
Intramolecular hydrogen bonding as a determinant of the inhibitory potency of N-unsubstituted...
9
Intramolecular hydrogen bonding as a determinant of the inhibitory potency of N-unsubstituted imidazole derivatives towards mammalian hemoproteinsw Lionel Perrin, a Franc¸ois Andre´, a Caroline Aninat, b Re´my Ricoux, c Jean-Pierre Mahy,* c Ning Shangguan, d Madeleine M. Joullie´ d and Marcel Delaforgez* a Received 8th October 2008, Accepted 27th October 2008 First published as an Advance Article on the web 27th November 2008 DOI: 10.1039/b817743k Enzymes involved in the mammalian microsomal metabolism of drugs are, in numerous cases, inhibited by compounds bearing an imidazolyl scaffold. However, the inhibition potency is highly dependent upon the accessibility of the imidazolyl nitrogen lone pair. In order to highlight some structural parameters of inhibitors that control this phenomenon, a series of compounds containing a nitrogen unsubstituted imidazolyl moiety with varying degrees of nitrogen lone pair accessibility was tested on human and rat hepatic cytochromes P450 and microperoxidase 8, an enzymatically active peptide derived from cytochrome c. In each case, we have shown that the accessibility of the imidazole lone pair determined the extent of inhibition. Nitrogen accessibility was tuned not only by varying the steric hindrance in the vicinity of the imidazolyl ring but also by modifying its surrounding hydrogen bonding network. Compounds in which there exists intramolecular hydrogen bonding between the imidazole moiety and an H-bond acceptor, such as an appropriately positioned amide carbonyl group, demonstrated enhanced inhibitory effects. Conversely, imidazole moieties which are in proximity to H-bond donors, such as an amide NH group, displayed reduced potency. This trend was observed in cyclo-peptide derivatives in which the intramolecular H-bond network was adjusted through the modification of the stereochemistry of a dehydrohistidine residue. It was observed that (Z)-isomers weakly bind heme, whereas (E)-isomers demonstrated higher degrees of metal binding. Therefore, enzymatic inhibition of heme-containing proteins by compounds bearing a dehydrohistidine motif seems to be closely related to its stereochemistry and hydrogen binding propensity. At neutral pH, these differences in binding affinities can be confidently attributed to the ambident H-bond properties of imidazole nitrogen atoms. This structure-activity relationship may be of use for the design of novel imidazolyl compounds as new P450 inhibitors or drug candidates. Introduction Metalloproteins represent an important class of enzymes which are involved in a variety of biochemical processes such as peroxidase, catalase, monooxygenase and reduc- tase activities, and oxygen transport and storage. These functions are related to the presence of a metal center tightly bound to amino acids such as methionine, cysteine or histidine. 1 In the case of hemoproteins, four N-pyrrolyl ligands coordi- nate the iron in the plane of the porphyrin ring, a fifth axial position is coordinated by a histidine, methionine or cysteine residue, and the sixth by an electron rich molecule such as dioxygen, cyanide, amine, nitroso derivatives or water. In certain cases, an external molecule containing an electron rich functional group may serve as a metal ligand, leading to inhibitory effects. The extent to which such molecules possess an affinity for complexation to the heme iron determines the degree of inhibition. 2,3 Considering the binding features of iron in hemoproteins, as well as the importance of their biological functions, these proteins are relevant targets for studying inhibitory effects. In particular, cytochromes P450 are highly attractive since a large panel of isoforms is able to recognize a wide variety of substrates. Nitrogen rich compounds, such as imidazole derivatives, represent a large class of cytochrome P450 rever- sible inhibitors. 2–5 Their inhibitory potency comes from their ability to form stable Fe(II)-N-imidazole complexes. 6 a URA 2096 du CNRS, Institut de Biologie et Technologies de Saclay, Commissariat a ` l’Energie Atomique, Ba ˆtiment 528, CEA-Saclay, 91191 Gif-sur-Yvette, France. E-mail: [email protected]; Fax: +33 1 69 08 87 17; Tel: +33 1 69 08 68 39 b INSERM U620; Universite ´ de Rennes 1, IFR 140, 2 Avenue du Pr. Le ´on Bernard, 35043 Rennes, France c Laboratoire de Chimie Bioorganique et Bioinorganique, CNRS, Institut de Chimie Mole ´culaire et des Mate ´riaux d’Orsay, UMR 8182, Ba ˆtiment 420, Universite ´ Paris-Sud XI, 91405 Orsay, France. E-mail: [email protected]; Fax: +33 1 69 15 72 81; Tel: +33 1 69 15 74 21 d Department of Chemistry, University of Pennsylvania, 231 South 34th Street, Philadelphia, Pennsylvania 19104-6323, USA w Electronic supplementary information (ESI) available: HPLC separation and MS-MS spectra of roquefortine and its metabolites. See DOI: 10.1039/b817743k z Marcel Delaforge, CEA-Saclay, iBiTec-S SB2SM, CNRS URA2096, baˆt. 528, 91191 Gif-sur-Yvette, France. 148 | Metallomics, 2009, 1, 148–156 This journal is c The Royal Society of Chemistry 2009 PAPER www.rsc.org/metallomics | Metallomics Published on 27 November 2008. Downloaded on 11/02/2014 10:03:51. View Article Online / Journal Homepage / Table of Contents for this issue
Transcript of Intramolecular hydrogen bonding as a determinant of the inhibitory potency of N-unsubstituted...

Intramolecular hydrogen bonding as a determinant of the inhibitory
potency of N-unsubstituted imidazole derivatives towards
mammalian hemoproteinsw
Lionel Perrin,aFrancois Andre,
aCaroline Aninat,
bRemy Ricoux,
c
Jean-Pierre Mahy,*cNing Shangguan,
dMadeleine M. Joullie
d
and Marcel Delaforgez*a
Received 8th October 2008, Accepted 27th October 2008
First published as an Advance Article on the web 27th November 2008
DOI: 10.1039/b817743k
Enzymes involved in the mammalian microsomal metabolism of drugs are, in numerous cases,
inhibited by compounds bearing an imidazolyl scaffold. However, the inhibition potency is highly
dependent upon the accessibility of the imidazolyl nitrogen lone pair. In order to highlight some
structural parameters of inhibitors that control this phenomenon, a series of compounds
containing a nitrogen unsubstituted imidazolyl moiety with varying degrees of nitrogen lone pair
accessibility was tested on human and rat hepatic cytochromes P450 and microperoxidase 8, an
enzymatically active peptide derived from cytochrome c. In each case, we have shown that the
accessibility of the imidazole lone pair determined the extent of inhibition. Nitrogen accessibility
was tuned not only by varying the steric hindrance in the vicinity of the imidazolyl ring but also
by modifying its surrounding hydrogen bonding network. Compounds in which there exists
intramolecular hydrogen bonding between the imidazole moiety and an H-bond acceptor, such as
an appropriately positioned amide carbonyl group, demonstrated enhanced inhibitory effects.
