
Investigating the role of articulatory organs and perceptual assimilation of native and non-native...
18
Investigating the Role of Articulatory Organs and Perceptual Assimilation of Native and Non-Native Fricative Place Contrasts ABSTRACT: The perceptual assimilation model (PAM; Best, C. T. [1995]. A direct realist view of cross-language speech perception. In W. Strange (Ed.), Speech perception and linguistic experience: Issues in cross-language research (pp. 171–204). Baltimore, MD: York Press.) accounts for developmental patterns of speech contrast discrimination by proposing that infants shift from untuned phonetic perception at 6 months to natively tuned perceptual assimilation at 11–12 months, but the model does not predict initial discrimination differences among contrasts. To address that issue, we evaluated the Articulatory Organ Hypothesis, which posits that consonants produced using different articulatory organs are initially easier to discriminate than those produced with the same articulatory organ. We tested English-learning 6- and 11-month-olds’ discrimination of voiceless fricative place contrasts from Nuu-Chah-Nulth (non- native) and English (native), with one within-organ and one between-organ contrast from each language. Both native and non-native contrasts were discriminated across age, suggesting that articulatory-organ differences do not influence perception of speech contrasts by young infants. The results highlight the fact that a decline in discrimination for non-native contrasts does not always occur over age. ß 2014 Wiley Periodicals, Inc. Dev Psychobiol 56: 210–227 2014. Keywords: perceptual narrowing; infants; cross-language speech perception; fricatives; perceptual assimilation model; native-language attunement; articulatory organs INTRODUCTION Classic cross-language speech perception findings with adult listeners are fairly clear: they often have difficulty categorizing and discriminating consonant contrasts not found in their native language, though there are a range of intriguing exceptions (e.g., Best, McRoberts, & Goodell, 2001; Best, McRoberts, & Sithole, 1988; Polka, 1991). Infants at 6–8 months of age, on the other hand, successfully discriminate not only most native contrasts but also many non-native ones, includ- ing those that adults of their language environment have difficulty discriminating (Aslin & Pisoni, 1980; Eilers, Gavin, & Wilson, 1979; Trehub, 1976; Werker, Gilbert, Humphrey, & Tees, 1981). Seemingly adult- like attunement to native consonant contrasts begins to Manuscript Received: 21 March 2013 Manuscript Accepted: 7 December 2013 Correspondence to: M. D. Tyler Contract grant sponsor: NIH Contract grant number: DC00403 Article first published online in Wiley Online Library (wileyonlinelibrary.com): 6 January 2014 DOI 10.1002/dev.21195 ß 2014 Wiley Periodicals, Inc. Developmental Psychobiology Michael D. Tyler 1,2 Catherine T. Best 1,3 Louis M. Goldstein 3,4 Mark Antoniou 1,5 1 MARCS Institute University of Western Sydney Penrith, New South Wales 2751, Australia E-mail: [email protected] 2 School of Social Sciences and Psychology University of Western Sydney Penrith, New South Wales, Australia 3 Haskins Laboratories New Haven, Connecticut 4 Department of Linguistics University of Southern California Los Angeles, California 5 Department of Linguistics and Modern Languages Chinese University of Hong Kong Hong Kong
-
Upload
westernsydney -
Category
Documents
-
view
0 -
download
0
Transcript of Investigating the role of articulatory organs and perceptual assimilation of native and non-native...

Investigating the Role ofArticulatory Organs andPerceptual Assimilation ofNative and Non-NativeFricative Place Contrasts
ABSTRACT: The perceptual assimilation model (PAM; Best, C. T. [1995]. Adirect realist view of cross-language speech perception. In W. Strange (Ed.),Speech perception and linguistic experience: Issues in cross-language research(pp. 171–204). Baltimore, MD: York Press.) accounts for developmental patternsof speech contrast discrimination by proposing that infants shift from untunedphonetic perception at 6 months to natively tuned perceptual assimilation at11–12 months, but the model does not predict initial discrimination differencesamong contrasts. To address that issue, we evaluated the ArticulatoryOrgan Hypothesis, which posits that consonants produced using differentarticulatory organs are initially easier to discriminate than those produced withthe same articulatory organ. We tested English-learning 6- and 11-month-olds’discrimination of voiceless fricative place contrasts from Nuu-Chah-Nulth (non-native) and English (native), with one within-organ and one between-organcontrast from each language. Both native and non-native contrasts werediscriminated across age, suggesting that articulatory-organ differences do notinfluence perception of speech contrasts by young infants. The results highlightthe fact that a decline in discrimination for non-native contrasts does not alwaysoccur over age. � 2014 Wiley Periodicals, Inc. Dev Psychobiol 56: 210–2272014.
Keywords: perceptual narrowing; infants; cross-language speech perception;fricatives; perceptual assimilation model; native-languageattunement; articulatory organs
INTRODUCTION
Classic cross-language speech perception findings with
adult listeners are fairly clear: they often have difficulty
categorizing and discriminating consonant contrasts not
found in their native language, though there are a range
of intriguing exceptions (e.g., Best, McRoberts, &
Goodell, 2001; Best, McRoberts, & Sithole, 1988;
Polka, 1991). Infants at 6–8 months of age, on the
other hand, successfully discriminate not only most
native contrasts but also many non-native ones, includ-
ing those that adults of their language environment
have difficulty discriminating (Aslin & Pisoni, 1980;
Eilers, Gavin, & Wilson, 1979; Trehub, 1976; Werker,
Gilbert, Humphrey, & Tees, 1981). Seemingly adult-
like attunement to native consonant contrasts begins to
Manuscript Received: 21 March 2013Manuscript Accepted: 7 December 2013Correspondence to: M. D. TylerContract grant sponsor: NIHContract grant number: DC00403Article first published online in Wiley Online Library
(wileyonlinelibrary.com): 6 January 2014DOI 10.1002/dev.21195 � � 2014 Wiley Periodicals, Inc.
Developmental Psychobiology
Michael D. Tyler1,2
Catherine T. Best1,3
Louis M. Goldstein3,4
Mark Antoniou1,5
1MARCS InstituteUniversity of Western Sydney
Penrith, New South Wales 2751, AustraliaE-mail: [email protected]
2School of Social Sciences and PsychologyUniversity of Western Sydney
Penrith, New South Wales, Australia
3Haskins LaboratoriesNew Haven, Connecticut
4Department of LinguisticsUniversity of Southern California
Los Angeles, California
5Department of Linguistics and ModernLanguages
Chinese University of Hong KongHong Kong

emerge by 10–12 months of age, when there is no
longer evidence for discrimination of many non-native
consonant distinctions (Werker & Lalonde, 1988;
Werker & Tees, 1984). For example, in a study that is
often cited in support of perceptual narrowing in early
development, there is a decline from 6, through 8, to 12
months of age, in the number of English-learning
infants who discriminated the Hindi unaspirated dental
versus retroflex stops / /-/ / and Nthlakampx velar
versus uvular ejectives /k0/-/q0/ (Werker & Tees, 1984).
A decline was also found for Zulu plosive versus
implosive bilabial stops /b/-/ / and voiced versus
voiceless lateral fricatives / /-/ / (Best & McRoberts,
2003). This pattern of decline is consistent with the
idea that there is perceptual narrowing in speech
perception over the first year, as a result of native
language experience.
Developmental decline in discrimination is not
observed for all non-native contrasts, however. Best,
McRoberts, LaFleur, and Silver-Isenstadt (1995) found
that English-learning infants at both 6–8 and 10–12
months of age successfully discriminated the non-native
Zulu dental versus lateral click contrast /|/-/||/, in
contrast to their replication of the earlier Werker and
Tees (1984) finding of developmental decline for
Nthlakampx velar versus uvular ejectives, which only
the 6-8-month-olds discriminated. Similarly, Best and
McRoberts (2003) found that both 6–8- and 10–12-
month-olds learning English discriminated a Tigrinya
bilabial versus alveolar ejective stop contrast /p0/-/t0/.A further complexity in the developmental story is
that some native contrasts appear to be initially difficult
to discriminate, with performance then improving over
time. For example, one study found that American-
English-learning infants’ discrimination of English /r/
and /l/ was mediocre at 6–8 months, equivalent to that
of Japanese-learning infants, but only American infants
showed improved discrimination at 10–12 months
(Kuhl et al., 2006). Another study found no evidence
for discrimination of the Tagalog (Filipino language)
alveolar-velar nasal stop contrast, /na/-/Ea/, by English-
learning infants aged 4–5, 6–8, or 10–12 months
(Narayan, Werker, & Beddor, 2010). On the other hand,
infants learning Filipino languages in which /na/-/Ea/ isa native contrast showed no evidence of discrimination
at 6–8 months, but succeeded at 10–12 months.
Additionally, the English alveolar stop versus interden-
tal fricative contrast /d/-/ð/ was discriminated equally
poorly by Canadian-English- and Canadian-French-
learning 6–8- and 10–12-month-olds (Polka, Colanto-
nio, & Sundara, 2001). Improved discrimination for the
English learners was achieved by 4 years of age
(Sundara, Polka, & Genesee, 2006), suggesting that
native-language attunement occurred between the ages
of 1 and 4 years. In contrast, the Canadian-French-
learning infants and children had poor discrimination of
the contrast at 6–8 months, 10–12 months, and 4 years,
and French-speaking adults were consistently less
accurate than the English-speaking adults. Moreover,
one study has even found a developmental decline for a
native contrast. Best and McRoberts (2003) found that
although both 6–8- and 10–12-month-olds were able to
discriminate English /s/-/z/, there was a significant
decline in the degree of discrimination at the older age.
On the surface, this latter finding seems to pose a
challenge for notions of perceptual narrowing.
Developmental theories of speech perception thus
face a difficult set of challenges. They must account
for why discrimination of both native and non-native
contrasts can initially be successful or unsuccessful,
may or may not improve or decline over time, and
why performance on native contrasts appears to follow
different developmental trajectories for different con-
trasts. But theoretical accounts can be difficult to
compare because they differ in focus, that is, one
account may focus on initial perceptual sensitivities
and another on developmental changes, and both
of these aspects might be investigated from the
perspective of native or non-native speech perception,
possibly using different procedures. We therefore
have concentrated on one model in the design of the
present study, the perceptual assimilation model
(PAM; Best, 1994a,b, 1995), but we will evaluate our
results in light of alternative models in the General
Discussion (the Native Language Magnet Theory-
Expanded, Kuhl et al., 2008; and Processing Rich
Information from Multi-dimensional Interactive Repre-
sentations, Curtin, Byers-Heinlein, & Werker, 2011;
Werker & Curtin, 2005). The present article has three
main goals: (1) To outline how PAM accounts for both
perceptual narrowing to native speech contrasts as well
as for variations in infants’ success at discriminating
different non-native contrasts at different ages. (2) To
test whether the articulatory organ hypothesis (AOH),
an expansion on PAM described in Best and McRo-
berts (2003), predicts infants’ initial discrimination of
native and non-native contrasts. (3) To provide much
needed data on infant discrimination of naturally
produced native and non-native fricative place of
articulation contrasts.
We follow Aslin and Pisoni (1980) in applying to
speech perception Gottlieb’s (1976) framework for
describing the possible trajectories that perceptual
abilities might display developmentally as a function of
language experience. At birth, discrimination of pho-
netic contrasts could range from perfect to poor. Those
discriminated perfectly at birth may undergo mainte-
nance with language experience or decline through lack
Developmental Psychobiology Infant Perception of Native and Non-Native Fricatives 211

