
Investigations of Selected Parameters for Growth of Larval and Juvenile Black Sea Bass Centropristis...
10
JOURNALOFTHE Vol. 31, No. 3 WORLD AQUACULTURE SOCIETY September, 2000 Investigations of Selected Parameters for Growth of Larval and Juvenile Black Sea Bass Centropristis stria& L. DAVID BERLINSKY AND MARK WATSON Department of Fisheries, Animal and Veterinary Science, Building #14, East Farm, University of Rhode Island, Kingston, Rhode Island 02881 USA GEORGE NARDI Great Bay Aquafarms, 153 Gosling Rd, Portsmouth, New Hampshire 03801 USA TERENCE M. BRADLEY' Department of Fisheries, Animal and Veterinary Science, BuiMing #14, East Farm, University of Rhode Island, Kingston, Rhode Island 02881 USA Abstract.-The black sea bass Centropristis striata L. endemic to the U. S. Atlantic and Gulf of Mexico coasts is a highly sought species that commands a high price in the marketplace. Investigations were undertak- en to determine the basic requirements for culture of sea bass larvae and juveniles. Adult black sea bass were captured from the wild and were found to be robust and to adapt well to captivity. Larvae were ob- tained by strip spawning of these adults and survived and grew at higher rates in seawater supplemented with algae (greenwater) and maintained at 22 C, than in seawater with or without supplemental algae at 18 C. Larvae were provided a diet of rotifers through 12 d post hatch (DPH) and weaned over a 3-d period to enriched Arremia replenished daily to a density of 10 individualdml. By 18 DPH, larvae began ingesting a formulated diet for marine finfish and were completely weaned from live feed by 25 DPH. which coincided with the onset of metamorphosis. Juveniles tolerated a range of salinity from 10-32 ppt with the highest growth rate observed at 20 ppt. Culture of larvae and juveniles on a commercial scale was successful using conditions similar to those employed in the laboratory. The findings presented here suggest the potential for commercial culture of this species in the USA. The black sea bass Centropristis striata L. is an important species in commercial and recreational fisheries of the U.S. Atlan- tic coast from the Gulf of Maine to the northeast Gulf of Mexico (Musick and Mer- cer 1977; Fig. 1). Populations of the Atlan- tic and Gulf of Mexico coasts have been divided into two subspecies, Centropristis striutu striata and Centropristis striata me- luna, respectively. Adults and juveniles of I Corresponding author. the Atlantic population undertake seasonal migrations offshore as water temperatures decline in the autumn months (Musick and Mercer 1977; Vaughn et al. 1995). Fish re- main offshore of the New England coast until May and return when water tempera- tures inshore reach approximately 10 C (Caruso 1995). Reproductively, black sea bass, like groupers, are protogynous her- maphrodites developing first as females and later at 3-4 yr of age as males (Wenner et al. 1986). Spawning progresses seasonally from south to north and is initiated as early as March in North Carolina and Virginia and in June and July in New England. Adults may return to their natal spawning grounds in subsequent years (Able et al. 1995). Black sea bass are highly sought and command a premium price in the ethnic live fish market and upscale restaurant trade. As temperate reef species, sea bass generally are inaccessible to bottom trawlers and the fishery primarily consists of capture by fish pots and angling. Demand typically exceeds supply in the northeast U.S. The market value and limited supply of black sea bass warranted evaluation of the potential of this species for commercial aquaculture. Previ- ous investigations of black sea bass were limited in scope, examining spawning and larval development and failed to assess the requisite conditions for culture (Hoff 1970; Roberts et al. 1976; Tucker 1984). In this 0 Copyright by the World Aquaculture Society ZOO0 426
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Investigations of Selected Parameters for Growth of Larval and Juvenile Black Sea Bass Centropristis...

JOURNALOFTHE Vol. 31, No. 3 WORLD AQUACULTURE SOCIETY September, 2000
Investigations of Selected Parameters for Growth of Larval and Juvenile Black Sea Bass Centropristis stria& L.
DAVID BERLINSKY AND MARK WATSON
Department of Fisheries, Animal and Veterinary Science, Building #14, East Farm, University of Rhode Island, Kingston, Rhode Island 02881 USA
GEORGE NARDI Great Bay Aquafarms, 153 Gosling Rd, Portsmouth, New Hampshire 03801 USA
TERENCE M. BRADLEY' Department of Fisheries, Animal and Veterinary Science, BuiMing #14, East Farm,
University of Rhode Island, Kingston, Rhode Island 02881 USA
Abstract.-The black sea bass Centropristis striata L. endemic to the U. S. Atlantic and Gulf of Mexico coasts is a highly sought species that commands a high price in the marketplace. Investigations were undertak- en to determine the basic requirements for culture of sea bass larvae and juveniles. Adult black sea bass were captured from the wild and were found to be robust and to adapt well to captivity. Larvae were ob- tained by strip spawning of these adults and survived and grew at higher rates in seawater supplemented with algae (greenwater) and maintained at 22 C, than in seawater with or without supplemental algae at 18 C. Larvae were provided a diet of rotifers through 12 d post hatch (DPH) and weaned over a 3-d period to enriched Arremia replenished daily to a density of 10 individualdml. By 18 DPH, larvae began ingesting a formulated diet for marine finfish and were completely weaned from live feed by 25 DPH. which coincided with the onset of metamorphosis. Juveniles tolerated a range of salinity from 10-32 ppt with the highest growth rate observed at 20 ppt. Culture of larvae and juveniles on a commercial scale was successful using conditions similar to those employed in the laboratory. The findings presented here suggest the potential for commercial culture of this species in the USA.
