
Insect pest management by the horse shoe bats (Rhinolophus species) in the forest ecosystem of...
19
SIRJ-BES Volume 1 Issue 5 (2014) ISSN 2348 - 5787 www.scrutinyjournals.com Scrutiny International Research Journal of Biological and Environmental Science (SIRJ-BES) Insect pest management by the horse shoe bats (Rhinolophus species) in the forest ecosystem of Kalakad Mundanthurai Tiger Reserve, India Selva Ponmalar S 1 and Juliet Vanitharani 2 Bat Research Laboratory, Zoology Department and Research Centre, Sarah Tucker College, Tirunelveli, India Article history: Submitted 17 June 2014; Accepted 28 June 2014; Available online 14 July 2014 Abstract Kalakad Mundanthurai Tiger Reserve (KMTR), which is the home for many endemic, rare and endangered species of plants and animals, is one of the priority hotspot for conservation. Insect pests, being the most destructive agents affecting forest and shade trees are by far the most numerous animal life inhabiting the forest. As the primary predators of nocturnal insects, bats play a significant role in all forested ecosystems. The insect eating bats allocate considerable time and effort with different flight strategies to forage the available insect prey . Among the insectivorous bats of the forest ecosystem, the species of Rhinolophus genus help to control insect pest populations throughout its geographic range. The studies on faecal pellet analysis has proved their control over the lepidopteran, coleopteran, dipteran, hymenopteran, homopteran, orthopteran, hemipteran trichopteran and neuropteran group of insect pests that infects the forest trees of KMTR. The valuable pest controlling service provided by the study species needs special protection through conservation measures. Key words: Foraging, insect pests, KMTR, primary predators, pest controller Corresponding author Dr. Juliet Vanitharani, Professor and Head, Zoology Department and Research Centre, Sarah Tucker College, Tirunelveli - 7, Tamilnadu, India Introduction Kalakad Mundanthurai Tiger Reserve (KMTR), which is situated in the south Western Ghats of India one among the 18 world biodiversity hotspots which is bound by forests in west, North and South and by villages in the East. This is the only area of
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Insect pest management by the horse shoe bats (Rhinolophus species) in the forest ecosystem of...

SIRJ-BES Volume 1 Issue 5 (2014) ISSN 2348 - 5787 www.scrutinyjournals.com
Scrutiny International Research Journal of Biological and
Environmental Science (SIRJ-BES)
Insect pest management by the horse shoe bats (Rhinolophus species) in the forest ecosystem of Kalakad
Mundanthurai Tiger Reserve, India
Selva Ponmalar S1 and Juliet Vanitharani2
Bat Research Laboratory, Zoology Department and Research Centre,
Sarah Tucker College, Tirunelveli, India
Article history: Submitted 17 June 2014; Accepted 28 June 2014; Available online 14 July 2014
Abstract
Kalakad Mundanthurai Tiger Reserve (KMTR), which is the home for many endemic, rare and
endangered species of plants and animals, is one of the priority hotspot for conservation. Insect pests, being the
most destructive agents affecting forest and shade trees are by far the most numerous animal life inhabiting the
forest. As the primary predators of nocturnal insects, bats play a significant role in all forested ecosystems. The
insect eating bats allocate considerable time and effort with different flight strategies to forage the available insect
prey . Among the insectivorous bats of the forest ecosystem, the species of Rhinolophus genus help to control
insect pest populations throughout its geographic range. The studies on faecal pellet analysis has proved their
control over the lepidopteran, coleopteran, dipteran, hymenopteran, homopteran, orthopteran, hemipteran
trichopteran and neuropteran group of insect pests that infects the forest trees of KMTR. The valuable pest
controlling service provided by the study species needs special protection through conservation measures.
Key words: Foraging, insect pests, KMTR, primary predators, pest controller
Corresponding author
Dr. Juliet Vanitharani,
Professor and Head,
Zoology Department and Research Centre,
Sarah Tucker College,
Tirunelveli - 7,
Tamilnadu, India
Introduction
Kalakad Mundanthurai Tiger Reserve (KMTR), which is situated in the south Western Ghats of India one among the 18 world biodiversity hotspots which is bound by forests in west, North and South and by villages in the East. This is the only area of

www.scrutinyjournals.com Ponmalar and Vanitharani / SIRJ-BES 1:5 (2014)
15
Western Ghats which has longest raining period of about 8 months. Insect pests are the most destructive agents affect forest and shade trees (Grindal and Brigham, 1999; Douce et al., 2002). They affect tree roots, stems, limbs, needles, leaves of healthy or weakened trees, hardwoods etc. Insectivorous bats play a significant role of pest management in all forested ecosystems (Barclay, 1985; Freeman, 1979; Fenton, 2003). Flight has enabled them to exploit a variety of foraging niches inaccessible to other mammals. Undoubtedly, they are the most gregarious and successful group of beneficial animals to the ecosystem and have lured the attention of ethologists.
