
Panax ginseng as an adjuvant treatment for Alzheimer's disease

Isolation and characterization of Panax ginsenggeranylgeranyl-diphosphate synthase genes respondingto drought stress
Shadi Rahimi & Yu-Jin Kim & Balusamy Sri Renuka Devi &Altanzul Khorolragchaa & Johan Sukweenadhi & Deok-Chun Yang
Accepted: 24 March 2015# Koninklijke Nederlandse Planteziektenkundige Vereniging 2015
Abstract Geranylgeranyl-diphosphate synthases(GGDPS) catalyze branch point enzymatic reactionsproducing isoprenoid-derived products which are nec-essary for plant growth and responses to a wide range ofbiotic and abiotic stresses. In our study, full lengthgeranylgeranyl-diphosphate synthase 1 (PgGGDPS1)and 2 (PgGGDPS2) cDNAwere isolated and character-ized from the flower of Panax ginseng and 4-year oldP. ginseng cv. Gumpoong. The cDNA had openreading frame of 1032 and 1116 bp with a deducedamino acid sequence of 343 and 371 residues forGGDPS1 and GGDPS2, respectively. The calculat-ed molecular mass of GGDPS1 and GGDPS2 wereapproximately 37.66 and 40.21 kDa with a predi-cated isoelectric point of 5.32 and 6.23 and predict-ed localization of plastid. A GenBank Blast Xsearch revealed that the deduced amino acid ofPgGGDPS1 shared a high degree of homology withGGDPS from Panax notoginseng. The transcriptionpattern of GGDPS genes was different at various
developmental stages. Both GGDPS genes werehighly expressed in aerial parts of the plant, espe-cially in rapidly growing tissues such as 4-year oldflower and stem tissues. Transcript level ofPgGGDPS1 was differentially induced in ginsengnot only during Pseudomonas syringae pv tomatoinfection but also after exposure to abiotic stresses.Our results suggested that the induction of GGDPSgenes specifically PgGGDPS1 by drought stressmay affect chlorophyll levels, intracellular GA con-tent and accumulation of carotenoids as the precur-sor for higher production of ABA and possiblystomatal closure as the barrier for water loss.
Keywords Drought . Gene characterization .
Geranylgeranyl diphosphate synthase . OrganExpression .Panax ginseng
AbbreviationsABA abscisic acidCLD chain length determinationEST expressed sequence tagsGA3 gibberellinGGDP geranylgeranyl diphosphateGGDPS geranylgeranyl-diphosphate synthaseGGR geranylgeranyl reductaseMJ methyl jasmonatesRT-PCR
reverse transcriptase-polymerase chainreaction
SA salicylic acidYE yeast extract
Eur J Plant PatholDOI 10.1007/s10658-015-0648-1
Y.<J. Kim :D.<C. Yang (*)Department of Oriental Medicinal Biotechnology, College ofLife Sciences, Kyung Hee University, Yongin 446-701,Republic of Koreae-mail: [email protected]
S. Rahimi :Y.<J. Kim : B. S. R. Devi :A. Khorolragchaa :J. Sukweenadhi :D.<C. YangGraduate School of Biotechnology and Ginseng Bank,College of Life Sciences, Kyung Hee University,Yongin 446-701, Republic of Korea

Introduction
Geranylgeranyl diphosphate (GGDP) is the key precur-sor for biosynthesis of several important terpenoidswhich are necessary for plant growth and response toenvironmental changes including diterpenes, gibberel-lins, carotenoids, and the isoprenoid side chain of chlo-rophyll (Fig. 1c). It is synthesized by branch pointenzymatic reaction of geranylgeranyl pyrophosphatesynthase which functions in condensation ofisopentenyl diphosphate (IPP) and other substrates suchas farnesyl diphosphate (FDP) to produce C20 GGDP.Prenyl diphosphate synthases, the enzymes utilizingIPP, are localized in three subcellular compartments:the cytosol, mitochondria, and plastids (Gray 1987;Kleinig 1989). In plants, few data are availableconcerning the enzymes of the prenyltransferase familyand their regulation at the molecular level. In plant cells,isoprenoids such as carotenoids, chlorophylls, and ent-kaurene have been shown to be synthesized fromGGDPin plastids (Bartley and Scolnik 1995; Hedden andKamiya 1997). Plant growth hormone of gibberellin isderived from GGDP with two-step cyclization viacopalyl diphosphate that produced a common precursor,ent-kaurene. Carotenoids with protective functionagainst harmful effects in photo-oxidative process arelipophilic isoprenoids produced by head-to-head con-densation of two GGDP molecules. Subsequentlyabscisic acid (ABA) as the plant stress hormone is thecarotenoid-derived signaling molecule. GGDP is re-duced to phytyl pyrophosphate which forms essentialphytol side chain for chlorophyll biosynthesis. GGDPsynthase (GGDPS) genes have been characterized fromCapsicum annuum (Kuntz et al. 1992; Badillo et al.1995), Sinapis alba (Laferriere and Beyer 1991),Lupinus albus (Aitken et al. 1995),Catharanthus roseus(Bantignies et al. 1995), and Arabidopsis (Scolnik andBartley 1994; Scolnik and Bartley 1995; Scolnik andBartley 1996; Zhu et al. 1997a, b). GGDPS genes havebeen cloned in a number of organisms including;Arabidopsis thaliana (Okada et al. 2000; Zhu et al.1997a, b), Taxus canadensis (Hefner et al. 1998),Helianthus annuus (Oh et al. 2000), Scoparia dulcisand Croton sublyratus (Sitthithaworn et al. 2001), orSolanum lycopersicon (Ament et al. 2006).
Panax ginseng Meyer, a perennial herbaceous plantfrom the Araliaceae family, is one of the most common-ly utilized medicinal plants (Lee and Kim 2014; Songet al. 2014). Various abiotic stresses such as drought,
salinity, and adverse temperatures might affect cropplants and due to long cultivation period of ginseng, thishighly valued medicinal plant is more susceptible toecological stresses. Ginseng plants activate distinct de-fense responses to adapt to adverse conditions duringtheir growth and development. In this report, we attemptto identify the GGDPS genes in P. ginseng and under-stand the biological significance of these genes in re-sponse to environmental stresses. In addition, themRNA abundance of these GGDPS genes at differentdevelopmental stages and in different organs wasexamined.
