupled. Therefore, ATP with its high-energy phosphate bonds is not
formed (Ralph, 2008; Mandal and Suzuki, 2002). Adverse effects of arsenate are partially due to
its transformation to trivalent arsenite. Since, arsenite has greater mobility, it is considered more
toxic than arsenate (Ehrlich, 2005; Tamaki and Frankenberger, 1992). Arsenite inhibits
numerous cellular enzymes through sulfhydryl group binding. It inhibits cellular glucose uptake,
gluconeogenesis, fatty acid oxidation, and decreases production of cellular ATP and further
production of acetyl CoA. Exposure to arsenicals either in vitro or in vivo in model organisms
caused the induction of heat shock proteins (hsp), superoxide dismutase (SOD), catalase (CAT),
glutathione reductase (GR), glutathione peroxidase enzymes (Hughes, 2002).
A wide variety of microorganisms are found in water bodies receiving industrial
effluents. Many of the microorganisms show adaptation to the toxic materials constantly released
into their environment. They have developed strategies to defy, tolerate, metabolize, and to
detoxify these toxic substances (Parsek et al. 1995). Several heavy metal tolerance mechanisms
have been reported in different types of cells (Reed and Gadd 1990), viz., cell wall binding,
Dow
nloa
ded
by [
McM
aste
r U
nive
rsity
] at
10:
10 0
1 A
ugus
t 201
3

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 5
decreased cell membrane permeability, active extrusion and complexation with chelating agents
(Gekeler et al. 1988; Rauser 1990). The most widespread resistance mechanisms found in
bacteria against arsenic are mediated through the ars operons (Mukhopadhyay and Rosen 2002).
The current study deals with the effects of arsenic exposure on an As-hypertolerant
Alishewanella sp. strain GIDC-5, isolated from industrial effluents of Gujarat Industrial
Development Corporation area of Sachin, Gujarat, India. Our results clearly demonstrate that in
addition to the ars operon, adjustments in antioxidant machinery and central carbon metabolic
pathway are the means of adaptations in Alishewanella sp. GIDC-5 to arsenite exposure.
Materials and Methods
Isolation of arsenite tolerant bacteria: - The water sample was collected from the effluents of the
Gujarat Industrial Development Corporation (G.I.D.C.) area, located at Sachin, Gujarat, India
(Latitude: 21° 29' 46" N; Longitude: 72° 76' 51" E). Arsenic-oxidizing bacteria were isolated by
enrichment culture method. Different dilutions of the water sample were inoculated in modified
M9 minimal medium (Sambrook and Russell, 2001) containing total P < 100 µM and
supplemented with 1 mM of Sodium arsenite (NaAsO2). The media composition included
Na2CO3.10H2O, 36g/L; KH2PO4, 0.015g/L; KCl, 8g/L; NH4Cl, 5g/L; NaCl, 2.5g/L; 2mM
MgSO4; 0.1mM CaCl2 and micronutrient cocktail. The micronutrient cocktail was constituted of
FeSO4.7H2O, 3.5 mg/L; ZnSO4.7H2O, 0.16 mg/L; CuSO4.5H2O, 0.08 mg/L; H3BO3, 0.5 mg/L;
CaCl2.2H2O, 0.03 mg/L and MnSO4.4H2O, 0.4 mg/L. Glucose was used as carbon source. pH of
the medium was adjusted to 7.4 and was sterilized by autoclaving for 20 min at 121°C. Flasks
were incubated at 28°C on a rotary shaker for 48-72 h. The grown culture was harvested and
Dow
nloa
ded
by [
McM
aste
r U
nive
rsity
] at
10:
10 0
1 A
ugus
t 201
3

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 6
transferred to fresh arsenite-amended medium. After 5-6 sub-culturing with increasing
concentrations of sodium arsenite from 1 to 20 mM (As(III) : NaAsO2; 0, 5, 10, 15, 18, 20), an
enrichment culture was established. Sequential purification of culture was done by streaking on
As(III)-supplemented modified M9 agar plates. Morphologically distinct colonies tolerating
maximum arsenite concentration were further purified and stored at -20°C as 50% glycerol stock
for future use. The strain was grown to mid-exponential phase in modified M9 minimal medium
at 28°C with shaking at 180 rpm, prior to use.
16S rRNA gene amplification and sequencing: - NaCl - CTAB method (Sambrook and Russell,
2001) was used for DNA isolation from grown cells. 16S rRNA gene fragment was amplified
with thermal cycler (Eppendorf, Germany) using degenerate universal primer pair 27f and 1492r
(Aksornchu et al. 2008). After an initial denaturation step at 94°C for 5 min, 30 cycles consisting
denaturation at 94°C for 1 min, annealing at 50°C for 1 min and extension at 72°C for 1.5 min
were performed. Final extension step was kept at 72°C for 10 min. PCR products were examined
by agarose gel electrophoresis. The amplified 16S rRNA fragments were used as templates for
cycle sequencing reaction (Bigdye v3.1) followed by DNA sequencing (ABI Prism 3130 genetic
analyzer, Applied Biosystems, U.S.A) using 27f and 1492r primers. The derived sequences were
analyzed by performing online BLASTn sequence homology test
(http://www.ncbi.nlm.nih.gov/BLAST/). The phylogenetic tree was constructed by using the
neighbor-joining method in the program Phylogeny.fr (Dereeper et al. 2008).
Biochemical and Phenotypic characterization: - Growth curve of GIDC-5 was plotted by
growing the cells in modified M9 medium at 28°C in 150 ml Erlenmeyer flasks, with 0, 15, 18
Dow
nloa
ded
by [
McM
aste
r U
nive
rsity
] at
10:
10 0
1 A
ugus
t 201
3

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 7
and 20 mM NaAsO2 with continuous shaking at 180 rpm in the orbital shaker. At different time
intervals, 2 ml culture broth was withdrawn till bacteria reached stationary phase. Growth was
monitored by measuring optical density (OD) of the cultures at 600 nm using spectrophotometer
(CARY 50 UV, Australia).
Phenotypic characters of bacterial cells were observed by performing Gram staining and
subsequently analyzing under compound microscope (100x). Biochemical characterization of
strain was done by performing various tests as described by Lanyi (1987) using cultures grown at
28°C in modified M9 broth.
Cross tolerance of bacterium isolate for other heavy metals was checked by determining
the minimal inhibitory concentrations (MIC) for the metals: As (V) 0.0–250 mM; Co, Cu, Ni,
Cd, Hg 0.0-0.5 mM, as described by Drewniak et al. (2008). Medium supplemented with the
respective metal compounds was inoculated with mid-log phase grown culture to a final density
of ~106 cells/ml and then incubated for 24 h.
Amplification and sequencing of arsenical resistance genes: - The amplification of genes for
arsenite oxidase (aioB), arsenite transporter (arsB), and arsenite membrane pump, (ACR3(1) and
ACR3(2)) were performed using four pairs of degenerate primers [#1F and #1R for aioB,
darsB1F and darsB1R for arsB, dacr1F and dacr1R for ACR3(1), dacr5F and dacr4R for
ACR3(2)] as described by Cai et al. (2009). The PCR products were purified, sequenced and
analyzed.
Transformation of arsenite and arsenate: - Conversion of supplemented arsenite to arsenate in
intracellular and extracellular fractions was determined by using Inductively Coupled Plasma
Dow
nloa
ded
by [
McM
aste
r U
nive
rsity
] at
10:
10 0
1 A
ugus
t 201
3

