
INTERINDIVIDUAL ASSOCIATIONS IN DOGS
30
INTERINDIVIDUAL ASSOCIATIONS IN DOGS by BURNEY J. LE BOEUF 1) (Department of Psychology, University of California, Berkeley, Calif., U. S. A.) (With 7 Figures) (Rec. 3I-1-i967) CONTENTS I. Introduction .269 II. Subjects, Materials and Methods.270 III. Results ..275 A. Heterosexual Interactions .275 i. Normative data from the anestrous, estrous, and post-estrous periods of females .. .275 2. The response of individual females to individualmales 277 3. Behavior in the circle when females were in estrus . . . . 277 4. A comparison of present and previous mating preferences 28I 5. Behavior in the circle when females were in nonestrus 282 6. Temporal changes during estrus .. .282 7. Individual differences among males . . . .285 8. Differences in the testing condition, tethered vs roving . . 286 B. Intrasexual Interactions ..286 i. Female-femalebehavior ..286 2. Male-male behavior ..288 C. Normative Comparisons ..290 IV. Discussion .. .29I Summary .. .292 References .. .293 Resume ...294 i) Present address: Crown College, University of California, Santa Cruz, California, U.S.A. This research was conducted at the Field Station for Research on Animal Behavior in Berkeley, California and was supported by United States Public Health Service Grant MH-o4noo to Dr FRANK A. BEACH. The work reportedis a condensation of a Ph.D. thesis submitted to the University of California at Berkeley. I wish to thank Drs P. R. MARLER and G. E. FRENCH for reading and commenting on the original manu- script and particularly, Dr FRANK A. BEACH, who overseeredthe work from its inception to the writing of the final draft.
-
Upload
khangminh22 -
Category
Documents
-
view
4 -
download
0
Transcript of INTERINDIVIDUAL ASSOCIATIONS IN DOGS

INTERINDIVIDUAL ASSOCIATIONS IN DOGS
by
BURNEY J. LE BOEUF 1)
(Department of Psychology, University of California, Berkeley, Calif., U. S. A.)
(With 7 Figures)
(Rec. 3I-1-i967)
CONTENTS
I. Introduction .269 II. Subjects, Materials and Methods .270
III. Results ..275
A. Heterosexual Interactions .275
i. Normative data from the anestrous, estrous, and post-estrous periods of females .. .275
2. The response of individual females to individual males 277 3. Behavior in the circle when females were in estrus . . . . 277 4. A comparison of present and previous mating preferences 28I 5. Behavior in the circle when females were in nonestrus 282 6. Temporal changes during estrus .. .282 7. Individual differences among males . . . .285 8. Differences in the testing condition, tethered vs roving . . 286
B. Intrasexual Interactions ..286 i. Female-female behavior ..286 2. Male-male behavior ..288
C. Normative Comparisons ..290
IV. Discussion .. .29I Summary .. .292 References .. .293 Resume ...294
i) Present address: Crown College, University of California, Santa Cruz, California, U.S.A. This research was conducted at the Field Station for Research on Animal Behavior in Berkeley, California and was supported by United States Public Health Service Grant MH-o4noo to Dr FRANK A. BEACH. The work reported is a condensation of a Ph.D. thesis submitted to the University of California at Berkeley. I wish to thank Drs P. R. MARLER and G. E. FRENCH for reading and commenting on the original manu- script and particularly, Dr FRANK A. BEACH, who overseered the work from its inception to the writing of the final draft.

INTERINDIVIDUAL ASSOCIATIONS IN DOGS 269
I. INTRODUCTION
Numerous allusions to individual preferences, friendships, personal bonds, sympathies and antipathies, etc. can be found in the literature on the domestic dog. These citations emanate from a broad source which includes dog owners, breeders, veterinarians (WHITNEY, I947; HANCOCK & ROWLANDS, I949;
HARROP, I96o), and well-known investigators of behavior (ENDERS, I945; TINBERGEN, I951; LORENZ, I952; KINSEY et al, I923; BEACH, I96I). The most frequent report deals with strong attractions between particular mem- bers of the opposite sex when the female is in estrus. Some authorities claim the male shows preferences for certain females while others report that it is the female that selects and refuses to mate with certain males. Whichever the case may be, one thing is certain: all of these references are based on subjective impressions or observations, which are unsubstantiated by ex- perimental data.
Two studies present quantitative data. FULLER & DuBuIs (I962) reported sexual behavior scores for a single fox terrier female observed separately with two males of the same breed during portions of her first estrus. Attempts to mate the bitch with a litter mate were unsuccessful, but when a strange male experienced as a stud was introduced copulation was effected within two minutes.
In a more extensive study using 5 male and 5 female beagles, BEACH & LE BOEUF (in press) observed differences in each female's readiness to mate with different masculine partners over two estrous periods. Each female mated readily with some males but repeatedly rejected the copulatory attempts of others. Females attempting to prevent mounting barked, growled, and snapped at males or attacked and bit them. The same males were not always rejected by all females; a male preferred by one female was frequently rejected by another. A female's preferences were consistent from her first estrous period through a subsequent one six months to a year later.
The foregoing study was not designed specifically to investigate the phenomena of selective mating and it is probable that certain features in the testing procedure obscured the full magnitude of female preferences. In some tests the manner in which the animals were released made it possible for a genital lock to occur during the first few seconds after the start of a test. This had two results. The female was tied before she had time to look the male over, so to speak, and once the animals were locked, no further scoring of preferential behavior was possible. When copulation did not occur during the first few seconds of a test, the female exerted obvious control over the mating situation, copulating readily with some males while repeatedly

270 BURNEY J. LE BOEUF
rejecting others, usually for the entire I5-minute observation period. Further- more, each female was permitted to lock just once a day each day of her estrous period. After a lock, no further tests were conducted on that day with other males. Consequently when the female locked with the first male tested one could not say what might have resulted if a different male had been used. As a result of this procedure, all males were not tested on the same days with the estrous bitch and some males copulated more frequently than others. Lastly, rejection behavior by the female was the only measure of individual preferences employed.
The primary purpose of the present experiments was to extend the study of BEACH & LE BOEUF and, specifically, to investigate interindividual asiso- ciations during, before, and after mating in a test situation which maximized the frequency of opportunities for interaction. The procedure was designed to facilitate the expression and description of individual preferences by: (a) permitting all males to have equal time with estrous females and vice versa, (b) alleviating the difficulties presented by occurrence of the genital lock, (c) permitting one animal in a test pair to control the frequency and nature of interaction and, in effect, choose among several partners, and (d) measuring preferences, in several different ways. In addition, the nature and frequency of associations of males with males and of females with females was investigated.
SUBJECTS, MATERIALS AND METHODS
S u b j e c t s'.
The subjects (Ss) were 9 pure bred, 3-year-old Beagles of which 5 were males and 4 were females. These same animals were used in the study by BEACH & LE BOEUF (in press). They lived together in a 3/4 acre enclosed field (see Figure i) from the time they were weaned to the beginning of the study, with only occasional separation of the sexes when females were in estrus. During this experiment when the females were not being tested, they were caged and the males ran free in the field. The Ss were fed after the completion of a day's tests.
S etti ng.
Tests were conducted in a fenced enclosure measuring approximately I,700 sq. ft which formed part of the unit where the Ss lived. Observations were made from a small cabin on the premises which was designed speci- fically for the purpose. The cabin contained recording instruments and afforded the observer an excellent view of the testing area (see Figures i and 2).

~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~tIC 0
- ~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~-
Figure 2. The testing area. A tethered animal is restricted to the area surrounded by the small circle (the large circle is not relevant to the present study).

