
Isolation and characterization of chitinase genes from pitchers ...
10
RESEARCH PAPER Isolation and characterization of chitinase genes from pitchers of the carnivorous plant Nepenthes khasiana Haviva Eilenberg, Smadar Pnini-Cohen, Silvia Schuster, Anna Movtchan and Aviah Zilberstein* Department of Plant Sciences, The George S. Wise Faculty of Life Sciences, Tel Aviv University, Ramat Aviv 69978, Israel Received 7 December 2005; Accepted 5 April 2006 Abstract The genus Nepenthes represents carnivorous plants with pitcher traps capable of efficient prey capture and digestion. The possible involvement of plant chitinases in this process was studied in Nepenthes khasiana. Two different types of endochitinases were identified in the liquid of closed traps exhibiting substrate specificity for either long chitin polymers or N-acetylglucosamine (GlcNAc) oligomers. Injection of chitin into such closed sterile pitchers induced the appearance of additional endochitinase isoenzymes, with substrate specificity only for long chitin polymers. No significant exochiti- nase (N-acetyl-b-glucosaminidase) or chitobiosidase activity could be detected in the non-induced or in- duced trap liquid. Four genes representing two sub- groups of basic chitinases, denoted as Nkchit1b and Nkchit2b, were isolated from the secretory region of N. khasiana pitchers. The main differences between the two subgroups are the presence of a proline-rich hinge region only in NkCHIT1b and a C-terminal putative vacuole targeting extension only in NkCHIT2b, indicat- ing different compartmentalization of the two enzymes. Reverse transcription–polymerase chain reaction (RT– PCR) evaluation of mRNA levels showed that the Nkchit2b genes are constitutively expressed in the secretory cells while transcription of Nkchit1b genes is induced by chitin injection. These results show for the first time the involvement of genes encoding chitinases in prey–trap interaction and their differential expression and activity during prey trapping. Key words: Carnivorous plants, chitinase, chitin induction, Nepenthes khasiana, pitcher liquid. Introduction Carnivorous plants grown in barren regions with high humidity and acidic soil have developed special organs to capture insects, digest them, and absorb the nutrients. The tropical carnivorous pitcher plant Nepenthes, whose name originated from the poisonous drug used by Helen in Homer’s Odyssey, encompasses ;80 species, with the largest diversity in the islands of Borneo and Sumatra. Additional scattered populations are found in northern India (N. khasiana), Madagascar (N. distillatoria), and the Seychelles (N. perveleii). Pitcher-shaped leaves, that capture insects, characterize all the species. The prey is attracted to the pitcher by its colour and nectar secretion. The upper part of the pitcher has a wax-coated inner epidermis that forms a slippery surface promoting the sliding down of the prey into the lower part of the pitcher, which has >6000 digestive cellular glands per cm 2 . These glands function in the perception of chemical stimuli, secretion of digestive enzymes, and nutrient absorption (Owen et al., 1999; Schulze et al., 1999). The presence of proteolytic enzymes in carnivorous plants had already been reported by Darwin (1875). The acidic trap liquid of Nepenthes was shown to have several hydrolytic enzymes such as proteases and phosphatases (Heslop-Harrison, 1975). Five cDNA clones of aspartic proteinase (AP) were isolated from Nepenthes alata and their expression was localized to different parts of the pitcher (An et al., 2002). A plasma membrane H + -ATPase and transporters of ammonium, amino acids, and peptides, involved in acidification of the pitcher fluid and nutrient absorption, respectively, were also identified in N. alata secretory glands (Schulze et al., 1999; An et al., 2001). The presence of proteases, RNases, nucleases, and phosphatases during trap development was studied in Sarracenia * To whom correspondence should be addressed. E-mail: [email protected] Abbreviations: CaMV, cauliflower mosaic virus; ChiA, Serratia marcescens chitinase; GlcNAc, N-acetylglucosamine; 4MU, 4-methylumbelliferyl; Nkchit1b, Nepenthes khasiana basic chitinase 1; pNP, p-nitrophenyl; RT–PCR, reverse transcription–polymerase chain reaction. Journal of Experimental Botany, Vol. 57, No. 11, pp. 2775–2784, 2006 doi:10.1093/jxb/erl048 Advance Access publication 7 July, 2006 ª The Author [2006]. Published by Oxford University Press [on behalf of the Society for Experimental Biology]. All rights reserved. For Permissions, please e-mail: [email protected] Downloaded from https://academic.oup.com/jxb/article/57/11/2775/682094 by guest on 11 January 2022
-
Upload
khangminh22 -
Category
Documents
-
view
4 -
download
0
Transcript of Isolation and characterization of chitinase genes from pitchers ...

RESEARCH PAPER
Isolation and characterization of chitinase genes frompitchers of the carnivorous plant Nepenthes khasiana
Haviva Eilenberg, Smadar Pnini-Cohen, Silvia Schuster, Anna Movtchan and Aviah Zilberstein*
Department of Plant Sciences, The George S. Wise Faculty of Life Sciences, Tel Aviv University,Ramat Aviv 69978, Israel
Received 7 December 2005; Accepted 5 April 2006
Abstract
The genus Nepenthes represents carnivorous plants
with pitcher traps capable of efficient prey capture and
digestion. The possible involvement of plant chitinases
in this process was studied inNepenthes khasiana. Two
different types of endochitinases were identified in the
liquid of closed traps exhibiting substrate specificity
for either long chitin polymers or N-acetylglucosamine
(GlcNAc) oligomers. Injection of chitin into such closed
sterile pitchers induced the appearance of additional
endochitinase isoenzymes, with substrate specificity
only for long chitin polymers. No significant exochiti-
nase (N-acetyl-b-glucosaminidase) or chitobiosidase
activity could be detected in the non-induced or in-
duced trap liquid. Four genes representing two sub-
groups of basic chitinases, denoted as Nkchit1b and
Nkchit2b, were isolated from the secretory region of N.
khasiana pitchers. The main differences between the
two subgroups are the presence of a proline-rich hinge
region only in NkCHIT1b and a C-terminal putative
vacuole targeting extension only in NkCHIT2b, indicat-
ing different compartmentalization of the two enzymes.
Reverse transcription–polymerase chain reaction (RT–
PCR) evaluation of mRNA levels showed that the
Nkchit2b genes are constitutively expressed in the
secretory cells while transcription of Nkchit1b genes
is induced by chitin injection. These results show for
the first time the involvement of genes encoding
chitinases in prey–trap interaction and their differential
expression and activity during prey trapping.
Key words: Carnivorous plants, chitinase, chitin induction,
Nepenthes khasiana, pitcher liquid.