of experience.1 Intermediate levels of initial discrimina-
tion can undergo facilitation (further improvement) or
maintenance with experience, or can decline through
lack of experience. Finally, poor discrimination will
remain unaffected in the absence of relevant experi-
ence, or will undergo induction (improvement) with
relevant experience [see Figure 5.4 in Aslin and Pisoni
(1980), for a depiction of this framework].
For PAM (Best, 1994a, b, 1995), the objects of
speech perception are articulatory gestures. In line with
Gibson’s (1979) ecological theory of perception, the
acoustic signal is thought to carry information about
those articulatory gestures, but information may also
come from other modalities such as vision or touch.
Although infants are exposed to low-frequency compo-
nents of maternal speech in the womb, which may result
in some prenatal attunement for prosodic properties
(e.g., intonation and rhythm) and possibly for some
vowels, infants’ initial consonant discrimination abilities
are generally unaffected by experience with their native
language environment. Successes or failures in phoneme
discrimination at a very early age therefore reflect
general pre-phonetic perceptual abilities. As they gain
experience with the ambient language environment,
infants begin to recognize that constellations of similar
articulatory gestures contrast with other constellations
and they begin to recognize the higher-order invariants
that characterize native phonetic categories.2 Later in
development (around 17–19 months), tuning of phonetic
categories is driven additionally by phonological catego-
ries that arise as a result of an expanding vocabulary
(see Best, Tyler, Gooding, Orlando, & Quann, 2009;
Mulak & Best, 2013; Studdert-Kennedy, 1991). Thus,
from a PAM perspective, discrimination of speech in
the first year of life is determined first by general pre-
phonetic perceptual abilities, and then additionally by
the recognition of native phonetic categories through
language-specific tuning, followed by emerging phono-
logical categories through lexical development. Impor-
tantly, it is not necessary for the infant to have had
experience with the specific phonetic characteristics of a
non-native phone for it to be perceived as an instance of
a native category. The non-native phone simply needs to
share some higher-order invariant information with the
native phonetic category.
As infant perception develops, it will be driven less
by pre-phonetic perceptual abilities and more by
phonetic (and phonological) categories. Perception via
those categories is highly efficient because they are
specially tuned to the properties of the ambient speech
environment. Phonetic categories are the result of
infants tuning in to those speech characteristics that are
necessary to distinguish them from other native speech
categories (i.e., higher-order invariants) and tuning out
within-category phonetic variability. Thus, while pho-
netic categories provide the infant with improved
speech perception efficiency, within-category differen-
ces become more difficult to detect. It is important to
note, though, that sensitivity to within-category differ-
ences is not lost altogether, according to PAM, and
they should remain detectable under the right condi-
tions.
Once an infant has developed phonetic categories, if
the phones of a contrast fall in regions of phonetic
space that have undergone language-specific tuning,
then the infant’s discrimination is likely to be affected,
either positively or negatively. If both phones in a
contrast (native or non-native) fall entirely within a
single developing native phonetic category then dis-
crimination will be poorer than if each phone falls
within a different native category or if one or the other
phone falls in a region of phonetic space that has not
(yet) undergone language-specific tuning.
In adult studies that have examined PAM, the
influence of attunement to the native language is
gauged by asking participants to categorize non-native
phones in terms of native speech categories and to rate
their goodness of fit. Patterns of assimilation of non-
native phones to native categories are then established
from those data and those assimilation patterns are
used to test PAM predictions for discrimination of
non-native contrasts (see, e.g., Best et al., 2001;
1Aslin and Pisoni (1980) used the term loss instead of
decline. We use the term decline because such contrasts may be
discriminable in adulthood under certain task conditions (for a
discussion, see Werker, 1995).2The term phonetic category is used widely in the literature,
but it is not clear whether all authors use the term in an
equivalent fashion, especially with respect to non-native speech
perception. In particular, some theorists appear to consider
phonetic categories to be innate and, therefore, that the infant’s
job is to discover which of those categories are relevant for the
language of their environment [for a lively debate on this issue,
see Nittrouer (2001), a comment by Aslin, Werker, and Morgan
(2002), and reply by Nittrouer (2002)]. In contrast, we conceive
of a universal phonetic space that is multidimensional and
encompasses all possible articulatory configurations of the
world’s languages (other theorists may prefer to consider acoustic
cues rather than articulatory configurations, but this does not alter
the general principle). Phonetic categories are not innate, but
rather emerge as the young infant carves out regions of phonetic
space that are utilized in the native-language. Initially those
native phonetic categories may be broad, such that there would
be little differentiation between good and poor exemplars, which
would then undergo additional fine-tuning throughout develop-
ment. Several adjacent points in the phonetic space may therefore
fall within a single phonetic category for an infant learning one
language, or at the intersection between two phonetic categories
for another language, or even in a region of phonetic space that
has not undergone language-specific tuning, for an infant learning
yet a third language.
212 Tyler et al. Developmental Psychobiology

Harnsberger, 2001; Strange et al., 1998; Tyler, Best,
Faber, & Levitt, in press). The challenge for predicting
infant discrimination from a PAM perspective is to
observe (and to predict) the point in development when
native-language phonetic tuning begins to exert an
influence on perception. This appears to occur, for most
contrasts, at some time between 6 and 12 months of
age. It is important to note that a shift to natively tuned
speech perception may occur without an observable
change in discrimination, or even with a change that
appears to be counterintuitive. For example, the
decline in discrimination for English /s/-/z/ (Best &
McRoberts, 2003) may reflect a shift from reliance on
pre-phonetic differences between stimuli at 6–8 months
to reliance on rudimentary natively tuned phonetic
categories at 10–12 months.
A shift to natively tuned perception would result in
a decline in discrimination for a pair of non-native
phones that are both assimilated to the same native
phonetic category. However, if they are assimilated to
two different native phonetic categories then discrimi-
nation could appear to be maintained at 12 months of
age, and might even improve between 6 and 12 months.
By this account, the shift to natively tuned perception
certainly does include many examples that are consis-
tent with the notion of perceptual narrowing, but it
might not always result in a decline in discrimination
for non-native phonemes.
Previous descriptions of PAM (e.g., Best, 1994a,
b, 1995) listed a range of ways that non-native phones
might be assimilated to the native phonological system
of adult perceivers (e.g., single category, category
goodness, two category, uncategorized–categorized, and
non-assimilable–non-assimilable). However, if the pho-
nological system does not develop until 17–19 months,
some of those assimilation types may not apply at
10–12 months.3 The question of interest in the present
study, for infants prior to the development of phono-
logical categories, is whether a non-native phone is
assimilated to a natively-tuned (i.e., a phonetic catego-
ry) or an untuned region of phonetic space. This results
in four possible non-native contrast assimilation types
for infants who have transitioned to natively tuned
perception: (1) Single-category assimilation occurs
when both phones in a non-native contrast are assimi-
lated to a single developing native phonetic category
and discrimination is predicted to be poor. (2) Two-
category assimilation occurs when each contrasting
non-native phone is assimilated to a different native
phonetic category and discrimination should be very
good. (3) Untuned–tuned assimilation occurs when one
non-native phone is assimilated to a developing native
phonetic category and the other falls in a region of
phonetic space that is not (yet) utilized for speech in
the native language, and discrimination should be very
good. (4) Untuned–untuned assimilation refers to the
case when both phones fall in an untuned region, and
in that case discrimination would depend on the infant’s
pre-phonetic perceptual abilities.
From this extension of PAM principles to infant
assimilation types, it is now possible to detail how
discrimination of non-native phones should develop as
a function of initial sensitivities and the contrast
assimilation type when infants transition to natively
tuned perception. The predictions are presented in
Table 1. A decline in discrimination over development
would be observed when a contrast that was initially
discriminated successfully becomes a single-category
assimilation after the development of phonetic catego-
ries. Contrasts that infants initially failed to discrimi-
nate would remain poor for single-category and
untuned–untuned assimilations, but would improve for
two-category and untuned–tuned assimilations. Initial
discrimination levels (from poor to good) would simply
be maintained for untuned–untuned contrasts, because
the non-native phones fall in a region of phonetic space
that has not undergone native-language tuning. Finally,
non-native contrasts that are initially discriminated well
will be maintained for two-category and untuned–tuned
assimilations, even though older infants may have
shifted to using higher-order information to achieve the
same performance.
PAM was devised to account for attunement to the
native language so it does not make clear predictions
about which contrasts will be easy or difficult to
discriminate at the earliest, that is, pre-phonetic period
of acquisition. Given the direct-realist foundations of
PAM, one possible source of predictions about this is
the AOH (Goldstein & Fowler, 2003; Studdert-Kennedy
& Goldstein, 2003), which was devised to explain
differences in contrast discrimination by young infants,
3For readers familiar with PAM, we have omitted the
category-goodness assimilation type because we have assumed
that phonetic categories are likely to be “broad and flat” at the
end of the first year of life, that is, to have little perceptual
differentiation within a category (for a discussion, see
Best, 1994a). Infants are unlikely to recognize differences in
category goodness until they have begun to develop phonological
categories between 15 and 19 months of age (see Best et al.,
2009). Furthermore, we propose that there is no difference
between uncategorized and non-assimilable phones for infants.
The difference between those two assimilation types is simply
whether they are heard as speech (i.e., fall inside the phonologi-
cal space) or as non-speech (see Best et al., 1988). We suggest
that the distinction between speech and non-speech might
develop, like category goodness, with the establishment of
phonological categories later in development. Alternatively, it
might appear a bit earlier, with the establishment of native
phonetic categories.
Developmental Psychobiology Infant Perception of Native and Non-Native Fricatives 213

