
Isolation and characterization of mercury resistant Bacillus sp. from soils with an extensive...
15
This article was downloaded by: [b-on: Biblioteca do conhecimento online UTL] On: 03 March 2014, At: 03:22 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Journal of Toxicology and Environmental Health, Part A: Current Issues Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/uteh20 Isolation and Characterization of Mercury-Resistant Bacteria From Sediments of Tagus Estuary (Portugal): Implications for Environmental and Human Health Risk Assessment Neusa L. L. Figueiredo a , João Canário b , Aida Duarte a , Maria Luísa Serralheiro c & Cristina Carvalho a a Instituto de Investigação do Medicamento (iMed.ULisboa), Faculdade de Farmácia , Universidade de Lisboa Av. Prof. Gama Pinto , Lisboa , 1649-003 , Portugal b Centro de Química Estrutural , Instituto Superior Técnico, University of Lisbon , Lisboa , Portugal c Centro de Química e Bioquímica, Faculdade de Ciências da Universidade de Lisboa , Campo Grande , Lisboa , 1749-016 , Portugal Published online: 20 Feb 2014. To cite this article: Neusa L. L. Figueiredo , João Canário , Aida Duarte , Maria Luísa Serralheiro & Cristina Carvalho (2014) Isolation and Characterization of Mercury-Resistant Bacteria From Sediments of Tagus Estuary (Portugal): Implications for Environmental and Human Health Risk Assessment, Journal of Toxicology and Environmental Health, Part A: Current Issues, 77:1-3, 155-168, DOI: 10.1080/15287394.2014.867204 To link to this article: http://dx.doi.org/10.1080/15287394.2014.867204 PLEASE SCROLL DOWN FOR ARTICLE Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensors make no representations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content. Any opinions and views expressed in this publication are the opinions and views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and should be independently verified with primary sources of information. Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use of the Content. This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http:// www.tandfonline.com/page/terms-and-conditions
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Isolation and characterization of mercury resistant Bacillus sp. from soils with an extensive...

This article was downloaded by: [b-on: Biblioteca do conhecimento online UTL]On: 03 March 2014, At: 03:22Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Journal of Toxicology and Environmental Health, PartA: Current IssuesPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/uteh20
Isolation and Characterization of Mercury-ResistantBacteria From Sediments of Tagus Estuary (Portugal):Implications for Environmental and Human Health RiskAssessmentNeusa L. L. Figueiredo a , João Canário b , Aida Duarte a , Maria Luísa Serralheiro c & CristinaCarvalho aa Instituto de Investigação do Medicamento (iMed.ULisboa), Faculdade de Farmácia ,Universidade de Lisboa Av. Prof. Gama Pinto , Lisboa , 1649-003 , Portugalb Centro de Química Estrutural , Instituto Superior Técnico, University of Lisbon , Lisboa ,Portugalc Centro de Química e Bioquímica, Faculdade de Ciências da Universidade de Lisboa , CampoGrande , Lisboa , 1749-016 , PortugalPublished online: 20 Feb 2014.
To cite this article: Neusa L. L. Figueiredo , João Canário , Aida Duarte , Maria Luísa Serralheiro & Cristina Carvalho (2014)Isolation and Characterization of Mercury-Resistant Bacteria From Sediments of Tagus Estuary (Portugal): Implications forEnvironmental and Human Health Risk Assessment, Journal of Toxicology and Environmental Health, Part A: Current Issues,77:1-3, 155-168, DOI: 10.1080/15287394.2014.867204
To link to this article: http://dx.doi.org/10.1080/15287394.2014.867204
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of theContent. Any opinions and views expressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon andshould be independently verified with primary sources of information. Taylor and Francis shall not be liable forany losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use ofthe Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Journal of Toxicology and Environmental Health, Part A, 77:155–168, 2014Copyright © Taylor & Francis Group, LLCISSN: 1528-7394 print / 1087-2620 onlineDOI: 10.1080/15287394.2014.867204
ISOLATION AND CHARACTERIZATION OF MERCURY-RESISTANT BACTERIA FROMSEDIMENTS OF TAGUS ESTUARY (PORTUGAL): IMPLICATIONS FORENVIRONMENTAL AND HUMAN HEALTH RISK ASSESSMENT
Neusa L. L. Figueiredo1, João Canário2, Aida Duarte1, Maria Luísa Serralheiro3,Cristina Carvalho1
1Instituto de Investigação do Medicamento (iMed.ULisboa), Faculdade de Farmácia,Universidade de Lisboa Av. Prof. Gama Pinto, 1649-003 Lisboa, Portugal2Centro de Química Estrutural, Instituto Superior Técnico, University of Lisbon, Lisboa, Portugal3Centro de Química e Bioquímica, Faculdade de Ciências da Universidade de Lisboa,Campo Grande 1749-016, Lisboa, Portugal
Mercury (Hg) contamination of aquatic systems has been recognized as a global and seri-ous problem affecting both human and environmental health. In the aquatic ecosystems,mercurial compounds are microbiologically transformed with methylation responsible forgeneration of methylmercury (MeHg) and subsequent biomagnification in food chain, conse-quently increasing the risk of poisoning for humans and wildlife. High levels of Hg, especiallyMeHg, are known to exist in Tagus Estuary as a result of past industrial activities. Theaim of this study was to isolate and characterize Hg-resistant bacteria from Tagus Estuary.Mercury-resistant (Hg-R) bacteria were isolated from sediments of two hotspots (Barreiroand North Channel) and one reserve area (Alcochete). Mercury contamination in these areaswas examined and bacterial susceptibility to Hg compounds evaluated by determination ofminimal inhibitory concentrations (MIC). The isolates characterization was based on mor-phological observation and biochemical testing. Bacteria characteristics, distribution, and Hgresistance levels were compared with metal levels. Barreiro and North Channel were highlycontaminated with Hg, containing 126 and 18 µg/g total Hg, respectively, and in Alcochete,contamination was lower at 0.87 µg/g total Hg. Among the isolates there were aerobic andanaerobic bacteria, namely, sulfate-reducing bacteria, and Hg resistance levels ranged from0.16 to 140 µg/ml for Hg2+ and from 0.02 to 50.1 µg/ml for MeHg. The distribution of thesebacteria and the resistance levels were consistent with Hg contamination along the depth ofthe sediments. Overall, results show the importance of the characterization of Tagus Estuarybacteria for ecological and human health risk assessment.
Mercury (Hg) is among the metals withhighest toxicological importance and iswidespread in the environment (Domingo,1994; Nichols et al., 1999). Naturally occurringHg results from degassing of the earth’s crustand evaporation from oceans (Boening, 2000).Mercury exists in different chemical species,with the major forms in water being mercuricmercury (Hg2+) and methylmercury (MeHg)(Morel et al., 1998). However, the increaseof aquatic systems with Hg contamination,resulting from anthropogenic activities, was
Address correspondence to Cristina Carvalho, Instituto de Investigação do Medicamento (iMed.ULisboa), Faculdade de Farmácia,Universidade de Lisboa Av. Prof. Gama Pinto, 1649-003 Lisboa, Portugal. E-mail: [email protected]
recognized as a global and serious problem,affecting both wildlife and human health(Campbell et al., 2003; Marvin-DiPasqualeand Agee, 2003; Sweet and Zelikoff, 2001).Mercury pollution and its effects throughbioaccumulation in food web is a seriousproblem because of the high toxicity associatedwith its species forms (Mathema et al., 2011).Ecologically, Hg pollution affects both highpredators and microbial community. Adverseeffects were observed in mammals (Ni et al.,2012; Sweet and Zelikoff, 2001), amphib-
155
Dow
nloa
ded
by [
b-on
: Bib
liote
ca d
o co
nhec
imen
to o
nlin
e U
TL
] at
03:
22 0
3 M
arch
201
4

156 N. L. L. FIGUEIREDO ET AL.
ians (Davidson et al., 2011) fish (Ung et al.,2010), and birds (Burger and Gochfeld, 1997;Herring et al., 2012; Heinz and Hoffman,2003), and include behavioral disturbances,immunotoxicity, neurological system deformityand abnormal functions, and developmentaland reproductive abnormalities such as fetalmalformations or inability of reproduce (Burgerand Gochfeld, 1997; Davidson et al., 2011;Heinz and Hoffman, 2003; Herring et al.,2012; Sweet and Zelikoff, 2011). The mostimportant case of human poisoning by Hginvolved the population of Minamata Bay(Japan), who suffered severe neurologicaldisorders termed Minamata disease as a conse-quence of contamination on Minamata Bay byindustrial activity (Ekino et al., 2007).
