
Centromeric Protein B Null Mice Are Viable with No Apparent Abnormalities
Upload
independentCategory
view
1download
0
REGULAR ARTICLE
Intestinal stem cells remain viable after prolonged tissuestorage
Megan K. Fuller & Denver M. Faulk & Nambirajan Sundaram & Maxime M. Mahe &
Kara M. Stout & Richard J. von Furstenberg & Brian J. Smith & Kirk K. McNaughton &
Noah F. Shroyer & Michael A. Helmrath & Susan J. Henning
Received: 22 January 2013 /Accepted: 27 May 2013 /Published online: 3 July 2013# Springer-Verlag Berlin Heidelberg 2013
Abstract Intestinal stem cells (ISCs) are responsible forrenewal of the epithelium both during normal homeostasisand following injury. As such, they have significant thera-peutic potential. However, whether ISCs can survive tissuestorage is unknown. We hypothesize that, although the ma-jority of epithelial cells might die, ISCs would remain viablefor at least 24 h at 4 °C. To explore this hypothesis, jejuna of
C57Bl6/J or Lgr5-LacZ mice were removed and eitherprocessed immediately or placed in phosphate-buffered sa-line at 4 °C. Delayed isolation of epithelium was performedafter 24, 30, or 48 h storage. At the light microscope level,despite extensive apoptosis of villus epithelial cells, smallintestinal crypts remained morphologically intact for 30 hand ISCs were identifiable via Lgr5-LacZ positivity. Elec-tron microscopy showed that ISCs retained high integrity for24 h. When assessed by flow cytometry, ISCs were moreresistant to degeneration than the rest of the epithelium,including neighboring Paneth cells, with higher viabilityacross all time points. Cultured isolated crypts showed noloss of capacity to form complex enteroids after 24 h tissuestorage, with efficiencies after 7 days of culture remainingabove 80 %. By 30 h storage, efficiencies declined butbudding capability was retained. We conclude that, withdelay in isolation, ISCs remain viable and retain their prolif-erative capacity. In contrast, the remainder of the epithelium,including the Paneth cells, exhibits degeneration andprogrammed cell death. If these findings are recapitulatedin human tissue, storage at 4 °C might offer a valuabletemporal window for the harvesting of crypts or ISCs fortherapeutic application.
Keywords Intestinal stem cells . Light and electronmicroscopy . Viability . Resistance to storage . Enteroidculture . Mouse
Introduction
The intestinal epithelium is the most rapidly proliferatingtissue in the mammalian body, with complete turnover ofthe majority of cells occurring every 3–4 days. Pioneeringwork of Cheng and Leblond in the early 1970s suggested that
This work was supported an AGA Student Research Fellowship Award(K.M.S.) and by the National Institutes of Health Grants R01-DK083325 (M.A.H.), P30 DK034987 (S.J.H., M.A.H.) and U01-DK085547 (S.J.H., M.A.H.). The last-mentioned grant is part of theIntestinal Stem Cell Consortium, a collaborative research projectfunded by the National Institute of Diabetes and Digestive and KidneyDiseases and the National Institute of Allergy and Infectious Diseases.Additionally, this work received support from the UNC Department ofSurgery (M.K.F.). The UNC Flow Cytometry Core Facility is supportedin part by an NCI Center Core Support Grant (P30CA06086) to theUNC Lineberger Comprehensive Cancer Center.
M. K. FullerDepartment of Surgery, University of North Carolina at ChapelHill, Chapel Hill, NC, USA
K. M. Stout :R. J. von Furstenberg : B. J. Smith :K. K. McNaughtonDepartment of Medicine and Cell Biology & Physiology,University of North Carolina at Chapel Hill, Chapel Hill, NC, USA
D. M. Faulk :N. Sundaram :M. M. Mahe :M. A. HelmrathDepartment of Surgery, Cincinnati Children’s Hospital MedicalCenter, Cincinnati, OH, USA
N. F. ShroyerDepartment of Pediatrics: Division of Gastroenterology,Hepatology & Nutrition, Cincinnati Children’s Hospital MedicalCenter, Cincinnati, OH, USA
S. J. Henning (*)University of North Carolina at Chapel Hill, 4341 MedicalBiomolecular Research Building (MBRB), CB# 7555, 111 MasonFarm Road, Chapel Hill, NC 27599-7555, USAe-mail: [email protected]
Cell Tissue Res (2013) 354:441–450DOI 10.1007/s00441-013-1674-y

this turnover arises from stem cells located deep in theintestinal crypts (Cheng and Leblond 1974). The generalconcept that these stem cells give rise to all differentiatedlineages within the epithelium has stood the test of time andhas now been confirmed by numerous groups (for reviews,see Garrison et al. 2009; King andDekaney 2013;Montgomeryand Breault 2008; Shaker and Rubin 2010; Wong 2004). Ad-ditionally, as more sophisticated tools have become available,accumulated evidence indicates that at least two relativelydistinct populations of intestinal stem cells (ISCs) exist. Themost extensively studied population is the one that is markedby the G-protein-coupled receptor Lgr5. This population isactively cycling and is normally responsible for the day-to-day homeostatic turnover of the epithelium (Barker et al.2012). In contrast, the other population, marked by Bmi1, m-Tert, Hopx, Lrig1 and Dclk1, is relatively quiescent and ap-pears to be responsible for epithelial regeneration after injury(Carlone and Breault 2012; Lund 2012; Potten et al. 2009;Scoville et al. 2008).
