
A systematic approach to creating terrain-capable hybrid soft ...
Upload
independentCategory
view
0download
0
B
IddMR
Opc
Stiw
Rnda
Cc
Mopf
FMOGO(R
R2
RIOS
T0S2FfRF
0©d
Research www.AJOG.org
ASIC SCIENCE: OBSTETRICS
solation and characterization of true mesenchymal stem cellserived from human term decidua capable of multilineageifferentiation into all 3 embryonic layersaria I. Macias, MSc; Jesús Grande, MD; Ana Moreno, MSc; Irene Domínguez, MSc;
afael Bornstein, MD, PhD; Ana I. Flores, PhD
emf
CMbflDr
K
BJECTIVE: The objective of the study was to isolate and characterize aopulation of mesenchymal stem cells (MSCs) from human term pla-ental membranes.
TUDY DESIGN: We isolated an adherent cell population from ex-raembryonic membranes. Morphology, phenotype, growth character-stics, karyotype, and immnunological and differentiation propertiesere analyzed.
ESULTS: The isolated placental MSCs were from maternal origin andamed as decidua-derived mesenchymal stem cells (DMSCs). DMSCsifferentiated into derivatives of all germ layers. It is the first report
apable of multilineage differentiation into all 3 embryonic layers. Am J Obstet Gyne
eiththisc
seeqpdd
wirco(efifluid,13-16 Wharton’s
oi: 10.1016/j.ajog.2010.06.045
xpanded DMSCs differentiated into all embryonic layers, including pul-onary cells. DMSCs showed higher life span than placental cells from
etal origin and proliferated without genomic instability.
ONCLUSION: The data suggest that DMSCs are true multipotentSCs, distinguishing them from other placental MSCs. DMSCs could
e safely used in the mother as a potential source of MSCs for pelvicoor dysfunctions and immunological diseases. Additionally, frozenMSCs can be stored for both autologous and allogeneic tissue
egeneration.
ey words: decidua, differentiation, mesenchymal stem cells,
bout placental MSC differentiation into alveolar type II cells. Clonally multipotency, placentaite this article as: Macias MI, Grande J, Moreno A, et al. Isolation and characterization of true mesenchymal stem cells derived from human term decidua
col 2010;203:495.e9-23.mdcsB
gppsdtctg
toctacmasfd
esenchymal stem cells (MSCs)have a spindle-shaped morphol-
gy, are isolated for their adherence tolastic, and phenotypically characterized
or the expression of a selection of mark-
rom the Research Center (Dr Flores, Msacias, and Ms Domiguez), Departments ofbstetrics and Gynecology (Dr Grande) andenetics (Ms Moreno), Hospital 12 dectubre; and Department of Hematology
Dr Bornstein), Hospital Central de la Cruzoja, Madrid, Spain.
eceived Jan. 22, 2010; revised May 27,010; accepted June 17, 2010.
eprints: Ana I. Flores, PhD, Centro denvestigacion, Laboratorio 1, Hospital 12ctubre, Avda Cordoba s/n, 28041 Madrid,pain. [email protected].
his study was supported in part by Grants FIS5/2288 and CP03/00004 (to A.I.F.) from thepanish Institute of Health Carlos III, Grants004-029 and 2008-108 from the MMAoundation (to A.I.F.), and a contract project
rom the Hospital 12 Octubre Biomedicalesearch Foundation (associated with A.I.F.IS 05/2288).
002-9378/$36.002010 Mosby, Inc. All rights reserved.
rs and the capacity of differentiationnto bone, fat, and cartilage.1,2 Besidesheir mesodermal differentiation, MSCsave been differentiated to cells of mul-iple organs such as muscle, neurons,epatocytes, and insulin-producing cells,
ndicating that MSCs are an importantource of stem cells for regenerative medi-ine and tissue engineering.3
Currently bone marrow (BM) repre-ents the main source of MSCs for bothxperimental and clinical studies. How-ver, BM-MSCs are present in lowuantities and collected by an invasiverocedure, and their proliferation andifferentiation capacity decrease withonor age.4,5
Placenta is an additional source of MSCsith several advantages, including easy
solation without invasive procedures, lowisk of viral infection, and lack of ethicaloncerns.6-9 Human placenta is composedf fetal and maternal tissues.10 Stem cellsmostly MSCs but also hematopoietic andpithelial stem cells) have been obtainedrom multiple parts of the placenta includ-ng the umbilical cord blood,11,12 amniotic
jelly,17-19 amniotic t
NOVEMBER 2010 Americ
embrane,20-22 chorionic membrane,23,24
ecidua,25-29 and chorionic villi.30-32 Pla-enta-derived MSCs have higher expan-ion25,28 and engraftment33 capacity thanM-MSCs.Placental MSCs are hypoimmuno-
enic and exhibit immunomodulatoryroperties,34,35 so they can be trans-lanted without the need for immuno-uppression. Moreover, placental MSCsisplay no evidence for teratoma forma-ion.14 Then MSCs from a single sourcean be used for multiple patients in au-oimmune diseases and as a vehicle forenes in gene therapy protocols.36-38
In this study, an adherent cell popula-ion was isolated from the extraembry-nic membranes of human term pla-enta and characterized in reference toheir morphology, immunophenotype,nd differentiation potential. Isolatedells belong to the maternal side of fetalembranes (ie, decidua parietalis) and
re a true population of mesenchymaltem cells with higher expansion and dif-erentiation capacity than other MSCs,istinguishing them from other placen-
al cell types.an Journal of Obstetrics & Gynecology 495.e9

cclsclchtktto
MPaHDoofC1(epa�bba
comtlcraltm
cafi2p1crf
dgdopw
IoPsePFPCntpPPC(anestD
IMAobut(
ftfS3n(
wc(tDib
ev
lbw(7dngif(
wgsp
EPawdwC(
EFwct2nahl
TTTlipsrtcnnatr
Research Basic Science: Obstetrics www.AJOG.org
4
Decidua-derived mesenchymal stemells (DMSCs) and clonally expandedells differentiated into the 3 embryonicayers. Additionally, this is the first studyhowing differentiation of human pla-enta-derived MSCs to alveolar epithe-ial type II cells. These maternal mesen-hymal-like cells are easy to isolate, haveigh expansion and differentiation po-ential while maintaining a normalaryotype, and would be suitable for es-ablishment of either allogenic or pa-ient-specific banks for future clinicalutcomes.
ATERIALS AND METHODSrocessing of placental membranesnd culture of primary cellsuman placentas were obtained at theepartment of Obstetrics and Gynecol-gy from healthy mothers during naturalr cesarean births under written in-ormed consent approved by the Ethicsommittee from Hospital Universitario2 de Octubre. Placental membranesamnion, chorion, and decidua pari-talis) were dissected and washed inhosphate-buffered saline (PBS) plusntibiotics (100 IU penicillin and 100g/mL streptomycin) to remove redlood cells. At times, the amnion mem-rane was peeled off and processed sep-rately from the chorion-decidua tissue.
Placental tissues were processed ac-ording to protocol described for amni-tic membrane,21 except for significantodifications. Tissue was digested with
rypsin-versene (Lonza, Basel, Switzer-and) twice, and cell suspensions wereentrifugated at 400 � g for 10 minutes,esuspended in PBS plus 2 mM EDTAnd 0.5% human serum albumin, andysed in 0.8% ammonium chloride solu-ion for 15 minutes to remove any re-
aining erythrocytes.Isolated cells were seeded at 1.16 � 105
ells/cm2 and cultured at 37°C, 5% CO2,nd 95% humidity in Dulbecco’s modi-ed Eagle’s medium supplemented withmM of glutamine, 0.1 mM of sodiumyruvate, 55 �M of �-mercaptoethanol,% nonessential amino acids, 1% peni-illin/streptomycin, 10% fetal bovine se-um, and 10 ng/mL of epithelial growth
actor. p95.e10 American Journal of Obstetrics & Gyneco
Nonadherent cells were washed off 5ays after seeding. Adherent cells wererown to confluence and passaged at aensity of 4-5 � 104 cells/cm2. Cloningf cells was carried out by plating 1 celler well in a 96 well plate. Four clonesere selected.
nmunophenotypic characterizationf placental cellslacental cells from early and late pas-ages were characterized by flow cytom-try with antibodies against CD45-erCP, CD73-PE, CD29-PE, CD44-ITC, CD117-PE, and CD90-FITC CD13-E (BD Pharmingen, San Diego, CA);D34-FITC, and CD133/1-PE (Milte-yi, Auburn, CA); CD105-FITC (Sero-
ec, Oxford, UK), BCRP1-FITC (Milli-ore, Bedford, MA); SSEA-1 PE, SSEA-3E, SSEA-4 PE, TRA-1-60 PE, TRA-1-81E, HLA-ABC-FITC, HLA-DR-FITC,D40-FITC, CD80-FITC, and CD86-PE
eBioscience, San Diego, CA). FITC-, PE-,nd PerCP-control isotypes were used asegative controls. Cell fluorescence wasvaluated in a FACScan (Becton Dickin-on, Lincoln Park, NJ) and analyzed withhe CellQuest Pro 9.0 software (Bectonickinson).
n vitro cell differentiation studiesesodermal differentiation
dipogenic differentiation was carriedut with the adipogenic differentiationullet kit (Lonza) according to the man-facturer’s instructions and differentia-
ion was evaluated by Oil Red O stainingSigma, St Louis, MO).