Mercury toxicity derives from the forma-tion of highly toxic organomercurial complexes,predominantly MeHg, which is highly neuro-toxic (Mathema et al., 2011; Ni et al., 2012).In humans, Hg affects the central nervous sys-tem, motor system, and renal, cardiovascular,immune, and reproductive systems (Ratcliffeet al., 1996; Zahir et al., 2005), by disrupt-ing cell function due to the affinity to thiolgroups of proteins (Ung et al., 2010). Amongmercurial compounds, MeHg is one of themost toxic forms as it passes through biologicmembranes and, once inside the cell, producesirreversible damage to nucleic acids, alteringnormal configuration and biological cell activ-ities (Mathema et al., 2011; NRC, 2000).
Although Hg compounds are toxic to all liv-ing organisms, constant exposure to mercurialcompounds enables the bacterial commu-nity to develop several types of resistancemechanisms, which allow them to deal withthe adverse effects of metal-mediated toxi-city (Mathema et al., 2011). In water andsediments, these mechanisms are responsiblefor bacterial Hg transformation, namely, reduc-tion of Hg2+, Hg methylation, and MeHgdemethylation. These conversion processesmay occur as the result of diverse biologicalpathways (Barkay and Wagner-Dobler, 2005).The best known pathway is the reductivedemethylation followed reduction of Hg2+—a
process mediated by enzymatic activity of pro-teins codified by a cluster of genes organizedin the mer operon (Nascimento and Chartone-Souza, 2003).
The methylation process is considered ofgreat concern, as it yields the highly toxic com-pound MeHg. Among the bacterial community,Hg methylation was found to predominantlybe associated with sulfate-reducing bacteria(SRB) (King et al., 2000, 2001, 2002). SRB areanaerobic bacteria that use sulfate as a termi-nal electron acceptor for degradation of organiccompounds, resulting in production of sulfide(Muyzer and Stams, 2008). Bacterial transfor-mation of Hg is influenced by several factors,such as bacterial community composition andavailability of reactive Hg species (Macaladyet al., 2000; Rasmussen and Sorensen, 1998).
Estuaries are important for their ecologi-cal potential that offers adequate conditionsfor development of both animal and plantspecies, promoting biodiversity. However, thesesystems are often impacted by anthropogenicactivities, namely, industrial discharge (Ferreira,1988). Tagus Estuary, one of the most impor-tant estuaries in Europe, covering an area of325 km2, was reported to be contaminatedsince 1985 due to two industrial areas locatedin the north and south margins, North Channeland Barreiro, which led to high levels of Hgin sediments, suspended matter, and water(Figuères et al., 1985). Despite the inactiva-tion of the most critical industrial units inthese areas, contamination still persists. Severalstudies noted high levels of Hg in fish (Limaet al., 1982) and microalgae (Ferreira, 1988)and in sediments (Canário et al., 2003, 2004)of several areas of Tagus Estuary. Inventoriesin sediments estimated 21 tons of total Hgand 23 kg of MeHg in the estuary (Canárioet al., 2005). Despite the knowledge of con-tamination levels, there is still a lack of informa-tion on the biogeochemical processes involvedin the Hg cycle in Tagus Estuary, namely,the reduction/methylation/demethylation pro-cesses. Thus, the aim of this study was to isolateand characterize Hg-resistant (Hg-R) bacteriafound in sediments of two industrial sites and
Dow
nloa
ded
by [
b-on
: Bib
liote
ca d
o co
nhec
imen
to o
nlin
e U
TL
] at
03:
22 0
3 M
arch
201
4

MERCURY-RESISTANT BACTERIA FROM ESTUARY SEDIMENTS 157
one natural reserve area in Tagus Estuary toassess the role of bacteria in Hg cycling.
MATERIAL AND METHODS
Study Area and Sample CollectionAccording to previous studies (Canário
et al., 2003, 2005), three sites of TagusEstuary were sampled (Figure 1): twohighly Hg-contaminated areas, Barreiro(lat.: 38◦40′45.40′′ N; long.: 9◦3′1.70′′ W)and North Channel (lat.: 38◦51′21.21′′ N;long.: 9◦3′40.51′′ W), and Alcochete (lat.:38◦45′41.58′′ N; long.: 8◦56′49.93′′ W),which was used as a control area due to lowcontamination (Figure 1). Barreiro station hadan intensive industrial activity since the 1960s,decade, which has now stopped, while NorthChannel industrial activity started around1940–1950 and continues to the present(LNEC, 2008). In contrast, Alcochete stationbelongs to a natural reserve of high ecologicalvalue having an important role in conservationof plant and animal species.
Sediments samples were collected duringthe summer. Sediments cores, approximately50 cm long, were collected twice (one for Hganalysis and other for bacterial isolation) and
rapidly sliced in layers of 3 cm along depth(Figure 2). Samples were stored in sealed tubes,in a refrigerated environment, and transportedto the lab for microbial isolation.
Analysis of Total Hg (HgT) and MeHg onSedimentsMercury contamination levels of total Hg
(HgT) in the solid portion and pore water,along the depth of sediment core, were deter-mined. HgT of solid fraction and pore waterwas determined following sediment centrifuga-tion by atomic absorption spectrometry usinga silicon ultraviolet (UV) diode detector LECOAMA-254 after pyrolysis of each sample ina combustion tube at 750◦C under an oxy-gen atmosphere and collection on a goldamalgamator (Costley et al., 2000). MeHgwas determined in dry sediments by alka-line digestion (KOH/MeOH), organic extrac-tion with dichloromethane (DCM) and precon-centration in aqueous sulfide solution, back-extraction into DCM, and quantification bygas chromatography with atomic fluorescencespectroscopy (GC-AFS) using an Agilent chro-matograph coupled with a pyrolizer unit anda PSA fluorescence detector (Canário et al.,2004). Recoveries and possible MeHg artifact
FIGURE 1. Map of Tagus Estuary with the location of the sampled areas (Barreiro, North Channel, and Alcochete).
Dow
nloa
ded
by [
b-on
: Bib
liote
ca d
o co
nhec
imen
to o
nlin
e U
TL
] at
03:
22 0
3 M
arch
201
4

158 N. L. L. FIGUEIREDO ET AL.
MH + MeHg (0.02µg/mL)
Overlyingwater
MH + MeHg (0.22µg/mL)
0.22µg/mLMeHg
Anaerobic Jar
100µL 100µL 100µL
0.22µg/mLMeHg
0.02µg/mL MeHg
1mL
500 rpm1min
AerobicBacteriaIsolation
SRBIsolation
AnaerobicBacteriaIsolation
Bacteria Isolates
0.5mL
MH(4.5 mL)
Postgate C(4.5 mL)
Postgate C + MeHg(0.02µg/mL)
Postgate C + MeHg(0.22µg/mL)
0.5mL
SedimentCollection
BacteriaIsolates
BacteriaIsolates
Postgate E
FIGURE 2. Schematic representation of samples collection and different type of Hg-R bacteria isolation from sediments of sampled areasof Tagus Estuary. Aerobic Hg-R bacteria and anaerobic Hg-R bacteria, including Hg-R SRB, were isolated.
formation were assessed by spiking several sam-ples with Hg2+ and MeHg standard solutionswith different concentrations. Recoveries variedbetween 97 and 103%, and no artifact MeHgformation was observed during the procedure.Precision, expressed as relative standard devi-ation of 4 replicate samples, was less than4% (p < .05). Certified reference materials forMeHg (IAEA-405; 54.9 ± 5.3 ng/g) and fortotal Hg (BCR-580; 132 ± 3 µg/g) were usedto ensure the accuracy of the procedure, andthe obtained values 55.9 ± 5 ng/g for IAEA-405 and 131 ± 2 µg/g for BCR-580 were notstatistically different.