.. ... ....... _ ... .:::::: .:
.....................................................................:...:::. .,..x,j ..*
_ S ' . . ,~~~~~~~~~~~~~~~~~~~~~~~~~~~~..... .. ...
Figure 3. A female wearing a chastity belt.

INTERINDIVIDUAL ASSOCIATIONS IN DOGS 27I
CABIN
STAKE AND CIRCLE
ENCLOSED TESTING AREA
N
/ CAGES _
Figure i. A schematic of the living unit and the enclosure where tests were conducted. The field area measures approximately 34 acre.
P r o c e d u r e.
The behavior of the following pair combinations was observed in the same test situation: (a) heterosexual pairs before, during, and after the female was in estrus, (b) female pairs before, during, and after one of them was in estrus, and (c) male-male pairs. The test situation provided a setup in which one animal could completely avoid contact with the other or terminate contact at any time. One S was tethered to a stake by a 5 ft chain attached to a fitted harness, and thus resitricted to a circular area with a diameter of approximately I2 ft (see Figure 2). The perimeter of this- circular area was outlined with gypsum to facilitate observations and scoring behavior. The other S, the roving animal, was released into the testing en- closure from the observer's cabin situated 20 ft from the tethering stake. The roving animal was allowed 5 minutes in the field, the duration of each test. It is reported that the type of interactions observed between an estrous female and a male dog do not change substantially if the test duration is extended beyond 5 minutes to 30 minutes (BEACH & LE BOEUF, in press). Daily tests with each pair of Ss included two observations, one while the animal was tethered and one while it was roving free.
Behavioral items recorded.
The following gross measures were recorded: (a) whether or not the roving animal went to the tethered animal and made physical contact, (b) the time spent by the roving animal in the circle with the tethered animal, and

272 BURNEY J. LE BOEUF
(c) the number of entrances and departures into and out of the circle by the roving animal. Behavioral interactions between the roving and the tethered S in the circle were described with categories similar to those used by BEACH & LE BOEUF (in press). In this schema, shown in Table i, an attempt was made to reduce behavioral interactions to discrete stimulus-response units or episodes. The term "episode" was used to refer to the initiating act of one animal plus the response of the other.
TABLE I
Categories of social and sexual behavior in dogs
Stimulus Units of Behavior* Code Description of Behavior
I Approach: moves to within 2 feet 2 Partial investigation: brings nose within i foot of anogenital region or makes
momentary contact with that area 3 Investigation: sniffs and/or licks anogenital area for at least 5 seconds 4A Pre-mount contact: partial mount, forepaws on back, chin over back 4B Prance: forepaws extended, body thrown back on haunches, head cocked to
one side; barking and vigorous tail wagging may be involved 4C Present: hindquarters directed to the other animal and tail is deviated laterally 5 Mount: forelimb clasp without pelvic thrusts 6 Clasp and thrust: Same as 5 with several pelvic thrusts 7 Behazior of intromission: same as 6 but thrusts are more rapid and include
pelvic oscillation, "stepping and treading" of the hindlegs
Response Units of Behavior **
Code Description of Behavior Ai Stand: Maintains erect posture for at least 5 seconds when mounted or in-
vestigated A2 Stand with Tail Deviation: same as Ai plus lateral deviation of tail B Face: whirls around to face other animal when investigated or mounted C Sit: the nonerect female does not rise when mounted or investigated D Avoid: Moves or pulls away, sits or lies down to avoid mount or investigation E Threat: barks, growls, snaps, lunges or any combination which prevents the
initiation or continuation of behavior by the other animal F Attack: chase, drive, bite or any combination
The estrous period.
A female's estrous period was defined and determined by a combination of behavioral signs, genital swelling and/or bleeding, and vaginal smears
* Males display all behaviors except 4C. Females display all behaviors described. Any activity not directed to the rear of the other animal but rather to the head or side is suffixed by an "H" or an "S".
** All response units are shown by the female. The male responds to a female by standing (AI) or by moving away (D). Females terminate Ai and A2 responses with B, D, E, or F, or they stand until dismounted.

INTERINDIVIDUAL ASSOCIATIONS IN DOGS 273
taken before each day's tests. A female was classified as being in behavioral estrus on those days in which the following conditions were satisfied: (a) swelling with or without vaginal discharge, (b) a vaginal smear of the late proestrous or estrous type, i.e., primarily cornified epithelium, with or without red blood cells, and with but few nucleated epithelial cells, (c) the female "stood", deviated her tail laterally and permitted a male to mount, clasp her hindquarters and execute several pelvic thrusts. With the first two conditions satisfied, one mount with clasp and thrusts by one male, in which the female stood and deviated her tail, was sufficient to classify the female as being in estrus on that day. The behavioral criterion of standing and "curling the tail", alone, has frequently been employed as an indication of estrus in the bitch (WHITNEY, I947; HANCOCK & ROWLANDS, I949; GRA-
HAM, I96I; ASDELL, I966). One female, Blanche, was tested while in natural estrus. The other three
females were brought into estrus by means of 2 intramuscular injections of estrogen 1) (.38 mglkg of estradiol benzoate, Schering "Progynon", 3.3 mg/cc in sesame oil), spaced 48 hours apart. The hormone was administered during the anestrous period, 4-6 months after a female's previous natural estrus. It was known from previous work that the bitch usually comes into estrus I-5 days after the second injection and remains, in estrus for approximately IO days.
Special equipment.
In heterosexual tests it was important to maintain the female stimulus to each male in as constant a condition as possible. To do this, and in order to run several combinations of tests per day, the occurrence of complete copulation with insertion and genital lock was prevented. This was effected by fitting each with a "chastity" belt which covered the entire anogenital area and prevented males from achieving insertion (see Figure 3). The belt was made of thin vinyl plastic, and small holes in the transparent material permitted the female to urinate and allowed for the transmission of olfactory and gustatory cues to the male. After a brief period of adaptation, females became accustomed to wearing the belt. It did not impede the movements of the female nor did it produce any apparent decrease in her interest in the male nor the male's interest in the female.
It was also necessary to adapt both males and females to the harnesses and the tether. This was accomplished during the same 6-day pre-experimental
i) The hormones were supplied by Dr PRESTON L. PERLMAN of the Biological Research Department, Schering Corporation, Bloomfield, New Jersey.

274 BURNEY J. LE BOEUF
period during which females were adapted to the belts. Harnesses were worn at all times; chastity belts were installed for tests only.
Observation periods and their duration.
Tests involving estrous females were started 8 days before the female received her first estrogen injection and continued without interruption (except for a few days when the weather conditions made observation impossible) until the ioth day after the female was last mounted. The 8 obser- vation days prior to injections were called the anestrous period; days with mounts in which the female stood and deviated the tail constituted the estrous period; and the io days following the last day of estrus was called the post- estrous period. Anestrus and post-estrus combined were called nonestrus.
Combinations or groups of Ss tested.
Since tests with each S included observations when the animal was tethered and when he or she was roving free, and since each female was tested during estrus and nonestrus, several combinations' of sex and female "reproductive" conditions resulted. These combinations and their abbreviations (called groups) are shown in Table 2 and will be referred to in a later section.
TABLE 2
Combinations of subjects tested
Groups Roving Animal Tethered Animal
M-EF male estrous female EF-M estrous female male M-NEF male nonestrous female NEF-M nonestrous female male NEF-EF nonestrous female estrous female EF-NEF estrous female nonestrous female NEF-NEF nonestrous female nonestrous female M-M male male
Time and order of testing.
Two females were brought into estrus at the start of the experiment while the other two females served as anestrous Ss. When testing was completed on the first two females, they served as anestrous Ss and a third female, used formerly as an anestrous S, was brought into estrus. The last S came into natural estrus during the post-estrous' tests of the first two females tested.
Heterosexual pairs and female-female pairs were observed on the same days with the former always tested first in the day. All tests involving