Conversely, imidazole moieties which are in proximity to H-bond donors, such as an amide NH
group, displayed reduced potency. This trend was observed in cyclo-peptide derivatives in which
the intramolecular H-bond network was adjusted through the modification of the stereochemistry
of a dehydrohistidine residue. It was observed that (Z)-isomers weakly bind heme, whereas
(E)-isomers demonstrated higher degrees of metal binding. Therefore, enzymatic inhibition of
heme-containing proteins by compounds bearing a dehydrohistidine motif seems to be closely
related to its stereochemistry and hydrogen binding propensity. At neutral pH, these differences in
binding affinities can be confidently attributed to the ambident H-bond properties of imidazole
nitrogen atoms. This structure-activity relationship may be of use for the design of novel
imidazolyl compounds as new P450 inhibitors or drug candidates.
Introduction
Metalloproteins represent an important class of enzymes
which are involved in a variety of biochemical processes
such as peroxidase, catalase, monooxygenase and reduc-
tase activities, and oxygen transport and storage. These
functions are related to the presence of a metal center tightly
bound to amino acids such as methionine, cysteine or histidine.1
In the case of hemoproteins, four N-pyrrolyl ligands coordi-
nate the iron in the plane of the porphyrin ring, a fifth axial
position is coordinated by a histidine, methionine or cysteine
residue, and the sixth by an electron rich molecule such as
dioxygen, cyanide, amine, nitroso derivatives or water. In
certain cases, an external molecule containing an electron rich
functional group may serve as a metal ligand, leading to
inhibitory effects. The extent to which such molecules possess
an affinity for complexation to the heme iron determines the
degree of inhibition.2,3
Considering the binding features of iron in hemoproteins, as
well as the importance of their biological functions, these
proteins are relevant targets for studying inhibitory effects.
In particular, cytochromes P450 are highly attractive since a
large panel of isoforms is able to recognize a wide variety of
substrates. Nitrogen rich compounds, such as imidazole
derivatives, represent a large class of cytochrome P450 rever-
sible inhibitors.2–5 Their inhibitory potency comes from
their ability to form stable Fe(II)-N-imidazole complexes.6
aURA 2096 du CNRS, Institut de Biologie et Technologies de Saclay,Commissariat a l’Energie Atomique, Batiment 528, CEA-Saclay,91191 Gif-sur-Yvette, France. E-mail: [email protected];Fax: +33 1 69 08 87 17; Tel: +33 1 69 08 68 39
b INSERM U620; Universite de Rennes 1, IFR 140,2 Avenue du Pr. Leon Bernard, 35043 Rennes, France
c Laboratoire de Chimie Bioorganique et Bioinorganique, CNRS,Institut de Chimie Moleculaire et des Materiaux d’Orsay, UMR8182, Batiment 420, Universite Paris-Sud XI, 91405 Orsay, France.E-mail: [email protected]; Fax: +33 1 69 15 72 81;Tel: +33 1 69 15 74 21
dDepartment of Chemistry, University of Pennsylvania, 231 South34th Street, Philadelphia, Pennsylvania 19104-6323, USA
w Electronic supplementary information (ESI) available: HPLCseparation and MS-MS spectra of roquefortine and its metabolites.See DOI: 10.1039/b817743kz Marcel Delaforge, CEA-Saclay, iBiTec-S SB2SM, CNRSURA2096, bat. 528, 91191 Gif-sur-Yvette, France.
148 | Metallomics, 2009, 1, 148–156 This journal is �c The Royal Society of Chemistry 2009
PAPER www.rsc.org/metallomics | Metallomics
Publ
ishe
d on
27
Nov
embe
r 20
08. D
ownl
oade
d on
11/
02/2
014
10:0
3:51
. View Article Online / Journal Homepage / Table of Contents for this issue

Numerous imidazole derivatives were investigated for their
ability to inhibit hemoproteins such as cytochromes P450,2,4–9
cyclo-oxygenases10 and NO-synthases.11 They are also known
and used for their potent inhibition of lanosterol demethylase
activity in fungi.7,8 In addition, this inhibitory action has
been fruitfully exploited to reduce the rate of metabolism of
certain drugs. For example, it has been demonstrated that
co-administration of ketoconazole with cyclosporine A9 or
ixabepilone12 limits their metabolism and enhances their
therapeutic effects. Finally, another example of heme-
imidazole complexes is found in the case of monooxygenase
inhibition by histamine, a natural derivative of histidine.13,14
It has been demonstrated that the stability of the reversible
imidazole–iron interaction depends upon the accessibility of
imidazolyl nitrogen atoms as well as the hydrophobicity of its
surrounding substituents.15–18 Among the imidazole-based
inhibitors, most of the molecules reported have one of their
two nitrogen atoms substituted.15 In this case, the inhibition
potency is enhanced by the coordination of the electron lone
pair of the remaining nitrogen of the imidazole ring. This effect
has been demonstrated, experimentally and theoretically, in
the case of the binding ofN-substituted imidazoles to the heme
of P450cam.19
We have previously demonstrated that small peptides, and
particularly cyclodipeptides, are recognized, and in some cases
metabolized, by hepatic cytochromes P450.20,21 In the present
study, we describe our efforts to demonstrate the influence of
the chemical environment of imidazole derivatives on their
P450 inhibition. In particular, since it is clear that the strength
of the coordination of the imidazole nitrogen on the iron atom
is related to the availability of its doublet, we examine the
influence of its involvement in a H-bond with a neighboring
hydrogen atom, which is supposed to decrease its availability
and its ability to bind to the iron atom. To this end, a
determination of the preferred mode of interaction between
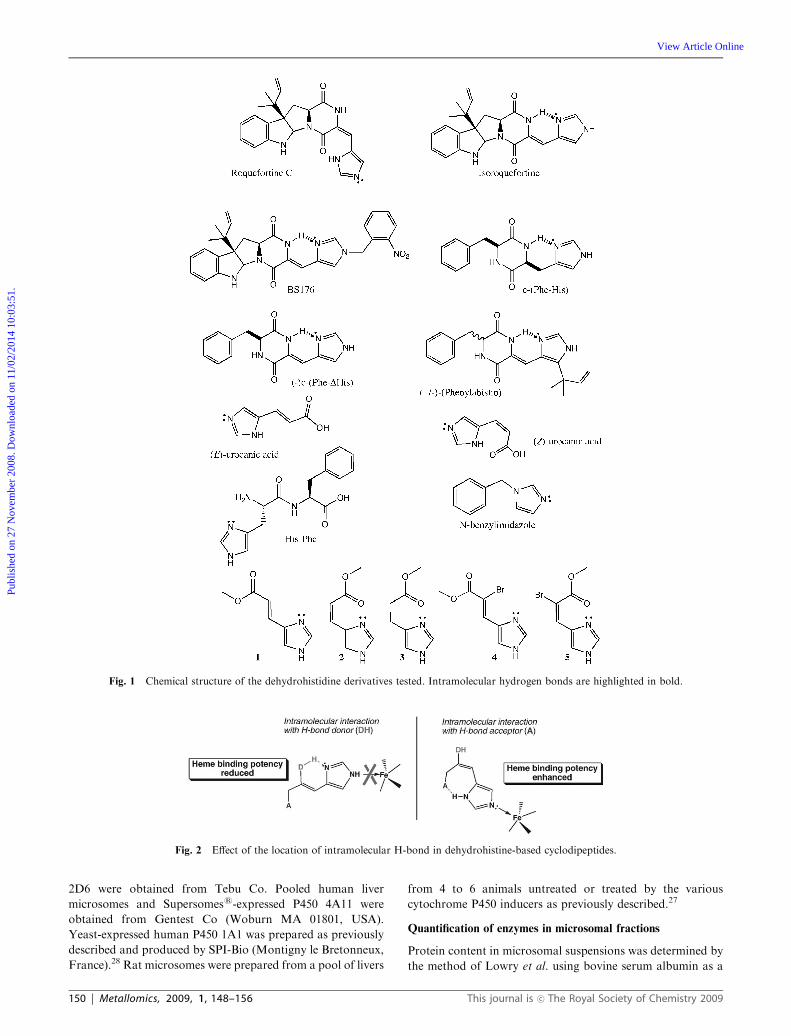
a series of dehydrohistidine-based molecules (Fig. 1) and
various P450 isoforms has been performed. In this series of
compounds, firstly a group of molecules which cannot estab-
lish any intramolecular hydrogen bonds was studied. This
subset includes (E)-urocanic acid, its methyl ester (1) and
a-bromo derivative (4), His-Phe and N-benzylimidazole. A
second series of compounds which potentially develop intra-
molecular hydrogen bonds consists of roquefortine, iso-
roquefortine, phenylahistin, BS176, c-(Phe-His), c-(Phe-DHis),
(Z)-urocanic acid, its cis methyl ester (2) and a-bromo deri-
vatives (5), as well as the saturated derivative 3. In the latter
group, two types of behavior may occur: one of the two
nitrogens of the imidazole ring may act either as a hydrogen
bond donor or acceptor. An example of the first case is found
in roquefortine, whereas an example of the second case is
illustrated by isoroquefortine.