Isolation and MorphologicalCharacterization of Bacteria StrainsInoculums were prepared through the dilu-
tion of sediments samples with 20 ml distilledsterile water. Figure 2 shows the scheme ofdifferent techniques used for the isolation ofdifferent Hg-resistant (Hg-R) bacteria: aerobicHg-R bacteria and anaerobic Hg-R bacteria,including Hg-R SRB.
Aerobic Hg-R Bacteria Isolation Thesamples were shaken and after centrifugationat 2650 × g for 1 min (4◦C); 100 µl of super-natant was plated on Mueller-Hinton (MH) agar
Dow
nloa
ded
by [
b-on
: Bib
liote
ca d
o co
nhec
imen
to o
nlin
e U
TL
] at
03:
22 0
3 M
arch
201
4

MERCURY-RESISTANT BACTERIA FROM ESTUARY SEDIMENTS 159
media without and with MeHg selective pres-sure in a concentration of 0.022 µg/ml and0.22 µg/ml (Figure 2). MeHg was used toestablish a selective pressure since the mainobjective of the study was focused on bacteriaresponsible for methylation/demethylation ofHg. Colony numbers were counted in all mediafor colony-forming units (CFU) quantification.Different colonies were selected on MH agarplus 0.22 µg/ml MeHg and after were storedin MH broth plus 15% of glycerol and 0.022µg/ml MeHg at –80◦C.
Anaerobic Hg-R Bacteria IsolationWashed sediment, 0.5 ml, was inoculated inserum bottles (Belco Glass, Inc.) containing4.5 ml MH and closed with rubber stopperswith a crimped metal seal. Media were pre-pared under nonsterile conditions and addedto serum bottles. Serum bottles were gassedwith N2 during and after media addition andthen sealed as they were withdrawn fromgassing needles. Metal seals were then crimpedand the bottled media were autoclaved(Figure 2). To avoid O2 contamination, allinoculations were performed with a hypoder-mic syringe and needle washed with N2 inanaerobic chamber (with N2 flux). After 3 d ofgrowing at 22◦C, 0.5-ml inoculums were trans-ferred to a new bottled medium supplementedwith 0.022 µg/ml MeHg and then to bot-tled medium supplemented with 0.22µg/mlMeHg, in order to select Hg-R bacteria. After3 d of growth, aliquots were inoculated on solidMH (0.22 µg/ml MeHg) in an anaerobic cham-ber and incubated in anaerobic jars (Oxoid)(anaerobic conditions were obtained usingAnaeroGen sachet [Oxoid]). Single colonieswere selected and stored in MH broth plus15% of glycerol added by 0.022 µg/ml MeHgat –80◦C.
Hg-R SRB Isolation The isolation of SRBwas performed using the same methodol-ogy already described, with the exception ofcolony isolation techniques, which were differ-ent (Figure 2) and involved the use of a selectivemedium, Postgate. This medium contains sul-fate that is reduced by SRB to sulfide, forminga black precipitate that indicates SRB growing.Postgate C (liquid medium) supplemented with
MeHg (0.022 and 0.22 µg/mL) was used toselect Hg-R SRB. Single colonies were isolatedaccording to the roll tube method describedby Miller and Wolin (1974), with some modifi-cations. Fifty-milliliter serum bottles containing6 ml melted agar medium (Postgate E with0.22 µg/ml MeHg) were inoculated with 0.1 mlPostgate C inoculum with added 0.22 µg/mlMeHg and adjusted to 10−5 CFU/ml. For softmix, bottles were manually rolled prior to inoc-ulation and placed in cold water to solidify.After 5–7 d of growth, single colonies wereselected and submitted again to isolation pro-cess just described. After 3 repetitions, sin-gle colonies were incubated into Postgate Cmedium containing 0.22 µg/ml MeHg andstored at 4◦C.
Morphological and BiochemicalCharacterization Cells at the early growthstages were examined with respect to mor-phology and gram staining characteristics.Biochemical characterization was carried outfollowing standard methodologies, such aslactose, glucose, and mannitol fermentation,enzymatic activity detection (catalase, oxidase,amylase, casease, phosphatase, and lipase),salt tolerance, or ability to growth on media(MH) containing 8% (w/v) NaCl. Commercialmicromethods for the biochemical identifi-cation were also applied and included BBLCrystal Identification Systems (BD) for aerobicand anaerobic bacteria identification andAPI20E/20NE (bioMérieux sa) comprising testsfor fermentation, oxidation, degradation, andhydrolysis of various substrates. Some organiccompounds (electron donors), such as formate,lactate, fumarate, and acetate, and electronacceptors (sulfate and nitrate) were used tocharacterize SRB.
Mercury Susceptibility TestingAerobic and anaerobic isolates suscepti-
bilities to Hg compounds were determinedby minimal inhibitory concentration (MIC)determination, using a modified micro dilu-tion broth method described by CLSI (2006).Microorganism cultures in brain heart infusionbroth (BHI) at a concentration of 108 CFU/ml
Dow
nloa
ded
by [
b-on
: Bib
liote
ca d
o co
nhec
imen
to o
nlin
e U
TL
] at
03:
22 0
3 M
arch
201
4

160 N. L. L. FIGUEIREDO ET AL.
(OD 0.5 at 595 nm) were diluted in MH inorder to obtain 3 × 106 CFU/ml. To achieveconcentrations ranging from 1.33 × 10−3 to1358 µg/ml HgCl2 (4.88 × 10−6 to 5.00µM) and from 1.23 × 10−5 to 125.5 µg/mlMeHgCl (4.88 × 10−8 to 0.50 µM), 100 µl ofaqueous solution of these compounds (2.72 to2715 µg/ml HgCl2 and 0.025 to 251.1 µg/mlMeHgCl) was diluted with 100 µL of bacterialsuspension into the first well of a sterile 96-wellmicroplate and then sequentially diluted 1:2 inthe following 10 wells. Bacterial suspension inthe absence of mercurial compounds was usedas a control in the 12th well. Duplicate sam-ples were performed for each concentrationtested. After incubation at 37◦C for 24 h underaerobic and anaerobic conditions, bacterialgrowth or its absence was observed. The MICwas defined as the minimum concentration oftest compound that inhibited visible growth.All data points represent the mean ± standarddeviation (STD) of two to three independentdeterminations.
SRB susceptibility to mercurial compoundswas determined following the same criteriaalready described, adjusting it into a macrodilution broth method using Postgate C bottledand gassed with N2. An adequate volume ofsterile Postgate C medium was added to eachsealed and sterile serum bottle using a sterileand N2-washed syringe. After 24 h, bacterialgrowth was determined by spectrophotomet-ric reading at 595 nm. It was only possible todetermine MIC50 (minimal inhibitory concen-tration that inhibited 50% of bacterial growth)once high concentrations of HgCl2 led to for-mation of colored precipitate, probably due tocinnabar (HgS) formation.