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 8
Optical Emission Spectrometry (ICP-OES) equipped with hydride generator. To prepare
intracellular and extracellular fractions, bacterial cells were inoculated in modified M9 minimal
medium with 5 mM of arsenite, and allowed to grow till late log phase (OD 1.3, at 600 nm). 20
ml grown cultures were collected and centrifuged at 7,168xg for 3 min. Supernatant was
collected as extracellular fractions. Intracellular fractions were washed, resuspnded in 100 mM
Phosphate buffer (pH 7.0), subsequently disrupted and arsenic speciation analysed by the method
described by Cumming et al. (1999). To differentiate between intracellular and extracellular
fractions, assay for cytosolic marker (G6PDH enzyme) was performed. Standard solutions of
arsenic availed from Merck (India) was used in the experiment. HCl (for As(V)) or mixture of
KIO3 and HCl (for As(III)) served as blank, whereas, heat killed bacterial cell fractions (intra and
extracellular) were used as controls.
Analysis of organic acids: - Cells of bacterial isolate were grown in amended M9 minimal
medium (containing 100 mM glucose as a carbon source) with and without 5 mM of NaAsO2. 5
ml of late log phase cultures with pH below 5.5 were collected and centrifuged at 9,200 ×g for 5
min, supernatant was filtered through 0.22 µm filters. Pellets were washed twice with 50 mM
Phosphate buffer (pH 7.0) and resuspended in 2 ml of same buffer. These cell suspensions were
sonicated and centrifuged at 10,800 ×g for 30 min at 4°C to obtain intracellular fractions.
Intracellular fractions were filtered (0.22 µm membranes) and used for HPLC analysis.
Detection and quantification of different organic acids was done on Knauer advanced
High Performance Liquid Chromatogram (HPLC) equipped with PDA detector (Merck,
Germany). The mobile phase was 0.02% ortho-phosphoric acid (Merck, Germany) in the
Dow
nloa
ded
by [
McM
aste
r U
nive
rsity
] at
10:
10 0
1 A
ugus
t 201
3

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 9
gradient of flow rate as described by Vyas and Gulati (2009). Eluates were detected at λ 210 nm
and identified by retention time and co-chromatographed by spiking the sample with the
authentic organic acids. Organic acid standards prepared in double distilled water were filtered
(0.22 µm nylon membranes) and their retention times were determined under similar conditions.
Comparison of peak areas with external standards was used for quantification.
Enzymatic preparations and assays: - Overnight grown cells in amended M9 minimal media
supplemented with various concentrations of As(III) (0, 1, 5 and 10 mM) were harvested during
mid-log to late-log growth phase by centrifugation at 9,200 ×g for 2 min at 4°C. The preparation
of cell-free extracts for pyruvate carboxylase (PYC), phosphoenol pyruvate carboxylase (PPC)
and glucose-6-phosphate dehydrogenase (G6PDH) assays was carried out according to Kodaki et
al. (1985). The cell pellet was washed with 80 mM phosphate buffer (pH 7.5) followed by
resuspension in same buffer containing 20% glycerol and 1 mM DTT (Dithiothreitol). The cells
were lysed by sonication (1 – 1.5 min at 4°C) and centrifuged at 9,200 ×g at 4°C for 30 min to
remove the cell debris. The supernatant was used as cell-free extract for the enzyme assays and
pellets were used as membrane fractions for different enzyme assays. In case of arsenite oxidase
and arsenate reductase enzymes, 50 mM Tris-Cl (pH 7.0) buffer was used, and assay was
performed in cell-free extracts and membrane fractions both, to localize the activity of respective
enzymes. The whole cell preparation for glucose dehydrogenase (GDH) assay was carried by
washing the harvested cells thrice with normal saline and resuspending it in 0.01 M phosphate
buffer (pH 6.0) with 5 mM MgCl2.
Dow
nloa
ded
by [
McM
aste
r U
nive
rsity
] at
10:
10 0
1 A
ugus
t 201
3

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 10
Total catalase (CAT) (EC 1.11.1.6) activity was determined in the homogenates by
measuring the decrease in absorption at 240 nm as the consumption of H2O2 (ε = 39.4 mM-1 cm-
1) according the method of Aebi (1984). Ascorbate peroxidase (APX) (EC 1.11.1.11) activity
was measured in fresh extract by following the decrease in absorbance, due to ascorbate
oxidation, at 290 nm (ε = 2.8 mM-1 cm-1) (Nakano and Asada 1981). Total superoxide dismutase
(SOD) (EC 1.15.1.1) activity was measured spectrophotometrically based on inhibition in the
photochemical reduction of nitroblue tetrazoilum (NBT) as described by Van Rossun et al.
(1997). Enzyme unit of SOD was calculated according to formula given by Constantine and
Stanley (1977). Glutathione reductase (GR) (EC 1.6.4.2) activity was measured by following
NADPH reduction at 340 nm following the method of Schaedle and Bassham (1977).
Gluatathione S-transferase (GST) (EC 2.5.1.18) activity was measured by observing the
conjugation of 1-chloro, 2, 4-dinitrobenzene (CDNB) with GSH (Boyland and Chasseaud 1969).
The molar extinction of CDNB was taken as 9.6 mM-1 cm-1 at 340 nm.
Citrate synthase, CS (EC 2.3.3.1) activity was estimated by measuring the absorbance of
5,5΄-dithiobis(2-nitrobenzoic acid) at 412 nm, which would change due to its reaction with the
thiol group of CoA (Serre, 1969). The assay mixture contained the following in 1.0 ml: Tris/HCl
(pH 8.0), 93 mM; acetyl-CoA, 0.16 mM; oxaloacetate, 0.2 mM; 5,5΄-dithiobis(2-nitrobenzoic
acid), 0.1 mM and cell lysate. The reaction was started by addition of oxaloacetate. The molar
absorption coefficient was taken as 13.6 mM-1cm-1 at 412 nm. Phosphoenol pyruvate
carboxylase, PPC (EC 4.1.1.31) and pyruvate carboxylase, PYC (EC 6.4.1.1) activities were
monitored by following NADH oxidation at 340 nm in an assay combined with malate
dehydrogenase (MDH); Glucose 6-phosphate dehydrogenase, G6PDH (EC 1.1.1.49) and
Dow
nloa
ded
by [
McM
aste
r U
nive
rsity
] at
10:
10 0
1 A
ugus
t 201
3