based on the principles of articulatory phonology
(Browman & Goldstein, 1990, 1992, 1995, 2000). The
articulatory organs are the lips, tongue tip, tongue
body, tongue root, velum, and larynx (specifically, the
glottis: the area circumscribed by the vocal cords).
These constricting organs are distinct and independent,
because the constrictions formed by these organs are
made with different parts of the vocal anatomy and a
constriction can be formed with one organ without
necessarily producing a constriction in another. In the
articulatory phonology view, the traditional phonetic
dimensions of place and manner of articulation corre-
spond, respectively, to the location and degree of a
constriction achieved by a specific articulatory organ.
Based on the infant lip and tongue motion imitation
studies of Meltzoff and Moore (1977), the AOH
proposes that infants can identify articulatory organs
from birth and, therefore, that contrasts involving
separate organs (between-organ contrasts; e.g., /b/ [lips]
vs. /d/ [tongue tip]) should be discriminated easily. In
contrast, those involving the same articulatory organ,
with differences in constriction degree (e.g., tongue tip
in /t/ vs. /s/), constriction location (e.g., /s/ vs. /u/), orphasing between organs (e.g., /t/ vs. /d/, phasing
between tongue tip and glottal gestures), require experi-
ence with language-specific distributions of those
parameters and should be much more difficult for
young infants to discriminate. Therefore, the AOH
would predict that initial discrimination of both native
and non-native contrasts is determined by whether the
contrast is between- or within-organ. One clear predic-
tion that can be made from applying AOH principles to
discrimination by young infants is that they should
never fail to discriminate a between-organ distinction,
even if it does not occur in the ambient language
environment. Furthermore, in general, between-organ
distinctions should be detected more easily than within-
organ distinctions prior to the influence of native-
language tuning.
Best and McRoberts (2003) tested whether discrimi-
nation of non-native within-organ contrasts would show
a developmental decline earlier and more dramatically
than non-native between-organ contrasts. They found a
decline in discrimination from 6–8 to 10–12 months on
discrimination of a series of within-organ glottal con-
trasts (plosive/implosive, plosive/ejective, and voiced/
voiceless) but no decline for a between-organ non-
native place of articulation contrast (lips vs. tongue
tip). Their finding of significantly poorer discrimination
at 10–12 than 6–8 months for native voiceless-voiced
/s/-/z/ was interpreted to suggest that organ effects may
not be restricted to non-native contrasts. Note, however,
that Kuhl et al. (2006) instead found an improvement
from 6–8 to 10–12 months in American-English-
learning infants’ discrimination of the primarily within-
organ /r/-/l/ contrast, which is not compatible with the
idea that discrimination of within-organ contrasts
declines even for native speech. In light of the Kuhl
et al. finding, we suggest here that the decline for
English /s/-/z/ observed by Best and McRoberts (2003)
may be due to a transition from untuned to natively
tuned perception. The focus of the present article is on
whether articulatory-organ-based differences can pre-
dict infants’ initial discrimination of non-native con-
trasts.
To summarize, PAM makes clear predictions about
how discrimination might change over development as
a result of native-language tuning. In principle, PAM
can account for the developmental trajectories observed
in the literature, but validation is difficult because it
requires information, such as categorization with good-
ness ratings, that is not accessible using current infant
research techniques. Firstly, it is necessary to know
whether the infant has transitioned to natively tuned
speech perception and, secondly, to know how the
contrast of interest has assimilated to the developing
native system. Tests of assimilation with adults can
provide some guidance, but there is no guarantee that
assimilation patterns at 12 months mirror those of the
mature adult perceiver. Indeed, by our reasoning they
would not be entirely parallel at those two ages because
the adult operates in a phonological domain that the
Table 1. PAM Predictions for Changes in Non-Native Contrast Discrimination Over Development as a Function of (i)
Discriminability Prior To Natively Tuned Speech Perception, and (ii) Assimilation to Developing Phonetic Categories After
Natively-Tuned Speech Perception (But Prior To the Development of a Phonological System)
After the Onset of Natively-Tuned Speech Perception
Single-Category Two-Category/Untuned–Tuned Untuned–Untuned
Pre-phonetic speech perception
Poor Maintenancea Induction Maintenance
Moderate Decline Facilitation Maintenance
Good Decline Maintenance Maintenance
aThat is, the contrast continues to be discriminated poorly.
214 Tyler et al. Developmental Psychobiology

