
Interspecific Differences in the Phenotypic Plasticity of Intertidal Barnacles in Response to...
10
BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, research libraries, and research funders in the common goal of maximizing access to critical research. Interspecific Differences in the Phenotypic Plasticity of Intertidal Barnacles in Response to Habitat Changes Author(s): Boris A. López, Roxana P. Ramírez, Sandra Y. Guaitro, and Daniel A. López Source: Journal of Crustacean Biology, 30(3):357-365. 2010. Published By: The Crustacean Society DOI: http://dx.doi.org/10.1651/09-3211.1 URL: http://www.bioone.org/doi/full/10.1651/09-3211.1 BioOne (www.bioone.org ) is a nonprofit, online aggregation of core research in the biological, ecological, and environmental sciences. BioOne provides a sustainable online platform for over 170 journals and books published by nonprofit societies, associations, museums, institutions, and presses. Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use . Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiries or rights and permissions requests should be directed to the individual publisher as copyright holder.
Transcript of Interspecific Differences in the Phenotypic Plasticity of Intertidal Barnacles in Response to...

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, researchlibraries, and research funders in the common goal of maximizing access to critical research.
Interspecific Differences in the Phenotypic Plasticity of Intertidal Barnacles inResponse to Habitat ChangesAuthor(s): Boris A. López, Roxana P. Ramírez, Sandra Y. Guaitro, and Daniel A. LópezSource: Journal of Crustacean Biology, 30(3):357-365. 2010.Published By: The Crustacean SocietyDOI: http://dx.doi.org/10.1651/09-3211.1URL: http://www.bioone.org/doi/full/10.1651/09-3211.1
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological, andenvironmental sciences. BioOne provides a sustainable online platform for over 170 journals and books publishedby nonprofit societies, associations, museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance ofBioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiriesor rights and permissions requests should be directed to the individual publisher as copyright holder.

INTERSPECIFIC DIFFERENCES IN THE PHENOTYPIC PLASTICITY OF INTERTIDAL BARNACLES IN
RESPONSE TO HABITAT CHANGES
Boris A. Lopez, Roxana P. Ramırez, Sandra Y. Guaitro, and Daniel A. Lopez
(BAL, correspondence, [email protected]); (RPR, [email protected]); (SYG, [email protected]); (DAL, [email protected]);
Departamento de Acuicultura y Recursos Acuaticos, Universidad de Los Lagos, Casilla 933, Osorno, Chile
A B S T R A C T
We evaluate the morphological variation of cirri of two intertidal barnacles in response to different wave-exposure regimes and
population densities during growth. Length, diameter and number of segments of the third and sixth cirri were determined in Jehlius
cirratus and Notochthamalus scabrosus, two dominant sympatric species in the rocky intertidal zone of Chile. Aggregated and isolated
individuals of both species were collected in two coastal localities. Reciprocal transplant experiments were undertaken between wave-
protected and wave-exposed zones, in addition to experimental modification of density during growth. For both species, individuals
inhabiting wave-protected zones had cirri that were longer, thinner, with a greater number of segments, than in those inhabiting wave-
exposed zones. The sixth cirri of J. cirratus were longer, thicker and had more segments in specimens growing at high densities, than in
individuals growing in isolation. In N. scabrosus, no density dependent effects were observed in cirral structure. Thus, phenotypic
responses differed between species, depending on the morphometric or meristic variable of the cirral pair evaluated. Forty-five days after
J. cirratus density conditions and wave-exposure regime were modified in the transplant experiments, changes were observed in cirral
length, diameter, and number of cirral segments. No modification was observed in the cirral characteristics over time in specimens
whose density was artificially reduced during growth. Results indicate that morphological structures associated with filtration and
respiration, vary according to density and degree of wave-exposure. We suggest that cirral phenotypic variability can serve as an
adaptive mechanism associated with the changing availability and access to food and contributes to explaining the success of these
organisms in environments with high spatial-temporal fluctuations in biotic and abiotic factors.
KEY WORDS: barnacles, cirral morphology, density dependence, phenotypic plasticity, transplants, wave-exposure
DOI: 10.1651/09-3211.1
INTRODUCTION
Phenotypic plasticity is the variation of morphologicalcharacteristics in individuals of the same species, inresponse to local environmental variables (Stearns, 1989).The capacity to present different phenotypes can improvesurvival and reproduction in populations that inhabitheterogeneous environments (Lively, 1999). Phenotypicplasticity has been reported in different species of sessile orsemi-sessile marine invertebrates that inhabit the intertidalzone, due to their exposure to high spatial-temporalfluctuations in physical, chemical and biological factors(Kaandorp, 1999; Okamura and Partridge, 1999; Arsenaultet al., 2001; Zhang, 2006). In the particular case ofbarnacles, it has been reported that exposure to predatorscan stimulate the rotation or reduction of the opercularaperture area (Lively, 1986a, b; Jarrett, 2008, 2009). Wave-exposure level also produces morphological changes in thecalcareous structures and the chitinous exoskeleton ofindividuals (Pentcheff, 1991; Arsenault et al., 2001; Miller,2007; Neufeld and Palmer, 2008; Hoch, 2008, 2009). Thecirri of individuals in wave-exposed environments tend tobe shorter and thicker than those inhabiting wave-protectedzones (Arsenault et al., 2001; Marchinko and Palmer, 2003;Li and Denny, 2004; Miller, 2007). This has beeninterpreted as a mechanism to increase particle captureefficiency (Arsenault et al., 2001). Cirral morphology alsovaries according to the bathymetric distribution of individ-uals, cirri being longer in individuals inhabiting the upper
intertidal zone, than in those in the middle-lower zone(Chan and Hung, 2005). Modifications in cirral appendagesare produced over short periods of weeks, when they aresubject to different water current intensities under con-trolled conditions (Marchinko, 2003). Nevertheless, thereare no records of field experiments that verify these results.
Another factor that produces morphological variations inbarnacle structure is the density at which individuals grow.In various species, dense aggregates of barnacles form afterintense recruitment, resulting in intraspecific competitionfor space and food, with evidence of shell elongation(Barnes and Powell, 1950; Bertness et al., 1998), decreasein survival, growth and fecundity (Wu et al., 1977; Wu,1980; Wethey, 1983, 1984; Bertness, 1989; Thomason et al.,2000; Hills and Thomason, 2003a, b; Lopez andGonzalez, 2003; Jenkins et al., 2008) and penis length(Hoch, 2008). Lopez et al. (2007a) reported that cirri ofJehlius cirratus (Darwin, 1854) are longer, thicker and havea greater number of segments in individuals growing ingroups than in those growing in isolation. Nevertheless, it isnot known whether phenotypic plasticity of the cirri occursin response to changes in density conditions during growth.Similarly, there is no evidence verifying differences inthese phenotypic plasticity patterns between species.