Materials and methods
Plant materials and growth conditions
P. ginseng adventitious roots were collected fromGinseng Bank, Kyung Hee University and grown for1 month in liquid Murashige & Skoog medium(Murashige and Skoog 1962) supplemented with 3 mgl−1 indole-3-butyric acid (IBA) at 25 °C. The roots weremaintained by regular subculture in every 4 weeks.Embryo, 1-month old seedling and different organs(leaf, stem, flower bud, main root, lateral root, rhizome)of 2-, 4- and 6-year old healthy ginseng plants werecollected from Ginseng Bank, Korea.
RNA purification and construction of a cDNA library
Total RNAwas isolated from the 4-year old Gumpoongcultivar and flower of P. ginseng and RNAwas isolatedusing the aqueous phenol extraction procedure de-scribed by Morris et al. (1990). Poly (A) + RNA wasisolated by an oligo (dT) cellulose column using thePoly (A) Quick mRNA isolation kit (Stratagene,USA).A commercial cDNA synthesis kitwas used to constructa library according to the manufacturer’s instructionmanual (Clontech, USA). Size-selected cDNA was li-gated into a λTriplEx2 vector and was packaged in vitrousing Gigapack III Gold Packaging Extract kits(Stratagene). pTriplEx phagemids were excised fromthe λpTriplEx2 and used as templates for sequenceanalysis. The 5′ ends of the cDNA inserts were se-quenced by an automatic DNA sequencer (ABI Prism3700, Perkin-Elmer, USA) (Kim et al. 2014).
Eur J Plant Pathol
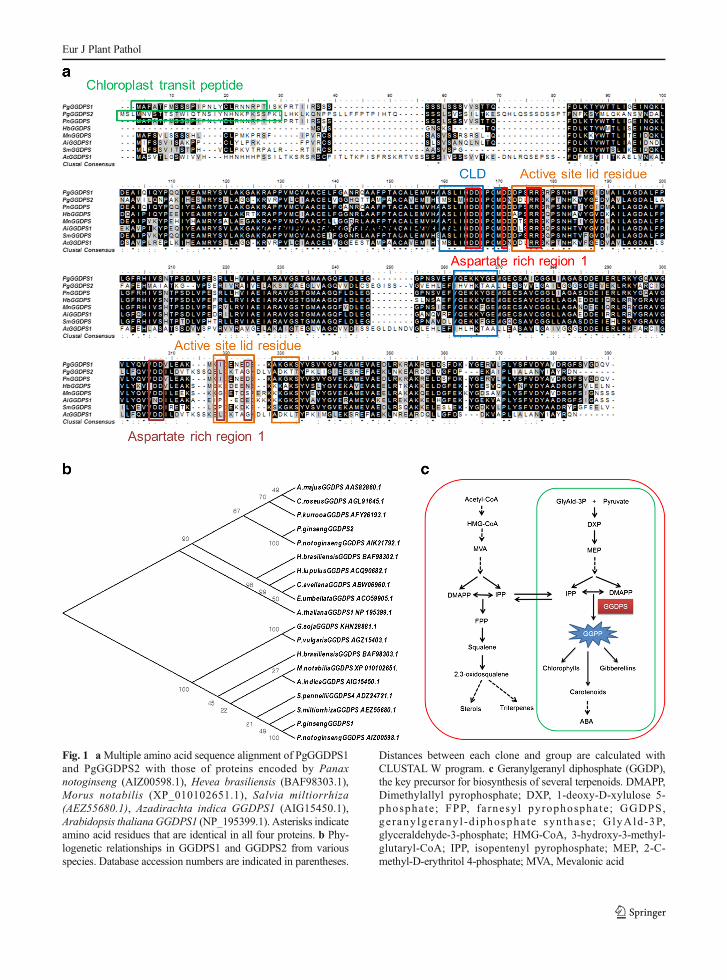
Fig. 1 aMultiple amino acid sequence alignment of PgGGDPS1and PgGGDPS2 with those of proteins encoded by Panaxnotoginseng (AIZ00598.1), Hevea brasiliensis (BAF98303.1),Morus notabilis (XP_010102651.1), Salvia miltiorrhiza(AEZ55680.1), Azadirachta indica GGDPS1 (AIG15450.1),Arabidopsis thaliana GGDPS1 (NP_195399.1). Asterisks indicateamino acid residues that are identical in all four proteins. b Phy-logenetic relationships in GGDPS1 and GGDPS2 from variousspecies. Database accession numbers are indicated in parentheses.
Distances between each clone and group are calculated withCLUSTAL W program. c Geranylgeranyl diphosphate (GGDP),the key precursor for biosynthesis of several terpenoids. DMAPP,Dimethylallyl pyrophosphate; DXP, 1-deoxy-D-xylulose 5-phosphate ; FPP, fa rnesy l pyrophosphate ; GGDPS,ge rany lge rany l -d iphospha te syn thase ; GlyAld-3P,glyceraldehyde-3-phosphate; HMG-CoA, 3-hydroxy-3-methyl-glutaryl-CoA; IPP, isopentenyl pyrophosphate; MEP, 2-C-methyl-D-erythritol 4-phosphate; MVA, Mevalonic acid
Eur J Plant Pathol

Sequence analysis
Homologous sequences of GGDPS EST were searchedagainst the GenBank databases using a BLASTX algo-rithm. A pTriplEx phagemid for GGDPS cDNA wasexcised from the λpTriplEx2 and used as a template forsequence analysis. Nucleotide and amino acid sequenceanalyses were performed using the DNASIS program(Hitachi, Japan) (Kim et al. 2014).
We identified and selected GGDPS genes based ontheir open reading frames encoding the specific proteinvia BlastX program (NCBI BlastX program). ClustalXwith default gap penalties was used to perform multiplealignment of GGDPS1 and GGDPS2 isolated fromP. ginseng and previously registered in other species.A phylogenetic tree was constructed by the neighbor-joining method, and the reliability of each node wasestablished by bootstrap methods using MEGA4 soft-ware. The protein properties were estimated usingProtParam (Artimo et al. 2012). Proposed signal peptideis predicted from PSORT (K. Nakai, Tokyo University,http://psort.ims.u-tokyo.ac.jp/).