The black sea bass Centropristis striata L. is an important species in commercial and recreational fisheries of the U.S. Atlan- tic coast from the Gulf of Maine to the northeast Gulf of Mexico (Musick and Mer- cer 1977; Fig. 1). Populations of the Atlan- tic and Gulf of Mexico coasts have been divided into two subspecies, Centropristis striutu striata and Centropristis striata me- luna, respectively. Adults and juveniles of
I Corresponding author.
the Atlantic population undertake seasonal migrations offshore as water temperatures decline in the autumn months (Musick and Mercer 1977; Vaughn et al. 1995). Fish re- main offshore of the New England coast until May and return when water tempera- tures inshore reach approximately 10 C (Caruso 1995). Reproductively, black sea bass, like groupers, are protogynous her- maphrodites developing first as females and later at 3-4 yr of age as males (Wenner et al. 1986). Spawning progresses seasonally from south to north and is initiated as early as March in North Carolina and Virginia and in June and July in New England. Adults may return to their natal spawning grounds in subsequent years (Able et al. 1995).
Black sea bass are highly sought and command a premium price in the ethnic live fish market and upscale restaurant trade. As temperate reef species, sea bass generally are inaccessible to bottom trawlers and the fishery primarily consists of capture by fish pots and angling. Demand typically exceeds supply in the northeast U.S. The market value and limited supply of black sea bass warranted evaluation of the potential of this species for commercial aquaculture. Previ- ous investigations of black sea bass were limited in scope, examining spawning and larval development and failed to assess the requisite conditions for culture (Hoff 1970; Roberts et al. 1976; Tucker 1984). In this
0 Copyright by the World Aquaculture Society ZOO0
426

REQUIREMENTS FOR BLACK SEA BASS CULTURE 427
FIGURE I . Phorograph of an adult black sea bass Centropristis striata approximately 40 ern total length.
report, we present our initial investigations of culture of black sea bass larvae and ju- veniles.
Materials and Methods Lurviculture
Larvae were obtained by strip spawning fish held in captivity for 4-9 mo. Ovulation was induced during the natural spawning season (May-June) by implantation or in- jection of 50 pg Luteinizing Hormone Re- leasing Hormone analog (LHRHa; Penin- sula Laboratories, Belmont, California, USA; Berlinsky et al. 1996). To assess con- ditions conducive to growth, larvae were initially reared at either 18 C with or with- out addition of microalgae (greenwater; Te- traselmis suecica and Isocrisis galbana; 200,000 planktodml) or at 22 C with mi- croalgae. Larvae were stocked at a density of 60 individuals& in 40-L glass tanks (three per treatment) containing Narragan- sett Bay seawater (32 ppt salinity) filtered to 15 pm and maintained under constant light (-100 lx). Daily, 15525% of the tank
volume was replaced. Six larvae from each of three replicate tanks (N = 18 per con- dition) were randomly collected every 3 d from hatching and length was determined to the nearest 0.1 mm using an image anal- ysis system (Optima, Bioscan Inc., Ed- monds, Washington, USA).
In a subsequent experiment larvae were reared in green water at 22 C and two feed- ing regimes were investigated with six rep- licate tanks each. The first regimen was identical to that reported in the Results sec- tion (Larviculture). In the second feeding regimen, larvae were maintained on rotifers and Artemia for an additional 4 d, such that transition from rotifers to Artemia and Ar- temia to formulated feed occurred on 16 and 25 DPH, respectively. Larvae were sampled 10, 15 and 25 d after active feed- ing was initiated (15, 20 and 30 DPH) for measurement of length as described above.
Feeding Trials
Early juvenile (15-30 mm). Twenty-five hatchery reared juveniles were randomly

428 BERLINSKY ET AL.
TABLE I . Composition of diets rested for sustaining survival and growth of early juvenile (15-30 mm) and sub-adult black sea bass.