RESULTS
Isolation and Characterization of Hg-RBacteria in SedimentsIn total, 93 different Hg-R bacteria from
sediments collected in Tagus Estuary were iso-lated on media (MH and Postgate) in thepresence of 0.22 µg/ml MeHg, a concentra-tion closely associated to resistance, according
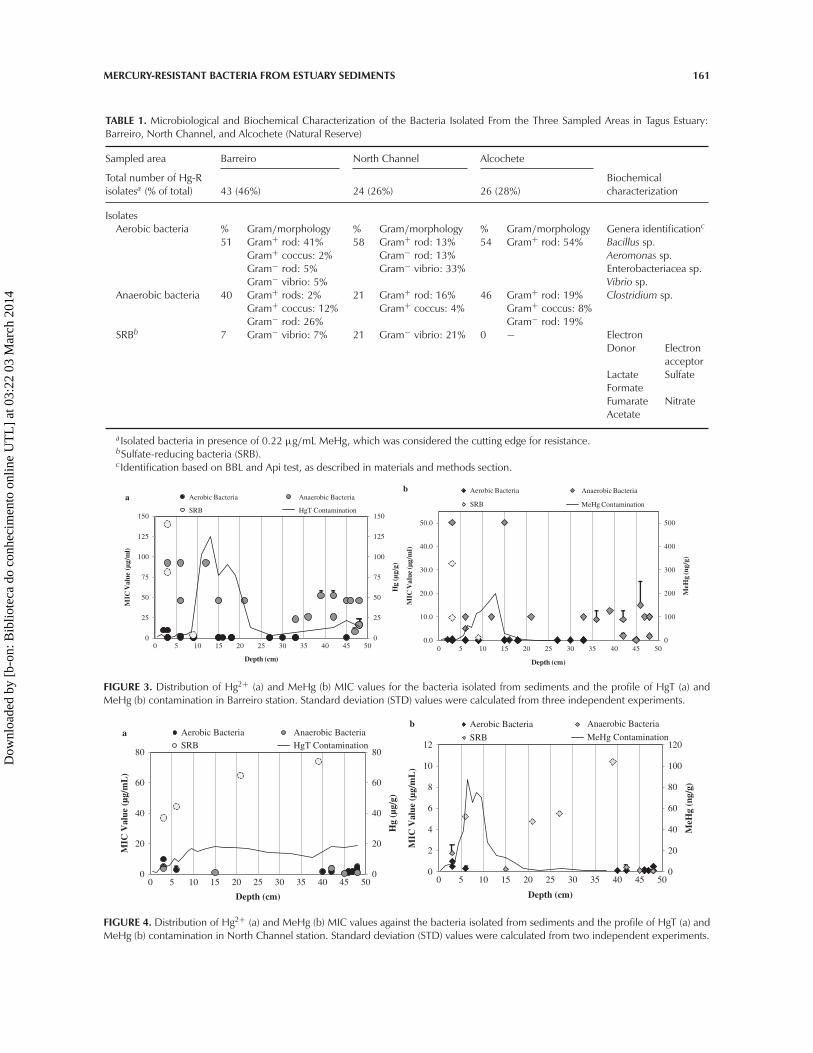
to the results of Sadhukhan and co-workers(1997), which indicated 2.5µg/ml as the fron-tier for Hg2+. Also François et al., (2011) cor-roborated this edge by using 2.72 µg/ml ofHg2+ as a typical concentration for the isolationof resistant strains. For MeHg the concentra-tion was decreased by one order of magnitudeaccording to the normal tolerance ratio thatisolates present for Hg2+/MeHg. Among theHg-R isolates, 43 were isolated from Barreiro,24 from North Channel, and 26 from Alcochetesediments (Table 1).
The Hg-R isolates were mostly gram-positive rods (44%). According to the BBL test,the aerobic gram-positive rods belong mainly toBacillus sp. and anaerobic gram-positive rodsmainly to Clostridium sp. Bacilli were mainlyisolated from Barreiro sediments (41%, 17/43).Gram-negative bacteria were predominatelyisolated from the two highly contaminatedareas (Barreiro and North Channel) (Table 1).Among the gram-negative isolates, genera suchas Vibrio, Aeromonas, and Enterobacteriaceaespecies were identified through BBL and API20E/20NE systems (data not shown). For SRBcharacterization, biochemical evaluation indi-cated that all isolates were able to use lactateas an electron donor and sulfate as an electronacceptor. Further, some bacteria had the capa-bility of using formate, fumarate, and acetate aselectron donor and nitrate as electron acceptor(Table 1).
Hg-R Bacteria Distribution and MercuryContamination ProfileFigures 3–5 exhibit Hg contamination and
Hg-R bacteria isolates distribution profile alongthe depth in different areas of collection. Thedistribution of Hg contamination in Barreirosediments (Figure 3) showed that HgT wasspread along the depth at mainly between8 and 22 cm, with a peak (126 µg/g) at13 cm (Figure 3a) whereas MeHg concen-trated in the first 20 cm with the peak(201 ng/g) also at 13 cm (Figure 3b). In thecase of North Channel, results demonstrateda contamination with MeHg also in the first20 cm (peak at 7 cm [87 ng/g]), whereas
Dow
nloa
ded
by [
b-on
: Bib
liote
ca d
o co
nhec
imen
to o
nlin
e U
TL
] at
03:
22 0
3 M
arch
201
4
MERCURY-RESISTANT BACTERIA FROM ESTUARY SEDIMENTS 161
TABLE 1. Microbiological and Biochemical Characterization of the Bacteria Isolated From the Three Sampled Areas in Tagus Estuary:Barreiro, North Channel, and Alcochete (Natural Reserve)
Sampled area Barreiro North Channel Alcochete
Total number of Hg-Risolatesa (% of total) 43 (46%) 24 (26%) 26 (28%)
Biochemicalcharacterization
IsolatesAerobic bacteria % Gram/morphology % Gram/morphology % Gram/morphology Genera identificationc
51 Gram+ rod: 41% 58 Gram+ rod: 13% 54 Gram+ rod: 54% Bacillus sp.Gram+ coccus: 2% Gram− rod: 13% Aeromonas sp.Gram− rod: 5% Gram− vibrio: 33% Enterobacteriacea sp.Gram− vibrio: 5% Vibrio sp.
Anaerobic bacteria 40 Gram+ rods: 2% 21 Gram+ rod: 16% 46 Gram+ rod: 19% Clostridium sp.Gram+ coccus: 12% Gram+ coccus: 4% Gram+ coccus: 8%Gram− rod: 26% Gram− rod: 19%
SRBb 7 Gram− vibrio: 7% 21 Gram− vibrio: 21% 0 − ElectronDonor Electron
acceptorLactateFormate
Sulfate
FumarateAcetate
Nitrate
aIsolated bacteria in presence of 0.22 µg/mL MeHg, which was considered the cutting edge for resistance.bSulfate-reducing bacteria (SRB).cIdentification based on BBL and Api test, as described in materials and methods section.
0
25
50
75
100
125
150
0
25
50
75
100
125
150
0 5 10 15 20 25 30 35 40 45 50
Hg
(µg/
g)
MIC
Val
ue (
µg/m
l)
Depth (cm)
a Aerobic Bacteria Anaerobic Bacteria
SRB HgT Contamination
0
100
200
300
400
500
0.0
10.0
20.0
30.0
40.0
50.0
0 5 10 15 20 25 30 35 40 45 50
MeH
g (n
g/g)
MIC
Val
ue (µ
g/m
l)
Depth (cm)
b Aerobic Bacteria Anaerobic Bacteria
SRB MeHg Contamination
FIGURE 3. Distribution of Hg2+ (a) and MeHg (b) MIC values for the bacteria isolated from sediments and the profile of HgT (a) andMeHg (b) contamination in Barreiro station. Standard deviation (STD) values were calculated from three independent experiments.
0
20
40
60
80
0
20
40
60
80
0 5 10 15 20 25 30 35 40 45 50
Hg
(µg/
g)
MIC
Val
ue (
µg/m
L)
Depth (cm)
a Aerobic Bacteria Anaerobic BacteriaSRB HgT Contamination
0
20
40
60
80
100
120
0
2
4
6
8
10
12
0 5 10 15 20 25 30 35 40 45 50
MeH
g (n
g/g)
MIC
Val
ue (
µg/m
L)
Depth (cm)
b Aerobic Bacteria Anaerobic Bacteria
SRB MeHg Contamination
FIGURE 4. Distribution of Hg2+ (a) and MeHg (b) MIC values against the bacteria isolated from sediments and the profile of HgT (a) andMeHg (b) contamination in North Channel station. Standard deviation (STD) values were calculated from two independent experiments.