These differences in H-bonding properties between
roquefortine and isoroquefortine are supported by NMR
analysis (M Joullie, personal observations). In roquefortine,
hydrogen bonding between the NH-group of imidazole
and the oxygen of the neighboring carbonyl group of the
diketopiperazine has been suggested,21 whereas intramolecular
hydrogen bonding between the sp2 nitrogen¼N– of the imidazole
group and the NH-amide group of the diketopiperazine (Fig. 1)
has been proposed for isoroquefortine.22 In terms of binding
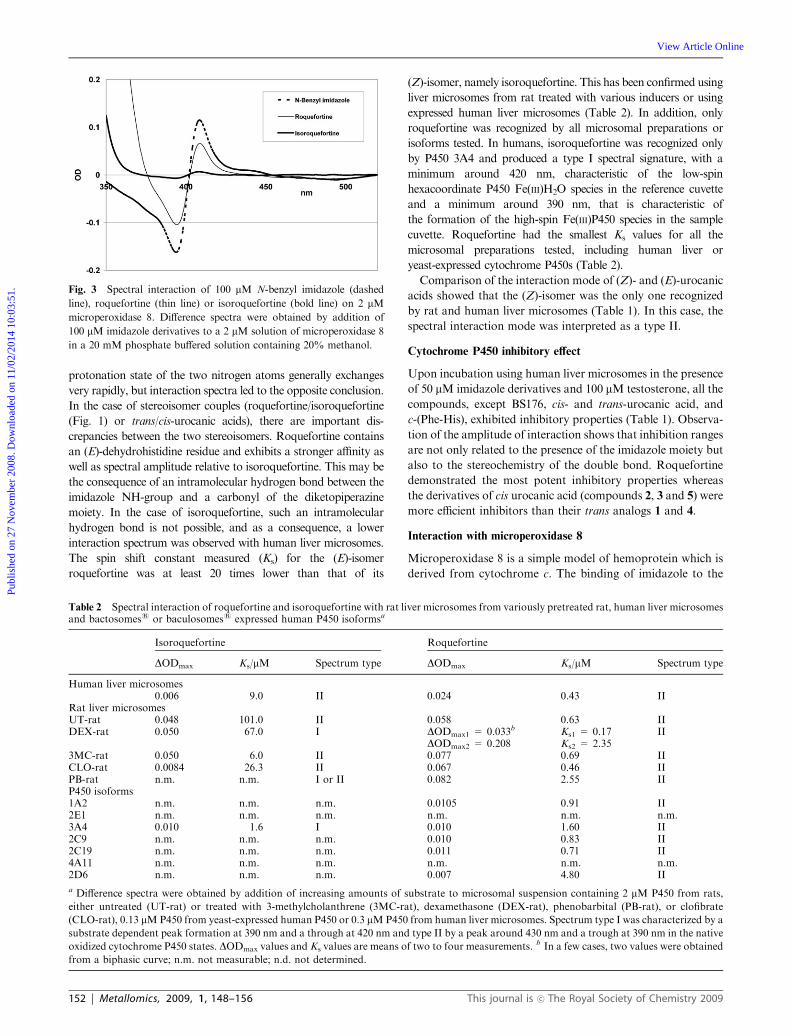
affinities, we have previously demonstrated the inhibitory
action of roquefortine towards cytochromes P45021 and
attributed this inhibitory action to the binding of the basic
sp2 nitrogen lone pair of roquefortine to the heme iron atom
(see mechanism in Fig. 2).
In order to gain insight into this structure–activity relation-
ship in the case of (iso)roquefortine and to extend it to a series
of analogs, a metabolism and a binding study to P450s has
been undertaken. Isoroquefortine should possess a lower
inhibitory potential than roquefortine due to the reduced
accessibility of its sp2 nitrogen. Similarly, the (Z)-isomer of
urocanic acid and its derivatives 2, 3 and 5 would be expected
to exhibit hydrogen bonding similar to that of roquefortine
(Fig. 1) and should display markedly increased inhibitory
activity relative to its (E)-isomer and its derivatives 1 and 4.
In order to validate this assumption, heme spectral interaction
and inhibitory studies were performed in the presence of
different human isoforms of cytochrome P450 and on rat or
human liver microsomes. Additionally, in order to dis-
criminate between P450 molecular recognition and heme
binding strength, the same compounds were tested on a simple
hemoprotein model, microperoxidase 8 (MP8).23,24
Microperoxidase 8 is obtained by hydrolytic digestion of
cytochrome c. It contains the heme prosthetic group as well
as the amino acid residues 14–21 of horse cytochrome c,
including His 18, whose imidazole group acts as the fifth axial
ligand of the iron. The absence of a heme iron ligand at the
sixth coordination position allows this site to be occupied by a
number of different ligands. In aqueous solution water
occupies the sixth coordination site.23 Due to these structural
characteristics, microperoxidase 8 is considered to be an
effective model for the study of the redox mechanism of
hemoproteins such as cytochromes, hemoglobin, myoglobin
and peroxidases.23
Experimental
Chemicals
Roquefortine, trans-(E)-urocanic acid, N-benzylimidazole,
His-Phe, testosterone, 6b-hydroxy-testosterone, NADPH,
NADP, glucose-6-phosphate (G6P), glucose-6-phosphate
dehydrogenase (G6PDH), dexamethasone (DEX), clofibrate
(CLO), sodium phenobarbital (PB), 3-methylcholanthrene
(3-MC), and microperoxidase 8 (MP8) were obtained from
Sigma Chemical. c-(Phe-His) was obtained from Bachem.,
BS176, compounds 1–5 and isoroquefortine were synthesized
as previously described.22 c-(Phe-DHis) and (�/+)-phenylahistin
were synthesized and kindly provided by Y. Hayashi25 and
cis-(Z)-urocanic acid by H. Morrison.26 All other chemicals were
of the highest quality commercially available.