Lansy Nursery Corey Clear Corey Hi-Pro Fry Biokyowa Diet. Inve Choice. Zeigler Trout Diet. Fredericton, B-Series Fry Aquaculture, Fredericton, New Grower. Gardner, New Brunswick, Diet. Chesterton, Grantsville, Utah, Brunswick, Pennsylvania,
Canada Missouri, USA USA Canada USA
Feed Size 500-700 pm 400-700 pm 500-800 pm 4 rnm 4 mm Protein (min.) 55% 55% 50% 52% 38% Lipid (min.) 17% 10% 18% 18% 12% Fiber (max.) 2% 3 70 2% 2% 4% Moisture 8-9% 5-6% 6 7 % 8-9% 8% Energy : Protein 8.7 kcaVg 7.2 kcal/g 9.2 kcal/g 9.1 kcal/g 1 1.3 kcaYg
Caloric Value 4.8 kcaVg 3.9 kcal/g 4.6 kcaYg 4.8 kcal/g 4.3 kcallg protein protein protein protein protein
assigned to each of nine 80-L tanks (three tanks per treatment) to compare growth af- forded by three commercial feeds (Bio- Kyowa, Corey and Lansy; see Table 1 for composition of feeds). Tanks were equipped with external filters, supplemental aeration and maintained under constant light (- 13 lx) and water temperature of 22 C for the duration of the trials. Approximately 10- 20% of the water volume was replaced dai- ly and uneaten feed was removed by si- phon. Concentrations of dissolved oxygen, ammonia, and nitrite were assayed weekly to monitor quality of the rearing water (Hach, Loveland, Colorado, USA). Fish were fed one of the three diets at a ration of approximately 15% body weight daily. After 4 wk, the length of each individual was measured to the nearest mrn and used to determine growth rate.
Juveniles (35-60 mm). Thirty-five fish were randomly assigned to each of six 160- L tanks (3 tanks per diet) equipped with external filters and supplemental aeration. Approximately 20% of the water volume was replaced daily and water quality was measured twice weekly as described above. Temperature was maintained at 23 C. Fol- lowing a 1-wk acclimation period, fish were manually fed three times daily either BioKyowa C (700-1000 pm) or Corey Hi- Pro (1000 pm) formulated feeds with the same composition as the BioKyowa and Corey diets in Table 1, at a ration of 9.5%
bodyweight daily. Fish were measured to the nearest mm at 30-d intervals.
Sub-adults (> I00 8). To expedite pro- gress in assessing the suitability of com- mercially available formulated feeds for sea bass, wild fish (18.4 2 0.2 cm and 105 2 2.4 g) were collected by commercial fish- erman using fish traps in coastal Rhode Is- land waters and maintained in a 1,400-L tank supplied with filtered flow-through seawater. Fish rapidly acclimated to captiv- ity and began feeding on a commercial for- mulated feed within 3 d. During a 3-wk ac- climation period the fish were fed a mixture of the two pelleted feeds (Corey Clear Choice and Zeigler Grower Diet) to be compared in the feeding trial (see Table 1 for diet composition). At the end of this pe- riod, fish were anesthetized (200 ppm 2- phenoxyethanol; Sigma Chemical Compa- ny, St. Louis, Missouri, USA) and weighed and measured to the nearest g and 0.1 cm, respectively. Twenty individuals were ran- domly assigned to each of six 230-L cir- cular tanks (3 tanks per diet) provided with filtered flow-through seawater and supple- mental aeration and maintained on a simu- lated natural photoperiod. Water tempera- ture was maintained at 15-16 C throughout the study. The fish were fed to excess five times daily using automatic feeders (Swee- ney, Welfare, Texas, USA) and any uning- ested feed was siphoned from the tank dai- ly.

REQUIREMENTS FOR BLACK SEA BASS CULTURE 429
TABLE 2. Rearing conditions and diets employed for large scale culture .f black sea bass at Great Bay Aquafarms, Portsmouth, New Hampshire, USA.
Photo- Salinity Tank vol. Feeds period
Age (DPH) C (ppt) Diet Diet size (mm) (m3) per day (h light)
0-36 19 29 1 36-45 19 29 Biomar' 0.28-0.5 1.5 10 16 45-5 1 19 29 Biomar 0.5-0.7, 0.5-0.7/0.7-1.0 mix 1.5 8 16 51-61 19 29 Biomar/Provimi* 0.5-0.7/0.7-1.0 mix, 1.5 1.5 6 16 6 1-80 19 29 Provimi 1.5, 132.0 mix 1.5 4 16 80-111 18 29 ProvimilCorey' I .5/2.0 mix. 2.0 4.5 4 14
I Biomar (Nersac, France): 56% protein; 7 or 16% fat. * Provimi (Zwolle, The Netherlands): 62% protein; 1 1 % fat.
Corey: 50% protein; 19% fat.
After 3, 6 and 9 wk, fish were anesthe- tized and total lengths and weights record- ed. At wk 14 the study was terminated and a sample of muscle tissue was collected from six fish fed each of the diets and stored at -20 C for proximate analysis. To afford the opportunity for comparison with tissues from fish feeding on natural prey items, tissue samples were collected from four fish of the same cohort collected in Rhode Island waters. Crude protein was as- sayed by Kjeldahl digestion and Nessleriza- tion (AOAC 1975). Total lipid was deter- mined by ch1oroform:ether extraction as de- scribed by Lee et al. (1996). Ash was mea- sured as the residue remaining after the oxidation of tissue at 600 C for 4 h (Web- ster et al. 1997).