Osteogenic differentiation was per-ormed with the osteogenic differentia-ion bullet kit (Lonza) for 3 weeks. Dif-erentiation was assessed by Alizarin Red
and Sigma Fast 5-bromo-4-chloro--indoyl-phosphate, 4-toluidine salt/4-itro blue tetrazolium chloride stainingSigma).
For chondrogenic differentiation, cellsere cultured for 21 days with the
hondrogenic differentiation bullet kitLonza) supplemented with 10 ng/mLransforming growth factor-�1 (Sigma).ifferentiation was evaluated by indirect
mmunofluorescent staining with anti-odies for aggrecan and chondroitin sul-
hate (Millipore). Additionally, secreted slogy NOVEMBER 2010
xtracellular matrix proteoglycans wereisualized by Alcian Blue staining.Myogenic differentiation was ana-
yzed on 1.9 � 104 cells seeded in fi-ronectin-coated plates. Differentiationas initiated by 10 �M 5=-azacitidine
Sigma) incubation for 24 hours and-14 days in skeletal muscle growth me-ium (Lonza) supplemented with 10g/mL of fibroblast growth factor. Myo-enic differentiation was evaluated bymmunofluorescence with antibodiesor fast skeletal myosin and �-actininSigma).
For cardiogenic differentiation, cellsere cultured in the skeletal musclerowth medium (Lonza) for 41 days andtained with alpha atrial natriureticolypeptide (ANP) antibody (Millipore).
ndodermal differentiationulmonary epithelial differentiation wasccomplished by incubation in small air-ay growth medium (Lonza) for 3-4ays. Differentiated cells were stainedith antibodies to prosurfactant protein(proSP-C) and surfactant protein B
SP-B) (Millipore).
ctodermal differentiationor neurogenic differentiation, cellsere seeded in 6-multiwell plates pre-
oated with poly-L-lysine (Nunc, Glos-rup, Denmark) at a concentration of.29 � 104 cells/cm2. After 24 hours,eurogenic differentiation media weredded.32 Cells were cultured for 24-72ours and stained with anti-� III-tubu-
in and anti-NF200 (Sigma).
elomerase activity (TRAP assay)elomerase activity was analyzed by theRAPeze telomerase detection kit (Mil-
ipore) according to the manufacturer’snstructions. The telomeric repeat am-lification protocol (TRAP) assay is aensitive and efficient polymerase chaineaction (PCR)-based detection methodo detect telomerase activity in humanells and not only the presence of ribo-ucleic acid (RNA) or protein compo-ents of telomerase. First, enzymaticallyctive telomerase present in the cell pro-ein extracts adds a number of telomericepeats (GGTTAG) onto the 3= end of a
ubstrate oligonucloetide (TS).
ppacnhmtS
FhFdAtsfiJ
SGwrtCS1DDDaffTP(
KPwn1thCddua
TtTP
tgttCfi(pi9oi7siss
IClht
isaa(indm
CFwpdaaVceac
A(mil2M
www.AJOG.org Basic Science: Obstetrics Research
Later the extended products are am-lified by PCR generating a ladder ofroducts with 6 base increments startingt 50 nucleotides. Telomerase-positiveells are used as a positive control. As aegative control, cell extracts wereeated before the assay at 85°C for 10inutes, according to the kit instruc-
ions. TRAP products were visualized byYBR Gold staining.
luorescence in situybridization (FISH)ISH analysis was carried out with Posei-on FISH Kit (Kreatech Diagnostics,msterdam, The Netherlands) following
he manufacturer’s instructions. Analy-is was performed on DMSCs isolatedrom 3 male placentas using a Zeiss Ax-oplan2 microscope (Carl Zeiss AG,ena, Germany
hort tandem repeats (STR) analysisenomic deoxyribonucleic acid (DNA)as extracted from 0.5 � 106 cells using a
esin-based procedure (Instagene Ma-rix; Bio-Rad Laboratories, Hercules,A). QF-PCR for the amplification ofTR markers for chromosomes X, Y, 13,8, and 21 (AMXY, X22, HPRT, SRY,XYS218, D13S631, D13S634, D13S258,13S305, D21S1414, D21S1411, D21S1446,21S1437, D18S535, D18S386, D18S391,
nd D18S390) was carried out by an Aneu-ast QF-PCR kit (Genomed Ltd, Kent, UK)ollowing the manufacturer’s instructions.he PCR products were analyzed on ABIRISM 3100-AVANT genetic analyzerApplied Biosystems, Foster City, CA).
aryotype analysislacental-derived cells at passages 15-20ere processed using standard cytoge-etic techniques. Cells were treated with0 �g/mL of Colcemid for 1.5 hours,rypsinized, treated briefly with 0.075 Mypotonic KCl solution, and fixed witharnoy’s fixative. Cells were thenropped on a microscope glass slide andried. Metaphase cells were G bandedsing Wright staining. At least 20 met-phases per sample were karyotyped.
otal RNA extraction and reverseranscription (RT)-PCRotal RNA was isolated using Master-
ure RNA isolation kit (Epicentre Bio- Xechnologies, Madison, WI). One micro-ram of total RNA was used for reverseranscription using ImProm II reverseranscriptase (Promega, Madison, WI).omplementary DNAs were then ampli-ed using MBL Taq DNA polymeraseDominion MBL) and tissue-specificrimers. Amplification was carried out
n a TC3000 thermocycler (Techne) at4°C for 4 minutes, followed by 30 cyclesf 94°C for 30 seconds, specific anneal-
ng temperature for 30 seconds, and2°C for 30 seconds. PCR products wereeparated on 2% agarose gel and visual-zed by SYBR Gold staining. Human-pecific PCR primers and conditions arehown in the Supplementary Table.
mmunostainingells were either fixed with 10% forma-
in for 90 minutes or 4% paraformalde-yde for 10 minutes at room tempera-ure and permeabilized with 0.3% Triton
FIGURE 1Morphology and growth characteri
, Initial adherent culture is a mixture of fibroblawhite arrows). B, Fibroblast-like cells form colonake a confluent culture at approximately day 2
ndividual placenta. The average doubling time frative cell number was 1.15 � 1013 obtained fro3 times, undergoing 21.31 cell population douacias. Decidua multipotent mesenchymal stem cells. Am J O
-100 for 10 minutes. Nonspecific bind- w
NOVEMBER 2010 American
ng was blocked by PBS plus 5% horseerum (Lonza) for 90 minutes. Primaryntibodies were incubated overnightt 4°C and fluorescein isothiocyanateFITC)-conjugated secondary antibod-es for 2 hours at room temperature. Theucleus was counterstained with 4=,6=-iamino-2-phenylindole (Sigma) for 1inute.
ell cryopreservation and thawingifty percent confluent placental cellsere trypsinized, counted, resus-ended in cell culture freezing me-ium-dimethylsulfoxide (Millipore) atconcentration of 1 � 106 cells/mL,
nd added to 2 mL cryovials (Nunc).ials were introduced in a freezing
ontainer (Mr Frosty; Nalgene, Roch-ster, NY), placed overnight at – 80°C,nd transferred to liquid N2. Thawedells in a water bath at 37°C were
s of placenta-derived cells
ike cells (black arrows) and epithelioid-like cellsat approximately day 10. C, Fibroblast-like cellsD and E, Growth characteristics of cells from 1passages 2 to 22 was 129.12 hours. D, Cumu-.57 � 106 initial cells. E, Cells were passageds.
t Gynecol 2010.
stic
st-lies0.omm 4
blingbste
ashed and then cultured and differ-
Journal of Obstetrics & Gynecology 495.e11

ec
RIilAac1smst
cao26ndmptva
ddweC
fCw
mPtma1frlpHca
aaTgw((cws
fmtasaknmpmt
t3p(m(vlDtpet
ucte
aoatf(Diuopio
ec3udgctpsdsMm(ocs
CpclRa
MO
Hl1Ra
MO
Research Basic Science: Obstetrics www.AJOG.org
4
ntiated as described for fresh cellultures.