Microsomal preparations
Human liver samples were kindly supplied by the surgery
service of the Franche Comte Hospital (University of Besancon)
and microsomes were prepared as previously described.27
Bactosomess-expressed P450 3A4, 2E1, 1A2, 2C19, 2C9 and
This journal is �c The Royal Society of Chemistry 2009 Metallomics, 2009, 1, 148–156 | 149
Publ
ishe
d on
27
Nov
embe
r 20
08. D
ownl
oade
d on
11/
02/2
014
10:0
3:51
.
View Article Online
2D6 were obtained from Tebu Co. Pooled human liver
microsomes and Supersomess-expressed P450 4A11 were
obtained from Gentest Co (Woburn MA 01801, USA).
Yeast-expressed human P450 1A1 was prepared as previously
described and produced by SPI-Bio (Montigny le Bretonneux,
France).28 Rat microsomes were prepared from a pool of livers
from 4 to 6 animals untreated or treated by the various
cytochrome P450 inducers as previously described.27
Quantification of enzymes in microsomal fractions
Protein content in microsomal suspensions was determined by
the method of Lowry et al. using bovine serum albumin as a
Fig. 1 Chemical structure of the dehydrohistidine derivatives tested. Intramolecular hydrogen bonds are highlighted in bold.
Fig. 2 Effect of the location of intramolecular H-bond in dehydrohistine-based cyclodipeptides.
150 | Metallomics, 2009, 1, 148–156 This journal is �c The Royal Society of Chemistry 2009
Publ
ishe
d on
27
Nov
embe
r 20
08. D
ownl
oade
d on
11/
02/2
014
10:0
3:51
.
View Article Online

standard.29 The P450 concentration was measured as described
by Omura and Sato.30
Substrate binding to P450s microsomal isoforms
and to microperoxidase 8
Substrate binding to cytochrome P450 was studied by
difference visible spectroscopy using 2 mM P450 from rat liver
microsomes or 0.3 mM P450 from human liver microsomes
in 0.1 M phosphate buffer (pH 7.4). Bactosomess and
Supersomess-expressed P450 were suspended in 0.1 M phos-
phate buffer (pH 7.4) to obtain a P450 concentration of
0.13 mM. The solution was equally divided between two
cuvettes (1 mL cuvette for microsomes and 100 mL cuvette
for Bactosomess and Supersomess). Low amounts of human
liver were used due to their high protein concentrations and
turbidity. After recording the baseline, aliquots (0.5–1 mL) ofsubstrate solutions were added to the sample cuvette, and the
same volume of solvent (DMSO) was added to the reference
cuvette. Difference spectra were recorded between 350 and
520 nm at room temperature, with a Perkin Elmer l18 UV
spectrophotometer equipped with a turbidity accessory. Ks
and DODmax were calculated by linear regression from double
reciprocal plots of 1/DOD versus 1/[substrate].31 Micro-
peroxidase 8 preparations at 2 mM were performed in
20 mM phosphate buffer solutions at pH 6, 7.4 or 8.1 containing
20% methanol in order to obtain a homogenous solution.23
Spectral interactions were measured in a differential mode as for
hepatic microsomal preparations. Control measurements were
done in simple beam spectrometry in order to evaluate the
absolute high spin to low spin transition upon addition of
imidazole derivative.23,24
Cytochrome P450 activities
Incubations of human liver microsomal suspensions (1 mg
protein/mL) were done in the presence of 100 mM of testoster-
one and 0.5 mM of NADPH in a 0.1 M phosphate buffer
(pH 7.4). Experiments were performed at 37 1C in the presence
of 50 mM of each tested compound. Incubations were stopped
after 15 min by the addition of an equivalent volume of
acetonitrile. The final mixture was cooled and centrifuged at
10 000 rpm for 10 minutes. HPLC analysis was performed as
previously indicated; testosterone and 6b-hydroxytestosteronewere detected at 254 nm.21
Inhibition of microperoxidase 8 activity
The peroxidase activity of MP8 was assayed by measurement
of the kinetics of oxidation of o-dianisidine by H2O2.32 The
assay system contained o-dianisidine (100 mM) and MP8
(0.22 mM) in phosphate buffer (0.1 M, pH 7.4) at 25 1C in
the presence or absence of the imidazole derivatives. The
reaction was initiated by addition of hydrogen peroxide
(90 mM). The concentration of oxidized o-dianisidine was
determined by the change in absorbance at 500 nm using a
molar extinction coefficient of 6360 M�1 cm�1.
Molecular modeling
Quantum mechanics calculations have been carried out at the
DFT-B3LYP level of theory.33–35 C, H, O and N atoms have
been represented by an all-electron triple-z quality basis set:
6-311G(d,p).36 A relativistic electron core potential and its asso-
ciated basis set have been used for Br.37 A d polarization function
has been added for Br (a = 0.389). All calculations have been
achieved with the Gaussian 98 suite of programs.38 Geometries
have been fully optimized without any symmetry restrictions. For
each molecule, all major conformers have been optimized and
compared. Only the most stable conformers of the configuration
of interest are presented. The nature of the extrema has been
checked by an analytical calculation of the frequencies.
Results
Recognition of imidazole derivatives by P450
Spectral evaluation of the binding of substrates, using human
liver microsomes, revealed a type II interaction for all the
substrates tested except for BS176, and trans-(E)-urocanic acid
(Table 1). This type II interaction, which is characterized by a
spectral absorption around 430 nm, is characteristic of the
formation of a heme–imidazole complex. The dissociation
constants can be calculated for compounds with values bet-
ween 0.4 and 20 mM (Table 1). These affinities are in the same
range as those of N-substituted imidazoles such as miconazole
for which Ks values were determined to be lower than 1 mM.39
The strongest amplitude spectrum is observed with N-benzyl-
imidazole (Fig. 3) for which the two imidazole nitrogen atoms are
chemically non-equivalent and the accessible nitrogen is in its sp2
hybridization state. For all the other compounds measured, the
Table 1 UV spectral interaction of imidazole derivatives with humanliver microsomes
Compound
Spectral interaction% inhibition
DODmax Ks/mMSpectrumtype
of testosteronehydroxylation
Roquefortine 0.024 0.4 II 88Isoroquefortine 0.006 9.0 II 66BS176 n.m. n.m. n.m. o5c-(Phe-His) 0.053 n.m. II 5c-(Phe-DHis) 0.030 n.m. II 50(�)-Phenylahistin 0.011 9.0 II 54trans-(E)-Urocanicacid
n.m. n.m n.m o5
cis-(Z)-Urocanicacid
0.047 20.0 II o5
Compound 1 0.015 1.4 II 19Compound 2 0.018 4.6 II 46Compound 3 0.017 1.4 II 27Compound 4 0.016 1.0 II 19Compound 5 0.019 0.95 II 71His-Phe 0.037 n.m. II 25N-Benzylimidazole 0.084 2.7 II 48
Spectral interactions were measured at room temperature using
1 mg prot/mL (0.3 nmol P450) of a pool of 10 human liver microsomes
in phosphate buffer. Spectrum type II was characterized by a substrate
dependent formation of a peak around 430 nm and a trough at 390 nm
in the native oxidized cytochrome P450 state. DODmax values were
expressed as nmol P450 and Ks values were determined from double
reciprocal plots from spectra obtained using 0.5 to 100 mM substrate.