EfSect of Salinity on Growth
A preliminary study was conducted to establish the minimum threshold of salinity tolerated. Ten fish, approximately 2.5 cm in length, were maintained in each of two 40- L tanks containing full salinity seawater (32 ppt) and supplemental aeration. Salinity was lowered 5 ppt daily by dilution with non-chlorinated tap water until 10 ppt was reached, at which point it was reduced fur- ther by 1-2 ppt daily. Because survival was reduced at salinities < 9 ppt, subsequent growth studies were conducted at 10, 20 and 32 ppt.
Twenty-six juveniles (3.7 2 0.2 g) were
randomly allocated into each of nine 80-L tanks (3 tanks each for 10, 20 and 32 ppt) equipped with external filters and supple- mental aeration. Temperature ranged from 21-22 C throughout the duration of the 8- wk study. Fish were fed a 1.0-mm diet (Corey Hi-Pro) at a daily ration of 3.5% body weight. Water quality was monitored and water replaced as above. At biweekly intervals fish were anesthetized and weighed to the nearest 0.1 g. Following the second inventory, the fish were transferred to 160-L tanks to compensate for the in- crease in fish density.
Large Scale Culture
Larvae also were cultured through the ju- venile stage on a larger scale at a commer- cial summer flounder hatchery (Great Bay Aquafanns, Portsmouth, New Hampshire, USA). Newly hatched larvae were reared in greenwater in a 1-m3 tank, fed a regimen of rotifers and Artemia as described above, and maintained under constant light. Dis- solved oxygen concentrations were main- tained at 90-150% saturation and the tank volume was replaced 8 times daily. For de- tails of the juvenile feeding regimens and rearing conditions, see Table 2. Fish were anesthetized as above and the weight was determined to the nearest 0.01 g on 70, 1 1 1, 264 DPH. The largest 10% of the fish in the population were retained after 264 DPH and weighed on 380 DPH.
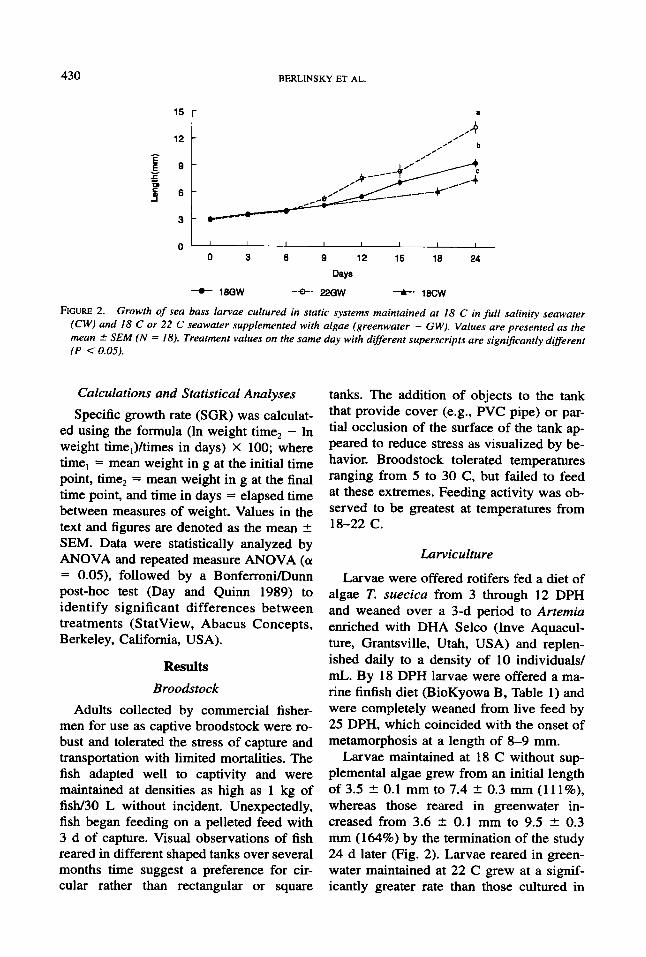
430
12
BERLINSKY ET AL.
A 00
/ b -
(P < 0.05).
Calculations and Statistical Analyses Specific growth rate (SGR) was calculat-
ed using the formula (In weight time, - In weight time,)/times in days) X 100; where time, = mean weight in g at the initial time point, time, = mean weight in g at the final time point, and time in days = elapsed time between measures of weight. Values in the text and figures are denoted as the mean 2 SEM. Data were statistically analyzed by ANOVA and repeated measure ANOVA (a = 0.05), followed by a Bonferromunn post-hoc test (Day and Quinn 1989) to identify significant differences between treatments (Statview, Abacus Concepts, Berkeley, California, USA).