Stress treatment
To investigate the response of the PgGGDPS gene tovarious elicitors, 100 μM methyl jasmonate (MJ),200 μM salicylic acid (SA) and 3 g l−1 yeast extract(YE) were applied to 30 days old subcultured adventi-tious roots, and harvested at 6, 12, 24, 48, 72 h aftertreatment. TheP. ginseng seeds used in the present studywere obtained from the Ginseng Bank, South Korea.The 4 week-old ginseng seedlings germinated in vitrocondition on solidified MS media (Duchefa biochem,Netherland) with 3 % sucrose, supplemented withgibberellic acid (10 mg l−1) at 25 °C under a 16 hphotoperiod and used for the treatment studies. For coldtreatment, ginseng plants were exposed to low temper-ature (4 °C). To induce drought stress, plants wereexposed to dehydration by withholding water. To eval-uate the water loss (g) during the drought treatment,plants were weighted before and after treatment andwater loss was calculated as Water loss (g) = Freshweight before treatment (g) - Fresh weight after treat-ment (g).
For salt treatment, ginseng plants were removed fromthe media and soaked in 150 mM NaCl. Ginseng plantswere inoculated with the virulent Pseudomonassyringae pv tomato strain DC3000. The bacteria were
grown at 28 °C in King’s B medium containing rifam-picin (50 mg l−1). To study bacterial infection, bacterialsuspensions (5×106 cfu ml−1) were infiltrated into theginseng leaves using a syringe. At appropriate timepoints, three independent leaves infiltrated with Pst wereharvested. For stress treatments, the plantlets were sub-jected to treatment for 6, 12, 24, 48 and 72 h. Controlplants were grown at 25 °C under a 16 h photoperiod.
RNA extraction and quantitative RT-PCR analysis
Total RNA was extracted from adventitious roots ofP. ginseng using RNeasy mini kit (Qiagen, Valencia,CA, USA). For RT-PCR, 200 ng of total RNAwas usedas a template for reverse transcription using oligo (dT)15primer (0.2mM) and AMVReverse Transcriptase (10 Uμl−1) (INTRON Biotechnology, Inc., South Korea) ac-cording to the manufacturer’s instructions. Real-timequantitative PCR was performed using 100 ng ofcDNA in a 10-μl reaction volume using SYBR®Green Sensimix Plus Master Mix (Quantace, Watford,England). Gene-specific primers for GGDPS1,(forward) 5′-CAG AAG CTG GAC GAA GCA ATTC-3′ and (reverse) 5′-CAT CCATGC ATG GCA AGTCAT-3′, GGDPS2, (forward) 5′-CTT AGA AAA TATGCT AGG TGT ATC GGG-3′and (reverse) 5′-AAATCC AAC AAG CTG ATC CCG A-3′ were used toperform quantitative RT-PCR. The thermal cycler con-ditions recommended by the manufacturer were used asfollows: 10 min at 95 °C, followed 40 cycles of 95 °Cfor 10 s, 58 °C for 10 s, and 72 °C 20 s. The fluorescentproduct was detected at the last step of each cycle.Relative quantity of the PgGGDPS genes transcriptionlevel was performed using Rotor-Gene 6000 real-timerotary analyzer (Corbett Life Science, Sydney,Australia), and calculated using the comparative cyclethreshold (CT) method according to the manufacturers’instructions for normalizing data. A constitutivelyexpressed β-actin gene with primer (forward) 5′-CGTGAT CTT ACA GAT AGC TTG ATG A-3′ and(reverse) 5′-AGA GAA GCT AAG ATT GAT CCTCC-3′was used as internal reference. Three independentexperiments were performed. The primer efficiencieswere determined according to the method by Livakand Schmittgen (2001) to validate the ΔΔCt method.The observed slopes were close to zero, indicating thatthe efficiencies of the gene and the internal control β -actin were equal.
Eur J Plant Pathol

Determination of endogenous gibberellin (GA3)and ABA hormones and chlorophyll and carotenoidcontent
Fresh leaf sample was homogenized with 70 % (v/v)methanol and stirred overnight at 4 °C. The extract wasfilltered through a Whatman filter and the methanolevaporated under vacuum. The aqueous phase was ad-justed to pH 8.5 with 0.1 M phosphate buffer and thenpartitioned with ethyl acetate three times. After removalof the ethyl acetate phase, the pH of the aqueous phasewas adjusted to 2.5 with 1 N HCl. The solution waspartitioned with diethyl ether three times, and thenpassed through andydrous sodium sulfate. After thatthe diethyl ether phase was evaporated under vacuumand the dry residue containing hormones was dissolvedin 2.0 ml of methanol and stored in vials at 4 °C. GA3and ABA were quantified by reversed phase liquidchromatography as described previously Kelen et al.2004. GA3 and ABA were purchased from Mb Cell(Seoul, Korea) and Sigma Chemical Company (St.Louis, USA).
The chlorophyll and carotenoid content of ginsengseedlings leaves were estimated according to previousmethod (Kirk and Allen 1965).
Statistical analysis
Means of three independent replicates are statisticallyanalyzed and compared with control at (*p<0.05,**p<0.01, ***p<0.001) using Student’s t-test.