Dow
nloa
ded
by [
b-on
: Bib
liote
ca d
o co
nhec
imen
to o
nlin
e U
TL
] at
03:
22 0
3 M
arch
201
4

162 N. L. L. FIGUEIREDO ET AL.
0.0
1.0
2.0
3.0
4.0
5.0
0.0
10.0
20.0
30.0
40.0
50.0
0 5 10 15 20 25 30 35 40 45 50
Hg
(µg/
g)
MIC
Val
ue (
µg/m
l)
Depth (cm)
aAerobic Bacteria Anaerobic Bacteria HgT Contamination
0
5
10
15
20
0.0
1.0
2.0
3.0
4.0
5.0
6.0
0 5 10 15 20 25 30 35 40 45 50
MeH
g (n
g/g)
MIC
Val
ue (
µg/m
l)
Depth (cm)
bAerobic Bacteria Anaerobic Bacteria MeHg Contamination
FIGURE 5. Distribution of Hg2+ (a) and MeHg (b) MIC values against the bacteria isolated from sediments and the profile of HgT (a) andMeHg (b) contamination in Alcochete station. Standard deviation (STD) values were calculated from two independent experiments.
HgT was spread along the depth maintain-ing a concentration of 15–18 µg/g between9 and 48 cm (Figures 4a and 4b). Alcochete,the less contaminated station, displayed levelsof Hg contamination two orders of magni-tude lower (HgT peak [0.87 µg/g] at 15 cm)than values registered in Barreiro and NorthChannel, with MeHg contamination locatedmore at the superficial layer of sediments(MeHg peak [4.1 ng/g] at 5 cm) (Figures 5aand 5b).
The distribution profile of the Hg-R iso-lated is in agreement with the contaminationprofile, as seen in Figures 3–5. In Barreiro, themajority of Hg-R bacteria were isolated in thefirst 15 cm, indicating that resistant bacteriawere concentrated on sediments surface closeto contaminated layers (Figures 3a and 3b).In North Channel, resistant bacteria distributedmore evenly with a numerical increase indepth (40–48 cm) (Figure 4). In Alcochete,Hg-R were located along the depth (Figure 5)without a clear separation between aerobicand anaerobic bacteria and Hg-R SRB wereabsent. These differences in the distributionbetween the three sampled areas showed thatbacterial communities are influenced differ-ently by levels of contamination. As observed inFigures 3–5, Hg-R bacteria are predominantlyin sediments layers adjacent to high Hg contentlayers, and normally there are only anaerobicbacteria or SRB—highly Hg-R bacteria—adjacent to Hg, especially MeHg peakconcentrations.
Mercury Resistance LevelsThe resistance levels of the isolates to Hg
were evaluated by determining MIC values ofinorganic mercury (HgCl2) and methylmercury(MeHgCl). Data are summarized in Table 2.The isolates exhibited MIC values ranging from0.16 to 140 µg/ml for Hg2+ and 0.02 to 50.1µg/ml for MeHg (Table 2). In general, the iso-lates were 10-fold more resistant to Hg thanto MeHg, and the magnitude of resistance wasaerobic bacteria < anaerobic bacteria < SRB(Table 2). The most resistant bacteria, includ-ing aerobic, anaerobic, and SRB, were isolatedfrom sediments of Barreiro and North Channel,and the highest MIC values for Hg2+ and MeHgwere observed on Barreiro isolates (140 and32.8 µg/ml, respectively) (Table 2).
DISCUSSION
The sampled areas of Tagus Estuary showedhigh Hg contamination, in particular MeHg.Sediments of Barreiro and North Channel werehighly contaminated, presenting HgT aver-age 33.2 µg/g (0.31–126 µg/g ) and 11.7µg/g (0.99–18 µg/g ), respectively (Figures 3and 4). These levels of contamination arein agreement with the tendency reported in2003 and 2005 by Canário et al (2003, 2005)and 20 years ago by Figuères et al. (1985).Barreiro contamination is comparable with thatreported in highly contaminated ecosystemssuch as Fort Churchill (Carson River, Sierra
Dow
nloa
ded
by [
b-on
: Bib
liote
ca d
o co
nhec
imen
to o
nlin
e U
TL
] at
03:
22 0
3 M
arch
201
4

MERCURY-RESISTANT BACTERIA FROM ESTUARY SEDIMENTS 163
TABLE 2. MIC Values Range Exhibited by the Isolates for Mercurial Compounds Depending on the Sampled Area of Tagus Estuary
MIC values range (µg/ml) MIC50a values range (µg/ml)
Aerobic bacteria Anaerobic bacteria SRB
Sampled area Hg2+ MeHg Hg2+ MeHg Hg2+ MeHg
Barreiro 0.16–9.87 0.02–0.54 8.11–92.7 0.10–50.1 3.69–140 1.12–32.8North Channel 1.13–10.0 0.11–1.12 0.65–4.06 0.07–1.75 36.9–81.3 5.19–15.2Alcochete 0.41 – 1.13 0.04–0.19 0.87–46.4 0.05–5.01 —
aMIC50: Minimal inhibitory concentration that inhibits 50% of bacterial growth.
Nevada), which presented Hg levels of 29.95µg/g (Oremland et al., 1995).
Data indicated that Hg contamination wasexerting metal tolerance/resistance within thebacterial community as evidenced by the ratiobetween total number of bacteria capable oftolerating 0.02 µg/ml MeHg and total numberof bacteria (7/9 in Barreiro and 4/5 in NorthChannel and Alcochete) (Table 3). Further, thenumber of bacteria resistant to 0.22 µg/ml rep-resented 77% of total CFU in Barreiro, whilein North Channel and Alcochete less than 5%of the total CFU were resistant to the sameconcentration of MeHg (Table 3). In addi-tion, a significant number of Hg-R bacteriawas found in sediments of Barreiro (46% oftotal number of bacteria isolated) when com-pared to North Channel and Alcochete (26%and 28%, respectively) (Table 1). Thus, col-onization of sediments by resistant bacteriawas influenced by the chronic high level ofHg contamination in Barreiro. The high lev-els of mercurial compounds also affected Hg-R bacteria prevalence in the neighborhoodof Hg contamination peaks (Figures 3–5).
These results are in accordance with sev-eral studies that reported that the num-ber of Hg-R bacteria in soil and aquaticenvironment depended upon Hg content ofthe environment (Barkay, 1987; Batten andScow, 2003; Ruggiero et al., 2011; Sadhukhanet al., 1997; Summers, 1986; Timoney et al.,1978).
The isolates included aerobic andanaerobic bacteria, and among anaerobicbacteria there were SRB (Table 1). Biochemicalidentification indicated that most of the iso-lates belonged to Bacillus, Aeromonas, Vibrio,Enterobacteriaceae, and Clostridium species,which are typical freshwater bacteria (Ivanovaet al., 2001; Matyar et al., 2000). Bacillusspp. were the most common Hg-R bacteriafound, which is not surprising, as Bacillususually predominate in sediments, includingcontaminated ecosystems such as sediments ofMinamata Bay (Nakamura and Silver, 1994),which led to these bacteria being consideredimportant indicators for Hg resistance moni-toring (Summers, 1986). Among Barreiro andNorth Channel anaerobic isolates detected,
TABLE 3. Mercury Contamination in the Three Sampled Areas of Tagus Estuary and Analysis of Total Number of Hg-Tolerant Bacteria andHg-R Bacteria
Total number of Hg-tolerant and Hg-resistantbacteria (CFU/cma) Mercury contamination range
Sampled areasTotal number ofbacteria (CFU/cma) 0.02 µg/ml MeHgb 0.22 µg/ml MeHgb HgT (µg/g)c MeHg (ng/g)c
Barreiro 9.45 × 103 7.28 × 103 7.28 × 103 0.31–126 0.76–201North Channel 5.08 × 104 4.19 × 104 9.80 × 102 0.99–18 1.0–87Alcochete 1.69 × 104 1.40 × 104 7.70 × 102 0.24–0.87 0.0–4.10
Note. CFU, colony-forming units.acm of sediment.bIncorporated in MH agar.cMercury content was quantified until ± 40 cm of depth.