INTERINDIVIDUAL ASSOCIATIONS IN DOGS 275
females were completed before observation of male-male pairs were initiated. Each male was tested IO times with each other male over a period of 30 days.
To control for possible order effects in the daily tests, individuals of one sex were run first to individuals of the other sex on alternate days. For example, an estrous female was run to males before they were run to her on one day and the procedure was reversed on the following day. When two females were being tested on the same day, they were alternately run first and second. Within each group, the order of release or tethering was varied in a way which prevented systematic error.
RESULTS
HETEROSEXUAL INTERACTIONS
Normative data f rom the anestrous, estrous, and post-estrous periods of females.
Table 3 shows the percentage of days that the roving dog visited the tethered dog and initiated a behavioral episode. Males visited females on 93.lYo of the days that the latter were in estrus, an increase over the number of days females were visited when they were in anestrus (78.6%) and post-estrus (72.0%). Similarly, females visited males more frequently when they were in estrus (76.2%) than in anestrus (27.8%) or post-estrus (4255%). Under each female condition, males visited females more often than females visited males.
The percentage of total test time spent by the roving dog in the circle with the tethered dog is shown in Table 4. The findings represented in this table support the conclusions suggested by Table 3. According to these two measures, the sexes are generally most attracted to each other when the female is in estrus.
Detailed examination of Table 3 and 4 shows that one female was far more responsive to males than were the other three females. Kate's visits to males totaled 89%, ioo% and ioo% of the tests during her anestrous, estrous, and post-estrous periods, respectively. The response of the other three females was much lower during their respective periods. Kate also spent more time with males, once she visited them, than did the other females. The magnitude of these individual dif ferences precludes further normative analyses. A more meaningful approach, for our purposes, is to examine the response of individuals, the source of the variation.
Behaviour XXIX I8

TABLE 3
Percentage of days that the roving dog visited the tethered dog
FEMALES
PEGGY SPOT BLANCHE KATE MEANS
Days ~~~~Tethered Roving Tethered Roving Tethered Roving Tethered Roving Yy Tethered Roving
8 i8 io 8 i8 1o 8 12 I0 8 12 I0 8 12 I0 8 12 I0 g 8 i0 9 8 IO. MALES A E PA E_P A E P A E PA E P A E P A E P A E P A E P IA E P
CLARK 75 100 100 12 22 0 100 100 30 0 I17 0 100 100 100 0 42 0 89 100 1001 100 100 100 9I.0 100 82.5 28.0 45.2 25.0
JOHN 12 78 20 12 83 I10 38 50 20 0 100 30 50 67 5 0 0 100 30 33 i00 8o 100 100 100 33.2 73.8 42.5 28.0 95.8 42.5
EDDIE, 62 100 40 12 100 90 i00 83 40 0 92 20 70 83 7 12 5 0 89 00 70 78 100 100 o. 9I.5 55.0 25.5 792 52.5
KEN 88 i00 90 25 II 0 88 100 40 0 50 0 90 100 100 0 83 0 i00 i00 i00 89 i00 i00 91.5 i00 82.5 28.5 6i.0 25.0
BROADTUS 100 100 100 25 i00 8o 88 i00 90 0 100 50 100 100 100 12 100 40 100 100 100 78 i00 i00 97.0 100 9-7.5 28.8 i00 6-7.5
MEANS 167.4 95.6 700] 17.2 63.2 36.0] 82.8 86.6 44.0] 0 7I.8 20.0 182.0 90.0 84.0 4.8 70.0 140] 82.2 100 90.0] 89.0 100 i00o 78.6 93.I 72.0 27.8 -76.2 42.5
TABLE 4
Percentage of total test time spent by the roving dog in the circle with the tethered dog
FEMALES
PEGGY SPOT BLANCHE KATE MEANS Tethered Roving Tethered Roving Tethered Rovi'ng Tethered Roving YQ Tethered ~ Y Roving
MALES A E P1 A E P. A E__Pi A E P. A E__PI_A E P A E Pi A E__P7A E P _A E P
CLARK 2-7 30 12 4 6 0 4 6 i 0 2 -O 1 I7 41 20 0 I 0 39 8o 35 32 68 44.2i.8 39.21I7.0 9. 0 19.2 11I.0
JOHN I125 I 0 14 <1I 15 <1I 0 32 I 451I 4 0 38 I 2 2422 35 5459 :2.028.8 6.8 8.8 34.51I5.2
EDDIE, 5 72 3 <I 74 12 7 37 I 0 28 I 6 25 14 <1 I 0 15 40 21 39 47 36 8.2 43.5 9.8 9.8 37.8 12.2
KEN 33 381I3 I I 0 i6 34 2 01I3 0 2879 38 01I5 0 24 8I 34 28 68 58 25.2'58.218 7. 242 1.
BROADUS 21 8 2 I I I 84 1 4 31i 62 1 5 0 5 7 3 31I 8-8 34 <I 44 2 27 69 22 30 58 46 2 7.5 75.2 20.5 -7.8 6o.8 16.2
MENS '74I8o .2 35.8 521i8308 3.8 0 26. o 1725682. 0 20.0 .62I4 58.8 26.8] 32.8 59.0 48.6] 26.9 48.9 I25.2 8.5 35.3 13.8
A= Anestrus E = Estrus P= Pos testrus

INTERINDIVIDUAL ASSOCIATIONS IN DOGS 277
The response of individual females to individual males.
The group analysis showed that females in estrus were attracted to males. Although this statement is true it could be misleading if taken to imply that all females were universally attracted to all males. Tables 3 and 4 show that three of the four females visited some males more frequently than others and also spent more time with them. For example, Peggy visited Broadus and Eddie every day that she was in estrus and spent 84% and 74% of her test time with them in the circle. In contrast, she visited Clark and Ken only 22% and II%o of the time and spent only 6% and i.% of her time with them. Spot and Blanche showed similar variation in their response to males. It should also be noted that no female responded differentially to males during anestrus. However, response tendencies evident during estrus tended to persist into the post-estrus period.
These data suggest that all females were 'ot equally attracted to all males. In fact, some females spent no more time with certain males when they were in estrus than when they were in anestrus. This can be seen in the combinations: Peggy with Clark and Ken; Spot with Clark; and Blanche with Clark and Eddie as shown in Table 4. Hlowever, these differences in the response magnitude of individual estrous females derive from two gross, correlated measures, only. Additional insight is provided by the behavior of the pairs of dogs when they were together in the circle.
Behavior in the circle when females were in estrus.
Before presenting these results, some of the measures recorded must be described. When released into the field, a male usually approached the female, investigated her anogenital region, and if she was in estrus, mounted from the rear, clasping the female's hindquartersi and executing several pelvic thrusts. After approximately 5-30 seconds of thrusting without introa mission, the male dismounted or was thrown off by the female. This pattern was often repeated as much as 20 times in 5 minutes, and was not unlike behavior observed normally when the male has difficulty obtaining insertion. Since insertion was prevented in this study, a mount with clasp and thrusts (MCT) was the most nearly complete form of sexual activity possible.. A female was maximally cooperative if she stood and deviated her tail laterally when mounted. Ordinarily, this response would have facilitated insertion and the establishment of a genital lock.
The positive behavior of an estrous female to a male sometimes took an active form which some have called "'courtship" (FULLER & DuBuis, I962)
or "'solicitation" behavior (WHITNEY, 1947). The latter term is used generally