12-month-old has not yet reached. In contrast, PAM
does not provide any concrete predictions about
whether particular contrasts might initially be perceived
well or poorly, but the AOH fills that gap, as it does
provide a clear prediction of moderate-to-good initial
discrimination of between-organ contrasts.
To test AOH predictions for initial discrimination,
we required a set of stimuli that would allow us to
compare discrimination of between- and within-organ
contrasts. Furthermore, to test PAM-based developmen-
tal predictions, it was necessary to investigate discrimi-
nation of both native and non-native contrasts. The
contrasts tested in Best and McRoberts (2003) involved
the same oral constriction but differed in a gesture of
the glottis (voiced versus voiceless lateral fricatives
/ /-/ /, voiceless aspirated versus ejective velar stops
/kha/-/k’a/, and unaspirated bilabial plosive versus
implosive stops /pu/-/ u/). Although they are certainly
within-organ contrasts, those particular contrasts do not
provide a simple test of organ-based predictions
because they involve coordination (phasing) of a
laryngeal gesture with a constriction made by a
different articulator (tongue tip, lips) at a different
location in the vocal tract. The English /r/-/l/ contrast,
tested by Kuhl et al. (2006), also involves phasing
between different organs. Although the constriction is
primarily within-organ, it also does not provide a
simple test because /r/ and /l/ are complex articulations
that involve secondary constrictions with other organs.
Specifically, English /r/ involves bunching of the tongue
body, lip rounding, and lip protrusion (Boyce & Espy-
Wilson, 1997; Guenther et al., 1999), whereas English
/l/ involves tongue tip contact (dental-alveolar), tongue
root retraction, and no lip rounding (Stone &
Lundberg, 1996). Finally, English /d/-/ð/ and /b/-/v/
(Polka et al., 2001) are within-organ and they are much
less complex than /r/ and /l/, but they do not provide a
minimal test of within-organ contrast perception be-
cause they each involve differences in both constriction
degree (stop/complete closure vs. fricative/critical clo-
sure) and location (alveolar ridge or lips vs. teeth).
For a basic test of organ-based predictions we
sought between-organ contrasts versus within-organ
contrasts that only involved a single active articulator,
that is, did not involve coordination between different
articulatory organs. We selected contrasts between
voiceless fricative pairs differing in constriction loca-
tion (within-organ) and/or articulatory organ (between-
organ). An additional advantage of selecting voiceless
fricative contrasts is that they appear to be more
difficult for infants to discriminate than other contrasts
such as stop-place distinctions (Eilers, Wilson, &
Moore, 1977; Holmberg et al., 1977 as cited in
Kuhl, 1980). Using such contrasts therefore provides a
situation in which young infants are most likely to have
difficulty discriminating both within- and between-
organ contrasts, in both the native and in non-native
languages. The choice of fricative contrasts for this
study will also provide much needed data on the
development of fricative place (constriction location)
perception in infants between 6 and 12 months of age.
The majority of studies on infant perception of non-
native consonant place contrasts have focused on stop
consonants and glides. Few studies have focused on
fricative place distinctions, not all of them assessing
developmental changes, and the findings have been
mixed regarding infants’ success on these contrasts
(e.g., Cristia, McGuire, Seidl, & Francis, 2011; Eilers
et al., 1977; Levitt, Jusczyk, Murray, & Carden, 1988;
Nittrouer, 2001), as we describe in the context of our
experimental stimuli later.
We tested discrimination of non-native fricative
place contrasts in Experiment 1 and of native fricative
place contrasts in Experiment 2. In each experiment
there were two groups of infants: (1) 6-month-olds,
who should not yet have undergone much, if any,
native-language tuning for consonant perception, and
(2) 11-month-olds, who should have begun to develop
native phonetic categories for consonants.
EXPERIMENT 1: DISCRIMINATION OF NON-NATIVE VOICELESS FRICATIVE CONTRASTS
The aim of our study was to test whether articulatory-
organ-based predictions can account for 6-month-olds’
discrimination and whether PAM can account for the
change in discrimination over development to
12 months of age. We began our investigation with
discrimination of non-native contrasts because they
allowed an exposure-free context for evaluating the
influence of articulatory organs, as the non-native
fricatives we selected for this experiment do not occur
in English even as allophones. Experiment 2 examines
organ effects from the complementary vantage point,
by using native fricatives to assess native-language
tuning due to exposure in the ambient input.
In Experiment 1, we minimized the influence of
native-language tuning by selecting between- and
within-organ non-native voiceless fricative place con-
trasts from an untuned region of phonetic space—
untuned–untuned assimilation types (see Tab. 1). There
are no fricative place contrasts in English that are
produced with the tongue body or the tongue body
versus tongue root. Thus, a set of fricatives produced
with those organs would be least likely to have been
influenced by native-language tuning, because they
would not overlap with phonetic distributions of any
Developmental Psychobiology Infant Perception of Native and Non-Native Fricatives 215
native English fricative (or for that matter, stop or
approximant) place of articulation contrasts.
The target language we selected for this purpose was
Nuu-Chah-Nulth, which has a set of voiceless fricatives
that meet our criteria (Maddieson, 1984). Nuu-Chah-
Nulth is a Wakashan language spoken on Vancouver
Island, Canada, which has velar /x/ and uvular /x/fricatives, both produced with the tongue body but at
different constriction locations, and a pharyngeal /£/fricative, produced with the tongue root (Carlson,
Esling, & Fraser, 2001). We used the between-
organ /x/-/£/ and within-organ /x/-/x/ contrasts to test
6–8- and 10–12-month-old Australian-English-learning
infants’ discrimination. The inclusion of the uvular
fricative /x/ in both contrasts provides a crucial added
control—it is the base segment that serves as a
comparator for both within-organ and between-organ
change in a repeated measures design. To maximize the
opportunity for infants to detect differences between
the control and test items, we used a stimulus-alternat-
ing preference paradigm (Best & Jones, 1998; Houston,
Horn, Qi, Ting, & Gao, 2007; Mattock, Molnar, Polka,
& Burnham, 2008), in which the test trials consisted of
tokens of the target fricative alternating with tokens of
the habituation stimulus.
Method
Participants. Forty-eight English-learning infants from
Sydney, Australia, formed the sample for Experiment 1.
Half were aged 6 months, Mage¼ 28.8 weeks, range:
24.6–31.9, the other half 11 months, Mage¼ 48.6
weeks, range: 45.3–52.6. There were 13 females and 11
males in each age group. An additional 28 infants were
tested but their data were eliminated from the sample
due to equipment failure (n¼ 6), failure to habituate
(n¼ 1), exposure to a language other than English
(n¼ 1), having fixation times shorter than 1 s on any
individual trial (n¼ 6; Cristia et al., 2011), or spontane-
ous recovery to the control stimulus (fixation duration
on the second control trial that was more twice as long
as the first control trial [i.e., the last habituation trial];
n¼ 14), indicating they had not actually fully habituat-
ed to the control stimuli (Kitamura, Panneton, &
Best, 2013).
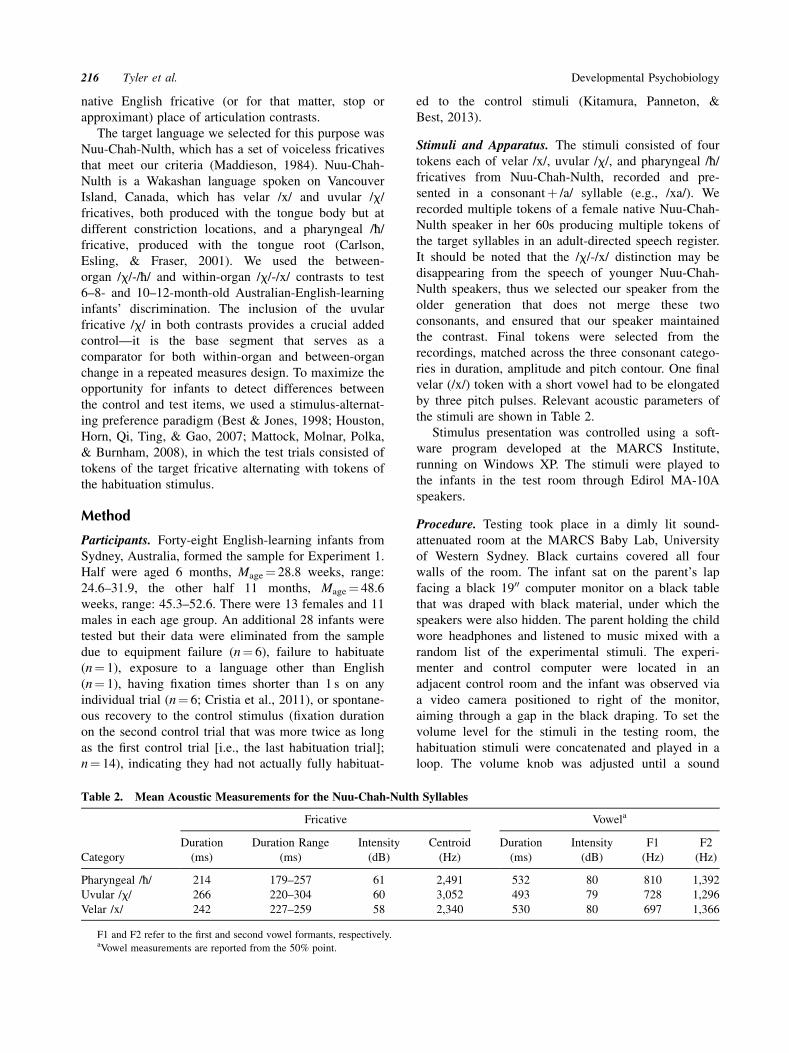
Stimuli and Apparatus. The stimuli consisted of four
tokens each of velar /x/, uvular /x/, and pharyngeal /£/fricatives from Nuu-Chah-Nulth, recorded and pre-
sented in a consonantþ /a/ syllable (e.g., /xa/). We
recorded multiple tokens of a female native Nuu-Chah-
Nulth speaker in her 60s producing multiple tokens of
the target syllables in an adult-directed speech register.
It should be noted that the /x/-/x/ distinction may be
disappearing from the speech of younger Nuu-Chah-
Nulth speakers, thus we selected our speaker from the
older generation that does not merge these two
consonants, and ensured that our speaker maintained
the contrast. Final tokens were selected from the
recordings, matched across the three consonant catego-
ries in duration, amplitude and pitch contour. One final
velar (/x/) token with a short vowel had to be elongated
by three pitch pulses. Relevant acoustic parameters of
the stimuli are shown in Table 2.
Stimulus presentation was controlled using a soft-
ware program developed at the MARCS Institute,
running on Windows XP. The stimuli were played to
the infants in the test room through Edirol MA-10A
speakers.
Procedure. Testing took place in a dimly lit sound-
attenuated room at the MARCS Baby Lab, University
of Western Sydney. Black curtains covered all four
walls of the room. The infant sat on the parent’s lap
facing a black 1900 computer monitor on a black table
that was draped with black material, under which the
speakers were also hidden. The parent holding the child
wore headphones and listened to music mixed with a
random list of the experimental stimuli. The experi-
menter and control computer were located in an
adjacent control room and the infant was observed via
a video camera positioned to right of the monitor,
aiming through a gap in the black draping. To set the
volume level for the stimuli in the testing room, the
habituation stimuli were concatenated and played in a
loop. The volume knob was adjusted until a sound
Table 2. Mean Acoustic Measurements for the Nuu-Chah-Nulth Syllables
Category
Fricative Vowela
Duration
(ms)
Duration Range
(ms)
Intensity
(dB)
Centroid
(Hz)
Duration
(ms)
Intensity
(dB)
F1
(Hz)
F2
(Hz)
Pharyngeal /£/ 214 179–257 61 2,491 532 80 810 1,392
Uvular /x/ 266 220–304 60 3,052 493 79 728 1,296
Velar /x/ 242 227–259 58 2,340 530 80 697 1,366
F1 and F2 refer to the first and second vowel formants, respectively.aVowel measurements are reported from the 50% point.
216 Tyler et al. Developmental Psychobiology

pressure level meter at the infants’ location registered
between 65 and 70 dB (A-weighting).
We used an infant-controlled visual fixation habitu-
ation procedure (see Best et al., 1988; Miller, 1983)
with stimulus-alternating test trials. A trial began when
the infant fixated on a flashing checkerboard pattern
(on the monitor facing the infant) for more than
400ms, at which point the checkerboard became
stationary and the auditory stimuli played. The experi-
menter pressed the space bar whenever the infant
fixated on the computer monitor, which was registered
by the experimental control program that also played
out the audio stimuli. If the infant looked away, the
stimuli stopped playing (after finishing any token that
had already begun playing) until the infant looked
back again. The trial ended when the infant looked
away for more than 2 s or if the total trial duration
reached 30 s.
Habituation trials consisted of all four tokens of the
uvular fricative /x/ presented in random order, with an
interstimulus interval of 1 s, and these trials were
repeated until the infant’s fixation time was lower than
a habituation criterion on two consecutive trials. The
habituation criterion was calculated as half the average
of the first two trials of the session. If the fixation time
was less than 1 s, the trial was repeated, and if the
habituation criterion was not reached by 20 trials, the
session was aborted. Test trials consisted of tokens of
the habituation stimuli alternating with tokens of the
test stimuli. These test trials were interleaved with
control trials, which were non-alternating, as they were
identical to the habituation trials. There were two
blocks of four test trials, with one between-organ and
one within-organ test trial and a control trial for each.
The last habituation trial was used as the control for
the first test trial. As there were two types of critical
test trial, between- versus within-organ, the presenta-
tion order was counterbalanced across infants; either
(1) control, between organ, control, within organ,
control, between organ, control, within organ, or (2)
control, within organ, control, between organ, control,
within organ, control, between organ. Mean fixation
times for each between- or within-organ test trial were
compared to the mean fixation time of the control trial
that immediately preceded it. This resulted in four
control–test pairs for each infant.
Results
To ascertain any effects of the counterbalancing factors
on the experimental variable, we conducted an initial
omnibus analysis of variance (ANOVA) on the data
using a 2� 2� (2)� (2)� (2) design with age (6- vs.
11-month-olds) and presentation order (counterbalanc-
ing factor: between organ followed by within organ vs.
within organ followed by between organ) as between-
subjects factors, and trial type (test vs. control), organ
(between vs. within), and block (first vs. second
presentation of a each control–test trial pair) as within-
subjects factors. This omnibus ANOVA found no
significant main effect or two-way interactions involv-
ing the presentation order counterbalancing factor;
however, one three-way interaction was significant,
indicating it mitigated the interactions involving two of
the experimental factors: Block�Presentation Order�Organ, F(1,44)¼ 6.21, p¼ .02, hp
2¼ .12. This interac-
tion indicates that, collapsed across age, combined
fixation times for test and control trials are generally
longer for the organ type that was presented first,
although that pattern reverses in the second block only
for the presentation order in which within organ is
followed by between organ. As this interaction does not
involve the trial type factor it is unlikely to have
affected the main results.
As for the experimental factors, there was not a
main effect of trial type but, importantly, there was a
significant interaction between block and trial type,
F(1,44)¼ 12.62, p¼ .001, hp2¼ .22. For the first block
the mean fixation times were longer for test than
control trials, Mtest¼ 7.49 s (SE¼ 0.69), Mcontrol¼ 5.03
s (SE¼ 0.41), but in the second block the pattern was
reversed, Mtest¼ 6.22 s (SE¼ 0.72), Mcontrol¼ 7.41 s
(SE¼ 0.96). This suggests that the infants either recov-
ered from habituation in the second block, or they
began to transition from a novelty to a familiarity
preference (see Cohen, 2004).
To investigate further whether infants’ fixation times
to control trials increased across the experiment, we
rearranged the data by trial order rather than factor.
Those data are presented in Figure 1. Note that the
columns representing test items are collapsed across
between- and within-organ trials, as they vary by
presentation order. The pattern of data in the figure
suggests that control-trial fixation times increase over
the course of the session, whereas test-trial fixation
times generally decrease. To test this, we subjected the
data to a 2� (4)� (2) ANOVA with age (6- vs. 11-
month-olds) as a between-subjects factor, and trial
position (1, 2, 3, or 4) and trial type (control vs. test)
as within-subjects factors. The pattern of increasing
fixation times for control trials and decreasing fixation
times for test trials was confirmed by a two-way
interaction between trial position and trial type,
F(3,138)¼ 5.82, p¼ .001, hp2¼ .11. This rearrange-
ment of the data also allows us to see whether we
introduced a confound by using the last habituation
trial as the first control trial in the first ANOVA. In a
recent review of habituation procedures, Oakes (2010)
Developmental Psychobiology Infant Perception of Native and Non-Native Fricatives 217