Incubations of 100 mM testosterone were performed in the presence of
50 mM imidazole derivatives.
This journal is �c The Royal Society of Chemistry 2009 Metallomics, 2009, 1, 148–156 | 151
Publ
ishe
d on
27
Nov
embe
r 20
08. D
ownl
oade
d on
11/
02/2
014
10:0
3:51
.
View Article Online
protonation state of the two nitrogen atoms generally exchanges
very rapidly, but interaction spectra led to the opposite conclusion.
In the case of stereoisomer couples (roquefortine/isoroquefortine
(Fig. 1) or trans/cis-urocanic acids), there are important dis-
crepancies between the two stereoisomers. Roquefortine contains
an (E)-dehydrohistidine residue and exhibits a stronger affinity as
well as spectral amplitude relative to isoroquefortine. This may be
the consequence of an intramolecular hydrogen bond between the
imidazole NH-group and a carbonyl of the diketopiperazine
moiety. In the case of isoroquefortine, such an intramolecular
hydrogen bond is not possible, and as a consequence, a lower
interaction spectrum was observed with human liver microsomes.
The spin shift constant measured (Ks) for the (E)-isomer
roquefortine was at least 20 times lower than that of its
(Z)-isomer, namely isoroquefortine. This has been confirmed using
liver microsomes from rat treated with various inducers or using
expressed human liver microsomes (Table 2). In addition, only
roquefortine was recognized by all microsomal preparations or
isoforms tested. In humans, isoroquefortine was recognized only
by P450 3A4 and produced a type I spectral signature, with a
minimum around 420 nm, characteristic of the low-spin
hexacoordinate P450 Fe(III)H2O species in the reference cuvette
and a minimum around 390 nm, that is characteristic of
the formation of the high-spin Fe(III)P450 species in the sample
cuvette. Roquefortine had the smallest Ks values for all the
microsomal preparations tested, including human liver or
yeast-expressed cytochrome P450s (Table 2).
Comparison of the interaction mode of (Z)- and (E)-urocanic
acids showed that the (Z)-isomer was the only one recognized
by rat and human liver microsomes (Table 1). In this case, the
spectral interaction mode was interpreted as a type II.
Cytochrome P450 inhibitory effect
Upon incubation using human liver microsomes in the presence
of 50 mM imidazole derivatives and 100 mM testosterone, all the
compounds, except BS176, cis- and trans-urocanic acid, and
c-(Phe-His), exhibited inhibitory properties (Table 1). Observa-
tion of the amplitude of interaction shows that inhibition ranges
are not only related to the presence of the imidazole moiety but
also to the stereochemistry of the double bond. Roquefortine
demonstrated the most potent inhibitory properties whereas
the derivatives of cis urocanic acid (compounds 2, 3 and 5) were
more efficient inhibitors than their trans analogs 1 and 4.
Interaction with microperoxidase 8
Microperoxidase 8 is a simple model of hemoprotein which is
derived from cytochrome c. The binding of imidazole to the
Fig. 3 Spectral interaction of 100 mM N-benzyl imidazole (dashed
line), roquefortine (thin line) or isoroquefortine (bold line) on 2 mMmicroperoxidase 8. Difference spectra were obtained by addition of
100 mM imidazole derivatives to a 2 mM solution of microperoxidase 8
in a 20 mM phosphate buffered solution containing 20% methanol.
Table 2 Spectral interaction of roquefortine and isoroquefortine with rat liver microsomes from variously pretreated rat, human liver microsomesand bactosomess or baculosomess expressed human P450 isoformsa
Isoroquefortine Roquefortine
DODmax Ks/mM Spectrum type DODmax Ks/mM Spectrum type
Human liver microsomes0.006 9.0 II 0.024 0.43 II
Rat liver microsomesUT-rat 0.048 101.0 II 0.058 0.63 IIDEX-rat 0.050 67.0 I DODmax1 = 0.033b Ks1 = 0.17 II
DODmax2 = 0.208 Ks2 = 2.353MC-rat 0.050 6.0 II 0.077 0.69 IICLO-rat 0.0084 26.3 II 0.067 0.46 IIPB-rat n.m. n.m. I or II 0.082 2.55 IIP450 isoforms1A2 n.m. n.m. n.m. 0.0105 0.91 II2E1 n.m. n.m. n.m. n.m. n.m. n.m.3A4 0.010 1.6 I 0.010 1.60 II2C9 n.m. n.m. n.m. 0.010 0.83 II2C19 n.m. n.m. n.m. 0.011 0.71 II4A11 n.m. n.m. n.m. n.m. n.m. n.m.2D6 n.m. n.m. n.m. 0.007 4.80 II
a Difference spectra were obtained by addition of increasing amounts of substrate to microsomal suspension containing 2 mM P450 from rats,
either untreated (UT-rat) or treated with 3-methylcholanthrene (3MC-rat), dexamethasone (DEX-rat), phenobarbital (PB-rat), or clofibrate
(CLO-rat), 0.13 mMP450 from yeast-expressed human P450 or 0.3 mMP450 from human liver microsomes. Spectrum type I was characterized by a
substrate dependent peak formation at 390 nm and a through at 420 nm and type II by a peak around 430 nm and a trough at 390 nm in the native
oxidized cytochrome P450 states. DODmax values and Ks values are means of two to four measurements. b In a few cases, two values were obtained
from a biphasic curve; n.m. not measurable; n.d. not determined.
152 | Metallomics, 2009, 1, 148–156 This journal is �c The Royal Society of Chemistry 2009
Publ
ishe
d on
27
Nov
embe
r 20
08. D
ownl
oade
d on
11/
02/2
014
10:0
3:51
.
View Article Online

heme functionality of microperoxidase 8 has already been
reported and discussed.23,24 N-Benzylimidazole was used as
a reference compound for the formation of a low spin
N-imidazole–iron complex (absorption at 409 nm) from the
high spin aqua-complex (broad absorption peak around
394 nm) (Fig. 3). In differential mode visible spectroscopy,
all the tested compounds except BS176 and phenylahistin gave
a 394 to 409 nm shift (Table 3). This spectral shift was
concentration-dependent. His-Phe, c-(Phe-His) and com-
pound 3, which have mobility between the amide function
and the imidazole moiety and can form an intramolecular
CQO� � �HN(Im) bond (Fig. 2), gave similar high spin to low
spin transition ratios with respect to N-benzylimidazole,
whereas c-(Phe-DHis), which forms an intramolecular
NH� � �N(Im) bond, exhibited lower interaction spectrum.