Among the forest insectivorous bats, the species of the Rhinolophus are rich in number; play a crucial role thereby keeps the insect population under check. They forage and feed insects throughout its geographic range. They roost in large numbers in caves, underground tunnels and abandoned old buildings. These insectivorous bats have many morphological adaptations that allow them to capture and handle prey in flight. Their dietary adaptations are commonly reflected in the skull (feeding apparatus) and wing (flight apparatus) morphology. Teeth and skull are the morphological indicators with food and the relationship in dietary diversity (Neuweiler, 2001). To suit their diet preference they show considerable diversity in wing morphology (Fenton, 1992) and flight style (Neuweiler, 1984). The present study focuses on the impact of rhinolophids as pest manager in the forests of KMTR.
Materials and Methods
The field work was conducted between May 2011 and December 2013 in KMTR (Latitude - 8.6833o N; Longitude - 77.3167o E; Elevation range 40 – 1800 m). Initially, the study sites were surveyed to collect baseline data on bat species and abundance. The distribution of the study species was confirmed by erecting mist nets, setting canopy, harp nets and hand nets. Digital recording of mist netted bats was done for closer look on morphological features. During day hours, the bat roost search was done in the abandoned buildings, crevices and cracks, tunnels, caves etc. The bats roosted in these area were collected using hand nets and mist nets to identify the species and their morphology. Morphological measurements were taken by following the standard techniques (Bates and Harrison, 1997). The bats were then released back in the place where they were caught. The documented results were then analysed to determine the potential habitats and ecomorphological features of rhinolophids, the horseshoe bats of KMTR.