Results Broodstock
Adults collected by commercial fisher- men for use as captive broodstock were ro- bust and tolerated the stress of capture and transportation with limited mortalities. The fish adapted well to captivity and were maintained at densities as high as 1 kg of fish/30 L without incident. Unexpectedly, fish began feeding on a pelleted feed with 3 d of capture. Visual observations of fish reared in different shaped tanks over several months time suggest a preference for cir- cular rather than rectangular or square
tanks. The addition of objects to the tank that provide cover (e.g., PVC pipe) or par- tial occlusion of the surface of the tank ap- peared to reduce stress as visualized by be- havior. Broodstock tolerated temperatures ranging from 5 to 30 C, but failed to feed at these extremes. Feeding activity was ob- served to be greatest at temperatures from 18-22 C.
Larviculture
Larvae were offered rotifers fed a diet of algae T. suecica from 3 through 12 DPH and weaned over a 3-d period to Artemia enriched with DHA Selco (Inve Aquacul- ture, Grantsville, Utah, USA) and replen- ished daily to a density of 10 individuals/ mL. By 18 DPH larvae were offered a ma- rine finfish diet (BioKyowa B, Table 1) and were completely weaned from live feed by 25 DPH, which coincided with the onset of metamorphosis at a length of 8-9 mm.
Larvae maintained at 18 C without sup- plemental algae grew from an initial length of 3.5 2 0.1 mm to 7.4 t 0.3 mm (1 l l%), whereas those reared in greenwater in- creased from 3.6 t 0.1 mm to 9.5 2 0.3 mm (164%) by the termination of the study 24 d later (Fig. 2). Larvae reared in green- water maintained at 22 C grew at a signif- icantly greater rate than those cultured in
REQUIREMENTS FOR BLACK SEA BASS CULTURE 43 1
200 1
0 3 6 9 11 14
Weeks
+ Corey -*- Zeigler
200 -
180 -
160 -
140 -
120 -
100 I I I I I I
0 3 6 9 11 14
Weeks
+ Corey -*- Zeigler
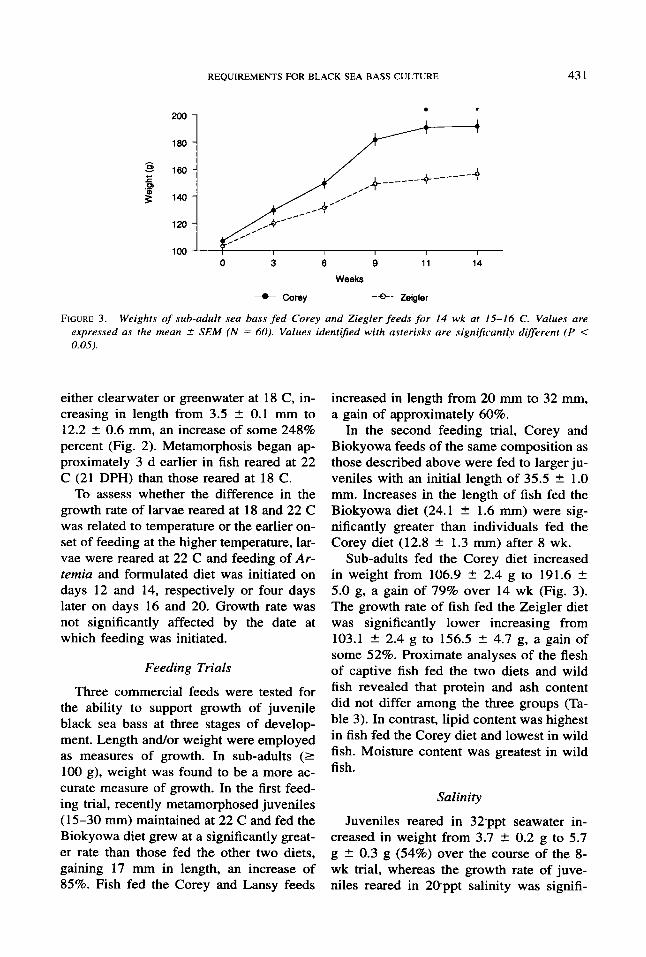
FIGURE 3 . Weights of sub-adult sea bass fed Corey and Ziegler feeds for 14 wk at 15-16 C. Values are expressed as the mean _f SEM (N = 60). Values identiJied with asterisks are significantly different ( P < 0.05).
either clearwater or greenwater at 18 C, in- creasing in length from 3.5 * 0.1 mm to 12.2 2 0.6 mm, an increase of some 248% percent (Fig. 2). Metamorphosis began ap- proximately 3 d earlier in fish reared at 22 C (21 DPH) than those reared at 18 C.
To assess whether the difference in the growth rate of larvae reared at 18 and 22 C was related to temperature or the earlier on- set of feeding at the higher temperature, lar- vae were reared at 22 C and feeding of Ar- temia and formulated diet was initiated on days 12 and 14, respectively or four days later on days 16 and 20. Growth rate was not significantly affected by the date at which feeding was initiated.
