
Intraspecific comparison of diet of California sea lions ...

ISSN 1067�4136, Russian Journal of Ecology, 2013, Vol. 44, No. 1, pp. 18–27. © Pleiades Publishing, Ltd., 2013.Original Russian Text © E.V. Antonova, E.M. Karimullina, V.N. Pozolotina, 2013, published in Ekologiya, 2013, No. 1, pp. 20–29.
18
The impact of ionizing radiation on living orga�nisms is modified by various factors (Vilenchik, 1991;Petin, Zhurakovskaya, and Komarova, 2012). Expo�sure to low radiation doses in combination with otherfactors may have antagonistic, additive, or synergisticeffects (Geras’kin et al., 2005; Burkhardt, Internicola,and Bernasconi, 2009), and what we observe in natureis an integrated response of organisms to the entirecomplex of relevant factors. The ecological approachto studies on the consequences of long�term radiationexposure for the biota makes it necessary to take intoaccount different types of intraspecific variation in theresponse to radiation and evaluation of the interactionof factors acting in different combinations (Pozolo�tina, 2003).
The Eastern Ural Radioactive Trace (EURT),which was formed after the 1957 nuclear accident atthe Mayak Radiochemical Plant, is a unique test rangefor assessing the consequences of chronic radiationexposure for living organisms. Radioactive 90Sr is atracer for radionuclides released during the accident.Ten years after the accident, this territory was con�taminated again by radioactive silt and sand trans�
ferred by wind from the shore of shallowed LakeKarachay (reservoir V�9 for liquid radioactive wastes),with 137Cs being the tracer radionuclide. Several vil�lages in the frontal part of the EURT were relocated,and the Eastern Ural State Reserve was established inthis area (Nikipelov et al., 1989).
Genetic effects of chronic radiation exposure indifferent plant species from the EURT zone aredescribed in numerous publications, including papersfrom our laboratory. They concern the levels of chro�mosome aberrations, allozyme diversity in cenopopu�lations of model species, and the frequencies of muta�tions and nonheritable modifications in the seed prog�eny of plants from the EURT (Shevchenko,Pechkurenkov, and Abramov, 1992; Kal’chenko,Rubanovich, and Shevchenko,1995; Lysenko,Kal’chenko, and Shevchenko, 1999; Pozolotina,2003; Antonova and Pozolotina, 2007; Pozolotinaet al., 2008; Pozolotina, Antonova, and Karimullina,2010; Abramov, Stepanova, and Famelis, 2010).However, taking into account the variety of ecoge�netic characteristics of plants, it is apparent that newspecies should be involved in radioecological research.
Intraspecific Variation in Melandrium album along a Radioactive Contamination Gradient at the Eastern Ural Radioactive Trace
E. V. Antonova, E. M. Karimullina, and V. N. PozolotinaInstitute of Plant and Animal Ecology, Ural Branch, Russian Academy of Sciences,
ul. Vos’mogo Marta 202, Yekaterinburg, 620144 Russia
e�mail: [email protected]
Received January 18, 2012
Abstract—Specific ecogenetic features of plants and parameters of seed progeny have been studied in Melan�drium album cenopopulations from test plots established along the radioactive contamination gradient in thezone of the Eastern Ural Radioactive Trace (EURT), where absorbed radiation doses in buffer and impactplots exceeded the background level by factors of 2–18 and 330–418, respectively. It has been shown thatinterannual variation in parameters characterizing the viability of seed progeny largely depends on fluctua�tions of weather conditions. This dependence is more distinct in cenopopulations from the EURT than incontrol samples, which is indicative of interaction between radiation and other relevant ecological factors.The radioresistance of seed progeny is contingent on its viability, being independent of radiation load at thesites of maternal plant growth. Hermaphrodite plants occur only in irradiated M. album cenopopulations.This character is inherited: all F1 descendants of such plants grown in a vegetation experiment had herma�phrodite flowers. The mutability of seed progeny formed in the EURT zone exceeds the background level.The occurrence and frequencies of recorded anomalies has proved to vary by years and, therefore, can beinterpreted as nonheritable modifications.
Keywords: Eastern Ural Radioactive Trace (EURT), absorbed radiation doses, ERICA Tool, Melandriumalbum, chronic radiation exposure, weather conditions, radiobiological effects, intraspecific variation, her�maphrodites
DOI: 10.1134/S1067413613010025

RUSSIAN JOURNAL OF ECOLOGY Vol. 44 No. 1 2013
INTRASPECIFIC VARIATION IN Melandrium album 19
Analysis of plant species differing in the mode ofreproduction, morphological characters, initialradioresistance, and requirements for basic ecologicalfactors (temperature, humidity, etc.) and comparisonof their specific responses to chronic radiation expo�sure provide the possibility of revealing the diversity ofadaptive mechanisms in plants.
The object of this study was the white campion,Melandrium album (Mill.) Garcke (synonyms: Silenelatifolia Poirt, Lychnis alba Miller, Silene alba), a dioe�cious, herbaceous annual or biennial plant of thefamily Caryophyllaceae, a diploid (2n = 24, x = 12). Itis classified as a hemicryptophyte or therophyte and amesophyte but is tolerant of low air humidity. Theplant is photophilic and does not bloom in shadedareas. In nature, M. album usually grows in meadowsand at forest margins and reproduces mainly by seeds(Baker, 1947; Gulenkova and Pyatunina, 1997). Thisspecies is widely used as a model for studies in evolu�tion and phylogenetics (Bernasconi et al., 2009; Rau�tenberg et al., 2010), reproductive biology and ecology(Jolivet and Bernasconi, 2007), and, in particular, onsexual dimorphism (Taylor, Trimble, and McCauley,1999; Steven, Delph, and Brodie, 2007; Delph et al.,2010). Such interest received from specialists indifferent fields confirms the uniqueness of this species.However, its radiobiological characteristics practicallyhave not been studied.
Combinations of temperature and precipitationlevels, frosts and thaws, etc. change every year, therebycreating specific conditions for the development ofplants and formation of their seed progeny. In theEURT zone, the spectrum of these factors is comple�mented by elevated background radiation. The qualityof seed progeny, including its radioresistance, is vari�able. One�year observations are insufficient for objec�tively evaluating the consequences of chronic radia�tion exposure, since it is necessary to take into accountshifts in the limits of variation in key parameterscharacterizing the state of plants depending onweather conditions (Pozolotina, 2003).