pointed out that this practice may overestimate dishabi-
tuation to test trials because the last habituation trial is
low by design. From Figure 1, however, it can be seen
that although infants in our study show increased
fixation in the first test trial relative to the first control
trial (last habituation trial), in the second control trial
(following test trial 1) they return to the same level as
or lower than the first. It is only in the second block of
four trials that fixations to control trials increase and
those to test trials decrease, relative to baseline,
suggestive of a transition to a familiarity preference.
On the basis of this analysis, we chose to analyze the
data again, but using data from only the first block,
which should be the most sensitive to discrimination
abilities and less likely than the second block to exhibit
fatigue effects.
The results of that more focused 2� 2� (2)� (2)
ANOVA revealed a main effect of trial type,
F(1,44)¼ 18.01, p< .001, hp2¼ .29, such that infants
oriented longer overall to test trials than control trials.
Mean fixation times are presented in Figure 2. There
was also a significant Presentation Order�Organ
interaction, F(1,44)¼ 4.70, p¼ .036, hp2¼ .10, such
that mean fixation times were longer overall for test/
control trial pairs where the test trial involved the organ
difference that was presented first (e.g., for the within
organ! between organ presentation order, participants
oriented longer toward within-organ test/control trials
than to between-organ test/control trials). This effect
can be seen most clearly in Figure 1. For both 6- and
11-month-olds, the combined fixation times for test 1
and control 1 are longer than for test 2 and control 2.
Discussion
Our initial omnibus ANOVA, using two blocks of four
trials, showed a significant interaction between trial
type and block, such that infants apparently changed
their preference from looking longer to test than control
trials in Block 1, to looking longer at control than test
trials in Block 2. The reason for this shift is not clear,
but there are two reasonable possible explanations.
First, infants may have tired of the task during the
course of the test phase and preferred to listen to the
more familiar control trials that were the habituation
stimuli (Cohen, 2004), or they may have simply
recovered from habituation by block 2 due to the length
of the test phase. Whatever the source for the shift in
preference, the interaction showed that infant behavior
had changed by the second half of the test phase.
Therefore, we re-analyzed without the block factor,
using only the first and most informative block of the
test phase, for which infants showed the expected
preference for trials containing novel stimuli inter-
spersed with the habituation stimuli.
Results of that more focused ANOVA indicate that
infants looked longer to test trials than control trials in
the first exposure to the two novel test stimuli in the
first half of the test phase, suggesting that infants at
both 6 and 11 months can discriminate both the
between-organ /x/-/£/ and within-organ /x/-/x/ con-
trasts. Therefore, there was no evidence in this experi-
ment that 6-month-olds discriminated between-organ
contrasts more successfully than within-organ contrasts,
as had been predicted by the AOH.
0
1
2
3
4
5
6
7
8
9
10
11
12
6 month-olds 11 month-olds
Mea
n fix
atio
n tim
e (s
)
Control 1
Test 1
Control 2
Test 2
Control 3
Test 3
Control 4
Test 4
FIGURE 1 Mean fixation durations for 6- and 11-month-olds for each trial in the order that it
was presented in Experiment 1. Note that the test trials are averaged across the organ factor.
The control trials (black columns) show a generally increasing pattern of fixation durations and
the test trials (gray columns) a generally decreasing one. Error bars represent standard errors
of the mean.
218 Tyler et al. Developmental Psychobiology

Before explaining how PAM might account for
the observed developmental pattern between 6 and
11 months, it is useful to consider the results of an
adult perceptual assimilation study we conducted with
the Nuu-Chah-Nulth fricatives. In that study we found
that the voiceless pharyngeal fricative /£/ and the
voiceless uvular fricative /x/ were both assimilated to
English /h/ (Tyler & Best, 2010). There was a signifi-
cant difference in goodness-of-fit to /h/, which means
that the between-organ contrast was a category-good-
ness assimilation for the adults. The voiceless velar
fricative /x/ was uncategorized, with responses shared
primarily among /t /, /k/, and /h/, so the within-organ
contrast was an uncategorized–categorized assimilation.
According to PAM (Best, 1995), discrimination of
category-goodness assimilations should be moderate to
very good, while discrimination of uncategorized–
categorized assimilations should be very good.
Returning now to the current infant findings, we can
suggest possible assimilation types for the 11-month-
old infants, who should be transitioning to natively
tuned perception. For the within-organ contrast, /x/-/x/,the velar fricative /x/ would fall in an untuned region of
phonetic space and the uvular fricative /x/ would be
assimilated to the English [h] phonetic category—an
untuned–tuned contrast—leading to very good discrimi-
nation via natively tuned perception. In contrast, the
between-organ contrast, /x/-/£/, would have become a
single-category assimilation, because both phones
would have assimilated to English [h], and thus a
decline in discrimination would be predicted over
development. Although Figure 2 is suggestive of a
decline in discrimination for the /x/-/£/ contrast, therewas no significant interaction between age group,
organ, and trial type, so it must be concluded that
discrimination was maintained. Is it possible that
the transition to natively tuned perception occurs a little
later for fricatives than for other consonants, as
suggested by the lack of discrimination of the stop-
fricative contrast /d/-/ð/ by Canadian-English-learning
6–8- and 10–12-month-olds (Polka et al., 2001;
Sundara et al., 2006)? If that were the case, then we
would expect to see a significant decline in discrimina-
tion of this Nuu-Chah-Nulth contrast, relative to
6-month-olds, at a slightly older age such as 14 or
15 months or even 4 years (Sundara et al., 2006).
However, to fully clarify the effects of native language
experience on perception during the first year, specifi-
cally of voiceless fricative place distinctions, we need
also to examine English-learning 6- and 11-month-old
infants’ discrimination of comparable within- versus
between-organ English fricative contrasts, which was
the goal of Experiment 2.
EXPERIMENT 2: DISCRIMINATION OFNATIVE VOICELESS FRICATIVE CONTRASTS
In Experiment 1, we selected between- and within-
organ non-native contrasts from a region of an English-
learning infant’s phonetic space that is unlikely to have
undergone native-language tuning. Here, in Experiment
2, we investigated whether articulatory organ differ-
ences can explain 6-month-olds’ discrimination and
0
1
2
3
4
5
6
7
8
9
10
Between Organ / /-/ /
Within Organ / /-/x/
Between Organ / /-/ /
Within Organ / /-/x/
6 month-olds 11 month-olds
Mea
n fix
atio
n tim
e (s
)
Control
Test
FIGURE 2 Mean fixation durations for between- (/x/-/£/) and within-organ (/x/-/x/) Nuu-Chah-Nulth contrasts at 6 and 11 months of age. Error bars represent standard errors of the mean.
Developmental Psychobiology Infant Perception of Native and Non-Native Fricatives 219

whether PAM can explain developmental changes in
infants’ perception of voiceless fricatives they have
experienced in ambient speech, by testing 6- and 11-
month-olds’ discrimination of between- versus within-
organ contrasts from their native language environment
of English.
The voiceless fricatives of Australian English (and
the primary articulatory organs) are /f/ (lips), /u/, /s/
(both tongue tip), / / (tongue tipþ tongue body), and
/h/ (glottis) (see Cox & Palethorpe, 2007, for a phonetic
description of Australian English). Clearly the within-
organ contrasts must involve the tongue tip and the
between-organ contrast must involve either the lips or
the glottis. Surprisingly, there is very little data on
infant perception of English fricative contrasts differing
in constriction location. In a widely cited study, using a
conditioned headturn procedure, Eilers et al. (1977)
found that while both 6-8- and 12-14-month-olds
discriminated /sa/-/ a/, neither discriminated /ua/-/fa/,and the only the older group discriminated /fi/-/ui/.
On the surface, the failure of young infants to
discriminate between-organ /f/-/u/ in either vowel
context is problematic for the articulatory organ-based
predictions. However, Holmberg et al. (1977, as cited
in Kuhl, 1980), also using a conditioned headturn
procedure, found that 6-month-olds discriminated
/ua/-/fa/ and /sa/-/ a/, and Levitt et al. (1988) reported
discrimination of /ua/-/fa/ and /ða/-/va/ by 2-month-
olds using a modified high-amplitude sucking para-
digm. As /f/ and /u/ naturally have less acoustic energy
than /s/ and / / (see, e.g., Jongman, Wayland, &
Wong, 2000), infants in the Eilers et al. study may have
been capable of detecting the spectral information that
signals the articulatory difference, but the intensity in
her stimuli may not have been sufficiently high to allow
robust discrimination (this has also been suggested by
Kuhl et al., 2008).
Here we tested 6- and 11-month-olds’ discrimination
of between-organ /ua/-/fa/ and within-organ /ua/-/sa/.To our knowledge, no study to date has tested infant
discrimination of /ua/-/sa/, perhaps because infants are
likely to respond to the large differences in acoustic
energy between those two fricatives. Such differences
in acoustic energy would be particularly striking in a
stimulus-alternating preference procedure, such as the
one used here, because tokens of low-energy /ua/ andhigh-energy /sa/ would alternate during the test phase.
If such a large difference in acoustic energy were
presented to infants they may show a preference for the
energy-varying test trials (e.g., alternating /ua/-/sa/)over the energy-stable control trials (e.g., repeated
tokens of /ua/), but it would not be possible to tease
apart whether discrimination was due simply to detec-
tion of acoustic energy differences or to other differ-
ences that are more specific to phonetic properties of
fricative place distinctions. To reduce the possibility
that infants would discriminate /ua/-/sa/ solely on
energy differences between the consonants, without
relying on the dynamic spectral information for place,
and also to minimize the possibility that they would
fail to discriminate /ua/-/fa/ simply due to their low
amplitudes, again without responding to the dynamic
spectral differences of interest, we amplified the frica-
tive portions of /f/ and /u/ and reduced that for /s/ (see
the method section for the stimulus measurements and
naturalness judgments). Paralleling our approach in
Experiment 1, we habituated English-learning infants
to /u/ and then tested their discrimination of it from
both the labiodental /f/ (between-organ contrast) and
alveolar /s/ (within-organ) fricatives using a stimulus-
alternating preference paradigm.
Method
Participants. The sample consisted of 48 English-
learning infants from Sydney, Australia. There were
two age groups, 6-month-olds, Mage¼ 28.6 weeks,
range: 25.4–32.1, and 11-month-olds, Mage¼ 48.5
weeks, range: 46.1–53.6. There were 13 females and 11
males in the 6-month-old group, and 10 females and 14
males in the 11-month-old group. An additional 33
infants were tested and their data were eliminated from
the sample due to failure to habituate (n¼ 4), equip-
ment failure (n¼ 8), experimenter error (n¼ 2), a
subsequent diagnosis of developmental delay (n¼ 1),
having fixation times shorter than 1 s on any individual
trial (n¼ 6), or recovery on the second control trial
(same criterion as Experiment 1), suggesting failure to
completely habituate (n¼ 12).
Stimuli and Apparatus. The English fricative stimuli
consisted of four tokens each of English /f/, /u/, and /s/,
recorded from an Australian female speaker in her late
50s, chosen to match the voice quality of the speaker
whose stimuli were used in Experiment 2. The
consonants were recorded and presented in a consonant
þ /a/ syllable (e.g., /fa/) that was produced in an adult-
directed speech register.
To ensure that any observed discrimination would be
due to acoustic characteristics of the fricatives, and not
to extraneous stimulus variables, we used Praat soft-
ware (Boersma & Weenink, 2009) to adjust the
durations of the consonants and vowels, using overlap-
add resynthesis, to ensure that they had a similar mean
and range as the stimuli used in Experiment 1. To
reduce the possibility that infants might discriminate
the /u/-/s/ contrast using intensity differences alone, /f/
and /u/ were amplified by 5 dB and /s/ was reduced by
220 Tyler et al. Developmental Psychobiology