Results
Isolation and Characterization of cDNA EncodingPgGGDPS Genes
As part of a genomic project to identify genes of themedicinal plant P. ginseng, a cDNA library consistingabout 20,000 cDNAs were previously constructed.Moreover, a full genomic DNA sequence was analyzedusing the genomic DNA sequence retrieved from gin-seng genome database (http://im-crop.snu.ac.kr/new/index.php). There are fewer studies of GGDPS, thus,this study will provide more information about GGDPSto characterize it from other plants. cDNA encoding twogeranylgeranyl pyrophosphate synthases, designatedPgGGDPS1 and PgGGDPS2 were isolated. As is
shown in Fig. 1a, the cDNAs had open reading frameof 1032 and 1116 bp with a deduced amino acidsequence of 343 and 371 residues for GGDPS1 andGGDPS2, respectively. The calculated molecular massof these proteins are approximately 37.66 and 40.21 kDa with a predicated isoelectric point of 5.32 and6.23 for GGDPS1 and GGDPS2, respectively. In thededuced amino acid sequence of PgGGDPS1 andPgGGDPS2 proteins, the total number of negativelycharged residues (Asp and Glu) was 45 and 42 whilethe total number of positively charged residues (Arg andLys) were 37 in both proteins (ProtParam). A GenBankBlast search (http://www.ncbi.nlm.nih.gov/BLAST/)resulting in highly conserved domains of Trans-Isoprenyl Diphosphate Synthases possessed by all thepreviously cloned plant and indicated that PgGGDPS1and PgGGDPS2 belonged to the GGDPS family. Greenbox in Fig. 1a is the proposed signal peptide predictedby PSORT program. The predicted PgGGDPS1 andPgGGDPS2 transit peptide was also identified byTargetP (http://www.cbs.dtu.dk/services/TargetP/),showing that PgGGDPS1 and PgGGDPS2 had atransit peptide at its N-terminal which may suggestchloroplast localization (Emanuelson et al. 2000). Thecatalytic site of GGDPS enzyme consists of a largecentral cavity formed by mostly antiparallel alpha heli-ces with two aspartate-rich regions (DDXX(XX)D) lo-cated on opposite walls. These residues mediate bindingof prenyl phosphates via bridging Mg2+ ions, inducingproposed conformational changes that close the activesite to solvent, protecting and stabilizing reactivecarbocation intermediates. 25 active site lid resi-dues in Orange box (Fig. 1a) shield highly reactivecarbocationic intermediates from solvent; conforma-tional change may accompany substrate binding.Chain length determination (CLD) region in blue boxwith 10 conserved residues dictate product specificity(chain length) by altering the binding pocket depth.Typically, at the 2nd position of the CLD region, FDPsynthases have a Phe; GGDP and higher chain lengthsynthases have a Ser, Ala, or Thr and as it was shownGGDPS1 and GGDPS2 in P. ginseng have Ser at thisposition (amino acid 129). The active site contains dis-tinct allylic (aspartate-rich region 1) and homoallylic(aspartate-rich region 2) binding regions. seven con-served residues that compose the aspartate-rich region1 (red box) make substrate-Mg2+ binding site 1.Aspartates bind the diphosphate moieties of the allylicsubstrate through Mg2+ bridges. Aspartate-rich region 2
Eur J Plant Pathol

(brown box) which contribute in substrate-Mg2+ bind-ing site 2 includes six conserved residues bound to thehomoallylic substrate IPP.
It was revealed that the deduced amino acid ofPgGGDPS1 shares a high degree of sequence homologywith those of proteins encoded by Panax notoginseng(AIZ00598.1) (identity 99 % & similarity 99 %), Heveabrasiliensis (BAF98303.1) (identity 82 % & similarity85 %), Morus notabilis (XP_010102651.1) (identity73 % & similarity 99 %), Salvia miltiorrhiza(AEZ55680.1) (identity 72 % & similarity 99 %),
Azadirachta indica GGDPS1 (AIG15450.1) (identity73 % & similarity 98 %), Arabidopsis thaliana GGDPS1 (NP_195399.1) (identity 44 % & similarity 82 %),however PgGGDPS2 showed higher homology withAtGGDPS1 (NP_195399.1) (identity 78 % & similarity79 %) . Clustal X and the MEGA 5 Program were usedfor the construction of phylogenetic tree based onGGDPS amino acid sequences (Fig. 1b).
Distribution of GGDP synthase genes transcriptsin different organs
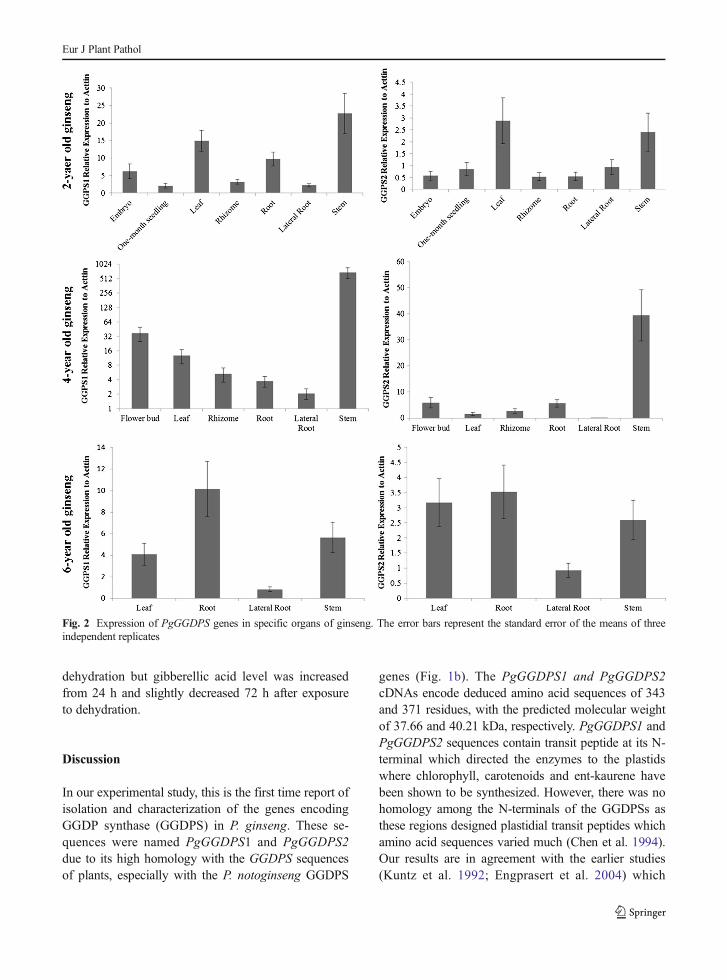
To determine the mRNA level of PgGGDPS genes atdifferent developmental stages and various organs ofginseng, we performed quantitative RT-PCR analysisof expression of PgGGDPS1 and PgGGDPS2 in em-bryo stage, 1-month old seedling and 2-year, 4-year and6-year old leaf, stem, flower, main root, lateral root,rhizome samples (Fig. 2). Among 2-year collected or-gans, relatively higher levels of PgGGDPS1 andPgGGDPS2 transcripts were observed in stem and leaf.4-year old flower and stem contain highest level ofPgGGDPS1 and PgGGDPS2 transcripts. PgGGDPS1and PgGGDPS2 transcriptome level appears to be highin root tissue when compared to other 6-year old organs.