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 11
isocitrate dehydrogenase, ICDH (1.1.1.42) activities were determined by following the reduction
of NADP at 340 nm; isocitrate lyase, ICL (4.1.3.1) activity was monitored by measuring
glyoxylate formation at 324 nm with the aid of phenylhydrazine-HCl; and glucose
dehydrogenase, GDH (EC 1.1.5.2) was assayed by following the coupled reduction of 2,6-
dichlorophenolindophenol at 600 nm, as described by Buch et al. (2008).
Arsenate reductase (AR) and arsenite oxidase (AIO) assays were performed both in
membrane and cytosolic fractions. AR activity was measured using a coupled assay system that
measures the arsenate-dependent oxidation of NADPH (Mukhopadhyay et al. 2000). This
activity was measured as a change in absorbance at 340 nm. The quantity of NADPH oxidized
was calculated using an extinction coefficient of 6,200 M-1 cm-1. The assay of arsenite oxidase
enzyme was done by following the method of Anderson et al. (1992). The reduction of the
artificial electron acceptor 2,4-dichlorophenolindophenol was monitored at 600 nm in the
presence of 1, 5 and 10 mM As(III).
All enzyme activities were determined at 28 ± 2°C, against appropriate controls lacking
the substrate or the enzyme source in the reaction mixture. One unit of specific enzyme activity
was defined as the amount of protein required to convert 1 nM substrate min-1mg-1 total protein,
unless specified. Total protein concentration of crude extracts and whole-cell suspensions was
measured by a modified Lowry method (Peterson 1977) using BSA as standard. All enzyme
activities are expressed in nM min-1 mg-1 total protein except CAT, GST, APX, GR, MDH, AIO
and AR are depicted in µM min-1 mg-1 total protein and SOD is expressed in U mg-1 total protein.
The values are depicted as Mean ± S.E.M of 3 independent observations. Statistical significance
Dow
nloa
ded
by [
McM
aste
r U
nive
rsity
] at
10:
10 0
1 A
ugus
t 201
3

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 12
level is indicated as p-values: ***, p<0.001, **, p<0.01, *, p<0.05, ns, p is non-significant. All
parameters are compared to the control i.e., culture without arsenite in medium.
Results
Isolation and characterization of arsenite tolerant bacterium: -
The isolation procedure described above yielded eight arsenite tolerant bacterial isolates
from industrial effluents of Sachin, Gujarat, India. Isolate GIDC-5 showed hypertolerance to
both the species of arsenic, As(III): 18 mM and As(V): 220 mM. Also, GIDC-5 showed a
tolerance level of 48 mM As(III) when grown in LB medium. Cross metal tolerance experiments
demonstrated that GIDC-5 could grow in presence of 0.5 mM copper, cobalt and cadmium each.
NCBI-BLAST analysis of the partial 16S rRNA sequence of GIDC-5, showed 99%
identity to 16S rRNA genes of Alishewanella genera (Fig. 1; Accession no.: HQ659190).
Microbiological tests indicated Alishewanella sp. GIDC-5 to be a Gram negative, bacilli shaped,
aerobic, motile bacterium. It can survive in high salt concentrations of up to 12% NaCl. Catalase
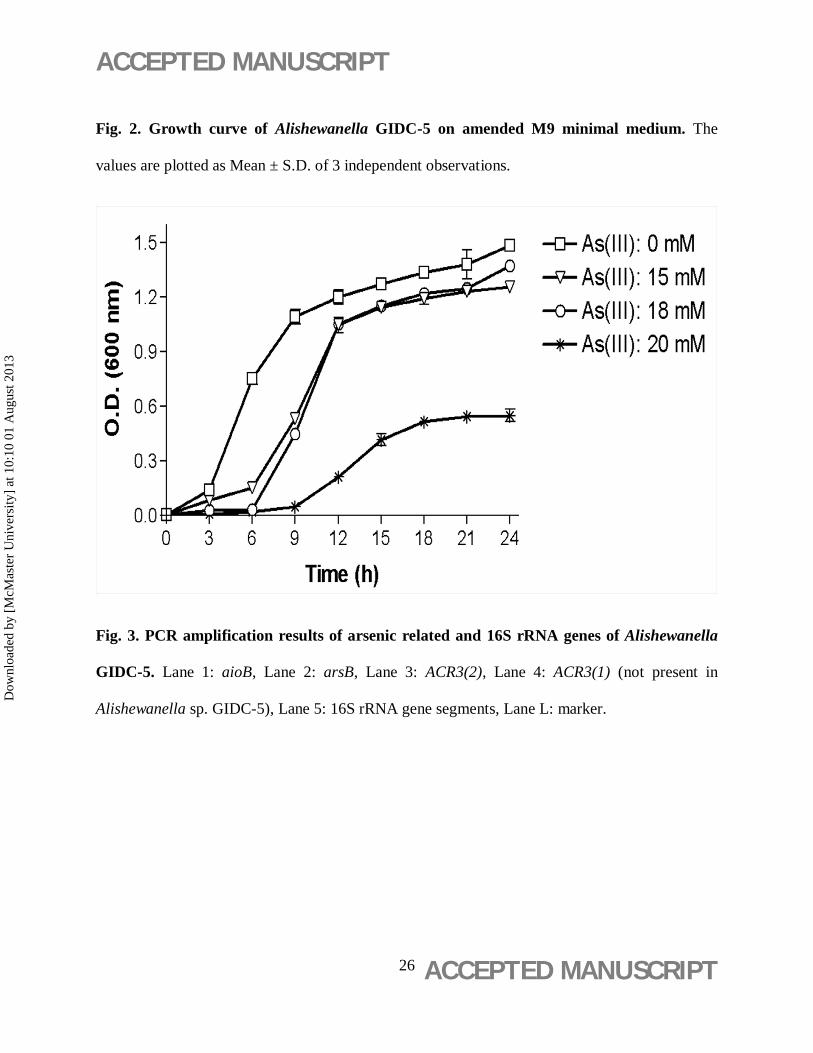
and oxidase tests were positive for GIDC-5. Results of growth kinetic study are shown in Fig. 2.
Stationary phase was obtained at 21 – 24 h in presence of As(III).
DNA fragments for arsenical resistance genes were amplified by PCR using genomic
DNA and gene specific primers. Positive amplification for arsenite oxidase (aioB, ~450bp),
arsenite transporter (arsB, ~700 bp) and arsenite membrane pump (ACR3(2), ~750 bp) genes was
obtained (Fig. 3). The nucleotide sequences of putative ACR3(2) gene are deposited to NCBI
Gene Bank database (Fig. 4; HQ659195 and HQ659196).
Transformation of arsenite and arsenate: -
Dow
nloa
ded
by [
McM
aste
r U
nive
rsity
] at
10:
10 0
1 A
ugus
t 201
3