The use of preparations that maintain ISCs in their niche,such as isolated crypts or organoids (i.e., crypts with adher-ent myofibroblasts), has long been recognized as havingvaluable therapeutic applications for situations in which acritical mass of the intestine is damaged. Several groups havereported the functional engraftment of tissue-engineeredsmall intestine grown from organoid units (Avansino et al.2005; Bitar and Raghavan 2012; Levin and Grikscheit2012). Initially, a critical limiting factor in such applica-tions was the inability to expand intestinal organoidsin vitro (Gupta et al. 2006). Recently, however, cultureconditions have been reported that allow the net expan-sion of murine crypts (Fuller et al. 2012; Sato et al.2009), thus bringing therapeutic transplantation signifi-cantly closer to reality. Nevertheless, to date, all studiesof ISC/crypt expansion and transplantation have utilizedfreshly isolated, healthy tissue (Lahar et al. 2011; Satoet al. 2011; Yui et al. 2012). Whereas such an approach iseasy in the laboratory, this might rarely be the case in theclinic. Therapeutic sources would typically include tissuecollected either postmortem or at surgery. In either case, theability to store tissue prior to crypt or ISC isolation wouldconstitute a significant advantage.
Data from other organ systems suggest that therapeutical-ly useful cells are present postmortem. Hematopoeitic stemcells have long been known to survive with delayed bonemarrow harvest (Liu et al. 1978; Mugishima et al. 1985;Yesus et al. 1981). More recently, hepatocytes have beenfound to be viable and capable of engraftment after harvestup to 27 h postmortem (Erker et al. 2010). On the basis ofthese findings, we hypothesize that, following the storageof intestinal tissue at 4 °C, ISCs would remain viable, even ifother components of the epithelium degenerate. To investi-gate this hypothesis, viability has been assessed by light and
electron microscopy, flow cytometry and crypt culture. Theresults indicate that, just as in other tissues, stem cells of thesmall intestine are remarkably resistant to apoptosis andnecrosis during storage up to 30 h postmortem.
Materials and methods
Animals and reagents
All animal procedures were approved by the InstitutionalAnimal Care and Use Committee at the two respective insti-tutions. Male C57BL/6 J mice were obtained from JacksonLaboratories (Bar Harbor, Me., USA). Heterozygote breederpairs of Lgr5-LacZ mice (Barker et al. 2007) were generous-ly provided by Lexicon Pharmaceuticals (The Woodlands,Tex., USA). All mice were housed at 4–5 per cage on a 12 hlight/dark cycle in facilities approved by the American Asso-ciation for Accreditation of Laboratory Animal Care and were6–8 weeks of age at the time of usage. Mice were killed by anoverdose of isoflurane (until breathing stopped) followed bycervical dislocation. The jejunum was then removed and usedas described in Experimental design below.
Reagents used for histology were: anti-cleaved caspase-3(Cell signaling, no. 9661, Danvers, Mass., USA) and 5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside (X-gal; Sig-ma, no. 7240-90-6, St. Louis, Mo., USA). Antibodies used forflow cytometric analysis included: Alexa Fluor 647 (Alexa-647)-conjugated anti-CD31 (Biolegend, no. 102415, SanDiego, Calif., USA), Alexa-647-conjugated anti-CD45(Biolegend no. 103124), Pacific-Blue-conjugated anti-CD24(Biolegend, no. 101820), fluorescein isothiocyanate (FITC)-conjugated bovine anti-rabbit IgG (Santa Cruz Biotechnology,no. SC-2365, Santa Cruz, Calif., USA) and FITC-conjugatedlectin from Ulex europaeus (UEA; Sigma, no. L9006).
Experimental design
In the first experiment, viability of the entire jejunal epithe-lium was assessed following immediate dissociation, ascompared with 24 h storage under various conditions: (1)storage in phosphate-buffered saline (PBS) at 4 °C afterinitial flushing of the intestine; (2) storage in PBS at 4 °Cwithout flushing of the intestinal contents; (3) storage of theentire mouse at 4 °C. Jejunal epithelial cells were isolated viathe EDTA/dispase method as previously described, exceptthat dithiothreitol was omitted (Formeister et al. 2009). Eachepithelial preparation was stained with trypan blue and thentrypan-positive (dead) and trypan-negative (alive) cells werecounted in a hemocytometer. Viability was expressed as thenumber of alive cells as a percentage of total (alive plusdead) cells. Whereas this approach gives useful informationregarding the overall pattern of epithelial viability, absolute
442 Cell Tissue Res (2013) 354:441–450

values for percent viable cells might nevertheless be an over-estimation if some proportion of the dead cells (i.e., part ofthe denominator) becomes degraded to the point of beingunrecognizable as cells assessed for overall viability bytrypan blue.
For all subsequent experiments, the jejunum of C57BL6/Jor Lgr5-LacZmice was dissected, flushed with ice-cold PBSand split into four aliquots for immediate processing (0 h)and delayed processing (24, 30 and 48 h). For delayedprocessing, flushed jejunum was kept in a 15 ml flaskscontaining PBS at 4 °C for 24, 30, or 48 h. When theintestine of each mouse jejunum was split into groups,sections were taken diagonally or small transverse sec-tions were cut, all of which were systematically splitbetween time points in order to prevent regional differ-ences between the time points.
Histology/microscopy
Tissue was pinned flat and fixed with 4 % paraformaldehyde(PFA) for 3 h at room temperature (RT) or processed forwhole-mount β-galactosidase (X-gal) staining. After whole-mount staining and/or fixation, tissue was immersed in 70 %ethanol for at least 24 h and then paraffin-embedded into asingle block for each time point. Longitudinal sections of6 μm in thickness were utilized for morphology andimmunohistochemistry.