(Z)- and (E)-urocanic acid and compounds 4 and 5 showed
similar spectral transitions of approximately 50% of that of
N-benzylimidazole. The strongest differences between E and
Z configurations were observed in the roquefortine series.
Roquefortine led to around 60% transition spectra whereas
isoroquefortine showed around 5% (pH 7.4). Similarly the
cis-urocanic methyl ester 2 led to a 4 times stronger inter-
action spectrum than its trans isomer 1. BS176, which is a
N-substituted isoroquefortine, was unable to significantly
modify the microperoxidase 8 spin state.
The concentration of imidazole–iron complexes may be
related to the decrease of the peroxidase activity, since
N-benzylimidazole, His-Phe, c-(Phe-His), compound 3,
roquefortine and (Z)-urocanic acid inhibited more than 60%
of the initial rate of o-dianisidine peroxidation. Isoroquefortine,
c-(Phe-DHis) and (E)-urocanic acid and its derivatives 1, 2, 4
and 5 showed either no or low inhibition of such activity
(Table 3). BS176, which is unable to form a significant
concentration of microperoxidase complex, is still able to
inhibit microperoxidase activity. We have observed that this
compound can be oxidized under such conditions, it could
then act as a competitive inhibitor for the oxidation of
ortho-dianisidine.
Cytochrome P450 metabolism of imidazole derivatives
Liver microsomes from control or dexamethasone pretreated
rats in combination with a NADPH-generating system were
used for the metabolism study. Under these conditions, only
substituted cyclodipeptides were metabolized. Isoroquefortine,
and phenylahistin, were significantly metabolized (turnover
values being around 2 nmol metabolized min�1 nmol P450�1),
whereas c-(Phe-His), c-(Phe-DHis) and His-Phe were not.
Metabolism of urocanic acid derivatives was not studied.
Using human liver microsomes, more than 60% of
isoroquefortine was metabolized in three major mono-
hydroxylated metabolites, the main one being hydroxylated
on the isoprenyl function (see figures A and B in the ESIw)whereas roquefortine yielded almost no metabolite (see figure
A in ESIw). This isoroquefortine metabolism is only observed
in the presence of NADPH and was inhibited by ketoconazole,
a classical P450 3A inhibitor. Other P450 inhibitors such as
furafylline (1A2), quinidine (2D6) and sulfaphenazole (2C9)
had no significant effects on the formation of metabolites.
Molecular modeling
In order to obtain more information concerning the relative
stability of the different tautomers of roquefortine, urocanic
acids and their derivatives, the energy-minimized structures of
these molecules have been determined by quantum mechanics
at the DFT-level. Fig. 4 indicates the most stable isomers of
each protonation state and their relative energy in kcal mol�1.
In the gas phase, the more favorable conformation of iso-
roquefortine is slightly more stable than that of roquefortine
by 2.8 kcal mol�1. This can be explained by a lesser cyclic
constraint in isoroquefortine (H-bond based on a 6 membered
ring) than in roquefortine (H-bond based on a 7-membered
ring). However, considering the precision of the calculation
level, these two isomers have to be considered as isoenergetic.
For both isomers, the more stable tautomer is the one that can
develop an intramolecular hydrogen bond. The loss of this
hydrogen bond leads to structures with increased energies
of 7.3 kcal mol�1 (isoroquefortine) and 12.6 kcal mol�1
(roquefortine). In these two molecules, the imidazole ring
has a different role with respect to the establishment of a
hydrogen bond. In isoquefortine, imidazole acts as an H-bond
acceptor, whereas it acts as a donor in roquefortine. Hence,
roquefortine can act as a Lewis base and efficiently bind a
metal center whereas isoroquefortine cannot.
The same analysis can be performed in the case of urocanic
acids and their methyl ester and a-brominated derivatives.
Whereas the two tautomers of (E)-urocanic acid are iso-
energetic, one isomer of (Z)-urocanic acid is much more stable
than the others. This isomer is characterized by an H-bond
Table 3 UV spectral interaction of imidazole derivatives with micro-peroxidase 8
Compound
Spectral interactiona Inhibition ofperoxidaseactivitydDOD409–394 nm Percentb
N-Benzylimidazole 0.28 100 58Roquefortine 0.17 61 76Isoroquefortine 0.014 5 0BS176 o0.005 n.d.c 60c-(Phe-His) 0.29 103 80c-(Phe-DHis) 0.057 20 0(�)-Phenylahistin n.d.c n.d.c 0trans-(E)-Urocanic acid 0.16 57 18cis-(Z)-Urocanic acid 0.19 68 69Compound 1 0.03 11 25Compound 2 0.12 43 39Compound 3 0.29 105 55Compound 4 0.07 37 30Compound 5 0.12 44 13His-Phe 0.34 121 41
a Difference spectra were obtained by addition of 100 mM imidazole
derivatives to a 2 mM solution of microperoxidase 8 in a 20 mM
phosphate buffered solution containing 20% methanol. b Percentage
of the spectral interaction obtained with N-benzylimidazole which
possesses only one accessible tertiary nitrogen. c n.d. no detectable
peak shift. d Percentage of the initial velocity of peroxidase activity
obtained using 100 mM o-dianisidine as substrate, 90 mM H2O2,
0.22 mM microperoxidase 8, 100 mM imidazole derivatives in 0.1 M
phosphate buffer, pH 7.4 at 25 1C. Peroxidase activity was 23 nmol
o-dianisidine consumed min�1 nmol microperoxidase�1. Values are
the mean of two to four independent determinations.
This journal is �c The Royal Society of Chemistry 2009 Metallomics, 2009, 1, 148–156 | 153
Publ
ishe
d on
27
Nov
embe
r 20
08. D
ownl
oade
d on
11/
02/2
014
10:0
3:51
.
View Article Online

between the imidazole (acting as an H-bond donor) and the
carboxylic function (acting as the complementary acceptor).
Hence the imidazole ring of this isomer can behave as a Lewis
base and efficiently bind the iron atom of heme. Noteworthily,
the reverse situation, in which the imidazole ring acts as
an acceptor and the carboxylic as a donor is slightly less
favorable. The same computational study applied to the
urocanic acid methyl esters and the a-brominated derivatives
leads to very similar results. In each case, the Z conformation
in which the imidazole group acts as an H-bond donor and the
carbonyl group as an H-bond acceptor is the most stable
configuration.
Finally, the conformational sampling of the dihydro isomer
3 revealed the absence of an intramolecular H-bond. This
result shows that the energy that could be gained through
H-bonding cannot compensate the cost associated with the
formation of a 7-membered ring.