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 13
ICP-OES was performed to study the amount of accumulation, expulsion and
transformation of As(III) and As(V) in the Alishewanella sp. GIDC-5. G6PDH activity in
extracellular fractions was found to be 0.04 ± 0.001 µM min-1 mg-1 total protein, and in cytosolic
fractions was 33.18 ± 0.07 µM min-1 mg-1 total protein. Intracellular As(III) concentration was
0.072 ± 0.034 mM, much lower than extracellular concentration, 2.539 ± 0.53 mM. Similarly,
intracellular and extracellular As(V) concentrations were 0.12 ± 0.017 mM and 2.077 ± 0.019
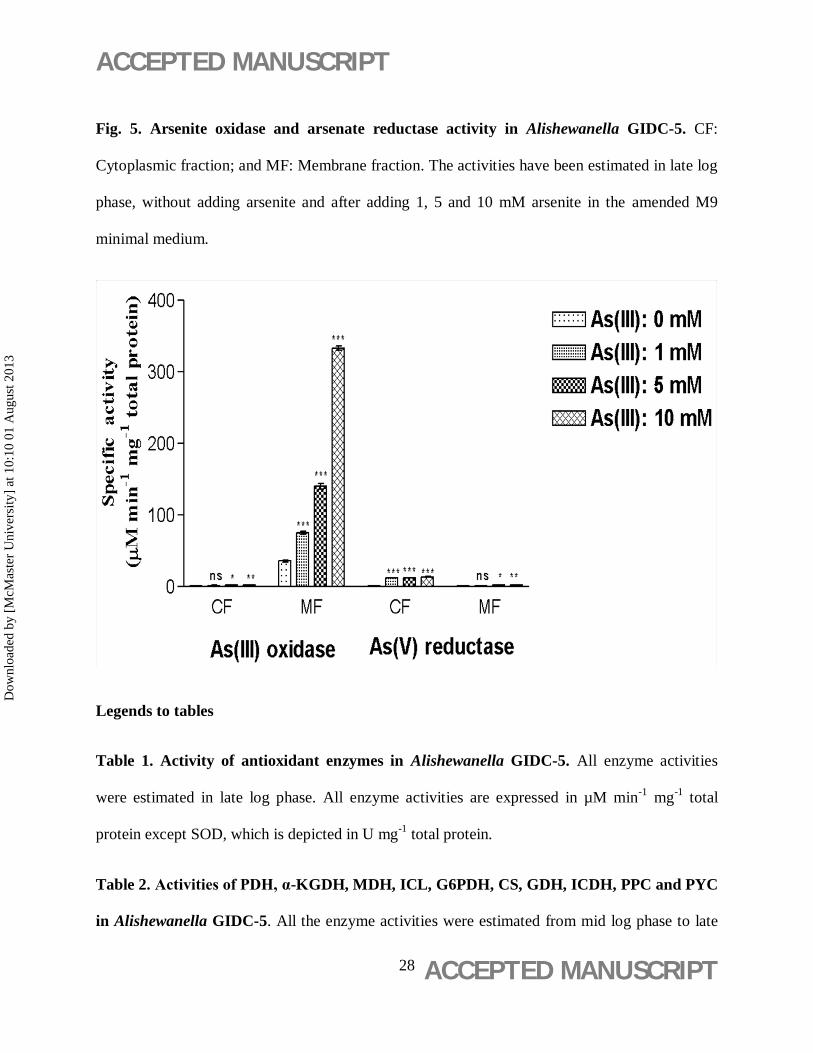
mM, respectively. As shown in Fig. 5, Arsenite oxidase (AIO) was more prominent in membrane
fractions and a positive correlation between As(III) concentrations and activity of membrane
bound arsenite oxidase was observed. In presence of 5 mM As(III), the AIO activity in
membrane fractions was observed to be 140.05 ± 3.9 µM min-1 mg-1 total protein , which is
significantly higher than that in cytoplasmic fractions (2.31 ± 0.13 µM min-1 mg-1 total protein).
Also, it is 3.9 fold higher than activity of control in membrane fractions. Contrastingly, arsenate
reductase (AR) activity was observed mainly in the cytosolic fractions of the cell. At 5 mM
As(III) concentration, AR activity was observed to be 12.59 ± 0.07 µM min-1 mg-1 total protein
in cytosol, which was significantly higher than 2.55 ± 0.1 µM min-1 mg-1 total protein compared
to membrane fraction of cell. It is 16.7 fold higher than activity of control in cytosolic fractions.
At 10 mM As(III) concentrations, both AIO and AR activities were further increased in
respective fractions. At this concentration, AR activity (13.63 ± 0.12 µM min-1 mg-1 total
protein) was 18.1 fold and AIO activity (333.07 ± 3.11 µM min-1 mg-1 total protein) was 9.3 fold
increased compared to control. No AIO and AR activities were detected in any of the heat-killed
controls.
Effect of As(III) on antioxidant enzymes: -
Dow
nloa
ded
by [
McM
aste
r U
nive
rsity
] at
10:
10 0
1 A
ugus
t 201
3