The chthamalid barnacles J. cirratus and Notochthama-lus scabrosus (Darwin, 1854) are sympatric species in theintertidal zone of rocky beaches along the Chileancoastline. Jehlius cirratus is located between 2.4 and
JOURNAL OF CRUSTACEAN BIOLOGY, 30(3): 357-365, 2010
357

6.5 m above low-tide level and N. scabrosus, between 0.3and 3.2 m above low-tide level (Brattstrom, 1990; Pitomboand Ross, 2002; Lopez et al., 2007b). Morphologicalvariations have been observed in the shell and cirralappendages of J. cirratus and attributed to primarysubstrate limitations with evidence that individuals growingat high densities have more elongated shells, a smallerbasal area, and greater hard-part weight than specimens thatgrow in isolation (Lopez and Gonzalez, 2003).
The aim of this study is to determine interspecificvariations in the morphological changes of intertidalbarnacle cirri, that could be generated both by densityduring growth and wave-exposure levels. In addition, wepropose to verify the level of cirral phenotypic plasticity,when density conditions and wave-exposure level aremodified experimentally in the field.
MATERIALS AND METHODS
In order to test the hypothesis that wave-exposure level and density ofindividuals during growth modify the cirral structures of the intertidalbarnacles J. cirratus and N. scabrosus, two types of evaluation wereundertaken: descriptive and experimental.
Descriptive Approach: Interspecific Variations in Effect ofWave-Exposure Levels and Densities During Growth on Cirral
Phenotypic Plasticity
Specimens of J. cirratus and N. scabrosus were collected between May andAugust 2005, from the middle-low intertidal zone (0.5 and 3.5 m) of twolocalities: Pucatrihue (40u339S; 73u439W), a wave-exposed zone with anaverage tidal amplitude of 1.5 m and Metri Bay (21u369S, 72u439W), awave-protected zone, with an average tidal amplitude of 7m (Fig. 1). Thecriterion used to define areas as wave-exposed or wave-protected, was theheight of prevailing waves within the approximate ranges identified by Hoch(2008). In Pucatrihue, wave heights of 2-4 m or more was frequent duringapproximately 80% of the study period, whereas in Metri Bay, the prevailingwave height was always below 1 m. Prevailing current velocity was alsodetermined in both sites. Current values only exceptionally exceed 1 m/s inMetri Bay, whereas in Pucatrihue, values over 2 m/s were frequent. Currentmeasurements were undertaken consecutively over 7 days, and maximumcurrent velocity was recorded twice daily. This was carried out during twospring tide periods over one month. Individuals growing isolated (no contactwith neighbouring specimens) and individuals growing in groups wereconsidered in each site. Densities observed for both species were 0.51 6
0.38 individuals/cm2 in specimens growing in isolation and 4.13 6 0.18individuals/cm2 in specimens growing in groups. Size range in specimens ofJ. cirratus fluctuated between 3.5 and 4.5 mm carino-rostral length and in N.scabrosus, between 2.7 and 3.6 mm carino-rostral length. Carino-rostrallength corresponds to maximum distance between the carinal plate and therostral plate, and is a density-independent size measurement (Lopez andGonzalez, 2003). Forty specimens per species, for each locality andgrouping level, were analyzed during growth. In addition to carino-rostrallength, the following variables were measured with respect to the third andsixth pairs of cirri from the left side of each of these specimens: total length,diameter, and number of segments.
Experimental Approach: Effect of Modifications in Wave-Exposure Leveland Density During Growth on Phenotypic Plasticity of Cirri of J. Cirratus
Experiment 1: Transplants Between Localities with Different Wave-Exposure Levels.—Reciprocal transplants were undertaken of specimensof J. cirratus attached to substrata, such as rocks or stones (15 to 20 cmdiameter, approximately). The grouping level of specimens was the same(4.13 6 0.18 individuals/cm2) and they were transported from a wave-protected zone (Metri Bay) to a wave-exposed zone (Pucatrihue) and viceversa. Transplants began in Spring, 2006 (October and November); non-transplanted specimens from each zone, of the same size and growing atthe same density, were used as controls. Size range of the transplanted
specimens and controls fluctuated between 2.0 and 4.5 mm carino-rostrallength. Individuals were collected every 15 days for a total of 45 days afterthe transplants, in order to verify variations in cirral morphology. Fivespecimens were removed per site and collection period.
Experiment N2: Density Modification During Growth.—The naturaldensity of J. cirratus specimens growing in groups (4.1 6 0.2individuals/cm2) was reduced in the upper-middle intertidal zone of MetriBay (3 m). Specimens were eliminated mechanically, at random, creatingartificially solitary individuals, with no mural plate contact. Groupedspecimens, in physical contact with neighbouring individuals, togetherwith specimens growing naturally in isolation, were used as controls. Thisexperiment began in winter, 2006 (between June and July). The cirri ofspecimens within a size range of 3.5 and 5.5 mm carino-rostral length,were analyzed every 15 days, over a period of 45 days. In each treatmentand control, 5 specimens per experimental period were analyzed.
Measurement of Morphometric and Meristic Characteristics ofCirral Structure.—In both the descriptive and experimental evaluations,specimens collected were fixed in 20% alcohol and transported to theMarine Aquaculture Laboratory of the Universidad de Los Lagos, Osorno,Chile, where morphometric and meristic analyses of the cirri were carriedout. Individuals were dried, and the third and sixth pairs of cirri from theleft side of each specimen, extracted. Both cirral branches from the leftside were measured, and the average value was used for statisticalanalyses. Carino-rostral length was also measured with a graduated ocularlens. Each cirral pair was placed in lugol with sea water for staining andobserved under a stereoscopic microscope equipped with a camera lucida.Scaled drawings of the branches (endopod and exopod) of each cirral pairwere prepared and measurements taken from the drawings. The followingvariables were determined: total cirral length, that is, distance between thebase and the distal extreme; cirral diameter, between the first and secondarticulation of the external branch; and number of segments of eachbranch, from the base to the distal extreme.
Statistical Analysis
Descriptive Approach.—Average values were calculated for each variablemeasured in the external and internal cirral branches. The length anddiameter values of the cirri were transformed logarithmically, using Log10
(x + 1), while radicular transformation was applied to number of segments.Previous suppositions of normality and homocedasticity were verifiedthrough Shapiro-Wilk and Levene analyses, respectively (Zar, 1999). In
Fig. 1. Geographic location of the two localities where studies werecarried out; Pucatrihue (40u339S; 72u439W), wave-exposed zone and MetriBay (41u369S, 72u439W), wave-protected zone, in southern Chile.
358 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 30, NO. 3, 2010

order to verify interspecific variations in the morphometric and meristicresponses of the cirri to different densities during growth and wave-exposure levels, two-way analysis of covariance (ANCOVA) was appliedseparately, for each species (Zar, 1999). In the case of density, factorswere: a) density during growth (isolated and grouped); and b) species (J.cirratus and N. scabrosus). For wave-exposure, the first factor was level ofexposure (exposed and protected) and the second, species (J. cirratus andN. scabrosus). In all analyses, carino-rostral length of specimens of bothspecies, was considered as a covariate. Statistical analyses were carried outusing the STATISTICA 7.0 (Statsoft, 2004) statistical package.