Dow
nloa
ded
by [
b-on
: Bib
liote
ca d
o co
nhec
imen
to o
nlin
e U
TL
] at
03:
22 0
3 M
arch
201
4

164 N. L. L. FIGUEIREDO ET AL.
SRB are normally associated with Hg methyla-tion. Some SRB are more likely to methylateHg, especially SRB, which also use acetate asan electron donor (King et al., 2000). TheseSRB were detected, suggesting their probableinvolvement in Hg methylation processes.Other bacteria described as an Hg methylatorare Clostridium sp. (Yamada and Tonomura,1972), frequently found among the anaerobicisolates identified.
In general, in Barreiro and North Channelthere was more diversity among the Hg-R,while in Alcochete the isolates were mostlygram-positive rods (Table 1). This might berelated to the fact that chronic exposure tohigh Hg concentrations selectively affects dif-ferent bacterial communities and increases Hg-R bacteria diversity (Ruggiero et al., 2011;Vishnivetskaya et al., 2011). However, theinfluence of other factors, such as tempera-ture, pH, organic matter content, and eutro-phization (Macalady et al., 2000), as well asother contaminants including antibiotics (Lima-Bittencourt et al., 2007) and chemical com-pounds, such as heavy metals (Nascimento andChartone-Souza, 2003), needs to be consid-ered. For instance, high levels of As, Pb, Zn,Cr, Cu (Vale, 1990; Caçador et al., 1996;Vale et al., 2008), and cadmium (Cd) (Vale,1990) were noted in sediments of TagusEstuary, which may also affect bacterial diver-sity. In addition, Hg contamination influencedHg-R bacterial distribution. Mercury contami-nation in Barreiro was high, between 10 and20 cm (Figures 3a and 3b), with higher lev-els of Hg-R bacteria in this segment than inadjacent layers (0–8 and 25–45 cm). On theother hand, in North Channel contaminationstarted close to the surface, with MeHg con-tamination increasing sharply at 3 cm andpeaking at 7 cm, and therefore exerting aselective pressure for Hg-R bacteria that arescarce on the first sediment slices and morefrequent with depth (40–45 cm) (Figures 4aand 4b). In Alcochete sediments, Hg-R bac-teria were isolated along the depth; however,the most resistant bacteria were isolated on thesurface, where Hg contamination was higher(Figures 5a and 5b). Thus, Hg contamination
exerted a selective pressure on Hg-R bacteriain these areas, affecting their distribution alongthe depth, which is in agreement with a studyfrom Ruggieno and coworkers (2011) that con-cluded that Hg contamination altered microbialcommunity.
This is also supported by the isolates sus-ceptibility to Hg compounds. The most resistantHg-R bacteria were isolated from Barreiro andNorth Channel; resistance levels reached 9.87µg/ml and 10 µg/ml among aerobic bacte-ria and 140 µg/ml and 81.3 µg/ml amonganaerobic bacteria (SRB) (Table 2). The resis-tance levels correlated positively with contami-nation levels, with high contamination in NorthChannel and Barreiro exerting strong selec-tive pressure for generation and persistenceof Hg-R bacteria, while low contamination inAlcochete station did not favor selection. Thecomparison of bacteria isolated in Tagus Estuaryand bacteria isolated in other polluted placesindicate that in our study resistance was ofthe same magnitude or higher. For example,Hg2+ MIC for Bacillus spp. was found by oth-ers to range from 0.01 to 36.9 µg/ml (Oslonet al., 1979, Timoney et al., 1978, Pan Hou,2010), and bacteria from Tagus showed val-ues between 0.18 and 5.01 µg/ml. Austin andcoworkers (1977) studied a mixture of bacte-ria isolated from estuary sediments and waterand MIC of Hg2+ was 0.01 µg/ml, whereasour data demonstrated Hg2+ MIC averageson the order of 1.8 µg/ml for aerobic bac-teria and 38.69 µg/ml for anaerobic bacteriaincluding SRB.
Thus, the results presented in this studysupport the contention that chronic and highHg contamination in some areas of TagusEstuary (Barreiro and North Channel) exertsa selective effect on Hg-R bacteria, whichmay be involved in Hg cycling. The dif-ferent chemical forms of Hg are not con-fined, and various processes affect the envi-ronmental cycling of Hg. Concentrations of Hgin water may be extremely low but micro-bial activity in aquatic ecosystems convertsa small proportion of deposited Hg in inor-ganic form to MeHg, which bioaccumulatesto relatively high concentrations in fish and
Dow
nloa
ded
by [
b-on
: Bib
liote
ca d
o co
nhec
imen
to o
nlin
e U
TL
] at
03:
22 0
3 M
arch
201
4

MERCURY-RESISTANT BACTERIA FROM ESTUARY SEDIMENTS 165
especially in top predatory fish. Conversionof inorganic Hg into MeHg is responsible forneurotoxic, cardiotoxic, and teratogenic effects.The MeHg concentrations result from a bal-ance between methylation and demethyla-tion processes, which is not fully understood,but sediments are potential sources of MeHgfor food webs. From a human health per-spective, exposure to MeHg rather than totalHg is more important, since MeHg is readilyavailable for absorption into the bloodstreamand crosses the blood–brain barrier, producingneurotoxicity.
Bacteria as a community have a rathercomplex role in Hg conversion, as bacte-ria such as SRB and Clostridium sp. wereshown to methylate Hg (King et al., 2000;Eckstrom et al., 2003; Yamada and Tonomura,1972) whereas Bacillus sp. has the abilityto reduce the metal and volatilize it in theform of Hg0 (Sadhukahan et al., 1997). Moststudies focused on identifying bacteria withmethylation capacities, although for remedi-ation purposes and human health protec-tion Hg-R bacteria perform reduction andvolatilization, which is crucial. Therefore, iso-lates need to be individually tested to evaluateenzymatic conversion capacity and detoxifyingprocesses.
CONCLUSIONS
This study is the first that identifies andcharacterizes Hg-R bacteria from Tagus Estuaryand investigates their involvement in Hgcycling in this polluted ecosystem. Datashowed that Hg-R bacteria exist and are fre-quent in Tagus Estuary sediments. Further, thisstudy provided evidence that contaminationby Hg compounds influenced significantlynot only diversity of Hg-R bacteria but alsotheir distribution, and resistance levels tomercurial compounds. Overall, data indicateinvolvement of these bacteria in Hg conversion(reduction/methylation/demethylation) andimportance of this for aquatic ecosystems forthe biosphere in general and human health riskin particular.
FUNDING
This work was funded by FCT throughthe project PROFLUX (PTDC/MAR/102748/
2008) and by iMed.UL through FCT’s strate-gic project: PEst-OE/SAU/UI4013/2011. N. L.Figueiredo is grateful to FCT for the BI,research grant of PROFLUX. The authors thankthe members of the Laboratory of MolecularBiology in the Faculty of Pharmacy, Universityof Lisbon, for their important collaboration.
REFERENCES
Austin, B., Allen, D. A, Mills, A. L., and Colwell,R. R. 1977. Numerical taxonomy of heavymetal-tolerant bacteria isolated from estuary.Can. J. Microbiol. 23: 1433–1447.
Barkay, T. 1987. Adaptation of aquatic micro-bial communities to mercury2+ stress. Appl.Environ. Microbiol. 53: 2725–2732.
Barkay, T., and Wagner-Dobler, I. 2005.Challenges and achievements in controllingmercury toxicity in the environment. Adv.Appl. Microbiol. 57: 1–40.
Batten, K. M., and Scow, K. M. 2003. Sedimentmicrobial community and methylmercurypollution at four mercury mine-impactedsites. Microbiol. Ecol. 46: 429–441.