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 14
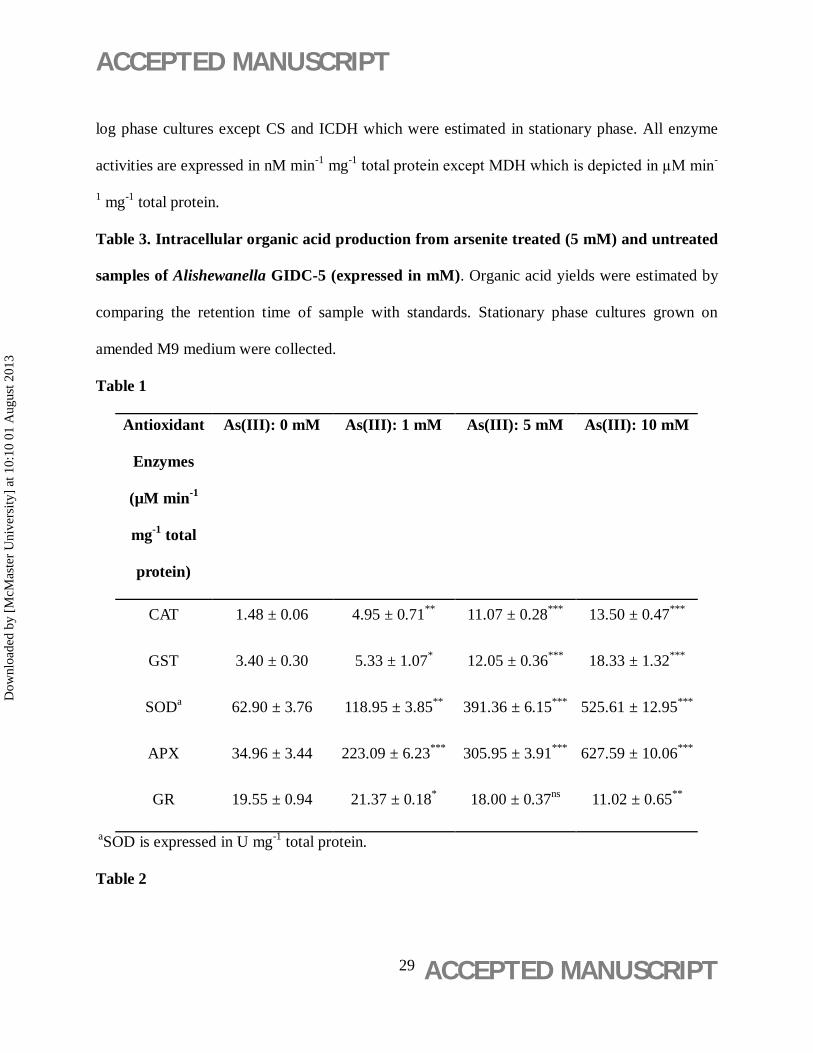
A positive effect of As(III) was observed on the activities of various antioxidant enzymes
viz. catalase (CAT), ascorbate peroxidase (APX), superoxide dismutase (SOD) and glutathione
S-transferase (GST), except glutathione reductase (GR) whose total activity was decreased at 10
mM As(III) concentration. At 5 mM As(III) a 7.5 fold increase in total CAT activity, 8.75 fold in
APX, 6.22 fold in SOD and 3.54 fold in GST activity was observed, in comparison to control.
GR activity was 1.8 times lower in presence of 10 mM As(III), respectively (Table 1).
Effect of As(III) on carbon metabolic pathway: -
Carbon metabolic pathways and organic acid concentrations were analyzed for
Alishewanella sp. GIDC-5 in response to arsenite exposure. GDH activity was 1.9 fold higher in
sample treated with 5 mM As(III) as compared to untreated control. But, G6PDH activity at
same As(III) concentration was 2.5 fold decreased. Similarly, a 1.6 and 1.2 fold inhibition in the
CS and ICDH activities was observed at 5 mM As(III). Major effect of increasing concentrations
of As(III) was observed on PYC, PDH and α-KGDH enzymes, which were 10.9, 8.6 and 7.5 fold
decreased compared to control, respectively. In contrast to all these enzyme activities which
were made vulnerable by As(III), PPC, MDH and ICL activities were increased 1.7, 3.1 and 1.9
fold in presence of 5 mM As(III) compared to respective controls (Table 2).
Intracellular organic acid levels of Alishewanella cells growing in presence of 5 mM
As(III) were quantified in HPLC (Table 3). The results demonstrate that the levels of citric (1.8
times), malic (2.2 times), fumaric (2.0 times), succinic (2.6 times) and acetic (2.8 times) acids
were significantly decreased; whereas, gluconic (2.6 times), α-ketoglutaric (2.2 times), pyruvic
(1.3 times) and oxaloacetic (5.4 times) acid levels were increased.
Dow
nloa
ded
by [
McM
aste
r U
nive
rsity
] at
10:
10 0
1 A
ugus
t 201
3

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 15
Discussion
An As-hypertolerant Alishewanella sp. GIDC-5 was isolated from an effluent treatment
plant in the industrial area near Sachin, Gujarat (India). Groundwater contamination due to
arsenic mobilization is a problem of geomicrobiology (Oremland and Stolz 2005). Hence,
isolation and study of microbes involved in arsenic transformation will add to the knowledge of
how microbes interact with arsenic-bearing minerals and control arsenic pollution in
groundwater. Thus, present study was an attempt to analyze the arsenite tolerance of
Alishewanella sp. GIDC-5 by ascertaining a link between its multiple gene products (antioxidant
enzymes, arsenite oxidase and arsenate reductase) and central carbon metabolic pathways.
Phylogenetic analysis of 16S rRNA gene of GIDC-5 clearly indicates that it belongs to
the genera Alishewanella. Further, phylogenetic study of its arsenite efflux pump gene, ACR3(2),
reveals that this gene is similar to other arsenite tolerating microbes reported. In addition to this
it explain the divergence of this bacterium from other bacteria of Alishewanella genus. The most
studied mechanism of arsenic detoxification in bacteria is ars operon. Ars-mediated tolerance
involves the reduction of arsenate (AsV) to arsenite (AsIII) via cytoplasmic AsV reductase (ArsC),
followed by the extrusion of AsIII by a membrane-associated ArsB efflux pump that is efficient at
removing AsIII (Mobley and Rosen, 1982). Most arsenite tolerant bacteria possess both arsenate
reductase and arsenite oxidase activities (Silver and Phung 2005; Mateos et al. 2006). ICP-OES
analysis of As-treated Alishewanella sp. GIDC-5 revealed the presence of extracellular and
intracellular As(III) and As(V). AIO activity was localized to membrane fractions, and was ~25
folds higher than AR activity, which was more prominent in cytosolic fractions. This indicates
that biological oxidation of arsenite could be a detoxification mechanism (Tamaki and
Dow
nloa
ded
by [
McM
aste
r U
nive
rsity
] at
10:
10 0
1 A
ugus
t 201
3

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 16
Frankenberger 1992) converting more toxic As(III) (present in the inhabiting environment of
GIDC-5) into less toxic As(V), before it could cross the cell membrane. Arsenate, As(V), a
structural analogue of phosphate, thus formed entered the cells of GIDC-5 via phosphate-specific
transporters present on membranes of bacterial cells (Rosen, 2002). Prominent AR activity was
found in cytosolic fractions of GIDC-5 cells, suggesting that As(V) thus formed was again
transformed back to As(III) in the cells. As(III) inside the cytoplasm could be extruded out via
arsenite efflux pump (arsB) and arsenite transporter (ACR3(1)). PCR analysis for marker genes
(aioB, arsB and acr3(1), further corroborated the presence of an ars-operon, a genetic
mechanism for arsenic tolerance and arsenic transformation, in GIDC-5.
Arsenic lead bacteria to increase the rate of H2O2 production which increase intracellular
ROS concentrations, by depleting GSH concentrations and membrane lipid peroxidation
reactions. This in turn induces major components of the oxidative stress response (Huang et al.
2010). Interestingly, specific activities of catalase (CAT), ascorbate peroxidase (APX),
superoxide dismutase (SOD) and glutathione S-transferase (GST) were found to increase
significantly, suggesting that the overall ROS-scavenging pathway is active in GIDC-5 in
response to arsenite exposure. Earlier studies have reported that cytoplasmic arsenate reductase
(arsC) uses glutathione (GSH) as a reactant, thus, the generated arsenite bound to GSH to form
As(III)-GS3 complex by the action of GST enzyme, followed by extrusion of the complex
through ABC transporter (Ghosh et al. 1999). This complex is a potent inhibitor of Glutathione
reductase (GR) activity in cells (Kala et al. 2000; Styblo et al. 1997). Notably, As(III) treated
GIDC-5 showed dose dependent increase in GST and decrease in GR specific activity,
suggesting the formation of arsenite-glutathione complex (As(III)-GS3) for extrusion by ABC
Dow
nloa
ded
by [
McM
aste
r U
nive
rsity
] at
10:
10 0
1 A
ugus
t 201
3