Feeding Trials
Three commercial feeds were tested for the ability to support growth of juvenile black sea bass at three stages of develop- ment. Length andor weight were employed as measures of growth. In sub-adults (2 100 g), weight was found to be a more ac- curate measure of growth. In the first feed- ing trial, recently metamorphosed juveniles (1 5-30 mm) maintained at 22 C and fed the Biokyowa diet grew at a significantly great- er rate than those fed the other two diets, gaining 17 mm in length, an increase of 85%. Fish fed the Corey and Lansy feeds
increased in length from 20 mm to 32 mm, a gain of approximately 60%.
In the second feeding trial, Corey and Biokyowa feeds of the same composition as those described above were fed to larger ju- veniles with an initial length of 35.5 2 1.0 mm. Increases in the length of fish fed the Biokyowa diet (24.1 2 1.6 mm) were sig- nificantly greater than individuals fed the Corey diet (12.8 2 1.3 mm) after 8 wk.
Sub-adults fed the Corey diet increased in weight from 106.9 2 2.4 g to 191.6 2 5.0 g, a gain of 79% over 14 wk (Fig. 3). The growth rate of fish fed the Zeigler diet was significantly lower increasing from 103.1 * 2.4 g to 156.5 * 4.7 g, a gain of some 52%. Proximate analyses of the flesh of captive fish fed the two diets and wild fish revealed that protein and ash content did not differ among the three groups (Ta- ble 3). In contrast, lipid content was highest in fish fed the Corey diet and lowest in wild fish. Moisture content was greatest in wild fish.
Salinity
Juveniles reared in 32-ppt seawater in- creased in weight from 3.7 * 0.2 g to 5.7 g 2 0.3 g (54%) over the course of the 8- wk trial, whereas the growth rate of juve- niles reared in 20-ppt salinity was signifi-

432 BERLINSKY ET AL.
TABLE 3. Proximate analyses of muscle tissue sampled from wild fish or captive sub-adult black sea bass fed two different diets. Values are expressed as the mean 2 sem (N = 4 for wild: 6 for Zeigler and Corey). Values in the same row,followed by different letters are statistically different (P < 0.05).
Component (%) Wild Zeigler Corey
Protein 21.9 ? 0.84 21.3 2 0.7 22.8 2 0.3 Lipid 1.5 2 0.2 a 3.3 ? 0.6 b 5.8 ? 0.5 c Moisture 78.4 t 0.9 a 75.1 2 0.6 b 72.7 2 0.5 c Ash 1 . 1 t 0.1 1.2 t 0.1 1.2 2 0.1
cantly higher, increasing from 3.8 5 0.2 g to 7.7 2 0.5 g (102%) (Fig. 4). Interesting- ly, the growth rate of fish maintained in 10- ppt salinity seawater was intermediate and not significantly different from that of fish reared at 32- or 20-ppt salinity.
Large Scale Culture The survival of larvae and juveniles
reared on a small commercial scale was comparable to that obtained in laboratory studies. The weight of the largest 10% of the individuals in the population increased from 0.78 g to 156 g over a period of 310 d (70-380 DPH) for a specific growth rate of 1.7% per d.
Discussion The present report describes our initial
efforts toward culture of black sea bass. As anticipated from anecdotal evidence provid- ed by local commercial fishermen, the adults were found to be robust and were
7
readily handled and transported with limit- ed mortalities. Other than maintaining en- vironmental temperature, no special condi- tions were required for successful transport of the animals. The fish readily adapted to formulated feeds and tolerated a wide range of temperatures. Together these character- istics indicate that capture and domestica- tion of broodstock is not likely to be a con- straint to culture of this species.
Growth of larvae was superior in rearing water supplemented with algae (greenwa- ter) than in non-supplemented water. The beneficial effects of greenwater observed in the present study are consistent with those reported for a number of marine species. Previous reports that cod Gadus morhua (van der Meeren 1991) and halibut Hippo- glossus hippoglossus (Reitan et al. 1991) larvae ingest substantial quantities of mi- croalgae during the first days after hatching suggest that algae may serve as an initial source of feed. Incidental ingestion of algae
0
:: 0 0 4 7 9
Weeks
-e 1oppt -+- 2oppt - S P P t
FIGLIRE 4. The effect of 10, 20 or 32 ppt seawater on growth of juvenile black sea bass. Values are denoted as the mean 2 SEM ( N = 78) and those with different superscripts on the same day are significantly different ( P < 0.05).

REQUIREMENTS FOR BLACK SEA BASS CULTURE 433
may provide a source of micro-nutrients and exogenous enzymes as well as stimu- late onset of feeding (Naas et al. 1992) and enzyme synthesis (Hjelmeland et al. 1988). Greenwater also maintains the nutritional value of rotifers and reduces the concentra- tion of nitrogenous compounds in the rear- ing water (Lavens et al. 1995).