Transcription level of PgGGDPS genes in responseto various biotic and abiotic stresses
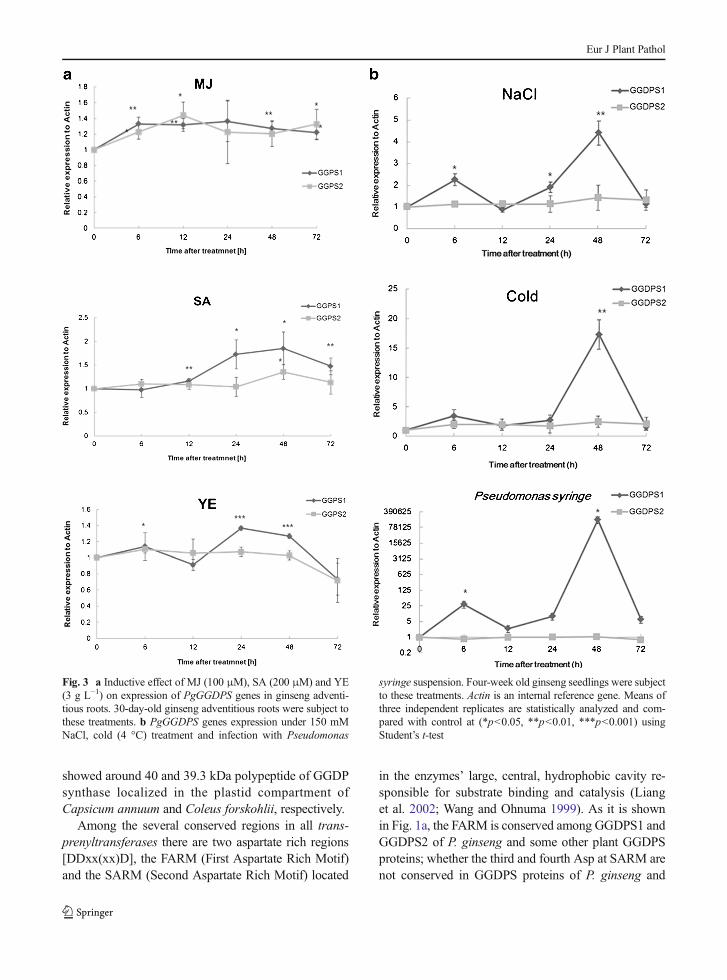
To determine the transcription of the PgGGDPS genesin response to various elicitors, we performed qRT-PCRanalysis ofGGDPS genes on ginseng adventitious rootstreated with 100 μM MJ, 200 μM SA and 3 g l−1 YE(Fig. 3a). Both PgGGDPS1 and PgGGDPS2 were up-regulated during whole treatment time of MJ elicitor. Inthe case of SA treatment, PgGGDPS1 expression in-creased after 12 h. Meanwhile, PgGGDPS2 transcrip-tion was highest at 48 h. The transcription profile of
GGDPS1 showed higher transcript level at 24 and 48 hafter YE supplementation. To recognize pathogen caninduce the transcriptome profile of PgGGDPS genes, weinfected 30 day-old ginseng seedlings withPseudomonassyringe suspension and checked the transcription patternof this gene using quantitative RT-PCR analysis. Theexpression level of PgGGDPS1 was highest at 48 h afterbacterial infection but PgGGDPS2 expression did notshow significant expression compared to control.
To elucidate the expression patterns of PgGGDPSgenes under abiotic stresses, 30 day-old ginseng seed-lings grown in vitro conditions were treated with differentabiotic stimuli (Fig. 3b). The expression patterns ofPgGGDPS genes at different time points after treatmentswere analyzed using real-time PCR. The expression pro-file of GGDPS1 increased up to 4.4-fold at 48 h of posttreatment with salt and declined at 72 h after treatmentcompared to the control. Under chilling stress (4 °C), theGGDPS1 gene expression increased to highest expres-sion level (17.3-fold) at 48 h and decreased at 72 h aftertreatment. However, both salt and cold stresses were notaffected PgGGDPS2 transcription.
Effect of dehydration treatment on water contentand mRNA levels of PgGGDPS genes
Wemeasured the water loss rate of dehydrated seedlingsof ginseng plant (Fig. 4). Ginseng slightly lost water 6 to24 h after exposing to drought. We recorded lowerrelative water content 48 h after drought treatment. Weexamined the expression level of GGDPS genes underdrought treatment (Fig. 4). During drought treatment,the transcript level of PgGGDPS1 and PgGGDPS2showed maximum accumulation at 24 h and graduallydecreased at 72 h.
Effect of dehydration stress on chlorophylland carotenoid content and endogenous hormones
To further clarify the function of GGDPS, we deter-mined the amounts of chlorophyll and carotenoids underdrought condition (Fig. 5). Carotenoids were accumu-lated 12 to 72 h after treatment. Total chlorophyll con-tents were increased at 12 to 72 h of treatment comparedwith their controls which showed similar pattern tocarotenoids accumulation.
Endogenous ABA and GA hormone levels were alsomeasured in dehydrated seedlings (Fig. 5). ABA contentsignificantly increased 48 and 72 h after exposing to
Eur J Plant Pathol
dehydration but gibberellic acid level was increasedfrom 24 h and slightly decreased 72 h after exposureto dehydration.
Discussion
In our experimental study, this is the first time report ofisolation and characterization of the genes encodingGGDP synthase (GGDPS) in P. ginseng. These se-quences were named PgGGDPS1 and PgGGDPS2due to its high homology with the GGDPS sequencesof plants, especially with the P. notoginseng GGDPS
genes (Fig. 1b). The PgGGDPS1 and PgGGDPS2cDNAs encode deduced amino acid sequences of 343and 371 residues, with the predicted molecular weightof 37.66 and 40.21 kDa, respectively. PgGGDPS1 andPgGGDPS2 sequences contain transit peptide at its N-terminal which directed the enzymes to the plastidswhere chlorophyll, carotenoids and ent-kaurene havebeen shown to be synthesized. However, there was nohomology among the N-terminals of the GGDPSs asthese regions designed plastidial transit peptides whichamino acid sequences varied much (Chen et al. 1994).Our results are in agreement with the earlier studies(Kuntz et al. 1992; Engprasert et al. 2004) which
Fig. 2 Expression of PgGGDPS genes in specific organs of ginseng. The error bars represent the standard error of the means of threeindependent replicates
Eur J Plant Pathol
showed around 40 and 39.3 kDa polypeptide of GGDPsynthase localized in the plastid compartment ofCapsicum annuum and Coleus forskohlii, respectively.