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 17
transporters. However, these transporters are not capable of removing the entire amount of
As(III) as revealed by intracellular ICP-OES studies. We further investigated the effect of
leftover intracellular As(III) on central carbon metabolism (Glycolysis, TCA and Glyoxylate
cycles) in GIDC-5 cells. Different studies have shown that G6PDH (the enzyme mediating the
intracellular glucose oxidative phosphorylation) supplies the reducing power for antioxidant
regeneration (Lundberg et al. 1999), and plays a role in the mechanism of protection against
oxidative stress (Angelova et al. 2005). Therefore, we investigated the importance of G6PDH
during arsenite exposure. G6PDH showed a dose dependent decrease in its specific activity,
implying that As(III) affect the normal Embden Meyerhoff pathway (EMP) of pyruvate
production. Whereas increased glucose dehydrogenase (GDH) activity, followed by increased
gluconic acid concentration after arsenite exposure suggests that Alishewanella sp. GIDC-5
diverts glucose towards direct oxidative pathway.
Pyruvate enters the tricarboxylic acid cycle (TCA) via the reactions catalyzed by PDH
and pyruvate carboxylase (PYC), which are regulated to coordinate the utilization of pyruvate for
anabolism (via the anaplerotic production of oxaloacetate by PYC) and catabolism (via acetyl-
CoA production by PDH) (Zubay 1988). As(III) treated GIDC-5 showed pyruvate
dehydrogenase (PDH) and pyruvate carboxylase (PYC) enzymes inhibition. Whereas, two fold
increased PPC activity was observed, suggesting that As(III) treated GIDC-5 could overcome the
inhibitory effect of metalloid on PDH enzyme by adopting an alternative pathway for
Oxaloacetic acid (OAA) production via PPC. The activities of CS, ICDH and α-KGDH enzymes
were significantly reduced followed by reduced levels of citric acid. Since, Acetyl CoA
concentration is also limiting in the cells, which could be the consequence of PDH inhibition and
Dow
nloa
ded
by [
McM
aste
r U
nive
rsity
] at
10:
10 0
1 A
ugus
t 201
3

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 18
pyruvate accumulation in the cells. Therefore, it is possible that the major control of citrate
synthase activity is by changes in the concentration of its substrates acetyl CoA. ICDH and α-
KGDH decarboxylate their respective substrates (isocitrate and α-KG) with the concomitant
formation of respective products (α-ketoglutarate and succinyl coA) and the reducing factors
NADPH or NADH. A reduction in the activity of ICDH would target isocitrate to isocitrate lyase
and lead to the production of glyoxylate (Hamel and Appanna 2001). Analysis of TCA bypass
pathway revealed that the activity of ICL enzyme, responsible for production of malic acid via
glyoxylate was up-regulated, simultaneously with reduced concentrations of succinic, fumaric
and malic acids, which suggests the metabolic shift from citric acid pathway to glyoxylate
pathway in arsenite exposed cells. Arsenite also significantly induced the activity of MDH
enzyme and resulted in intracellular OAA accumulation. The changes in the enzymatic activity
of the TCA cycles evidenced that this crucial metabolic pathway plays an important role in
Alishewanella sp. GIDC-5 to maintain an appropriate energy balance in order to adapt arsenite
stress.
In conclusion, arsenite oxidation, arsenite efflux mechanism, antioxidant enzymes and
central carbon metabolic pathway participates in Alishewanella sp. GIDC-5 to tolerate arsenite
stress in industrial effluent.
Acknowledgments
The work was supported by the Department of Biotechnology, Ministry of Science and
Technology, Government of India and Gujarat State Biotechnology Mission (GSBTM), Gujarat.
No Author has any conflict of interest.
References
Dow
nloa
ded
by [
McM
aste
r U
nive
rsity
] at
10:
10 0
1 A
ugus
t 201
3

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 19
1. Aebi H 1984. Catalase in vitro. Meth Enzymol. 105:121-126.
2. Akai J, Izumi K, Fukuhara H, Masuda H, Nakano S, Yoshimura T, Ohfuji H, Anawar
HM, Akai K 2004. Mineralogical and geomicrobiological investigations on groundwater
arsenic enrichment in Bangladesh. Appl Geochem. 19: 215–230.
3. Aksornchu P, Prasertsan P, Sobhon V 2008. Isolation of arsenic tolerant bacteria from
arsenic-contaminated soil. Songklanakarin J Sci Technol. 30:95–102.
4. Anderson GL, Williams J, Hille R 1992. The purification and characterization of arsenite
oxidase from Alcaligenes faecalis, a molybdenum-containing hydroxylase. J Biol Chem.
267:23674–23682.
5. Angelova MB, Pashova SB, Spasova BK, Vassilev SV, Slokoska LS 2005. Oxidative
stress response of filamentous fungi induced by hydrogen peroxide and paraquat.
Mycological Res 109:150–158.
6. Blacksmith Institute 2007. The World's Worst Polluted Places, The top ten of the dirty
thirty. New York, 70 p.
7. Boyland E, Chasseaud LF 1969. The Role of Glutathione and Glutathione S Transferase
in Mercaptic Acid Biosynthesis. Adv Enzymol. 32:173 – 219.
8. Bruno K 1995. Chemical Pollution: Gujarat’s toxic corridor. In: The Hindu Society of the
Environment, Kasturi & Sons Ltd., Chennai, India, p. 163-166.
Dow
nloa
ded
by [
McM
aste
r U
nive
rsity
] at
10:
10 0
1 A
ugus
t 201
3

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 20
9. Buch A, Archana G, Naresh Kumar G 2008. Metabolic channeling of glucose towards
gluconate in phosphate-solubilizing Pseudomonas aeruginosa P4 under phosphorus
deficiency. Res Microbiol. 159:635–642.
10. Cai L, Liu G, Rensing C, Wang G 2009. Genes involved in arsenic transformation and
resistance associated with different levels of arsenic contaminated soils. BMC Microbiol.
9:4-16.
11. Constantine N, Stanley KR 1977. Superoxide dismutases. 1. Occurrence in higher plants.
Pl Physiol 59:309-314.
12. CPCB 1996. Inventorisation of hazardous waste generation in five districts (Ahmedabad,
Vadodara, Bharuch, Surat and Valsad) of Gujarat. Central Pollution Control Board
(Ministry of Environment & Forests, Government of India). ISBN: 8186396632.
13. Cummings DE, Frank C Jr., Scott F, Rosenzweig RF 1999. Arsenic mobilization by the
dissimilatory Fe(III)-reducing bacterium Shewanella alga BrY. Environ Sci Technol.
33:723-729.
14. Dopp E, Hartmann LM, Florea AM, van Recklinghausen U, Pieper R, et al. 2004. Uptake
of inorganic and organic derivatives of arsenic associated with induced cytotoxic and
genotoxic effects in Chinese hamster ovary (CHO) cells. Toxicol Appl Pharmacol
201:156-165.
Dow
nloa
ded
by [
McM
aste
r U
nive
rsity
] at
10:
10 0
1 A
ugus
t 201
3