Introduction
Carnivorous plants grown in barren regions with highhumidity and acidic soil have developed special organs tocapture insects, digest them, and absorb the nutrients. Thetropical carnivorous pitcher plant Nepenthes, whose nameoriginated from the poisonous drug used by Helen inHomer’s Odyssey, encompasses ;80 species, with thelargest diversity in the islands of Borneo and Sumatra.Additional scattered populations are found in northernIndia (N. khasiana), Madagascar (N. distillatoria), andthe Seychelles (N. perveleii). Pitcher-shaped leaves, thatcapture insects, characterize all the species. The prey isattracted to the pitcher by its colour and nectar secretion.The upper part of the pitcher has a wax-coated innerepidermis that forms a slippery surface promoting thesliding down of the prey into the lower part of the pitcher,which has >6000 digestive cellular glands per cm2. Theseglands function in the perception of chemical stimuli,secretion of digestive enzymes, and nutrient absorption(Owen et al., 1999; Schulze et al., 1999).
The presence of proteolytic enzymes in carnivorousplants had already been reported by Darwin (1875). Theacidic trap liquid of Nepenthes was shown to have severalhydrolytic enzymes such as proteases and phosphatases(Heslop-Harrison, 1975). Five cDNA clones of asparticproteinase (AP) were isolated from Nepenthes alata andtheir expression was localized to different parts of thepitcher (An et al., 2002). A plasma membrane H+-ATPaseand transporters of ammonium, amino acids, and peptides,involved in acidification of the pitcher fluid and nutrientabsorption, respectively, were also identified in N. alatasecretory glands (Schulze et al., 1999; An et al., 2001). Thepresence of proteases, RNases, nucleases, and phosphatasesduring trap development was studied in Sarracenia
* To whom correspondence should be addressed. E-mail: [email protected]: CaMV, cauliflower mosaic virus; ChiA, Serratia marcescens chitinase; GlcNAc, N-acetylglucosamine; 4MU, 4-methylumbelliferyl; Nkchit1b,Nepenthes khasiana basic chitinase 1; pNP, p-nitrophenyl; RT–PCR, reverse transcription–polymerase chain reaction.
Journal of Experimental Botany, Vol. 57, No. 11, pp. 2775–2784, 2006
doi:10.1093/jxb/erl048 Advance Access publication 7 July, 2006
ª The Author [2006]. Published by Oxford University Press [on behalf of the Society for Experimental Biology]. All rights reserved.For Permissions, please e-mail: [email protected]
Dow
nloaded from https://academ
ic.oup.com/jxb/article/57/11/2775/682094 by guest on 11 January 2022

purpurea, representing another family of pitcher-possessingplants (Gallie and Chang, 1997). The expression of thesehydrolytic enzymes was shown to be developmentallyregulated during the pitcher opening stages. Later on, inmature opened pitchers, these enzymes are induced by an,as yet undefined, signal transduction mechanism in re-sponse to nucleic acids, proteins, and reduced nitrogen(Gallie and Chang, 1997). Digestion of colloidal chitin bythe secreted liquid of the Nepenthes pitcher and by anaqueous extract of Drosera peltata indicated the possibleinvolvement of chitinolytic enzymes in the digestion ofinsects by carnivorous plants (Amagase et al., 1972).Recently, chitinase activity was detected in the carnivorousgenus Utricularia (Sirova et al., 2003) and in Droserarotundifolia L. (Matusikova et al., 2005). However, as yet,no full-length sequences of chitinase genes or cDNAs havebeen isolated from carnivorous plants.
Plant chitinases are to be considered pathogen-related(PR) proteins, since their activity is induced by fungal,bacterial, and viral infections, as well as by more generalsignals of stress such as wounding, salicylic acid, ethylene,and heavy metal salts (Collinge et al., 1993; Graham andSticklen, 1994). Moreover, chitinases also have importantnon-defensive functions in growth and development pro-cesses (Kasprzewska, 2003). Chitinases are involved in theregulation of nodulation signal molecules (Goormachtiget al., 1998), they affect embryogenesis (De Jong et al.,1992; Helleboid et al., 2000), and they might participate inprogrammed cell death (Passarinho et al., 2001). Chitin andchitosan are major components of cell walls in insects andfungi. Since insects make up the bulk of the prey found inpitcher traps, chitinases appear to play a double role incarnivorous plants—defence and carnivory. The isolationand characterization of four N. khasiana chitinase genesand their corresponding cDNA sequences is described here.To gain a better understanding of the role of chitinases inpitchers, exogenous chitin was applied to the pitchers asa means to mimic prey capture, and the expression patternof chitinases was characterized.
Materials and methods
Plant growth conditions
Nepenthes khasiana plants were grown in the Botanical Gardens ofTel Aviv University in a greenhouse under a 16 h/8 h light/darkregime at 25 8C. The plants were watered twice a week with double-distilled water. Closed pitchers were used for all analyses describedherein.
Trap liquid sampling and induction of chitinase activity by
chitin injection
Trap liquid was collected from closed young pitchers by a sterilesyringe, in order to preserve sterility, and stored at –70 8C in aliquots.Approximately 1 mg of colloidal chitin (Vessey and Pegg, 1973), pH5.0, was injected with a sterile syringe into the side of a closed pitcherthat was first surface-sterilized with 70% ethanol. Aliquots of the trap
liquid were collected from the closed pitcher at increasing intervalsafter injection.
Chitinase assays using various substrates
Endochitinase activity was assayed by first separating sterile trapliquid or callus extracts on native 12–15% acrylamide gels using25 mM TRIS–HCl, pH 8.3, 192 mM glycine as running buffer (Davis,1964) and then by overlaying an additional acrylamide gel containingglycol-chitin (0.01% w/v) as a substrate (Trudel and Asselin, 1989).After 20 h incubation at 37 8C, the gels were stained for 5 min with0.01% (w/v) Calcofluor white M2R. Chitinase activity was visualizedas dark spots on the glycol-chitin gel by UV light (260 nm).
Chitinolytic activity was also characterized in gels by using 4-methylumbelliferyl (4MU) b-D-N,N9,N$-triacetylchitotrioside asa substrate. After electrophoresis, the gel was washed twice with100 mM sodium citrate buffer, pH 5.5, and then overlaid with200 lM substrate solubilized in 1% plant agar. The released 4MUwas detected by UV light.
The activity of N-acetyl-b-glucosaminidase (exochitinase), chito-biosidase, and endochitinase in pitcher liquid was determined inmicrotitre plates by using as substrates the dimer 4MU-N-acetyl-b-D-glucosaminide [4MU-(GlcNAc)1; Sigma], trimer 4MU-b-D-N,N9-diacetylchitobioside [4MU-(GlcNAc)2, Sigma], or tetramer4MU-b-D-N,N9,N$-triacetylchitotrioside [4MU-(GlcNAc)3, Sigma],respectively. Chitinase specificity was estimated by the cleavage ofthe b-1,4-bond that releases 4MU from the different oligomers(Kuranda and Robbins, 1991). The chitinase reaction contained 20 llof trap liquid mixed with 30 ll of 300 lM substrate in 0.1 M sodiumcitrate buffer, pH 3.0 and incubated at 37 8C for 1 h. The reaction wasstopped by addition of 150 ll of 0.64 M glycine-NaOH buffer,pH 10.4. The production of 4MU was measured by a fluorescencespectrophotometer (excitation at 360 nm, emission at 460 nm).Activity units were defined as nM 4MU released min�1.