ESULTSnitial primary culture of adherent cellss a heterogeneous population of epithe-ioid and fibroblast-like cells (Figure 1,). Fibroblast-like cells formed coloniest around day 10 (Figure 1, B) and be-ame confluent at around day 20 (Figure, C). Between the first and the third pas-age, fibroblast-like cells became a ho-
ogeneous population exhibiting thispindle-like morphology until aroundhe 20th to the 22nd passage.
Growth characteristics during theomplete expansion period were evalu-ted (n � 9). An exponential growth wasbserved between the second and the2nd passage. Initial cell number was.88 � 5.23 � 106 and cumulative cellumber was 2.54 � 4.97 � 1014. Averageoubling time was 96 � 18 hours. Theedian total number of passages and cell
opulation doublings were 20, respec-ively. Growth characteristics of 1 indi-idual placenta are shown in Figure 1, Dnd E.
Placental-derived cell phenotype wasetermined by flow cytometry (Appen-ix, Supplementary Figure 1). These cellsere positive for many BM-MSC mark-
rs like CD44, CD90, CD117, CD73,
FIGURE 2Pluripotency marker expressionby RT-PCR analysis
ells from 3 different placentas (lanes 1-3) atassages 12 (lane 2) and 21 (lanes 1 and 3); RTontrol (lane 4); �2-microglobulin was used asoading control of complementary DNA.T, reverse transcriptase; RT-PCR, reverse transcriptase polymer-se chain reaction.
acias. Decidua multipotent mesenchymal stem cells. Am Jbstet Gynecol 2010.
D29, CD13, and CD105 and negative s
95.e12 American Journal of Obstetrics & Gyneco
or hematopoietic markers CD34, CD45,D133, and BCRP1. Moreover, cellsere negative for STRO-1 (not shown).Of note, MSC marker expression wasaintained until the end of the culture.
lacenta-derived cells were negative forhe embryonic stem cells (ESC) surface
arkers SSEA-1, SSEA-4, TRA-1-60,nd TRA-1-81 (Supplementary Figure), but expressed the ESC transcriptionactors Oct-4, Rex1, and organogenesisegulator, GATA-4 (Figure 2). Immuno-ogically, placental-derived cells wereositive for HLA-ABC but negative forLA-DR and the costimulatory mole-
ules CD40L, CD80, and CD86 (Figure 3nd Supplementary Figure 1).
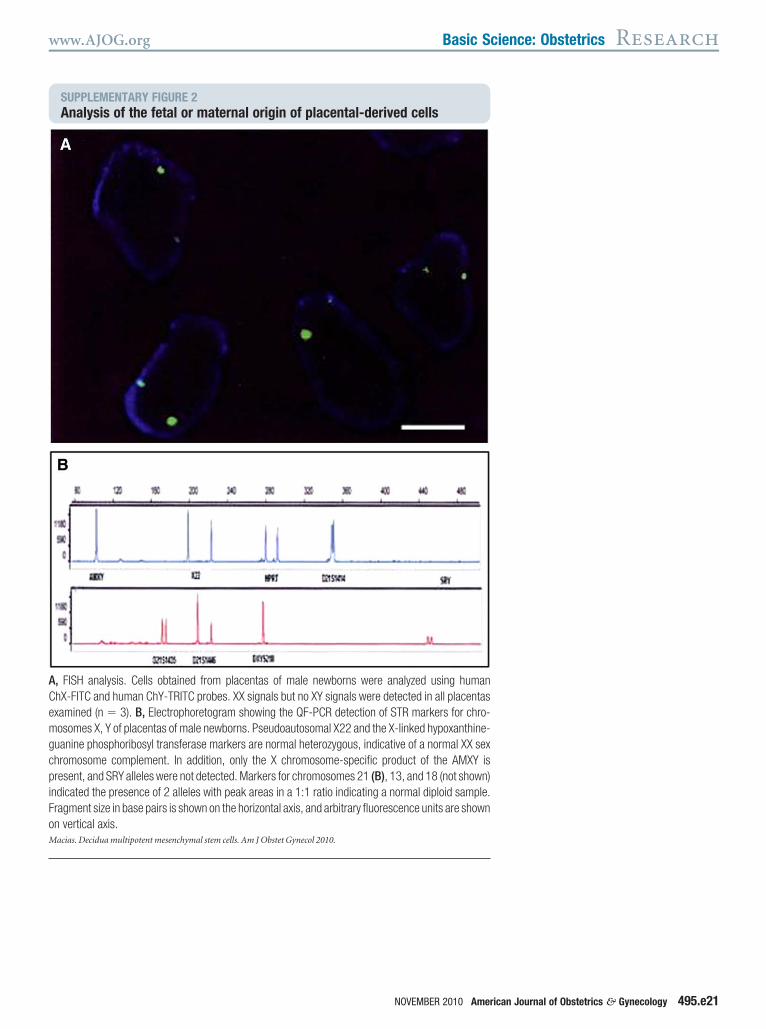
Placental extraembryonic membranesre composed of both fetal (chorion andmnion) and maternal (decidua) cells.o determine the maternal or fetal ori-in, cells from 4 male newborn placentasere analyzed for the presence of male
X, Y) and female (X, X) chromosomesSupplementary Figure 2). In all 4 pla-entas, XX signals but not XY signalsere detected by both FISH and STR
tudies, suggestive of a maternal origin.To validate these results, STR markers
rom placental cells of 1 male and 1 fe-ale newborn were compared with ma-
ernal and fetal blood. Nonadherent cellsnd adherent cells at passages 0 and 7howed a maternal origin for all STRsnalyzed (data not shown). In addition,aryotype analysis of 2 additional maleewborn placentas showed 46 XX chro-osomes. Considering the morphology,
henotype, immunological profile, andaternal origin, we named the placen-
al-derived cells as DMSCs.Because Oct-4 expression is restricted
o cells capable of differentiating into theembryonic layers, differentiation ca-
acity of DMSCs along the mesodermalosteocytes, adipocytes, chondrocytes,yocytes, cardiomyocytes), endodermal
respiratory epithelial cells [ie, type II al-eolar cells]), and ectodermal (neurons)ineages was evaluated (Figures 4 and 5).
MSC control cultures did not expresshe differentiation markers or their ex-ression was increased under the differ-ntiation culture conditions, suggestinghat DMSCs differentiated into the tis-
ues studied. Besides, dermal fibroblasts Elogy NOVEMBER 2010
sed as a negative control neitherhanged morphology under differentia-ion conditions nor expressed the differ-ntiation markers (data not shown).
Adipogenic differentiation was evalu-ted by visualization of lipid-rich vacu-les inside the cells (Figure 4, B1 and B2)nd expression of peroxisome prolifera-or-activated receptor (PPAR)-�-2 andatty acid binding protein (FABP4, AP4)Figure 5). On osteogenic conditions,MSCs fibroblast morphology changed
nto a cobblestone-like appearance (Fig-re 4, C1), showed calcium crystal dep-sition (Figure 4, C2) and high alkalinehosphatase activity (not shown), and
ncreased expression of osteonectin andsteocalcin genes (Figure 5).Under chondrogenic conditions, cells
xhibited an important morphologichange associated with the formation of-dimensional extracellular matrix (Fig-re 4, D1), positive staining for chon-roitin sulphate (Figure 4, D2) and ag-recan (not shown), and expression ofollagen II and Sox9 (Figure 5). Addi-ionally, secreted extracellular matrixroteoglycans were also stained (nothown). Occasionally cells gathered andetached from the culture plate, formingpherical cell masses with shiny surfaces.
yogenic differentiation caused a dra-atic change in DMSCs morphology
Figure 4, E1) and increased expressionf myogenin (Figure 5). Differentiatedells were positive for fast skeletal myo-in (not shown) and �-actinin (Figure 4,
FIGURE 3Immunogenic characterizationof placenta-derived cells
uman leukocyte antigen molecules were ana-yzed by RT-PCR in 4 different placentas (lanes-4); RT (control lane 5).T, reverse transcriptase; RT-PCR, reverse transcriptase polymer-se chain reaction.
acias. Decidua multipotent mesenchymal stem cells. Am Jbstet Gynecol 2010.