The purpose of this study was to evaluate specificecogenetic features of M. album plants from theEURT zone and interannual variation in the quality oftheir seed progeny formed in the gradient of radioac�tive contamination and to analyze the causes of thisvariation.
MATERIAL AND METHODS
Studies were performed in the frontal part of theEURT, at different distances from the epicenter of theaccident (indicated in parentheses). Two impact plotswere established along the EURT central axis:Impact�1 (9–12 km) at the site of relocated villagenear Lake Berdenish (55°46′ N, 60°53′ E) and Impact 2(6–8 km) in the vicinity of a track road (55°45′ N,60°50′ E). Plot Buffer�2 (15–18 km) was also on theEURT central axis, at the site of relocated village nearLake Uruskul’ (55°49′ N, 60°55′ E); plot Buffer�1(17 km) was at the EURT periphery, near the boun�dary of the Eastern Ural State Reserve (55°50′ N,60°52′ E). Three background plots were beyond thezone of any technogenic pollution: Background�1(112 km) in the vicinity of Rassokha (56°41′ N, 61°02′ E),Background�2 (125 km) in the vicinity of Sheelit(56°47′ N, 61°18′ E), and Background�3 (240 km) inthe vicinity of Zaikovo (57°33′ N, 62°42′ E). Adetailed geobotanical description of the plots was pub�lished previously (Pozolotina et al., 2008, 2009).
Gamma�radiation exposure dose rates were mea�sured with a DRG�01T dosimeter at the soil surface(Table 1). To calculate absorbed doses of β�radiation,M. album phytomass and soil samples (by layers) werecollected at the sites of maternal plant growth. Sam�ples from each plot were taken in 5–25 replications.Concentrations of 90Sr in soil and plant samples weredetermined radiochemically, by daughter 90Y, andthose of 137Cs, by spectrometry in a multichannel γ�analyzer with a germanium semiconductor detector(Pozolotina et al., 2008). Absorbed radiation doses forthe entire plants were calculated with the ERICA Toolprogram package (Beresford et al., 2007; 2008; Brown
Table 1. Gamma�radiation exposure dose rates at the soil surface and 90Sr + 137Cs specific radioactivities in the upper (0–5 cm) soil layer in the EURT zone and background plots
Plot Exposure dose rate, µR/h
Soil specific radioactivity, Bq/kg air�dry weight
90Sr 137Cs
X ± SE min–max X ± SE min–max
Background�1 5–10 6.5 ± 1.3 1.1–10 28.0 ± 8.7 3.4–73
Background�2 9–12 6.3 ± 1.45 2.9–13.5 19.8 ± 10.6 3.9–81.5
Buffer�1 12–16 588 ± 335 60–2551 320 ± 77.6 95.4–651
Buffer�2 10–21 10320 ± 7871 1704–33922 405 ± 206 87–989
Impact�1 14–176 85434 ± 11158 171–298411 3564 ± 625 151–20303
Impact�2 86–127 214948 ± 40971 103278–284909 12667 ± 3488 3096–18682
20
RUSSIAN JOURNAL OF ECOLOGY Vol. 44 No. 1 2013
ANTONOVA et al.
et al., 2008) in the version of June 2011. We used theTier 2 model, which allowed us to put in empiricalvalues of radionuclide transfer coefficients, thefresh/dry biomass ratio for the test plant species, andother parameters. Separate accounts were taken of thecontributions of 90Sr + 90Y and 137Cs by inputtingaverage values of their specific radioactivity in theupper (0–5 cm) soil layer, because β�radiation fromdeeper soil horizons is absorbed completely.
The seeds of M. album were collected from the plotsin 2005, 2007, 2008, and 2010. The inflorescences ofM. album plants in 2009 were damaged en masse bylarvae of the moth Hadena bicruris (Hufnagel, 1766),a pollinator of this plant, which feed on immatureseeds and thereby drastically reduce seed production(Wolfe, 2002). Hence, we failed to collect enoughseeds for experiments. In the laboratory, the seeds weregerminated by the method of paper roll culture at con�stant temperature (24°С) and 12�hour photoperiod,and the seed progeny was analyzed for viability (therates of seedling survival, true leaf emergence, androot growth) and mutability (the occurrence fre�quency of morphological modifications). Radiosensi�tivity of the seeds was estimated by means of acuteirradiation at doses of 100, 200, 300, 400 Gy (dose rate41.2 cGy/s) in an Igur γ�radiation device with a 137Сssource. After irradiation the seeds were germinatedfollowing the above criteria. Comparing the viability ofirradiated and nonirradiated seeds from different sam�ples, it was possible to estimate their adaptive potential.Individual variation in viability and radiosensitivity wasevaluated by germinating the seeds of 10–20 plantsfrom different samples. A total of 22749 seeds wereincluded in experiments.
The data collected over several years were used toanalyze interannual variation in plant viability andradiosensitivity within M. album cenopopulations.The effect of weather conditions in different years wasestimated by taking into account monthly average andeffective temperatures, monthly average precipitation,and the sum of precipitation at effective temperatures,
using Selyaninov’s hydrothermal index (The Encyclo�pedia…, 2005) as an integrated parameter. Meteoro�logical data on the background area were obtainedfrom Yekaterinburg weather station (28440; 56°48′ N,60°38′ E); on plots in the EURT zone, from theVerkhnii Ufalei weather station (28541; 56°05′ N,60°18′ E).
Working hypotheses were verified using one� andtwo�way ANOVA, correlation analysis, Kruskal–Wallis(H) and Mann–Whitney (U) nonparametric rank tests,and Fisher’s multiple comparison test (F). Their statisti�cal significance was verified using the Statistica 6.0 pro�gram package (StatSoft Inc., 2001).
RESULTS AND DISCUSSION
Radiation Loads on Melandrium album Plants
As follows from Table 1, γ�radiation exposure doserate increases as the epicenter of the accident isapproached, exceeding the background level by a fac�tor of 8.5–17 near the track road. Specific radioactiv�ities of the main dose�forming radionuclides (90Sr and137Cs) in the soil widely vary in the EURT zone,exceeding the background levels by factors of 91–33070 and 11–640, respectively. Such a mosaic pat�tern of contamination is due to nonuniformity ofradioactive fallout after the accident and subsequentland reclamation measures taken in this area (Pozolo�tina et al., 2008; Molchanova et al., 2009).