Experimental Approach.—Prior to analyzing the phenotypic responses ofthe cirri, one-way variance analysis was undertaken to verify that sizes(measured as carino rostral length), were similar between transplantedindividuals v/s controls and between the different density and growthcategories. Average values for each variable were calculated for both cirralbranches. The normality and homoscedasticity of data were evaluated,using the Shapiro Wilk and Levene tests, respectively (Zar, 1999). Toevaluate cirral structural changes in both experiments (artificial modifi-cation of density and transplants between sites with different wave-exposure), a two-way variance analysis measurement was used, after datatransformation; logarithmic transformation, Log10 (x + 1), was used forlength and cirral diameter and the square root transformation, for numberof cirral segments. When significant variations were identified betweentreatments, an a posteriori Tukey analysis (Sokal and Rohlf, 1995) wasundertaken. All statistical analyses were carried out using the STATIS-TICA 7.0 (Statsoft, 2004) statistical package.
RESULTS
Descriptive Approach: Interspecific Variations in Effect ofWave-Exposure Levels and Densities During Growth on
Cirral Phenotypic Plasticity
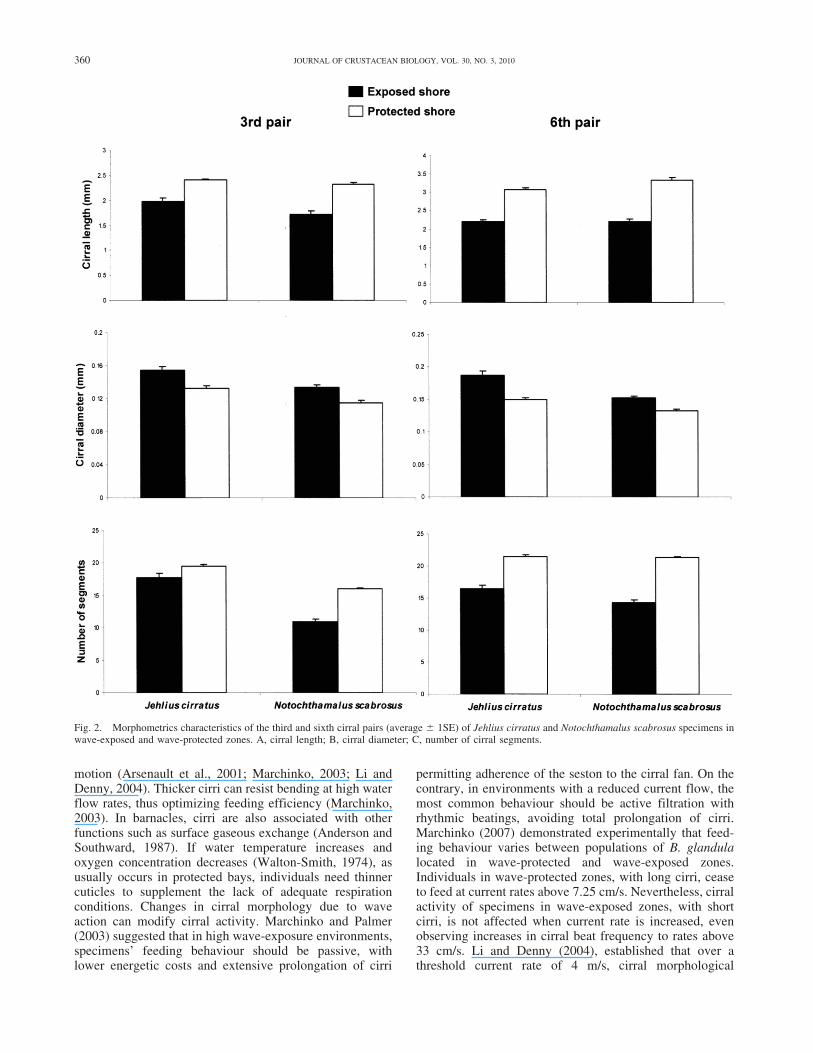
Specimens of both species had longer, thinner cirri, and agreater number of segments in wave-protected than inwave-exposed environments (Fig. 2). Nevertheless, cirrallength of N. scabrosus specimens, proportional to size, wasgreater than that of individuals of J. cirratus (F 5 8.39; d.f.5 1; 152; P 5 0.004); while cirral diameter and number ofcirral segments, was greater in J. cirratus than in N.scabrosus (F 5 7.02; d.f. 5 1; 152; P 5 0.008).
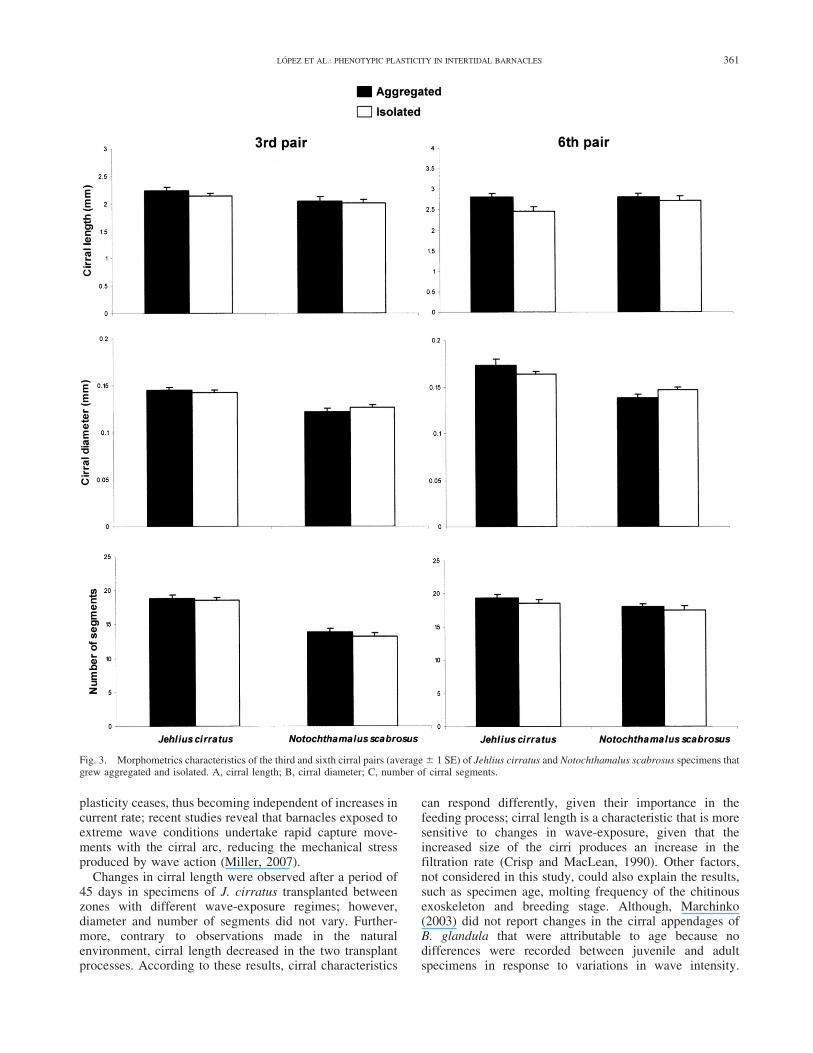
Density during growth affected the morphometric andmeristic characteristics of the sixth cirral pair of J. cirratus.Individuals of this species growing in groups presentedlonger cirri (F 5 6.91; d.f. 5 1; 152; P 5 0.009), and agreater number of segments (F 5 4.043; d.f. 5 1; 152; P 50.046) than those growing isolated (Fig. 3). No morpho-logical variations resulting from grouping during growth,were identified in the sixth cirral pair of N. scabrosus. Inthe third cirral pair, density did not generate differences inlength, diameter or number of cirral segments in either ofthe two species.
