
Involvement of proteinase activated receptor-2in the vascular response to sphingosine 1-phosphate
15
Clinical Science (2014) 126, 545–556 (Printed in Great Britain) doi: 10.1042/CS20130272 Involvement of proteinase activated receptor-2 in the vascular response to sphingosine 1-phosphate Fiorentina ROVIEZZO ∗ , Antonella DE ANGELIS†, Luana DE GRUTTOLA ∗ , Antonio BERTOLINO ∗ , Nikol SULLO†, Vincenzo BRANCALEONE‡, Mariarosaria BUCCI ∗ , Raffaele DE PALMA§, Konrad URBANEK†, Bruno D’AGOSTINO†, Angela IANARO ∗ , Raffaella SORRENTINO ∗ and Giuseppe CIRINO ∗ ∗ Dipartimento di Farmacia, Universit` a di Napoli Federico II, Napoli, Italy †Dipartimento di Medicina Sperimentale, Sezione di Farmacologia L. Donatelli, Seconda Universit` a degli Studi di Napoli, Napoli, Italy ‡Dipartimento di Chimica, Universit` a degli Studi della Basilicata, Pontenza, Italy §Dipartimento di Internistica Clinica e Sperimentale, Seconda Universit` a degli Studi di Napoli, Napoli, Italy Abstract S1P (sphingosine 1-phosphate) represents one of the key latest additions to the list of vasoactive substances that modulate vascular tone. PAR-2 (proteinase activated receptor-2) has been shown to be involved in cardiovascular function. In the present study, we investigated the involvement of PAR-2 in S1P-induced effect on vascular tone. The present study has been performed by using isolated mouse aortas. Both S1P and PAR-2 agonists induced endothelium-dependent vasorelaxation. L-NAME (N G -nitro-L-arginine methyl ester) and wortmannin abrogated the S1P-induced vasorelaxatioin, while significantly inhibiting the PAR-2-mediated effect. Either ENMD1068, a PAR-2 antagonist, or gabexate, a serine protease inhibitor, significantly inhibited S1P-induced vasorelaxation. Aortic tissues harvested from mice overexpressing PAR-2 displayed a significant increase in vascular response to S1P as opposed to PAR-2-null mice. Immunoprecipitation and immunofluorescence studies demonstrated that S1P 1 interacted with PAR-2 and co-localized with PAR-2 on the vascular endothelial surface. Furthermore, S1P administration to vascular tissues triggered PAR-2 mobilization from the plasma membrane to the perinuclear area; S1P-induced translocation of PAR-2 was abrogated when aortic rings were pre-treated with ENMD1068 or when caveolae dysfunction occurred. Similarly, experiments performed in cultured endothelial cells (human umbilical vein endothelial cells) showed a co-localization of S1P 1 and PAR2, as well as the ability of S1P to induce PAR-2 trafficking. Our results suggest that S1P induces endothelium-dependent vasorelaxation mainly through S1P 1 and involves PAR-2 transactivation. Key words: aorta, nitric oxide, protease-activated receptor-2 (PAR-2), sphingosine 1-phosphate (S1P), vascular tone INTRODUCTION S1P (sphingosine 1-phosphate) is a phosphorylated product of sphingosine, the core structure of a lipid class termed sphingo- lipids [1–3]. S1P has been found to exert a diverse set of physiolo- gical and pathophysiological responses in mammalian tissues through the activation of heterotrimeric G-proteins [4,5]. So far, five independent S1P receptor subtypes have been identified and termed S1P 1 –S1P 5 . The isoforms S1P 1 , S1P 2 and S1P 3 are mainly expressed in blood vessels, vascular endothelial cells and smooth muscle cells [6–8]. S1P and S1P receptors have critical effects on Abbreviations: DAPI, 4 ′ ,6-diamidino-2-phenylindole; eNOS, endothelial nitric oxide synthase; HUVEC, human umbilical vein endothelial cell; IL, interleukin; L-NAME, N G -nitro-L-arginine methyl ester; LPS, lipopolysaccharide; NOS, nitric oxide synthase; PAR-2, proteinase activated receptor-2; PE, phenylephrine; PI3K, phosphoinositide 3-kinase; PTX, pertussis toxin; qPCR, quantitative real-time PCR; S1P , sphingosine 1-phosphate; vWF, von Willebrand factor. Correspondence: Professor Giuseppe Cirino (email [email protected]). morphogenesis and embryonic development of the vasculature as well as of the heart [2]. S1P has direct vasoactive effect both on isolated vessel and in vivo [7,9]. eNOS (endothelial nitric oxide synthase) activation is a common feature of the S1P response in vascular endothelial cells [9,10]. Indeed, S1P induces vascular relaxation by activating eNOS via a pertussis toxin-sensitive G protein-coupled receptor pathway [9–13]. In addition, S1P pro- tects endothelial cells from apoptosis or programmed cell death and promotes endothelial cell survival, proliferation and mi- gration by activating the PI3K (phosphoinositide 3-kinase)/Akt pathway [8–11]. The ability of S1P to transactivate heterologous www.clinsci.org 545 Clinical Science www.clinsci.org
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Involvement of proteinase activated receptor-2in the vascular response to sphingosine 1-phosphate

Clinical Science (2014) 126, 545–556 (Printed in Great Britain) doi: 10.1042/CS20130272
Involvement of proteinase activated receptor-2in the vascular response to sphingosine1-phosphateFiorentina ROVIEZZO∗, Antonella DE ANGELIS†, Luana DE GRUTTOLA∗, Antonio BERTOLINO∗, Nikol SULLO†,
Vincenzo BRANCALEONE‡, Mariarosaria BUCCI∗, Raffaele DE PALMA§, Konrad URBANEK†,
Bruno D’AGOSTINO†, Angela IANARO∗, Raffaella SORRENTINO∗ and Giuseppe CIRINO∗
∗Dipartimento di Farmacia, Universita di Napoli Federico II, Napoli, Italy
†Dipartimento di Medicina Sperimentale, Sezione di Farmacologia L. Donatelli, Seconda Universita degli Studi di Napoli, Napoli, Italy
‡Dipartimento di Chimica, Universita degli Studi della Basilicata, Pontenza, Italy
§Dipartimento di Internistica Clinica e Sperimentale, Seconda Universita degli Studi di Napoli, Napoli, Italy
Abstract
S1P (sphingosine 1-phosphate) represents one of the key latest additions to the list of vasoactive substances that
modulate vascular tone. PAR-2 (proteinase activated receptor-2) has been shown to be involved in cardiovascular
function. In the present study, we investigated the involvement of PAR-2 in S1P-induced effect on vascular tone. The
present study has been performed by using isolated mouse aortas. Both S1P and PAR-2 agonists induced
endothelium-dependent vasorelaxation. L-NAME (NG-nitro-L-arginine methyl ester) and wortmannin abrogated the
S1P-induced vasorelaxatioin, while significantly inhibiting the PAR-2-mediated effect. Either ENMD1068, a PAR-2
antagonist, or gabexate, a serine protease inhibitor, significantly inhibited S1P-induced vasorelaxation. Aortic
tissues harvested from mice overexpressing PAR-2 displayed a significant increase in vascular response to S1P as
opposed to PAR-2-null mice. Immunoprecipitation and immunofluorescence studies demonstrated that S1P1
interacted with PAR-2 and co-localized with PAR-2 on the vascular endothelial surface. Furthermore, S1P
administration to vascular tissues triggered PAR-2 mobilization from the plasma membrane to the perinuclear area;
S1P-induced translocation of PAR-2 was abrogated when aortic rings were pre-treated with ENMD1068 or when
caveolae dysfunction occurred. Similarly, experiments performed in cultured endothelial cells (human umbilical vein
endothelial cells) showed a co-localization of S1P1 and PAR2, as well as the ability of S1P to induce PAR-2
trafficking. Our results suggest that S1P induces endothelium-dependent vasorelaxation mainly through S1P1 and
involves PAR-2 transactivation.
Key words: aorta, nitric oxide, protease-activated receptor-2 (PAR-2), sphingosine 1-phosphate (S1P), vascular tone
INTRODUCTION
S1P (sphingosine 1-phosphate) is a phosphorylated product of
sphingosine, the core structure of a lipid class termed sphingo-
lipids [1–3]. S1P has been found to exert a diverse set of physiolo-
gical and pathophysiological responses in mammalian tissues
through the activation of heterotrimeric G-proteins [4,5]. So far,
five independent S1P receptor subtypes have been identified and
termed S1P1–S1P5. The isoforms S1P1, S1P2 and S1P3 are mainly
expressed in blood vessels, vascular endothelial cells and smooth
muscle cells [6–8]. S1P and S1P receptors have critical effects on
Abbreviations: DAPI, 4′,6-diamidino-2-phenylindole; eNOS, endothelial nitric oxide synthase; HUVEC, human umbilical vein endothelial cell; IL, interleukin; L-NAME, NG -nitro-L-arginine
methyl ester; LPS, lipopolysaccharide; NOS, nitric oxide synthase; PAR-2, proteinase activated receptor-2; PE, phenylephrine; PI3K, phosphoinositide 3-kinase; PTX, pertussis toxin;
qPCR, quantitative real-time PCR; S1P, sphingosine 1-phosphate; vWF, von Willebrand factor.
Correspondence: Professor Giuseppe Cirino (email [email protected]).
morphogenesis and embryonic development of the vasculature as
well as of the heart [2]. S1P has direct vasoactive effect both on
isolated vessel and in vivo [7,9]. eNOS (endothelial nitric oxide
synthase) activation is a common feature of the S1P response in
vascular endothelial cells [9,10]. Indeed, S1P induces vascular
relaxation by activating eNOS via a pertussis toxin-sensitive G
protein-coupled receptor pathway [9–13]. In addition, S1P pro-
tects endothelial cells from apoptosis or programmed cell death
and promotes endothelial cell survival, proliferation and mi-
gration by activating the PI3K (phosphoinositide 3-kinase)/Akt
pathway [8–11]. The ability of S1P to transactivate heterologous
www.clinsci.org 545
Clin
ical
Sci
ence
ww
w.c
linsc
i.org

F. Roviezzo and others
receptors triggered by vascular endothelial growth factor, platelet
derived growth factor, epidermal growth factor and a membrane-
bound protease named MT-SP1 (membrane-type serine protease
1) has also been demonstrated [14–19].
