
Interaction of sodium and potassium with a cation-dependent adenosine triphosphatase system from rat...
10
BIOCHIMICA ET BIOPHYSICA ACTA 351 BBA 45353 INTERACTION OF SODIUM AND POTASSIUM WITH A CATION-DEPENDENT ADENOSINE TRIPHOSPHATASE SYSTEM FROM RAT BRAIN I~. AHMED AND J. D. JUDAH Division of Metabolic Research, Chicago Medical School, Chicago, Ill. (U.S.A.) and P. G. SCHOLEFIELD McGill University Cancer Research Unit, McGill University, Montreal (Canada) (Received December 2Ist, 1965) SUMMARY The kinetics of the hydrolysis of ATP by a rat-brain lipoprotein fraction have been investigated. The activity is greatly increased in the presence of Na + and K +, but reaches a maximum value and then suffers a diminution in the presence of an excess of either ion. Kinetic evidence indicates that these ions combine with the enzyme protein according to Michaelis-Menten kinetics, that 2 Na + are involved in the activation, and that the interaction of Na + and K + at each site is of a competitive nature. Inhibition of the (Na+-K+)-stimulated ATPase by ouabain, appears to result from a competition between ouabain and both K + and Na + while addition of oligo- mycin leads to kinetics which suggest competition of oligomycin with Na +. NH4+ may substitute for K + and, when present in excess, may also give rise to inhibitory effects. INTRODUCTION In view of the possible involvement of Na +- and K+-stimulated ATPase systems in the Na + transport mechanisms of cells 1-4, attempts have been made recently to analyze the kinetics of Na + and K + interaction with these enzyme systems 4-v. SKOU l investigated the effects of various Na+ and K + concentrations on the ATPase activity of rabbit brain and kidney preparations and noted that high concentrations of K + inhibited the stimulation by Na+. POST et al. ~ had made a similar observation in their study of erythrocyte membrane ATPase and more detailed analyses of the inter- actions between Na + and K + effects have recently been published by GREEN AND TAYLOR 5 and by SQUIRES 6. Studies, in which ~2P-labelled ATP was employed, strongly suggest that Na + is required for a phosphorylation step and that K + is required for the subsequent dephosphorylation3, 8-1°. GREEN AND TAYLOR 5 have examined their kinetic data in an attempt to distinguish between a (Na+-K+)-dependent ATPase requiring the simultaneous attachment of both ions to the enzyme protein for ATP- splitting activity on the one hand, and a sequential Na+-activated phosphorylation followed by a K+-activated dephosphorylation on the other. Their analysis suggested Biochim. Biophys. Acta, 12o (1966) 351-36o.
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Interaction of sodium and potassium with a cation-dependent adenosine triphosphatase system from rat...

BIOCHIMICA ET BIOPHYSICA ACTA 351
BBA 4 5 3 5 3
INTERACTION OF SODIUM AND POTASSIUM WITH A CATION-DEPENDENT
ADENOSINE TRIPHOSPHATASE SYSTEM FROM RAT BRAIN
I~. A H M E D AND J. D. J U D A H
Division of Metabolic Research, Chicago Medical School, Chicago, Ill. (U.S.A.)
and P. G. SCHOLEFIELD McGill University Cancer Research Unit, McGill University, Montreal (Canada)
(Received December 2Ist, 1965)
SUMMARY
The kinetics of the hydrolysis of ATP by a rat-brain lipoprotein fraction have been investigated. The activity is greatly increased in the presence of Na + and K +, but reaches a maximum value and then suffers a diminution in the presence of an excess of either ion. Kinetic evidence indicates that these ions combine with the enzyme protein according to Michaelis-Menten kinetics, that 2 Na + are involved in the activation, and that the interaction of Na + and K + at each site is of a competitive nature. Inhibition of the (Na+-K+)-stimulated ATPase by ouabain, appears to result from a competition between ouabain and both K + and Na + while addition of oligo- mycin leads to kinetics which suggest competition of oligomycin with Na +. NH4+ may substitute for K + and, when present in excess, may also give rise to inhibitory effects.
INTRODUCTION
In view of the possible involvement of Na +- and K+-stimulated ATPase systems in the Na + transport mechanisms of cells 1-4, attempts have been made recently to analyze the kinetics of Na + and K + interaction with these enzyme systems 4-v. SKOU l investigated the effects of various Na+ and K + concentrations on the ATPase activity of rabbit brain and kidney preparations and noted that high concentrations of K + inhibited the stimulation by Na+. POST et al. ~ had made a similar observation in their study of erythrocyte membrane ATPase and more detailed analyses of the inter- actions between Na + and K + effects have recently been published by GREEN AND TAYLOR 5 and by SQUIRES 6. Studies, in which ~2P-labelled ATP was employed, strongly suggest that Na + is required for a phosphorylation step and that K + is required for the subsequent dephosphorylation3, 8-1°. GREEN AND TAYLOR 5 have examined their kinetic data in an attempt to distinguish between a (Na+-K+)-dependent ATPase requiring the simultaneous attachment of both ions to the enzyme protein for ATP- splitting activity on the one hand, and a sequential Na+-activated phosphorylation followed by a K+-activated dephosphorylation on the other. Their analysis suggested
Biochim. Biophys. Acta, 12o (1966) 351-36o.