Experimental Approach: Effect of Modifications in Wave-Exposure Levels and Density During Growth on the
Phenotypic Plasticity of Cirri of J. Cirratus
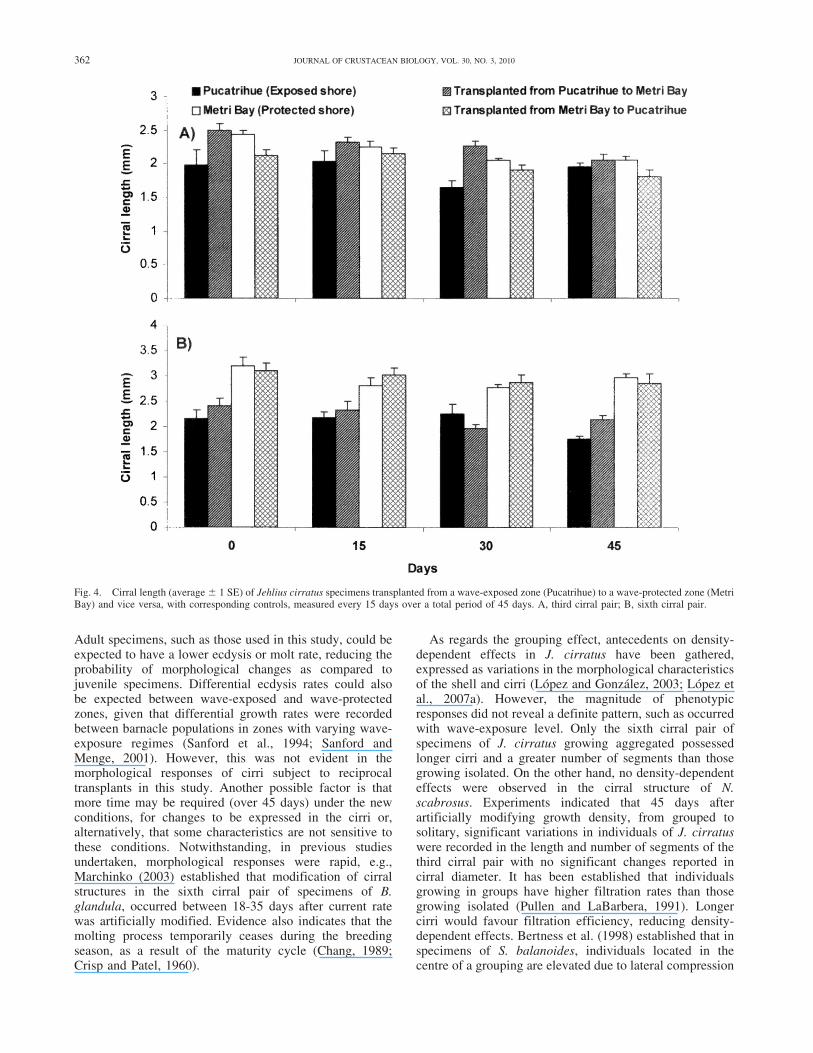
Changes in wave-exposure regimes modified the cirralmorphology of J. cirratus specimens, depending on thedirection of the wave-exposure change carried out duringthe transplant and on the cirral pair evaluated. In specimenstransplanted from a wave-protected zone (Metri Bay) to awave-exposed zone (Pucatrihue), a relative decrease inlength of the third cirral pair occurred after 45 days, incomparison to the non-transplanted controls (F 5 13.33;
d.f. 5 1; 112; P , 0.05) (Fig. 4). This did not occur in thesixth cirral pair (F 5 0.81; d.f. 5 1; 112; P 5 0.371). Therewere no statistically significant differences between thecarino-rostral length of transplanted specimens and thecontrols (F 5 0.2; d.f. 5 1; 118; P 5 0.656). Nodifferences in diameter and number of cirral segmentsoccurred between treatment and controls, during theexperimental period (Table 1).
In the case of specimens transplanted from a wave-exposed zone (Pucatrihue) to a wave-protected zone (MetriBay), a relative decrease in size of the third cirral pair wasalso observed, compared with control specimens (F 513.04; d.f. 5 1; 112; P , 0.05). This did not occur in thesixth cirral pair (F 5 1.54; d.f. 5 1; 112; P 5 0.216). Adecrease in the diameter of the third cirral pair wasobserved in both the control specimens and the transplantedspecimens (F 5 12.88; d.f. 5 3; 112; P , 0.05). Number ofsegments in the third cirral pair was statistically greater intransplanted specimens than in control specimens, al-though, in both treatments, no changes were recorded after45 days (Table 1). This did not occur in the sixth cirral pair.The transplanted and control specimens had the samecarino-rostral length (F 5 1.9; d.f. 5 1; 118; P 5 0.171).
Changes in density during growth did not modify cirralmorphological characteristics. Forty-five days after densitywas artificially modified, whereby specimens growing ingroups were isolated, lengths and segment numbers of thethird and sixth pairs of cirri were statistically greater thanthose control individuals growing isolated (F 5 6.21; d.f. 52; 48; P , 0.05) (Fig. 5). Nevertheless, there were nodifferences in these variables between the groupedindividuals and those that initially grew in groups, butwere then artificially transformed into solitary individuals.There were no significant differences in diameter in thecirral types evaluated during the experimental period(Table 2). The experimental and control specimens didnot differ in carino-rostral length (F 5 0.16; d.f. 5 2; 57; P5 0.852).