Discussion
Imidazole derivatives are largely used as hemoprotein inhibi-
tors, and the most effective compounds generally contain
N-substituted imidazoles.15 A few unsubstituted imidazole
derivatives exhibit weak inhibitory effects mainly related to
the rapid proton exchange between the two imidazole nitrogens.
To our knowledge, this report represents the first study
of the hemoprotein inhibitory properties of dehydrohistidine-
based derivatives in which nitrogen atoms are not substituted.
Our studies demonstrate that these compounds interact with
the heme iron of cytochrome P450 and microperoxidase 8.
Formation of the iron–imidazole complex is sufficiently stable
to perturb the catalytic cycle of such hemoproteins. Inhibitory
potency is dependent upon the formation of intramolecular
hydrogen bonds between either imidazole nitrogen and
the surrounding CO or NH groups of amide functions.
Comparison of the inhibitory effects between c-(Phe-His)
and c-(Phe-DHis) (Table 3) clearly demonstrates that this
intramolecular interaction is associated with the planarity
imposed on the molecule by the Ca–Cb double bond of the
dehydro-residue. The (E)-configuration allows strong hydrogen
bonding between the diketopiperazine carbonyl and the
imidazole NH group. Therefore, the imidazole sp2 nitrogen
is accessible to bind the heme iron. In contrast, in the
(Z)-configuration, the basic nitrogen atom of imidazole is
inaccessible due to hydrogen bonding with a vicinal NH
group, which lowers the binding potency to the heme iron
atom. This interpretation of the effect of the molecular
environment on the binding of ligand to heme is strongly
supported by quantum mechanical optimized structures of the
different isomers of the molecules herein studied.
Such discrepancies in the affinities of (Z)- and (E)-dehydro-
histidines arise from differences in imidazole nitrogen
accessibility. As indicated by NMR and molecular modeling,
(Z)-isomers possess an imidazole nitrogen doublet, which is
involved in a hydrogen bond with a NH group of the
diketopiperazine (Fig. 2 and 4). This conformation greatly
minimizes the availability of the imidazole nitrogen doublet
for heme iron binding. The accessible nitrogen doublet of the
imidazole shows only a low affinity for iron association.15–18 In
contrast, in (E)-isomers such as roquefortine, the hydrogen
atom of the protonated imidazole nitrogen is bonded to the
oxygen of the diketopiperazine carbonyl group (Fig. 2 and 4),
making the lone pair of the remaining nitrogen atom available
for binding to heme iron.
In general, (Z)-dehydrohistidine isomers are preferentially
formed by dehydration of histidine derivatives.40 In the same
way, natural dehydro-amino acids are predominantly found in
the (Z)-configuration.41–44 Interestingly, some methods for
the preparation of dehydro-diketopiperazines have been
recently patented.45 Alternatively, examples of natural
Fig. 4 Relative energies of tautomers of roquefortine/isoroquefortine and urocanic acids. The relative energies are given with respect to the more
stable isomer and are expressed in kcal mol�1.
154 | Metallomics, 2009, 1, 148–156 This journal is �c The Royal Society of Chemistry 2009
Publ
ishe
d on
27
Nov
embe
r 20
08. D
ownl
oade
d on
11/
02/2
014
10:0
3:51
.
View Article Online

(E)-dehydrohistidine are few, and roquefortine is an exception
that has been of interest due to its antibacterial activities.10
The first total synthesis of roquefortine was recently reported
using a novel elimination strategy.46
In the urocanic acid series, cis isomers exhibit higher
inhibition strength towards P450s and MP8. This result is in
agreement with the structure of the most stable tautomers of
these cis-compounds in which the imidazole ring acts as an
H-bond donor with respect to the intramolecular carbonyl
group. Experimentally, the inhibition potency of these
compounds is reduced by changing the stereochemistry of
the double bond from cis to trans (Z to E). In trans isomers,
no intramolecular H-bond can be developed, hence both
imidazole tautomers are in equilibrium, which results in a
decrease of binding strength. Finally, compound 3 highlights
the sterical constraints induced by a,b-saturation on the
nitrogen lone pair accessibility. Indeed, the planarity of the
dehydro-compounds causes the Ca–H or Cb–H to point
towards a direction similar to that of the imidazole lone pair
and, hence, decreases the binding strength of the molecule
compared to compound 3 in which the planarity is lost.
Our results suggest intriguing possibilities for the develop-
ment of hemoprotein inhibitors, which may be deactivated by
simple structural modification. Such structural modifications
can easily be obtained under light exposure or saturation
reduction. In the case of urocanic acid isomers, further
research is in progress in an attempt to correlate hemoprotein
inhibition properties to in vivo modifications of physiological
parameters and precisely address the impact of the substitution
on the a and/or b position in the dehydro series.
Acknowledgements
Part of this work was supported by the AC 009E Environne-
ment et Sante program of the French Ministere de l’Ecologie,
de l’Energie , du Developpement durable et de l’Amenagement
du Territoire. We also wish to thank Ms S. Blondel for
her excellent technical assistance. The synthetic work was
supported by NSF (CHEM 0515443).
References
1 S. J. Lippard and J. M. Berg, Principles of Bioinorganic Chemistry,Science Book University, Mill Valley, 1994.
2 P. R. Ortiz de Montellano and M. A. Correia, in CytochromeP450: Structure, Mechanism and Biochemistry, ed. P. R. Ortiz deMontellano, Plenum Press, New York, 1995, pp. 305–364.
3 S. D. Zaric, D. M. Popovic and E. W. Knapp, Biochemistry, 2001,40, 7914–7928.
4 B. Testa and P. Jenner, Drug Metab. Rev., 1981, 12, 1–117.5 M. Murray and A. J. Ryan, Xenobiotica, 1983, 13, 707–714.6 W. Zhang, Y. Ramamoorthy, T. Kilicarslan, H. Nolte,R. F. Tyndale and E. M. Sellers, Drug Metab. Dispos., 2002, 30,314–318.
7 H. Van den Bossche, P. Marichal, J. Gorrens and M. C. Coene, Br.J. Clin. Pract. Suppl., 1990, 71, 41–46.
8 E. Albengres, H. Le Louet and J. P. Tillement, Drug Saf., 1998, 18,83–97.
9 M. A. Abraham, P. P. Thomas, G. T. John, V. Job, V. Shankarand C. K. Jacob, Transplant. Proc., 2003, 35, 215–216.
10 P. Cozzi, G. Branzoli, C. Carganico, C. Ferti, A. Pillan,D. Severino and R. Tonani, Eur. J. Med. Chem., 1991, 26, 423–433.
11 D. J. Wolff, G. A. Datto, R. A. Samatovicz and R. A. Tempsick,J. Biol. Chem., 1993, 268, 9425–9429.
12 S. Goel, M. Cohen, S. N. Comezoglu, L. Perrin, F. Andre,D. Jayabalan, L. Iacono, A. Comprelli, V. T. Ly, D. Zhang,C. Xu, W. G. Humphreys, H. McDaid, G. Goldberg,S. B. Horwitz and S. Mani, Clin. Cancer Res., 2008, 14,2701–2709.