PARs (proteinase-activated receptors) are a family of seven-
transmembrane G-protein-coupled receptors [20]. PARs are ac-
tivated through an unique mechanism of proteolysis that involves
cleavage at a specific site, different for each receptor, unmasking
a new N-terminal that acts as a tethered ligand [20,21]. At the
present stage, four subtypes of PARs have been characterized:
PAR-1, PAR-3 and PAR-4 selectively activated by thrombin and
PAR-2 that is activated by trypsin, tryptase, tissue factor, Factor
VII and Factor X. PAR-2 has been shown to be involved in cardi-
ovascular function [20]. Functional studies have shown that activ-
ation of the receptor by using a small agonist peptide derived from
the tethered ligand sequence (PAR-2AP) causes an endothelium-
dependent vasodilatation that involves NO and prostanoids [22–
24]. Subsequent studies have clearly shown that PAR-2 is ex-
pressed both in the human endothelial and smooth muscle cells
[25]. Systemic administration of PAR-2AP in both rats and mice
causes hypotension [23–25]. Interestingly, in humans in vivo
PAR-2AP also causes vasodilatation [26]. Unlike PAR-1, PAR-
2 expression in endothelial cells is up-regulated by inflammat-
ory stimuli such as TNFα (tumour necrosis factor α), bac-
terial endotoxin [LPS (lipopolysaccharide)] and IL (interleukin)-
1α [26–30]. Likewise, LPS administration to rats increases
PAR-2 expression on both endothelium and smooth muscle
cells, which translates into an increased relaxation to PAR–2AP
[26,27].
Several lines of evidence suggest a tight link between pro-
teases and sphingolipids [31–34]. This interaction has been
widely investigated for PAR-1, where it has been demonstrated a
specific interaction between PAR-1 and S1P3 [35]. Since PAR-2 is
involved in vascular homoeostasis [27–30], we have investigated
if PAR-2 also interacts with S1P.
MATERIALS AND METHODS
Reagents
SIP and ENMD-1068 were purchased from Enzo Life Sciences;
gabexate mesilate, wortmannin, trypsin, DAPI, PTX (pertussis
toxin) and a rabbit polyclonal anti-vWF (von Willebrand factor)
antibody were purchased from Sigma; W146 was purchased
from Avanti Polar Lipids; the antibody to PAR-2 (SAM-11) was
purchased from Santa Cruz Biotechnology; the anti-(phospho-
eNOS) polyclonal antibody and rabbit anti-eNOS antibody were
purchased from Calbiochem; and the antibody to S1P1 was pur-
chased from Exalpha Biologicals. All salts used for Krebs solu-
tion preparation were purchased from Carlo Erba Reagenti. The
PAR-2 agonist peptide (SLIGRL-NH2) was a gift from Professor
V. Santagada and G. Caliendo (Dipartimento di Chimica Farma-
ceutica e Tossicologica, Universita di Napoli “Federico II”, Na-
poli, Italy); normal donkey serum and donkey anti-(mouse IgG)
and anti-(rabbit IgG) secondary antibodies were from Jackson
ImmunoResearch; fluorescein-conjugated Griffonia simplicifolia
lectin I was purchased from Vector Laboratories (CA, U.S.A.)
Experimental animals
The study was carried out in 6–8-week-old male CD1 mice
(Charles River breeding Laboratories); mice overexpressing
PAR-2 (PAR2tg) and PAR-2 null mice (PAR2ko) were provided
by Professor Lungarella (Faculty of Medicine, University of Si-
ena, Siena, Italy). Detailed descriptions of the mice can be found
in the Supplementary Materials and methods section as http://
www.clinsci.org/cs/126/cs1260545add.htm). Mice were housed
in a controlled environment and provided with standard rodent
chow and water.
Animal care was in compliance with Italian regulations on
protection of animals used for experimental and other scientific
purposes (D.M. 116192) as well as with the EEC regulations (O.J.
of E.C. L 358/1 12/18/1986).
Vasomotor reactivity studies
Mice were anaesthetized with isofluorane (5 %) followed by ex-
sanguination. The adequacy of anaesthesia was monitored by lack
of twitch after foot pinch. Aortas were carefully removed and
dissected free from connective tissue. Aortic rings were used for
functional studies in an isolated organ bath filled with oxygenated
Krebs’ solution at 37 ◦C, linked to isometric force transducers
(Fort 10; World Precision Instrument). Concentration-dependent
response curves to S1P (1×10− 8–3×10− 5 mol/l) were cumulat-
ively obtained following submaximal contractions in response to
PE (phenylephrine) in the presence or absence of endothelium.
S1P-induced vasorelaxation was assessed in the presence of (i)
PTX (that uncouples the receptor from its Gi-protein; 1 mg/l);
(ii) L-NAME (NG-nitro-L-arginine methyl ester) [aNOS (nitric
oxide synthase) inhibitor; 10− 4 mol/l] and wortmannin (PI3K
inhibitor; 10− 7 mol/l); (iii) a specific S1P1 receptor antagonist
(W146; 10 µmol/l); and (iv) a monoclonal anti-PAR-2 primary
antibody (SAM-11) [36] at a 1:1000 dilution. This antibody is
directed to N-terminal amino acids 37–50, where the extracellular
cleavage site of PAR-2 is located.
Preliminary experiments were performed in order to define
the optimal concentrations of inhibitor used. The inhibitor con-
centrations used do not affect PE-induced contraction.
PAR-2 activation was evaluated by using: (i) trypsin, that
causes proteolytic activation of PAR-2; and (ii) PAR-2AP
(SLIGRL-NH2), the N-terminal receptor sequence, that acts as a
‘tethered’ ligand. PAR-2 activation was pharmacologically mod-
ulated by using specific inhibitors such as: (i) a serine protease
inhibitor, gabexate mesilate (10 µmol/l); (ii) a PAR-2 antagonist
(ENMD-1068; 100 µmol/l) [36]; (iii) wortmannin (10− 7 mol/l);
and (iv) SAM-11 (1:1000 dilution) [36]. In another set of exper-
iments, we tested the effect of: (i) gabexate mesilate or ENMD-
1068 on S1P-induced vasorelaxation; and (ii) W146 or PTX on
PAR-2AP-induced vasorelaxation.
Western blot analysis
Western blot analysis for eNOS and phospho-eNOS was per-
formed on aortic tissues incubated with S1P (10− 6 mol/l) or
546 C© The Authors Journal compilation C© 2014 Biochemical Society

S1P/PAR-2 cross-talk and vascular function
Figure 1 S1P and PAR-2AP induce endothelium-mediated vessel relaxation
(A) Cumulative administration of S1P on PE-contracted aortic rings causes a concentration- and endothelium-dependentrelaxation. (B) Quantitative PCR demonstrates the presence of S1P1, S1P2 and S1P3 receptors in mouse aorta. (C)Incubation with both W146 (10 µmol/l), an S1P1 antagonist, and (D) PTX (1 mg/l), which specifically uncouples S1P1 fromits Gi -protein, significantly inhibits S1P induced vasorelaxation. (E) Cumulative administration of PAR-2AP (SLIGRL-NH2), apeptide that mimics the tethered ligand exposed following PAR-2 proteolytic activation, induces endothelium- dependentvasorelaxation. (F) Incubation of aortic tissues with PTX, prior to PAR-2AP challenge, does not modify the relaxation. Dataare expressed as means +
− S.E.M., n = 6/group. ∗∗∗P < 0.001 compared with its vehicle.
PAR-2AP (10− 6 mol/l). In another set of experiments aortic tis-
sues were pre-treated with wortmannin (10− 7 mol/l). Details are
described in the Supplementary Materials and methods section.
Immunoprecipitation
Aortic tissues or HUVECs (human umbilical vein endothelial
cells) were used for S1P1 immunoprecipitation and PAR-2 co-
precipitation was monitored. The isotype control antibody was
used as negative control. Details are described in the Supplement-
ary Materials and methods section.
Immunofluorescence
Aortic rings or HUVECs untreated or stimulated with S1P
(1 µmol/l) or trypsin (3 units/ml) were rinsed with PBS and fixed
with phosphate-buffered formalin. In another set of experiments
aortic rings were incubated with ENMD-1068 (100 µmol/l;
10 min) or filipin (1 mg/l; 30 min at 37) prior to adding S1P
www.clinsci.org 547

F. Roviezzo and others
Figure 2 S1P and PAR-2AP induce vasorelaxation by activating eNOS via the PI3K/Akt pathway
(A) S1P-induced relaxation is abrogated by both L-NAME (100 µmol/l) and wortmannin (10− 7 mol/l). (B) L-NAME or wort-mannin significantly inhibit PAR-2AP-induced relaxation. All data are shown as means +
− S.E.M., n = 6/group. ∗∗∗P < 0.001
compared with its vehicle. (C) Representative Western blots of eNOS or Akt phosphorylation in aortic rings incubated withboth S1P (10− 6 mol/l) and PAR-2AP (10− 6 mol/l) without or with wortmannin (W; 10− 7mol/l). Histograms indicate therelative densitometry compared with total eNOS protein. Data are expressed as means +
− S.E.M., n = 6/group. ∗∗P < 0.01
compared with vehicle. V, vehicle.
(1 µmol/l). Filipin treatment disrupted caveolar structures, alter-
ing the physical distribution of the cholesterol in the membrane
by forming filipin–cholesterol complexes [37]. Tissues were fixed
in formalin, embedded in paraffin and cut into sections of 4 µm in
thickness. Details are described in the Supplementary Materials
and methods section.
mRNA quantification by qPCR (quantitative
real-time PCR)
S1P receptor mRNA was determined by qPCR. Details are de-
scribed in the Supplementary Materials and methods section.
Calculations and statistical analysis
All results are expressed as means +− S.E.M. and n indicates the
number of animals from which tissues were harvested. Relaxa-
tions (expressed as percentage of contraction) were determined
for each individual concentration–response curve by non-linear
regression analysis. The concentration–response curves of the
different groups were compared by ANOVA for repeated meas-
urements followed by Bonferroni correction. When appropriate,
unpaired Student’s t test was used. A value of P < 0.05 was con-
sidered significant.