DISCUSSION
Phenotypic plasticity, in response to wave-exposure level,has been established in the cirral appendages of variousspecies of barnacles, such as Balanus glandula Darwin,1854, Tetraclita japonica Pilsbry, 1916, Chthamalus dalliPilsbry, 1916 and Semibalanus cariosus Pallas, 1788(Marchinko, 2003; Marchinko and Palmer, 2003; Chanand Hung, 2005). In this study, significant variations in thelength, diameter and number of cirral segments wereobserved in the intertidal species J. cirratus and N.scabrosus with the cirri being comparatively longer,thinner, and with more segments in the wave-protectedzone than in the wave-exposed zone. This variation mayresult from the need to maximize particle capture rates,depending on the environmental current velocity. In wave-protected areas, it would be beneficial to possess longercirri, because they can filter a greater volume of water percirral arc movement, thus increasing filtration rateefficiency. In high wave-exposure zones, shorter andthicker cirri would reduce the mechanical drag of the wave
LOPEZ ET AL.: PHENOTYPIC PLASTICITY IN INTERTIDAL BARNACLES 359
motion (Arsenault et al., 2001; Marchinko, 2003; Li andDenny, 2004). Thicker cirri can resist bending at high waterflow rates, thus optimizing feeding efficiency (Marchinko,2003). In barnacles, cirri are also associated with otherfunctions such as surface gaseous exchange (Anderson andSouthward, 1987). If water temperature increases andoxygen concentration decreases (Walton-Smith, 1974), asusually occurs in protected bays, individuals need thinnercuticles to supplement the lack of adequate respirationconditions. Changes in cirral morphology due to waveaction can modify cirral activity. Marchinko and Palmer(2003) suggested that in high wave-exposure environments,specimens’ feeding behaviour should be passive, withlower energetic costs and extensive prolongation of cirri
permitting adherence of the seston to the cirral fan. On thecontrary, in environments with a reduced current flow, themost common behaviour should be active filtration withrhythmic beatings, avoiding total prolongation of cirri.Marchinko (2007) demonstrated experimentally that feed-ing behaviour varies between populations of B. glandulalocated in wave-protected and wave-exposed zones.Individuals in wave-protected zones, with long cirri, ceaseto feed at current rates above 7.25 cm/s. Nevertheless, cirralactivity of specimens in wave-exposed zones, with shortcirri, is not affected when current rate is increased, evenobserving increases in cirral beat frequency to rates above33 cm/s. Li and Denny (2004), established that over athreshold current rate of 4 m/s, cirral morphological
Fig. 2. Morphometrics characteristics of the third and sixth cirral pairs (average 6 1SE) of Jehlius cirratus and Notochthamalus scabrosus specimens inwave-exposed and wave-protected zones. A, cirral length; B, cirral diameter; C, number of cirral segments.
360 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 30, NO. 3, 2010
plasticity ceases, thus becoming independent of increases incurrent rate; recent studies reveal that barnacles exposed toextreme wave conditions undertake rapid capture move-ments with the cirral arc, reducing the mechanical stressproduced by wave action (Miller, 2007).
Changes in cirral length were observed after a period of45 days in specimens of J. cirratus transplanted betweenzones with different wave-exposure regimes; however,diameter and number of segments did not vary. Further-more, contrary to observations made in the naturalenvironment, cirral length decreased in the two transplantprocesses. According to these results, cirral characteristics
can respond differently, given their importance in thefeeding process; cirral length is a characteristic that is moresensitive to changes in wave-exposure, given that theincreased size of the cirri produces an increase in thefiltration rate (Crisp and MacLean, 1990). Other factors,not considered in this study, could also explain the results,such as specimen age, molting frequency of the chitinousexoskeleton and breeding stage. Although, Marchinko(2003) did not report changes in the cirral appendages ofB. glandula that were attributable to age because nodifferences were recorded between juvenile and adultspecimens in response to variations in wave intensity.
Fig. 3. Morphometrics characteristics of the third and sixth cirral pairs (average 6 1 SE) of Jehlius cirratus and Notochthamalus scabrosus specimens thatgrew aggregated and isolated. A, cirral length; B, cirral diameter; C, number of cirral segments.
LOPEZ ET AL.: PHENOTYPIC PLASTICITY IN INTERTIDAL BARNACLES 361
Adult specimens, such as those used in this study, could beexpected to have a lower ecdysis or molt rate, reducing theprobability of morphological changes as compared tojuvenile specimens. Differential ecdysis rates could alsobe expected between wave-exposed and wave-protectedzones, given that differential growth rates were recordedbetween barnacle populations in zones with varying wave-exposure regimes (Sanford et al., 1994; Sanford andMenge, 2001). However, this was not evident in themorphological responses of cirri subject to reciprocaltransplants in this study. Another possible factor is thatmore time may be required (over 45 days) under the newconditions, for changes to be expressed in the cirri or,alternatively, that some characteristics are not sensitive tothese conditions. Notwithstanding, in previous studiesundertaken, morphological responses were rapid, e.g.,Marchinko (2003) established that modification of cirralstructures in the sixth cirral pair of specimens of B.glandula, occurred between 18-35 days after current ratewas artificially modified. Evidence also indicates that themolting process temporarily ceases during the breedingseason, as a result of the maturity cycle (Chang, 1989;Crisp and Patel, 1960).
As regards the grouping effect, antecedents on density-dependent effects in J. cirratus have been gathered,expressed as variations in the morphological characteristicsof the shell and cirri (Lopez and Gonzalez, 2003; Lopez etal., 2007a). However, the magnitude of phenotypicresponses did not reveal a definite pattern, such as occurredwith wave-exposure level. Only the sixth cirral pair ofspecimens of J. cirratus growing aggregated possessedlonger cirri and a greater number of segments than thosegrowing isolated. On the other hand, no density-dependenteffects were observed in the cirral structure of N.scabrosus. Experiments indicated that 45 days afterartificially modifying growth density, from grouped tosolitary, significant variations in individuals of J. cirratuswere recorded in the length and number of segments of thethird cirral pair with no significant changes reported incirral diameter. It has been established that individualsgrowing in groups have higher filtration rates than thosegrowing isolated (Pullen and LaBarbera, 1991). Longercirri would favour filtration efficiency, reducing density-dependent effects. Bertness et al. (1998) established that inspecimens of S. balanoides, individuals located in thecentre of a grouping are elevated due to lateral compression
Fig. 4. Cirral length (average 6 1 SE) of Jehlius cirratus specimens transplanted from a wave-exposed zone (Pucatrihue) to a wave-protected zone (MetriBay) and vice versa, with corresponding controls, measured every 15 days over a total period of 45 days. A, third cirral pair; B, sixth cirral pair.