352 K. AHMED, J. D. JUDAH, P. G. SCHOLEFIELD
t h a t it was not possible, on the basis of thei r da ta , to exclude one poss ib i l i ty in favour of the other. However , SQUIRES 6, from his analysis, in which the Hill equat ion was employed , suggested the possible invo lvement of two or more coopera t ive ly inter- ac t ing b inding sites for each ligand.
In a previous publ ica t ion the p repa ra t ion of a r a t -b ra in l ipoprote in fract ion conta in ing high levels of the ca t ion -ac t iva ted ATPase sys tem was descr ibed 1'. In the present paper the results of an a t t e m p t to s t u d y the mechanism of in te rac t ion of Na+ and K + wi th the ca t ion-dependen t ATPase of this p repa ra t ion are described. The na tu re of the inhibi t ions of the ca t ion-s t imula ted ATPase b y ouaba in and oligo- myc in has also been inves t igated.
MATERIALS AND METHODS
Prepa ra t i on of T r i s - A T P and recrys ta l l iza t ion of the var ious sal ts used were as descr ibed before ~. The ATPase sys tem was the ra t -b ra in l ipoprote in descr ibed pre- viouslyg, 11.
The incuba t ion med ium for ATPase assay was 30 mM Tris-HC1 buffer (pH 7.4) ; 5 mM T r i s - A T P ; 5 mM MgSO4; KC1 and NaC1 concent ra t ions were as ind ica ted in the text . The final volume was 2 ml and the incubat ion t e m p e r a t u r e was 38°. Ra t - b ra in l ipoprote in was suspended in a med ium consist ing of 0.25 M manni to l , 20 mM Tris-HC1 (pH 7.4), and I mM E D T A (Tris salt) (pH 7-4)- The react ion was s t a r t ed b y adding 0.2 ml (53/xg protein) of this suspension and s topped b y adding 2 ml cold IO % (w/v) t r ichloroacet ie acid. The t ime of incubat ions was va r ied according to the design of ind iv idua l exper iments . The amoun t of Pi p roduced in each exper iment was of the order of I ~mole. The t ime course of ATPase ac t i v i t y was l inear under the condi t ions of the exper iments . The results are expressed as/~moles PI formed per mg prote in per min. AP i refers to the ATPase ac t i v i t y in the presence of Mg 2+, Na + and K + m i n u s t ha t in presence of Mg 2+ alone. In a sys tem conta in ing I iO mM Na + and IO mM K + the ra t io of the ATPase act iv i t ies (Mg 2+, Na + and K+/Mg 2+) was 8.
Pi was e s t ima ted b y the me thod of FISKE AND SUBBAROW 12. Pro te in was de t e rmined b y the m e t h o d of LowRY et al. 13.
RESULTS
The effects of increasing concent ra t ions of K + on the ATPase a c t i v i t y ob ta ined w i th this p r epa ra t i on at var ious levels of Na + are shown in Fig. I. The effects of in- c reas ing concen t ra t ions of Na + on the ac t iv i t ies observed at var ious levels of K + are shown in Fig. 2. I t is appa ren t t ha t in this p repa ra t ion increase in the concent ra t ion of K + increases the ATPase ac t iv i ty and t ha t subsequen t ly an inh ib i to ry effect is ob ta ined . Similar ly, add i t ion of N a + leads to an increase in the ac t i v i t y bu t on in- creasing the N a + concent ra t ion a m a x i m u m veloci ty is ob ta ined beyond which a fur ther increase in N a + concent ra t ion leads to an inhibi t ion of the enzyme act iv i ty . The inhib i t ion has usual ly been assumed to be compet i t ive in na tu r e bu t posi t ive evidence on this poin t seems to be lacking. Eqn. I was ob ta ined b y GREEN AND TAYLOR 5 from one of thei r analyses in which i t was assumed tha t sepa ra te b inding sites a re involved and t ha t compet i t ion between bo th ions m a y occur at bo th sites.