Kinetic properties of endochitinases were determined according toTronsmo and Harman (1993) in microtitre plates using p-nitrophenyl(pNP)-b-(GlcNac)3 as the substrate. Reactions (80 ll) containingincreasing substrate concentrations ranging from 0 to 250 lM in50 mM potassium phosphate buffer, pH 6.7, and 30 ll of 5.6-foldconcentrated pitcher liquid or 0.0005 U of commercial Serratiamarcescens chitinase (Sigma), were incubated for 30 min at 50 8C.Reactions were terminated by adding 50 ll of 0.4 M sodiumcarbonate, which also enhanced the colour of p-nitrophenol formedby the enzymatic cleavage of the substrate. Absorbance at 405 nmwas measured by a Spectromax 190 microplate reader.
Isolation of genomic DNA
Leaf tissue (1 g) was homogenized in 5 ml of buffer consisting of 1vol. of DNA extraction buffer (0.35 M sorbitol, 0.1 M TRIS–HCl, pH7.5, 5 mM EDTA, and 0.02 M Na-bisulphite), 1 vol. of nuclei lysisbuffer (0.2 M TRIS–HCl, pH 7.5, 50 mM EDTA, 2 M NaCl, 2%CTAB), and 0.4 vol. of 5% sarkosyl. The homogenate was incubatedfor 20 min at 65 8C and then extracted twice with 1 vol. ofchloroform:isoamyl alcohol (24:1 v:v). Three volumes of 6 M NaIwere added to the aqueous phase and the genomic DNA was furtherpurified by using a ‘High Pure’ filter tube (High Pure PlasmidIsolation Kit, Boehringer Mannheim).
Isolation of chitinase genes
Conserved amino acid sequences of plant chitinases were used todesign degenerate primers for polymerase chain reaction (PCR)amplification of Nepenthes chitinase genes. The following degenerateprimers were used: 59-TTIGGICAA/GACIT/AG/CICAC/TGAG/AAC-39(direct primer), and 59-GAG/TICCICCA/GTTIATIATA/GTTT/A/C/GGT-39 (reverse primer). PCR was carried out using N.khasiana genomic DNA as a template at an annealing temperature of
2776 Eilenberg et al.
Dow
nloaded from https://academ
ic.oup.com/jxb/article/57/11/2775/682094 by guest on 11 January 2022

50 8C. Based on the isolated sequences, sets of marginal primers withreverse orientation were synthesized to isolate the rest of the genesequences further by an inverse PCR strategy. In brief, genomic DNA(5 lg) digested with HindIII, BamHI, or NdeI was self-ligated to formcircles. About 100 ng of this circular DNA was used as a template forPCR amplification at annealing temperatures between 60 8C and65 8C using TaKaRa Ex Taq and the specifically designed primers.
Isolation of total RNA
Total RNA was isolated from the lower part of the pitcher by a specialhot borate/proteinase K method (Schulze et al., 1999).
Isolation of mRNA and cDNA synthesis
To identify differential expression of the defined chitinase genes,mRNA was purified from total RNA by using oligo(dT) conjugatedto magnetic DynaBeads (Dynal, Norway) according to the manu-facturer’s instructions. Thereafter total cDNA was synthesized byM-MLV reverse transcriptase (Promega) using oligo(dT)12–18 asa primer. The primer was annealed at 22 8C for 10 min and thereverse transcription reaction was performed at 40 8C for 60 min. A1 ll aliquot of the reverse transcription reaction (50 ll) was taken toPCR amplification. The following gene-specific primers were usedfor the identification of the transcribed genes in the trap secretorycells: Nkchit1b direct primer 59-CGACGGTCCATATGCATGGG-GATACTGTTTCAAG-39, reverse primer 59-CAAATGGCCACTGGGTGTAGCAGTCCAAGTTATC-39; Nkchit2b direct primer 59-CGTGGGGATATTGCTATCTCAG-39, and reverse primer 59-CTACTCGGTGG CCCACAAAAG-39.
Constructs and plant transformation
Nkchit1b was synthesized by PCR amplification using genomic DNAtemplate, Taq polymerase (Ex.Taq.,TaKaRa), and specific primerscorresponding to the 59 and 39 ends of the coding sequence. Nkchit2bcDNA was synthesized by a reverse transcription (RT)–PCR strategy.Briefly, mRNA isolated from trap tissue was reverse transcribedby M-MLV reverse transcriptase (Promega). The resulting cDNAwas PCR amplified using specific primers designed according tothe 59 and 39 end of the coding sequence. Nkchit1b and Nkchit2bPCR products were cloned downstream of the constitutive cauli-flower mosaic virus (CaMV) 35S promoter in the pPCV702 binaryvector, which was used for Agrobacterium-mediated transformationof tobacco (Nicotiana tabacum var. Samsun NN) leaves (Konczet al., 1989).
Callus generation from transgenic tobacco leaves and
estimation of chitinase activity
Leaf discs were excised from sterile young tobacco plants and grownon Murashige and Skoog (MS) medium containing 3% sucrose,0.8% agar, 5 lg ml�1 a-naphthalene acetic acid (NAA) and 1 lg ml�1
benzylaminopurine (BA). The calli were grown at 25 8C in thedark. Crude extracts of transgenic calli were prepared by grindingthe sterile calli in a glass homogenizer without addition of buffer.The homogenates were centrifuged for 5 min at 14 000 rpm in anEppendorf minifuge and the supernatants were used for the enzymeactivity assay on gels.
Results
Pitcher structure
A longitudinal section of N. khasiana closed mature pitcher(Fig. 1A) shows the glandular bottom part of the pitcher andthe secretory glands (Fig. 1B, C). The glands are situated
within defined depressions of the epidermis (Fig. 1D)partially covered by small epidermal ridges (Fig. 1E).Numerous secretory cells are visualized in each gland (Fig.1D). The glandular secretory zone extends only to abouthalf of the pitcher length, and in the upper half it isexchanged for a waxy epidermal zone with a distinctnetwork of red pigmented cells (Fig. 1F).
Induction of chitinase activity in trap liquid
Plants respond to pathogen attack by induced accumulationof lytic enzymes such as chitinases and b-1,3-glucanases(Punja, 2001). To follow the response of the pitcher toa mimicked pathogen attack and prey sensing, changes inprotein pattern (Fig. 2A) and chitinase activity (Fig. 2B) inthe liquid of a closed trap were analysed before and afterchitin injection. Trap liquid samples were collected prior toand after the injection of soluble colloidal chitin (1 mg perpitcher, pH 5.0) into closed pitchers. Total protein concen-tration in the collected samples was very low, belowdetectable levels of Coomassie staining when determinedon gels or Whatman paper (Marder et al., 1986) or bymeasuring absorbance at 280 nm. Consequently, samples
Fig. 1. Pitcher structure of Nepenthes khasiana. (A) Longitudinalsection of the lower part of a pitcher. Bar=7.5 mm. (B) Bottom part ofpitcher visualized under low magnification. Bar=2.5 mm. (C) Magnifi-cation of secretory glands. Bar=300 lm. (D) Glands full of secretorygranules (arrow). Bar=100 lm. (E) Epidermal ridges (arrow) coveringevery gland. Bar=100 lm. (F) Waxy epidermal zone covering the innersurface of the upper part of a pitcher. Bar=2 mm.