2). Cardiomyocyte differentiation was

M
www.AJOG.org Basic Science: Obstetrics Research
FIGURE 4Differentiation potential of DMSCs into tissues from the 3 germ layers
acias. Decidua multipotent mesenchymal stem cells. Am J Obstet Gynecol 2010.
NOVEMBER 2010 American Journal of Obstetrics & Gynecology 495.e13

ogcgd(m
v
icaGt4man
tmlit
drgmrwcpeDdfsaSttc
onsdsrcDs
m1sXm(a
FaaDMEecdfA
M
T(fogetdplAR
M
Research Basic Science: Obstetrics www.AJOG.org
4
btained 40 days after initiating myo-enic culture. Although no beating myo-ytes were observed, cells expressedenes and proteins associated with car-iac tissue, like atrial natriuretic peptideFigures 4, F1 and F2, and 5) and ho-eobox Nkx2.5 (Figure 5).DMSCs differentiation into type II al-
eolar cells was evaluated for a more ep-
igure 4. A, DMSCs under proliferation culture cond B2, by staining with Oil Red O (Sigma, St Louind C2, calcium deposition was evaluated by AMSCs associated with the formation of 3-dimeyogenic differentiation caused a dramatic chan2, positive immunostaining for �-actinin. F1,valuated by positive staining with alpha-ANP anytoplasmic organelles that resemble lamellar bramatic change in morphology and H2, positiveor B1 and G1, 50 �m.NP, atrial natriuretic polypeptide; DMSCs, decidua-derived mese
FIGURE 5Expression of tissue specific genesbefore and after differentiation
issue-specific genes were analyzed by RT-PCRlanes 2-4, n � 3). Adipogenic differentiation waactor and fatty acid binding protein (FABP4, AP4)f osteonectin and osteocalcin genes. Chondrogen II and Sox9. Myogenic differentiation increasntiation was observed by expression of ANP andiation was demonstrated by the expression ofifferentiation media induced the expression ofhate dehydrogenase (GAPDH) expression was u
ane represents the RT control (lane 5).NP, atrial natriuretic polypeptide; DMSCs, decidua-derived meseT, reverse transcriptase; RT-PCR, reverse transcriptase polymer
acias. Decidua multipotent mesenchymal stem cells. Am J O
acias. Decidua multipotent mesenchymal stem cells. Am J Obste
95.e14 American Journal of Obstetrics & Gyneco
thelioid morphology of cells showingytoplasmic organelles with the appear-nce of lamellar bodies (Figure 4,1). Respiratory-epithelial differentia-
ion was confirmed by proSP-C (Figure, G2) and SP-B (data not shown) im-unostaining and expression of SP-B
nd GATA-6 genes (Figure 5). Undereurogenic conditions, DMSCs changed
ions. B1, Adipogenic differentiation was evaluateO). C1, Under osteogenic conditions, DMSCs charin Red S staining. D1, Chondrogenic differentinal extracellular matrix and D2, positive immunon the morphology of the DMSCs to a myofibroblasdiomyocyte differentiation was obtained after 4dy. G1, Pulmonary epithelioid morphology wass (arrows) and G2, positive immunostaining for
ning to �-III tubulin. Control cells were clearly neg
mal stem cells; proSP-C, prosurfactant protein C.
DMSCs
fore (lane 1) and after differentiation of DMSCsaluated by expression of PARP�-2 transcriptionteogenic differentiation increases the expressiondifferentiation induced the expression of colla-
the expression of myogenin. Cardiogenic differ-meobox Nkx2.5. Respiratory epithelial differen-actant protein B and GATA-6 genes. NeuronalII tubulin and NF-200. Glyceraldehyde-3-phos-as loading control of complementary DNA. Right
mal stem cells; PARP, peroxisome prolieratior-activated receptor;hain reaction.
t Gynecol 2010.
t Gynecol 2010.
logy NOVEMBER 2010
o a more rounded, highly refractileorphology with prominent process-
ike extensions (Figure 4) and were pos-tive to �-III tubulin and NF-200 pro-eins (Figures 4, H2 and 5).
Significantly, nearly 100% of DMSCsifferentiated into the desired cell types,egardless of the passage number, sug-esting a homogeneous populationaintained during the entire culture pe-
iod. However, to discard that DMSCsere heterogeneous cell populations
ontaining a mixture of progenitors ca-able of differentiation into certain lin-ages and to prove the “stemness” ofMSCs, we obtained 4 clone lines from 2ifferent placentas and tested their dif-
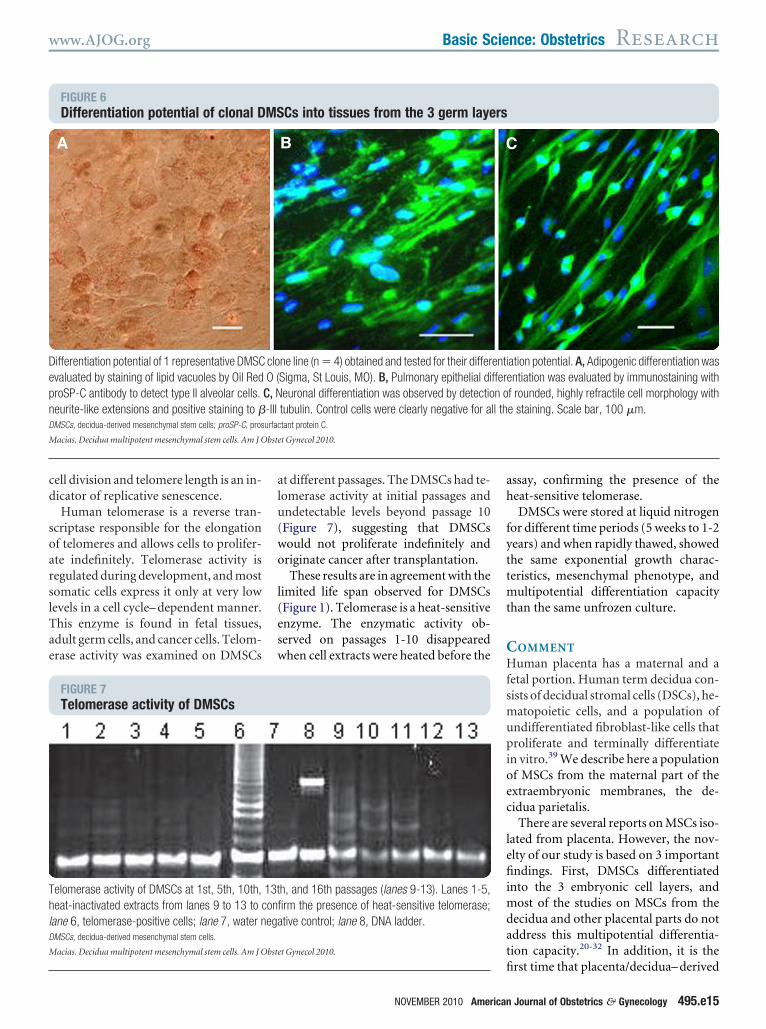
erentiation potential. All 4 cloneshowed differentiation capacity to deriv-tives of all germ layers (Figure 6 andupplementary Figure 3), suggestinghat DMSCs are a homogeneous popula-ion of multipotent mesenchymal stemells.