J?iochim. Biophys. Acta, i2o (1966) 351 360

B R A I N L I P O P R O T E I N ATPase A C T I V I T Y 353
E 1.5o
~- 0.75 a_- <3
o, - - ,~o ," ,o~0 m mola r K +
Fig . i . T h e e f f ec t s of v a r i o u s K + c o n c e n t r a t i o n s in t h e p r e s e n c e o f d i f f e r e n t N a + c o n c e n t r a t i o n s o n t h e A T P a s e a c t i v i t y . T h e c o n d i t i o n s o f t h e e x p e r i m e n t w e r e as d e s c r i b e d u n d e r MATERIALS AND METHODS, I n c u b a t i o n t i m e , 15 r a i n a t 38° . C u r v e i , 5 m M N a + ; C u r v e 2, 20 m M N a + ; C u r v e 3, 5o m M N a +.
i.o
~ 3
2 0.75
_E E
£ o..
0 ,50
::L
<] 0.25
w I
0.3 1.0 3.0 I0.0
m mo lo r N o "
F i g . 2. T h e e f fec t s of v a r i o u s N a + c o n c e n t r a t i o n s in t h e p r e s e n c e of d i f f e r e n t K + c o n c e n t r a t i o n s o11 t h e A T P a s e a c t i v i t y . F o r c o n c e n t r a t i o n s of N a + u p t o 2 r a M , t h e i n c u b a t i o n t i m e w a s 9 0 r a i n a n d f o r t h e r e s t o f t h e e x p e r i m e n t 45 r a in . T e m p e r a t u r e : 38 ° t h r o u g h o u t . C u r v e I, o . i m M K + ; C u r v e 2, 0. 3 m M K + ; C u r v e 3, i . o m M K +.
KEI v ( i )
In this equation a and b represent the concentrations of Na+ and K + respectively, K 1 is the Km value for Na +, K 2 is the Kl value for K + at this site, K 3 is the Km value
Biochim. Biophys. Acta, 12o (1966) 3 5 1 - 3 6 o

354 K. AHMED, J. D. JUDAH, P. G. SCHOLEFIELD
for K +, Ka is t he K , va lue for N a + at th is si te a n d K is t h e ra te cons tan t . I n conf i rma- t ion of t he resul ts r e p o r t e d by GREEN AND TAYLOR 5, a t t e m p t s to a p p l y Eqn . I to
famil ies of curves such as those shown in Figs. I and 2 p r o v e d to be unsa t i s fac to ry .
I t was the re fo re dec ided to a t t e m p t to ana lyse s epa ra t e ly t h e fo l lowing four d i s t inc t effects : (i) t he s t i m u l a t i o n b y Na+, (ii) t he inh ib i t ion b y N a +, (iii) t he s t i m u l a t i o n
b y K +, a n d (iv) t he inh ib i t ion b y K +. P r e l i m i n a r y e x p e r i m e n t s i nd i ca t ed t h a t t he s t i m u l a t i o n b y K + was mos t suscep t ib le to k ine t ic analysis . T y p i c a l resul ts o b t a i n e d in an e x p e r i m e n t des igned to i l lus t r a t e th is effect are shown in Tab l e I. A t c o n s t a n t
levels of A T P , Mg 2+ a n d N a + t h e response to K + is t h a t which w o u l d be p r e d i c t e d f r o m the M i c h a e l i s - M e n t e n 14 concep t . U n d e r t he cond i t ions e m p l o y e d the a p p a r e n t
Michael is c o n s t a n t for K + was o.195 m M and the m a x i m u m ve loc i ty , if no inh ib i t ion
b y h igh levels of K + were to occur , was 1 . 3 8 / , m o l e s per m g p ro te in per min. In t he e x p e r i m e n t r e p o r t e d in th is table , t h e effect of ouaba in a t c o n c e n t r a t i o n s of o.o5 and
o . I / * M was i n v e s t i g a t e d and t h e resul t s o b t a i n e d are also p r e s e n t e d in Tab l e I. T h e y
TABLE I
T H E E F F E C T S O F K + A N D O U A B A I N ON B R A I N C A T I O N - A C T I V A T E D A T P a s e A C T I V I T Y
Standard incubation conditions described under MATERIALS AND METHODS were employed. The incubation was carried out for 30 rain, the concentration of Na + being maintained at io mM throughout.
K + Ouabain Velocity (mM) (/~M) (AP,, #moles~rag protein
per rain)
Observed Theoretical*
o.15 - - o.556 o.598 o.2 - - o.721 o.695 o. 3 - - o.839 o.833 0.6 - - I.O44 I.O36 o.15 0.05 0.399 o.416 0.2 0.05 0.484 0.504 o. 3 0.05 o.661 0.639 0.6 0.05 0.789 o.871 o.15 o.I o.319 o.319 0.2 o.I 0.379 0-395 o. 3 o.I 0.529 o.518 0.6 o.I 0.763 0.753
* Calculated assuming that under the experimental conditions the maximum velocity would be 1.375/*moles per mg protein per min, the Km for K + is o.195 mM and that ouabain is a com- petitive inhibitor of K+ activation with a K~ value of 0.065 #M.
a re in good a g r e e m e n t w i t h those p r e d i c t e d a s suming t h a t ouaba in is a c o m p e t i t i v e i nh ib i t o r of K + in th is s y s t e m and t h a t i t has a Michael is c o n s t a n t of 0.065/*M. T h e close co r re l a t ion b e t w e e n t h e obse rved and t heo re t i c a l resul t s seems to j u s t i f y t he a s s u m p t i o n t h a t ouaba in is, in fact , a c o m p e t i t i v e inh ib i to r of t he a c t i v a t i o n by K + of th is A T P a s e sys tem.