Among the several conserved regions in all trans-prenyltransferases there are two aspartate rich regions[DDxx(xx)D], the FARM (First Aspartate Rich Motif)and the SARM (Second Aspartate Rich Motif) located
in the enzymes’ large, central, hydrophobic cavity re-sponsible for substrate binding and catalysis (Lianget al. 2002; Wang and Ohnuma 1999). As it is shownin Fig. 1a, the FARM is conserved among GGDPS1 andGGDPS2 of P. ginseng and some other plant GGDPSproteins; whether the third and fourth Asp at SARM arenot conserved in GGDPS proteins of P. ginseng and
Fig. 3 a Inductive effect of MJ (100 μM), SA (200 μM) and YE(3 g L−1) on expression of PgGGDPS genes in ginseng adventi-tious roots. 30-day-old ginseng adventitious roots were subject tothese treatments. b PgGGDPS genes expression under 150 mMNaCl, cold (4 °C) treatment and infection with Pseudomonas
syringe suspension. Four-week old ginseng seedlings were subjectto these treatments. Actin is an internal reference gene. Means ofthree independent replicates are statistically analyzed and com-pared with control at (*p<0.05, **p<0.01, ***p<0.001) usingStudent’s t-test
Eur J Plant Pathol

other plant GGDPSs but PgGGDPS1 and PnGGDPSwhich share highest homology sequence have similaramino acids of Isoleucine and Aspartate in SARM andin the same way, PgGGDPS2 and AtGGDPS1 also haveLeucine and Glycine. However, further studies are need-ed to express two PgGGDPS genes in bacteria and
demonstrate activities for the encoded recombinantproteins.
The temporal expression analysis from different sam-ples in various growth stages demonstrated thatGGDPS1 and GGDPS2 mRNA were ubiquitouslyexpressed in all organs we examined though an
Fig. 4 Effect of dehydration treatment on water content andmRNA levels of PgGGDPS genes in ginseng seedlings. Four-week old ginseng seedlings were subject to this treatment. Actin
is an internal reference gene. Means of three independent repli-cates are statistically analyzed and compared with control at(*p<0.05, ** p<0.01, ***p<0.001) using Student’s t-test
Fig. 5 Time course of changes in chlorophyll and carotenoid con-tent, abscisic acid and gibberellic acid hormone levels in dehydratedginseng seedlings. Four-week old ginseng seedlings were subject to
this treatment. Actin is an internal reference gene. Means of threeindependent replicates are statistically analyzed and compared withcontrol at (*p<0.05, **p<0.01, ***p<0.001) using Student’s t-test
Eur J Plant Pathol

expression level of GGDPS2 is low. It is likely that theGGPP synthase 1 has similar functions as that ofA. thaliana GGPS-1, which locates in the chloroplastand engages in the biosynthesis of biologically impor-tant isoprenoids such as carotenoids, chlorophylls andgibberellins (Okada et al. 2000). GGDPS1 andGGDPS2 were strongly expressed in aerial parts of theplant, especially in rapidly growing tissues such as 4-year old stem and flower tissue which is likely to reflecttheir roles and specific functions in each tissue. Theresults indicate that these GGDPS1 enzyme was devel-opmentally and tissue-regulated showing higher expres-sion in 2-year old leaf than 4 and 6-year old leaf.Meanwhile, GGDPS1 expression was induced in roottissues of 6-year old ginseng. Root tissues haveetioplasts rather than chloroplasts and it can be sug-gested that GGDPS1 and GGDPS2 will be translocatedinto etioplasts in ginseng root tissues, a possible subcel-lular location for GA biosynthesis in root tissue(Tanimoto 1991). It may suggest that GGDPS1 andGGDPS2 similar to GGPS1 and GGPS3 proteins inArabidopsis (Okada et al. 2000) were translocated intochloroplasts in the upper part of the plant, and intoleukoplasts (etioplasts) in the root tissues. GGDPS wasstrongly expressed in the leaves and barely expressed inthe roots of C. forskohlii (Engprasert et al. 2004). Asmall gene family was found for GGDP synthasesencoding five isozymes and one related protein inArabidopsis (Okada et al. 2000). Among them GGRand GGDPS1 were ubiquitously expressed, whileGGDPS2, GGDPS3, and GGDPS4 were expressed spe-cifically in the flower, root, and flower, respectively.These results indicate that GGDPS is thought to useIPP, which is produced through the non-mevalonatepathway in plastids and the GGDPS1 and GGDPS2proteins in ginseng supply GGDP for the biosynthesisof chlorophyll, carotenoids, and ent-kaurene, precursorsof gibberellin, in the plastids.
Ginseng plants have evolved effective mechanismsto deal with the ever-changing environment during theirgrowth and development. In order to determine themolecular mechanisms of ginseng plant adapted to bi-otic and abiotic challenges over long years of life cycle,transcript profiling of PgGGDPS genes were studied byquantitative real time-PCR analysis. The PgGGDPS1and PgGGDPS2 genes transcription was affected by100 μM methyl jasmonate, 200 μM salicylic acid and3 g l−1 YE yeast extract in the adventitious roots ofP. ginseng as suggesting that GGDPS1 was elicitor-
responsive. It was previously proved that GGDPS wasregulated by salicylic acid (SA) and inhibited by methyljasmonate (MeJA) in Salvia miltiorrhiza (Kai et al.2010) and in S. miltiorrhiza hairy root cultures, theaccumulation of diterpenoid tanshinones was enhancedby a yeast elicitor and silver ion Ag + (Ge and Wu2004). PgGGDPS1 expressions seem to be stronglyup-regulated by salt and chilling treatment althoughlonger treatment of chilling resulted in down-regulation of its expression while transcriptome profileof GGDPS1 was induced in Pst-infected ginseng seed-lings. Our results confirmed earlier report (Hua et al.2012) that S. miltiorrhiza GGDPS can be induced byvarious factors, such as NaCl and pathogen infection.PgGGDPS1 and PgGGDPS2 responses against abioticand biotic stresses demonstrated different results, as itwas shown GGDPS1was inducible by cold and saltstress and pathogen attack.
GGDP biosynthesis is the key step in terpenoid path-way and GGDP synthase catalyzed the branch pointenzymatic reaction that is necessary for plant growth.GGDP as the precursor for the biosynthesis of the chlo-rophyll side chain can directly affect chlorophyll syn-thesis (Lichtenthaler 1987).