Boening, D. 2000. Ecological effects, trans-port, and fate of mercury: A general review.Chemosphere 40: 1335–1351.
Burger, J., and Gochfeld, M. 1997. Risk, mer-cury levels, and birds: Relating adverse labo-ratory effects to field biomonitoring. Environ.Res. 75: 160–172.
Caçador, I., Vale, C., and Catarino, F., 1996.Accumulation of Zn, Pb, Cu, Cr and Ni insediments between roots of the Tagus Estuarysalt marshes, Portugal. Est. Coast. Shelf Sci.42: 393–403.
Campbell, L. M., Dixon, R. G., and Hecky,R. E. 2003. A review of mercury in LakeVictoria, East Africa: Implications for humanand ecosystem health. J. Toxicol. Environ.Health B 6: 325–356.
Canário, J., Vale, C., and Caetano, M. 2005.Distribution of monomethylmercury andmercury in surface sediments of the Tagus
Dow
nloa
ded
by [
b-on
: Bib
liote
ca d
o co
nhec
imen
to o
nlin
e U
TL
] at
03:
22 0
3 M
arch
201
4

166 N. L. L. FIGUEIREDO ET AL.
Estuary (Portugal). Mar. Pollut. Bull. 50:1142–1145.
Canário, J., Antunes, P., Lavrado, J., and Vale, C.2004. Simple method for monomethylmer-cury determination in estuarine sediments.Trends Anal. Chem. 23: 798–805.
Canário, J., Vale, C., and Caetano, M. 2003.Mercury in contaminated sediments andpore waters at a contaminated enriched insulphate (Tagus Estuary). Environ. Pollut. 126:425–433.
Chiang, Y.-C., Yang, C.-Y., Li, C., Ho, Y.-C.,Lin, C.-K. and Tsen, H.-Y. 2006. Identificationof Bacillus spp., Escherichia coli, Salmonellaspp., Staphylococcus spp. and Vibrio spp.With 16S ribosomal DNA-based oligonu-cleotide array hybridization. Int. J. FoodMicrobiol. 107: 131–137.
Clinical and Laboratory Standards Institute.2006. Methods for dilution antimicrobial sus-ceptibility tests for bacteria that grow aer-obically; Approved standard—Seventh edi-tion. Clinical and Laboratory StandardsInstitute document M7-A7 [ISBN 1-56238-587-9]. Wayne, PA: Clinical and LaboratoryStandards Institute.
Costley, C. T., Mossop, K. F., Dean, J. R.,Garden, L. M., Marshall, J., and Carroll, J.2000. Determination of mercury in environ-mental and biological samples using pyrol-ysis atomic absorption spectrometry withgold amalgamation. Anal. Chim. Acta 405:179–183.
Davidson, M. A., Croteau, M. C., Millar, C.S., Trudeau, V. L., and Lean, D. R. 2011.Fate and developmental effects of dietaryuptake of methylmercury in Silurana tropi-calis tadpoles. J. Toxicol. Environ. Health A74: 364–379.
Domingo, J., 1994. Metal-induced develop-mental toxicity in mammals: A review. J.Toxicol. Environ. Health A 42:123–141.
Ekstrom, E. B., Morel, F. M., and Benoit, J. M.2003. Mercury methylation independent ofthe acetyl-coenzyme: a pathway in sulfate-reducing bacteria. Appl. Environ. Microbiol.69: 5414–5422.
Ekino, S., Susa, M., Ninomiya, T., Imamura,K., and Kitamura, T. 2007. Minamatadisease revisited: An update on the acute
and chronic manifestations of methylmercury poisoning. J. Neurol. Sci. 262:131–144.
Ferreira, J. G. 1988. Mercúrio em algas macró-fitas no Estuário do Tejo. PhD thesis, NewUniversity of Lisbon, Lisbon, Portugal8.
Figuères, G., Martin, J. M., Meybeck, M.,and Seyler, P. 1985. A comparative studyof mercury contamination in the TagusEstuary (Portugal) and major French Estuaries(Gironde, Loire, Rhône). Est. Coast. Shelf Sci.20:183–203.
François, F., Lombard, C., Guigner, J. M.,Soreau, P., Brian-Jaisson, F., Martino, G.,Vandervennet, M., Garcia, D., Molinier,A.L., Pignol, D., Peduzzi, J., Zirah, S., andRebuffat, S. 2011. Isolation and character-ization of environmental bacteria capableof extracellur biosorption of mercury. Appl.Environ. Microbiol. 78:1097–1106.
Heinz, G., and Hoffman, D. 2003. Embryotoxicthresholds of mercury: Estimates from indi-vidual mallard eggs. Arch. Environ. Contam.Toxicol. 44: 257–264.
Herring, G., Ackerman, J., and Herzog, M.2012. Mercury exposure may suppress base-line corticosterone levels in juvenile birds.Environ. Sci. Technol. 46: 6339–6346.
Ivanova, E. P., Zhukova, N. V., Gorshkova,N. M., and Chaikina, E. L. 2001.Characterization of Aeromonas and Vibriospecies isolated from a drinking waterreservoir. J. Appl. Microbiol. 90:919–927.
King, J. K., Harmon, S. M., Fu, T. T., andGladden, J. B. 2002. Mercury removal,methylmercury formation, and sufate-reducing bacteria profiles in wetlandmesocosms. Chemosphere 46: 859–870.
King, J. K., Kostka, J. E., Fricher, M. E., Saunders,F. M., and Jahnke, R. A. 2001. A quantita-tive relationship that demonstrates mercurymethylation rates in marine sediments arebased on the community composition andactivity of sulfate-reducing bacteria. Environ.Sci. Technol. 35: 2491–2496.
King, J. K., Kostka, J. E., Frischer, M. E., andSaunders, F. M. 2000. Sulfate-reducing bac-teria methylate mercury at variable rates inpure culture and in marine sediments. Appl.Environ. Microbiol. 66: 2430–2437.
Dow
nloa
ded
by [
b-on
: Bib
liote
ca d
o co
nhec
imen
to o
nlin
e U
TL
] at
03:
22 0
3 M
arch
201
4

MERCURY-RESISTANT BACTERIA FROM ESTUARY SEDIMENTS 167
LNEC (Laboratório Nacional de EngenhariaCivil). 2008. Avaliação das alternativas delocalização da travessia do Tejo. Ambiente—Qualidade do sedimento do Estuário.Relatório 0/2008 NEC.
Liebert, C., Wireman, J., Smith, T., andSummers, A. 1997. Phylogeny of mer-cury resistance (mer) operons of Gram-negative bacteria isolated from the fecal floraof primates. Appl. Environ. Microbiol. 63:1066–1076.
Lima-Bittencourt, C. I., Cursino, L., Gonçalves-Dornelas, H., Pontes, D. S., Nardi, R. M.D., Callisto, M., Chartone-Souza, E., andNascimento, A. M. A. 2007. Multiple antimi-crobial resistance in Enterobacteriaceae iso-lates from pristine freshwater. Genet. Mol.Res. 6: 510–521.
Lima, C., Figuères, G., Martin, J.M., Meybeck,M., and Seyler, P. 1982. Mercury in the TagusEstuary: An acute or obsolete problem? CAN/
UNESCO/UNEP/IOC http://books.google.com/books?vid=ISBN9251023867, Lisbon.
Macalady, J. L., Mack, E. E., Nelson, D.,and Scow, K. M. 2000. Sediment micro-bial community structure and mercurymethylation in mercury-polluted Clear Lake,California. Appl. Environ. Microbiol. 66:1479–1488.
Mason, R. P., and Benoit, J. M. 2003.Organomercury compounds in the environ-ment. In Organometallic compounds in envi-ronment, ed. P. Graig, 57–99. West Sussex,UK: Wiley.
Mathema, V., Thakuri, C. and Sillanpaa, M.2011. Bacterial mer operon-mediated detox-ification of mercurial compounds: a shortreview. Arch. Microbiol. 193: 837–844.