6 dB (though this still left /s/ with somewhat higher
amplitude than the other two fricatives, to maintain
naturalness). Vowel intensities for all tokens were
adjusted to the same level of acoustic intensity. The
stimulus measurements based on the modified tokens
are presented in Table 3.
Procedure. The procedure was the same as in Experi-
ment 1.
Results
As in Experiment 1, we first analyzed the data using a
2� 2� (2)� (2)� (2) omnibus ANOVA with age
(6- vs. 11-month-olds) and presentation order (between
organ!within organ vs. within organ! between or-
gan) as between-subjects factors, and trial type (test vs.
control), organ (between vs. within), and block (first
vs. second presentation of test trials for each contrast)
as within-subjects factors. Again there was no signifi-
cant main effect of trial type or any other factors, but
there was a significant Block�Trial Type interaction,
F(1,44)¼ 13.94, p¼ .001, hp2¼ .24. For the first block
the mean fixation times were longer for test than
control trials, Mtest¼ 7.39 s (SE¼ 0.63), Mcontrol¼ 5.23 s
(SE¼ 0.31), but in the second block the pattern was
reversed, Mtest¼ 5.88 s (SE¼ 0.53), Mcontrol¼ 7.08 s
(SE¼ 0.62), as we had seen in Experiment 1. An
additional 2� (4)� (2) ANOVA with the data in trial
order showed the same Trial Position�Trial Type
interaction as in Experiment 1. As can be seen in
Figure 3, there is a generally increasing pattern of
fixation times for control trials and a generally decreas-
ing one for test trials. Furthermore, the fixations to the
second control trial appear to be similar to the first
control trial (i.e., the last habituation trial), indicating
that the infants were not simply showing regression to
the mean. Therefore, as in Experiment 1, we analyzed
results for the first block only.
A 2� 2� (2)� (2) ANOVA on the first block
only revealed a significant main effect of trial type,
Table 3. Mean Acoustic Measurements for the English Syllables
Category
Fricative Vowela
Duration
(ms)
Duration Range
(ms)
Amplitude
(dB)
Centroid
(Hz)
Duration
(ms)
Amplitude
(dB SPL)
F1
Hz)
F2
(Hz)
Labiodental /f/ 222 180–254 41 5,991 517 73 786 1,270
Dental /u/ 247 219–302 42 5,995 520 73 780 1,296
Alveolar /s/ 245 176–274 50 8,135 529 73 805 1,269
F1 and F2 refer to the first and second vowel formants, respectively.aVowel measurements are reported from the 50% point.
0
1
2
3
4
5
6
7
8
9
10
11
6 month-olds 11 month-olds
Mea
n fix
atio
n tim
e (s
)
Control 1
Test 1
Control 2
Test 2
Control 3
Test 3
Control 4
Test 4
FIGURE 3 Mean fixation durations for 6- and 11-month-olds for each trial in the order that it
was presented in Experiment 2. Note that the test trials are averaged across the organ factor. The
control trials (black columns) show a generally increasing pattern of fixation durations and the test
trials (gray columns) a generally decreasing one. Error bars represent standard errors of the mean.
Developmental Psychobiology Infant Perception of Native and Non-Native Fricatives 221

F(1,44)¼ 11.00, p¼ .002, hp2¼ .20, with higher fixa-
tion for test trials than for control trials, across ages
and fricative contrasts. No other main effects or
interactions were significant and the mean fixation
times are presented in Figure 4.
Discussion
As in Experiment 1, infants in Experiment 2 looked
longer overall to test than control trials. Again, there
was no evidence for an influence of articulatory-organ-
based differences in 6-month-olds’ discrimination. Our
findings are inconsistent with Eilers et al. (1977), who
found that infants up to 12 months of age failed to
discriminate /u/-/f/, but they are consistent with Holm-
berg (1977, as cited in Kuhl, 1980) and Levitt et al.
(1988), who found that young infants discriminated
/u/-/f/.PAM would explain the developmental pattern in
one of two ways. Either, (1) the 11-month-olds
continue to discriminate the contrasts using the
same pre-phonetic abilities as the 6-month-olds, or;
(2) the 11-month-olds have transitioned to natively-
tuned perception and detect the differences using
their developing phonetic categories. There is some
suggestion in Figure 4 that the 11-month-olds dishabi-
tuated more strongly than the 6-month-olds, which
would lend support to the second possibility, but
there was no significant interaction between age
group and trial type so that possibility must remain
speculative.
GENERAL DISCUSSION
As we noted at the outset, the challenge for theoretical
approaches to experiential effects on infant speech
perception is reconciling seemingly divergent patterns
of discrimination and their development over time. In
this study, we aimed to test whether the AOH (Gold-
stein & Fowler, 2003; Studdert-Kennedy & Goldstein,
2003) might successfully predict 6-month-olds’ dis-
crimination of native and non-native speech contrasts,
to outline how the PAM (Best, 1994a,b, 1995; Best
et al., 1988) accounts for developmental changes in
discrimination, and to provide much needed new data
on infant discrimination of fricative place contrasts.
In our two experiments, we chose non-native
(Experiment 1) and native (Experiment 2) voiceless
fricative place contrasts that would provide a testing
ground for articulatory-organ-based discrimination pre-
dictions. We selected non-native contrasts that fell in a
region of the phonetic space that is unlikely to have
undergone native-language tuning for English-learning
infants, to narrow the possible influences of native-
language exposure on initial discrimination and to test
among the developmental mechanisms put forward in
the literature. The native contrasts, on the other hand,
are naturally constrained because they should both
eventually develop into two phonological categories,
even for within-organ contrasts.
Overall, English-learning 6- and 11-month-olds
looked significantly longer to test than control trials in
both Experiment 1 (Nuu-Chah-Nulth) and Experiment
0
1
2
3
4
5
6
7
8
9
10
Between Organ / /-/f/
Within Organ / /-/s/
Between Organ / /-/f/
Within Organ / /-/s/
6 month-olds 11 month-olds
Mea
n fix
atio
n tim
e (s
)
Control
Test
FIGURE 4 Mean fixation durations for between- (/u/-/f/) and within-organ (/u/-/s/) English
contrasts at 6 and 11 months of age. Error bars represent standard errors of the mean.
222 Tyler et al. Developmental Psychobiology

2 (English). On the basis of the AOH (Goldstein &
Fowler, 2003; Studdert-Kennedy & Goldstein, 2003),
we had predicted that between-organ contrasts
should always be discriminable and that discrimination
of within-organ contrasts should initially be poorer
than for between-organ contrasts, prior to native-
language attunement. That prediction was not upheld
in either experiment. Therefore, although organ-based
approaches provided a potentially useful way of
predicting which contrasts an infant might initially
discriminate well, we did not find support for this idea
in our experiments (see similar findings in Narayan
et al., 2010).
Turning to predictions for changes in discrimination
over time, the results showed maintenance of all
contrasts in both experiments from 6 to 11 months.
Here we will discuss how this result fits with PAM and
with two additional theories/frameworks of infant
speech perception that are most relevant to the issues at
hand: (1) the Native Language Magnet Theory-Expand-
ed (NLM-e; Kuhl et al., 2008); and (2) Processing Rich
Information from Multi-dimensional Interactive Repre-
sentations (PRIMIR; Curtin et al., 2011; Werker &
Curtin, 2005).
NLM-e (Kuhl et al., 2008) proposes that develop-
mental changes in speech perception result from infants
tracking the statistical regularities in the acoustics of
ambient speech (the native language environment).
Infants can initially discriminate “all phonetic units in
the world’s languages” (Kuhl et al., 2008, p. 998), but
there may be variation in the level of discrimination
achieved according to acoustic factors such as salience
(e.g., Burnham, 1986; see also Cristia et al., 2011), and
amplitude variation across different consonants. If
discrimination is not initially high for a given contrast,
it should improve over development if it is a native
contrast, but there should necessarily be a decline in
discrimination of all non-native contrasts, regardless of
how well they were discriminated initially. The results
for the native contrasts are consistent with NLM-e
because the initial high discrimination of the English
contrasts was maintained over development. However,
the results for Nuu-Chah-Nulth appear inconsistent
with NLM-e because we did not observe a decline over
development for either non-native contrast.
Like NLM-e, PRIMIR (Curtin et al., 2011; Werker
& Curtin, 2005) states that the learning mechanism
responsible for infant attunement to the language
environment is statistical learning, however, it takes
no stand regarding the type of information that is
“tracked” or perceived by the infant (e.g., acoustic or
articulatory). The framework is comprised of three
multidimensional interactive representational spaces.
Phonetic and non-phonetic (i.e., indexical) information
are stored in the General Perceptual Space, in which
clusters of phonetic and indexical categories form over
time through processes of statistical learning. The
Word Form Space stores exemplars of sound sequences
that become attached to meaning as the conceptual
system develops, and from that linkage the Phoneme
Space emerges as the children come to recognize
recurring segments across their developing lexicons. In
addition to these spaces, dynamic filters direct infants’
attention to particular subsets of the available informa-
tion. The filters are initial biases, task demands, and
developmental level. During the first year of life,
infants should demonstrate facilitation or maintenance
for native contrasts, similar to NLM-e, depending on
initial biases and task demands. A decline should be
observed for non-native contrasts, although PRIMIR
differs from NLM-e in positing that discrimination may
still be observed later in life if the demands of the task
are sufficiently low (e.g., Werker & Logan, 1985). A
decline in discrimination for only some native or non-
native contrasts but not others would be explained in
the PRIMIR framework via the intersection of the
formation of new representational spaces (Word Form
and Phoneme spaces) and the influence of the dynamic
filters, but to our knowledge the framework is not yet
sufficiently well specified to predict which particular
contrasts should show maintenance and which should
show decline.
According to PRIMIR, infants use the General
Perceptual Space and any phonetic categories devel-
oped during the first year of life for discrimination.
Globally, the predictions for PRIMIR would appear to
be similar to those of PAM. However, there is one key
difference between PRIMIR and PAM when consider-
ing discrimination of non-native contrasts. For PRI-
MIR, phonetic categories are established through
processes of statistical learning so non-native phones
could only be assimilated to a native category if they
fall within its statistical distribution of phonetic proper-
ties. PAM, on the other hand, proposes a Gibsonian
(Gibson & Gibson, 1955) process of perceptual learn-
ing. From this viewpoint, perceivers extract the infor-
mation relevant to their needs directly from the distal
objects and events in their environment. With complex
events, such as speech, they often must learn through
experience which dimensions and combinations of
information are crucial in their environment and thus
should be selectively attended to, versus which other
aspects of available information are irrelevant and need
not be attended to or extracted. By that mechanism
infants might assimilate a non-native phone to a native
category, even if it lies outside of any specific native
statistical distribution of phonetic properties, so long as
the infant recognized a higher-order invariant that was
Developmental Psychobiology Infant Perception of Native and Non-Native Fricatives 223