I f t h e s t i m u l a t i o n b y K + a n d t h e inh ib i t ion b y N a + a c t u a l l y occur a t t h e s a m e s i te t h r o u g h a c o m p e t i t i v e i n t e r a c t i o n a t th is site, t h e n t h e inh ib i t ion b y N a + should also be p r e d i c t e d in t e r m s of t h e M i c h a e l i s - M e n t e n concep t . I n Fig. 3, t h e rec iproca l
Biochim. Biophys. Aeta, 12o (1966) 351-36o

BRAIN LIPOPROTEIN A T P a s e ACTIVITY 355
of the ATPase activity is plotted against the concentration of Na + used to inhibit the reaction according to the method of DIXON ~5. It is apparent that the expected linear relationship is obtained. The effect of o.I/~M ouabain on the ATPase activity
I 0 0
80 ~ 2
V-I
4.0 ~~1~e i
t I I 0 40 80 120
m m o l o r N o +
Fig. 3. The effects of inhibi tory concentrat ions of 1%a + and of ouabain on the ATPaso activity. Incuba t ion time, 3 ° min at 38°. Velocity is expressed as APi (/*moles per mg protein per rain). Curve I, no addit ions; Curve 2, o.I ,uM ouabain added.
was examined and the results obtained also appear in this figure. If ouabain and high concentrations of Na + act simultaneously as competitive inhibitors of activation by K + then the effect of ouabain should be capable of interpretation in terms of its function as a second competitive inhibitor. In a plot by the method of DIXON 15, the presence of a second competitive inhibitor should lead to a change in the intercept and a change in the slope but the relationship between the reciprocal of the velocity and the concentration of inhibitor should remain a linear one. All these points seem
0.35
0.30
0.25
0.20 V
0.15
0.10
0.05
0
A B 6 . ~
o
0 q
y . i
LO 2.O
~ 4 5
2 3 3.s ,/2_,
i 1.0 2.0 0 J
[No']
C
2
I~0 20 I
[~o-] Fig. 4. Kinetic analysis of the effect of various Na + concentrat ions and of ouabain and oligomycin on the ATPase activity. Incuba t ion time, 12o min at 38°. Curve i, no addit ions; Curve 2, 0. 4/~M ouabain added; Curve 3, 2/~g oligomycin per vessel (2 ml). A, plot of v (API, /~moles per mg protein per rain) versus concentrat ion of Na +. B, plot of i/v versus I / [Na+]. C, plot of I/%/vversus I / [Na+]. The concentrat ion of K+ was mainta ined at I mM throughout .
Biochim. Biophys. Acta, 12o (i966) 351-36o

356 K. AHMED, J. D. JUDAH, P. G. SCHOLEFIELD
to be verified in the results shown in Fig. 3. Oligomycin (I /~g/2 ml) inhibited the ATPase reaction but competit ion with K + could not be demonstrated.
When all other conditions were kept constant and the concentration of Na ÷ was gradually increased, the results shown in Fig. 4A were obtained. Increasing concentrations of Na + led to an increase in velocity and the gradient of the resulting curve continually increased. This sigmoid relationship indicates tha t simple Miehaelis- Menten kinetics cannot accurately describe the effect of Na + on the ATPase activity, a conclusion which is made more apparent from Fig. 4 B where the kinetic data are plotted by the method of LINEWEAVER AND BURK 1~. The relationship is obviously not the usual linear one. I t should also be noted tha t both ouabain and oligomycin were added to this system, tha t both produced inhibitory effects and tha t the resulting graphical relationships were also non-linear. However, if the square root of the reciprocal of the velocity is plotted against the reciprocal of the Na + concentration, then the straight lines shown in Fig. 4C are obtained, not only in the absence of in- hibitors but also in the presence of ouabain or of oligomycin. This result immediately suggests tha t the stimulation by Na + involves the interaction of not I but 2 Na~ with the enzyme protein. By analogy with the results presented in Table I and Fig. 3, the inhibitory effects of K + at a constant concentrat ion of Na + were therefore in- vestigated to determine whether these inhibitory effects could also be shown to depend on the interaction of two ions with the enzyme protein. The results presented
I
2
• ~ 0.50 o
E
E ~. o . z 5 fi-
0.75
t 5 2tO m mo lo r K ÷
Fig. 5. The effects of i nh ib i to ry concen t ra t ions of K+ and of ouaba in on ATPase ac t iv i ty . Incuba- t ion t ime, 20 rain a t 38°. 5 mM Na + was p resen t th roughou t . Curve i, no add i t ions ; Curve 2, 0. 5 #M ouaba in added.