Nepenthes khasiana chitinases 2777
Dow
nloaded from https://academ
ic.oup.com/jxb/article/57/11/2775/682094 by guest on 11 January 2022

were separated on a 12% SDS–polyacrylamide gel andvisualized by silver staining (Fig. 2A). Chitin injectioninduced the synthesis of at least four new proteins andenhanced the production of two proteins that were alsopresent before induction (Fig. 2A, arrows and dots, re-spectively). All the major induced bands were isolated andsubjected to tandem mass spectrometry (MS/MS) analysis(Q-STAR, Applied Biosystems), but no oligopeptidesshowing resemblance to known chitinase sequences wereidentified. When chitinase activity was estimated usingglycol-chitin as a substrate following separation on nativegels, at least three new chitinases (Fig. 2B, lanes 2 and 3indicated by arrows) appeared after 20 h of chitin inductionshowing different migration rates from the non-inducedchitinase (lane 1). Injection of an equal volume of sterilewater into the trap also induced chitinase activity but toa lesser extent (data not shown). Hence, chitin injection intothe trap induced secretion of new proteins including chitinases.
Characterization of the trap liquid chitinase activity
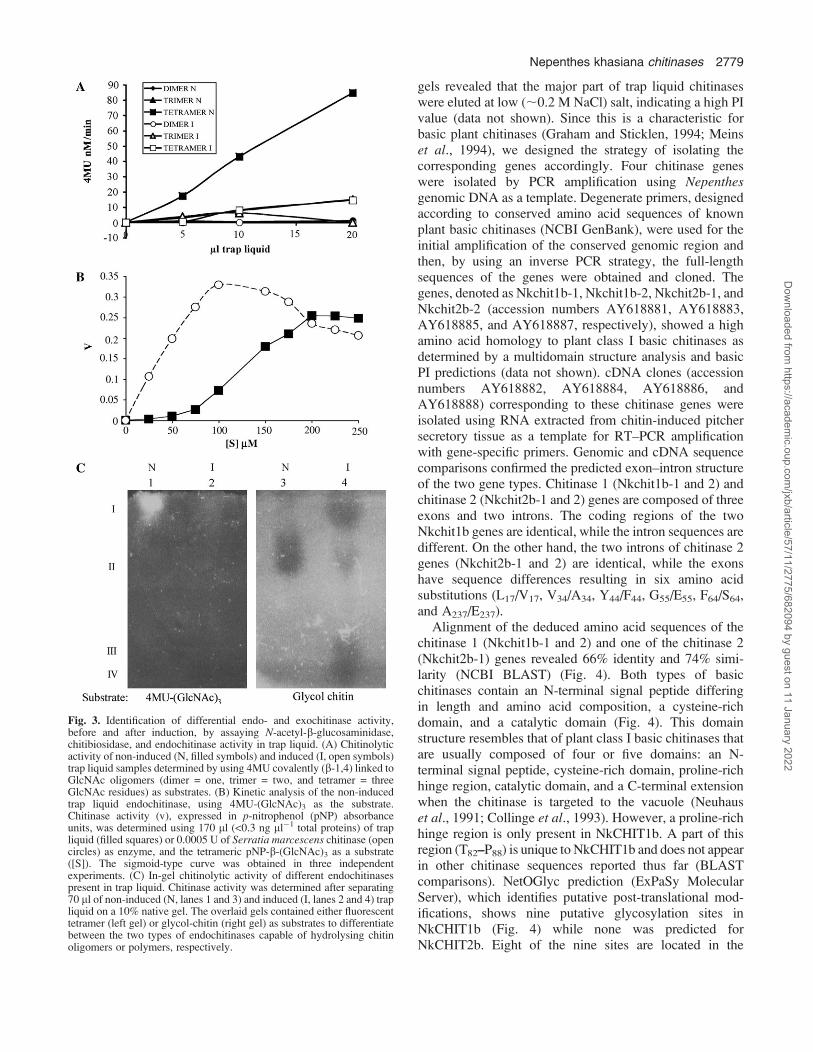
To verify endo- and exochitinase activities, differentiatingGlcNAc oligomeric substrates were used. The release of4MU from 4MU-(GlcNAc)1 (dimer), 4MU-(GlcNAc)2
(trimer), or 4MU-(GlcNAc)3 (tetramer) indicates the pres-ence of N-acetyl-b-glucosaminidase (exochitinase), chito-biosidase, or endochitinase activity, respectively. Noexochitinase activity was detected in either the non-inducedor induced trap liquid (Fig. 3A) since there was no releaseof fluorescent 4MU from 4MU-(GlcNAc)1. Similarly, thelack of ability to release 4MU from 4MU-(GlcNAc)2
indicated no chitobiosidase activity. However, a significantrelease of 4MU from 4MU-(GlcNAc)3 was identified onlyin non-induced trap liquid (Fig. 3A). This endochitinaseactivity of the non-induced liquid exhibited sigmoidkinetics as compared with S. marcescens endochitinase(commercial preparation), used as a control (Fig. 3B).Sigmoid enzyme kinetics may indicate positive substrateco-operativity, thus only allowing estimation of enzymeaffinity towards the substrate, i.e. K0.5 instead of Km. Theestimated K0.5 value for 4MU-(GlcNAc)3 was 130 lM.Only a trace amount of endochitinase activity, as evident by4MU-(GlcNAc)3 hydrolysis, was observed in the inducedliquid (Fig. 3A). This seemed to be in contrast to the resultsshown in Fig. 2B where induction of additional endochi-tinase activities was observed after chitin injection whenglycol-chitin was used as the substrate. To clarify this point,equal amounts of non-induced and induced trap liquid wereseparated on native gels which were subsequently overlaidwith either 4MU-(GlcNAc)3- or glycol-chitin-containinggels as substrates for the chitinase assay. Comparison of thelocation and intensity of the activity spots in the two gels(Fig. 3C) re-confirmed that the induced endochitinasescould only hydrolyse longer chitin polymers (lane 4), butnot the tetrameric substrate (lane 2). Thus, a specificchitinase capable of releasing trimeric GlcNAc oligomers
from 4MU-(GlcNAc)3 is only present in the non-inducedclosed trap. Together, these enzymatic assays showed thatat least two distinct endochitinases, differing in substratespecificity (Fig. 3A, C, lanes 1 and 3), exist in the non-induced liquid, whereas additional endochitinases, that canonly hydrolyse chitin polymers, appeared in the traps afterchitin induction (Fig. 3C, lane 4).