The increase in radionuclide contents in the soil isnot accompanied by a strictly proportional increase intheir contents in plants. As shown previously (Pozolo�tina, Antonova, and Karimullina, 2010), the range oftransfer coefficient (TC) values varies depending onphysicochemical parameters of soils, the level of theirradioactive contamination, and species�specific fea�tures of plants. According to our calculations, TC for90Sr in M. album vary from 0.236 to 0.283 in bufferplots and from 0.262 to 0.870 in impact plots (back�ground TC = 0.413).
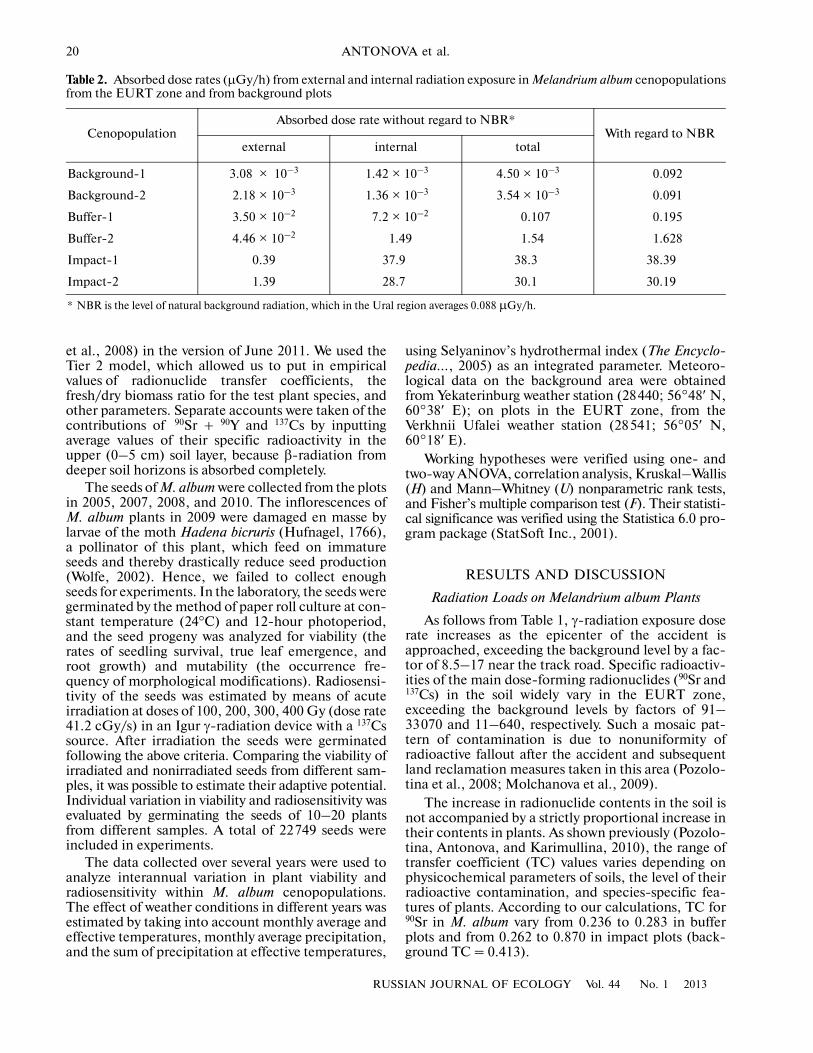
Table 2. Absorbed dose rates (µGy/h) from external and internal radiation exposure in Melandrium album cenopopulationsfrom the EURT zone and from background plots
CenopopulationAbsorbed dose rate without regard to NBR*
With regard to NBRexternal internal total
Background�1 3.08 × 10–3 1.42 × 10–3 4.50 × 10–3 0.092
Background�2 2.18 × 10–3 1.36 × 10–3 3.54 × 10–3 0.091
Buffer�1 3.50 × 10–2 7.2 × 10–2 0.107 0.195
Buffer�2 4.46 × 10–2 1.49 1.54 1.628
Impact�1 0.39 37.9 38.3 38.39
Impact�2 1.39 28.7 30.1 30.19
* NBR is the level of natural background radiation, which in the Ural region averages 0.088 µGy/h.

RUSSIAN JOURNAL OF ECOLOGY Vol. 44 No. 1 2013
INTRASPECIFIC VARIATION IN Melandrium album 21
Table 2 shows absorbed radiation rates for M. albumplants in different plots. It can be seen that, in all plotswithin the EURT zone, the contribution of internalradiation exposure markedly exceeds that of externalexposure. Since calculations made with the ERICATool program do not take into account the level of nat�ural background radiation (NBR), their results showabsorbed dose rates from 90Sr and 137Cs accumulatedin soils and plants. Absorbed radiation doses inclusiveof NBR exceed the background level by factors of 2 to18 in buffer plots and by factors of 330 to 418 in impactplots. The contribution of NBR to the total dose loadon M. album plants in background plots reached 95–96%, compared to 5–45% in buffer plots and 0.2–0.3% in impact plots.
Viability of Melandrium album Seed Progeny
The seed progeny of the 2005 yield was characteri�zed by a high seedling viability (Fig. 1a), with samplesfrom different cenopopulations showing no significantdifferences in this parameter (H(3) = 5.55, р = 0.142,n = 16). Such differences were revealed later, after theonset of apical and root meristem functioning. Thus,the rate of leaf formation in seedlings from irradiatedcenopopulations (Buffer�2 and Impact�1) was four tofive times lower than in the background sample(F�test, p 0.01). Root growth in all seedlings fromEURT cenopopulations was retarded, compared tothe control (F(4; 639) = 92.7, p 0.01), especially insamples from the buffer zone (p 0.001). A similarpicture was observed with respect to the relative num�
�
��
100
90
80
70
60
50
40
30
20
10
0
100
90
80
70
60
50
40
30
20
10
0
100
90
80
70
60
50
40
30
20
10
0
100
90
80
70
60
50
40
30
20
10
0Impact�1Background�2 Background�2
Background�2 Background�2Background�1
Background�3Buffer�1
Buffer�1
Buffer�1
Impact�1
Impact�2Impact�1
Buffer�2 Impact�2Impact�2
Buffer�2
Via
bili
ty,
%
1
2
(a) (b)
(c)(d)
Fig. 1. Parameters of viability of the seed progeny of Melandrium album plants from cenopopulations of the EURT zone and back�ground plots in (a) 2005, (b) 2007, (c) 2008, and (d) 2010: (1) seedling survival rate, (2) number of seedlings with true leaves.