Larvae reared at 22 C grew at a more rapid rate than those raised at 18 C. These results are consistent with a previous report in which larval black sea bass spawned from broodstock collected in the Gulf of Mexico grew to 9.0 mm in 24 d when reared at 21-23 C (Roberts et al. 1976). Water temperatures during the spawning season often times exceed those employed in the present study (22 C), suggesting that larvae might grow at a more rapid rate at even higher temperatures.
Larvae began feeding on rotifers approx- imately 3 DPH and were weaned to Artemia by 12 DPH when feeding of rotifers was discontinued. On day 18 larvae were of- fered a formulated feed and were weaned to this feed by day 25. The completion of weaning to formulated feed coincided with the onset of metamorphosis. Of note was the unusually wide mouth gape of the lar- vae which suggests that weaning to Artemia and formulated feed might be accomplished more rapidly. Feeding trials conducted with three formulated feeds in the present study suggest that the Biokyawa feed was supe- rior to the Corey and Lansy feeds for pro- moting juvenile growth.
In the present report, juvenile black sea bass fed a diet comprised of 55% protein and 10% lipid (BioKyowa) grew at signif- icantly higher rates than those fed diets with similar protein levels (50-55%) but with higher levels of lipid (2 17%). Investiga- tion of diets suitable for the European sea bass Dicentrarchus labrax demonstrated that optimal growth was obtained with pro- tein levels of 50% and lipid levels between 10-14% (Perez et al. 1997). In striped bass Morone saxatilis (Millikin 1982) and wolf- fish Anarhichas lupus (Stefanussen et al.
1993), protein levels greater than 50% pro- duced the highest rate of growth. The stud- ies presented here indicate that black sea bass are able to utilize commercially avail- able feeds containing relatively high levels of protein and lipid. Additional research is required to determine specific protein and lipid levels that will optimize growth.
Juvenile black sea bass tolerated environ- mental salinity levels as low as 10 ppt. Fur- thermore, growth rates were greater in ju- venile fish reared at 20 ppt than at 32 ppt. These findings are in agreement with pre- vious investigations demonstrating that many estuarine dependent juvenile fishes maintained in intermediate salinity water have lower metabolic rates and higher growth rates than those in full salinity sea- water (e.g., Atlantic menhaden Brevoortia ryrannus, Hettler 1976; golden line sea- bream Sparus sarba, Woo and Kelly 1995; striped bass Morone saxatilis, Otwell and Memner 1975; squeteague Cynoscion re- galis, Lankford and Targett 1994). Collec- tively, these studies support the concept that intermediate salinities minimize osmoregu- latory costs and allow increased allocation of energy to somatic growth (Febry and Lutz 1987).
The growth rate of black sea bass juve- niles was lower than anticipated. Over the course of a 310-d period of culture, juve- niles increased from an initial weight of 0.78 g to 156 g at 18 C, a specific growth rate of 1.71% per d. It appears quite prob- able that growth rates can be increased with optimization of husbandry, feed and envi- ronmental conditions.
The results of the investigations present- ed here suggest the potential of black sea bass for commercial aquaculture. The fish commands a premium price and supply from traditional capture fisheries is limited. Adults are robust and acclimate well to hatchery conditions and formulated feeds. Larvae grew best when cultured in green- water at a temperature of 22 C and juve- niles were readily weaned to formulated feeds. Furthermore, juveniles grew over a

434 BERLINSKY ET AL.
range of salinity from 10-32 ppt. Addition- al research will be required to optimize con- ditions for survival and growth of this promising species.
Acknowledgments This research was supported by Salton-
stall Kennedy grant #NA76FDO143. We would like to thank Dr. Michael Clancy for help with statistical analysis, Drs. Chong Lee and Jeeyoung Imm for assistance in conducting proximate analysis and Drs. Margie Gallagher (East Carolina Universi- ty) and TLI Smith (South Carolina Fish and Wildlife) for helpful discussions during these studies. mode Island Experiment Sta- tion Contribution #3800.
Literature Cited Able, K. W., M. P. Fahay, and G. R. Sheperd. 1995.
Early life history of black sea bass, Centropristis striata, in the mid-Atlantic Bight and a New Jer- sey estuary. Fishery Bulletin 93(3):429-445.
AOAC (Association of Omcial Analytical Chem- ists). 1975. Official methods of analysis, 12th edi- tion. Association of Official Analytical Chemists, Washington, D.C., USA.
Berlinsky, D. L., W. King V, T. I. J. Smith, R. D. Hamilton 11, J. Holloway Jr., and C. V. Sulli- van. 1996. Induced ovulation of southern flounder Puralichthys lerhosrigma using gonadotropin re- leasing hormone analogue implants. Journal of the World Aquaculture Society 27(2): 143-152.
Caruso, P. 1995. The biology and fisheries of black sea bass (Centroprisris srriara) in Massachusetts waters. Master's thesis. University of Rhode Is- land, Kingston, Rhode Island, USA.
Day, R. W. and G. P. Quinn. 1989. Comparisons of treatments after an analysis of variance in ecology. Ecological Monographs 59:433-463.