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 21
15. Dereeper A, Guignon V, Blanc G, Audic S, Buffet S, Chevenet F, Dufayard J-F, Guindon
S, Lefort V, Lescot M, Claverie J-M, Gascuel O 2008. Phylogeny.fr: robust phylogenetic
analysis for the non-specialist. Nucleic Acids Research 36 (Web Server Issue): W465-9.
16. Drewniak L, Styczek A, Majder-Lopatka M, Sklodowska A 2008. Bacteria, hypertolerant
to arsenic in the rocks of an ancient gold mine, and their potential role in dissemination of
arsenic pollution. Environ Poll. 156:1069–1074.
17. Ehrlich HL 2005. Geomicrobiology Marcel Dekker, Inc., New York.
18. Gekeler W, Grill E, Winnacker EL, Zenk MH 1988. Algae sequester heavy metals via
phytochelatin complexes. Arch Microbiol. 150:197–202.
19. Ghosh M, Shen J, and Rosen BP 1999. Pathways of As(III) detoxification in
Saccharomyces cerevisiae. Proc Natl Acad Sci USA 96:5001–5006.
20. Hamel RD, Appanna VD 2001. Modulation of TCA cycle enzymes and aluminum stress
in Pseudomonas fluorescens. J Inorg Biochem. 87:1–8.
21. Huang A, Teplitski M, Rathinasabapathi B, Ma L 2010. Characterization of arsenic-
resistant bacteria from the rhizosphere of arsenic hyperaccumulator Pteris vittata. Can J
Microbiol 56:236–246.
22. Hughes MF 2002. Arsenic toxicity and potential mechanisms of action. Toxicol Lett.
133:1–16.
Dow
nloa
ded
by [
McM
aste
r U
nive
rsity
] at
10:
10 0
1 A
ugus
t 201
3

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 22
23. Islam FS, Gault AG, Boothman C, Polya DA, Charnock JM, Chatterjee D, Lloyd JR
2004. Role of metal-reducing bacteria in arsenic release from Bengal delta sediments.
Nature, 430:68–71.
24. Jackson CR, Jackson EF, Dugas SL, Gamble K, Williams SE 2003. Microbial
transformations of arsenite and arsenate in natural environments. Rec Res Develop
Microbiol 7:103-118.
25. Kala SV, Neely MW, Kala G, Prater CI, Atwood DW, Rice JS, Lieberman MW 2000.
The MRP2/cMOAT transporter and Arsenic-Glutathione complex formation are required
for biliary excretion of arsenic. The J Biol Chem. 275:33404–33408.
26. Kinniburgh DG, Smedley PL 2001. Arsenic contamination of groundwater in
Bangladesh. Natural Environ Res Council. 2:630.
27. Kodaki T, Katagiri F, Asano M, Izui K, Katsuki H 1985. Cloning of
phosphoenolpyruvate carboxylase gene from a cyanobacterium, Anacystis nidulans, in
Escherichia coli. J. Biochem. 97:533-539.
28. Lanyi B 1987 Classical and rapid identification methods for medically important bacteria.
Methods Microbiol. 19:1–67.
29. Lundberg B, Wolf RE, Dinauer M, Xu Y, Fang F 1999. Glucose 6-phosphate
dehydrogenase is required for Salmonella typhimurium virulence and resistance to
reactive oxygen and nitrogen intermediates. Infect Immun. 65:5371–5375.
30. Mandal BK, Suzuki KT 2002 Arsenic round the world: a review. Talanta 58: 201-235.
Dow
nloa
ded
by [
McM
aste
r U
nive
rsity
] at
10:
10 0
1 A
ugus
t 201
3

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 23
31. Mateos LM, Ordonez E, Latek M, Gill JA (2006). Corynebacterium glutamicum as a
model bacterium for bioremediation of arsenic. Intrnl Microbiol. 9:207-215.
32. Mobley HL, Rosen BP 1982. Energetics of plasmid mediated arsenate resistance in
Escherichia coli. Proc Natl Acad Sci, USA. 79:6119–6122.
33. Mukhopadhyay R, Rosen BP 2002 Arsenate reductases in prokaryotes and eukaryotes.
Environ Health Perspect. 110:745–748.
34. Mukhopadhyay R, Shi J, Rosen BP 2000 Purification and characterization of ACR2p, the
Saccharomyces cerevisiae arsenate reductase. J Biol Chem. 275:21149–21157.
35. Nakano Y, Asada K 1981. Hydrogen peroxide is scavenged by ascorbate specific
peroxidase in spinach chloroplasts. Pl Cell Physiol 22:868-880.
36. Oremland RS, Stolz, JF 2005. Arsenic, microbes and contaminated aquifers. Trends
Microbiol 13:45-49.
37. Parsek MR, Mefall SM, Chakrabarty AM 1995. Microbial degradation of toxic
environmental pollutants, ecological and evolutionary considerations. Int Biodeterior
Biodegrad. 35:175-188.
38. Peterson GL 1979. Review of the Folin phenol quantitation method of Lowry,
Rosenberg, Farr and Randall. Anal Biochem. 100:201.
39. Ralph SJ 2008. Arsenic-based antineoplastic drugs and their mechanisms of action.
Metal-Based Drugs. doi:10.1155/2008/260146
Dow
nloa
ded
by [
McM
aste
r U
nive
rsity
] at
10:
10 0
1 A
ugus
t 201
3

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 24
40. Rauser WE 1990. Phytochelatins. Ann Rev Biochem 59: 61–86.
41. Reed RH, Gadd GM 1990. Metal tolerance in eukaryotic and prokaryotic algae. In: Shaw
AJ (ed) Heavy metal tolerance in plants: evolutionary aspects. CRC Press, Boca Raton,
FL, p. 105-118.
42. Rosen BP 2002. Biochemistry of arsenic detoxification. FEBS Lett 529: 86-92.
43. Sambrook J, Russell D 2001. Molecular cloning: A laboratory manual (3rd Eds.) Cold
Spring Harbor Laboratory Press, New York.
44. Schaedle M, Bassham JA 1977. Chloroplast Glutathione reductase. Pl Physiol. 59:1011-
1012.
45. Serre PA 1969. Citrate synthase. In: Lowenstein JM (ed) Methods in Enzymology, Vol.
13 Academic, New York, p. 3-11.
46. Silver S, Phung LT 2005. Genes and enzymes involved in bacterial oxidation and
reduction of inorganic arsenic. Appl Environ Microbiol. 71:599–608.
47. Styblo M, Seves SV, Cullen WR, Thomas, DJ 1997. Comparative inhibition of yeast
glutathione reductase by arsenicals and arsenothiols. Chem Res Toxicol. 10:27-33.
48. Tamaki S, Frankenberger WT Jr. 1992. Environmental biochemistry of arsenic. Rev
Environ Contam Toxicol 124:79–110.
49. Van Rossun MNPC, Alberda M, Van Der Plas LHW 1997. Role of oxidative damage in
tulip bulb scale micropropagation. Pl Sci. 130:207-216.
Dow
nloa
ded
by [
McM
aste
r U
nive
rsity
] at
10:
10 0
1 A
ugus
t 201
3