in Fig. 5 show the relationship between the actual velocity observed and the con- centrat ion of K + present, in the presence and absence of 0.5/~M ouabain. All a t tempts to demonstrate a linear relationship between the concentrat ion of inhibitor and the reciprocal of the velocity or the square root of the reciprocal of the velocity were unsuccessful. However, it should be mentioned tha t when the square root of the reciprocal of the velocity was plotted against the concentrat ion of K +, the results did tend to become linear. The reason for this deviation will be presented in the DISCUSSION.
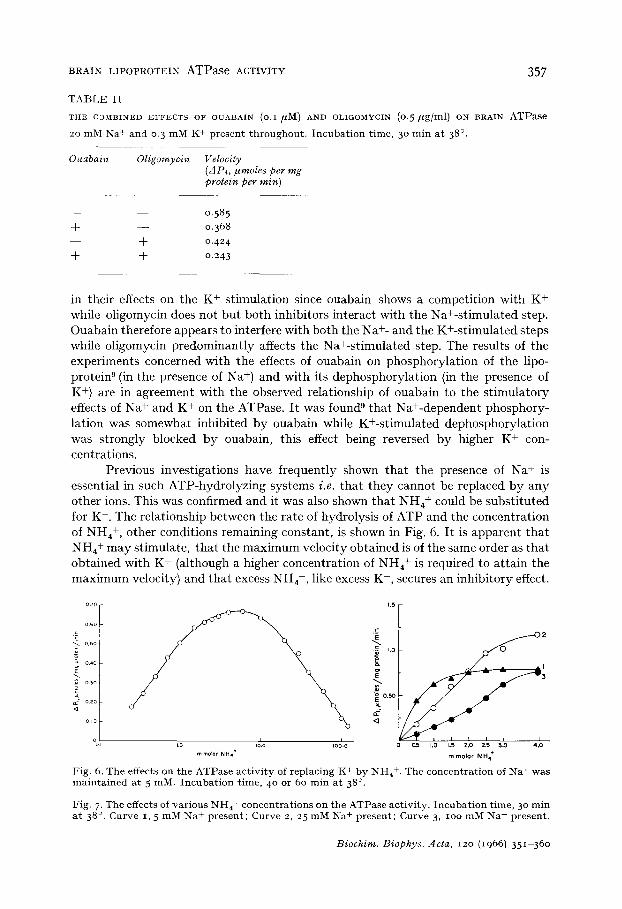
The combined effects of ouabain (o.I/zM) and oligomycin (0.5/~g/ml) on the brain ATPase preparat ion were also investigated and the results obtained are pre- sented in Table II . They demonstrate tha t ouabain and oligomycin secure inhibitory effects in the presence qf each other. I t appears tha t ouabain and oligomycin differ
Biochim. Biophys. Acta, 12o (1966) 351-36o
B R A I N L I P O P R O T E I N A T P a s e ACTIVITY 357
T A B L E I I
T H E C O M B I N E D E F F E C T S OF O U A B A I N (O.I /~M) AND OLIGOMYCIN (0. 5 # g / m l ) ON B R A I N A T P a s e
20 m M N a + a n d 0. 3 m M K + p r e s e n t t h r o u g h o u t . I n c u b a t i o n t i m e , 3 ° m i n a t 38° .
O~*abain Oligomycin Velocity (AP,, I~moles per mg protein per rain)
__ __ 0 . 5 8 5
+ - - 0 . 3 6 8
- - + 0 . 4 2 4
+ + 0 . 2 4 3
in their effects on the K + stimulation since ouabain shows a competition with K + while oligomycin does not but both inhibitors interact with the Na+-stimulated step. Ouabain therefore appears to interfere with both the Na +- and the K+-stimulated steps while oligomycin predominantly affects the Na+-stimulated step. The results of the experiments concerned with the effects of ouabain on phosphorylation of the lipo- protein s (in the presence of Na +) and with its dephosphorylation (in the presence of K +) are in agreement with the observed relationship of ouabain to the stimulatory effects of Na + and K + on the ATPase. It was found s that Na+-dependent phosphory- lation was somewhat inhibited by ouabain while K+-stimulated dephosphorylation was strongly blocked by ouabain, this effect being reversed by higher K + con- centrations.