Slides were evaluated by using an Axio Imager A1 mi-croscope and micrographs were taken with an AxioCamMRC 5 high-resolution camera (Carl Zeiss, Microimaging,Thornwood, N.Y., USA). Overall morphology was examinedwith hematoxylin and eosin staining (H&E). Apoptosis wasassessed with cleaved caspase-3. For caspase-3, heat-inducedepitope retrieval was performed in a decloaking chamber at120 °C for 30 s followed by 90 °C for 10 s (DAKO, no. S1699,Glostrup, Denmark). Sections were treated with 3 % H2O2 for10 min at RT and then incubated for 1 h in blocking buffer(DAKO, no. X0909, Glostrup, Denmark) or 10 % normal goatserum at RT. Subsequently, sections were incubated with pri-mary antibody at 1:800 (cleaved caspase-3) in antibody diluent(Thermo, no. TA125-ADQ, Waltham, Mass., USA) at 4 °Covernight. Sections were rinsed in TRIS buffer between allapplications of antibodies, reagents and endogenous peroxidaseblocking step. Secondary antibody, namely biotinylated goatanti-rabbit IgG (Jackson Immunoresearch, no. 111-065-144,West Grove, Pa., USA), was applied at 1:500 in antibodydiluent (Thermo, no. TA125-ADQ) for 1 h at RT. VectastainElite avidin-biotin horseradish peroxidase complex in TRISwas applied at 1:50 (Vector Labs, no. PK6100, Burlingame,Calif., USA) for 30 min at RT. Quatro chromogen, namely3,3′ diaminobenzidine (Thermo, no. TA125-QHDX) was thenapplied for 1 min. Finally, sections were counterstained with
either nuclear fast red or hematoxylin, dehydrated, cleared inxylene and DPX-coverslipped.
β-Galactosidase staining
To determine whether the Lgr5-positive crypt base stem cellsremained over time, whole-mount staining was performedwith intestinal tissue from male heterozygote Lgr5-LacZmice as described in the published protocol, with slightmodifications (Takeda et al. 2011). All segments werepinned flat and flushed with 3 mM dithiothreitol to removemucus. Briefly, the jejunum was fixed with glutaraldehydeand PFA, incubated with X-gal solution for 16 h overnight atRT and then further fixed with 4 % PFA. Tissue was thentransferred to cassettes in 70 % ethanol and embedded inparaffin.
Electron microscopy
For assessment of the structural integrity of the intestine atthe 0 and 24 h time points, 5-mm pieces of the jejunum wereimmersion-fixed in 2 % PFA/2.5 % glutaraldehyde in 0.15Msodium phosphate buffer, pH 7.4 and stored at 4 °C forseveral days before processing. Following several washesin 0.15 M sodium phosphate buffer, the samples were post-fixed in 1 % osmium tetroxide/1.25 % potassium ferrocya-nide in 0.15 M sodium phosphate buffer, pH 7.4, for 1 h(Russel and Burguet 1977). Samples were dehydrated in agraded series of ethanols followed by propylene oxide andthen infiltrated and embedded in Polybed 812 resin(Polysciences, Warrington, Pa., USA). Semithin transversesections (1 μm thick) were cut with a glass knife, mounted onslides, stained with 1 % toluidine blue and viewed by using alight microscope to select the region of interest. Ultrathinsections (70 nm) were cut by using a diamond knife,mounted on 200-mesh copper grids and stained with 4 %aqueous uranyl acetate and Reynold’s lead citrate (Reynolds1963). Samples were observed by using a LEO EM910transmission electron microscope operating at 80 kV (CarlZeiss Microscopy, Peabody, Mass., USA) and digital imageswere acquired by means of a Gatan Orius SC1000 charge-coupled device digital camera with Digital Micrograph3.11.0 (Gatan, Pleasanton, Calif., USA). The integrity ofthe cells at the crypt base was assessed by a pathologistwho was blinded to the experimental conditions.
Flow cytometry
Jejunal epithelial cells were isolated as described underExperimental design and 1 million cells were then incubatedin 100 μl PBS for 45 min on ice with no antibody, isotype-FITC Bovine IgG (0.5 μg), with CD31-Alexa674 (0.5 μg)and CD24-PB (0.25 μg), or with CD45-Alexa647 (0.5 μg),
Cell Tissue Res (2013) 354:441–450 443
CD31-Alexa647, CD24-PB and UEA-FITC (1:250). Sam-ples were then rinsed and resuspended in 500 μl PBS withthe addition of 1 μg propidium iodide (PI). Cells wereanalyzed by using a Beckman-Coulter (Dako) CyAn ADP.Debris was first excluded based on size via bivariate plotof forward scatter (FSC) versus side scatter. Doubletswere excluded by using successive gating on both abivariate plot of pulse-width versus FSC and a bivariateplot of FSC area versus FSC linear. CD45+ and CD31+
events, representing leukocytes and endothelial cells, respec-tively (von Furstenberg et al. 2011), were then excluded basedon positivity for Alexa-647. To determine which crypt basecell populations survived over the time course, the remainingcells were analyzed for PI, CD24 and UEA in order toassess viability of ISC and Paneth cell fractions as previ-ously described (von Furstenberg et al. 2011; Wong et al.2012). A separate aliquot was also analyzed for overallviability by trypan blue.
Tissue harvest for intestinal crypt culture
Crypts were isolated, cultured and quantified as previouslypublished (Fuller et al. 2012). The proximal jejunum ofmale C57Bl6/J mice was removed and processed (0 h) orplaced in PBS at 4 °C for 24, 30, or 48 h (n=6, 8, 8, 4,respectively). Tissue was manually shaken for crypt removalafter incubation with 3 mM EDTA. Pelleted crypts wereresuspended in growth-factor-reduced Matrigel (BD Biosci-ence) supplemented with epithelial growth factor (50 ng/ml),Noggin (100 ng/ml), R-spondin (500 ng/ml) and wnt3a(5 ng/ml). All concentrations are for total well volume. Withthe exception of wnt3a, these conditions are as described bySato et al. (2009). Crypts in 10 μl Matrigel were then platedin six experimental replicates per time point in 48-wellplates. Each was then overlaid with Advanced DMEM/F12(GIBCO, Carlsbad, Calif., USA) supplemented with HEPES(10 mM), N2 (1:100), B27 (1:50), L-glutamine (1:100) andpenicillin/streptomycin (1:100; Invitrogen). The final num-ber of crypts plated was counted per well and followed overthe course of 7 days for enterosphere and enteroid forma-tion. The definition of these structures followed recent no-menclature recommendations (Stelzner et al. 2012).Enteroid efficiencies were defined as the percentage ofenteroids formed divided by the number of crypts initiallyplated. Among formed enteroids, lateral crypt buds werecounted across a minimum of two wells and ten enteroidsper well per mouse.