Dietary habitat analysis of studied rhinolophids was done by faecal pellet analysis and examination of culled parts beneath the feeding roost. The microphotographs were taken to confirm the insect parts identification. Identifications were made upto the order level with the help of authenticated literatures available on Indian insects (Nair, 1989; Borror 1992). The percentage frequency of insects consumed during the study period were calculated covering the four seasons (Post monsoon, Summer, Premonsoon and Winter) applying the following formulae.
𝑷𝒆𝒓𝒄𝒆𝒏𝒕𝒂𝒈𝒆 𝒇𝒓𝒆𝒒𝒖𝒆𝒏𝒄𝒚 = 𝑵𝒖𝒎𝒃𝒆𝒓 𝒐𝒇 𝒐𝒄𝒄𝒖𝒓𝒓𝒆𝒏𝒄𝒆 𝒐𝒇 𝒄𝒂𝒕𝒆𝒈𝒐𝒓𝒊𝒆𝒔
𝑻𝒐𝒕𝒂𝒍 𝒐𝒄𝒄𝒖𝒓𝒓𝒆𝒏𝒄𝒆 𝒐𝒇 𝒂𝒍𝒍 𝒄𝒂𝒕𝒆𝒈𝒐𝒓𝒊𝒆𝒔× 𝟏𝟎𝟎

Table No. 1a: Morphological measurements and body weight of available Rhinolophids of KMTR
S. No Name of the Species
Sex No. of individuals
Status of bat
Body weight
(g)
Fore arm
(mm)
Ear (mm)
Tail (mm)
Hind foot (mm)
Head to
body (mm)
Tibia (mm)
1 Rhinolophus rouxii ♂ 14 Adult 12.03± 1.22
49.64± 1.35
13.44± 2.26
20.79± 3.02
5.49± 1.92
49.46± 2.48
23.82± 1.28
♀ 25 Adult 12.21± 1.51
49.95± 1.46
13.37± 2.46
22.86± 2.23
6.40± 1.61
50.56± 2.10
23.44± 1.38
2 Rhinolophus pusillus ♂ 6 Adult 4.67± 0.28
38.47± 0.99
15.95± 0.55
19.00± 0.57
6.32± 0.40
37.23± 1.59
14.72± 0.29
♀ 3 Adult 4.30± 0.20
38.07± 0.90
15.80± 0.66
19.00± 0.66
6.13± 0.25
37.27± 1.26
15.23± 0.72
3 Rhinolophus lepidus ♂ 5 Adult 5.26± 0.42
40.92± 1.91
12.58± 2.86
18.28± 1.86
5.64± 2.24
40.10± 2.01
16.10± 0.78
♀ 14 Adult 5.11± 0.40
38.92± 1.71
10.76± 0.49
17.62± 2.11
4.47± 0.45
40.23± 1.93
16. 40± 0.67
4 Rhinolophus beddomei
♂♀ 3 Adult 21.40± 1.01
57.43± 0.59
27.73± 1.27
37.87± 0.75
10.87± 2.44
62.73± 12.57
30.53± 0.21
Value: Mean ± Standard deviation
www.scrutinyjournals.com Ponmalar and Vanitharani / SIRJ-BES 1:5 (2014)
17
Results
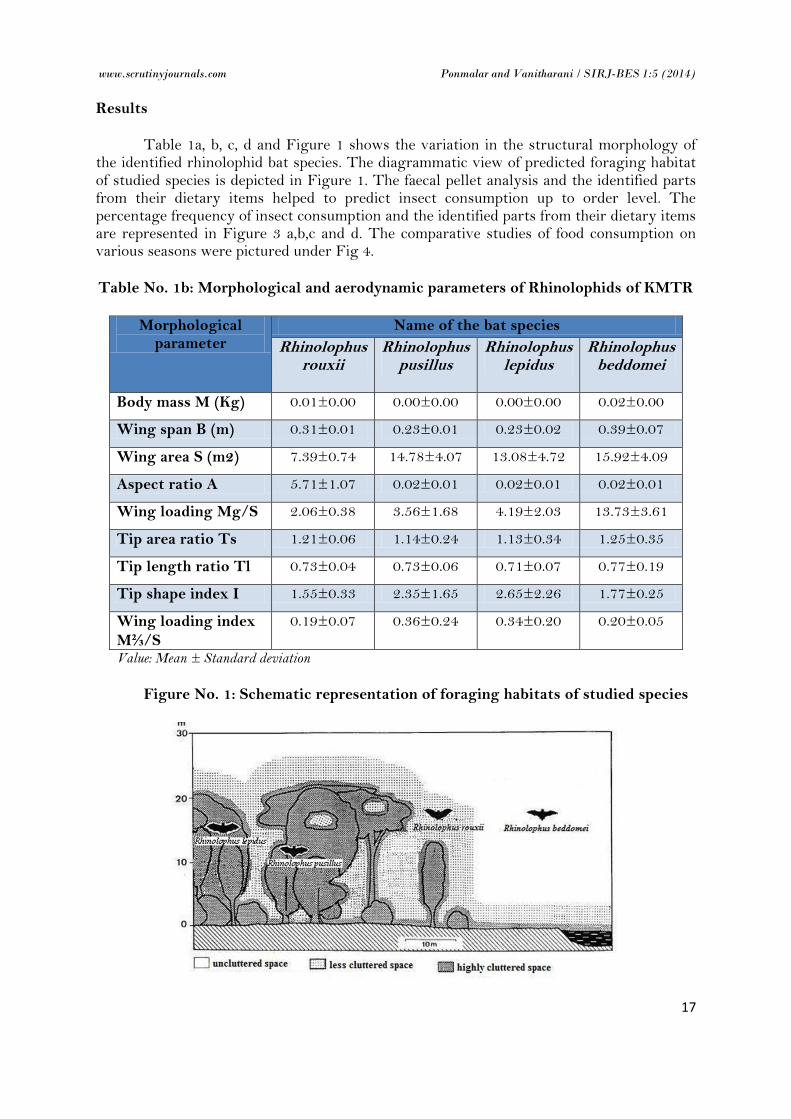
Table 1a, b, c, d and Figure 1 shows the variation in the structural morphology of the identified rhinolophid bat species. The diagrammatic view of predicted foraging habitat of studied species is depicted in Figure 1. The faecal pellet analysis and the identified parts from their dietary items helped to predict insect consumption up to order level. The percentage frequency of insect consumption and the identified parts from their dietary items are represented in Figure 3 a,b,c and d. The comparative studies of food consumption on various seasons were pictured under Fig 4.