Plants respond to survive under water-deficit condi-tions via a series of physiological, cellular, andmolecularprocesses culminating in stress tolerance.Many drought-inducible genes with various functions have been iden-tified. According to our results, dehydrated ginseng ex-hibited higher transcript level of PgGGDPS1 andPgGGDPS2 while chlorophyll as well as carotenoidsynthesis was affected by drought. In severe water stresscondition, photosynthesis rate will be reduced and phys-iological processes will be disturbed and finally plantwill be withered and dead.
Plant hormones are involved in diverse aspects ofgrowth and developmental processes as well as variousbiotic and abiotic stress responses in plants. It wasdemonstrated that the rising level of endogenous ABA,triggering stomata closure under drought or salt stressconditions (Verslues et al. 2006). As it was shown in ourstudy, ginseng plant had lost highest amount of water48 h after dehydration treatment. Under drought stress,upregulated PgGGDPS1 and PgGGDPS2may increaseaccumulation of carotenoid precursors 24 h after onsetof drought treatment which subsequently leads to higherproduction of ABA 48 h after dehydration treatment.During that time, endogenous ABA accumulated topossibly activate stomatal closure as the physical barrier
Eur J Plant Pathol

to inhibit water loss under drought stress. Also we haveshown that the increasing severity of drought stressinduced significant increase of intracellular GA as com-pared to control. In this way, this growth promotingphytohormone may alleviated the drought stress byincreasing carbohydrate content, amino-N and total ni-trogen which is already proved (El-Meleigy et al. 1999;Aliyev et al. 2000) in peanut plant and wheat seedling,respectively.
The results of the current study support the hypoth-esis that the induction of GGDPS genes specificallyPgGGDPS1 by drought stress may affect chlorophylllevels, accumulation of carotenoids as the precursor forhigher production of ABA to possibly activate stomatalclosure as the barrier for water loss and increase ofintracellular GA. Therefore we can suggest a crucialrole for this gene in protection of ginseng plant underenvironmental abiotic stresses. Further studies onPgGGDPS genes are needed for specific localizationwithin the cell and clarify the physiological functionsof each GGDPS isozymes in ginseng.
Acknowledgments This research was supported by iPET(312064-03-1-HD040), Korea Institute of Planning and Evalua-tion for Technology in Food, Agriculture, Forestry and Fisheries,Republic of Korea.
References
Aitken, S.M., Attucci, S., Ibrahim, R. K., &Gulick, P. J. (1995). AcDNA encoding geranylgeranyl pyrophosphate synthasefrom whitelupin. Plant Physiology, 108, 837–838.
Aliyev, R. Т., Coskuncelebi, K., Beyazoglu, O., & Hacieva, S. I.(2000). Alterations in the genome of wheat seedlings grownunder drought stress and the effect of gibberellic acid on thesealterations. Review Biology, 93, 183–189.
Ament, K., Van Schie, C. C., Bouwmeester, H. J., Haring, M. A.,& Schuurink, R. C. (2006). Induction of a leaf specificgeranylgeranyl pyrophosphate synthase and emission of −4,8, 12-trimethyltrideca-1, 3, 7, 11-tetraene in tomato are de-pendent on both jasmonic acid and salicylic acid signalingpathways. Planta, 224, 1197–1208.
Artimo, P., Jonnalagedda, M., Arnold, K., Baratin, D., Csardi, G.,de Castro, E., Duvaud, S., Flegel, V., Fortier, A., Gasteiger,E., et al. (2012). ExPASy: SIB bioinformatics resource portal.Nucleic Acids Research, 40, W597–W603.
Badillo, A., Steppuhn, J., Deruere, J., Camara, B., & Kuntz, M.(1995). Structure of a functional geranylgeranyl pyrophos-phate synthase gene from Capsicum annuum. PlantMolecular Biology, 27, 425–428.
Bantignies, B., Liboz, T., & Ambid, C. (1995). Nucleotide se-quence of a Catharanthus roseus geranylgeranyl pyrophos-phate synthase gene. Plant Physiology, 110, 336.
Bartley, G. E., & Scolnik, P. A. (1995). Plant carotenoids: pig-ments for photoprotection, visual attraction, and humanhealth. Plant Cell, 7, 1027–1038.
Chen, A. J., Kroon, P. A., & Poulter, C. D. (1994). Isoprenyldiphosphate synthases protein-sequence comparisons, a phy-logenetic tree and redictions of secondary structure. ProteinScience, 3, 600–607.
El-Meleigy, E. A., Hassanein, R. A., & Abdel Kader, D. Z. (1999).Improvement of drought tolerance in Araches hypogaea L.plant by some growth substances 1. Growth and productivity.Bulletin Faculty Science Association University, 28,159–185.
Emanuelson, O., Nielsen, H., Brunak, S., & von Heijne, G. (2000).Predicting subcellular localization of proteins based on theirN-terminal amino acid sequence. Journal of MolecularBiology, 300, 1005–1016.
Engprasert, S., Taura, F., Kawamukai, M., & Shoyama, Y. (2004).Molecular cloning and functional expression ofgeranylgeranyl pyrophosphate synthase from Coleusforskohlii Briq. BMC Plant Biology, 4, 18.
Ge, X. C., & Wu, J. Y. (2004). Tanshinone production and iso-prenoid pathways in Salvia miltiorrhiza hairy roots inducedby Ag+ and yeast elicitor. Plant Science, 168, 487–491.
Gray, J. C. (1987). Control of isoprenoid biosynthesis in higherplants. Advances in Botanical Research, 14, 25–91.
Hedden, P., & Kamiya, Y. (1997). Gibberellin biosynthesis: en-zymes, genes and their regulation. Annual Review of PlantPhysiology and Plant Molecular Biology, 48, 431–460.
Hefner, J., Ketchum, F. E. B., & Croteau, R. (1998). Cloning andfunctional expression of a cDNA encoding geranylgeranyldiphosphate synthase from Taxus canadensis and assessmentof the role of this prenyltransferase in cells induced for taxolproduction. Archives of Biochemistry and Biophysics, 360,62–74.
Hua, W., Song, J., Li, C., & Wang, Z. (2012). Molecular cloningand characterization of the promoter of SmGGPPs and itsexpression pattern in Salvia miltiorrhiza. Molecular BiologyReports, 39, 5775–5783.
Kai, G., Liao, P., Zhang, T., Zhou, W., Wang, J., Xu, H., Liu, Y., &Zhang, L. (2010). Characterization, expression profiling, andfunctional identification of a gene encoding geranylgeranyldiphosphate synthase from Salvia miltiorrhiza. Biotechnologyand Bioprocess Engineering, 15, 236–245.