Isolation of Nepenthes chitinase genes
Preliminary separation of trap liquid on a Mono Q ionexchange column followed by chitinase assays on activity
Fig. 2. Chitin injection into closed pitchers induced changes in proteinpattern and chitinase activity of pitcher liquid. Colloidal chitin (1 mg) wasinjected into closed young traps. (A) General protein pattern of the trapliquid: liquid samples (100 ll) taken prior to injection (lane 1) and 4d (lane 2) or 8 d (lane 3) after injection were separated on a 12% SDS–polyacrylamide gel and subsequently stained with a silver stain. Arrowsindicate chitin-induced bands, dots indicate chitin-enhanced bands. (B)Chitinase activity estimated on a native gel: samples (30 ll) taken prior toinjection (lane 1) and 20 h (lane 2) or 5 d (lane 3) after injection.Following electrophoresis, the gel was overlaid with an additional gelcontaining 0.01% (w/v) glycol-chitin and assayed for chitinase activity.Arrows show induced chitinase bands and the dot shows the chitin-enhanced band.
2778 Eilenberg et al.
Dow
nloaded from https://academ
ic.oup.com/jxb/article/57/11/2775/682094 by guest on 11 January 2022
gels revealed that the major part of trap liquid chitinaseswere eluted at low (;0.2 M NaCl) salt, indicating a high PIvalue (data not shown). Since this is a characteristic forbasic plant chitinases (Graham and Sticklen, 1994; Meinset al., 1994), we designed the strategy of isolating thecorresponding genes accordingly. Four chitinase geneswere isolated by PCR amplification using Nepenthesgenomic DNA as a template. Degenerate primers, designedaccording to conserved amino acid sequences of knownplant basic chitinases (NCBI GenBank), were used for theinitial amplification of the conserved genomic region andthen, by using an inverse PCR strategy, the full-lengthsequences of the genes were obtained and cloned. Thegenes, denoted as Nkchit1b-1, Nkchit1b-2, Nkchit2b-1, andNkchit2b-2 (accession numbers AY618881, AY618883,AY618885, and AY618887, respectively), showed a highamino acid homology to plant class I basic chitinases asdetermined by a multidomain structure analysis and basicPI predictions (data not shown). cDNA clones (accessionnumbers AY618882, AY618884, AY618886, andAY618888) corresponding to these chitinase genes wereisolated using RNA extracted from chitin-induced pitchersecretory tissue as a template for RT–PCR amplificationwith gene-specific primers. Genomic and cDNA sequencecomparisons confirmed the predicted exon–intron structureof the two gene types. Chitinase 1 (Nkchit1b-1 and 2) andchitinase 2 (Nkchit2b-1 and 2) genes are composed of threeexons and two introns. The coding regions of the twoNkchit1b genes are identical, while the intron sequences aredifferent. On the other hand, the two introns of chitinase 2genes (Nkchit2b-1 and 2) are identical, while the exonshave sequence differences resulting in six amino acidsubstitutions (L17/V17, V34/A34, Y44/F44, G55/E55, F64/S64,and A237/E237).
Alignment of the deduced amino acid sequences of thechitinase 1 (Nkchit1b-1 and 2) and one of the chitinase 2(Nkchit2b-1) genes revealed 66% identity and 74% simi-larity (NCBI BLAST) (Fig. 4). Both types of basicchitinases contain an N-terminal signal peptide differingin length and amino acid composition, a cysteine-richdomain, and a catalytic domain (Fig. 4). This domainstructure resembles that of plant class I basic chitinases thatare usually composed of four or five domains: an N-terminal signal peptide, cysteine-rich domain, proline-richhinge region, catalytic domain, and a C-terminal extensionwhen the chitinase is targeted to the vacuole (Neuhauset al., 1991; Collinge et al., 1993). However, a proline-richhinge region is only present in NkCHIT1b. A part of thisregion (T82–P88) is unique to NkCHIT1b and does not appearin other chitinase sequences reported thus far (BLASTcomparisons). NetOGlyc prediction (ExPaSy MolecularServer), which identifies putative post-translational mod-ifications, shows nine putative glycosylation sites inNkCHIT1b (Fig. 4) while none was predicted forNkCHIT2b. Eight of the nine sites are located in the
Fig. 3. Identification of differential endo- and exochitinase activity,before and after induction, by assaying N-acetyl-b-glucosaminidase,chitibiosidase, and endochitinase activity in trap liquid. (A) Chitinolyticactivity of non-induced (N, filled symbols) and induced (I, open symbols)trap liquid samples determined by using 4MU covalently (b-1,4) linked toGlcNAc oligomers (dimer = one, trimer = two, and tetramer = threeGlcNAc residues) as substrates. (B) Kinetic analysis of the non-inducedtrap liquid endochitinase, using 4MU-(GlcNAc)3 as the substrate.Chitinase activity (v), expressed in p-nitrophenol (pNP) absorbanceunits, was determined using 170 ll (<0.3 ng ll�1 total proteins) of trapliquid (filled squares) or 0.0005 U of Serratia marcescens chitinase (opencircles) as enzyme, and the tetrameric pNP-b-(GlcNAc)3 as a substrate([S]). The sigmoid-type curve was obtained in three independentexperiments. (C) In-gel chitinolytic activity of different endochitinasespresent in trap liquid. Chitinase activity was determined after separating70 ll of non-induced (N, lanes 1 and 3) and induced (I, lanes 2 and 4) trapliquid on a 10% native gel. The overlaid gels contained either fluorescenttetramer (left gel) or glycol-chitin (right gel) as substrates to differentiatebetween the two types of endochitinases capable of hydrolysing chitinoligomers or polymers, respectively.
Nepenthes khasiana chitinases 2779
Dow
nloaded from https://academ
ic.oup.com/jxb/article/57/11/2775/682094 by guest on 11 January 2022

proline-rich hinge region. Comparison of the C-terminalparts of the two types of chitinases revealed the presenceof a hydrophobic putative extension only in NkCHIT2b(Fig. 4). A BLAST search for amino acid sequenceidentity of NkCHIT1b and NkCHIT2b showed highest(66%) identity to Poa pratensis (AF000966) chitinase andto a basic endochitinase precursor (79%) of Vitis vinifera(P51613), respectively. Conserved amino acid residues,suggested to be of functional importance in all plantchitinases, were identified in both chitinases and areindicated by different symbols in Fig. 4. The eightcysteine residues (C), present at identical positions inthe chitin-binding region of most basic chitinases, areinvolved in the formation of disulphide bridges allowingthe correct folding of domains into a compact activeconformation. The threonine (T) and glutamine (Q)residues, that play a role in the active site geometry, aswell as the glutamic acid (E) and asparagine (N) that areimportant for catalysis (Graham and Sticklen, 1994;Hamel et al., 1997), are all present in both Nepentheschitinases. On the other hand, the conserved tyrosine (Y),
thought to affect substrate binding in the catalytic cleft(Verburg et al., 1993), is altered to phenylalanine (F) inNkCHIT2b. To date, the significance of the Y to Fsubstitution in Nepenthes remains to be determined.