278 BURNEY J. LE BOEUF
to refer to all female-initiated activities described in Table i and indicated by the symbols: 4A, 4B, 4C, 5, 6, and 7. Activities directed to the head, side, and anogenital region of the male were all considered equally.
The estrous female was not always cooperative. Besides avoiding the male, the female might activily prevent sexual activity by (a) barking, growling or snapping and lunging at him or by (b) biting andlor driving him away. These two responses, symbolized as E and F in Table i, will be
TABLE 5
Peggy's response to 5 males during an i8-day estrous period
Roving Day of No. MCT Total No. Female Negative Episodes Males -- ist MCT Days MCTs Solicitations Proportion Percent
Tethered
Clark 1 - 9 I4 3 24 II 321117 27.4
9-+ I4 3 I5 I 1/29 34
John 3 - 9 8 7 34 20 3/94 3.2
8 6 I3 2 0/54 0
Eddie 0 - 9 5 13 I27 70 8/285 2.8
---> 3 15 99 142 0/230 0
Ken I.->9 I4 I 5 0 149/197 75.6
9 o 8 - 0 0 0 0/7 0
Broadus I - 9 I I7 I63 30 2/34I .5 2 i6 94 177 0/176 0
X -*8 0 o 9 41 353 131 194II034 i8.8 40 221 322 I /496 .2
TABLE 6
Spot's response to 5 moles during a 12-day estrous period
Roving Day of No. MCT Total No. Female Negative Episodes
Males -- Ist MCT Days MCTs Solicitations Proportion Percent
Tethered
Clark 8-9 _ o 0 0 I8/I9 94.7
8 o 0 - 0 0 0 7/I I 63.6
John R >9 9 3 7 O 10/41 24.4
8 8 4 7 4 2/62 3.2
Eddie 8 - 9 7 3 I0 0 37177 48. I
8 8 2 1 5 0 7/83 8.4
Ken $-*9 7 2 2 0 i5o/i66 90.4
9 o 8 0 0 0 I 6/38 42. I
Broadus z - 9 2 I0 56 o II/I58 7.0
9 e t I 9 49 4 3/1I39 2.2
X t e -
< 9 I8 75 0 226/46I 49.0 9 -o t 15 71 8 35/333 10.5

INTERINDIVIDUAL ASSOCIATIONS IN DOGS 279
TABLE 7
Blanche's response to 5 males during a I2-day estrous period
Roving Day of No. MCT Total No. Female Negative Episodes Males -> Ist MCT Days MCTs Solicitations Proportion Percent
Tethered Clark ! -e 9 3 2 2 0 28/62 45.2
Q e 8 0 0 0 i/8 I2.5
John 8 - 5 7 63 II 13/I48 8.8 9 < 8 4 9 46 33 i/io6 1.0
Eddie 8 - 9 5 I 5 0 54/73 74.0 Q o 0 0 0 i/8 12.5
Ken > 9 I Io 67 9 1331303 43.9 Q e 8 4 5 I4 I4 I/49 2.0
Broadus 8 -> I 9 98 i6 I8/233 8.I Q o 8 2 8 40 2I 4/96 4.2
X 8 -~ 8 z Q29 235 36 246/8i9 30.0 9 - t 22 ioo 68 8/267 3.0
TABLE 8
Kate's response to 5 males during an 8-day estrous period
Roving Day of No. MCT Total No. Female Negative Episodes Males -> Ist MCT Days MCTs Solicitations Proportion Percent
Tethered Clark - 9 2 7 74 0 io/i6i 6.2
9 _1 8 I 7 64 4 2/I4I I14
John 8 -9 5 2 6 0 0/27 0 Q < 4 4 I2 i6 2/61 3.3
Eddie ,3*9 7 I I I i1/26 3.8 9 < 4 2 23 3 0/80 0
Ken 8 < Q 2 5 33 0 32/i65 194
9 o 2 6 22 7 6/1i I2 5 4 Broadus 8 - 9 4 2 I5 0 6/85 7.I
Q < 4 4 I I 2 i1/64 i.6
X 8 - 8>9 I7 I29 I 491464 io.6 9 < 8 23 I32 32 1 I /458 24
called "threat" and "'attack", respectively. Negative episodes are those in which the female responded with threat or attack.
Tables 5, 6, 7, and 8 show the response of each female to each male for 3 categories of behavior: the time and frequency of MCTs, the frequency of solicitation behavior, and the percentage of negative episodes. The data in these tables show that Peggy, Spot, and Blanche, each reacted positively to some males and negatively to others, i.e., they showed preferences. This is

280 BURNEY J. LE BOEUF
best indicated by considering one fenmale in detail. For example, Peggy clearly preferred Broadus and Eddie to Clark and Ken. She vis,ited Broadus and Eddie every day that she was in estrus and spent the majority of her test time with them when they were tethered (Table 3). On the contrary, she rarely visited Clark and Ken and when she did she remained with them for only a short time (Table 4). Table 5 shows that on these few visits, Ken never mounted. Clark mounted Peggy for 3 days but not until the 14th day of her i8-day estrous period. On the other hand, Peggy's visits to Broadus and Eddie resulted in numerous mounts as early as Day 2 and 3, and for a period of i6 and 15 days, respectively.
When Peggy was tethered and the males ran free, Ken and Clark did not mount until the 14th day of her estrous period. Once again, Peggy's response to these two males contrasted sharply with her response to Broadus and Eddie, who mounted sooner, on more days and more times per day. Peggy solicited Eddie and Broadus frequently, particularly when they were tethered and she was free. She rarely solicited Clark and never solicited Ken.
The proportion of negative episodes to total episodes initiated by the male when he was roving indicates that Clark and Ken did not mount frequently because they were prevented from doing so by the female. Twenty-seven percent of the episodes initiated by Clark and 76%v of those initiated by Ken, Peggy actively repulsed with threats and attack. Markedly lower percentages on the same measure were obtained by the other 3 males: John (3.2%), Eddie (2.8%), and Broadus (0.5%).
The response of the other females can be compared to Peggy's (Tables- 6, 7, and 8). Both Spot and Blanche were most positive to Broadus and some- what less so to John. Spot, like Peggy, was most uncooperative with Clark and Ken. She did not stand to be mounted and she rejected them 9 out of every iO times that they initiated an episode. Blanche was positive to Ken, in that he mounted often and was at times solicited, but she also rejected him frequently. The most interesting point is that Blanche displayed the greatest aversion to Eddie, one of Peggy's favorites. This suggests that the antipathy elicited by a male does not depend on an attribute peculiar to him to which all females respond, e.g., his outward appearance. Eddie obviously affected Peggy and Blanche dif ferently.
Kate did not show the antipathies shown by other females. She visited each male every day that she was in estrus and spent from 47% to 68% of her time with them. With Kate, no one male ranks far above or far below any other on any of the measures shown in Table 8.
Preferences were suggested in still another type of behavior shown by the female which some investigators have called "teasing" (WHITNEY, 1947).