22
RUSSIAN JOURNAL OF ECOLOGY Vol. 44 No. 1 2013
ANTONOVA et al.
ber of seedlings with lateral roots: their proportion inchronically irradiated cenopopulations varied from 6to 23%, compared to 42% in background cenopopula�tions (p 0.01).
In 2007, samples from different cenopopulationsdid not differ in the rates of seedling survival and leafformation (H(2) = 0.50–0.97, p = 0.23–0.78, n = 24)(Fig. 1b). Similar data on seedling survival (H(3) = 2.7–2.94, p = 0.44, n = 104) and leaf formation (U�test;p = 0.08, n1 = 25, n2 = 79) were also obtained in 2008(Fig. 1c). Root length in the seed progeny from EURTcenopopulations was greater than in seedlings fromthe background sample (F(3; 3283) = 49.9, p 0.01).
In 2010, all M. album seed progeny were highly via�ble, irrespective of the site of maternal plant growth(Fig. 1d), with samples from different cenopopulationsbeing similar in the rate of seedling survival (F(5; 18) =2.11, p = 0.11) and root length (F�test, p = 0.89).However, factor “site of maternal plant growth”proved to have a significant effect on the rate of trueleaf formation, either during analysis of the entire dataset (F(5; 18) = 6.99, p = 0.0009) or in pairwise compari�sons between background (p = 0.0018), buffer (p 0.01), and impact samples (p = 0.035).
The seeds from cenopopulation Buffer�1 in 2010were distinctly differentiated into three size fractions(large, medium, and small seeds), while no such dif�ferentiation was observed in other samples. To evaluatethe relationship between seed size and quality, we ger�minated these three fractions separately from eachother. The survival rate of seedlings proved to be highin the group grown from large seeds (73.9 ± 8.9%),intermediate in the group from medium�sized seeds(50.3 ± 10%), and low in the group from small seeds(29.7 ± 1.3%), with differences between the groupsbeing statistically significant (H(2) = 8.9, p = 0.013, n =12). Similar differences between the groups wereobserved with respect to growth parameters such as theproportion of seedlings with true leaves and the growthrate of the main and lateral roots. These results wellagree with data presented in the review by Moles andWestoby (2004), which provide evidence for the directdependence of seedling survival on seed weight. InM. album, seed size depends on the biotic factor,namely, pollination: large seeds are formed when theflower is pollinated by pollen from one male plant, andmedium�sized seeds, when the pollen comes from twomale plants (Burkhardt, Internicola, and Bernasconi,2009). The fact that plants from chronically irradiatedcenopopulation produce a certain proportion of verysmall seeds with low viability may be evidence forradiation mutagenesis, which is responsible for distur�bances in processes of seed maturation resulting indevelopment of dwarfism characters (Breslavets,1946).
Thus, our 4�year observations provided no definiteanswer concerning the effect of chronic radiationexposure on the viability of M. album seed progeny.
�
�
�
Statistically significant inhibition of growth parame�ters in the gradient of radioactive contamination wasrecorded only in 2005. No such effect took place insubsequent years; on the contrary, even stimulation ofroot growth was observed. Since the radiation load onplants barely changed during the study period and,according to geobotanical data, the state of phyto�cenoses was also stable, it appeared that changes in theset of weather conditions may be the main factorresponsible for interannual variation in the quality ofM. album seeds.
Effect of Weather Conditions on the Quality of Melandrium album Seed Progeny
During the 4�year observation period, the survivalof seed progeny from background samples was foundto directly depend on the monthly average and effec�tive temperatures of June in the year of seed formation(R = 0.85–0.91, p = 0.001–0.032). Weather condi�tions in other months had no effect on the survival ofseedlings. In impact cenopopulations, seed qualityshowed dependence on weather parameters in June,including the monthly average temperature and thesum of effective temperatures (R = 0.88–0.89, p =0.0002–0.0004) and also monthly precipitation andthe sum of precipitation at effective temperatures (R =–0.70 to –0.78, p = 0.005–0.02). In July and August,precipitation was the main factor (R = –0.61 to –0.71,p = 0.014–0.048): the lower the amount of precipita�tion, the higher the quality of seed progeny.
When weather conditions in all three summermonths (over the season) were included in analysis,neither temperature nor precipitation was found tohave a significant effect on the viability of seedlings(true leaf formation) in background samples (R =0.19–0.80, p = 0.054–0.72; R = –0.17 to –0.73, p =0.097–0.74). However, the growth of roots showeddependence on the average temperature over the sea�son (R = 0.89, p = 0.025) and the sum of effective tem�peratures over the season (R = 0.87, p = 0.024). Insamples from impact cenopopulations, both factorshad a significant effect on all growth parameters of theseed progeny (R = 0.66–0.92, p = 0.00005–0.027; R =–0.81 to –0.92, p = 0.00006–0.0023).
Selyaninov’s index takes into account both tem�perature and precipitation. We used it to compare theresponses of background and impact samples toweather conditions in June and revealed significantdifferences between them (Fig. 2). Moreover, parame�ters of seedling viability in background samplesshowed no dependence on this index, either in eachsummer month or over the season (R = –0.74 to⎯0.11, p = 0.09–0.90). In chronically irradiated sam�ples from the EURT zone, such a situation wasobserved only in August (R = –0.60 to –0.49, p =0.053–0.118), with correlations between seedling via�bility and Selyaninov’s index in other months and inthe summer season as a whole being negative and sta�

RUSSIAN JOURNAL OF ECOLOGY Vol. 44 No. 1 2013
INTRASPECIFIC VARIATION IN Melandrium album 23
tistically significant (R = –0.70 to –0.94, p =0.00001–0.02).
Thus, similar tendencies were revealed in back�ground and impact cenopopulations of M. album: inboth cases, the quality of seeds was found to have (1) apositive correlation with temperature in the year oftheir formation, (2) a negative correlation with theamount of precipitation, and (3) a negative correlationwith Selyaninov’s index. Since regression lines for thecorrelations with precipitation and Selyaninov’s inte�grated index are similar, it appears that precipitation isthe leading factor and that it is the low amount of pre�cipitation at the end of the season that provided for theformation of high�quality seeds. It should be empha�sized that the dependence of seed quality on weatherconditions was better manifested in the seed progenyfrom the EURT zone than from background plots.