RESULTS
S1P and PAR-2AP induce endothelium-dependent
vasorelaxation
Cumulative administration of S1P to aortic rings caused a
concentration-dependent vasorelaxation. The S1P effect was
endothelium-dependent since its removal abrogates S1P-induced
548 C© The Authors Journal compilation C© 2014 Biochemical Society

S1P/PAR-2 cross-talk and vascular function
Figure 3 S1P-induced vasorelaxation involves PAR-2 activation
(A) Cumulative administration of trypsin, that activates PAR-2 through a proteolytic mechanism, induces aortic relaxation.Trypsin-induced vasorelaxation is significantly inhibited by previous incubation with gabexate (10 µmol/l), a serine proteaseinhibitor, or ENMD-1068 (100 µmol/l), a specific PAR-2 antagonist. (B) PAR-2AP-induced relaxation is significantly reducedby incubation with ENMD-1068, but is not affected by gabexate. (C) Both ENMD-1068 and gabexate inhibit S1P-inducedvasorelaxation. (D) ENMD-1068 significantly inhibits SEW-2871 (an S1P1 agonist)-induced relaxation. Data are expressedas means +
− S.E.M., n = 6/group. ∗∗P < 0.01 and ∗∗∗
P < 0.001 compared with its vehicle. U, units.
vasodilatation (Figure 1A). Mouse aortas mainly express three
S1P receptor subtypes, namely S1P1, S1P2 and S1P3 with S1P1
being expressed most (Figure 1B). The main role of S1P1 has
been confirmed by functional experiments. Indeed, incubation
with W146, a specific antagonist of S1P1, abrogated S1P-induced
vasorelaxation (Figure 1C). S1P-mediated vasorelaxation was
PTX-sensitive as well (Figure 1D).
Similarly, PAR-2AP caused an endothelium-dependent re-
laxation (Figure 1E). Unlike S1P, incubation with PTX did not
modify PAR-2AP-induced vasodilatation (Figure 1F). These data
imply that both signals originate from independently engaged re-
ceptors.
S1P and PAR-2 induce relaxation by activating
eNOS
Endothelium removal abrogates both S1P- and PAR-2AP-
induced vascular relaxation, implying a role for endothelium-
derived mediators. In order to evaluate the contribute of NO,
we used L-NAME. The NOS inhibitor L-NAME abrogated S1P-
induced relaxation (Figure 2A) and significantly inhibited PAR-
2AP-induced vasorelaxation (Figure 2B). In order to gain fur-
ther insights into the NO pathway, we evaluated the effect of
wortmannin, an inhibitor of PI3K activity. S1P-induced vasore-
laxation was abrogated by wortmannin (Figure 2A). Similarly,
wortmannin caused a significant inhibition of PAR-2AP-induced
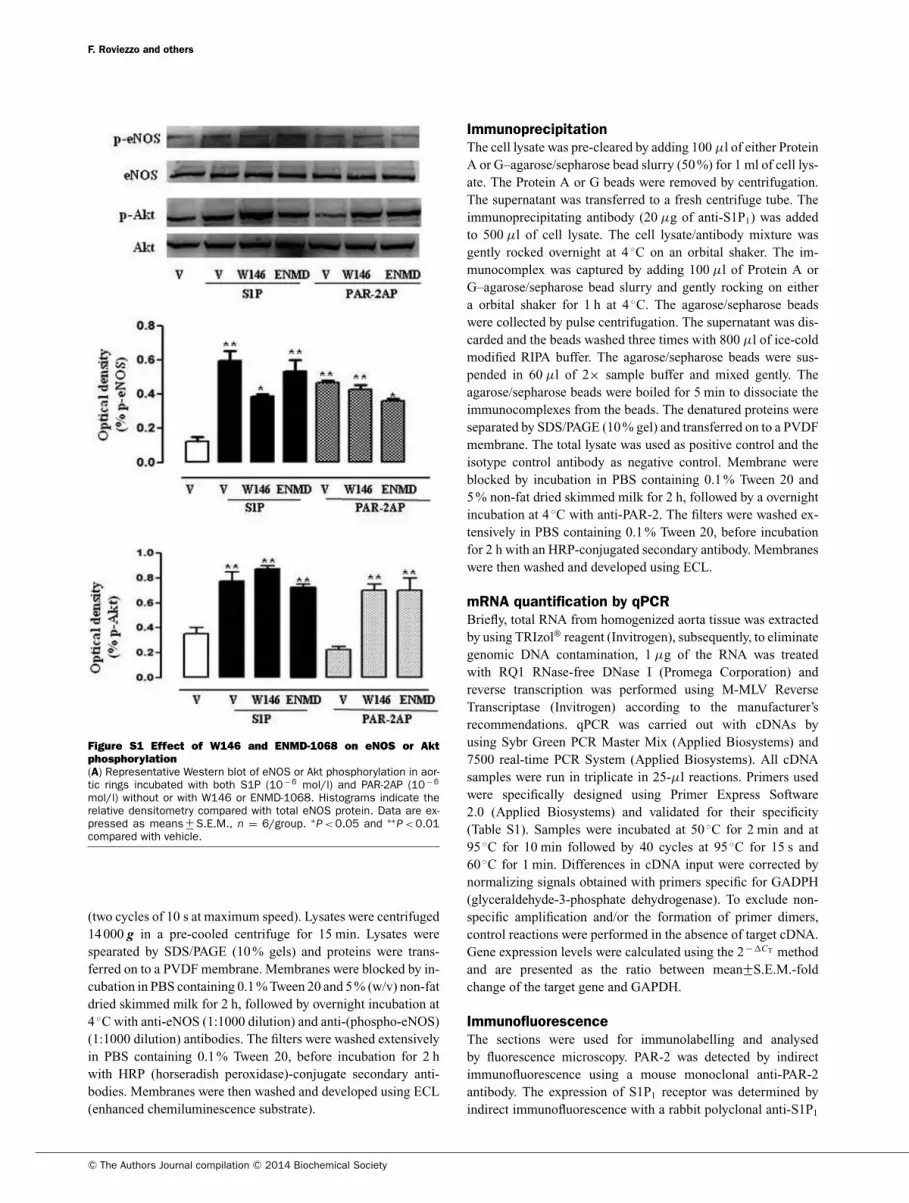
vasorelaxation (Figure 2B). Western blot analysis showed that
both S1P and PAR-2AP increased eNOS phosphorylation, which
was reversed by wortmannin pre-treatment (Figure 2C).
Aortic tissue challenged with S1P displayed a significant Akt
phosphorylation as well. Conversely, PAR-2AP did not.
Gabexate and ENMD-1068 inhibit S1P1- mediated
vasorelaxation
Next, we performed experiments by using specific antagonists of
PAR-2 and S1P1 in order to evaluate an interaction between these
receptors. PAR-2 activation was evaluated by using: (i) trypsin,
which causes proteolytic activation of PAR-2; or (ii) PAR-2AP, the
N-terminal receptor sequence, which acts as a ‘tethered’ ligand.
www.clinsci.org 549
F. Roviezzo and others
Cumulative administration of trypsin induced a concentration-
dependent vasorelaxation (Figure 3A). Both gabexate, a ser-
ine protease inhibitor, and ENMD-1068, a selective antagonist
of PAR-2, significantly inhibited trypsin-induced vasorelaxation
(Figure 3A). Conversely, ENMD-1068 but not gabexate inhibited
PAR-2AP-induced vasorelaxation (Figure 3B).
Incubation of aortic rings with gabexate or ENMD-1068 signi-
ficantly inhibited S1P-induced relaxation too (Figure 3C). These
data suggest that S1P may promote PAR-2 activation. This hypo-
thesis was supported further by the finding that ENMD-1068 sig-
nificantly inhibited SEW-2871 (S1P1 selective agonist)-induced
relaxation (Figure 3D).
In another set of experiments, we also tested the effects of
these inhibitors on S1P- and PAR-2-induced signalling. In par-
ticular, we performed a Western blot analysis study on aortic
tissues exposed in vitro to S1P or PAR-2AP, in the presence or
absence of antagonists, e.g. W146 or ENMD-1068 respectively.
These experiments confirmed the presence of two signalling path-
ways downstream, since neither ENMD-1068 blocked S1P sig-
nalling nor W146 blocked PAR-2 (Supplementary Figure S1 at
http://www.clinsci.org/cs/126/cs1260545add.htm).
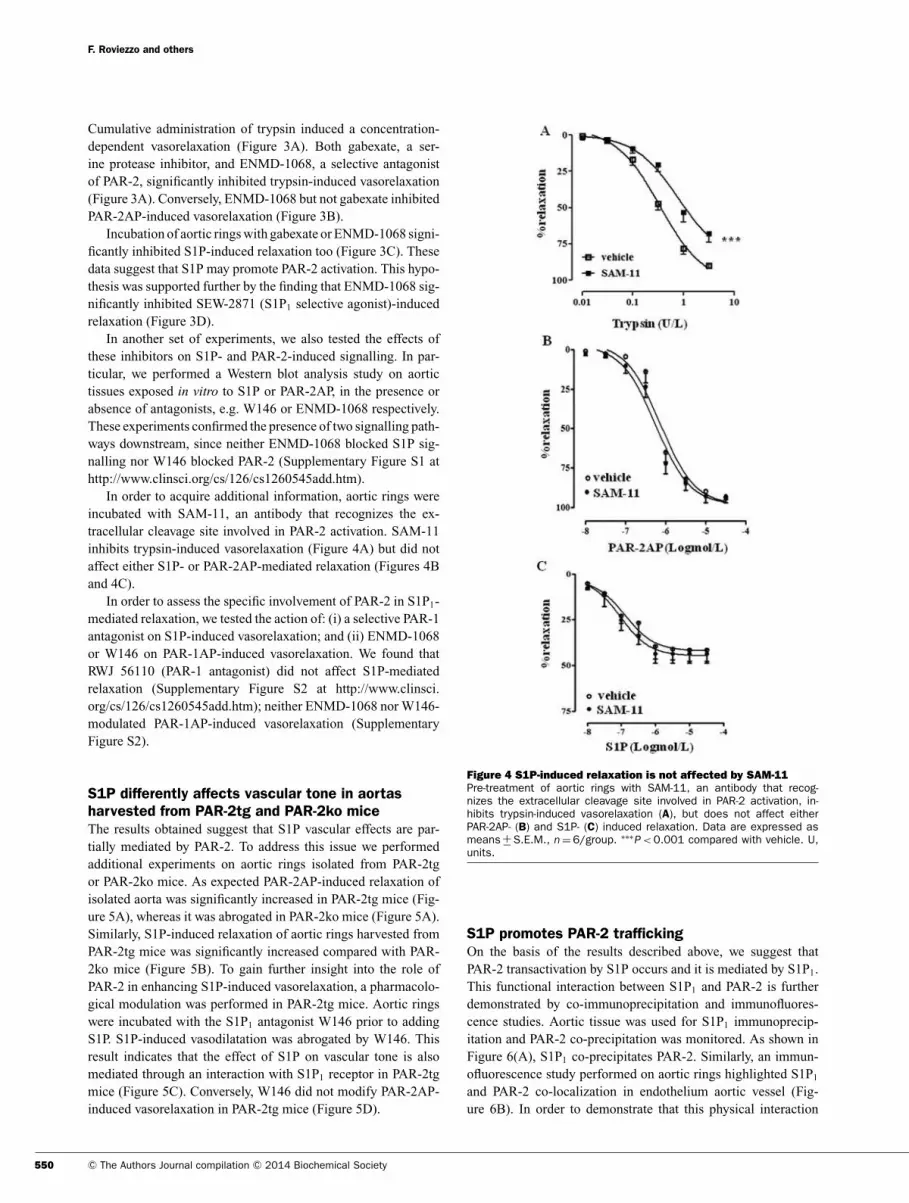
In order to acquire additional information, aortic rings were
incubated with SAM-11, an antibody that recognizes the ex-
tracellular cleavage site involved in PAR-2 activation. SAM-11
inhibits trypsin-induced vasorelaxation (Figure 4A) but did not
affect either S1P- or PAR-2AP-mediated relaxation (Figures 4B
and 4C).