Matyar, F., Kaya, A., and Dinçer, S. 2008.Antibacterial agents and heavy metal resis-tance in gram-negative bacteria isolatedfrom seawater, shrimp and sediment inIskenderun Bay, Turkey. Sci. Total Environ.407: 279–285.
Marvin-DiPasquale, M., and Agee, J.L. 2003.Microbial mercury cycling in sediments ofthe San Francisco Bay-Delta. Estuaries 26:1517–1528.
Miller, T., and Wolin, M. J. 1974. A serumbottle modification of the Hungate tech-nique for cultivating obligate anaerobes.Appl. Microbiol. 27: 985–987.
Monfort, P., and Baleux, B. 1990. Dynamicsof Aeromonas hydrophila, Aeromonas sobriaand Aeromonas caviae in a sewage treat-ment pond. Appl. Environ. Microbiol. 56:1999–2006.
Morel, F. M. M, Kraepiel, A. M. L., and Amyot,M. 1998. The chemical cycle and bioaccu-mulation of mercury. Annu. Rev. Ecol. Syst.29: 543–566.
Muccillo-Baisch, A., Mielean, N., Carrazzoni,D., Soares, M., Goulart, G., and Baisch, P.2012. Health effects of ingestion of mercury-polluted urban soil: An animal experiment.Environ. Geochem. Health 34: 43–53.
Muyzer, G., and Stams, A. F. 2008. Theecology and biotechnology of sulphate-reducing bacteria. Nature Rev. Microbiol. 6:441–454.
Nakamura, K., and Silver, S. 1994. Molecularanalysis of mercury-resistant Bacillus isolatesfrom sediment of Minamata Bay, Japan. Appl.Environ. Microbiol. 60:4596–4599
Nakamura K., Fujisaki T. and Tamashiro H.1986. Characteristics of Hg-resistant bacte-ria isolated from Minamata Bay sediment.Environ. Res. 40:58–67.
Narita, M., Chiba, K., Nishizawa, H., Ishii, H.,Huang, C.-C., Kawabata, Z., Silver, S., andEndo, G. 2003. Diversity of mercury resis-tance determinants among Bacillus strainsisolated from sediment of Minamata Bay.FEMS Microbiol. Lett. 223: 73–82.
Nascimento, A. M. A., and Chartone-Souza,E. 2003. Operon mer: Bacterial resistanceto mercury and potential for bioremediationof contaminated environments. Genet. Mol.Res. 2: 92–101.
National Research Council. 2000. Toxicologicaleffects of methylmercury. Washington, DC:National Academy Press.
Ni, M., Li, X., Rocha, J. B. T., Farina, M., andAschner, M. 2012. Glia and methylmercuryneurotoxicity. J. Toxicol. Environ. Health A 75:1091–1101.
Dow
nloa
ded
by [
b-on
: Bib
liote
ca d
o co
nhec
imen
to o
nlin
e U
TL
] at
03:
22 0
3 M
arch
201
4

168 N. L. L. FIGUEIREDO ET AL.
Nichols, J., Bradbury, S., and Swartout, J. 1999.Derivation of wildlife values for mercury. J.Toxicol. Environ. Health B 2: 325–355.
Olson, B., Barkay, T., Nies, D., Bellama, J. M.,and Colwell, R. R. 1999. Plasmid media-tion of mercury volatilization and methyla-tion by estuarine bacteria. J. Ind. Microbiol.Biotechnol. 22: 418–427.
Oremland, R., Miller, L., Dowdle, P., Connell,T., and Barkay, T. 1995. Methylmercuryoxidative degradation potentials in contam-inated and pristine sediments of the CarsonRiver, Nevada. Appl. Environ. Microbiol. 61:2745–2753.
Pan-Hou, H. 2010. Application of mercury-resistant genes in bioremediation of mer-curials in environments. Yakugaku Zasshi130:1143–1156.
Rasmussen, L.D., and Sorensen, S. J. 1998.The effect of long-term exposure to mercuryon bacterial community in marine sediment.Curr. Microbiol. 36: 291–297.
Ratcliffe, H. E., Swanson, G. M., and Fischer,L. J. 1996. Human exposure to mercury: Acritical assessment of the evidence of adversehealth effects. J. Toxicol. Environ. Health 49:221–270.
Ruggiero, P., Terzano, R., Spagnuolo, M.,Cavalca, L., Colombo, M., Andreoni, V., Rao,M. A., Perucci, P., and Monaci, E., 2011.Mercury bioavailability and impact on bacte-rial communities in a long-term polluted soil.J. Environ. Monit. 13: 145–156.
Sadhukahan, P. C., Ghosh, S., Chaudhuri, J.,Ghosh, D. K., and Mandal, A. 1997. Mercuryand organomercurial resistance in bacte-ria isolated from freshwater fish of wetlandfisheries around Calcutta. Environ. Pollut.97:71–78.
Schaefer, J. K., Yagi, J., Reinfelder, J. R.,Cardona, T., Ellickson, K. M., Tel-Or, S.,and Barkay, T. 2004. Role of the bacterialorganomercury lyase (MerB) in controllingmethylmercury accumulation in mercury-contaminated natural waters. Environ. Sci.Technol. 38: 4304–4311.
Sweet, L. I., and Zelikoff, J. T. 2001. Toxicologyand immunotoxicology of mercury: A com-parative review in fish and humans. J. Toxicol.Environ. Health B 4: 161–205.
Summers, A. O. 1986. Organization, expres-sion and evolution of genes for mercury resis-tance. Annu. Rev. Microbiol. 40:607–634.
Timoney, J. F., Port, J., Giles, J., and Spanier,J. 1978. Heavy-metal and antibiotic resis-tance in the bacterial flora of sediments ofNew York Bight. Appl. Environ. Microbiol. 36:465–472.
Ung, C., Lam, S., Hlaing, M., Winata, C.,Korzh, S., Mathavan, S., and Gong, Z.2010. Mercury-induced hepatotoxicity inzebrafish: In vivo mechanistic insights fromtranscriptome analysis, phenotype anchor-ing and targeted gene expression validation.BMC Genomics 11: 1471–2164.
Vale, C., Canário, J., Caetano, M., Lavrado,J., and Brito, P. 2008. Estimation of theanthropogenic fraction of elements in surfacesediments of the Tagus Estuary (Portugal).Mar. Pollut. Bull. 56: 1353–1376.
Vale, C. 1990. Temporal variations of particu-late metals in the Tagus River Estuary. Sci.Total Environ. 97: 137–154.
Vishnivetskaya, T., Mosher, J., Paumbo, A.,Yang, Z., Podar, M., Brown, S., Brooks,S., Gu, B., Southworth, G., Drake, M.,Brandt, C., and Elias, D. 2011. Mercuryand other heavy metals influence bacte-rial community structure in contaminatedTennessee streams. Appl. Environ. Microbiol.77: 302–311.
Wireman, J., Liebert, C., Smith, T., andSummers, A. 1997. Association of mercuryresistance with antibiotic resistance in thegram-negative fecal bacteria of primates.Appl. Environ. Microbiol. 63: 4494–4503.
Yamada, M., and Tonomura, K. 1972.Formation of methylmercury compoundsfrom inorganic mercury by Clostridiumcochlearium. J.Ferment. Technol. 50:159–166.
Zahir, F., Rizwi, S., Haq, S., and Khan, R. 2005.Low dose mercury toxicity and human health.Environ. Toxicol. Pharmacol. 20: 351–360.
Zhang, W., Chen, L., and Liu, D., 2012.Characterization of a marine-isolatedmercury-resistant Pseudomonas putidastrain SP1 and its potential application inmarine mercury reduction. Appl. Microbiol.Biotechnol. 93:1305–1314.
Dow
nloa
ded
by [
b-on
: Bib
liote
ca d
o co
nhec
imen
to o
nlin
e U
TL
] at
03:
22 0
3 M
arch
201
4




