362 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 30, NO. 3, 2010
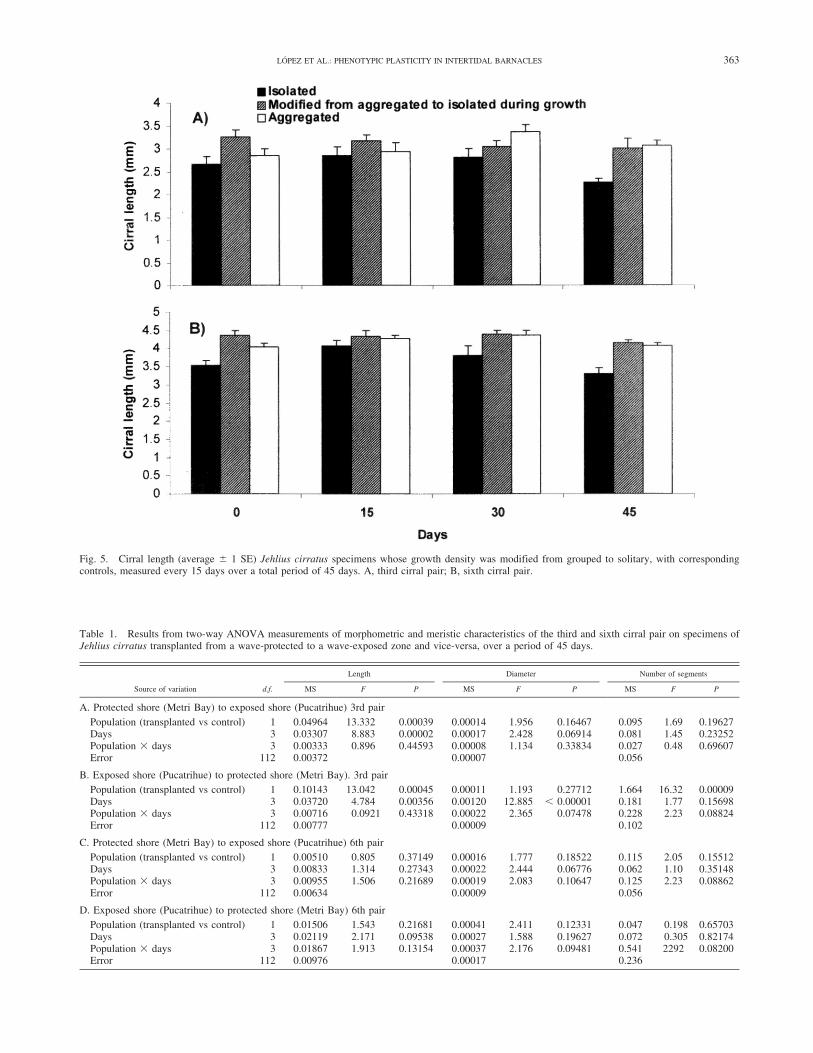
Fig. 5. Cirral length (average 6 1 SE) Jehlius cirratus specimens whose growth density was modified from grouped to solitary, with correspondingcontrols, measured every 15 days over a total period of 45 days. A, third cirral pair; B, sixth cirral pair.
Table 1. Results from two-way ANOVA measurements of morphometric and meristic characteristics of the third and sixth cirral pair on specimens ofJehlius cirratus transplanted from a wave-protected to a wave-exposed zone and vice-versa, over a period of 45 days.
Source of variation d.f.
Length Diameter Number of segments
MS F P MS F P MS F P
A. Protected shore (Metri Bay) to exposed shore (Pucatrihue) 3rd pair
Population (transplanted vs control) 1 0.04964 13.332 0.00039 0.00014 1.956 0.16467 0.095 1.69 0.19627Days 3 0.03307 8.883 0.00002 0.00017 2.428 0.06914 0.081 1.45 0.23252Population 3 days 3 0.00333 0.896 0.44593 0.00008 1.134 0.33834 0.027 0.48 0.69607Error 112 0.00372 0.00007 0.056
B. Exposed shore (Pucatrihue) to protected shore (Metri Bay). 3rd pair
Population (transplanted vs control) 1 0.10143 13.042 0.00045 0.00011 1.193 0.27712 1.664 16.32 0.00009Days 3 0.03720 4.784 0.00356 0.00120 12.885 , 0.00001 0.181 1.77 0.15698Population 3 days 3 0.00716 0.0921 0.43318 0.00022 2.365 0.07478 0.228 2.23 0.08824Error 112 0.00777 0.00009 0.102
C. Protected shore (Metri Bay) to exposed shore (Pucatrihue) 6th pair
Population (transplanted vs control) 1 0.00510 0.805 0.37149 0.00016 1.777 0.18522 0.115 2.05 0.15512Days 3 0.00833 1.314 0.27343 0.00022 2.444 0.06776 0.062 1.10 0.35148Population 3 days 3 0.00955 1.506 0.21689 0.00019 2.083 0.10647 0.125 2.23 0.08862Error 112 0.00634 0.00009 0.056
D. Exposed shore (Pucatrihue) to protected shore (Metri Bay) 6th pair
Population (transplanted vs control) 1 0.01506 1.543 0.21681 0.00041 2.411 0.12331 0.047 0.198 0.65703Days 3 0.02119 2.171 0.09538 0.00027 1.588 0.19627 0.072 0.305 0.82174Population 3 days 3 0.01867 1.913 0.13154 0.00037 2.176 0.09481 0.541 2292 0.08200Error 112 0.00976 0.00017 0.236
LOPEZ ET AL.: PHENOTYPIC PLASTICITY IN INTERTIDAL BARNACLES 363
of neighbouring individuals and have privileged access tothe particle flow, increasing their feeding rates incomparison to those specimens located on the margin ofthe grouping. Furthermore, it has been verified that densityin barnacles can affect opercular valve closure (‘‘hidingbehaviour’’). This reduces active feeding time (Dill andGillet, 1991; Mauck and Harkless, 2001). Anotherinterpretation for results related to cirral modificationduring growth, is based on the supposition that densitylevels were not sufficient to be expressed as morphologicalvariations. Lopez et al. (2007a), reported the existence of athreshold density value (1.73 6 0.18 individual/cm2) forindividuals of J. cirratus, above which phenotypic changeswould occur in the cirri.
Results also reveal interspecific differences in themodification of cirral appendages. The barnacle J. cirratuswas more sensitive to fluctuations in wave-exposureconditions and density than N. scabrosus. This can beinterpreted in terms of the bathymetric distribution of bothspecies; J. cirratus inhabits the upper intertidal zone, whileN. scabrosus inhabits the middle-lower intertidal zonewhere tidal fluctuations are lower and food availability maybe greater. This would result in less pressure to modifycirral structures. Marchinko and Palmer (2003) reportedinterspecific differences in the cirral phenotypic responsesto wave-exposure level in the balanomorph barnacles C.dalli, S. balanoides, B. glandula, and a lepadormoph,Pollicipes polymerus Sowerby, 1833; variations in cirrallength, cirral diameter, setal length, and intersetal spacewere more pronounced in balanomorph barnacles whilevariations in P. polymerus were less pronounced due to themorphological variations and feeding strategy of thisspecies. The results of this study also reveal variations inthe responses of the cirri evaluated (third and sixth pairs),identifying different functions in the feeding process ofbarnacles (Anderson and Southward, 1987; Anderson,1994). Cirri of the sixth pair specialize in the collectionof suspended food, extending completely outside theopercular plate, as a result of which they would beexpected to be more sensitive to environmental changes.On the contrary, the food filtering manipulation function ofthe third pair is more passive; consequently fewervariations would be expected. Nevertheless, this responsewas not observed in the present study, where modifications
were observed in the third pair of cirri of specimens of J.cirratus following changes in wave-exposure regimes anddensity, suggesting that cirral functionality could varyaccording to species.
Results suggest that certain characteristics of the cirralappendages are more adaptable or respond more rapidly tothe experimental changes, both in density during growth andwave-exposure, this being directly related to maximizationof food particle capture rates. This would suggest thatmorphological variation depends on the range of environ-mental conditions present in the habitat and would alsoexplain the adaptive ability of both species to successfullycolonize different substrate levels, with different spatial-temporal variability gradients. Also, our results indicate thatan analysis of cirral morphology is not a prudent means bywhich taxonomists can delineate species.
ACKNOWLEDGEMENTS
This study was financed by project Nu00205F and Nu0109, ResearchDepartment of the Universidad de Los Lagos and FONDEF ProjectD03I1116. The collaboration of Claudio Sepulveda, Alexis Santibanez,Sergio Arriagada, Veronica Dıaz, Daniela Perez and Jose Uribe, in fieldactivities is gratefully acknowledged, as is the assistance of Susan Angusin the translation of the manuscript. Finally, our thanks to the twoanonymous referees, whose comments contributed to improving themanuscript.