Similar data were obtained in our previous studyperformed in the EURT zone on Stellaria graminea,another species from the family Caryophyllaceae(Pozolotina, Antonova, and Karimullina, 2010). Acomparison between these two species showed that in2010, when summer was hot and dry, the seeds ofS. graminea failed to develop, while M. album pro�duced high�quality seeds. This could be explained byinterspecific differences in ecological features (resis�tance to drought and high temperatures) and in themore of reproduction: S. graminea is capable of bothseed and vegetative reproduction, while M. albumreproduces mainly by seeds.
Radiosensitivity of Melandrium album Seed Progeny
In 2005, the dose–effect relationship after acuteirradiation of seeds from cenopopulation Impact�1was described by the classic S�shaped curve, withseedling mortality reaching a maximum at doses of300–400 Gy (F�test, p 0.01). On the other hand, thesurvival of seedlings from buffer plots remained almostunchanged after seed irradiation at any dose tested(F(4; 15) = 0.621–1.09, p = 0.396–0.655). Seeds frombackground plots occupied an intermediate positionwith respect to radiosensitivity.
Analysis of data on root length showed that theeffect of irradiation on this parameter depended on theinitial rate of root growth. Root length in seedlingsfrom buffer cenopopulations was relatively small evenwithout preliminary irradiation of seeds; after irradia�tion, it changed only slightly, decreasing to the lowerlimit of the norm. In samples Background andImpact�2, root length gradually decreased with anincrease in the dose of acute irradiation, with thisdependence being statistically significant (F(4; 15) =25.4–151.1, p 0.001). The dose of 200 Gy could beconsidered critical, since it resulted in the practicalcessation of root and leaf growth. It is noteworthy thatthe proportion of seedlings with lateral roots in sam�ples from Background and Impact�2 populationsreached 41.6% and 23.2%, respectively, compared toonly 6–9% in populations Buffer�1 and Buffer�2.
In 2007, seeds from both background populationsproved to be resistant to acute irradiation according tothe rates of seedling survival and leaf formation. In thesample from population Impact�2, however, thedose–effect curve was saw�toothed, with 20% stimu�
�
�
70
60
50
40
30
20
10
02.60.2 2.42.22.01.81.61.41.21.00.80.60.4
Root length, mm
1
2
Selyaninov’s index
у(impact) = –25.45x + 60.98R2 = 0.88
у(background) = –9.34x + 46.18R2 = 0.103
Fig. 2. Dependence of root length on Selyaninov’s index in Melandrium album seedlings from (1) impact and (2) backgroundcenopopulations (data obtained in June).
24
RUSSIAN JOURNAL OF ECOLOGY Vol. 44 No. 1 2013
ANTONOVA et al.
lation of seedling survival taking place after irradiationat 100 and 300 Gy (U�test, p = 0.027, n = 8). In 2008,statistically significant inhibition of seedling survivalafter acute irradiation of seeds was observed in all sam�ples (F(1; 34–39) = 6.74–22.95, p = 0.00003–0.013),except for Impact�2 (F(1; 28) = 2.9, p = 0.099); the for�mation of true leaves was also retarded in all samples(F(1; 29–39) = 22.98–190.1, p 0.00002).
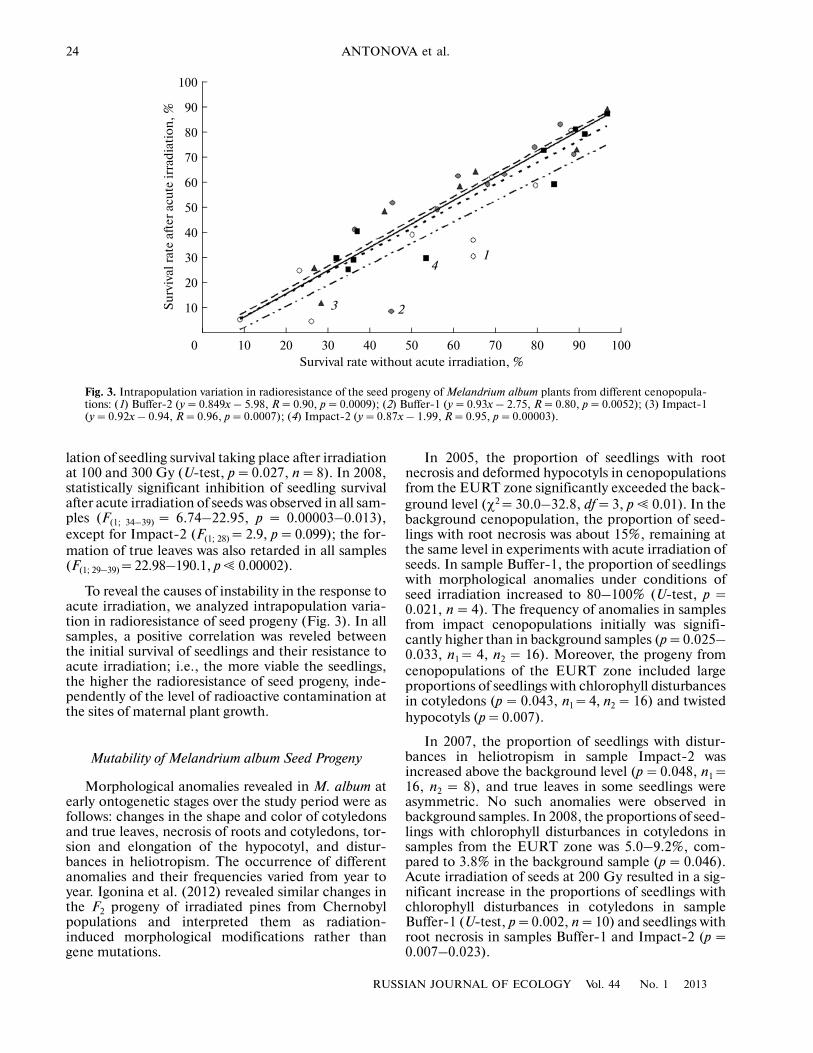
To reveal the causes of instability in the response toacute irradiation, we analyzed intrapopulation varia�tion in radioresistance of seed progeny (Fig. 3). In allsamples, a positive correlation was reveled betweenthe initial survival of seedlings and their resistance toacute irradiation; i.e., the more viable the seedlings,the higher the radioresistance of seed progeny, inde�pendently of the level of radioactive contamination atthe sites of maternal plant growth.