Future clinical use of DMSCs dependsn their expansion to obtain enough cellumbers for transplantation withoutpontaneous transformation. DMSCsemonstrated a decrease in their expan-ion capacity at around passage 20 whileetaining their morphology and mesen-hymal phenotype, suggesting thatMSCs do not display an aptitude for
pontaneous transformation.To confirm these observations, chro-osome stability of DMSCs at passages
5-20 was determined (n � 3). The re-ults showed a normal karyotype of 46X chromosomes without any nu-erical and/or structural abnormalities
Supplementary Figure 4). Telomeresre progressively shortened with each
the visualization of lipid vacuoles inside the cellsd from a fibroblast to a cobblestone morphology,n induced an important morphologic change inrescence staining for chondroitin sulphate. E1,orphology, as evaluated by light microscopy andays of incubation in myogenic media and F2,
erved by light microscopy for the appearance ofSP-C. H1, Neuronal induction media caused a
ve for these staining. Scale bar, 200 �m, except
ndit d bys, M ngeliza ationsio fluoge i t mCar 0 dtibo obsodie prostai ati
nchy
on
bes ev. Osenicedho
surf�-Ised
nchyase c
bste
cd
soarslTae
alu(wo
l(esw
ah
fyttmt
CHfsmupioec
lefiimdat
DepnD
M
ThlD
M
www.AJOG.org Basic Science: Obstetrics Research
ell division and telomere length is an in-icator of replicative senescence.Human telomerase is a reverse tran-
criptase responsible for the elongationf telomeres and allows cells to prolifer-te indefinitely. Telomerase activity isegulated during development, and mostomatic cells express it only at very lowevels in a cell cycle– dependent manner.his enzyme is found in fetal tissues,dult germ cells, and cancer cells. Telom-rase activity was examined on DMSCs
FIGURE 6Differentiation potential of clonal D
ifferentiation potential of 1 representative DMSCvaluated by staining of lipid vacuoles by Oil RedroSP-C antibody to detect type II alveolar cells.eurite-like extensions and positive staining to �MSCs, decidua-derived mesenchymal stem cells; proSP-C, pros
acias. Decidua multipotent mesenchymal stem cells. Am J O
FIGURE 7Telomerase activity of DMSCs
elomerase activity of DMSCs at 1st, 5th, 10th,eat-inactivated extracts from lanes 9 to 13 to c
ane 6, telomerase-positive cells; lane 7, water nMSCs, decidua-derived mesenchymal stem cells.
acias. Decidua multipotent mesenchymal stem cells. Am J O
t different passages. The DMSCs had te-omerase activity at initial passages andndetectable levels beyond passage 10Figure 7), suggesting that DMSCsould not proliferate indefinitely andriginate cancer after transplantation.These results are in agreement with the
imited life span observed for DMSCsFigure 1). Telomerase is a heat-sensitivenzyme. The enzymatic activity ob-erved on passages 1-10 disappearedhen cell extracts were heated before the
Cs into tissues from the 3 germ laye
ne line (n � 4) obtained and tested for their differSigma, St Louis, MO). B, Pulmonary epithelial dieuronal differentiation was observed by detectiotubulin. Control cells were clearly negative for atant protein C.
t Gynecol 2010.
h, and 16th passages (lanes 9-13). Lanes 1-5,rm the presence of heat-sensitive telomerase;tive control; lane 8, DNA ladder.
t Gynecol 2010.
fi
NOVEMBER 2010 American
ssay, confirming the presence of theeat-sensitive telomerase.DMSCs were stored at liquid nitrogen
or different time periods (5 weeks to 1-2ears) and when rapidly thawed, showedhe same exponential growth charac-eristics, mesenchymal phenotype, and
ultipotential differentiation capacityhan the same unfrozen culture.
OMMENTuman placenta has a maternal and a
etal portion. Human term decidua con-ists of decidual stromal cells (DSCs), he-
atopoietic cells, and a population ofndifferentiated fibroblast-like cells thatroliferate and terminally differentiate
n vitro.39 We describe here a populationf MSCs from the maternal part of thextraembryonic membranes, the de-idua parietalis.
There are several reports on MSCs iso-ated from placenta. However, the nov-lty of our study is based on 3 importantndings. First, DMSCs differentiated
nto the 3 embryonic cell layers, andost of the studies on MSCs from the
ecidua and other placental parts do notddress this multipotential differentia-ion capacity.20-32 In addition, it is the
ation potential. A, Adipogenic differentiation wasntiation was evaluated by immunostaining with
f rounded, highly refractile cell morphology withe staining. Scale bar, 100 �m.
MS rs
clo entiO ( ffere
C, N n o-III ll thurfac
bste
13tonfiega
bste
rst time that placenta/decidua– derived
Journal of Obstetrics & Gynecology 495.e15

stMcDttpfMt
cDetSmCeDatfl
mTcpOtGi
toTtstsrHustieM
cfbsuc
led(djo
pptcdvivsa
fotsoDtsor
sctocmscsnDeosfcc
caiiim
cliipt
wftcomtCpwe
mcsehhoflaeiDc
ftDmcwsag
WCpusoOmbWit
Research Basic Science: Obstetrics www.AJOG.org
4
tem cells are differentiated into alveolarype II cells. Second, DMSCs are a true
SC population demonstrated bylonally expanded cell lines. Third,MSCs coexpressed a set of transcrip-
ion factors under regular culture condi-ions, suggesting a more immaturehenotype and higher life span and dif-
erentiation potential than other adultSCs and other placental MSCs from fe-
al origin.DMSCs are a different population of
ells when compared with DSCs becauseMSCs are multipotent cells and do not
xpress HLA-DR, CD34, and STRO-1 orhe pluripotency markers SSEA-1,SEA-4, and TRA-1-81.29,40,41 First-tri-ester human decidua cells expressedD146,26 a marker of perivascular and
ndothelial cells42 not expressed inMSCs, suggesting a different cell type,
t a different stage of differentiation, orhe isolation and culture conditions in-uence on the markers expressed.Decidua basalis MSCs express ESCarkers like SSEA-1, -3, and -4 andRA-1-81.27 Under our isolation andulture conditions, DMSCs do not ex-ress these ESC markers but expressct-4 and Rex-1, two transcription fac-
ors involved in pluripotency; andATA-4, a transcription factor involved
n organogenesis.These data may be indicative of an in-
ermediate phenotype between embry-nic and adult stem cells for DMSCs.he coexpression of these 3 transcrip-
ion factors is not often reported in adulttem cells, except under restrictive cul-ure conditions (low serum and low den-ity), resembling the in vivo microenvi-onment of primitive stem cells.43-49
owever, DMSCs express these markersnder regular culture conditions of higherum and high density, suggesting a dis-inctive MSC population with a moremmature phenotype and higher differ-ntiation potential than other adult
SCs.9,47,49,50
To conclusively assert whether theells described here are different or notrom other decidual stem cells, it woulde necessary to perform a comparativetudy between these cell populationssing similar isolation and culture
onditions. d95.e16 American Journal of Obstetrics & Gyneco
Besides mesodermic (bone, fat, carti-age, skeletal, and cardiac muscle) differ-ntiation, DMSCs differentiated to ecto-ermal (neural-like) and endodermallung) cells. Several reports describedifferentiation of ESCs to lung cells, but
ust a few reports described this capacityn adult stem cells.51-53
To our knowledge, this is the first re-ort showing differentiation of humanlacenta-derived MSCs to alveolar epi-helial type II cells, in both the primaryulture of DMSCs and the single-cell–erived clones. We did not address the inivo differentiation capacity of DMSCsn this study. However, the elevated initro differentiation potential suggests aimilar or even higher in vivo differenti-tion capacity.23
Other studies showed a high plasticityor mesenchymal-like cells derived fromther parts of human placenta,23,26,32 al-hough most of these studies did nothow differentiation into the 3 embry-nic layers as shown here for DMSCs.MSCs display a significant differentia-
ion potential into cells of all germ layers,imilar to amniotic fluid MSCs,14 amni-tic epithelial cells,21 and endometrialegenerative cells.54
Under normal culture conditions, noignificant differences in differentiationapacity of DMSCs were observed be-ween early and late passages, as previ-usly reported for cells obtained fromentral placenta.24,31,55,56 In addition, al-ost the entire culture showed expres-
ion of the differentiation markers, andonsistent results were obtained fromeveral placentas, suggesting a homoge-eous stem cell population for DMSCs.efinitely, the true multilineage differ-
ntiation potential of DMSCs was dem-nstrated by cloning out 4 cell lines andhowing that these clones can each dif-erentiate into cells of the 3 germ layers,onfirming the true mesenchymal stemell nature of DMSCs.7,31
BM-MSCs are immunomodulatoryells to be used in mismatched recipientsnd to reduce any previously existingmmune reaction.57-59 Likewise, cellssolated from fetal membranes are hypo-mmunogenic and exhibit immuno-
odulatory properties.10,35,60,61 DMSCs
o not express major histocompatibility tlogy NOVEMBER 2010
omplex class II and the T-cell costimu-atory molecules, suggesting that they aremmunoprivileged cells to be toleratedn allogeneic transplantation61,62 androbably involved in the fetomaternalolerance.28
These maternal-derived stem cellsould be an autologous source of MSCs
or future use on pelvic floor dysfunc-ions such as stress urinary and fecal in-ontinence and pelvic organ prolapse63
r for preemptive storage in autoim-une diseases previously developed by
he mother such as multiple sclerosis androhn’s disease.38 This is a valuable as-ect because cells from a fetal originould not be suitable for a regular pre-
mptive clinical storage.Placenta-derived MSCs from fetalembranes (amnion and chorion) and
horionic villi have a limited lifepan,23,24,64 making it difficult to obtainnough cell numbers for transplants inumans. Conversely, DMSCs have aigher life span to obtain large amountf cells but without exceeding the Hay-ick limit of 50 population doublingsnd exhibiting genomic stability underxtended culture periods and a decreasen telomerase activity, suggesting that
MSCs can be a safe product for futurelinical applications.