Previous investigations have frequently shown that the presence of Na + is essential in such ATP-hydrolyzing systems i .e. that they cannot be replaced by any other ions. This was confirmed and it was also shown that NH4+ could be substituted for K +. The relationship between the rate of hydrolysis of ATP and the concentration of NH4+, other conditions remaining constant, is shown in Fig. 6. It is apparent that NH4+ may stimulate, that the maximum velocity obtained is of the same order as that obtained with K + (although a higher concentration of NH4+ is required to attain the maximum velocity) and that excess NH~ +, like excess K--, secures an inhibitory effect.
0.60
0.50
0.40
~ o.3o
~-- 0.20 <]
%.,
E
~ 0..~
d_-
,.' ,o' . . . . ".o m molor NH4¢
0.5 1.0 1.5 2.0 ?-.~ 3.0 4.0
m mo lo r NH~
F i g . 6. T h e e f fec t s o n t h e A T P a s e a c t i v i t y of r e p l a c i n g K + b y N H 4 + . T h e c o n c e n t r a t i o n o f N a + w a s m a i n t a i n e d a t 5 m M . I n c u b a t i o n t i m e , 4 ° o r 60 m i n a t 38° .
F i g . 7. T h e e f f ec t s of v a r i o u s N H 4 + c o n c e n t r a t i o n s o n t h e A T P a s e a c t i v i t y . I n c u b a t i o n t i m e , 3 ° r a i n a t 38° . C u r v e i , 5 m M N a + p r e s e n t ; C u r v e 2, 25 m M N a + p r e s e n t ; C u r v e 3, i o o m M N a + p r e s e n t .
Biochim. Biophys. Acta, 12o (1966) 3 5 1 - 3 6 o

358 K. AHMED, J. D. JUDAH, P. G. SCHOLEFIELD
The results presented in Fig. 7 c lear ly indicate tha t a complex in te rac t ion also occurs between NH4+, Na + and the enzyme.
DISCUSSION
Two different types of explana t ions have been offered to account for the s t imula- t ion of ATPase ac t iv i ty b y Na + and K+. They are : (i) t ha t there are separa te b inding sites, one p r imar i ly involving Na + and the o ther p r imar i ly involving K + (ref. I ) ; (ii) t ha t there is bu t one b inding site which is occupied b y Na + dur ing phosphory la t ion of the enzyme and la ter b y K + dur ing the ensuing dephosphorylat ionS, 9. I t is also possible t ha t while the occupancy of the Na + site b y a Na + ion is the i m p o r t a n t factor in phosphory la t ion , the presence or absence of K + at ano ther site has l i t t le effect on the phosphory la t ion process. S imi lar ly the presence of Na + at the Na+-ac t iva t ion site m a y have l i t t le effect on the K+-s t imula ted dephosphory la t ion .
The d a t a p resen ted in Figs. I and 2 clear ly suppor t the content ion t h a t the overal l ac t iv i ty of the bra in ca t ion -ac t iva ted l ipoprote in ATPase a c t i v i t y is de- penden t not only on the presence of Na + and K + bu t also upon an appropr i a t e balance between these two ions. This lends suppor t to the suggestion t ha t there exists a site on the enzyme at which each ion mus t specifically become a t t ached and at which the o ther ion m a y interfere wi th this a t t achmen t . Fur the rmore , all the results ob ta ined in the present and in previous work indicate t ha t the na ture of the in te rac t ion be tween Na + and K + at each of these enzyme sites is a compet i t ive one. However , it is not easy to ob ta in definit ive proof t ha t this in terac t ion is compet i t ive . Var ia t ion in the concent ra t ion of e i ther ion would be expec ted to influence bo th sites, one a t t ach- ment leading to an increased enzyme a c t i v i t y and the o ther a t t a c h m e n t leading to the d i sp lacement of an essential ion and thus to a decrease in enzyme act iv i ty . The s imul taneous opera t ion of all these var ious c i rcumstances as represented b y Eqn. I renders the kinet ic approach ra the r difficult. However , if the s t imu la to ry site has a very low Km value for K +, then one m a y safely inves t iga te the s t imu la to ry effect of K + by using only low concentrat ions . Conversely, one m a y inves t iga te the inh ib i to ry effect by rou t ine ly using only high concent ra t ions of K + with the as- surance tha t under these condi t ions the site of K + ac t iva t ion has been a lmost com- p le te ly s a tu r a t ed with K +. I t should be po in ted out, however, t ha t this approach is only feasible if app rop r i a t e concent ra t ions of Na + are present because there m a y be in te rac t ion of Na + with the site a t which K + combina t ion leads to enzyme ac t iv i ty . In pract ice it has been possible to find app rop r i a t e exper imenta l condi t ions (see Table I) where the addi t ion of low concent ra t ions of K + leads to a s t imula t ion of enzyme ac t iv i ty and no evidence of inh ib i to ry effects by K + is apparen t . The assign- ment of an appa ren t value for the m a x i m u m veloci ty and of an appa ren t Michaelis cons tan t for K + should therefore enable one to predict the veloci ty of ATPase ac t i v i t y at a n y given concent ra t ion of K +. The agreement between the observed and the theore t ica l value ca lcula ted from such an equat ion, as seen in Table I, leaves no doub t t ha t the in terac t ion of K + with the enzyme takes place according to the Michael is -Menten concept.