Mutability of Melandrium album Seed Progeny
Morphological anomalies revealed in M. album atearly ontogenetic stages over the study period were asfollows: changes in the shape and color of cotyledonsand true leaves, necrosis of roots and cotyledons, tor�sion and elongation of the hypocotyl, and distur�bances in heliotropism. The occurrence of differentanomalies and their frequencies varied from year toyear. Igonina et al. (2012) revealed similar changes inthe F2 progeny of irradiated pines from Chernobylpopulations and interpreted them as radiation�induced morphological modifications rather thangene mutations.
�
In 2005, the proportion of seedlings with rootnecrosis and deformed hypocotyls in cenopopulationsfrom the EURT zone significantly exceeded the back�ground level (χ2 = 30.0–32.8, df = 3, p 0.01). In thebackground cenopopulation, the proportion of seed�lings with root necrosis was about 15%, remaining atthe same level in experiments with acute irradiation ofseeds. In sample Buffer�1, the proportion of seedlingswith morphological anomalies under conditions ofseed irradiation increased to 80–100% (U�test, p =0.021, n = 4). The frequency of anomalies in samplesfrom impact cenopopulations initially was signifi�cantly higher than in background samples (p = 0.025–0.033, n1 = 4, n2 = 16). Moreover, the progeny fromcenopopulations of the EURT zone included largeproportions of seedlings with chlorophyll disturbancesin cotyledons (p = 0.043, n1 = 4, n2 = 16) and twistedhypocotyls (p = 0.007).
In 2007, the proportion of seedlings with distur�bances in heliotropism in sample Impact�2 wasincreased above the background level (p = 0.048, n1 =16, n2 = 8), and true leaves in some seedlings wereasymmetric. No such anomalies were observed inbackground samples. In 2008, the proportions of seed�lings with chlorophyll disturbances in cotyledons insamples from the EURT zone was 5.0–9.2%, com�pared to 3.8% in the background sample (p = 0.046).Acute irradiation of seeds at 200 Gy resulted in a sig�nificant increase in the proportions of seedlings withchlorophyll disturbances in cotyledons in sampleBuffer�1 (U�test, p = 0.002, n = 10) and seedlings withroot necrosis in samples Buffer�1 and Impact�2 (p =0.007–0.023).
�
100
70
60
50
40
30
20
10
100605040200 10 30 908070
90
80
Survival rate without acute irradiation, %
Su
rviv
al r
ate
afte
r ac
ute
irra
dia
tio
n,
%
1
23
4
Fig. 3. Intrapopulation variation in radioresistance of the seed progeny of Melandrium album plants from different cenopopula�tions: (1) Buffer�2 (y = 0.849x – 5.98, R = 0.90, p = 0.0009); (2) Buffer�1 (y = 0.93x – 2.75, R = 0.80, p = 0.0052); (3) Impact�1(y = 0.92x – 0.94, R = 0.96, p = 0.0007); (4) Impact�2 (y = 0.87x – 1.99, R = 0.95, p = 0.00003).

RUSSIAN JOURNAL OF ECOLOGY Vol. 44 No. 1 2013
INTRASPECIFIC VARIATION IN Melandrium album 25
In 2010, disturbances in the shape of cotyledons inM. album seedlings occurred most frequently. Theirfrequency in samples from EURT cenopopulationswas significantly higher than in background samples(p = 0.007–0.009, n1 = 8, n2 = 4), which did not differsignificantly in this parameter (U�test, p = 0.124, n = 4).Moreover, the proportion of seedlings with twistedhypocotyls in samples from cenopopulations chroni�cally exposed to radiation significantly exceeded thebackground value (1.5%), with the difference being2.4�fold in sample Buffer�1 (p = 0.041) and 1.5�fold insample Impact�1 (p = 0.027).
Seeds from cenopopulation Buffer�1 were dividedinto three size groups (see above), which producedseedlings with different frequencies of anomalies. Forexample, mutation monopteros (Lutova et al., 2000)was recorded in only 1% of viable seedlings obtainedfrom large seeds, compared to 11.2% and 32.5% ingroups of seedlings from medium�sized and smallseeds, respectively.
Samples from the EURT frontal part (plots Buffer�2 and Inpact�2) were found to contain hermaphroditeplants (Fig. 4), which did not occur in backgroundareas. Seeds collected from such plants were sown inautumn in uncontaminated experimental plots withuniform agricultural background. All plants thatemerged next spring were hermaphrodites, indicatingthat this trait was hereditary. Sexual dimorphism inM. album is under control of two gene regions on theY chromosome (Ye et al., 1991), and the induction ofmutations (deletions) in this chromosome by means ofγ�irradiation of pollen results in a sharp increase in theoccurrence frequency of hermaphrodite plants (Lar�don et al., 1999). Cases of sex reversal in M. albumunder the effect of radiation are described, with her�maphrodite flowers appearing as a result of gynoeciumdevelopment in male flowers (Davies and Wall, 1961).In general, sex ratio regulation in M. album popula�
tions is itself an individual ecological problem(Purrington, 1993; Lyons et al., 1995; Taylor, Trimble,and McCauley, 1999).
On the whole, the results of this study show that themutability of M. album plants and their seed progenyfrom cenopopulations of the EURT zone is increasedsignificantly, compared to the background level. Inview of previous data on the mutational origin of rootnecrosis (Tomilov et al., 2001), cotyledon necrosis(Tomilova et al., 2001), and twisted (wavy) shoots(Karpinskaya and Buzovkina, 2005), we attribute theincreased frequencies of these anomalies at earlystages of M. album seedling development to themutagenic effect of small radiation doses. Similarresults were obtained in our previous studies oncenopopulations of S. graminea, Taraxacum officinale,and other plant species growing in the EURT zone(Pozolotina et al., 2008; Pozolotina, Antonova, andKarimullina, 2010).
CONCLUSIONS
(1) Dose loads on M. album plants in the EURTfrontal part exceed the background level by one tothree orders of magnitude. Parameters characterizingthe viability of M. album seed progeny vary from yearto year, with their variation depending not so much onthe level of radiation exposure as on changes inweather conditions or, more specifically, on the inter�action of these two factors. The dependence of seedquality on weather conditions is much stronger insamples from the EURT zone than in backgroundsamples.