Kelen, M., Çubuk Demìralay, E., Demìralay, E., Sen, S., & özkan,G. (2004). Separation of Abscisic acid, Indole-3-Acetic acid,Gibberellic acid in 99 R (Vitisberlandierix Vitisrupestris)and rose oil (Rosa damascene Mill.) by Reversed PhaseLiquid Chromatography. Turkish Journal of Chemistry,28, 603–610.
Kim, Y. J., Jang, M. G., Noh, H. Y., Lee, H. J., Sukweenadhi, J.,Kim, J. H., Kim, S. Y., Kwon, W. S., & Yang, D. C. (2014).Molecular characterization of two glutathione peroxidasegenes of Panax ginseng and their expression analysis againstenvironmental stresses. Gene, 535, 33–41.
Kirk, J. T. O., & Allen, R. L. (1965). Dependence of chloroplastpigment synthesis on protein synthesis: Effect of actidione.Biochemical Biophysical Research Communications, 21,523–530.
Eur J Plant Pathol

Kleinig, H. (1989). The role of plastids in isoprenoid biosynthesis.Annual Review of Plant Physiology and Plant MolecularBiology, 40, 39–59.
Kuntz, M., Romer, S., Suire, C., Hugueney, P., Weil, J. H.,Schantz, R., & Camara, B. (1992). Identification of acDNA for the plastid-located geranylgeranyl pyrophosphatesynthase from Capsicum annuum: correlative increase inenzyme activity and transcript level during fruit ripening.Plant Journal, 2, 25–34.
Laferriere, A., & Beyer, P. (1991). Purification of geranylgeranyldiphosphate synthase from Sinapis alba etioplasts.Biochimica et Biophysica Acta, 1077, 167–172.
Lee, C. H., & Kim, J. H. (2014). A review on the medicinalpotentials of ginseng and ginsenosides on cardiovasculardiseases. Journal of Ginseng Research, 38, 161–166.
Liang, P. H., Ko, T. P., & Wang, A. H. J. (2002). Structure,mechanism and function of prenyltransferases. EuropeanJournal of Biochemistry, 269, 3339–3354.
Lichtenthaler, H. K. (1987). Chlorophylls and carotenoids: pig-ments of photosynthetic biomembranes. Methods inEnzymology, 148, 350–82.
Livak, K. J., & Schmittgen, T. D. (2001). Analysis of relative geneexpression data using real-time quantitative PCR and the2(−Delta Delta CT) method. Methods, 25, 402–408.
Morris, P. C., Kumar, A., Bowles, D. J., & Cuming, A. C. (1990).Osmotic stress and abscisic acid regulate the expression ofthe Em gene of wheat. European Journal of Biochemistry,190, 625–630.
Murashige, T., & Skoog, F. (1962). A revised medium for rapidgrowth and bio assay with tobacco tissue cultures. PlantPhysiology, 15, 473–499.
Oh, S. K., Kim, I. J., Shin, D. H., Yang, J., Kang, H., & Han, K. H.(2000). Cloning, characterization, and heterologous expres-sion of a functional geranylgeranyl pyrophosphate synthasefrom sunflower (Helianthus annuus L.). Journal of PlantPhysiology, 157, 535–542.
Okada, K., Saito, T., Nakagawa, T., Kawamukai, M., & Kamiya,Y. (2000). Five geranylgeranyldiphosphate synthasesexpressed in different organs are localized into three subcel-lular compartments in Arabidopsis. Plant Physiology, 122,1045–1056.
Scolnik, P. A., & Bartley, G. E. (1994). Nucleotide sequence of anArabidopsis cDNA for geranylgeranyl pyrophosphate syn-thase. Plant Physiology, 104, 1469–1470.
Scolnik, P. A., & Bartley, G. E. (1995). Nucleotide sequence of aputative geranylgeranyl pyrophosphate synthase (GenBankaccession no. L40577) from Arabidopsis (PGR 95–018).Plant Physiology, 108, 1343.
Scolnik, P. A., & Bartley, G. E. (1996). Two more members ofArabidopsis geranylgeranyl pyrophosphate synthase family(PGR 96–014). Plant Physiology, 110, 1435.
Sitthithaworn, W., Kojima, N., Viroonchatapan, E., Suh, D. Y.,Iwanami, N., Hayashi, T., Noji, M., Saito, K., Niwa, Y., &Sankawa, U. (2001) Geranylgeranyl diphosphate synthasefrom Scoparia dulcis and Croton sublyratus. Plastid localiza-tion and conversion to a farnesyl diphosphate synthase bymutagenesis. Chem Pharm Bull, 49, 197–292.
Song, M. Y., Kim, B. S., & Kim, H. (2014). Influence of Panaxginseng on obesity and gut microbiota in obese middle-agedKorean women. Journal of Ginseng Research, 38, 106–115.
Tanimoto, E. (1991). Gibberellin requirement for the normalgrowth of roots. In N. Takahashi, B. O. Phinney, & J.MacMillan (Eds.), Gibberellins (pp. 229–240). New York:Springer.
Verslues, P. E., Agarwal, M., Katiyar-Agarwal, S., Zhu, J., & Zhu,J. K. (2006). Methods and concepts in quantifying resistanceto drought, salt and freezing, abiotic stresses that affect plantwater status. Plant Journal, 45, 523–539.
Wang, K., & Ohnuma, S. I. (1999). Chain-length determinationmechanism of isoprenyl diphosphate synthases and implica-tions for molecular evolution. Trends Biochemical Science,24, 445–451.
Zhu, X. F., Suzuki, K., Okada, K., Tanaka, K., Nakagawa, T.,Kawamukai, M., & Matsuda, H. (1997a). Cloning and func-tional expression of a novel geranylgeranyl pyrophosphatesynthase gene from Arabidopsis thaliana in Escherichia coli.Plant Cell Physiology, 38, 357–361.
Zhu, X. F., Suzuki, K., Saito, T., Okada, K., Tanaka, K.,Nakagawa, T., Matsuda, H., & Kawamukai, M. (1997b).Geranylgeranyl pyrophosphatesynthase encoded by the new-ly isolated gene GGPS6 from Arabidopsis thaliana is local-ized in mitochondria. Plant Molecular Biology, 35, 331–341.
Eur J Plant Pathol
Copyright © 2022 FDOKUMEN




