REFERENCES
Anderson, D. T. 1994. Barnacles. Structure, Function, Development andEvolution. Chapman and Hall. London, UK.
———, and A. J. Southward. 1987. Cirral activity of barnacles, pp. 135-174. In, A. J. Southward (ed.), Crustacean Issues 5: Barnacle Biology.A. A. Balkema, Rotterdam.
Arsenault, D. J., K. B. Marchinko, and A. R. Palmer. 2001. Precise tuningof barnacle leg length to coastal wave action. Proceedings of the RoyalSociety of London B 268: 2149-2154.
Barnes, H., and H. T. Powell. 1950. The development, generalmorphology, and subsequent elimination of barnacle population aftera heavy initial settlement. Journal of Animal Ecology 19: 175-179.
Bertness, M. D. 1989. Positive and negative density-dependent mortalityand the population structure of Semibalanus balanoides in sheltered bayhabitat. Ecology 70: 257-268.
———, S. D. Gaines, and S. M. Yeh. 1998. Making mountains out ofbarnacles: The dynamic of acorn barnacle hummocking. Ecology 74:1382-1394.
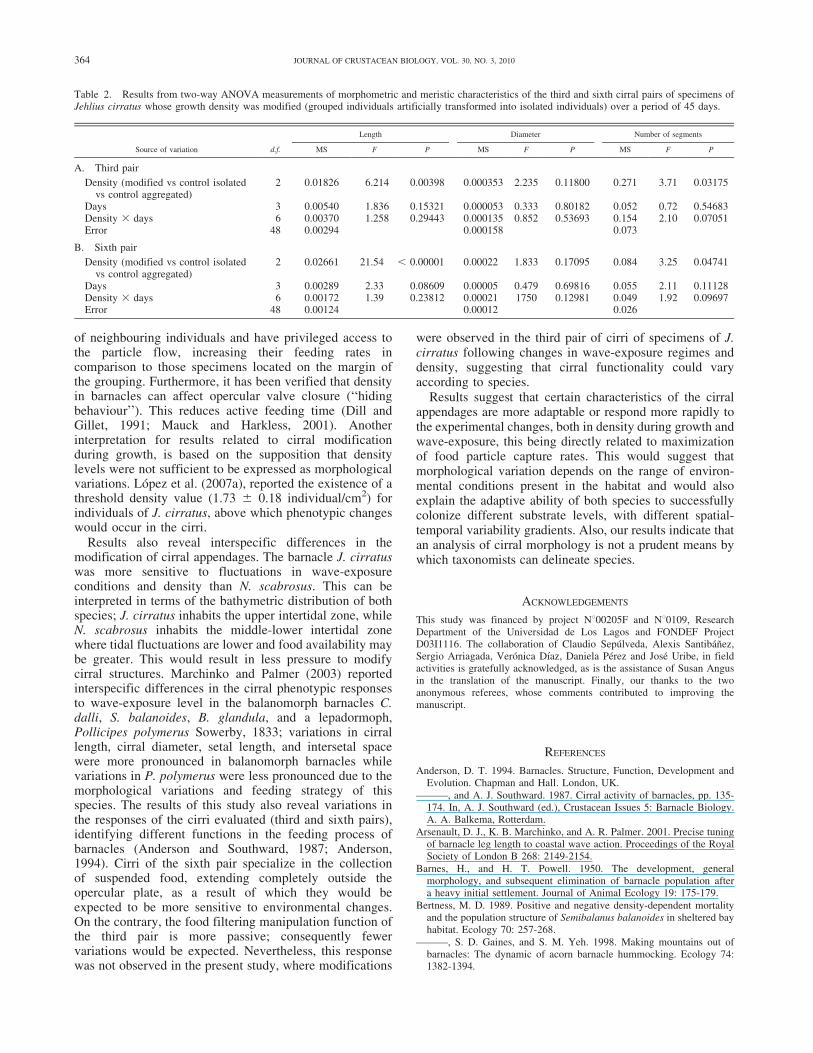
Table 2. Results from two-way ANOVA measurements of morphometric and meristic characteristics of the third and sixth cirral pairs of specimens ofJehlius cirratus whose growth density was modified (grouped individuals artificially transformed into isolated individuals) over a period of 45 days.
Source of variation d.f.
Length Diameter Number of segments
MS F P MS F P MS F P
A. Third pair
Density (modified vs control isolatedvs control aggregated)
2 0.01826 6.214 0.00398 0.000353 2.235 0.11800 0.271 3.71 0.03175
Days 3 0.00540 1.836 0.15321 0.000053 0.333 0.80182 0.052 0.72 0.54683Density 3 days 6 0.00370 1.258 0.29443 0.000135 0.852 0.53693 0.154 2.10 0.07051Error 48 0.00294 0.000158 0.073
B. Sixth pair
Density (modified vs control isolatedvs control aggregated)
2 0.02661 21.54 , 0.00001 0.00022 1.833 0.17095 0.084 3.25 0.04741
Days 3 0.00289 2.33 0.08609 0.00005 0.479 0.69816 0.055 2.11 0.11128Density 3 days 6 0.00172 1.39 0.23812 0.00021 1750 0.12981 0.049 1.92 0.09697Error 48 0.00124 0.00012 0.026
364 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 30, NO. 3, 2010

Brattstrom, H. 1990. Intertidal ecology of the northern part of the ChileanArchipelago. Sarsia 75: 105-160.
Chan, B. K., and O. S. Hung. 2005. Cirral length of the acorn barnacleTetraclita japonica (Cirripedia : Balanomorpha) in Hong Kong: effect ofwave exposure and tidal height. Journal of Crustacean Biology 25: 329-332.
Chang, E. S. 1989. Endocrine regulation of moulting in crustacean.Review of Aquatic Science 1: 131-157.
Crisp, D. J., and B. S. Patel. 1960. The moulting cycle in Balanusbalanoides (L). The Biological Bulletin 118: 31-47.
———, and F. J. MacLean. 1990. The relation between the dimensions ofthe cirral net, the beat frequency and the size and age of the animal inBalanus balanoides and Elminius modestus. Journal of the MarineBiological Association of the United Kingdom 70: 351-389.
Darwin, C. R. 1854. A monograph on the Sub-class Cirripedia, with figuresof all species. The Balanidae, the Verrucidae. Ray. Society, London.
Dill, L. M., and J. F. Gillett. 1991. The economic logic of barnacle Balanusglandula (Darwin) hiding behaviour. Journal of Experimental MarineBiology and Ecology 153: 115-127.
Hills, J. M., and J. C. Thomason. 2003a. The ‘‘ghost of settlement past’’determines mortality and fecundity in the barnacle, Semibalanusbalanoides. Oikos 101: 529-583.
———, and ———. 2003b. Recruitment density can determine adultmorphology and fecundity in the barnacle, Semibalanus balanoides.Biofouling 19: 205-213.