Identification of chitinase(s) present in pitchersecretory cells
Figure 2B shows that chitin injection into closed trapsinduced the appearance of at least three new trap liquidchitinases. Therefore, the presence of Nkchit1b andNkchit2b transcripts in the trap secretory region, beforeand after chitin injection, was estimated. Following RNAextraction from the pitcher secretory region, RT–PCR wasperformed with Nkchit1b- and Nkchit2b-specific primers.Figure 5 clearly demonstrates that the Nkchit2b transcript ispresent in the secretory region of both non-induced (Fig.5A) and induced (Fig. 5B) pitchers while the Nkchit1btranscript is only present in the secretory region afterinduction. Both primer sets yielded longer products whengenomic DNA, instead of cDNA, was used as a template,due to the presence of introns in the amplified region. Thus,
Fig. 4. Amino acid sequence alignment of the two types of N. khasiana basic endochitinases, NkCHIT1b-1 and NkCHIT2b-1. Identical residues areshown in black. The putative leader peptide is marked by a continuous line above it, the cysteine-rich domain by a broken line above it, the proline-richregion by asterisks, and the C-terminal extension by crosses. Vertical arrows pointing upward mark the six amino acid residues differing betweenNkCHIT2b-1 and NkCHIT2b-2. Predicted glycosylation sites (ExPASy—Biochemical Pathways) are marked by open triangles. Residues suggested tobe of functional importance are marked above the sequence by the following symbols: cysteines—involved in disulphide bridges (open circles), glutamicacid and asparagine—important for enzymatic catalysis (filled circles), threonine and glutamine—involved in maintenance of active site geometry (opensquares), tyrosine—suggested to affect substrate binding in the catalytic cleft (filled square).
2780 Eilenberg et al.
Dow
nloaded from https://academ
ic.oup.com/jxb/article/57/11/2775/682094 by guest on 11 January 2022

according to these results, it might be assumed that eitherthe uppermost or lowest endochitinase activity spot shownin Fig. 3C, lane 4, represents Nkchit1b while the activityspots present in both lanes 3 and 4 (middle band) (Fig. 3C)correspond to Nkchit2b, which was shown to be transcribedin the secretory region of both non-induced and inducedpitchers. To confirm this conclusion, the Nkchit1b genomicclone and Nkchit2b cDNA, each driven by the 35Spromoter, were separately introduced into transgenictobacco plants. It was assumed that in tobacco plants thealien chitinases would undergo similar post-translationalmodifications. Figure 6 demonstrates chitinase activity incalli derived from the transgenic tobacco plants. Theintroduced NkCHIT1b migrated slowly, as did the inducedupper band in Fig. 3C, lane 4, while NkCHIT2b co-migrates with the endogenous tobacco chitinase andresembles the migration of the middle band in Fig. 3C,lanes 3 and 4. The slow migration of NkCHIT1b on nativegels in comparison with NkCHIT2b could be due topossible glycosylation of the proline-rich region (Fig. 4).
Discussion
Nepenthes pitchers are highly specialized organs thatcapture and digest insects for an extra supply of organicprecursors when grown in harsh environments. The trappedinsects are digested in the pitcher fluid and then theresulting nutrients are absorbed via activated transporters.Prey digestion has been shown to involve the secretion ofproteases, RNases, and phosphatases (Matthews, 1960;Amagase, 1972; Amagase et al., 1972). The lack ofcomplete digestion of ants by purified proteases fromNepenthes traps (Amagase et al., 1972) had raised thepossibility that additional exoskeleton-digesting enzymes,such as chitinases, are present in the traps. Incubation ofconcentrated Nepenthes trap liquid with colloidal chitinresulted in considerable digestion of chitin (Amagase et al.,1972). However, this observed activity was partiallyattributed to the presence of chitinase-producing symbioticmicro-organisms in the open traps. The present studyclearly defines the involvement of plant chitinases by usingclosed Nepenthes pitchers that allowed the estimation of netplant chitinase activity in sterile pitcher liquid prior to anyprey intervention and the appearance of secondary micro-organisms. To mimic signals induced by insect capture,colloidal chitin was injected into the closed pitchers understerile conditions and the liquid was then analysed atdifferent time intervals.
The presence of at least two different chitinases in the N.khasiana pitcher liquid was shown under non-inducedconditions. These chitinases differed in their migration rateon native gels and in substrate specificity estimated by theability to release (GlcNAc)3 from the tetramer 4MU-(GlcNAc)3 or to hydrolyse glycol-chitin (Fig. 3A, C, lanes
Fig. 5. Differential presence of Nkchit1b and Nkchit2b transcripts in thepitcher secretory region before and after induction. RT–PCR analysis wasperformed on mRNA isolated from closed pitchers either before (A) or 4d after (B) chitin injection. Lanes 1–3 represent Nkchit1b amplification,and lanes 4–8 represent Nkchit2b amplification with gene-specificprimers. Lanes 1 and 4, reactions with reverse transcriptase (+RT); lanes2 and 5, reactions without reverse transcriptase (–RT); lanes 3 and 6,amplification with genomic DNA as a template; lane 7, Nkchit2b 59primer control; and lane 8, Nkchit2b 39 primer control.
Fig. 6. In-gel activity assay of NkCHIT1b and NkCHIT2b expressed incalli derived from transgenic tobacco plants. Crude extracts of transgeniccalli constitutively expressing NkCHIT2b (lanes 1 and 2) or NkCHIT1b(lanes 3 and 4) and wild-type NN (lanes 5 and 6) were loaded onto a 10%native gel. The amounts of loaded protein are indicated below the lanes.After electrophoresis, the gel was overlaid with an additional gelcontaining 0.01% (w/v) glycol-chitin and incubated for 2 h at 37 8C.Chitinase activity was detected by UV light after staining with 0.01%(w/v) Calcofluor white M2R for 5 min.
Nepenthes khasiana chitinases 2781
Dow
nloaded from https://academ
ic.oup.com/jxb/article/57/11/2775/682094 by guest on 11 January 2022

1 and 3). The sigmoid velocity curve, observed whenincreasing tetramer concentrations were used as the sub-strate (Fig. 3B), indicated enzyme–substrate co-operativityand the requirement for a certain threshold substrateconcentration in the trap to increase the activity rate ofthis chitinase significantly. The second endochitinase,identified in the pitcher liquid under non-induced condi-tions, could only hydrolyse long chitin polymers (Fig. 3C,lane 3).