We did not assess the risk of teratomasormation, but given that DMSCs even-ually stopped growing indicates thatMSCs do not have an intrinsic transfor-ation potential.65 Furthermore, DMSCs
anbecryopreservedfor long-termstorageithout loosing their properties and tryp-
in used to isolate DMSCs is currentlyvailable as a guanosine monophosphate–rade enzyme.66
According to the first internationalorkshop on Placenta Derived Stem
ells, it is important to clearly define theart of the placenta and the methodssed to isolate the cells.10 Our resultshowed that DMSCs are from maternalrigin (ie, from the decidua parietalis).ther groups have isolated MSCs fromaternal origin (ie, from the decidua
asalis25,27,28,56 and parietalis26,28,29).e can not discard the fact that freshly
solated cells were a mixture of ma-ernal and fetal cells, but after days in cul-
ure, only maternal cells survive and
gfm
Pmmcstelfvgasco
AWcog
R1imCt2CIt3aJ4s5M6cp7fJ8tc9fo41Cotr
1CE1Zplm1Mht1at11flm71Mham21Bs21spc21eh12Dfa12Sl2Nci22Apn22ahM2Crc2
u22tph22dsp2tcR3cdB3Icp3Yaft3KtmD3gdc3phm3ctJ3Me3mtT3SmslB4BmrH4Oc
www.AJOG.org Basic Science: Obstetrics Research
row, as observed for MSCs isolatedrom chorionic villi and chorionic
embranes.24,25,28,31,55,56
The first international Workshop onlacenta Derived Stem Cells defined theinimal criteria that human amnioticesenchymal stromal cells and human
horionic mesenchymal stromal cellshould fulfill.10 DMSCs follow all ofhem except for the fetal origin. How-ver, the fact that stem cells can be iso-ated from both fetal and maternal originrom the same tissue is an important ad-antage for their future use in autolo-ous and allogeneic transplantation. Inddition, placental tissue is an unre-tricted available material without ethi-al concerns to obtain elevated numbersf human adult stem cells. f
CKNOWLEDGMENTe thank Dr Irene Vegh (Centro de Investiga-
ion, Hospital 12 de Octubre) for critical readingf the manuscript and Manuel Macias for En-lish grammar revision.
EFERENCES. Dominici M, Le Blanc K, Mueller I, et al. Min-
mal criteria for defining multipotent mesenchy-al stromal cells. The International Society forellular Therapy position statement. Cyto-
herapy 2006;8:315-7.. Horwitz EM, Le Blanc K, Dominici M, et al.larification of the nomenclature for MSC: the
nternational Society for Cellular Therapy posi-ion statement. Cytotherapy 2005;7:393-5.. Liu ZJ, Zhuge Y, Velazquez OC. Traffickingnd differentiation of mesenchymal stem cells.Cell Biochem 2009;106:984-91.. Fehrer C, Lepperdinger G. Mesenchymaltem cell aging. Exp Gerontol 2005;40:926-30.. Vaananen HK. Mesenchymal stem cells. Anned 2005;37:469-79.. Hemberger M, Yang W, Natale D, et al. Stemells from fetal membranes—a workshop re-ort. Placenta 2008;29(Suppl A):S17-9.. Marcus AJ, Woodbury D. Fetal stem cells
rom extra-embryonic tissues: do not discard.Cell Mol Med 2008;12:730-42.. Matikainen T, Laine J. Placenta—an alterna-ive source of stem cells. Toxicol Appl Pharma-ol 2005;207:544-9.. Pappa KI, Anagnou NP. Novel sources of
etal stem cells: where do they fit on the devel-pmental continuum? Regen Med 2009;4:23-33.0. Parolini O, Alviano F, Bagnara GP, et al.oncise review: isolation and characterizationf cells from human term placenta: outcome ofhe first international Workshop on Placenta De-
ived Stem Cells. Stem Cells 2008;26:300-11. F1. Broxmeyer HE, Srour E, Orschell C, et al.ord blood stem and progenitor cells. Methodsnzymol 2006;419:439-73.2. McNiece IK, Almeida-Porada G, Shpall EJ,anjani E. Ex vivo expanded cord blood cellsrovide rapid engraftment in fetal sheep but
ack long-term engrafting potential. Exp He-atol 2002;30:612-6.3. Bossolasco P, Montemurro T, Cova L, et al.olecular and phenotypic characterization of
uman amniotic fluid cells and their differentia-ion potential. Cell Res 2006;16:329-36.4. De Coppi P, Bartsch G Jr, Siddiqui MM, etl. Isolation of amniotic stem cell lines with po-ential for therapy. Nat Biotechnol 2007;25:00-6.5. Kim J, Lee Y, Kim H, et al. Human amnioticuid-derived stem cells have characteristics ofultipotent stem cells. Cell Prolif 2007;40:5-90.6. Roubelakis MG, Pappa KI, Bitsika V, et al.olecular and proteomic characterization of
uman mesenchymal stem cells derived frommniotic fluid: comparison to bone marrowesenchymal stem cells. Stem Cells Dev007;16:931-52.7. Karahuseyinoglu S, Cinar O, Kilic E, et al.iology of stem cells in human umbilical cordtroma: in situ and in vitro surveys. Stem Cells007;25:319-31.8. Sarugaser R, Lickorish D, Baksh D, Hos-eini MM, Davies JE. Human umbilical corderivascular (HUCPV) cells: a source of mesen-hymal progenitors. Stem Cells 2005;23:20-9.9. Wang HS, Hung SC, Peng ST, et al. Mes-nchymal stem cells in the Wharton’s jelly of theuman umbilical cord. Stem Cells 2004;22:330-7.0. Marcus AJ, Coyne TM, Rauch J, Woodbury, Black IB. Isolation, characterization, and dif-
erentiation of stem cells derived from the ratmniotic membrane. Differentiation 2008;76:30-44.1. Miki T, Lehmann T, Cai H, Stolz DB, StromC. Stem cell characteristics of amniotic epithe-
ial cells. Stem Cells 2005;23:1549-59.2. Tamagawa T, Oi S, Ishiwata I, Ishikawa H,akamura Y. Differentiation of mesenchymalells derived from human amniotic membranes
nto hepatocyte-like cells in vitro. Hum Cell007;20:77-84.3. Portmann-Lanz CB, Schoeberlein A, Huber, et al. Placental mesenchymal stem cells asotential autologous graft for pre- and perinataleuroregeneration. Am J Obstet Gynecol006;194:664-73.4. Soncini M, Vertua E, Gibelli L, et al. Isolationnd characterization of mesenchymal cells fromuman fetal membranes. J Tissue Eng Regened 2007;1:296-305.5. Barlow S, Brooke G, Chatterjee K, et al.omparison of human placenta- and bone mar-
ow-derived multipotent mesenchymal stemells. Stem Cells Dev 2008;17:1095-107.6. Dimitrov R, Kyurkchiev D, Timeva T, et al.