consistent with a native category. The results for Nuu-
Chah-Nulth, therefore, could only be accounted for
within the PRIMIR framework via a combination of
initial biases and task demands—there could be no
influence of native-language attunement on discrimina-
tion of non-native contrasts.
For PAM, discrimination of both the Nuu-Chah-
Nulth and English contrasts at 6 months must be due to
initial biases. We argue that infants transition from
untuned to natively tuned perception between 6 and
12 months of age (or even later for certain consonant
contrasts; see Polka et al., 2001; Sundara et al., 2006).
Our data are not inconsistent with such a view, but we
did not observe a significant decline in discrimination
for the Nuu-Chah-Nulth uvular-pharyngeal contrast,
which would have been predicted for a single-category
contrast. This may be due to attunement to fricatives
occurring later in development (Polka et al., 2001;
Sundara et al., 2006), but it is also possible that the
11-month-olds showed a sensitivity to category good-
ness that was observed in adults by Tyler and Best
(2010). Such a finding would clearly falsify our
contention that native phonetic categories have little
within-category differentiation in infancy (see footnote
3). We acknowledge that more data are required to
evaluate our proposal.
In the context of perceptual narrowing, the critical
lesson is that attunement to the native language does
not involve a decline in discrimination for all non-
native phonetic contrasts. From the PAM perspective,
those heard as non-speech, that is, not assimilated as
speech elements (e.g., clicks for English-learning
infants; Best et al., 1988) will continue to be discrimi-
nated using domain-general perception, whereas those
heard as speech will be discriminated through the filter
of the native phonological system. If two non-native
phones are assimilated to two different native catego-
ries, then discrimination should remain excellent across
age (i.e., a PAM two-category assimilation; see Best,
1995), even if the non-native phones do not phonetical-
ly resemble any native phone (e.g., adult English-
speakers’ perception of Zulu voiced versus voiceless
lateral fricatives / /-/ /: Best et al., 2001). Note,
however, that the same contrast may be assimilated
differently by 11-month-olds than by adults, for exam-
ple as a single-category contrast or as an untuned–
untuned contrast, either of which could lead to a
temporary decline in discrimination (e.g., decline at
11 months in discrimination of the Zulu lateral fricative
voicing contrast / /-/ /: Best & McRoberts, 2003).
To continue toward an explanation of infant speech
discrimination abilities, we require data from cross-
language studies, such as those conducted by Polka
et al. (2001) and Kuhl et al. (2006), where infants from
different language-learning environments are presented
with the same stimuli. If infants are born with universal
perceptual biases, then young infants should show
similar discrimination patterns regardless of their
language environment. Effects of native-language tun-
ing are much easier to interpret when infants from
different language environments are presented with
exactly the same stimuli and divergent patterns of
discrimination are observed. For example, while we
have shown that English infants, whose language input
does not favor tuning to tongue-body and tongue-root
fricatives, discriminate the Nuu-Chah-Nulth stimuli, it
would be interesting to test Arabic-learning infants
because Arabic has a uvular and a pharyngeal fricative.
If the discrimination patterns we observed here are
universal at 6 months but language-specific at
11 months, then we should observe a developmental
decline for the velar-uvular (/x/-/x/) contrast by the
Arabic-learning infants. In contrast, the uvular-pharyn-
geal (/x/-/£/) contrast should be maintained for this
group as it would be developing into an Arabic two-
category distinction with native-language tuning. It
would be particularly interesting to extend these cross-
language comparisons to infants learning languages
with smaller consonant inventories, particularly inven-
tories with notable “gaps” in certain types of conso-
nants. In this regard, indigenous Australian languages
such as Arrente and Pitjantjatjara would be of particular
interest, as they lack fricative phonemes altogether, yet
on the other hand they tend to have multiple coronal
places for stops, nasals and approximants that are all
lacking in English. That is, Australian languages lack
an entire series of contrasts found in English (voiced
and voiceless fricatives), but conversely they have other
series of place contrasts lacking in most other lan-
guages including English (coronal stop place distinc-
tions).4
To conclude, articulatory-organ differences do not
appear to influence perception of speech contrasts by
young infants, and it is still not clear which factors
determine initial levels of discrimination, or whether
those levels are observed universally. Nevertheless, our
results provide another example of maintenance of
both non-native (Experiment 1) and native contrasts
(Experiment 2) across the second half of the first year
of life, for both within- and between-organ contrasts.
Moreover, our findings on 6- and 11-month-olds’
4Pragmatic issues, however, make this type of study very
unlikely. Many indigenous Australian languages are dying or
have died out already; the children are no longer acquiring them
naturally from birth. Even in languages that infants are still
learning, both practical and ethical concerns mean that conduct-
ing experiments would be highly problematic at best.
224 Tyler et al. Developmental Psychobiology

discrimination of non-native fricative contrasts from an
uncommitted region of phonetic space have shown
that perceptual attunement to the native language does
not necessarily entail perceptual narrowing, which
predicts a decline in discrimination of non-native
contrasts from a region of phonetic space that the
infants could not have encountered in their ambient
language environment.
NOTES
This research was supported by NIH grant DC00403 (PI: C.
Best). We thank Bryan Gick and Ian Wilson greatly for their
assistance in obtaining the Nuu-Chah-Nulth stimuli, Christian
Kroos for assistance with Praat stimulus adjustments for
Experiment 2, and all of the parents and babies who
participated in the study. We also grateful for the research
assistance of Suzana Bicanic, Lidija Krebs-Lazendic, Anna
Notley, Matthew Richardson, Dianne West, Susan Wijngaar-
den, and Bethany Wootton.
REFERENCES
Aslin, R. N., & Pisoni, D. B. (1980). Some developmental
processes in speech perception. In: G. H. Yeni-Komshian,
J. Kavanagh, & C. A. Ferguson (Eds.), Child phonology
(Vol. 2, pp. 67–96). New York, NY: Academic Press.
Aslin, R. N., Werker, J. F., & Morgan, J. L. (2002). Innate
phonetic boundaries revisited (L). Journal of the Acousti-
cal Society of America, 112, 1257–1260.
Best, C. T. (1994a). Learning to perceive the sound pattern of
English. In: C. Rovee-Collier & L. P. Lipsitt (Eds.),
Advances in infancy research (Vol. 9, pp. 217–304).
Norwood, NJ: Ablex.
Best, C. T. (1994b). The emergence of language-specific
phonemic influences in infant speech perception. In:
J. C. Goodman & H. C. Nusbaum (Eds.), The development
of speech perception (pp. 167–224). Cambridge, MA: MIT
Press.
Best, C. T. (1995). A direct realist view of cross-language
speech perception. In: W. Strange (Ed.), Speech percep-
tion and linguistic experience: Issues in cross-language
research (pp. 171–204). Baltimore, MD: York Press.
Best, C. T., & Jones, C. (1998). Stimulus-alternation prefer-
ence procedure to test infant speech discrimination. Infant
Behavior and Development, 21, 295.
Best, C. T., & McRoberts, G. W. (2003). Infant perception of
non-native consonant contrasts that adults assimilate in
different ways. Language and Speech, 46, 183–216.
Best, C. T., McRoberts, G. W., & Goodell, E. (2001).
Discrimination of non-native consonant contrasts varying
in perceptual assimilation to the listener’s native phono-
logical system. Journal of the Acoustical Society of
America, 109, 775–794.
Best, C. T., McRoberts, G. W., LaFleur, R., & Silver-
Isenstadt, J. (1995). Divergent developmental patterns for
infants’ perception of two non-native consonant contrasts.
Infant Behavior and Development, 18, 339–350.
Best, C. T., McRoberts, G. W., & Sithole, N. M. (1988).
Examination of perceptual reorganization for non-native
speech contrasts: Zulu click discrimination by English-
speaking adults and infants. Journal of Experimental
Psychology: Human Perception and Performance, 14,
345–360.
Best, C. T., Tyler, M. D., Gooding, T. N., Orlando, C. B., &
Quann, C. A. (2009). Development of phonological
constancy: Toddlers’ perception of native- and Jamaican-
accented words. Psychological Science, 20, 539–542.
Boersma, P., & Weenink, D. (2009). Praat: Doing phonetics
by computer (Version 4.5.4.4). Institute of Phonetic
Sciences, University of Amsterdam [Computer program].
Retrieved January 12, 2007, from http://www.praat.org/
Boyce, S. E., & Espy-Wilson, C. Y. (1997). Coarticulatory
stability in American English /r/. Journal of the Acoustical
Society of America, 101, 3741–3753.
Browman, C. P., & Goldstein, L. (1990). Gestural specifica-
tion using dynamically-defined articulatory structures.
Journal of Phonetics, 18, 299–320.
Browman, C. P., & Goldstein, L. (1992). Articulatory
phonology: An overview. Phonetica, 49, 155–180.
Browman, C. P., & Goldstein, L. (1995). Gestural syllable
position effects in American English. In: F. Bell-Berti &
L. J. Raphael (Eds.), Producing speech: Contemporary
issues (pp. 19–33). Woodbury, NY: AIP Press.
Browman, C. P., & Goldstein, L. (2000). Competing con-
straints on intergestural coordination and self-organization
of phonological structures. Bulletin de la Communication
Parlee, 5, 25–34.
Burnham, D. K. (1986). Developmental loss of speech
perception: Exposure to and experience with a first
language. Applied Psycholinguistics, 7, 207–240.
Carlson, B. F., Esling, J. H., & Fraser, K. (2001). Nuuchah-
nulth. Journal of the International Phonetic Association,
31, 275–279.
Cohen, L. B. (2004). Uses and misuses of habituation and
related preference paradigms. Infant and Child Develop-
ment, 13, 349–352.
Cox, F., & Palethorpe, S. (2007). Australian English. Journal
of the International Phonetic Association, 37, 341–350.
Cristia, A., McGuire, G. L., Seidl, A., & Francis, A. L.
(2011). Effects of the distribution of acoustic cues on
infants’ perception of sibilants. Journal of Phonetics, 39,
388–402.
Curtin, S., Byers-Heinlein, K., & Werker, J. F. (2011).
Bilingual beginnings as a lens for theory development:
PRIMIR in focus. Journal of Phonetics, 39, 492–504.
Eilers, R. E., Gavin, W., & Wilson, W. R. (1979). Linguistic
experience and phonemic perception in infancy: A cross-
linguistic study. Child Development, 50, 14–18.
Eilers, R. E., Wilson, W. R., & Moore, J. M. (1977). Develop-
mental changes in speech discrimination in infants. Journal
of Speech, Language, and Hearing Research, 20, 766–780.
Developmental Psychobiology Infant Perception of Native and Non-Native Fricatives 225