INTERINDIVIDUAL ASSOCIATIONS IN DOGS 28I
A female in estrus often terminates an investigation or mount by running away from the male. The male usually gives chase and after a short run, the female comes to an abrupt halt and may stand once again. If the male does not follow her, the female often returns to the male and solicits his attention (FULLER & DuBuis, I962; LE BOEUF, I967). Females frequently ran to, away from, and back to tethered males in this study and these data are shown in Table 9. The measure is the mean number of circle entrances by the female on days in which she visited tethered males.
TABLE 9
Mean number of circle entrances on days in which female visited the male
Roving Females
Tethered Peggy Spot Blanche Kate Males A E P A E P A E P A E P
Clark I I .5 0 0 I 0 0 I 0 1.3 2.5 I.,
John I.8 I 0 3.2 I 0 4.2 I 1.2 2.5 1.3
Eddie I 2.5 T.1 0 3.4 I.5 I I 0 L.I 2.2 1.1
Ken 2 I 0 0 I.7 0 0 2.9 0 1.2 2.8 1.2
Broadus I 3.0 I 0 4.4 I .4 I 4.2 I I .4 2.1 1.2
Means I.2 2.0 .6 0 2.7 o.8 0.4 2.7 0.4 T.2 2.4 1.2
A = anestrus E = estrus P = post-estrus
All females ran in and out of the circle most frequently when they were in the estrous condition. The mean number of circle entrances during this period for each female, with the exception of Kate, reflects the same preferences for particular males which were evident in the measures previously discussed. The conclusion is that females "'teased" preferred males only, or put more objectively, they returned to preferred males more often after leaving them than they did to nonpreferred males.
> A comparison of present and previous mating pref- e r en c e s.
The various differential responses of estrous females to certain males suggest preferences which are remarkably similar to those shown previously by these same females to the same males during their first and second natural estrous periods. Table io, adapted from BEACH & LE BOEUF (in

282 BURNEY J. LE BOEUF
press), shows the percentage of male acts (symbols i through 4A in Table i) to which females responded with threat or attack. Scores from both estrous periods are combined to form the rejection coefficients shown. The similarity and consistency of present and previous mating preferences are even more surprising when one considers the numerous differences already pointed out between the two studies, and the fact that a span of two years separated the first estrous periods from those observed in the present study. The present results also suggest that a female's, preferences are not attenuated by the artificial induction of estrus with exogenous hormones.
TABLE io Rejection coefficients from two estrous periods for each female with each
male (adapted from Beach and LeBoeuf, in press)
Female
Male Peggy Spot Blanche Kate ... . . .... ..... .... .... ..... ... .. ..... ...... .. .... ..... . ..... ... .. ................ ............ .... . . .. . . .......................................................................
Clark 33 6i 55 5 John 10 IO I5 2
Eddie 0 I2 58 12
Ken 8i 33 59 30
Broadus 0 4 6 o
Behavior in the circle when females were in non- estrous.
Only a small number of episodes took place during the anestrous and post-estrous periods of each female. No mounting occurred and solicitation was rare during these periods. A few negative episodes were observed during anestrus but they were distributed among several males. No male was rejected during post-estrus. More will be said about the anestrous and post-estrous periods in the next section.
Temporal changes during estrus.
In addition to displaying individual biases to males, females differed in more general ways amongst themselves. Each female responded to the administration lof exogenous hormones with different latencies to the onset of estrus and its duration (see Figures 4-7). Kate not only failed to show preferences, but was more "social" than other females when she was not in estrus (see Tables 3, 4 and 9). Peggy solicited frequently when she was in estrus while Spot solicited only rarely. Spot responded negatively to most

INTERINDIVIDUAL ASSOCIATIONS IN DOGS 283
8 0 I I I I I i
70 - PEGGY 0- MOUNTS BY MALES
A- SOLICITATION BY FEMALE
60 - X- NEGATIVE EPISODES
50 -
U ~~ ~~~~~~~A I z
8 40 r
30 A
30
/< II X
50~~~~~ __
1 2 3 4 56890 12 13 4 5 16 17 18 If ODAYS
Figure 4
80 I I I--- J I I I I I I I
70 - SPOT * - MOUNTS BY MALES
A- SOLICITATION BY FEMALE
S - NEGATIVE EPISODES
60-
50
:) 4 0-
130
20/1 \!
10 *~\ /
II I I~1 3 4I
DAYS
II 6 I1
Figure 5

284 BURNEY J. LE BOEUF
8 0 T ------------ - l I
70 BLANCHE *-MOUNTS BY MALES
A- SOLICITATION BY FEMALE
X - NEGATIVE EPISODES 60 -
540 -
30
10
IA---4.A---.-- --4i I I It
I 2 3 4 5 6 7 8 U 10 11 12
DAYS
Figure 6
80 --- -I I I I I I I I
70 KATE 0- MOUNTS BY MALES
A- SOLICITATION MY FEMALE |
U- XNEGATIVE EPISODES |\
60~~~~~~
1 2 3 4 5 6 7 8 - 0 11 1
l l ~~~~~DAYS
Figure 6
30~~~~~~~~~~~
60 -
50~~~~~~~~~~~~~~~~~~~~~~' D 40~~~~~~~~~~
CY~ ~ ~~~~~~~DY
Figures 4-7. Temporal changes in the frequency of MCTs, solicitation behavior and negative behavior. Estrogen was administered on the days indicated by the arrows.

INTERINDIVIDUAL ASSOCIATIONS IN DOGS 285
males far more frequently than did other females. The time course of the measures treated in Tables 5-8 are depicted in Figures 4-7. They show the temporal changes in the frequency with which females solicited, responded negatively or were mounted, from the time estrogen was administered (or proestrus began) to several days after the female ceased to allow mounting.
All females permitted relatively few mounts at first. Mount frequency rose, gradually with some females and more quickly with others, to a peak near the three quarter mark of the estrous period with Peggy and Spot and near the middle of the estrous period for Blanche and Kate.
The temporal order of negative behavior was quite different for each female. The peak was near the middle of Peggy's estrous period and did not coincide with those days in which she was most frequently mounted. Spot showed a bimodal distribution for mounts. However, she was most negative at the same time that she was most positive for mounts. Blanche, in natural estrus, was most negative during proestrus but the figure shows another peak of negative behavior which coincided with her high point for total mounts. Both Blanche and Peggy showed few rejection responses on the last few days of their estrous periods. Kate displayed a much different pattern of responce. She was most negative toward the end of her period when mount frequency was low. These data do not support the frequent suggestion (e.g., GRAHAM, 196T) that the female rejects the male only during the period immediately prior to and just after she is most "ready".
With Peggy, solicitation behavior closely paralleled mount frequency. Toward the end of her estrous period she continued to solicit when she was no longer mounted and she no longer rejected any male. Blanche behaved similarly, but she, unlike Peggy, did little soliciting when she was most frequently mounted. Kate solicited most frequently during the early part of her estrous period and was thus just the opposite from Blanche.
Individual differences among males.
There are many problems involved in determining whether males show preferences for certain females. For example, one must be able to show that all females are equally in estrus at the same time, and that the male's behavior is not just a reflection of the female's aversion to him. In the present study, it was not possible to control the circumstances in such a way that permits unequivocal statements about male preferences. However, marked differences in the response of individual males to all females were evident. Table 3 shows that John was an infrequent visitor of females regardless of their condition. In fact, he spent no more time with estrous females than Broadus spent with anestrous females (Table 4). This low