Injection of colloidal chitin into the closed pitchersmimicked prey trapping and induced the secretion of atleast three additional active chitinases (Fig. 2B). Such aninduced reaction resembles the plant response to pathogenattack when chitinases and other PR proteins are induced bydifferent pathogens and specific elicitors (Punja, 2001).Chitin and chitosan were shown to elicit chitinase activityin bacteria (e.g. Vibrio harveyi), fungi (e.g. Trichodermaharzianum), plant suspension cultures, and leaves of thecarnivorous plant Drosera rotundifolia (Haran et al., 1995;Svitil et al., 1997; Wu et al., 1997; Matusikova et al.,2005). Mechanical wounding was also shown to up-regulate chitinase isoenzymes differentially in tomato andwheat tissues (Botha et al., 1998; Wu and Bradford, 2003).In Nepenthes pitchers, the chitin-induced chitinases wereonly able to hydrolyse glycol-chitin (Fig. 2B, Fig. 3C lane4), but not 4MU-(GlcNAc)3 or shorter oligomers (Fig. 3A,C, lane 2). Similarly, when hydrolysis rates of S. marces-cens chitinases A and B were estimated using 4MU-chito-oligosaccharides as substrates, they did not fully correlatewith the whole chitinolytic activity measured by usingchitin or GlcNAc polymers as substrates (Brurberg et al.,1996, 2001), indicating the presence of endochitinases withspecificities for long polymers in addition to those that canhydrolyse short oligomers. The occurrence in the trap liquidof multiple forms of basic endochitinases, exhibitingdifferent substrate specificities, emphasizes the importantrole that chitinases play in the trap response to preycapturing. It is possible that in Nepenthes the combinedaction of the two endochitinases, initially present in theclosed sterile trap liquid, first cleaves the long polymers andthen releases GlcNAc oligomers from already producedends of the chitin exoskeleton of the intruding prey. Thesmall oligomers, in turn, induce the synthesis of a repertoireof new chitinases possessing the required endochitinasespecificities for further efficient digestion of the prey chitinskeleton whose action probably allows the penetration andinteraction of the inducible nucleases and proteases withtheir substrates (Gallie and Chang, 1997).
In tobacco, 10 clearly defined endo-type chitinases withdifferent substrate specificities, that act synergistically,appeared at the initial stages of the defence response(Brunner et al., 1998). Such a platform of different classesof chitinases can liberate hydrolysed products with variouslengths, which may possess variable abilities to elicitdefence reactions (Roby et al., 1987; Brunner et al., 1998).
Until this study, no Nepenthes chitinase genes have beencharacterized. Here the isolation of four Nepenthes chiti-nase genes (Nkchit1b-1 and 2, and Nkchit2b-1 and 2)belonging to two subgroups of basic chitinases, accordingto amino acid sequence homology and domain structure, isdescribed. Both Nkchit1b and Nkchit2b genes are com-posed of three exons and two introns and contain a putativeleader peptide sequence at the 59 end of the coding region.The Nkchit1b group contains two genes sharing identicalexons and different intron sequences. The two Nkchit2bgenes code for proteins differing from each other in sixamino acid residues but having identical introns. Since onlyone of the Nkchit1b promoters has been isolated, it isdifficult to estimate why the organism has retained twodifferent genes encoding an identical enzyme. Geneduplication could have occurred long ago, since the intronsequences differ significantly. On the other hand, the exonpreservation suggests an important function for theNkCHIT1b enzyme. It is also interesting to note thatNkCHIT1b and NkCHIT2b share 66% amino acid homol-ogy, while a higher identity exists between NkCHIT2b andseveral non-carnivorous plant chitinases, for example,chitinases from V. vinifera (79%), Ulmus pumila (74%),Persea americana (75%), and Medicago truncatula (73%).
NkCHIT2b can be considered as a constitutively ex-pressed housekeeping chitinase produced in both non-induced and induced secretory tissue (Fig. 5A, B). Accordingto the sequence of its C-terminal end (NGLLWATE),NkCHIT2b is probably targeted to vacuoles (Chrispeelsand Raikhel, 1992). NkCHIT1b, on the other hand, which issynthesized under induced conditions, lacks the C-terminaltargeting signal and is secreted into the pitcher fluid. It alsocontains a proline-rich domain with eight putative gly-cosylation sites.
This study has unravelled the role of chitinases ininducing the trap digesting machinery and also definedthe involvement of genes encoding basic chitinases in thismechanism. The differential expression of the newlycharacterized chitinases, before and after chitin induction,and the identification of chitinases with different substratespecificities suggest that the insect trap of carnivorousplants is an advanced and efficient modification of the plantdefence mechanism. In these traps, traces of GlcNAcoligomers, specifically produced by the sequential actionof long polymer- and short oligomer-cleaving enzymes,induce a massive production of endochitinases that digestthe trapped insect’s inner and outer skeleton to providecarbon and nitrogen sources.
Since some Nepenthes pitchers can contain >3.0 l ofsecreted liquid, chitin induction might be utilized as a toolfor inducing the synthesis of large quantities of solubleendochitinases. Moreover, GlcNAc oligomers were foundto play a role in various medical treatments, such as woundhealing (Cole et al., 1999; Schwaitzberg et al., 2004), andtheir selective production by plant-derived enzymes in the
2782 Eilenberg et al.
Dow
nloaded from https://academ
ic.oup.com/jxb/article/57/11/2775/682094 by guest on 11 January 2022

closed non-induced pitchers devoid of any animal virusesor bacterial contamination may offer a great biotechnolog-ical advantage.
Acknowledgements
This study was supported by Biotech M.A.H. (Makhteshim-Agan-Hazera) Brurim M.P. Shikmim 79837, Israel. We thankShoshan Haran for helpful discussions.
References
Amagase S. 1972. Digestive enzymes in insectivorous plants. III.Acid proteases in the genus Nepenthes and Drosera peltata.Journal of Biochemistry 72, 73–81.
Amagase S, Mori M, Nakayama S. 1972. Digestive enzymes ininsectivorous plants. IV. Enzymatic digestion of insects byNepenthes secretion and Drosera peltata extract: proteolytic andchitinolytic activities. Journal of Biochemistry 72, 765–767.
An CI, Fukusaki E, Kobayashi A. 2001. Plasma-membrane H+-ATPases are expressed in pitchers of the carnivorous plantNepenthes alata Blanco. Planta 212, 547–655.
An CI, Fukusaki E, Kobayashi A. 2002. Aspartic proteinases areexpressed in pitchers of the carnivorous plant Nepenthes alataBlanco. Planta 214, 661–667.
Botha A-MNM, Van der Westhuizen AJ, Botha FC. 1998.Chitinase isoenzymes in near-isogenic wheat lines challengedwith Russian wheat aphid, exogenous ethylene and mechanicalwounding. Botanical Bulletin of Academia Sinica 39, 99–106.
Brunner F, Stintzi A, Fritig B, Legrand M. 1998. Substratespecificities of tobacco chitinases. The Plant Journal 14,225–234.
Brurberg MB, Nes IF, Eijsink VG. 1996. Comparative studies ofchitinases A and B from Serratia marcescens. Microbiology 142,1581–1589.
Brurberg MB, Synstad B, Klemsdal SS, van Aalten DMF,Eijsink VG. 2001. Chitinases from Serratia marcescens. RecentResearch Developments in Microbiology 5, 187–204.
Chrispeels MJ, Raikhel NV. 1992. Short peptide domain targetproteins to plant vacuoles. Cell 68, 613–616.
Cole DJCR, ChanMW, Schwaitzberg SD, Byrne TK, Adams DB,Baron PL, O’Brien PH,Metcalf JS, DemchevaM, Vournakis J.1999. A pilot study evaluating the efficacy of a fully acetylatedpoly-N-acetyl glucosamine membrane formulation as a topicalhemostatic agent. Surgery 126, 510–517.