irst-trimester human decidua contains a pop- RNOVEMBER 2010 American
lation of mesenchymal stem cells. Fertil Steril010;93:210-9.7. Huang YC, Yang ZM, Chen XH, et al. Isola-ion of mesenchymal stem cells from humanlacental decidua basalis and resistance toypoxia and serum deprivation. Stem Cell Rev009;5:247-55.8. In’t Anker PS, Scherjon SA, Kleijburg-vaner Keur C, et al. Isolation of mesenchymaltem cells of fetal or maternal origin from humanlacenta. Stem Cells 2004;22:1338-45.9. Strakova Z, Livak M, Krezalek M, Ihna-ovych I. Multipotent properties of myofibroblastells derived from human placenta. Cell Tissuees 2008;332:479-88.0. Chang CM, Kao CL, Chang YL, et al. Pla-enta-derived multipotent stem cells induced toifferentiate into insulin-positive cells. Biochemiophys Res Commun 2007;357:414-20.1. Fukuchi Y, Nakajima H, Sugiyama D, Hirose
, Kitamura T, Tsuji K. Human placenta-derivedells have mesenchymal stem/progenitor cellotential. Stem Cells 2004;22:649-58.2. Igura K, Zhang X, Takahashi K, Mitsuru A,amaguchi S, Takashi TA. Isolation and char-cterization of mesenchymal progenitor cellsrom chorionic villi of human placenta. Cyto-herapy 2004;6:543-53.3. Brooke G, Tong H, Levesque JP, Atkinson. Molecular trafficking mechanisms of multipo-
ent mesenchymal stem cells derived from hu-an bone marrow and placenta. Stem Cellsev 2008;17:929-40.4. Jones BJ, Brooke G, Atkinson K, McTag-art SJ. Immunosuppression by placental in-oleamine 2,3-dioxygenase: a role for mesen-hymal stem cells. Placenta 2007;28:1174-81.5. Li C, Zhang W, Jiang X, Mao N. Human-lacenta-derived mesenchymal stem cells in-ibit proliferation and function of allogeneic im-une cells. Cell Tissue Res 2007;330:437-46.6. Baksh D, Song L, Tuan RS. Adult mesen-hymal stem cells: characterization, differentia-ion, and application in cell and gene therapy.Cell Mol Med 2004;8:301-16.7. El-Badri NS, Maheshwari A, Sanberg PR.esenchymal stem cells in autoimmune dis-
ase. Stem Cells Dev 2004;13:463-72.8. Tyndall A, Uccelli A. Multipotent mesenchy-al stromal cells for autoimmune diseases:
eaching new dogs old tricks. Bone Marrowransplant 2009;43:821-8.9. Richards RG, Brar AK, Frank GR, HartmanM, Jikihara H. Fibroblast cells from term hu-an decidua closely resemble endometrial
tromal cells: induction of prolactin and insulin-ike growth factor binding protein-1 expression.iol Reprod 1995;52:609-15.0. Garcia-Pacheco JM, Oliver C, Kimatrai M,lanco FJ, Olivares EG. Human decidual stro-al cells express CD34 and STRO-1 and are
elated to bone marrow stromal precursors. Molum Reprod 2001;7:1151-7.1. Oliver C, Cowdrey N, Abadia-Molina AC,livares EG. Antigen phenotype of cultured de-idual stromal cells of human term decidua. J
eprod Immunol 1999;45:19-30.Journal of Obstetrics & Gynecology 495.e17

4tct4mebft74Pmpct4HeaS4Pmpho4tgh4WecL
4Rih25bn15Dmi5mdC5atp25tl5ahti15Mm5do
5ft5Map6mct6csp26oS6pt56�ae26Hsm46Tsw
Research Basic Science: Obstetrics www.AJOG.org
4
2. Schwab KE, Gargett CE. Co-expression ofwo perivascular cell markers isolates mesen-hymal stem-like cells from human endome-rium. Hum Reprod 2007;22:2903-11.3. Battula VL, Bareiss PM, Treml S, et al. Hu-an placenta and bone marrow derived mes-
nchymal stem cells cultured in serum-free,asic FGF-containing medium express cell sur-
ace frizzled-9 and SSEA-4 and give rise to mul-ilineage differentiation. Differentiation 2007;5:279-91.4. D’Ippolito G, Diabira S, Howard GA, Menei, Roos BA, Schiller PC. Marrow-isolated adultultilineage inducible (MIAMI) cells, a uniqueopulation of postnatal young and old humanells with extensive expansion and differentia-ion potential. J Cell Sci 2004;117:2971-81.5. Guillot PV, Gotherstrom C, Chan J, Kurata, Fisk NM. Human first-trimester fetal MSCxpress pluripotency markers and grow fasternd have longer telomeres than adult MSC.tem Cells 2007;25:646-54.6. Pochampally RR, Smith JR, Ylostalo J,rockop DJ. Serum deprivation of humanarrow stromal cells (hMSCs) selects for a sub-opulation of early progenitor cells with en-anced expression of OCT-4 and other embry-nic genes. Blood 2004;103:1647-52.7. Poloni A, Rosini V, Mondini E, et al. Charac-erization and expansion of mesenchymal pro-enitor cells from first-trimester chorionic villi ofuman placenta. Cytotherapy 2008;10:690-7.8. Ratajczak MZ, Machalinski B, Wojakowski, Ratajczak J, Kucia M. A hypothesis for an
mbryonic origin of pluripotent Oct-4(�) stemells in adult bone marrow and other tissues.
eukemia 2007;21:860-7. r95.e18 American Journal of Obstetrics & Gyneco
9. Roche S, Richard MJ, Favrot MC. Oct-4,ex-1, and Gata-4 expression in human MSC
ncrease the differentiation efficiency but notTERT expression. J Cell Biochem 2007;101:71-80.0. Li C, Zhou J, Shi G, et al. Pluripotency cane rapidly and efficiently induced in human am-iotic fluid-derived cells. Hum Mol Genet 2009;8:4340-9.1. Berger MJ, Adams SD, Tigges BM, et al.ifferentiation of umbilical cord blood-derivedultilineage progenitor cells into respiratory ep-
thelial cells. Cytotherapy 2006;8:480-7.2. Carraro G, Perin L, Sedrakyan S, et al. Hu-an amniotic fluid stem cells can integrate andifferentiate into epithelial lung lineages. Stemells 2008;26:2902-11.3. Kannan S, Wu M. Respiratory stem cellsnd progenitors: overview, derivation, differen-iation, carcinogenesis, regeneration and thera-eutic application. Curr Stem Cell Res Ther006;1:37-46.4. Meng X, Ichim TE, Zhong J, et al. Endome-rial regenerative cells: a novel stem cell popu-ation. J Transl Med 2007;5:57.5. Semenov OV, Koestenbauer S, Riegel M, etl. Multipotent mesenchymal stem cells fromuman placenta: critical parameters for isola-ion and maintenance of stemness aftersolation. Am J Obstet Gynecol 2010;202:93.e1-13.6. Wulf GG, Viereck V, Hemmerlein B, et al.esengenic progenitor cells derived from hu-an placenta. Tissue Eng 2004;10:1136-47.7. Kode JA, Mukherjee S, Joglekar MV, Har-ikar AA. Mesenchymal stem cells: immunobi-logy and role in immunomodulation and tissue
egeneration. Cytotherapy 2009;11:377-91. clogy NOVEMBER 2010
8. Le Blanc K. Immunomodulatory effects ofetal and adult mesenchymal stem cells. Cyto-herapy 2003;5:485-9.9. Ringden O, Uzunel M, Rasmusson I, et al.esenchymal stem cells for treatment of ther-
py-resistant graft-versus-host disease. Trans-lantation 2006;81:1390-7.0. Bailo M, Soncini M, Vertua E, et al. Engraft-ent potential of human amnion and chorion
ells derived from term placenta. Transplanta-ion 2004;78:1439-48.1. Chang CJ, Yen ML, Chen YC, et al. Pla-enta-derived multipotent cells exhibit immuno-uppressive properties that are enhanced in theresence of interferon-gamma. Stem Cells006;24:2466-77.2. Yen BL, Huang HI, Chien CC, et al. Isolationf multipotent cells from human term placenta.tem Cells 2005;23:3-9.3. Gargett BE, Chan RW. Endometrial stem/rogenitor cells and proliferative disorders ofhe endometrium. Minerva Ginecol 2006;58:11-26.4. Okamoto K, Miyoshi S, Toyoda M, et al.Working’ cardiomyocytes exhibiting plateauction potentials from human placenta-derivedxtraembryonic mesodermal cells. Exp Cell Res007;313:2550-62.5. Schallmoser K, Bartmann C, Rohde E, et al.uman platelet lysate can replace fetal bovineerum for clinical-scale expansion of functionalesenchymal stromal cells. Transfusion 2007;7:1436-46.6. Alviano F, Fossati V, Marchionni C, et al.erm amniotic membrane is a high throughputource for multipotent mesenchymal stem cellsith the ability to differentiate into endothelial
ells in vitro. BMC Dev Biol 2007;7:11.