286 BURNEY J. LE BOEUF
response was not occasioned by the negative behavior of females for John was rarely threatened or attacked. Broadus was also rarely rejected by females, yet he mounted each female much sooner than John (Tables 5-8). Unlike John, Broadus rarely failed to visit a tethered female and he spent more time with them than did other males (Tables 3 and 4). Broadus was the most popular male in the sense that he was allowed to mount earlier and more often; he was more frequently solicited and he was least often rejected.
Clark and Ken were most often rejected. With two females (Spot and Blanche), both males were rejected approximately an equal percentage of time. However, Ken's scores were based on many more episodes. This disparity indicates something which was clear from observing the behavior. Ken was vigorous and persistent in his attempts to copulate despite being repeatedly rebuffed. In contrast, one or two snarls or snaps at the start of each test were sufficient to keep Clark away for the remainder of the period (see Table 4).
Dif ferences in the testing condition, tethered vs r o v i n g.
Tables 3 and 4 showed that the outcome of a test depended on whether the male or the female was tethered. Tables 5-8 provide additional data on this point. Females were mounted more frequently when they were tethered and the male was free. This is not surprising since some females did not go to some males when they had the choice. In four situations where a female was mounted by a nonpreferred male, it was always when the male was free and the female tethered (Tables 5-7: Peggy and Ken; Spot and Ken; Blanche and Clark, Eddie). On the other hand, females solicited more and rejected less when they were free and the males were tethered. The first day a female was mounted and the number of days that a male continued to mount were not affected by which partner was tethered.
INTRASEXUAL INTERACTIONS
Female-f emale behavior.
The collection of symmetric data from each female during anestrus, estrus and post-estrus was made difficult by the fact that the various conditions could not be held constant. To facilitate the following analysis, anestrous and post-estrous data were combiner and will be called non-estrous data.
Table ii shows the percentage of visits and test time spent by females in one condition with females in the same or opposite condition. The estrous female group visited other females most frequently and spent the greatest amount of time with them. They were, in turn, least often visited by others.

INTERINDIVIDUAL ASSOCIATIONS IN DOGS 287
TABLE I I
Percentage of days roving females visited tethered females and the percentage of total test time spent in the circle when the females were grouped according
to "reproductive" condition
Per cent Per cent Groups Visits Total Time
EF-NEF 74 I5
NEF-EF 42 6 NEF-NEF 55 8
Interactions between two females were infrequent as compared to those observed between heterosexual pairs or in tests involving two males. More than 3 episodes per visit day between two females was rare. Episodes were most often initiated by the roving female (74 %) rather than the restricted female (26 %o), regardless of either animal's reproductive condition. Most episodes involved brief investigation of the head or anogenital region. Mount- ing between females was also observed. In all cases, the roving female mounted the tethered female. This activity was most common when the nonestrous female was free and an estrous female was tethered. Estrous females rarely mounted tethered nonestrous females.
Females rarely responded negatively to mounts or investigation by other females except on one occasion. For two days, Peggy and Blanche were run to each other when the former was in estrus and the latter was in proestrus. Peggy courted Blanche vigorously, initiating a total of 79 episodes in these two days. Of these, 38 were investigatory, I4 involved mounting or attempts to mount, and 27 were terminated negatively by Blanche. The negative behavior displayed by Blanche to Peggy was as intense as that directed to males. Despite the threats and attacks, Peggy, like a sexually aggressive male, persisted in her attempts to investigate and mount. These data are not included in Tables i i and I2 because of the small number of observations.
Table 12 shows the response of each female roving to other tethered females. Kate showed the highest percentage of visits under each condition. All females visited most frequently in the EF-NEF situation except for Blanche, who visited most frequently when both she and the tethered female were in nonestrus (NEF-NEF). The percentage of test time that each female spent in the circle yields a similar picture.
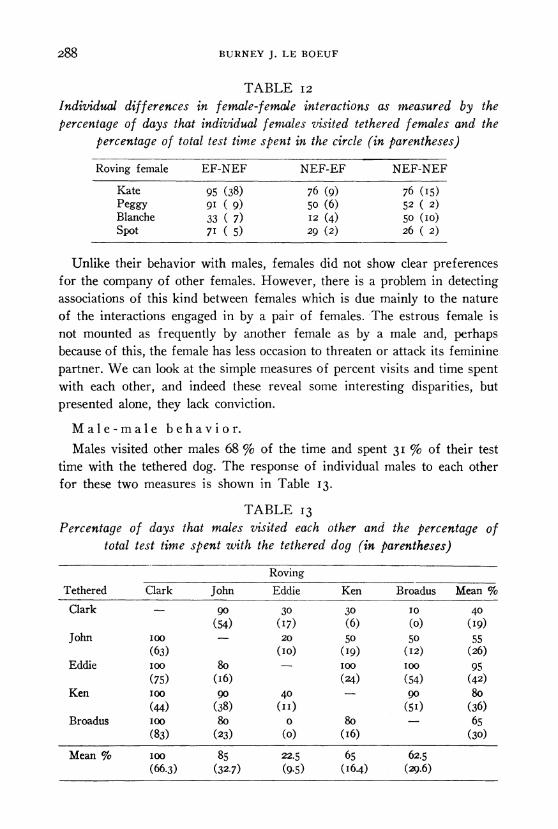
288 BURNEY J. LE BOEUF
TABLE I2
Individual differences in female-f emale interactions as measured by the percentage of days that individual fenales visited tethered females and the
percentage of total test time spent in the circle (in parentheses)
Roving female EF-NEF NEF-EF NEF-NEF
Kate 95 (38) 76 (9) 76 (I5) Peggy 91 ( 9) 5o (6) 52 ( 2) Blanche 33 ( 7) I2 (4) 50 (IO) Spot 71 ( 5) 29 (2) 26 ( 2)
Unlike their behavior with males, females did not show clear preferences for the company of other females. However, there is a problem in detecting associations of this kind between females which is due mainly to the nature of the interactions engaged in by a pair of females. The estrous female is not mounted as frequently by another female as by a male and, perhaps because of this, the female has less occasion to threaten or attack its feminine partner. We can look at the simple measures of percent visits and time spent with each other, and indeed these reveal some interesting disparities, but presented alone, they lack conviction.
Male-male behavior.
Males visited other males 68 % of the time and spent 3I % of their test time with the tethered dog. The response of individual males to each other for these two measures is shown in Table 13.
TABLE I3
Percentage of days that males visited each other and the percentage of total test time spent with the tethered dog (in parentheses)
Roving
Tethered Clark John Eddie Ken Broadus Mean %
Clark 90 30 30 10 40
(54) (I7) (6) (o) (I9) John IOO - 20 50 50 55
(63) (Io) (i9) (12) (26) Eddie IOO 80 100 100 95
(75) (I6) (24) (54) (42)
Ken IOO 90 40 90 8o (44) (38) (II) (50 (36)
Broadus ioo 8o 0 8o - 65 (83) (23) (o) (i6) (30)
Mean %o IOO 85 22.5 65 62.5 (66.3) (32.7) (9.5) (I64) (29.6)

INTERINDIVIDUAL ASSOCIATIONS IN DOGS 289
A comparison of the mean percentages in Table 13 indicates that the males who visited other males most frequently were themselves least fre- quently visited. For example, Clark made contact with all males every day that he was tested, but other males visited him only 40 % of the time. On the other hand, Eddie did not visit others very often (22.5 %) but was himself visited in virtually every test (95 %). This relationship is also apparent for the time measure.
A partial explanation for the discrepancies between cells in Table I3 is suggested by the behavior males displayed to each other. Interactions between males frequently involved elements of ritualized aggression which were unlike the negative behavior shown by estrous females to males. Aggressive en- counters between males resembled several descriptions in the literature (DAR-
WIN, I872; BUYTENDIJK, I936; LORENZ, I952, I953). One male, usually the free animal, approached the other, growling and walking stiffly with his tail held high, wagging briskly, and arched over his back. The other dog frequently responded in kind while the two stood nose to nose or walked together side by side. Encounters such as these were as brief as a few seconds or often as long as 2-3 minutes, and rarely terminated in a serious fight. Typically, the behavior subsided gradually with no dog "losing face". Mutual genital investigation was commonly observed at this time. If a male did not bristle when approached aggressively, he responded by retreating, cringing, or rolling over on his back while the other stood growling over him. Aggressive encounters such as these, where one dog clearly submitted to the other, were called unilateral. All other interactions involved head or anogenital investiga- tion. Male-male mounting was not observed during any of the tests.
TABLE I4 The proportion of visit days in which aggressive behavior was observed between males. The figures in parentheses refers to the number of episodes in which aggression was unilateral, i.e., one animal cringed or moved away
from the other; the arrow points to the "Winner" in the encounter
Roving Tethered Clark John Eddie Ken Broadus
Clark 8/9 0/3 0/3 o/I
John IOIO - 2/2 (2) 2/5 (2) 3/5
Eddie O/IO 2/8 (2) 5/I0 (2) IO/IO (3) t ~~~~t
Ken 6/io (6) 3/9 (3) 1/4 I/9 t
E3roadus 8/io (8) o/8 o/o o/8