If the concent ra t ion of K + is kep t cons tan t a t a level which secures only s t imu- l a to ry effects, and the concent ra t ion of Na + is fur ther increased, the resul t ing s i tua t ion should be one in which Na + not only sa tu ra tes the site at which Na + ac t iva t ion occurs
Biochim. Biophys. Acta, 12o (1966) 351-36o

BRAIN LIPOPROTEIN ATPase ACTIVITY 359
bu t also begins to compete effect ively for the site a t which K ÷ ac t iva t ion occurs. Fo r this reason, a plot of the reciprocal of the veloci ty of ATPase ac t iv i ty agains t the N a + concent ra t ion according to the m e t h o d of DIXON 15 should yield a l inear re la t ion- ship such as is seen in Fig. 3-
Increase in the Na + concent ra t ion at a cons tan t concent ra t ion of K + leads in i t ia l ly to a s t imula t ion b y Na +, as seen in Fig. 4 A. However , i t should be no ted t h a t even at concent ra t ions of Na + as low as 0.5 mM the re la t ionship be tween the ATPase ac t i v i t y and Na + concent ra t ion is represented b y a curve of increasing gradient . The LINEWEAVER--BuRK plot of these resul ts fai led to produce a s t ra igh t line, as seen in Fig. 4 B. An a t t e m p t was therefore made to inves t iga te whether 2 Na ÷ might be involved in the ac t iva t ion process. The corresponding kinet ic equat ion is:
KEt ~, = (2)
In the der iva t ion of this equat ion it is assumed tha t (i) N a + has the same affinity for each of the two sites; (ii) the inh ib i to ry effects resul t ing from in terac t ion of K + at e i ther of these two sites are exac t ly the same; (iii) Na + or K + a t t a c h themselves to any of the three sites qui te i ndependen t ly of the presence or absence of ions a t t a c h e d to a n y of the o ther si tes; and t ha t (iv) the species of the enzyme which is capable of b reak ing down the ATP is t ha t which has a t t a c h e d to it 2 Na + and I K +, each at an a p p r o p r i a t e site. A t an app rop r i a t e concent ra t ion of K + the second t e rm in the denomina to r of t i le r igh t -hand side of Eqn. 2 might become cons tan t and a l inear re la t ionship should ob ta in if the square root of the veloci ty of ATPase ac t iv i ty is employed in a LINEWEAVER--BURK analysis. The va l id i ty of this app roach is seen in Fig. 4 C.
There seems to be l i t t le doub t t ha t this p repa ra t ion of b ra in ATPase is a c t i va t ed as a resul t of the combina t ion of the enzyme with I K + and 2 Na +. The involvement of two sites for one ion or for one molecular species in t r anspor t phenomena is not unusual , hav ing been suggested prev ious ly in connect ion with glycerol t r anspo r t 17, sugar t r anspor t 1~, 19, amino acid t r anspor t 2°, 21, and ion t r anspor t 22.
The compet i t ive na tu re of the in terac t ion of K + and ouaba in wi th the enzyme is c lear ly shown in the results quoted in Table I. On the o ther hand, the results p resen ted in Fig. 4C show equal ly c lear ly t ha t the na tu re of the in terac t ion of ouaba in a n d N a + wi th the enzyme also appears to be compet i t ive . In considering this anoma- lous resul t two factors should be borne in mind. Fi rs t , the concentra t ions of Na ÷ used in the exper iments quoted in Fig. 4C are ex t r eme ly low, the m a x i m u m being 2 mM. Under the incuba t ion condi t ions usua l ly employed for examin ing (Na+-K+) - s t imu la t ed ATPase ac t iv i ty , the Na + concent ra t ion is usual ly IOO mM, at which level ouaba in should have ve ry l i t t le effect on the in terac t ion of the enzyme with N a +. The observed effect of ouaba in on in tac t cell p repara t ions m a y therefore be inter- p re ted p r imar i ly in t e rms of the "compe t i t i ve" na tu re of the in te rac t ion between ouaba in and K +. Secondly, it should be borne in mind t ha t a resul t which is kinet i - cal ly ident ica l wi th compet i t ive inhibi t ion does not necessa r i lymean t ha t the inhib i tor competes wi th the subs t ra te for a t t a c h m e n t to one specific site on t i le enzyme surface. The kinet ic equat ions only imply t ha t the inhibi tor and the subs t r a t e molecules