In order to assess the specific involvement of PAR-2 in S1P1-
mediated relaxation, we tested the action of: (i) a selective PAR-1
antagonist on S1P-induced vasorelaxation; and (ii) ENMD-1068
or W146 on PAR-1AP-induced vasorelaxation. We found that
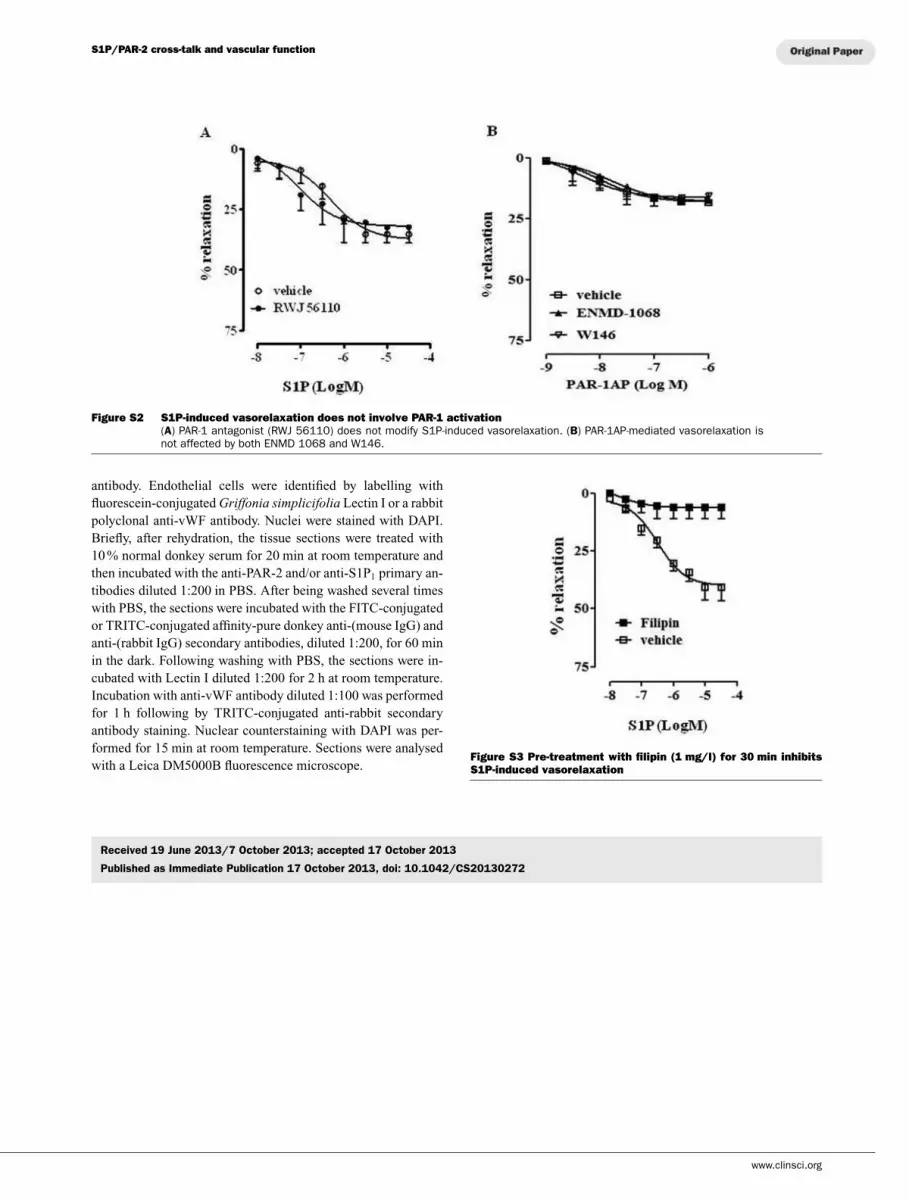
RWJ 56110 (PAR-1 antagonist) did not affect S1P-mediated
relaxation (Supplementary Figure S2 at http://www.clinsci.
org/cs/126/cs1260545add.htm); neither ENMD-1068 nor W146-
modulated PAR-1AP-induced vasorelaxation (Supplementary
Figure S2).
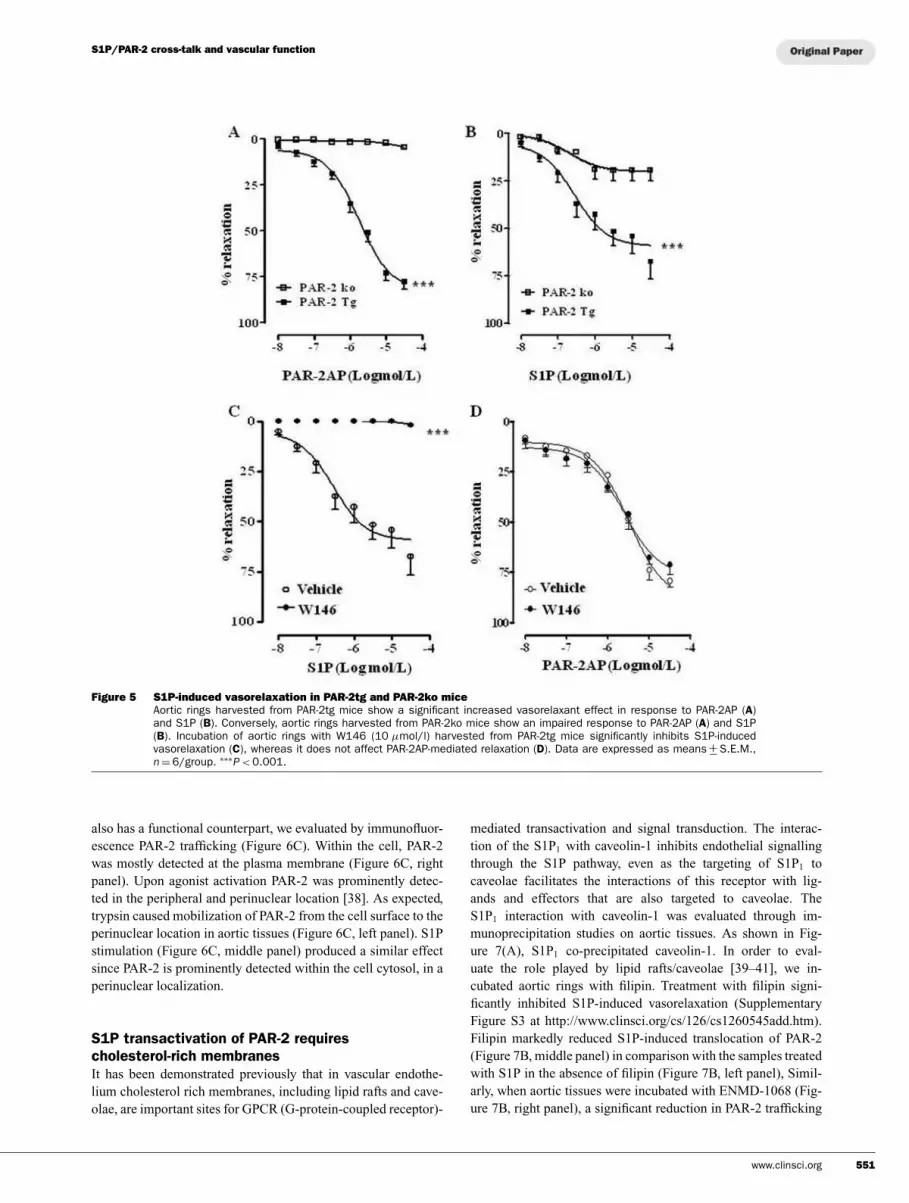
S1P differently affects vascular tone in aortas
harvested from PAR-2tg and PAR-2ko mice
The results obtained suggest that S1P vascular effects are par-
tially mediated by PAR-2. To address this issue we performed
additional experiments on aortic rings isolated from PAR-2tg
or PAR-2ko mice. As expected PAR-2AP-induced relaxation of
isolated aorta was significantly increased in PAR-2tg mice (Fig-
ure 5A), whereas it was abrogated in PAR-2ko mice (Figure 5A).
Similarly, S1P-induced relaxation of aortic rings harvested from
PAR-2tg mice was significantly increased compared with PAR-
2ko mice (Figure 5B). To gain further insight into the role of
PAR-2 in enhancing S1P-induced vasorelaxation, a pharmacolo-
gical modulation was performed in PAR-2tg mice. Aortic rings
were incubated with the S1P1 antagonist W146 prior to adding
S1P. S1P-induced vasodilatation was abrogated by W146. This
result indicates that the effect of S1P on vascular tone is also
mediated through an interaction with S1P1 receptor in PAR-2tg
mice (Figure 5C). Conversely, W146 did not modify PAR-2AP-
induced vasorelaxation in PAR-2tg mice (Figure 5D).
Figure 4 S1P-induced relaxation is not affected by SAM-11
Pre-treatment of aortic rings with SAM-11, an antibody that recog-nizes the extracellular cleavage site involved in PAR-2 activation, in-hibits trypsin-induced vasorelaxation (A), but does not affect eitherPAR-2AP- (B) and S1P- (C) induced relaxation. Data are expressed asmeans +
− S.E.M., n = 6/group. ∗∗∗P < 0.001 compared with vehicle. U,
units.