290 BURNEY J. LE BOEUF
The data in Table I4, showing the number of days that aggressive be- havior was observed and the direction it took relative to the number of visit days for each combination of males, suggest that a dog's readiness to visit a tethered dog was related to the aggressive relationship between the two. Table I4 shows that Clark and John were often engaged in aggressive encounters with others from which they did not retreat. They were always victorious in unilateral encounters. Males rarely visited males to whom they had submitted in aggressive encounters. On the few occasional visits that occurred (e.g., to John), they approached in a submissive manner, cringing, and with their tails between their legs. Taken together, Clark and John visited the other three males 93% of the time while they were, in turn, visited only 32% of the time. The time the subjects spent with each other provides additional data on this point. One gets the impression that Clark and John visited others frequently because they had nothing to fear. The contrary seems to hold for the infrequent visits of the other three males. Interestingly, Clark and John almost always visited each other and nearly always engaged in mutual aggresision. However, neither of them was ever clearly victorious over the other. Among the other three males, Eddie never visited Broadus, but Broadus always visited Eddie and on every occasion they assumed an aggressive posture with each other. Eddie backed down in three of these encounters.
Some pairs, although in frequent contact, were conspicuous for the absence of aggressive interactions. Clark was never aggressive to Eddie nor was John ever aggressive to Broadus. Ken and Broadus were aggressive to each other only once in I7 encounters. This is reminiscent of the "intrasex loyalties" in wolves that GINSBURG (I965) writes about.
The general pattern and direction of aggressive behavior observed sub- stantiates previous casual observations made on the dominance relationship among these 5 males. Clark and John clearly dominated in all encounters with the other males but they "fought" to a draw in frequent encounters between themselves. The other males were rarely aggressive to each other and no single male dominated any other.
NORMATIVE COMPARISONS
Although this paper has dealt primarily with the individual differences stressed in preceding sections, some interesting normative information can be obtained from a comparison of the performance of various test combinations or groups on general measures of attraction such as the percentage of days the tethered animal was visited and the total test time the free animal spent with the restricted one. These data are presented in Table I5. As might be

INTERINDIVIDUAL ASSOCIATIONS IN DOGS 29I
expected, heterosexual groups containing an estrous female scored highest on both measures, with the M-EF group initiating more contacts and for a longer period of time than its counterpart group, EF-M. Viewed as a group, esitrous females visited nonestrous females nearly as often as they visited males. However, they spent less time with the former. The lowest groups on both measures were composed of roving nonestrous females.
TABLE I 5 Group comparisons for the measures, percentage of days the roving dog visited the tethered dog and the percentage of total time spent in the circle
Per cent Per cent Groups Visits Total Time
M-EF 93 49 EF-M 76 35 M-NEF 75 i6 EF-NEF 74 I5 M-M 68 3I
NEF-NEF 55 8 NEF-EF 42 6 NEF-M 35 II
DISCUSSION
The observation that some bitches in physiological estrus mate less readily with some males than others suggest that female dogs enjoy a higher level of sexual freedom from ovarian hormones than was previously thought. Carnivores in general have usually been classed with lower mammals such as rodents with respect to the complexity and modifiability of an individual's sexual behavior pattern. BEACH (1947) pointed out that sexual activation and performance in lower mammals depends heavily on gonadal hormones, whereas in primates hormonal control is not as rigid, and a considerable degree of sexual responsiveness and flexibility is permitted in the absence of secretions from the reproductive glands. For example, a close relationship between high levels of ovarian hormones and sexual receptivity is the rule in rats and guinea pigs, while in the rhesus monkey and the chimpanzee copula- tion is not always restricted to the female's fertile period. Under laboratory conditions, mating may occur at any stage in the female's reproductive cycle (TINKLEPAUGH, I933; YERKES, I936; YERKES & ELDER, I936; MICHAELS,
I965; KUEHN, I965), thus indicating that the female's sexual responsiveness is not completely dependent on ovarian hormones. The sexuality of the bitch
Behaviour XXIX I9

292 BURNEY J. LE BOEUF
is also influenced by non-hormonal factors but in d different way. The bitch copulates only when levels of estrogen are high, but under these conditions she may display preferences and aversions in her choice of males. Selective behavior in mating adds a degree of flexibility to the behavior of dogs which, as far as we know, is lacking in other lower mammals such as the rat. Therefore, the present evidence makes it appear that the dependence of sexual activity upon ovarian hormones in a carnivore, the dog, is less pronounced than in rodents but stronger than in primates.
Certain definitional problems are created by selective mating tendencies of the bitch. The commonly used dichotomy of classifying the bitch as "receptive" or "not receptive" does not hold because receptivity is not a general female state applicable to all males. The female may be receptive to one male and not to another. This state of affairs may create problems for the breeder who has to differentiate between a female that is not yet in estrus and one that is in estrus to other males but not to the one with which she is being tried.
The present study does not explain the formation of mating preferences, however, it appears that their establishment does not depend on prior copula- tory experience since some females exhibited preferences during their first estrous period (BEACH & LE BOEUF, in press). Preferences do not seem to be directly related to the dominance relationship among males, as the latter was defined in this study. One of the two most aggressive males, John, was readily acceptable to all females while the other, Clark, was not. The circum- stances in which the animals develop may be very important. In the present study, all Ss were reared together from weaning to the time observations began. It is possible that the experience gained during development is neces- sary for the establishment of associations which appear as mating preferences in adults.
It is worthwhile to stress the fact that the present observations were made on one group of dogs. The sample size was small, one breed was studied, and pairs of animals were tested briefly under restricted circumstances. These considerations limit the generalizations that can be made, particularly with regard to the day-to-day behavioral relevance of the observations.
SUMMARY
Interindividual behavior in a group of dogs reared together in a large field was studied to determine whether social and sexual interactions were more frequent in some pairs than in others. The experimental setting was designed to maximize the display of individual sympathies and antipathies.
The basic test provided a situation in which behavioral interactions between a pair of animals was contingent on one animal approaching the other. One subject was restricted