A
J Obstet Gynecol 2010.
www.AJOG.org Basic Science: Obstetrics Research
PPENDIX
SUPPLEMENTARY TABLERT-PCR primer sequences, annealiGene Temperature, °C
�2-microglobulin 58.3...................................................................................................................
GAPDH 57.3...................................................................................................................
Oct-4 59.4...................................................................................................................
Rex-1 57.3...................................................................................................................
GATA-4 58.3...................................................................................................................
HLA ABC (I) 60.4...................................................................................................................
HLA DR (II) 58.3...................................................................................................................
AP4 57.3...................................................................................................................
PPAR�2 57.3...................................................................................................................
Osteocalcin 59.4...................................................................................................................
Osteonectin 58.3...................................................................................................................
Collagen type II 59.4...................................................................................................................
Sox 9 57.3...................................................................................................................
Myogenin 59.0...................................................................................................................
ANP 62.7...................................................................................................................
NKx2.5 57.3...................................................................................................................
GATA-6 57.3...................................................................................................................
Surfactant B 58.3...................................................................................................................
�III tubulin 59.4...................................................................................................................
Neurofilament 200 56.3...................................................................................................................
ANP, atrial natriuretic polypeptide; GAPDH, glyceraldehyde-3-
Macias. Decidua multipotent mesenchymal stem cells. Am
ng temperature, and products sizeFrangment, pb Primer sense (5’¡3’) Primer reverse (5’¡3’)
301 tgactttgtcacagcccaag tctctgctccccacctctaa.............................................................................................................................................................................................................................................................
512 gagtcaacggatttggtcgt tgtggtcatgagtccttcca.............................................................................................................................................................................................................................................................
208 gagaaggagaagctggagca agctaagctgcagagcctca.............................................................................................................................................................................................................................................................
403 ggaggaatacctggcattga ttaggatgtgggctttcagg.............................................................................................................................................................................................................................................................
576 tccctcttccctcctcaaat tgggttaagtgcccctgtag.............................................................................................................................................................................................................................................................
345 gcggctactacaaccagagc gtggcctcatggtcagagat.............................................................................................................................................................................................................................................................
220 agacaagttcaccccaccag agcatcaaactcccagtgct.............................................................................................................................................................................................................................................................
181 tactgggccaggaatttgac gtggaagtgacgcctttcat.............................................................................................................................................................................................................................................................
586 tgcagtggggatgtctcata gggctccataaagtcaccaa.............................................................................................................................................................................................................................................................
175 gtgcagagtccagcaaaggt tcagccaactcgtcacagtc.............................................................................................................................................................................................................................................................
490 cacgatgttgtcaaggatgg gggagggtgaagaaaaggag.............................................................................................................................................................................................................................................................
426 ggctcccagaacatcaccta tctgcccagttcaggtctct.............................................................................................................................................................................................................................................................
244 ttgagccttaaaacggtgct ctggtgttctgagaggcaca.............................................................................................................................................................................................................................................................
429 cctacagatgcccacaacct gaggccgcgttatgataaaa.............................................................................................................................................................................................................................................................
208 tgaagataacagccagggaggaca agaggcgaggaagtcaccatcaaa.............................................................................................................................................................................................................................................................
302 acgcccttctcagtcaaaga ttttcggctctagggtcctt.............................................................................................................................................................................................................................................................
259 actaacccacaggcaggttg ggtacaaaacggctccaaaa.............................................................................................................................................................................................................................................................
244 gtccagctgcaaaggaaaag agaggatggaagtgggctct.............................................................................................................................................................................................................................................................
367 acctcaaccacctggtatcg gggatccactccacgaagta.............................................................................................................................................................................................................................................................
233 aggaaccagatgatgccaag tggaggcttgctgtcttttt.............................................................................................................................................................................................................................................................
phosphate dehydrogenase; RT-PCR, reverse transcriptase polymerase chain reaction.
NOVEMBER 2010 American Journal of Obstetrics & Gynecology 495.e19

P ning (green line) vs the specific antibody staining profile (purple area). A representative example of moretM
Research Basic Science: Obstetrics www.AJOG.org
4
SUPPLEMENTARY FIGURE 1Phenotypic and immunogenic char
lots show isotype immunoglobulin G control stai
acterization of placenta-derived cells
han 3 experiments is shown.acias. Decidua multipotent mesenchymal stem cells. Am J Obstet Gynecol 2010.
95.e20 American Journal of Obstetrics & Gynecology NOVEMBER 2010
ACemgcpiFoM
www.AJOG.org Basic Science: Obstetrics Research
SUPPLEMENTARY FIGURE 2Analysis of the fetal or maternal or
, FISH analysis. Cells obtained from placentahX-FITC and human ChY-TRITC probes. XX signxamined (n � 3). B, Electrophoretogram showosomes X, Y of placentas of male newborns. Ps
uanine phosphoribosyl transferase markers arehromosome complement. In addition, only theresent, and SRY alleles were not detected. Mark
ndicated the presence of 2 alleles with peak areragment size in base pairs is shown on the horizon vertical axis.acias. Decidua multipotent mesenchymal stem cells. Am J O
igin of placental-derived cells
s of male newborns were analyzed using humanals but no XY signals were detected in all placentas
ing the QF-PCR detection of STR markers for chro-eudoautosomal X22 and the X-linked hypoxanthine-normal heterozygous, indicative of a normal XX sexX chromosome-specific product of the AMXY is
ers for chromosomes 21 (B), 13, and 18 (not shown)as in a 1:1 ratio indicating a normal diploid sample.ntal axis, and arbitrary fluorescence units are shown
bstet Gynecol 2010.
NOVEMBER 2010 American Journal of Obstetrics & Gynecology 495.e21

DCair1M
Research Basic Science: Obstetrics www.AJOG.org
4
SUPPLEMENTARY FIGURE 3Differentiation potential of clonal D
ifferentiation potential of 3 DMSC clone lines (n6 was derived from another different placenta. And staining of lipid vacuoles by Oil Red O (S
mmunostaining with proSP-C antibody to detectefractile cell morphology with neurite-like extens00 �m.acias. Decidua multipotent mesenchymal stem cells. Am J O
MSCs into tissues from the 3 germ layers
amed G2, C3, and C6). G2 and C3 were obtained from the same placenta as clone shown in Figure 6;, D, and G, Adipogenic differentiation was evaluated by visualization of lipid vacuoles inside the cells (G)
igma, St Louis, MO) (A and D). B, E, and H, Pulmonary epithelial differentiation was evaluated bytype II alveolar cells. C, F, and I, Neuronal differentiation was observed by detection of rounded, highly
ions and positive staining to �-III tubulin. Control cells were clearly negative for all the staining. Scale bar,
bstet Gynecol 2010.
95.e22 American Journal of Obstetrics & Gynecology NOVEMBER 2010

RsM
www.AJOG.org Basic Science: Obstetrics Research
NOVEMBER 2010 American
SUPPLEMENTARY FIGURE 4Molecular karyotyping of DMSCs
epresentative G-banding analysis of DMSCs (n � 3). Example illustrates DMSCs at passage 17howing a normal karyotype of 46 XX chromosomes.acias. Decidua multipotent mesenchymal stem cells. Am J Obstet Gynecol 2010.
Journal of Obstetrics & Gynecology 495.e23
Copyright © 2022 FDOKUMEN




