S1P promotes PAR-2 trafficking
On the basis of the results described above, we suggest that
PAR-2 transactivation by S1P occurs and it is mediated by S1P1.
This functional interaction between S1P1 and PAR-2 is further
demonstrated by co-immunoprecipitation and immunofluores-
cence studies. Aortic tissue was used for S1P1 immunoprecip-
itation and PAR-2 co-precipitation was monitored. As shown in
Figure 6(A), S1P1 co-precipitates PAR-2. Similarly, an immun-
ofluorescence study performed on aortic rings highlighted S1P1
and PAR-2 co-localization in endothelium aortic vessel (Fig-
ure 6B). In order to demonstrate that this physical interaction
550 C© The Authors Journal compilation C© 2014 Biochemical Society
S1P/PAR-2 cross-talk and vascular function
Figure 5 S1P-induced vasorelaxation in PAR-2tg and PAR-2ko mice
Aortic rings harvested from PAR-2tg mice show a significant increased vasorelaxant effect in response to PAR-2AP (A)and S1P (B). Conversely, aortic rings harvested from PAR-2ko mice show an impaired response to PAR-2AP (A) and S1P(B). Incubation of aortic rings with W146 (10 µmol/l) harvested from PAR-2tg mice significantly inhibits S1P-inducedvasorelaxation (C), whereas it does not affect PAR-2AP-mediated relaxation (D). Data are expressed as means +
− S.E.M.,n = 6/group. ∗∗∗
P < 0.001.
also has a functional counterpart, we evaluated by immunofluor-
escence PAR-2 trafficking (Figure 6C). Within the cell, PAR-2
was mostly detected at the plasma membrane (Figure 6C, right
panel). Upon agonist activation PAR-2 was prominently detec-
ted in the peripheral and perinuclear location [38]. As expected,
trypsin caused mobilization of PAR-2 from the cell surface to the
perinuclear location in aortic tissues (Figure 6C, left panel). S1P
stimulation (Figure 6C, middle panel) produced a similar effect
since PAR-2 is prominently detected within the cell cytosol, in a
perinuclear localization.
S1P transactivation of PAR-2 requires
cholesterol-rich membranes
It has been demonstrated previously that in vascular endothe-
lium cholesterol rich membranes, including lipid rafts and cave-
olae, are important sites for GPCR (G-protein-coupled receptor)-
mediated transactivation and signal transduction. The interac-
tion of the S1P1 with caveolin-1 inhibits endothelial signalling
through the S1P pathway, even as the targeting of S1P1 to
caveolae facilitates the interactions of this receptor with lig-
ands and effectors that are also targeted to caveolae. The
S1P1 interaction with caveolin-1 was evaluated through im-
munoprecipitation studies on aortic tissues. As shown in Fig-
ure 7(A), S1P1 co-precipitated caveolin-1. In order to eval-
uate the role played by lipid rafts/caveolae [39–41], we in-
cubated aortic rings with filipin. Treatment with filipin signi-
ficantly inhibited S1P-induced vasorelaxation (Supplementary
Figure S3 at http://www.clinsci.org/cs/126/cs1260545add.htm).
Filipin markedly reduced S1P-induced translocation of PAR-2
(Figure 7B, middle panel) in comparison with the samples treated
with S1P in the absence of filipin (Figure 7B, left panel), Simil-
arly, when aortic tissues were incubated with ENMD-1068 (Fig-
ure 7B, right panel), a significant reduction in PAR-2 trafficking
www.clinsci.org 551

F. Roviezzo and others
Figure 6 S1P induces PAR-2 trafficking in aortic tissues
(A) S1P1 was immunoprecipitaed and immunoblotted with antibody against PAR-2. (B) The expression of PAR-2 and S1P1
in tissue sections is shown by fluorescence microscopy. Co-expression of PAR-2 (left panel, green) and S1P1 (middle panel,red) is apparent in several cells in the vessel wall. The right panel shows the merge of the left and middle panels. Endothelialcells are labelled with an anti-CD31 antibody (white). Nuclei are stained with DAPI (4′,6-diamidino-2-phenylindole) (blue).(C) The immunolabelling for PAR-2 is shown in aortic rings incubated with trypsin (3 units/ml; 10 min) or S1P (1 µmol/l;10 min). The areas in the rectangles are illustrated at higher magnification in the lower panels. In the samples exposed totrypsin (left panels) or S1P (middle panels), the increased accumulation of PAR-2 (green, arrows) is visible in the cytoplasmof the endothelial cells (lectin I, red). In control tissue (right panels), PAR-2 is present mostly at the cell membrane level(arrowheads).The subcellular localization is better visible in the insets. Nuclei are stained with DAPI (blue).
occured following S1P challenge. Taken together, these data sug-
gest a key role for the endothelium in the cross-talk between S1P1
and PAR-2.
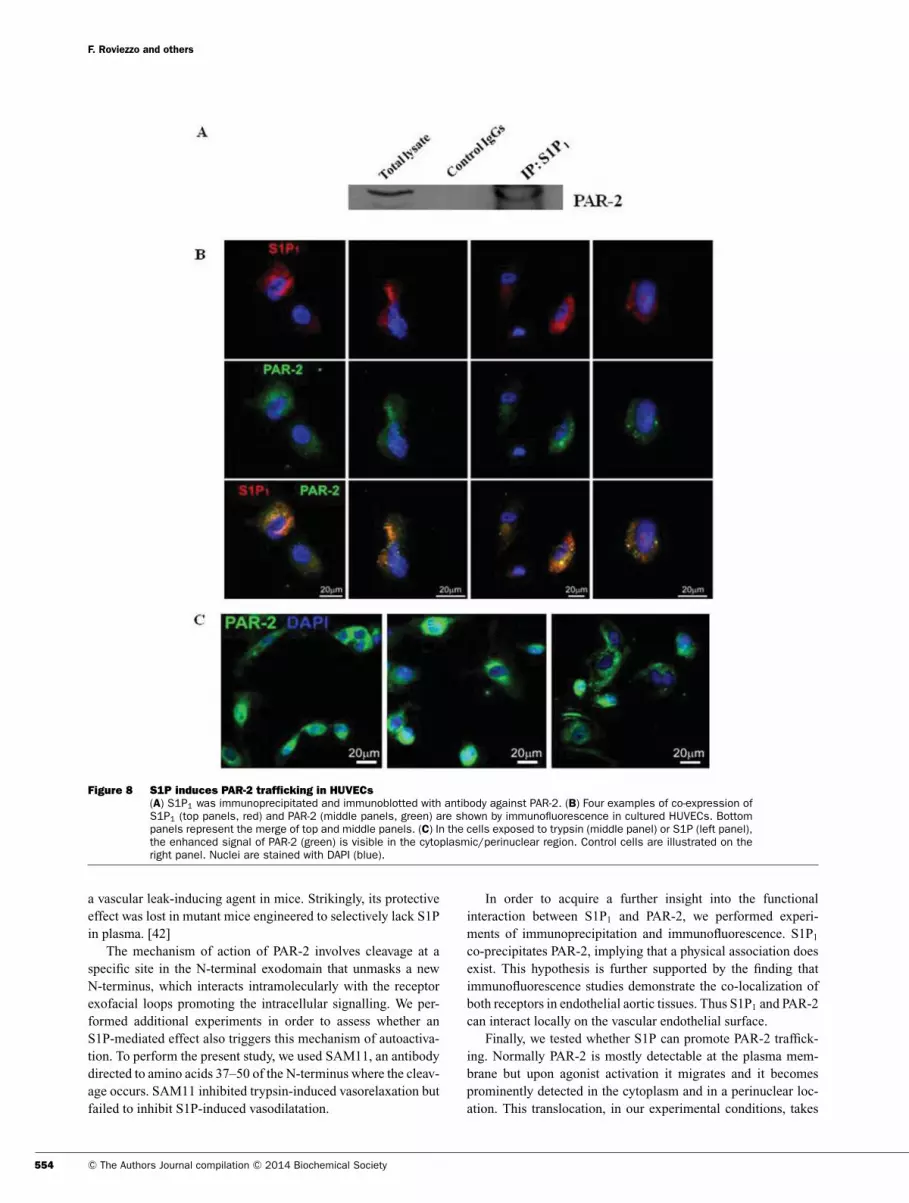
Finally, we tested whether this signalling pathway was detect-
able in a human cellular model. To this end, we studied cultured
endothelial cells (HUVECs). We observed the co-precipitation of
S1P1 and PAR-2 (Figure 8A) and the co-expression of S1P1 (Fig-
ure 8B, upper panels, red) and PAR-2 (Figure 8B, middle panels,
green). By evaluating, through immunofluorescence, PAR-2 traf-
ficking we demonstrate that S1P induced a functional activation
of PAR-2, (Figure 8C). When HUVECs were exposed to tryp-
sin (Figure 8C, middle panel) or S1P (Figure 8C, left panel) it
is possible to detect the increase in fluorescence due to PAR-2
(green) staining in the cytoplasmic/perinuclear region as well as
in vascular tissues when compared with the vehicle (Figure 8C,
right panel).
DISCUSSION
The role of S1P in the physiology of cardiovascular system
has been widely investigated leading to the demonstration of a
finely tuned integration of S1P sources and receptors to modulate
552 C© The Authors Journal compilation C© 2014 Biochemical Society

S1P/PAR-2 cross-talk and vascular function
Figure 7 Caveolae dysfunction affects S1P–PAR-2 interaction
(A) CAV-1 was immunoprecipitated and immunoblotted with antibody against S1P1. (B) Aortic rings treated with S1P only(left panels) and samples incubated with filipin (middle panels) or ENMD-1068 (right panels) and then exposed to S1Pare shown. S1P-induced cytosolic accumulation of PAR-2 (left panel) was significantly reduced by filipin (middle panel) orENMD-1068 (right panel). Endothelial cells are labelled with an anti-vWF antibody (red). Cell nuclei are labelled with DAPI(blue).
vascular tone [6–8]. PAR-2 involvement in cardiovascular dis-
eases has also been demonstrated [28,30]. Although numerous
previous studies have described a tight link between proteases
and sphingolipids and the modulation of vascular tone by S1P
and PAR-2, the cross-talk between these receptors has not been
as yet investigated.
Administration of S1P or PAR-2AP induces vascular relaxa-
tion that is abrogated following mechanical removal of endothe-
lium. Thus both agonists require an intact endothelium to display
a vascular action, suggesting common endothelium-derived me-
diators. Since it is known that S1P-induced relaxation requires
endothelial-derived NO through the PI3K/Akt/eNOS pathway
[11–13], we investigated the role played by the this pathway.