INTERINDIVIDUAL ASSOCIATIONS IN DOGS 293
by a tether to a circular area with a diameter of approximately I2 feet while the other subject, the roving animal, was released into the outdoor field for 5 minutes, the duration of each test. Measures of association and a description of behavior were made and then the roles of each subject were reversed. Test combinations consisted of paired females, paired males and heterosexual pairs. Females were observed before, during, and after they were in behavioral estrus.
Members of the opposite sex were most attracted to each other when the females were in estrus. However, all females were not equally attracted to all males. Three of four females showed preferences for some males over others. A preferred male enjoyed several advantages over nonpreferred males: (a) he was most frequently sought and solicited by the estrous female and she spent more time with him, (b) he was permitted to mount the female earlier in her estrous period, more frequently per day and, on more days, and (c) he was rarely prevented from investigating or mounting the female by being growled or barked at or by being bitten. Nonpreferred males were consistently rejected in this manner. One of the 5 males was readily accepted by all females. Otherwise, males preferred by one female were not uniformly preferred by the rest of them. In one case, a male preferred by one female was most frequently rejected by another. One female was equally receptive to all males.
The preferences and aversions revealed by these subjects were evident the first time the females came into estrus and in subsequent estrous periods, whether natural or induced by exogenous hormones. These results were interpreted as indicating that sexual receptivity in the bitch is not completely dependent on ovarian hormones.
Interactions between females were infrequent and brief as compared to those between two males or heterosexual pairs containing an estrous female. Strong individual associations between female pairs were not evident.
Male-male interactions commonly involved elements of ritualized aggression while female-female interactions did not. The response of one male to another depended in part on which one had previously retreated in an aggressive encounter with the other. The submissive male in these stereotyped encounters was frequently visited, but he himself, rarely approached others.
Some males preferred the company of some males to others. Members of a pair sympathetic to each other made frequent contacts of long duration and showed no aggression to each other.
REFERENCES
ASDELL, S. A. (I966). Dog Breeding. - Boston: Little, Brown and Company. BEACH, F. A. (I947). Evolutionary changes in the physiological control of mating behavior
in mammals. - Psychol. Rev. 54, 297-3I5. (I96I). Sex differences in the physiological bases of mating behavior in mammals.
- In: ALEXANDER SIMON (Ed.), The physiology of emotions. Illinois: Charles C. Thomas. & LE BOEUF, B. J. (in press). Mating preferences in female dogs. - Anim Behav.
BUYTENDIJK, F. J. J. (1936). The mind of the dog. - Boston: Houghton Mifflin. DARWIN, C. (I872). The expression of the emotions in man and animals. - London:
J. Murray. ENDERS, R. K. (I945). Induced changes in the breeding habits of foxes. - Sociometry
8, 53-55. FULLER, J. L. & DuBuIs, E. M. (I962). The behaviour of dogs. - In: E. S. E. HAFEZ
(Ed.), The behavior of domestic animals. Baltimore: Williams and Wilkins Company. GINSBURG, B. E. (I965). Coaction of genetical and nongenetical factors influencing sexual
behavior. - In: F. A. BEACH (Ed.), Sex and behavior. New York: John Wiley and Sons, Inc.

294 BURNEY J. LE BOEUF
GRAHAM, R. P. (ig6i). The mating and whelping of dogs (3rd ed.). - London: Poptilar Dogs.
HANCOCK, J. L. & ROWLANDS, I. W. (i949). The physiology of reproduction in the dog. - Vet. Rec. 6i, 77I-779.
HARROP, A. E. (I960). Reproduction in the dog. - Baltimore: Williams and Wilkins. KINSEY, A. C., POMEROY, W. B., MARTIN, C. E. & GEBHARD, P. H. (1953). Sexual
behavior in the human female. - Philadelphia: W. B. Saunders Company. KUEHN, R. E. (I965). Mating behavior in adult Macacca mulatta. - Paper presented at
the AAAS meetings in Berkeley, California, December. LE BOEUF, B. J. (I967). Heterosexual attraction in dogs. - Psychon. Sci. 7, 313-314. LORENZ, K. Z. (1952). King Solomon's Ring. - New York: Thomas Y. Crowell.
(953). Man meets dog. - Baltimore: Penguin. MICHAELS, R. P. (I965). Endocrine factors in sexual and grooming behavior in rhesus
monkeys. - Paper presented at the gth International Ethological Conference in Zurich, Switserland, September.
TINBERGEN, N. (I95I). The study of instinct. - London: Oxford Univ. Press. TINKELPAUGH, 0. L. (1933). Sex cycles and other cyclic phenomena in a chimpanzee
during adolescence, maturity and pregnancy. - J. Morph. 54, 52I-546. WHITNEY, L. F. (I947). How to breed dogs (revised). - New York: Orange Judd
Publishing Company. YERKES, R. M. (1936). A chimpanzee family. - J. genet. psychol. 48, 362-370.
& ELDER, J. H. (1936). Oestrus, receptivity, and mating in the chimpanzee. - Comp. Psychol. Monogr. I3, I-39.
RflSUMP:
La conduite individuelle dans un groupe de chiens eleves en meme temps dans un grand champ a ete etudiee pour determiner si les interactions, tant sociales que sexuelles, etaient plus frequentes chez certains couples que chez d'autres. L'atmosphere pour l'experience etait organisee pour deployer au maximum les sympathies et antipathies individuelles.
L'experience de base donnait une situation oiu la conduite entre un couple d'animaux etait contingente d'un animal approchant l'autre. Un sujet etait retenu par une chaine dans un endroit circulaire d'un diametre de 4 metres environ, pendant que l'animal libre etait lache dans un champ exterieur pour la duree de chaque experience qui est de cinq minutes. Des mesures d'association et une description de la conduite ont ete faites et ensuite le r6le de chaque sujet etait inverse. Des experiences combinees ont ete faites avec des paires de femelles, des paires de males, et des paires heterosexuelles. Les femelles etaient sous surveillance, avant, pendant et apres leur oestrouse behaviorale.
Les sujets de sexe oppose etaient attires l'un par l'autre lorsque les femelles etaient en oestrouse. Cependant, toutes les femelles n'etaient pas attirees de maniere egale par tous les males. Trois des quatre femelles ont montre une preference pour certains males plut6t que pour d'autres. Un male prefere avait plusieurs avantages sur les males delaisses: a) -il etait plus frequemment recherche et sollicite par la femelle oestrouse et elle
restait plus longuement avec lui. b) -il etait autorise a couvrir la femelle plus t6t dans sa periode oestrouse, plus
souvent dans une journee et pour plus de jours. c) il etait rarement empeche par des grognements ou aboiements ou par des coups
de crocs, d'etudier ou de couvrir la femelle. Les males delaisses etaient rejetes de cette maniere. Un des cinq males etait toujours accepte par toutes les femelles. D'autre part, les males preferes par une femelle n'etaient pas uniformement pre- feres par toutes. Dans un cas, un male prefere par une femelle etait plus fre-

INTERINDIVIDUAL ASSOCIATIONS IN DOGS 295
quemment rejete par une autre. Une des femelles etait receptive a tous les males de maniere egale.
Les preferences et aversions revelees par ces sujets etaient evidentes la premiere fois ou les femelles etaient en oestrouse et pendant les periodes oestrouses, naturelles ou provoques par des hormones exogenes. Ces resultats etaient interpretes comme indiquant que le comportement sexuel de la chienne n'est pas completement dependant des hormones ovariennes.
Les interactions entre femelles etaient breves et peu frequentes, comparees a celles de deux males ou de couples heterosexuels, comprenant une femelle oestrouse. De fortes associations entre paires de femelles n'etaient pas probantes.
Les interactions de male a male engageaient generalement des elements d'agression rituelle, alors que les interactions de femelle 'a femelle n'en engageaient pas. La reponse d'un male a un autre dependait en partie de celui que s'etait retire auparavant lors d'une rencontre aggressive avec l'autre. Le male soumis dans ces rencontres stereotypees etait souvent recherche, mais lui-menme approchait rarement les autres sujets.
Certains males preferaient la compagnie de certains males a d'autres. Les membres d'une mteme paire sympathisant entre eux avaient des contacts frequents de longue duree et ne montraient aucune agression l'un envers l'autre.




