Statistics and quantification
To assess the effect of storage conditions on epithelial via-bility, a one-way analysis of variance with Bonferroni post-test corrections for multiple comparisons was performed in
GraphPad Prism 5 (LaJolla, Calif., USA). For analysis ofviability over time by cell type (ISC v Paneth), a two-wayanalysis of variance was performed in GraphPad Prism 5.To assess the maintenance of culture efficiency and cryptbudding over time of tissue storage at 4 °C, a one-wayanalysis of variance with Bonferroni post-test correctionsfor multiple comparisons was performed in GraphPadPrism 5. P-values less than 0.05 were considered statisticallysignificant.
Results
Viable intestinal cells are present at least up to 24 h postmortem
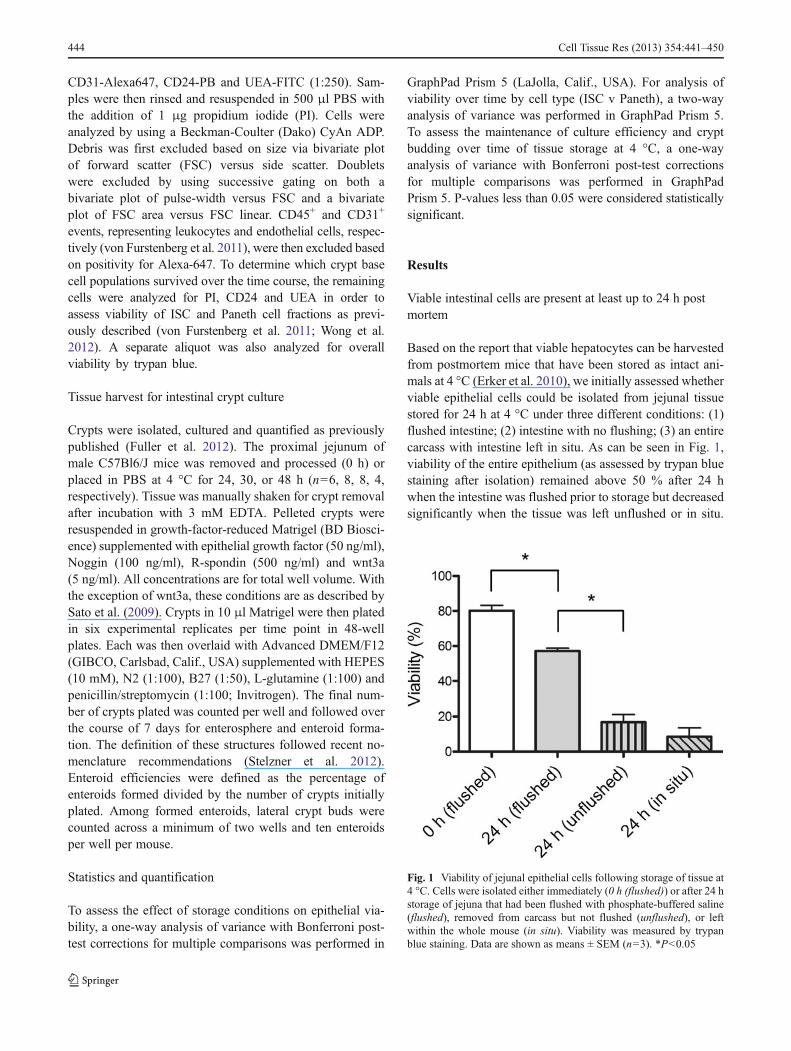
Based on the report that viable hepatocytes can be harvestedfrom postmortem mice that have been stored as intact ani-mals at 4 °C (Erker et al. 2010), we initially assessed whetherviable epithelial cells could be isolated from jejunal tissuestored for 24 h at 4 °C under three different conditions: (1)flushed intestine; (2) intestine with no flushing; (3) an entirecarcass with intestine left in situ. As can be seen in Fig. 1,viability of the entire epithelium (as assessed by trypan bluestaining after isolation) remained above 50 % after 24 hwhen the intestine was flushed prior to storage but decreasedsignificantly when the tissue was left unflushed or in situ.
Fig. 1 Viability of jejunal epithelial cells following storage of tissue at4 °C. Cells were isolated either immediately (0 h (flushed)) or after 24 hstorage of jejuna that had been flushed with phosphate-buffered saline(flushed), removed from carcass but not flushed (unflushed), or leftwithin the whole mouse (in situ). Viability was measured by trypanblue staining. Data are shown as means ± SEM (n=3). *P<0.05
444 Cell Tissue Res (2013) 354:441–450

Therefore, in all further experiments, the jejunum wasflushed prior to storage in PBS at 4 °C.
Morphology of small intestinal crypts remains intactthrough 30 h at 4 °C
Tissue architecture was initially assessed by H&E stainingperformed after 0, 24, 30 and 48 h after storage. The results(Fig. 2a–e’) demonstrated the overall maintenance of cryptarchitecture for 30 h postmortem. Meanwhile, by 30 h, thevilli began to slough off from their laminal cores and werenearly completely absent by 48 h. At 48 h, some tissuesections exhibited the conservation of crypts (Fig. 2d). How-ever, in other areas, crypts detached from the submucosawith demonstrable cellular shrinkage (Fig. 2e).