As expected also in our experimental conditions, S1P induces
a significant phosphorylation of both Akt and eNOS. In per-
fect tune with these data, S1P-induced vasorelaxtion is abrogated
by L-NAME or wortmannin. Conversely, PAR-2 while inducing
a significant phosphorylation of eNOS, does not promote Akt
phosphorylation. Functional data indicate a significant inhibition
of PAR-2AP-induced vasorelaxation by both wortmannin and L-
NAME. Thus S1P and PAR-2 modulate vascular tone activating
signalling pathway that converges on PI3K, but differs in the
downstream signalling. We went on in the present study to evalu-
ate the effect of a pharmacological modulation of both receptors
by using specific and selective receptor antagonists.
Mouse aorta expresses S1P1, S1P2 and S1P3 receptor types but
S1P1 is the most expressed and mediates S1P-induced vascular
relaxation. Either W146, a S1P1 antagonist, or PTX, that spe-
cifically uncouples S1P1 from its Gi-protein, abrogates S1P- in-
duced vasorelaxation. Neither W146 nor PTX affects PAR-2AP
or trypsin-induced vasorelaxation. Conversely, ENMD-1068, the
PAR-2 antagonist, significantly inhibits S1P-induced vasorelaxa-
tion. Indeed, ENMD-1068 significantly impairs S1P-induced vas-
orelaxation. Therefore all together these data demonstrate that (i)
both signals originate from independently engaged receptors, (ii)
S1P activation involves PAR-2, (iii) S1P1 is the main subtype
receptor involved.
To further sustain our hypothesis, we performed additional
experiments by using aortas harvested from either PAR-2tg or
PAR-2 ko mice. The results obtained further confirmed that PAR-
2 is involved in the vascular effect of S1P and that its positive
modulation amplifies S1P effect on vascular tone. Indeed, ves-
sels harvested from PAR-2tg mice and challenged in vitro with
S1P displayed an enhanced response, whereas in vessels harves-
ted from PAR2ko mice the S1P vascular effect was significantly
reduced.
Our hypothesis of a functional cross-talk between PAR-2 and
S1P is also sustained by literature data. In fact, it has been shown
that SLIGRL, the same PAR-2 activating peptide we have used
in our experiments (PAR-2AP), prevents the lethal response to
www.clinsci.org 553
F. Roviezzo and others
Figure 8 S1P induces PAR-2 trafficking in HUVECs
(A) S1P1 was immunoprecipitated and immunoblotted with antibody against PAR-2. (B) Four examples of co-expression ofS1P1 (top panels, red) and PAR-2 (middle panels, green) are shown by immunofluorescence in cultured HUVECs. Bottompanels represent the merge of top and middle panels. (C) In the cells exposed to trypsin (middle panel) or S1P (left panel),the enhanced signal of PAR-2 (green) is visible in the cytoplasmic/perinuclear region. Control cells are illustrated on theright panel. Nuclei are stained with DAPI (blue).
a vascular leak-inducing agent in mice. Strikingly, its protective
effect was lost in mutant mice engineered to selectively lack S1P
in plasma. [42]
The mechanism of action of PAR-2 involves cleavage at a
specific site in the N-terminal exodomain that unmasks a new
N-terminus, which interacts intramolecularly with the receptor
exofacial loops promoting the intracellular signalling. We per-
formed additional experiments in order to assess whether an
S1P-mediated effect also triggers this mechanism of autoactiva-
tion. To perform the present study, we used SAM11, an antibody
directed to amino acids 37–50 of the N-terminus where the cleav-
age occurs. SAM11 inhibited trypsin-induced vasorelaxation but
failed to inhibit S1P-induced vasodilatation.
In order to acquire a further insight into the functional
interaction between S1P1 and PAR-2, we performed experi-
ments of immunoprecipitation and immunofluorescence. S1P1
co-precipitates PAR-2, implying that a physical association does
exist. This hypothesis is further supported by the finding that
immunofluorescence studies demonstrate the co-localization of
both receptors in endothelial aortic tissues. Thus S1P1 and PAR-2
can interact locally on the vascular endothelial surface.
Finally, we tested whether S1P can promote PAR-2 traffick-
ing. Normally PAR-2 is mostly detectable at the plasma mem-
brane but upon agonist activation it migrates and it becomes
prominently detected in the cytoplasm and in a perinuclear loc-
ation. This translocation, in our experimental conditions, takes
554 C© The Authors Journal compilation C© 2014 Biochemical Society

S1P/PAR-2 cross-talk and vascular function
place upon challenge of aortic rings with either trypsin or PAR-
2AP. S1P incubation with aortic rings replicates this effect,
demonstrating that S1P triggers PAR-2 translocation. To further
validate this hypothesis, we performed a reversal experiment by
using ENMD-1068, the PAR-2 antagonist. ENMD-1068 abrog-
ates PAR-2 translocation caused by S1P. Therefore S1P causes
activation of PAR-2 by inducing its translocation.
It is known that PAR-2 and S1P1 are widely localized in the
caveolae [39–41]. The compartmentalization of signalling pro-
teins is an important cellular tool to afford efficacy and specificity
of inter-molecular communication processes. This is particularly
the case in endothelial cells signalling machinery, where the dis-
tinct fluid phase of caveolae seems to help draw together proteins
involved in a variety of signalling pathways, therefore facilitat-
ing protein–protein and protein–membrane interactions neces-
sary for signal transduction [43]. In order to further demonstrate
that PAR-2/S1P1 interaction is necessary for signalling to oc-
cur, we treated aortic rings with filipin an agent that depletes
caveolae of cholesterol content, altering their fluidity. Treat-
ment of aortic rings with filipin blunts S1P ability to induce
PAR-2 translocation. Thus S1P1/PAR-2 have physical proximity,
as demonstrated by the immunofluorescence studies and their
endothelial compartmentalization is crucial for their functional
interaction.
In HUVECs, the same phenomenon is clearly visible thereby
indicating that the mechanisms we have defined in mouse aorta
is shared by human endothelial cells.
In conclusion, the findings in the present paper demonstrate
that not only is PAR-2 activated by S1P, but it also works in con-
cert with S1P1 to regulate vascular homoeostasis. Further studies
will be necessary to better clarify the molecular basis of this func-
tional interaction. However, this cross-talk could play a major role
in vascular diseases where both signals have been demonstrated to
play a role such as angiogenesis, restenosis and neointima form-
ation, and atherosclerotic remodelling [7,13,20,29]. The exist-
ence of this cross-talk between S1P1 and PAR-2 further supports
the concept of a tight link between proteases and cardiovascular
function.
CLINICAL PERSPECTIVES
� S1P exerts a diverse set of vascular responses through the
activation of heterotrimeric G-proteins that in turn modulate
the activity of various downstream signalling molecules. PAR-
2 has been shown to be involved in vascular inflammation as
well as in other inflammatory-based diseases.� In the present study, we demonstrate that S1P-mediated vascu-
lar effect involves PAR-2 activation. The hypothesis of the ex-
istence of this transactivation between S1P1 and PAR-2 further
strengthens the concept of a tight link between cardiovascular
function and the inflammatory processes.� We believe that this aspect of the study will be of great interest
since the presence of this cross-talk between S1P and PAR-2
further supports the concept that protease receptors may play
an important role in cardiovascular inflammation.
AUTHOR CONTRIBUTION
Fiorentina Roviezzo and Giuseppe Cirino conceived and designed
the experiments and wrote the paper; Luana De Gruttola, Anto-
nio Bertolino and Nikol Sullo performed the vascular functional
experiments; Antonella De Angelis, Konrad Urbanek and Bruno
D’Agostino performed the immunofluorescence experiments; Raf-
faele De Palma performed the RT–PCR; Mariarosaria and Bucci
Vincenzo Brancaleone performed the cell and molecular experi-
ments; Angela Ianaro and Raffaella Sorrentino analysed data and
contributed to the writing of the paper.
FUNDING
This work was supported by the Ministero della Universita e Ricerca
Scientifica, Italy [grant number PRIN 2009].