(2) Sustainable adaptation to radiation exposurehas not been revealed in M. album samples from theEURT zone. The seed progeny of this species is highlyviable, which provides for its increased resistance toadditional acute irradiation regardless of the level of
(а) (b) (c)
Fig. 4. Flowers of Melandrium album: (a) female, (b) hermaphrodite, (c) male.

26
RUSSIAN JOURNAL OF ECOLOGY Vol. 44 No. 1 2013
ANTONOVA et al.
radioactive contamination in the habitat of maternalplants.
(3) Hermaphrodite plants occur in M. albumcenopopulations growing in the EURT zone, whereasin the norm this species is dioecious, with distinct sex�ual dimorphism. Analysis of F1 progeny has shownthat seeds from hermaphrodite parents produce onlyhermaphrodite plants; i.e., this trait is heritable.Mutability of M. album seed progeny in samples fromEURT cenopopulations is in most cases higher than inbackground samples. Additional acute irradiation isinstrumental in revealing hidden genetic disturbances.
ACKNOWLEDGMENTS
This study was supported by the Program of Inter�disciplinary Research between Institutes of the UralBranch of the Russian Academy of Sciences (projectno. 12�M�24�2016), Scientific School Support Pro�gram (project no. NSh�5325.2012.4), and Program forGoal�Oriented Basic Research Cooperation betweenInstitutes of the Ural Branch of the Russian Academyof Sciences and Russian State Corporations (projectno. 12�4�002�NC), and Russian Foundation for BasicResearch (project no. 12�04�31769).
REFERENCES
Abramov, V.I., Stepanova, A.A., and Famelis, S.A., Radio�biological Effects in Plants from the Territory of the EasternUral Radioactive Trace, Radiats. Biol. Radioekol., 2010,vol. 50, no. 3, pp. 345–351.Antonova, E.V. and Pozolotina, V.N., Clonal Structure ofDandelion Cenopopulations under Conditions of Radioac�tive Contamination in the Urals, Radiats. Biol. Radioekol.,2007, vol. 47, no. 3, pp. 349–354.Baker, H.G., Accounts of Melandrium, M. dioicum andM. album for the Biological Flora of the British Isles Spon�sored by the British Ecological Society, J. Ecol., 1947, vol. 35,pp. 271–292.Beresford, N., Brown, J., Copplestone, D., et al.,D�ERICA: An Integrated Approach to the Assessment andManagement of Environmental Risks from Ionizing Radia�tion: Description of Purpose, Methodology and Application,European Commission under the Euratom Research andTraining Programme on Nuclear Energy within the SixthFramework Programme, 2007.Beresford, N.A., Barnett, C.L., Howard, B.J., et al., Deri�vation of Transfer Parameters for Use within the ERICATool and the Default Concentration Ratios for TerrestrialBiota, J. Environ. Radioact., 2008, vol. 99, no. 9, pp. 1393–1407.Bernasconi, G., Antonovics, J., Biere, A., et al., Silene As aModel System in Ecology and Evolution, Heredity, 2009,vol. 103, no. 1, pp. 5–14.
Breslavets, L.P., Rasteniya i luchi rentgena (Plants and X�Rays), Leningrad: Akad, Nauk SSSR, 1946.Brown, J.E., Alfonso, B., Avila, R., et al., The ERICATool, J. Environ. Radioact., 2008, vol. 99, no. 9, pp. 1371–1383.
Burkhardt, A., Internicola, A., and Bernasconi, G., Effectsof Pollination Timing on Seed Paternity and Seed Mass inSilene latifolia (Caryophyllaceae), Ann. Bot., 2009, vol. 104,no. 4, pp. 767–773.
Davies, D.R. and Wall, E.T., The Effect of Gamma Irradi�ation on the Sex Ratio of Melandrium Species, Heredity,1961, vol. 16, pp. 131–136.
Delph, L.F., Arntz, A.M., Scotti�Saintagne, C., et al., TheGenomic Architecture of Sexual Dimorphism in the Dioe�cious Plant Silene latifolia, Evolution, 2010, vol. 64, no. 10,pp. 2873–2886.
Geras’kin, S.A., Kim, J.K., Dikarev, V.G., et al., Cytoge�netic Effects of Combined Radioactive (Cs�137) andChemical (Cd, Pb, and 2,4�D Herbicide) Contaminationon Spring Barley Intercalar Meristem Cells, Mutat. Res.Genet. Toxicol. Environ. Mutagen., 2005, vol. 586, no. 2,pp. 147–159.
Gulenkova, M.A. and Pyatunina, S.K., The White Cam�pion, in Biologicheskaya flora Moskovskoi oblasti (Biologi�cal Flora of Moscow Oblast), Moscow: Polieks, 1997,vol. 13, pp. 88–96.
Igonina, E.V., Fedotov, I.S., Korotkevich, A.Yu., andRubanovich, A.V., Morphological Anomalies in theProgeny of Irradiated Scots Pine (Pinus sylvestris L.) fromChernobyl Populations, Radiats. Biol. Radioekol., 2012,vol. 52, no. 1, pp. 90–102.
Jolivet, C. and Bernasconi, G., Within/Between Popula�tion Crosses Reveal Genetic Basis for Siring Success inSilene latifolia (Caryophyllaceae), J. Evol. Biol., 2007, vol. 20,no. 4, pp. 1361–1374.
Kal’chenko, V.A., Rubanovich, A.V., and Shevchenko, V.A.,Genetic Processes in Chronically Irradiated Populations ofCentaurea scabiosa L. from the Eastern Ural RadioactiveTrace, Radiats. Biol. Radioekol., 1995, vol. 37, no. 5,pp. 236–243.
Karpinskaya, L.I. and Buzovkina, I.S., Genetic Control ofthe Wavy Shoots Character in Radish Raphanus sativus L.Russ. J. Genet., 2005, vol. 41, no. 9, pp. 1028–1034.
Lardon, A., Georgiev, S., Aghmir, A., et al., Sexual Dimor�phism in White Campion: Complex Control of CarpelNumber Is Revealed by Y Chromosome Deletions, Genetics,1999, vol. 151, pp. 1173–1185.