To assess the viability of the crypt compartments further,we used tissue from Lgr5-LacZ mice to identify apoptoticcells simultaneously by staining with antibodies to cleavedcaspase-3 and Lgr5-positive ISCs by LacZ staining (Fig. 2f–j’). Consistent with the H&E staining of wild-type mice, thecrypts of Lgr5-LacZmice retained good architecture for 30 hbut showed signs of degeneration by 48 h. Moreover, for24 h, crypts exhibited no signs of apoptosis, despite exten-sive activated caspase-3 staining on the villi. By 48 h post-mortem, near complete loss of the apoptotic villi could beseen and at this time, the crypts began to display caspasepositivity (Fig. 2i, j). As the ISCs reside within the crypt baseand are responsible for the maintenance of the small intesti-nal epithelium, we further characterized our tissue sectionsfor the presence of crypt base columnar ISCs marked byLgr5. Lgr5-LacZ-expressing stem cells (Barker et al. 2007)were clearly evident for 30 h but could not be identified by48 h delayed isolation (Fig. 2f–j).
Small intestinal stem cells are resistant to degeneration
The electron microscopic images in Fig. 3 show that, at theultrastructural level, the ISCs of the crypt base retained highintegrity after 24 h storage of tissue. In contrast, some of theadjacent Paneth cells became necrotic at this time, althoughothers still appeared to be relatively healthy with intactelectron-dense secretory vesicles.
Based on the report that the CD24loUEA− populationrepresented a relatively pure fraction of ISC by flow cytom-etry (Wong et al. 2012), we examined the viability of thatfraction by using PI positivity to identify dead cells. In thesame epithelial preparations, the viability of Paneth cells wasassessed by utilizing the CD24loUEA+ fraction. As can beseen in Fig. 4a, a decline occurred in the percent of viablestem cells and Paneth cells over time postmortem (P<0.001).However, ISCs were significantly more viable (Fig. 4c) thantheir neighboring Paneth cells (P<0.001; Fig. 4d). Thisdifference is most pronounced by 30 h (Fig. 4a).
Small intestinal stem cells retain proliferative capacityafter storage at 4 °C
As ISCs appeared viable postmortem, we further assessedtheir functionality via in vitro crypt culture. Freshly isolatedcrypts showed culture efficiencies of over 80 % and remark-ably, no decline was seen in culture efficiency after a 24-hdelay in isolation (Fig. 5). Efficiency dropped to 36 % whencrypts were isolated from tissue that had been stored for 30 hand no crypts formed enteroids when storage was extendedto 48 h. However, if enteroids form, they retain their prolif-erative capacity as evidenced by no statistically significantdifferences in their ability to form buds (Fig. 6a, P>0.3).
Fig. 2 Architecture of flushed jejunal tissue stored for 4 °C for 0, 24,30, or 48 h. a–e’ Hematoxylin and eosin for demonstration of morphol-ogy. f–j’ Cleaved caspase-3 staining (brown) for apoptosis and co-staining for Lgr5-LacZ (blue). Each time point is shown at low power(a–j) and high power (a’–j’). For the 48-h time point, two differentregions are shown to illustrate the variability observed. Because the
tissue is fragile at this stage, good sections were difficult to obtain.Nevertheless, d, i show a region in which the mucosa is nearly denudedbut the muscle layers have remained intact and negative for cleavedcaspase-3, whereas e, j show a region in which the muscle layers arealso destroyed. Bar 100 μm
Cell Tissue Res (2013) 354:441–450 445

Multilobulated structures that equally progressed fromenterospheres to enteroids were observed in tissue that had
been stored for 0, 24, or 30 h. Moreover, the observation ofapproximately six buds per enteroid from tissue that had
Fig. 3 Electron microscopicimages of jejunal tissue storedfor 0 or 24 h at 4 °C. Top tobottom Crypts at increasingmagnification (left 0 h, right24 h, open arrows crypt basecolumnar ISCs, closed arrowsPaneth cells with electron densegranules)
Fig. 4 Viability of ISCs and Paneth cells isolated from jejunal tissuestored at 4 °C for 0, 24, or 30 h. a ISC viability was computed as live(propidium-iodide-negative, PI−) CD24loUEA− cells as a percentage oftotal CD24loUEA− cells and Paneth cell viability as live (PI−)CD24loUEA+ cells as a percentage of total CD24loUEA+ cells. Datadisplayed as means ± SEM (n=4). b Example of gating strategy as
illustrated by a 30-h sample, with gates termed as SC for stem cell andPC for Paneth cell (UEA lectin from Ulex europaeus, FITC fluoresceinisothiocyanate). The CD24loUEA− (SC) and CD24loUEA+ (PC) sub-populations are gated forward onto single parameter histograms (c, d,respectively) to determine the pecentage of cells negative for PI (live)
446 Cell Tissue Res (2013) 354:441–450

been stored for 30 h (Fig. 6b) not only is within the range ofthat observed for fresh tissue in the current study but is alsoconsistent with the extent of budding reported for cryptsisolated from fresh tissue in a previous study (Fuller et al.2012).
Discussion
Current advances in tissue engineering and cellular transplan-tation models require analysis of potential tissue sources, bothautologous and heterologous. Some tissue types have provenviable after delayed isolation and useful for therapy (Erkeret al. 2010; Mugishima et al. 1985). For the intestine, numer-ous studies have shown that this tissue is remarkablyrejuvenative after various types of in vivo damage, the ISCsubpopulations being relatively resistant to cell death (Barkeret al. 2012; Potten and Hendry 1975; Wright 2000). However,the survival of intestinal tissue postmortem has not been
systematically investigated. In this study, we have sought todetermine whether viable and therapeutically useful ISCs arepresent and functional after delayed isolation.