REFERENCES
1 Pitson, S. M. (2011) Regulation of sphingosine kinase andsphingolipid signalling. Trend Biochem. Sci. 2, 97–107
2 Hannun, Y. A. and Obeid, L. M. (2008) Principles of bioactivelipid signalling: lessons from sphingolipids. Nat. Rev. Mol. CellBiol. 9, 139–150
3 Maceyka, M., Milstien, S. and Spiegel, S. (2009)Sphingosine-1-phosphate: the Swiss army knife of sphingolipidsignaling. J. Lipid Res. 50, S272–S276
4 Sanchez, T. and Hla, T. (2004) Structural and functionalcharacteristics of S1P receptors. J. Cell Biochem. 92, 913–922
5 Bartke, N. and Hannun, Y. A. (2009) Bioactive sphingolipids:metabolism and function. J. Lipid Res. 50, S91–S96
6 Levkau, B. (2008) Sphingosine-1-phosphate in the regulation ofvascular tone: a finely tuned integration system of S1P sources,receptors, and vascular responsiveness. Circ. Res. 103,231–233
7 Lucke, S. and Levkau, B. (2010) Endothelial functions ofsphingosine-1-phosphate. Cell Physiol. Biochem. 26, 87–96
8 Ozaki, H., Hla, T. and Lee, M. J. (2003) Sphingosine-1-phosphate signaling in endothelial activation. J. Atheroscler.Thromb. 10, 125–131
9 Dantas, A. P., Igarashi, J. and Michel, T. (2003) Sphingosine1-phosphate and control of vascular tone. Am. J. Physiol. HeartCirc. Physiol. 284, H2045–H2052
10 Igarashi, J. and Michel, T. (2008) S1P and eNOS regulation.Biochim. Biophys. Acta 1781, 489–495
11 Igarashi, J. and Michel, T. (2001) Sphingosine 1-phosphate andisoform-specific activation of phosphoinositide 3-kinase β.Evidence for divergence and convergence of receptor-regulatedendothelial nitric-oxide synthase signaling pathways. J. Biol.Chem. 276, 36281–36288
12 Roviezzo, F., Bucci, M., Delisle, C., Brancaleone, V., Di Lorenzo,A., Mayo, I. P., Fiorucci, S., Fontana, A., Gratton, J. P. and Cirino,G. (2006) Essential requirement for Sphingosine kinase activityin eNOS-dependent NO release and vasorelaxation. FASEB J. 20,340–342
13 d’Emmanuele di Villa Bianca, R., Sorrentino, R., Roviezzo, F.,Imbimbo, C., Palmieri, A., De Dominicis, G., Montorsi, F., Cirino,G. and Mirone, V. (2006) Sphingosine-1-phosphate inducesendothelial NOS activation through phosphorylation in humancorpus cavernosum. J. Pharmacol. Exp. Ther. 316, 703–708
14 Tanimoto, T., Jin, Z. G. and Berk, B. C. (2002) Transactivation ofvascular endothelial growth factor (VEGF) receptor Flk-1/KDR isinvolved in sphingosine 1-phosphate-stimulated phosphorylationof Akt and endothelial nitric-oxide synthase (eNOS). J. Biol.Chem. 277, 42997–43001
www.clinsci.org 555

F. Roviezzo and others
15 Endo, A., Nagashima, K., Kurose, H., Mochizuki, S., Matsuda, M.and Mochizuki, N. (2002) Sphingosine 1-phosphate inducesmembrane ruffling and increases motility of human umbilical veinendothelial cells via vascular endothelial growth factor receptorand CrkII. J. Biol. Chem. 277, 23747–23754
16 Hobson, J. P., Rosenfeldt, H. M., Barak, L. S., Olivera, A.,Poulton, S., Caron, M. G., Milstien, S. and Spiegel, S.(2001) Role of the sphingosine-1-phosphate receptorEDG-1 in PDGF-induced cell motility. Science 291,1800–1803
17 Alderton, F., Rakhit, S., Kong, K. C., Palmer, T., Sambi, B., Pyne,S. and Pyne, N. J. (2001) Tethering of the platelet-derived growthfactor β receptor to G-protein-coupled receptors: a novel platformfor integrative signaling by these receptor classes in mammaliancells. J. Biol. Chem. 276, 28578–28585
18 Tanimoto, T., Lungu, A. O. and Berk, B. C. (2004) Sphingosine1–phosphate transactivates the platelet-derived growth factor β
receptor and epidermal growth factor receptor in vascularsmooth muscle cells. Circ. Res. 94, 1050–1058
19 Benaud, C., Oberst, M., Hobson, J. P., Spiegel, S., Dickson, R. B.and Lin, C. Y. (2002) Sphingosine 1-phosphate, present inserum-derived lipoproteins, activates matriptase. J. Biol. Chem.277, 10539–10546
20 Macfarlane, S. R., Seatter, M. J., Kanke, T., Hunter, G. D. andPlevin, R. (2001) Proteinase-activated receptors. Pharmacol.Rev. 53, 245–282
21 Vu, T. K., Hung, D. T., Wheaton, V. I. and Coughlin, S. R. (1991)Molecular cloning of a functional thrombin receptor reveals anovel proteolytic mechanism of receptor activation. Cell 64,1057–1106
22 Hirano, K. and Kanaide, H. (2003) Role of protease-activatedreceptors in the vascular system. J. Atheroscler. Thromb. 10,211–225
23 Sobey, C. G. and Cocks, T. M. (1998) Activation of proteaseactivated receptor-2 (PAR-2) elicits nitric oxide-dependentdilatation of the basilar artery in vivo. Stroke 29,1439–1444
24 Robin, J., Kharbanda, R., Mclean, P., Campbell, R. and Vallance,P. (2003) Protease-activated receptor 2-mediated vasodilatationin humans in vivo: role of nitric oxide and prostanoids.Circulation 107, 954–959
25 D’Andrea, M. R., Derian, C. K., Leturcq, D., Baker, S. M.,Brunmark, A., Ling, P., Darrow, A. L., Santulli, R. J., Brass, L. F.and Andrade-Gordon, P. (1998) Characterization of proteaseactivated receptor-2 immunoreactivity in normal human tissues.J. Histochem. Cytochem. 46, 157–164
26 Nystedt, S., Ramakrishnan, V. and Sundelin, J. (1996)The proteinase activated receptor 2 is induced byinflammatory mediators in human endothelial cells.Comparison with the thrombin receptor. J. Biol. Chem. 271,14910–14915
27 Cirino, G. and Cicala, C. (2003) Upregulation of proteinase-activated receptors (PARs) and cardiovascular function. DrugDev. Res. 60, 20–23
28 Cocks, T. M. and Moffatt, J. D. (2000) Protease-activatedreceptors: sentries for inflammation? Trends Pharmacol. Sci. 21,103–108
29 Roviezzo, F., Bucci, M., Brancaleone, V., Di Lorenzo, A., Geppetti,P., Farneti, S., Parente, L., Lungarella, G., Fiorucci, S. and Cirino,G. (2005) Proteinase-activated receptor-2 mediates arterialvasodilation in diabetes. Arterioscler. Thromb. Vasc. Biol. 25,2349–2354
30 Cicala, C., Pinto, A., Bucci, M., Sorrentino, R., Walker, B., Harriot,P., Cruchley, A., Kapas, S., Howells, G. L. and Cirino, G. (1999)Protease-activated receptor-2 involvement in hypotension innormal and endotoxemic rats in vivo. Circulation 99, 2590–2597
31 Kang, H., Kwak, H. I., Kaunas, R. and Bayless, K. J. (2011) Fluidshear stress and sphingosine1-phosphate activate calpain topromote membrane type 1 matrix metalloproteinase (MT1-MMP)
membrane translocation and endothelial invasion intothree-dimensional collagen matrices. J. Biol. Chem. 286,42017–42026
32 Zebrakovska, I., Masa, M., Srp, J., Horn, M., Vavrova, K. andMares, M. (2011) Complex modulation of peptidolytic activity ofcathepsin D by sphingolipids. Biochim. Biophys. Acta 1811,1097–1104
33 Kim, E. S., Kim, J. S., Kim, S. G., Hwang, S., Lee, C. H. andMoon, A. (2011) Sphingosine 1-phosphate regulates matrixmetalloproteinase-9 expression and breast cell invasion throughS1P3–Gαq coupling. J. Cell Sci. 124, 2220–2230
34 Kojima, K. and Inouye, K. (2011) Activation of matriptasezymogen. J. Biochem. 150, 123–125
35 Niessen, F., Schaffner, F., Furlan-Freguia, C., Pawlinski, R.,Bhattacharjee, G., Chun, J., Derian, C. K., Andrade-Gordon, P.,Rosen, H. and Ruf, W. (2008) Dendritic cell PAR1–S1P3
signalling couples coagulation and inflammation. Nature 452,654–658
36 Kelso, E. B., Lockhart, J. C., Hembrough, T., Dunning, L., Plevin,R., Hollenberg, M. D., Sommerhoff, C. P., McLean, J. S. andFerrell, W. R. (2006) Therapeutic promise of proteinase-activatedreceptor-2 antagonism in joint inflammation. J. Pharmacol. Exp.Ther. 316, 1017–1024
37 Kaiser, R. A., Oxhorn, B. C., Andrews, G. and Buxton, I. L. (2002)Functional compartmentation of endothelial P2Y receptorsignaling. Circ. Res. 91, 292–299
38 Dery, O., Thoma, M. S., Wong, H., Grady, E. F. and Bunnett, N. W.(1999) Trafficking of proteinase-activated receptor-2 andβ -arrestin-1 tagged with green fluorescent protein.β -Arrestin-dependent endocytosis of a proteinase receptor.J. Biol. Chem. 274, 18524–18535
39 Awasthi, V., Mandal, S. K., Papanna, V., Rao, L. V. and Pendurthi,U. R. (2007) Modulation of tissue factor-factor VIIa signaling bylipid rafts and caveolae. Arterioscler. Thromb. Vasc. Biol. 27,1447–1455
40 Hachem, J. P., Houben, E., Crumrine, D., Man, M. Q., Schurer, N.,Roelandt, T., Choi, E. H., Uchida, Y., Brown, B. E., Feingold, K. R.and Elias, P. M. (2006) Serine protease signaling of epidermalpermeability barrier homeostasis. J. Invest. Dermatol. 126,2074–2086
41 Igarashi, J. and Michel, T. (2000) Agonist-modulated targeting ofthe EDG-1 receptor to plasmalemmal caveolae. eNOS activationby sphingosine 1-phosphate and the role of caveolin-1 insphingolipid signal transduction. J. Biol. Chem. 275,32363–32370
42 Camerer, E., Regard, J. B., Cornelissen, I., Srinivasan, Y.,Duong, D. N., Palmer, D, Pham, T. H., Wong, J. S., Pappu,R. and Coughlin, S. R. (2009) Sphingosine-1-phosphatein the plasma compartment regulates basal andinflammation-induced vascular leak in mice. J. Clin. Invest. 119,1871–1879
43 Gonzalez, E., Kou, R., Lin, A. J., Golan, D. E. and Michel, T.(2002) Subcellular targeting and agonist-induced site-specificphosphorylation of endothelial nitric-oxide synthase. J. Biol.Chem. 277, 39554–39560
Received 19 June 2013/7 October 2013; accepted 17 October 2013
Published as Immediate Publication 17 October 2013, doi: 10.1042/CS20130272
556 C© The Authors Journal compilation C© 2014 Biochemical Society

Clinical Science (2014) 126, 545–556 (Printed in Great Britain) doi: 10.1042/CS20130272
SUPPLEMENTARY ONLINE DATA
Involvement of proteinase activated receptor-2in the vascular response to sphingosine1-phosphateFiorentina ROVIEZZO∗, Antonella DE ANGELIS†, Luana DE GRUTTOLA∗, Antonio BERTOLINO∗, Nikol SULLO†,
Vincenzo BRANCALEONE‡, Mariarosaria BUCCI∗, Raffaele DE PALMA§, Konrad URBANEK†,
Bruno D’AGOSTINO†, Angela IANARO∗, Raffaella SORRENTINO∗ and Giuseppe CIRINO∗
∗Dipartimento di Farmacia, Universita di Napoli Federico II, Napoli, Italy
†Dipartimento di Medicina Sperimentale, Sezione di Farmacologia L. Donatelli, Seconda Universita degli Studi di Napoli, Napoli, Italy
‡Dipartimento di Chimica, Universita degli Studi della Basilicata, Pontenza, Italy
§Dipartimento di Internistica Clinica e Sperimentale, Seconda Universita degli Studi di Napoli, Napoli, Italy
MATERIALS AND METHODS
Generation of PAR2tg mice
A 140-kb human BAC clone (identification no. 23C20) con-
taining the entire genomic sequence of human PAR-2 was ob-
tained from the Lawrence Berkley National Laboratory Hu-
man Genome Center (University of California, Berkeley, CA,
U.S.A.). The PAR2 gene is under the control of the endo-
genous human promoter and flanked by a minimum of 30 kb
of genomic sequence. This BAC DNA was microinjected
into the pronuclei of FVB/N eggs. F0 pups were screened
for incorporation of the transgene using the human-specific
PAR-2 primer set 5′-GTAAGCTTGATGGCACTACCC and 3′-
TCTGATCATCAGCACATAGGC and was confirmed by South-
ern blotting. Copy number of the transgene was estimated at
5–10 copies/diploid cell based on the intensity of the band in a
Southern blot compared with a single copy gene. Charles River
Breeding Laboratories were used for controls.