Biochim. Biophys. Acta, 12o (1966) 351-36o

360 K. AHMED, J. D. JUDAH, P. G. SCHOLEFIELD
cannot be a t t a c h e d to the enzyme at the same time. If ouaba in and K +, for example , occupied ad jacen t sites and if there was an extensive overlap, then i t is possible to envisage condi t ions where the a t t a c h m e n t of K + would p reven t the s imul taneous a t t a c h m e n t of ouaba in and where the a t t a c h m e n t of ouaba in would prevent the s imul taneous a t t a c h m e n t of K +. The kinet ic analysis of this t ype of in te rac t ion would be indis t inguishable from a s i tua t ion in which ouabain and K + compe ted with each o ther for one single site. I t is therefore concluded t ha t Na +, K +, and ouaba in become a t t a c h e d to the ATPase prote in at sites which are ve ry close to each other, t ha t Na+ m a y have an affini ty for the K + site and K + an affinity for the Na + site, t ha t the enzyme requires 2 Na + and I K + to become a t t a c h e d in the same ne ighbourhood on the enzyme surface for ca t a ly t i c ac t iv i ty to be manifested, t ha t when ouaba in becomes a t t a c h e d it p revents the a t t a c h m e n t of ei ther Na + or K + and conversely t ha t when Na + or K + are a t t a c h e d in this v ic in i ty t hey p reven t the a t t a c h m e n t of ouabain. Such a s i tua t ion would be in keeping wi th all the kinet ic resul ts which have been observed in the present inves t igat ion.
ACKNOWLEDGEMENTS
I t is a pleasure for one of us (P.G.S.) to t h a n k the Chicago Medical School for an a p p o i n t m e n t as a Visi t ing Professor which made possible this coopera t ion wi th the Division of Metabol ic Research of the Chicago Medical School. I t is also a pleasure to t h a n k the Na t iona l Cancer In s t i t u t e of Canada for suppor t of the work in Mon- t rea l and to t h a n k the U.S. Publ ic Hea l t h Service (Grant No. AM 7226), the Life Insurance Medical Research Fund , and the Otho S.A. Sprague Memorial F o u n d a t i o n of Chicago for suppor t of the work carr ied out in Chicago.
R E F E R E N C E S
I J. C. SKOU, Biochim. Biophys. Acta, 58 (1962) 314 . 2 J. C. SKOU, Physiol. Rev., 45 (1965) 596. 3 J. D. JUDAH AND K. AHMED, Biol. Rev. Cambridge Phil. Soc., 39 (1964) 16o. 4 R. L. POST, C. R. MERRIT, C. R. KINSOLVlNG AND C. D. ALBRIGHT, J. Biol. Chem., 235 (196o)
1796. 5 A. L. GREEN AND C. t3. TAYLOR, Biochem. Biophys. Res. Commun., 14 (1964) 118. 6 R. F. SQUIRES, Biochern. Biophys. Res. Cornmun., 19 (1965) 27. 7 H. J. SCHATZMANN. Biochirn. Biophys, Acta, 94 (1965) 89. 8 J. S. CHARNOCK AND R. L. POST, Nature, 199 (1963) 91o. 9 K. AHMED AND J. D. JUDAH, Biochirn. Biophys. Acta, lO 4 (1965) 112.
IO R. GIBBS, P. M. RODDY AND E. TITUS, J. Biol. Chem., 24o (1965) 2181. i i K. AHMED AND J. D. JUDAH, Bioehim. Biophys. Acta, 93 (1964) 6o3. 12 C. H. FISKE AND Y. SUBBAROW, J. Biol. Chem., 66 (1925) 375. 13 O. H. LowRY, N. J. ROSHNBROUGH, A. L. FARR AND R. J. RANDALL, J. Biol. Chem., 193 (195 I)
265 . 14 L. MICHAHLIS AND M. [. MENTEN, Biochem. Z., 49 (1913) 333. 15 M. DIXON, Biochem. J., 55 (195o) 17o. 16 H. LINEWEAVER AND D. J. BURK, J. Am. Chem. Soc., 56 (I934) 658. 17 W. D. STEIN, Nature, 191 (1961) 352. 18 W. D. STEIN, Biochirn. Biophys. Acta, 59 (1962) 66. 19 A. I~OTYK AND W. WILBRANDT, Helv. Physiol. Pharmacol. Acta, 21 (1963) C33. 20 J. A. JACQUEZ, Biochim. Biophys. Acta, 79 (1964) 318- 21 N. BI~OIN AND P. G. SCHOLEFIELD, Biochirn. Biophys. Acta, lO 4 (1965) 566. 22 G. VIDAVHR, Biochemistry, 3 (1964) 662.
Biochim. Biophys. Acta, 12o (1966) 351-36o




