13 L. J. Brandes, G. M. Queen and F. S. LaBella, Cancer Chemother.Pharmacol., 2000, 45, 298–304.
14 L. L. Von Moltke, D. J. Greenblatt, S. X. Duan, J. S. Harmatz,C. E. Wright and R. I. Shader, J. Clin. Psychopharmacol., 1996, 16,104–112.
15 T. D. Rogerson, C. F. Wilkinson and K. Hetarski, Biochem.Pharmacol., 1977, 26, 1039–1042.
16 K. K. Hajek, N. I. Cook and R. F. Novak, J. Pharmacol. Exp.Ther., 1982, 223, 97–104.
17 M. Murray and L. Zaluzny, Biochem. Pharmacol., 1988, 37,415–420.
18 C. F. Wilkinson, K. Hetnarski, G. P. Cantwell and F. J. Di Carlo,Biochem. Pharmacol., 1974, 23, 2377–2386.
19 A. Verras, I. D. Kuntz and P. R. Ortiz de Montellano, J. Med.Chem., 2004, 47, 3572–3579.
20 M. Delaforge, G. Bouille, M. Jaouen, C. K. Jankowski,C. Lamouroux and C. Bensoussan, Peptides, 2001, 22,557–565.
21 C. Aninat, Y. Hayashi, F. Andre and M. Delaforge, Chem. Res.Toxicol., 2001, 14, 1259–1265.
22 B. M. Schiavi, D. J. Richard and M. M. Joullie, J. Org. Chem.,2002, 67, 620–624.
23 H. M. Marques, M. S. Shongwe, O. Q. Munro and T. J. Egan,S. Afr. J. Chem., 1997, 50, 166–180.
24 D. A. Baldwin, H. M. Marques and J. M. Pratt, J. Inorg. Biochem.,1986, 27, 245–254.
25 Y. Hayashi, S. Orikasa, K. Tanaka, K. Kanoh and Y. Kiso, J. Org.Chem., 2000, 65, 8402–8405.
26 T. Mohammad and H. Morrison, OPPI Briefs, 2000, 32,581–584.
27 P. Kremers, P. Beaune, T. Cresteil, J. de Graeve, S. Columelli,J. P. Leroux and J. E. Gielen, Eur. J. Biochem., 1981, 118, 599–606.
28 M. A. Peyronneau, J. P. Renaud, G. Truan, P. Urban, D. Pomponand D. Mansuy, Eur. J. Biochem., 1992, 207, 109–116.
29 O. H. Lowry, N. J. Rosebrough, A. L. Farr and R. J. Randall,J. Biol. Chem., 1951, 193, 265–275.
30 T. Omura and R. Sato, J. Biol. Chem., 1964, 239, 2370–2378.31 J. B. Schenkman, S. G. Sligar and D. L. Cinti, Pharmacol. Ther.,
1981, 12, 43–71.32 Y. Kawamura-Konishi, A. Asano, M. Yamazaki, H. Tashiro and
H. Suzuki, J. Mol. Catal. B: Enzym., 1998, 14, 181–190.33 A. D. Becke, Phys. Rev. A, 1988, 38, 3098–3100.34 A. D. Becke, J. Chem. Phys., 1993, 98, 5648–5652.35 C. Lee, W. Yang and R. G. Parr, Phys. Rev. B, 1988, 37, 785–789.36 R. Krishnan, J. S. Binkley, R. Seeger and J. A. Pople, J. Chem.
Phys., 1980, 72, 650–654.37 A. Bergner, M. Dolg, W. Kuchle, H. Stoll and H. Preuß, Mol.
Phys., 1993, 80, 1431–1441.38 M. J. Frisch, G. W. Trucks, H. B. Schlegel, G. E. Scuseria,
M. A. Robb, J. R. Cheeseman, V. G. Zakrzewski,J. A. Montgomery Jr, R. E. Stratmann, J. C. Burant,S. Dapprich, J. M. Millam, A. D. Daniels, K. N. Kudin,M. C. Strain, O. Farkas, J. Tomasi, V. Barone, M. Cossi,R. Cammi, B. Mennucci, C. Pomelli, C. Adamo, S. Clifford,J. Ochterski, G. A. Petersson, P. Y. Ayala, Q. Cui,K. Morokuma, P. Salvador, J. J. Dannenberg, D. K. Malick,A. D. Rabuck, K. Raghavachari, J. B. Foresman, J. Cioslowski,J. V. Ortiz, A. G. Baboul, B. B. Stefanov, G. Liu, A. Liashenko,P. Piskorz, I. Komaromi, R. Gomperts, R. L. Martin, D. J. Fox,T. Keith, M. A. Al-Laham, C. Y. Peng, A. Nanayakkara,M. Challacombe, P. M. W. Gill, B. Johnson, W. Chen,M. W. Wong, J. L. Andres, C. Gonzalez, M. Head-Gordon,E. S. Replogle and J. A. Pople, Gaussian 98 (Revision A.11),Gaussian Inc., Pittsburgh PA, 2001.
39 K. Morita, T. Ono and H. Shimakawa, J. Pharmacobiodyn., 1988,11, 106–114.
40 H. Kanzaki, D. Imura, T. Nitoda and K. Kawazu, J. Biosci.Bioeng., 2000, 90, 86–89.
41 R. Cardillo, C. Fuganti, G. Gatti, D. Ghiringhelli and P. Grasseli,Tetrahedron Lett., 1974, 15, 3163–3166.
This journal is �c The Royal Society of Chemistry 2009 Metallomics, 2009, 1, 148–156 | 155
Publ
ishe
d on
27
Nov
embe
r 20
08. D
ownl
oade
d on
11/
02/2
014
10:0
3:51
.
View Article Online

42 C. G. Shin, M. Hayakawa, K. Mikami and J. Yoshimura, Tetra-hedron Lett., 1977, 18, 863–866.
43 B. Kopp-Holtwiesche and H. J. Rehm, J. Environ. Pathol. Toxicol.Oncol., 1990, 10, 41–44.
44 O. P. Larsen, J. C. Frisvad and S. R. Jensen, Phytochemistry, 1992,31, 1613–1615.
45 E. Couadouros, A. Magos and A. Strongilos, Methodsof preparation of dehydro-diketopiperazines in liquid phase or ofsolid supported phase. WO 2005/003102 A1, 2005.
46 N. Shangguan, W. J. Hehre, W. S. Ohlinger, M. P.Beavers and M. M. Joullie, J. Am. Chem. Soc., 2008, 130,6281–6287.
156 | Metallomics, 2009, 1, 148–156 This journal is �c The Royal Society of Chemistry 2009
Publ
ishe
d on
27
Nov
embe
r 20
08. D
ownl
oade
d on
11/
02/2
014
10:0
3:51
.
View Article Online



![Effects of neuronal and inducible NOS inhibitor 1-[2-(trifluoromethyl) phenyl] imidazole (TRIM) in unpredictable chronic mild stress procedure in mice](https://static.fdokumen.com/doc/165x107/631245a93ed465f0570a2648/effects-of-neuronal-and-inducible-nos-inhibitor-1-2-trifluoromethyl-phenyl-imidazole.jpg)
