Febry, R. and P. Lutz. 1987. Energy partitioning in fish: activity related cost of osmoregulation in a euryhaline cichlid. Journal of Experimental Biol-
Hettler, W. F. 1976. Influence of temperature and sa- linity on routine metabolic rate and growth of young Atlantic menhaden. Journal of Fish Biolo- gy 8:55-65.
Hjelmeland, K., B. Pedersen, and E. Nelson. 1988. Trypsin content in intestines of hemng larvae, CZupea harengus, ingesting inert polystyrene spheres or live crustacea prey. Marine Biology 98: 33 1-335.
Hoff, F. 1970. Artificial spawning of black sea bass, Centroprisris srriara meluna Ginsburg, aided by
ogy 128163435.
chorionic gonadotrophic hormones. Florida De- partment of Natural Research, Marine Resources Laboratory, Special Scientific Report No. 25.
Lankford, T. and T. Targett. 1994. Suitability of es- tuarine nursery zones for juvenile weakfish (Cy- noscion regalis): effects of temperature and salin- ity on feeding, growth and survival. Marine Bi-
Lavens, P., P. Sorgeloos, P. Dhert, and B. Devresse. 1995. Larval foods. Pages 373-397 in N. Brom- age and R. J. Roberts, editors. Broodstock man- agement and egg and larval quality. Blackwell Science, London, England.
Lee, C. M., B. Trevino and M. Chaiyawat. 1996. A simple and rapid solvent extraction method for de- termining total lipids in fish tissue. Journal of the Association of Official Analytical Chemists Inter- national 79(2):487-492.
Millikin, M. 1982. Effects of dietary protein concen- tration on growth, feed efficiency, and body com- position of age-0 striped bass. Transactions of the American Fisheries Society 1 1 1:373-378.
Muslck, J. and L. Mercer. 1977. Seasonal distribu- tion of black sea bass, Cenrropristis striata, in the Mid-Atlantic Bight with comments on the ecology and fisheries of the species. Transactions of the American Fisheries Society 106: 12-25.
Naas, K., T. Naess, and T. Harboe. 1992. Enhanced first feeding of halibut larvae (Hippoglossus hip- poglossus L.) in green water. Aquaculture 105:
Otwell, W. S. and J. V. Merrimer. 1975. Survival and growth of juvenile striped bass, Morone SM-
arilis, in a factorial experiment with temperature, salinity and age. Transactions of the American Fisheries Society 104:560-566.
Perez, L., H. Gonzalez, M. Jover, and J. Fernandez- Carmona. 1997. Growth of European sea bass fingerlings (Dicentruchus labrax) fed extruded di- ets containing varying levels of protein, lipid and carbohydrate. Aquaculture 156: 183-193.
Reitan, K., S. B o b , and Y. Olsen. 1991. Ingestion and assimilation of microalgae in yolk sac larvae of halibut, Hippoglossus hippoglossus (L.). Pages 332-334 in F! Lavens, I? Sorgeloos, E. Jaspers and E Ollevier, editors. Larvi '91-Fish & Crustacean Larviculture Symposium. European Aquaculture Society, Special Publication 15. Gent, Belgium.
Roberts, D., B. Harpster, W. Havens, and K. Hal- Scott. 1976. Facilities and methodology for the culture of the southern sea bass (Centropristis me- lanu). Proceedings of the Seventh Annual Work- shop of the World Mariculture Society, San Diego, California, USA.
Stefanussen, D., 0. Lie and E. Moksness. 1993. Growth of juvenile wolffish (Anarhichas lupus) fed practical fish diets. Aquaculture 114: 103-1 11.
Tucker, J. W., Jr. 1984. Hormone-induced ovulation
ology 119161 1-620.
143-1 56.

REQUIREMENTS FOR BLACK SEA BASS CULTURE 435
of black sea bass and rearing of larvae. Progres- sive Fish Culturist 46(3):201-204.
van der Meeren, T. 199 I . Algae as first food for cod larvae, Gudus morhus L.: filter feeding or inges- tion by accident? Journal of Fish Biology 39:225- 237.
Vaughan, D., M. Collins, and D. Schmidt. 1995. Population characteristics of the black sea bass Cenrropristis srriuru from the southeastern U.S. Bulletin of Marine Science 56(1):250-267.
Webster, C. D., L. G. Tiu, and J. H. Tidwell. 1997. Growth and body composition of juvenile hybrid
bluegill Lepomis cyunellus X L. macrochirus fed practical diets containing various percentages of protein. Journal of the World Aquaculture Society
Wenner, C., W. Roumillat, and C. Waltz. 1986. Contributions to the life history of black sea bass, Centropristis striutu, off the Southeastern United States. Fishery Bulletin 84(3):723-741.
Woo, N. and S. Kelly. 1995. Effects of salinity and nutritional status on growth and metabolism of Spurus surbu in a closed seawater system. Aqua- culture 135:229-238.
28(3) :230-240.




