Our initial experiment explored various storage condi-tions with the rationale that, depending on the nature of theharvest (e.g., heterologous tissue postmortem versus autolo-gous tissue via surgery), opportunity might be limited toprepare the tissue for storage. In this experiment, our assess-ment has been based on the viability of the entire epithelium.The data (Fig. 1) make a convincing case that, in order toensure reasonable viability after 24 h storage at 4 °C, theintestinal tissue needs not only to be removed from the bodybut also to be flushed prior to storage. Whether the loss ofviability in the tissue stored in situ or without flushing couldhave reflected slower cooling or adverse effects of intestinalcontents remains to be investigated. Likewise, the possibilitythat the ISC fraction is resistant, even under adverse storageconditions, remains open to question. Nevertheless, on thebasis on these initial observations, we chose to use flushedtissue for all subsequent experiments.
Our images of tissue architecture after storage for varioustimes revealed dramatic differences between villi and crypts.Whereas the villi showed extensive apoptosis at 24 h andalmost complete disintegration by 48 h, the crypts remainedessentially intact. Moreover, healthy ISCs at the crypt baseswere observed, both at the light microscopic (by Lgr5-LacZ)and electron microscopic levels. Similar morphology of ap-optotic villus loss with crypt preservation has also beennoted in models of intestinal ischemia and reperfusion (Itohet al. 2002; Park and Haglund 1992; Udassin et al. 1994).Although no studies have looked specifically at the ISCsfollowing ischemia and reperfusion, thymidine incorpora-tion has been noted to occur at the base of the crypt in onestudy at the time of regeneration (Park and Haglund 1992).Given the common findings of villus degeneration and cryptpreservation, the same protective mechanisms might be inplace both ex vivo following storage at 4 °C and in vivofollowing ischemia at body temperature. As many of the
Fig. 5 Survival and growth of murine crypts isolated from tissue storedat 4 °C for 0, 24, 30, or 48 h. Data are shown as enteroid-formingefficiency after 7 days (7 d) of crypt culture, with means ± SEM (n=7,7, 7, 4). *P<0.05
Fig. 6 Proliferative capacity ofmurine ISC after storage at 4 °Cas measured by enteroidbudding after 7 days of cryptculture. a Composite data for 0,24 and 30 h displayed asmeans ± SEM (n=6, 6, 5)P>0.3. b Representativeenteroid from 30-h condition(black arrows crypt buds).Bar 200 μm
Cell Tissue Res (2013) 354:441–450 447

pathologic diseases resulting in intestinal failure are second-ary to ischemia, our data suggest that ISCs remain viable inthese disorders.
In addition to the morphologic evidence of ISC viability,we have found that they are capable of supporting cryptgrowth in vitro after delayed isolation up to 30 h. Further-more, once growth is established in vitro, cells maintain theirproliferative capacity, as evidenced by the lack of significantdifference in the number of buds observed in enteroidsderived from 30 h tissue as compared with those of freshtissue. The successful expansion of crypts in vitro is consis-tent with our finding that the ISCs are more resistant todegeneration than their neighboring Paneth cells over time,as demonstrated via flow cytometry and electron microsco-py. However, we cannot say whether these dying Panethcells still play a role in protecting their neighboring stemcells, as has been shown in vivo for live Paneth cells (Parryet al. 2012). Protection by compromised Paneth cells isconsistent with a study that explored bone marrow transplantof both apoptotic support cells and live hematopoietic stemcells and revealed improved engraftment and declined ratesand severity of graft-versus-host disease (Pessach et al.2012). Interestingly, despite a lack of nutrients, the majorityof cell death appears to occur via apoptosis in a controlledfashion.
As viable crypts are present and expandable, they canprovide a tissue source for cellular transplantation or tissueengineering of the small intestine. Furthermore, as culturetechniques improve, single ISCs might provide therapeuticbenefits. Proof of principle for cellular transplantation wasrecently reported by Yui et al. (2012) who successfullytransplanted colonoids into a mouse model of colitis. Theaim of their study was to replace a damaged mucosal surfacearea by colonoid delivery via the anus; engraftment ofcolonoids and restoration of colonic architecture wereachieved (Yui et al. 2012). Although, as noted in a recentcommentary (Shaker and Rubin 2012), much work stillneeds to be done to establish functionality, this cellularapproach might be beneficial for some patients. However,for other patients, a tissue engineering approach to providean additional absorptive surface area might be necessary.Tissue-engineered small intestines are now well character-ized and have shown abrogation of nutritional deficits in asmall animal model and feasibility in a large animal model(Avansino et al. 2005; Grikscheit et al. 2004; Sala et al.2009). Whereas transplantation might provide therapy forshort bowel syndrome and intestinal failure, the finding ofviable ISCs after prolonged storage might also help us tounderstand the in vivo regeneration of the bowel followingdamage.
An exciting area for future investigation will be to deter-mine the cell that is responsible for our observed renewalin vitro. Current evidence supports the existence of actively
cycling “workhorse” ISCs and slowly cycling “quiescent”ISCs (Carlone and Breault 2012; Li and Clevers 2010; Pottenet al. 2009; Scoville et al. 2008). Additionally, understress, these cells might interconvert to repopulate the crypt(Montgomery et al. 2011; Takeda et al. 2011; Tian et al. 2011;Yan et al. 2012). In other states of damage and adaptation,such as resection, doxorubicin, radiation, or development, amarked expansion is noted within the stem cell compartment(Dehmer et al. 2011; Dekaney et al. 2007, 2009; VanLandeghem et al. 2012). We hypothesize that the same mech-anisms might be in play as cells survive delayed isolation andthen are able to proliferate in vitro. From the irradiationmodel,an expansion is noted in both the active and quiescent stempools but which cell population initiates the proliferativeresponse to damage is unclear (Van Landeghem et al. 2012).Likewise, although we detect ISCs marked by Lgr5 after 30-hdelayed isolation, we cannot say definitively that these cellsare responsible for initiating growth in vitro. More research isneeded to define the inter-relationships among crypt cellsduring damage and repair. Nevertheless, despite not knowingwhich stem cell compartment is responsible, we can say thatisolation in a delayed fashion provides viable ISCs that arecapable of growth and proliferation in vitro and might betherapeutically beneficial.