Gibson, J. J. (1979). The ecological approach to visual
perception. Boston, MA: Houghton Mifflin.
Gibson, J. J., & Gibson, E. J. (1955). Perceptual learning:
Differentiation or enrichment? Psychological Review, 62,
32–41.
Goldstein, L., & Fowler, C. A. (2003). Articulatory phono-
logy: A phonology for public language use. In:
N. O. Schiller & A. S. Meyer (Eds.), Phonetics and
phonology in language comprehension and production:
Differences and similarities (pp. 159–208). Berlin,
Germany: Mouton de Gruyter.
Gottlieb, G. (1976). The roles of experience in the develop-
ment of behavior and the nervous system. In: G. Gottlieb
(Ed.), Neural and behavioral plasticity (pp. 25–54). New
York, NY: Academic Press.
Guenther, F. H., Espy-Wilson, C. Y., Boyce, S. E., Matthies,
M. L., Zandipour, M., & Perkell, J. S. (1999). Articulatory
tradeoffs reduce acoustic variability during American
English /r/ production. Journal of the Acoustical Society
of America, 105, 2854–2865.
Harnsberger, J. D. (2001). On the relationship between
identification and discrimination of non-native nasal con-
sonants. Journal of the Acoustical Society of America,
110, 489–503.
Houston, D. M., Horn, D. L., Qi, R., Ting, J. Y., & Gao, S.
(2007). Assessing speech discrimination in individual
infants. Infancy, 12, 119–145.
Jongman, A., Wayland, R., & Wong, S. (2000). Acoustic
characteristics of English fricatives. Journal of the Acous-
tical Society of America, 108, 1252–1263.
Kitamura, C., Panneton, R., & Best, C. T. (2013). The
development of language constancy: Attention to native
versus non-native accents. Child Development, 64, 1686–
1700. doi: 10.1111/cdev.12068
Kuhl, P. K. (1980). Perceptual constancy for speech-sound
categories in early infancy. In: G. H. Yeni-Komshian,
J. Kavanagh, & C. A. Ferguson (Eds.), Child phonology
(Vol. 2, pp. 41–66). New York, NY: Academic Press.
Kuhl, P. K., Conboy, B. T., Coffey-Corina, S., Padden, D.,
Rivera-Gaxiola, M., & Nelson, T. (2008). Phonetic learn-
ing as a pathway to language: New data and native
language magnet theory expanded (NLM-e). Philosophical
Transactions of the Royal Society B: Biological Sciences,
363, 979–1000.
Kuhl, P. K., Stevens, E., Hayashi, A., Deguchi, T., Kiritani,
S., & Iverson, P. (2006). Infants show a facilitation effect
for native language phonetic perception between 6 and 12
months. Developmental Science, 9, F13–F21.
Levitt, A., Jusczyk, P. W., Murray, J., & Carden, G. (1988).
Context effects in two-month-old infants’ perception of
labiodental/interdental fricative contrasts. Journal of Ex-
perimental Psychology: Human Perception and Perfor-
mance, 14, 361–368.
Maddieson, I. (1984). Patterns of sounds. Cambridge, MA:
Cambridge University Press.
Mattock, K., Molnar, M., Polka, L., & Burnham, D. (2008).
The developmental course of lexical tone perception in the
first year of life. Cognition, 106, 1367–1381.
Meltzoff, A. N., & Moore, M. K. (1977). Imitation of facial
and manual gestures by human neonates. Science, 198,
75–78.
Miller, C. L. (1983). Developmental changes in male/female
voice classification by infants. Infant Behavior and Devel-
opment, 6, 313–330.
Mulak, K., & Best, C. T. (2013). Development of word
recognition across speakers and accents. In: L. Gogate &
G. Hollich (Eds.), Theoretical and computational models
of word learning: Trends in psychology and artificial
intelligence (pp. 242–269). Hershey, PA: IGI Global-
Robotics.
Narayan, C. R., Werker, J. F., & Beddor, P. S. (2010). The
interaction between acoustic salience and language experi-
ence in developmental speech perception: Evidence from
nasal place discrimination. Developmental Science, 13,
407–420.
Nittrouer, S. (2001). Challenging the notion of innate phonet-
ic boundaries. Journal of the Acoustical Society of
America, 110, 1598–1605.
Nittrouer, S. (2002). A reply to “Innate phonetic boundaries
revisited” [J. Acoust. Soc. Am. 112, 1257–1260 (2002)]
(L). Journal of the Acoustical Society of America, 112,
1261–1264.
Oakes, L. M. (2010). Using habituation of looking time to
assess mental processes in infancy. Journal of Cognitive
Development, 11, 255–268.
Polka, L. (1991). Cross-language speech perception in
adults: Phonemic, phonetic, and acoustic contributions.
Journal of the Acoustical Society of America, 89, 2961–
2977.
Polka, L., Colantonio, C., & Sundara, M. (2001). A cross-
language comparison of /d/–/ð/ perception: Evidence for a
new developmental pattern. Journal of the Acoustical
Society of America, 109, 2190–2201.
Stone, M., & Lundberg, A. (1996). Three-dimensional
tongue surface shapes of English consonants and vowels.
Journal of the Acoustical Society of America, 99, 3728–
3737.
Strange, W., Akahane-Yamada, R., Kubo, R., Trent, S. A.,
Nishi, K., & Jenkins, J. J. (1998). Perceptual assimilation
of American English vowels by Japanese listeners. Journal
of Phonetics, 26, 311–344.
Studdert-Kennedy, M. (1991). Language development from
an evolutionary perspective. In: N. A. Krasnegor,
D. M. Rumbaugh, M. Schiefelbusch, & M. Studdert-
Kennedy (Eds.), Language acquisition: Biological and
behavioral determinants (pp. 5–28). Hillsdale, NJ: Law-
rence Erlbaum Associates.
Studdert-Kennedy, M., & Goldstein, L. (2003). Launch-
ing language: The gestural origin of discrete infinity.
In: M. H. Christiansen & S. Kirby (Eds.), Language
evolution (pp. 235–254). Oxford, UK: Oxford University
Press.
Sundara, M., Polka, L., & Genesee, F. (2006). Language-
experience facilitates discrimination of /d-ð/ in monolin-
gual and bilingual acquisition of English. Cognition, 100,
369–388.
226 Tyler et al. Developmental Psychobiology

Trehub, S. E. (1976). The discrimination of foreign speech con-
trasts by infants and adults. Child Development, 47, 466–472.
Tyler, M. D., & Best, C. T. (2010). Discrimination of non-
native fricative contrasts by English monolinguals and
Arabic-English. In: N. Voudouris & V. Mrowinski (Eds.),
Combined abstracts of 2010 Australian psychology confer-
ences (p. 39). Melbourne, Victoria, Australia: Australian
Psychological Society Ltd.
Tyler, M. D., Best, C. T., Faber, A., Levitt, A. (in press).
Perceptual assimilation of non-native vowel contrasts.
Phonetica. doi: 10.1159/000356237
Werker, J. F. (1995). Exploring developmental changes in
cross-language speech perception. In: D. N. Osherson
(Series Ed.), L. R. Gleitman & M. Liberman (Vol. Eds.),
An invitation to cognitive science, Vol. 1 Language
(2nd ed., pp. 87–106). Cambridge, MA: MIT Press.
Werker, J. F., & Curtin, S. (2005). PRIMIR: A developmental
framework of infant speech processing. Language Learn-
ing and Development, 1, 197–234.
Werker, J. F., Gilbert, J. H. V., Humphrey, K., & Tees,
R. C. (1981). Developmental aspects of cross-language
speech perception. Child Development, 52, 349–355.
Werker, J. F., & Lalonde, C. E. (1988). Cross-language
speech perception: Initial capabilities and developmental
change. Developmental Psychology, 24, 672–683.
Werker, J. F., & Logan, J. S. (1985). Cross-language evidence
for three factors in speech perception. Perception &
Psychophysics, 37, 35–44.
Werker, J. F., & Tees, R. C. (1984). Cross-language speech
perception: Evidence for perceptual reorganization during
the first year of life. Infant Behavior and Development, 7,
49–63.
Developmental Psychobiology Infant Perception of Native and Non-Native Fricatives 227



