Lutova, L.A., Provorov, N.A., Tikhodeev, O.N., et al.,Genetika razvitiya rastenii (Genetics of Plant Develop�ment), St. Petersburg: Nauka, 2000.
Lyons, E.E., Shahmahoney, N., and Lombard, L.A., Evo�lutionary Dynamics of Sex�Ratio and Gender Dimorphismin Silene latifolia: 2. Sex�Ratio and Flowering Status in aPotentially Male�Biased Population, J. Hered., 1995, vol. 86,no. 2, pp. 107–113.
Lysenko, E.A., Kal’chenko, V.A., and Shevchenko, V.A.,Variation in Polymorphic Systems of Centaurea scabiosa L.under the Effect of Chronic Irradiation, Radiats. Biol.,Radioekol., 1999, vol. 39, no. 6, pp. 623–629.
Molchanova, I., Pozolotina, V., Karavaeva, E., et al.,Radioactive Inventories within the East Ural RadioactiveState Reserve on the Southern Urals, Radioprotection, 2009,vol. 44, no. 5, pp. 747–757.
Moles, A.T. and Westoby, M., Seedling Survival and SeedSize: A Synthesis of the Literature, J. Ecol., 2004, vol. 92,no. 3, pp. 372–383.

RUSSIAN JOURNAL OF ECOLOGY Vol. 44 No. 1 2013
INTRASPECIFIC VARIATION IN Melandrium album 27
Nikipelov, B.V., Mikerin, E.I., Romanov, G.N., et al., The1957 Radiation Accident in the Southern Urals and Subse�quent Recovery Operation, in Recovery Operation in theEvent of a Nuclear Accident or Radiological Emergency,Vienna: IAEA, 1989, pp. 373–403.Petin, V.G., Zhurakovskaya, G.P., and Komarova, L.N.,Radiobiologicheskie osnovy sinergicheskikh vzaimodeistvii vbiosfere (Radiobiological Foundations of Synergistic Inter�actions in the Biosphere), Moscow: GEOS, 2012.Pozolotina, V.N., Otdalennye posledstviya deistviya radiatsiina rasteniya (Remote Consequences of Radiation Impacton Plants), Yekaterinburg: Akademkniga, 2003.Pozolotina, V.N., Molchanova, I.V., Karavaeva, E.N.,et al., Sovremennoe sostoyanie nazemnykh ekosistem zonyVostochno�Ural’skogo radioaktivnogo sleda (Current State ofTerrestrial Ecosystems in the Zone of the Eastern UralRadioactive Trace), Yekaterinburg: Goshchitskii, 2008.Pozolotina, V.N., Antonova, E.V., Karimullina, E.M.,et al., Consequences of Chronic Radiation Exposure for theFlora of the Eastern Ural Radioactive Trace, Radiats. Biol.Radioekol., 2009, vol. 49, no. 1, pp. 97–106.Pozolotina, V.N., Antonova, E.V., and Karimullina, E.M.,Assessment of Radiation impact on Stellaria graminea Cen�opopulations in the Zone of the Eastern Ural RadioactiveTrace, Russ. J. Ecol., 2010, vol. 41, no. 6, pp. 459–468.Pozolotina, V.N., Molchanova, I.V., Karavaeva, E.N.,et al., Radionuclides in Terrestrial Ecosystems of the Zoneof Kyshtym Accident in the Urals, J. Environ. Radioact.,2010, vol. 101, pp. 438–442.Purrington, C.B., Parental Effects on Progeny Sex�Ratio,Emergence, and Flowering in Silene latifolia (Caryophyl�laceae), J. Ecol., 1993, vol. 81, no. 4, pp. 807–811.Rautenberg, A., Hathaway, L., Oxelman, B., et al., Geo�graphic and Phylogenetic Patterns in Silene Section Melan�drium (Caryophyllaceae) As Inferred from Chloroplast andNuclear DNA Sequences, Mol. Phylogenet. Evol., 2010,vol. 57, no. 3, pp. 978–991.
Shevchenko, V.A., Pechkurenkov, V.L., and Abramov, V.I.,Radiatsionnaya genetika prirodnykh populyatsii. Geneti�cheskie posledstviya Kyshtymskoi avarii (Radiation Geneticsof Natural Populations: Genetic Consequences of theKyshtym Accident), Moscow: Nauka, 1992.
Steven, J.C., Delph, L.F., and Brodie, E.D., Sexual Dimor�phism in the Quantitative�Genetic Architecture of Floral,Leaf, and Allocation Traits in Silene latifolia, Evolution,2007, vol. 61, no. 1, pp. 42–57.
Taylor, D.R., Trimble, S., and McCauley, D.E., EcologicalGenetics of Gynodioecy in Silene vulgaris: Relative Fitnessof Females and Hermaphrodites During the ColonizationProcess, Evolution, 1999, vol. 53, no. 3, pp. 745–751.
The Encyclopedia of World Climatology (Encyclopedia ofEarth Sciences Series), Oliver, E.J., Ed., Dordrecht:Springer, 2005.
Tomilov, A.A., Tomilova, N.V., Ogarkova, O.A., and Tara�sov, V.A., Identification of a Gene Involved in the Control ofthe Root System Development in Arabidopsis thaliana,Russ. J. Genet., 2001, vol. 37, no. 1, pp. 30–38.
Tomilova, N.V., Tomilov, A.A., Ogarkova, O.A., and Tara�sov, V.A., Identification of a Mutant Gene ControllingNecrotic Cotyledons in Developing Seedlings of Arabidop�sis thaliana, Russ. J. Genet., 2001, vol. 37, no. 4, pp. 392–400.
Vilenchik, M.M., Radiobiologicheskie effekty i okruzhay�ushchaya sreda (Radiobiological Effects and the Environ�ment), Moscow: Energoatomizdat, 1991.
Wolfe, L.M., Why Alien Invaders Succeed: Support for theEscape�from�Enemy Hypothesis, Am. Nat., 2002, vol. 160,no. 6, pp. 705–711.
Ye, D., Oliveira, M., Veuskens, J., et al., Sex Determinationin the Dioecious Melandrium: The X/Y Chromosome Sys�tem Allows Complementary Cloning Strategies, Plant Sci.,1991, vol. 80, pp. 93–106.
Copyright © 2022 FDOKUMEN




