Acknowledgments The authors wish to thank Dr. Joe Galanko fromthe UNC Center for Gastrointestinal Biology and Disease for assistancewith the statistical analysis, Ashley Ezzel from the Cell and MolecularPhysiology Core at UNC for assistance with the histology, VictoriaMadden, Steven Ray and Dr. Robert Bagnell of the UNC School ofMedicine Microscopy Services Lab for sample preparation and usefulinput concerning the electron microscopy and Drs. ChristopherDekaney, Scott Magness and P. Kay Lund for useful discussions.
References
Avansino JR, Chen DC, Hoagland VD, Woolman JD, Haigh WG,Stelzner M (2005) Treatment of bile acid malabsorption using ilealstem cell transplantation. J Am Coll Surg 201:710–720
Barker N, van Es JH, Kuipers J, Kujala P, van den Born M, CozijnsenM, Haegebarth A, Korving J, Begthel H, Peters PJ, Clevers H(2007) Identification of stem cells in small intestine and colon bymarker gene Lgr5. Nature 449:1003–1007
Barker N, van Oudenaarden A, Clevers H (2012) Identifying the stemcell of the intestinal crypt: strategies and pitfalls. Cell Stem Cell11:452–460
Bitar KN, Raghavan S (2012) Intestinal tissue engineering: currentconcepts and future vision of regenerative medicine in the gut.Neurogastroenterol Motil 24:7–19
Carlone DL, Breault DT (2012) Tales from the crypt: the expanding roleof slow cycling intestinal stem cells. Cell Stem Cell 10:2–4
Cheng H, Leblond CP (1974) Origin, differentiation and renewal of thefour main epithelial cell types in the mouse small intestine. V.Unitarian theory of the origin of the four epithelial cell types. Am JAnat 141:537–562
448 Cell Tissue Res (2013) 354:441–450

Dehmer JJ, Garrison AP, Speck KE, Dekaney CM, Van Landeghem L,Sun X, Henning SJ, Helmrath MA (2011) Expansion of intestinalepithelial stem cells during murine development. PLoS One6:e27070
Dekaney CM, Fong JJ, Rigby RJ, Lund PK, Henning SJ, Helmrath MA(2007) Expansion of intestinal stem cells associated with long-term adaptation following ileocecal resection in mice. Am JPhysiol Gastrointest Liver Physiol 293:G1013–G1022
Dekaney CM, Gulati AS, Garrison AP, Helmrath MA, Henning SJ(2009) Regeneration of intestinal stem/progenitor cells followingdoxorubicin treatment of mice. Am J Physiol Gastrointest LiverPhysiol 297:G461–G470
Erker L, Azuma H, Lee AY, Guo C, Orloff S, Eaton L, Benedetti E,Jensen B, Finegold M, Willenbring H, Grompe M (2010)Therapeutic liver reconstitution with murine cells isolated longafter death. Gastroenterology 139:1019–1029
Formeister EJ, Sionas AL, Lorance DK, Barkley CL, Lee GH, MagnessST (2009) Distinct sox9 levels differentially mark stem/progenitorpopulations and enteroendocrine cells of the small intestine epi-thelium. Am J Physiol Gastrointest Liver Physiol 296:G1108–G1118
Fuller MK, Faulk DM, Sundaram N, Shroyer NF, Henning SJ,Helmrath MA (2012) Intestinal crypts reproducibly expand inculture. J Surg Res 178:48–54
Garrison AP, Helmrath MA, Dekaney CM (2009) Intestinal stem cells. JPediatr Gastroenterol Nutr 49:2–7
Grikscheit TC, Siddique A, Ochoa ER, Srinivasan A, Alsberg E, HodinRA, Vacanti JP (2004) Tissue-engineered small intestine improvesrecovery after massive small bowel resection. Ann Surg 240:748–754
Gupta A, Dixit A, Sales KM, Winslet MC, Seifalian AM (2006) Tissueengineering of small intestine—current status. Biomacromolecules7:2701–2709
Itoh H, Yagi M, Hasebe K, Fushida S, Tani T, Hashimoto T, Shimizu K,Miwa K (2002) Regeneration of small intestinal mucosa afteracute ischemia-reperfusion injury. Dig Dis Sci 47:2704–2710
King SL, Dekaney CM (2013) Small intestinal stem cells. Curr OpinGastroenterol 29:140–145
Lahar N, Lei NY,Wang J, Jabaji Z, Tung SC, Joshi V, Lewis M, StelznerM, Martin MG, Dunn JC (2011) Intestinal subepithelialmyofibroblasts support in vitro and in vivo growth of human smallintestinal epithelium. PLoS One 6:e26898
Levin DE, Grikscheit TC (2012) Tissue-engineering of the gastrointes-tinal tract. Curr Opin Pediatr 24:365–370
Li L, Clevers H (2010) Coexistence of quiescent and active adult stemcells in mammals. Science 327:542–545
Liu PI, Ogawa M, Crook L, Ochia R, Upshur JK (1978) Proliferativefunction of cadaveric bone marrow cells. Am J Hematol 5:145–150
Lund PK (2012) Fixing the breaks in intestinal stem cells after radia-tion: a matter of DNA damage and death or DNA repair andregeneration. Gastroenterology 143:1144–1147
Montgomery RK, Breault DT (2008) Small intestinal stem cell markers.J Anat 213:52–58
Montgomery RK, Carlone DL, Richmond CA, Farilla L, KranendonkMEG, Henderson DE, Baffour-Awuah NY, Ambruzs DM, FogliLK, Algra S, Breault DT (2011) Mouse telomerase reverse tran-scriptase (mtert) expression marks slowly cycling intestinal stemcells. Proc Natl Acad Sci USA 108:179–184