Generation of PAR2ko mice
Four mouse genomic clones were isolated using mouse PAR2
primers. The short arm of the vector (Xba–Kpn fragment) was
cloned into the ppNTT neo vector. A hygromycin selectable
marker and a lox P sequence from the vector pBS-PGKhyg/loxP
modified with Sse8783 linkers were incorporated. Finally, the
long arm of the vector (Xho–Sse) was ligated in to produce the
final 16-kb targeting vector. RW4 embryonic stem cells were
electroporated with the vector and screened for homologous re-
combination using standard selection conditions of 10–12 days
of G418 treatment. Surviving clones were screened by South-
ern blot analysis and positive clones were injected into C57
blastocysts. Chimaeric males were back-crossed to C57 females
to test for germline transmission and to 129SvJ females to cre-
ate an inbred strain of homozygous KO mice. Animals were
genotyped using primers specific for the mouse PAR2 gene (5′-
GCATTGAACATCACCACCTG, 3′-GGATAGCCCTCTGCCT-
Correspondence: Professor Giuseppe Cirino (email [email protected]).
Table S1 Primers used in qRT-PCR
FW, forward; RV, reverse.
Primer Sequence
Mouse S1P1 FW 5′ -TTCCACCGGCCCATGTACTA-3′
Mouse S1P1 RV 5′ -CAAACATACTCCCTTCCCGC-3′
Mouse S1P2 FW 5′ -AACTCCCGTGCAGTGGTTTG-3′
Mouse S1P2 RV 5′ -CATCAGCATTCGGCAGCTTT-3′
Mouse S1P3 FW 5′ -GCGACACCTGACCATGATCA-3′
Mouse S1P3 RV 5′ -GCAGTCGGGAAAGTTCTCCAG-3′
Mouse GAPDH FW 5′ -TGCACCACCAACTGCTTAGC-3′
Mouse GAPDH RV 5′ -TGATGGCATGGACTGTGGTC-3′
TTTC) as well as for the selection marker (5′-GCAGCCA-
ATATGGGATCG, 3′-ATCAGAGCAGCCGATTGTCT).
Cell culture
HUVECs were purchased from Promocell. All experiments were
performed on low-passage cell cultures. Cells were grown in
endothelial growth medium-2 (EBM-2, 2 % FBS, VEGF, R3-
IGF-1, hEGF, hFGF, hydrocortisone, ascorbic acid, heparin and
GA-1000) (Clonetics) at 37 ◦C and in a 5 % (v/v) CO2 atmosphere.
To examine trafficking cells were maintained at 37 ◦C in DMEM
containing 0.1 % BSA. Cells were observed after addition of
trypsin (10 nM) or S1P (1 μM). Cells were washed and fixed
with 4 % (v/v) paraformaldehyde in 100 mM PBS, pH7.4.
Western blot analysis
Thoracic aorta was dissected and cleaned from fat and connect-
ive tissue. Rings, of the same size as described before, were
homogenized in RIPA buffer (50 mM Tris/HCl, pH 7.4, 1% Tri-
ton X-100, 0.25 %, sodium deoxycolate, 150 mM NaCl, 1 mmol/l
EDTA, 1 mmol/l PMSF, 10 μg/ml aprotinin, 20 μmol/l leupeptin
and 50 mmol/l sodium fluoride) using a Polytron homogenizer
www.clinsci.org
F. Roviezzo and others
Figure S1 Effect of W146 and ENMD-1068 on eNOS or Akt
phosphorylation
(A) Representative Western blot of eNOS or Akt phosphorylation in aor-tic rings incubated with both S1P (10− 6 mol/l) and PAR-2AP (10− 6
mol/l) without or with W146 or ENMD-1068. Histograms indicate therelative densitometry compared with total eNOS protein. Data are ex-pressed as means +
− S.E.M., n = 6/group. ∗P < 0.05 and ∗∗
P < 0.01compared with vehicle.
(two cycles of 10 s at maximum speed). Lysates were centrifuged
14 000 g in a pre-cooled centrifuge for 15 min. Lysates were
spearated by SDS/PAGE (10 % gels) and proteins were trans-
ferred on to a PVDF membrane. Membranes were blocked by in-
cubation in PBS containing 0.1 % Tween 20 and 5 % (w/v) non-fat
dried skimmed milk for 2 h, followed by overnight incubation at
4 ◦C with anti-eNOS (1:1000 dilution) and anti-(phospho-eNOS)
(1:1000 dilution) antibodies. The filters were washed extensively
in PBS containing 0.1 % Tween 20, before incubation for 2 h
with HRP (horseradish peroxidase)-conjugate secondary anti-
bodies. Membranes were then washed and developed using ECL
(enhanced chemiluminescence substrate).
Immunoprecipitation
The cell lysate was pre-cleared by adding 100 μl of either Protein
A or G–agarose/sepharose bead slurry (50 %) for 1 ml of cell lys-
ate. The Protein A or G beads were removed by centrifugation.
The supernatant was transferred to a fresh centrifuge tube. The
immunoprecipitating antibody (20 μg of anti-S1P1) was added
to 500 μl of cell lysate. The cell lysate/antibody mixture was
gently rocked overnight at 4 ◦C on an orbital shaker. The im-
munocomplex was captured by adding 100 μl of Protein A or
G–agarose/sepharose bead slurry and gently rocking on either
a orbital shaker for 1 h at 4 ◦C. The agarose/sepharose beads
were collected by pulse centrifugation. The supernatant was dis-
carded and the beads washed three times with 800 μl of ice-cold
modified RIPA buffer. The agarose/sepharose beads were sus-
pended in 60 μl of 2× sample buffer and mixed gently. The
agarose/sepharose beads were boiled for 5 min to dissociate the
immunocomplexes from the beads. The denatured proteins were
separated by SDS/PAGE (10 % gel) and transferred on to a PVDF
membrane. The total lysate was used as positive control and the
isotype control antibody as negative control. Membrane were
blocked by incubation in PBS containing 0.1 % Tween 20 and
5 % non-fat dried skimmed milk for 2 h, followed by a overnight
incubation at 4 ◦C with anti-PAR-2. The filters were washed ex-
tensively in PBS containing 0.1 % Tween 20, before incubation
for 2 h with an HRP-conjugated secondary antibody. Membranes
were then washed and developed using ECL.
mRNA quantification by qPCR
Briefly, total RNA from homogenized aorta tissue was extracted
by using TRIzol® reagent (Invitrogen), subsequently, to eliminate
genomic DNA contamination, 1 μg of the RNA was treated
with RQ1 RNase-free DNase I (Promega Corporation) and
reverse transcription was performed using M-MLV Reverse
Transcriptase (Invitrogen) according to the manufacturer’s
recommendations. qPCR was carried out with cDNAs by
using Sybr Green PCR Master Mix (Applied Biosystems) and
7500 real-time PCR System (Applied Biosystems). All cDNA
samples were run in triplicate in 25-μl reactions. Primers used
were specifically designed using Primer Express Software
2.0 (Applied Biosystems) and validated for their specificity
(Table S1). Samples were incubated at 50 ◦C for 2 min and at
95 ◦C for 10 min followed by 40 cycles at 95 ◦C for 15 s and
60 ◦C for 1 min. Differences in cDNA input were corrected by
normalizing signals obtained with primers specific for GADPH
(glyceraldehyde-3-phosphate dehydrogenase). To exclude non-
specific amplification and/or the formation of primer dimers,
control reactions were performed in the absence of target cDNA.
Gene expression levels were calculated using the 2− �CT method
and are presented as the ratio between mean+−S.E.M.-fold
change of the target gene and GAPDH.
Immunofluorescence
The sections were used for immunolabelling and analysed
by fluorescence microscopy. PAR-2 was detected by indirect
immunofluorescence using a mouse monoclonal anti-PAR-2
antibody. The expression of S1P1 receptor was determined by
indirect immunofluorescence with a rabbit polyclonal anti-S1P1
C© The Authors Journal compilation C© 2014 Biochemical Society
S1P/PAR-2 cross-talk and vascular function
Figure S2 S1P-induced vasorelaxation does not involve PAR-1 activation
(A) PAR-1 antagonist (RWJ 56110) does not modify S1P-induced vasorelaxation. (B) PAR-1AP-mediated vasorelaxation isnot affected by both ENMD 1068 and W146.
antibody. Endothelial cells were identified by labelling with
fluorescein-conjugated Griffonia simplicifolia Lectin I or a rabbit
polyclonal anti-vWF antibody. Nuclei were stained with DAPI.
Briefly, after rehydration, the tissue sections were treated with
10 % normal donkey serum for 20 min at room temperature and
then incubated with the anti-PAR-2 and/or anti-S1P1 primary an-
tibodies diluted 1:200 in PBS. After being washed several times
with PBS, the sections were incubated with the FITC-conjugated
or TRITC-conjugated affinity-pure donkey anti-(mouse IgG) and
anti-(rabbit IgG) secondary antibodies, diluted 1:200, for 60 min
in the dark. Following washing with PBS, the sections were in-
cubated with Lectin I diluted 1:200 for 2 h at room temperature.
Incubation with anti-vWF antibody diluted 1:100 was performed
for 1 h following by TRITC-conjugated anti-rabbit secondary
antibody staining. Nuclear counterstaining with DAPI was per-
formed for 15 min at room temperature. Sections were analysed
with a Leica DM5000B fluorescence microscope.Figure S3 Pre-treatment with filipin (1 mg/l) for 30 min inhibits
S1P-induced vasorelaxation
Received 19 June 2013/7 October 2013; accepted 17 October 2013
Published as Immediate Publication 17 October 2013, doi: 10.1042/CS20130272
www.clinsci.org




