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 25
50. Vyas P, Gulati A 2009. Organic acid production in vitro and plant growth promotion in
maize under controlled environment by phosphate-solubilizing fluorescent Pseudomonas.
BMC Microbiol. 9:174-189.
51. Wang S, Mulligan CN 2006. Occurrence of arsenic contamination in Canada: sources,
behavior and distribution. Sci Total Environ. 366:701-721.
52. Zubay G 1988. Biochemistry. Macmillan Publishing Corp., New York.
Legends to figures
Fig. 1. Phylogram of arsenic-resistant bacteria derived from 16S rRNA sequence data of
Alishewanella sp. GIDC-5 and closely related species. The scale bar represents 0.8
substitutions per nucleotide.
Dow
nloa
ded
by [
McM
aste
r U
nive
rsity
] at
10:
10 0
1 A
ugus
t 201
3
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 26
Fig. 2. Growth curve of Alishewanella GIDC-5 on amended M9 minimal medium. The
values are plotted as Mean ± S.D. of 3 independent observations.
Fig. 3. PCR amplification results of arsenic related and 16S rRNA genes of Alishewanella
GIDC-5. Lane 1: aioB, Lane 2: arsB, Lane 3: ACR3(2), Lane 4: ACR3(1) (not present in
Alishewanella sp. GIDC-5), Lane 5: 16S rRNA gene segments, Lane L: marker.
Dow
nloa
ded
by [
McM
aste
r U
nive
rsity
] at
10:
10 0
1 A
ugus
t 201
3

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 27
Fig. 4. Phylogram of arsenic-resistant bacteria derived from arsenic efflux pump gene
ACR3(2) gene sequence data of Alishewanella sp. GIDC-5 and closely related species. The
scale bar represents 0.6 substitutions per nucleotide. Bootstrap values (expressed as percentages
of 1000 replications) greater than 50% are given at the nodes.
Dow
nloa
ded
by [
McM
aste
r U
nive
rsity
] at
10:
10 0
1 A
ugus
t 201
3
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 28
Fig. 5. Arsenite oxidase and arsenate reductase activity in Alishewanella GIDC-5. CF:
Cytoplasmic fraction; and MF: Membrane fraction. The activities have been estimated in late log
phase, without adding arsenite and after adding 1, 5 and 10 mM arsenite in the amended M9
minimal medium.
Legends to tables
Table 1. Activity of antioxidant enzymes in Alishewanella GIDC-5. All enzyme activities
were estimated in late log phase. All enzyme activities are expressed in µM min-1 mg-1 total
protein except SOD, which is depicted in U mg-1 total protein.
Table 2. Activities of PDH, α-KGDH, MDH, ICL, G6PDH, CS, GDH, ICDH, PPC and PYC
in Alishewanella GIDC-5. All the enzyme activities were estimated from mid log phase to late
Dow
nloa
ded
by [
McM
aste
r U
nive
rsity
] at
10:
10 0
1 A
ugus
t 201
3
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 29
log phase cultures except CS and ICDH which were estimated in stationary phase. All enzyme
activities are expressed in nM min-1 mg-1 total protein except MDH which is depicted in µM min-
1 mg-1 total protein.
Table 3. Intracellular organic acid production from arsenite treated (5 mM) and untreated
samples of Alishewanella GIDC-5 (expressed in mM). Organic acid yields were estimated by
comparing the retention time of sample with standards. Stationary phase cultures grown on
amended M9 medium were collected.
Table 1
Antioxidant
Enzymes
(µM min-1
mg-1 total
protein)
As(III): 0 mM As(III): 1 mM As(III): 5 mM As(III): 10 mM
CAT 1.48 ± 0.06 4.95 ± 0.71** 11.07 ± 0.28*** 13.50 ± 0.47***
GST 3.40 ± 0.30 5.33 ± 1.07* 12.05 ± 0.36*** 18.33 ± 1.32***
SODa 62.90 ± 3.76 118.95 ± 3.85** 391.36 ± 6.15*** 525.61 ± 12.95***
APX 34.96 ± 3.44 223.09 ± 6.23*** 305.95 ± 3.91*** 627.59 ± 10.06***
GR 19.55 ± 0.94 21.37 ± 0.18* 18.00 ± 0.37ns 11.02 ± 0.65** aSOD is expressed in U mg-1 total protein.
Table 2
Dow
nloa
ded
by [
McM
aste
r U
nive
rsity
] at
10:
10 0
1 A
ugus
t 201
3

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 30
Metabolic
enzymes (nM
min-1 mg-1
total protein)
As(III): 0 mM As(III): 1 mM As(III): 5 mM As(III): 10 mM
PDH 19.04 ± 0.04 5.82 ± 0.44*** 2.21 ± 0.21*** 1.20 ± 0.01***
α-KGDH 18.62 ± 0.29 11.72 ± 0.55*** 2.49 ± 0.11*** 1.05 ± 0.12***
MDHb 2.06 ± 0.07 5.12 ± 0.25*** 6.39 ± 0.18*** 10.00 ± 0.22***
ICL 4.67 ± 0.11 6.24 ± 0.80ns 8.72 ± 0.52** 8.55 ± 0.65**
G6PDH 84.14 ± 1.58 40.25 ± 8.41*** 33.18 ± 0.07*** 13.56 ± 2.92***
CS 19.51 ± 0.15 14.27 ± 0.62*** 12.27 ± 0.76*** 9.84 ± 0.70***
GDH 58.57 ± 1.46 56.37 ± 1.46ns 114.03 ± 6.04** 166.98 ± 7.16**
ICDH 109.74 ± 11.20 95.61 ± 0.14** 89.39 ± 0.07** 87.96 ± 0.39**
PPC 13.75 ± 0.35 18.70 ± 0.4** 23.40 ± 0.70*** 28.18 ± 0.88***
PYC 88.45 ± 2.57 23.20 ± 0.75*** 8.15 ± 0.90*** 2.31 ± 0.02***
b MDH is expressed in µM min-1 mg-1 total protein
Table 3
Intracellular
Organic acids
(mM)
As(III): 0 mM As(III): 5 mM
Gluconic acid 3.20 ± 0.00 8.40 ± 0.31***
Dow
nloa
ded
by [
McM
aste
r U
nive
rsity
] at
10:
10 0
1 A
ugus
t 201
3

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 31
Pyruvic acid 2.53 ± 0.25 3.31 ± 0.19*
Succinic acid 2.68 ± 0.01 ND
Citric acid 4.50 ± 0.08 2.50 ± 0.00***
α-KG 1.37 ± 0.09 3.04 ± 0.03***
Fumaric acid 2.09 ± 0.08 1.03 ± 0.02***
Malic acid 5.87 ± 0.44 2.66 ± 0.06***
Oxaloacetic acid 0.65 ± 0.05 3.50 ± 0.10*** ND: not detected
Dow
nloa
ded
by [
McM
aste
r U
nive
rsity
] at
10:
10 0
1 A
ugus
t 201
3




