Mugishima H, Terasaki P, Sueyoshi A (1985) Bone marrow fromcadaver donors for transplantation. Blood 65:392–396
Park PO, Haglund U (1992) Regeneration of small bowel mucosa afterintestinal ischemia. Crit Care Med 20:135–139
Parry L, Young M, El Marjou F, Clarke AR (2012) Evidence for acrucial role of Paneth cells in mediating the intestinal response toinjury. Stem Cells 31:776–785
Pessach I, Shimoni A, Nagler A (2012) Apoptotic cells in allogeneichematopoietic stem cell transplantations: “Turning trash intogold”. Leuk Lymphoma 53:2130–2135
Potten CS, Hendry JH (1975) Differential regeneration of intestinalproliferative cells and cryptogenic cells after irradiation. Int JRadiat Biol Relat Stud Phys Chem Med 27:413–424
Potten CS, Gandara R, Mahida YR, Loeffler M, Wright NA (2009) Thestem cells of small intestinal crypts: where are they? Cell Prolif42:731–750
Reynolds ES (1963) The use of lead citrate at high pH as an electron-opaque stain in electron microscopy. J Cell Biol 17:208–212
Russel L, Burguet S (1977) Ultrastructure of Leydig cells as revealed bysecondary tissue treatment with a ferrocyanide-osmium mixture.Tissue Cell 9:751–766
Sala FG, Kunisaki SM, Ochoa ER, Vacanti J, Grikscheit TC (2009)Tissue-engineered small intestine and stomach form from autolo-gous tissue in a preclinical large animal model. J Surg Res156:205–212
Sato T, Vries RG, Snippert HJ, van de Wetering M, Barker N, StangeDE, van Es JH, Abo A, Kujala P, Peters PJ, Clevers H (2009)Single Lgr5 stem cells build crypt-villus structures in vitro withouta mesenchymal niche. Nature 459:262–265
Sato T, Stange DE, Ferrante M, Vries RG, van Es JH, Van den Brink S,Van Houdt WJ, Pronk A, Van Gorp J, Siersema PD, Clevers H(2011) Long-term expansion of epithelial organoids from humancolon, adenoma, adenocarcinoma, and Barrett’s epithelium.Gastroenterology 141:1762–1772
Scoville DH, Sato T, He XC, Li L (2008) Current view: intestinal stemcells and signaling. Gastroenterology 134:849–864
Shaker A, Rubin DC (2010) Intestinal stem cells and epithelial-mesenchymal interactions in the crypt and stem cell niche. TranslRes 156:180–187
Shaker A, Rubin DC (2012) Stem cells: one step closer to gut repair.Nature 485:181–182
Stelzner M, Helmrath M, Dunn JCY, Henning SJ, Houchen CW, Kuo C,Lynch J, Li L, Magness ST, Martin MG, Wong MH, Yu J (2012) Anomenclature for intestinal in vitro cultures. Am J PhysiolGastrointest Liver Physiol 302:1359–1363
Takeda N, Jain R, LeBoeuf MR, Wang QH, Lu MM, Epstein JA (2011)Interconversion between intestinal stem cell populations in distinctniches. Science 334:1420–1424
Tian H, Biehs B, Warming S, Leong KG, Rangell L, Klein OD, deSauvage FJ (2011) A reserve stem cell population in smallintestine renders Lgr5-positive cells dispensable. Nature478:255–259
Udassin R, Vromen A, Haskel Y (1994) The time sequence of injuryand recovery following transient reversible intestinal ischemia. JSurg Res 56:221–225
Van Landeghem L, Santoro MA, Krebs AE, Mah AT, Dehmer JJ,Gracz AD, Scull BP, McNaughton K, Magness ST, Lund PK(2012) Activation of two distinct Sox9-EGFP-expressing intes-tinal stem cell populations during crypt regeneration afterirradiation. Am J Physiol Gastrointest Liver Physiol 302:1111–1132
von Furstenberg RJ, Gulati AS, Baxi A, Doherty JM, Stappenbeck TS,Gracz AD, Magness ST, Henning SJ (2011) Sorting mouse jejunalepithelial cells with cd24 yields a population with characteristicsof intestinal stem cells. Am J Physiol Gastrointest Liver Physiol300:G409–G417
Wong MH (2004) Regulation of intestinal stem cells. J Invest DermatolSymp Proc 9:224–228
Wong VWY, Stange DE, Page ME, Buczacki S, Wabik A, Itami S, vande Wetering M, Poulsom R, Wright NA, Trotter MWB, Watt FM,Winton DJ, Clevers H, Jensen KB (2012) Lrig1 controls intestinalstem-cell homeostasis by negative regulation of Erbb signalling.Nat Cell Biol 14:401–408
Cell Tissue Res (2013) 354:441–450 449

Wright NA (2000) Epithelial stem cell repertoire in the gut: clues to theorigin of cell lineages, proliferative units and cancer. Int J ExpPathol 81:117–143
Yan KS, Chia LA, Li X, Ootani A, Su J, Lee JY, Su N, Luo Y, HeilshornSC, Amieva MR, Sangiorgi E, Capecchi MR, Kuo CJ (2012) Theintestinal stem cell markers Bmi1 and Lgr5 identify two function-ally distinct populations. Proc Natl Acad Sci USA 109:466–471
Yesus YW, Kenneally C, Taylor HM (1981) Preservation of hemato-poietic stem cells in cadaveric marrow. Am J Clin Pathol 76:205–207
Yui S, Nakamura T, Sato T, Nemoto Y, Mizutani T, Zheng X, IchinoseS, Nagaishi T, Okamoto R, Tsuchiya K, Clevers H, Watanabe M(2012) Functional engraftment of colon epithelium expandedin vitro from a single adult Lgr5(+) stem cell. Nat Med 18:618–623
450 Cell Tissue Res (2013) 354:441–450
Copyright © 2022 FDOKUMEN




