Hoch, J. M. 2008. Variation in penis morphology and mating ability in theacorn barnacle, Semibalanus balanoides. Journal of ExperimentalMarine Biology and Ecology 359: 126-130.
———. 2009. Adaptative plasticity of the penis in a simultaneoushermaphrodite. Evolution 63: 1946-1953.
Jarrett, J. N. 2008. Inter-population variation in shell morphology of thebarnacle Chthamalus fissus. Journal of Crustacean Biology 28: 16-20.
———. 2009. Predator-induced defense in the barnacle Chthamalus fissus.Journal of Crustacean Biology 29: 329-333.
Jenkins, S. R., J. Murua, and M. T. Burrows. 2008. Temporal changes inthe strength of density-dependent mortality and growth in intertidalbarnacles. Journal of Animal Ecology 77: 573-584.
Kaandorp, J. A. 1999. Morphological analysis of growth forms ofbranching marine sessile organisms along environmental gradients.Marine Biology 134: 295-306.
Li, N. K., and M. W. Denny. 2004. Limits to phenotypic plasticity: Floweffects on barnacle feeding appendages. The Biological Bulletin 206 (3):121-124.
Lively, C. M. 1986a. Competition, comparative life histories, andmaintenance of shell dimorphism in a barnacle. Ecology 67: 858-864.
———. 1986b. Predators-induced shell dimorphism in the acorn barnacleChthamalus anisopoma. Evolution 40: 232-242.
———. 1999. Developmental strategies in spatially variable environ-ments: Barnacle dimorphism and strategic models of selection, pp. 245-258. In, R. Tollrain and C. D. Harvell (eds.), The Ecology and Evolutionof Inducible Defenses. Princeton University Press, Princeton.
Lopez, D. A., and M. L. Gonzalez. 2003. Density-dependent effects inJehlius cirratus (Darwin, 1854) (Cirripedia: Chthamalidae) underdifferent growth conditions. Marine Ecology 24: 289-302.
———, S. Y. Guaitro, and D. A. Lopez. 2007a. Density-dependent effectson the cirral structure of the barnacle Jehlius cirratus (Darwin 1854)(Cirripedia). Crustaceana 80: 793-801.
———, B. A. Lopez, and M. L. Gonzalez. 2007b. Bibliographic index onaquatic biodiversity of Chile: Crustacea, Cirripedia, Thoracica.Ciencia y Tecnologıa del Mar 30: 161-165.
Marchinko, K. B. 2003. Dramatic phenotypic plasticity in barnacle legs(Balanus glandula Darwin): magnitude, age dependence, and speed ofresponse. Evolution 56: 1281-1290.
———. 2007. Feeding behavior reveals the adaptive nature of plasticity inbarnacle feeding limbs. The Biological Bulletin 213: 12-15.
———, and A. R. Palmer. 2003. Feeding in flow extremes dependence ofcirrus form on wave-exposure in four barnacle species. Zoology 106:127-141.
Mauck, R. A., and K. C. Harkless. 2001, The effect of group membershipon hiding behaviour in the northern rock barnacle, Semibalanusbalanoides. Animal Behaviour 62: 743-748.
Miller, L. P. 2007. Feeding in extreme flows: behaviour compensates formechanical constraints in barnacle cirri. Marine Ecology Progress Series349: 227-234.
Neufeld, C. J., and A. R. Palmer. 2008. Precisely proportioned: intertidalbarnacles alter penis form to suit coastal wave action. Proceedings of theRoyal Society of London B 275: 1081-1087.
Okamura, B., and J. C. Partridge. 1999. Suspension feeding adaptations toextreme flow environments in a marine bryozoan. Biological Bulletin196: 205-215.
Pallas, P. S. 1788. Voyages de M.P.S. Pallas, En Differentes Provinces deL’empire de Russie, et Dans L’asie Septentrionale. Cinq Volumes in 4,et Un de Planches.
Pentcheff, N. D. 1991. Resistance to crushing from wave-borne debris inthe barnacle Balanus glandula. Marine Biology 110: 399-408.
Pilsbry, H. A. 1916. The sessile barnacles (Cirripedia) contained in thecollections of the U.S. National Museum; including a monograph of theAmerican species. Bulletin of the United States National Museum 93: 1-366.
Pitombo, F. B., and A. Ross. 2002. A checklist of the intertidal andshallow-water sessile barnacles of the Eastern Pacific, Alaska to Chile.Contributions to the Study of East Pacific Crustaceans 1: 97-106.
Pullen, J., and M. LaBarbera. 1991. Modes of feeding in aggregations ofbarnacles and the shape of aggregations. The Biological Bulletin 181:442-452.
Sanford, E., and B. A. Menge. 2001. Spatial and temporal variation inbarnacle growth in a coastal upwelling system. Marine EcologyProgress Series 209: 143-157.
———, D. Bermudez, M. D. Bertness, and S. D. Gaines. 1994. Flow, foodsupply and acorn barnacle population dynamics. Marine EcologyProgress Series 104: 49-62.
Sokal, R. R., and F. J. Rohlf. 1995. Biometry: The Principles and Practices ofStatistics in Biological Research. Third edition. W. H. Freeman, New York.
Sowerby, G. B. 1821-1834. The Genera of Recent and fFossil Shells.London. 1: pls. 1-126 + text (not numbered) (1821-1825); 2: pls. 127-162 + text (not numbered) (1825-1834).
Statsoft, Inc. 2004. STATISTICA (data analysis software system), version7. www.statsoft com.
Stearns, S. C. 1989. The evolutionary significance of phenotypic plasticity.BioScience 39: 436-445.
Thomason, J. C., J. M. Hills, and P. Mapson. 2000. The consequence ofseasonal reproductive strategies for the interpretation of settlementtrials. Biofouling 16: 171-179.
Walton-Smith, F. G. 1974. CRC Handbook of marine science. CRC Press,Cleveland, Ohio, U.S.A.
Wethey, D. S. 1983. Intrapopulation variation in growth of sessileorganism: natural population of the intertidal barnacle Balanusbalanoides. Oikos 40: 14-23.
———. 1984. Effects of crowding on fecundity in barnacles: Balanusbalanoides, Balanus glandula, and Chthamalus dalli. Canadian Journalof Zoology 62: 1788-1795.
Wu, R. S. S. 1980. Effects of crowding on the energetics of the barnacleBalanus glandula (Darwin). Canadian Journal of Zoology 58: 559-556.
———, C. D. Levings, and D. J. Randall. 1977. Differences in energypartitioning between crowded and uncrowded individual barnacles.Canadian Journal of Zoology 55: 643-647.
Zar, J. H. 1999. Biostatistical Analysi.s Fourth edition. Prentice Hall, NewJersey.
Zhang, Y. 2006. Balancing food availability and hydrodynamic constraint:phenotypic plasticity and growth in Simulum noelleri blackfly larvae.Oecologia 147: 39-46.
RECEIVED: 26 August 2009.ACCEPTED: 2 October 2009.
LOPEZ ET AL.: PHENOTYPIC PLASTICITY IN INTERTIDAL BARNACLES 365




