Collinge DB, Kragh KM, Mikkelsen JD, Nielsen KK,Rasmussen U, Vad K. 1993. Plant chitinases. The Plant Journal3, 31–40.
Darwin C. 1875. Insectivorous plants. John Murray, London.Davis BJ. 1964. Disc electrophoresis. II. Method and application to
human serum proteins. Annals of the New York Academy ofSciences 121, 404–427.
deJongA,CordewenerJ,LoSchiavoF,TerziM,VandekerckhoveJ,Van Kammen A, De Vries SC. 1992. A carrot somatic embryomutant is rescued by chitinase. The Plant Cell 4, 425–433.
Gallie DR, Chang SC. 1997. Signal transduction in the carnivorousplant Sarracenia purpurea. Plant Physiology 115, 1461–1471.
Goormachtig S, Lievens S, Van de Velde W, Van Montagu M,Holsters M. 1998. Srchi 13, a novel early nodulin from Sesbaniarostrata, is related to acidic class III chitinases. The Plant Cell 4,425–433.
Graham LS, Sticklen MB. 1994. Plant chitinases. CanadianJournal of Botany 72, 1057–1083.
Hamel F, Boivin R, Tremblay C, Bellemare G. 1997. Structuraland evolutionary relationships among chitinases of floweringplants. Journal of Molecular Evolution 44, 614–624.
Haran S, Schikler H, Oppenheim A, Chet I. 1995. Newcomponents of the chitinolytic system of Trichoderma harzianum.Mycological Research 99, 441–446.
Helleboid S, Hendriks T, Bauw G, Inze D, Vasseur J, Hilbert JL.2000. Three major somatic embryogenesis related proteins inCichorium identified as PR proteins. Journal of ExperimentalBotany 51, 1189–1200.
Heslop-Harrison Y. 1975. Enzyme release in carnivorous plants.Frontiers in Biology 43, 525–578.
Kasprzewska A. 2003. Plant chitinases—regulation and function.Cellular and Molecular Biology Letters 8, 809–824.
Koncz C,Martini N,Mayerhofer R, Koncz-Kalman Z, KorberH,Redei GP, Schell J. 1989. High-frequency T-DNA-mediated genetagging in plants. Proceedings of the National Academy of Sciences,USA 86, 8467–8471.
Kuranda MJ, Robbins PW. 1991. Chitinase is required for cellseparation during growth of Saccharomyces cerevisiae. Journal ofBiological Chemistry 266, 19758–19767.
Marder JB, Mattoo AK, Edelman M. 1986. Identification andcharacterization of the psbA gene product: the 32-kDa chloroplastmembrane protein. Methods of Enzymology 118, 384–396.
Mathews R. 1960. A ribonuclease from Nepenthes spp. Biochimia etBiophysica Acta 28, 552–553.
Matusikova I, Salaj J, Moravcikova J, Mlynarova L, Nap J-P,Libantova J. 2005. Tentacles of in vitro-grown round-leaf sundew(Drosera rotundifolia L.) show induction of chitinase activity uponmimicking the presence of prey. Planta 222, 1020–1027.
Meins Jr F, Fritig B, Linthorst HJM,Mikkelsen JD, Neuhaus JM,Ryals J. 1994. Plant chitinase genes. Plant Molecular BiologyReporter 12, S22–S28.
Neuhaus JM, Sticher L, Meins F, Boller T. 1991. A short C-terminal sequence is necessary and sufficient for the targeting ofchitinases to the plant vacuole. Proceedings of the NationalAcademy of Sciences, USA 88, 10362–10366.
Owen Jr TP, Lennon KA, Santo MJ, Anderson AN. 1999.Pathways for nutrient transport in the pitchers of the carnivorousplant Nepenthes alata. Annals of Botany 84, 459–466.
Passarinho PA, Van Hengel AJ, Fransz PF, de Vries SC. 2001.Expression pattern of the Arabidopsis thaliana AtEP3/AtchitIVendochitinase gene. Planta 212, 556–567.
Punja Z. 2001. Genetic engineering of plants to enhance resistance tofungal pathogens—a review of progress and future prospects.Canadian Journal of Plant Pathology 23, 216–235.
Roby D, Gadelle A, Toppan A. 1987. Chitin oligosaccharides aselicitors of chitinase activity in melon plants. Biochemical andBiophysical Research Communications 143, 885–892.
Schulze W, Frommer WB, Ward JM. 1999. Transporters forammonium, amino acids and peptides are expressed in pitchers ofthe carnivorous plant Nepenthes. The Plant Journal 17, 637–646.
Schwaitzberg SD, Chan MW, Cole DJ, Read M, Nichols T,Bellinger D, Connolly RJ. 2004. Comparison of poly-N-acetylglucosamine with commercially available topical hemostats forachieving hemostasis in coagulopathic models of splenic hemor-rhage. Journal of Trauma-Injury Infection and Critical Care 57,Supplement, S29–S32.
Sirova D, Adamec L, Vrba J. 2003. Enzymatic activities in trapsof four aquatic species of the carnivorous genus Utricularia. NewPhytologist 159, 669–675.
Svitil AL, Ni9 Chadhain SM, Moore JA, Kirchman DL. 1997.Chitin degradation proteins produced by marine bacterium Vibrioharveyi growing on different forms of chitin. Applied andEnvironmental Microbiology 63, 408–413.
Nepenthes khasiana chitinases 2783
Dow
nloaded from https://academ
ic.oup.com/jxb/article/57/11/2775/682094 by guest on 11 January 2022

Tronsmo A, Harman G. 1993. Detection and quantification ofN-acetyl-b-D-glucosaminidase, chitibiosidase, and endochitinasein solutions and on gels. Analytical Biochemistry 208,74–79.
Trudel J, Asselin A. 1989. Detection of chitinase activity afterpolyacrylamide gel electrophoresis. Analytical Biochemistry 178,362–366.
Verburg JG, Rangwala SH, Samac DA, Luckow VA, Huynh QK.1993. Examination of the role of tyrosine-174 in the catalyticmechanism of the Arabidopsis thaliana-chitinase: comparison ofvariant chitinases generated by site-directed mutagenesis and
expressed in insect cells using baculovirus vectors. Archives ofBiochemistry and Biophysics 300, 223–230.
Vessey JC, Pegg GF. 1973. Chitinase in Verticillium. Transactionsof the British Mycological Society 60, 133–143.
Wu CT, Bradford KJ. 2003. Class I chitinase and beta-1,3-glucanase are differentially regulated by wounding, methyljasmonate, ethylene and gibberellin in tomato seeds and leaves.Plant Physiology 133, 263–273.
Wu H, Echt CS, Popp MP, Davis JM. 1997. Molecular cloning,structure and expression of an elicitor-inducible chitinase genefrom pine trees. Plant Molecular Biology 33, 979–987.
2784 Eilenberg et al.
Dow
nloaded from https://academ
ic.oup.com/jxb/article/57/11/2775/682094 by guest on 11 January 2022




