Table No. 1b: Morphological and aerodynamic parameters of Rhinolophids of KMTR
Morphological parameter
Name of the bat species
Rhinolophus rouxii
Rhinolophus pusillus
Rhinolophus lepidus
Rhinolophus beddomei
Body mass M (Kg) 0.01±0.00 0.00±0.00 0.00±0.00 0.02±0.00
Wing span B (m) 0.31±0.01 0.23±0.01 0.23±0.02 0.39±0.07
Wing area S (m2) 7.39±0.74 14.78±4.07 13.08±4.72 15.92±4.09
Aspect ratio A 5.71±1.07 0.02±0.01 0.02±0.01 0.02±0.01
Wing loading Mg/S 2.06±0.38 3.56±1.68 4.19±2.03 13.73±3.61
Tip area ratio Ts 1.21±0.06 1.14±0.24 1.13±0.34 1.25±0.35
Tip length ratio Tl 0.73±0.04 0.73±0.06 0.71±0.07 0.77±0.19
Tip shape index I 1.55±0.33 2.35±1.65 2.65±2.26 1.77±0.25
Wing loading index
M⅔/S
0.19±0.07 0.36±0.24 0.34±0.20 0.20±0.05
Value: Mean ± Standard deviation
Figure No. 1: Schematic representation of foraging habitats of studied species
www.scrutinyjournals.com Ponmalar and Vanitharani / SIRJ-BES 1:5 (2014)
18
Discussion
Bats (Chiroptera), are the major contributors to mammalian biodiversity, comprising about 20% of mammalian species globally (Simmons, 2005). In tropical ecosystem the insect eating bats fulfill key ecosystem services as control agents of arthropod populations (Kalka et al., 2008; Williams-Guillén et al., 2008). The present study is focused on the impact of identified horseshoe bats of KMTR (Rhinolophus rouxii, Rhinolophus pusillus, Rhinolophus lepidus and Rhinolophus beddomei). These bats were captured from foraging area, roosting caves and tunnels at Sengaltheri, Kodayar, Mundanthurai and Servalar hills.
Morphological features
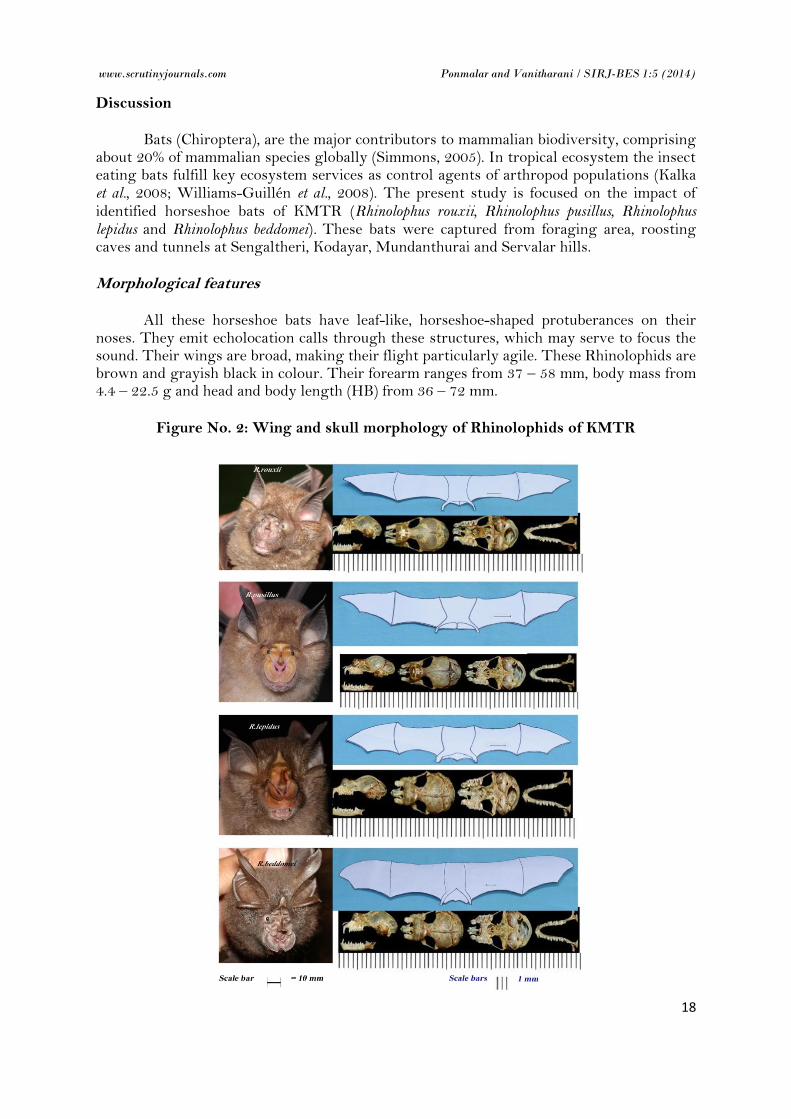
All these horseshoe bats have leaf-like, horseshoe-shaped protuberances on their noses. They emit echolocation calls through these structures, which may serve to focus the sound. Their wings are broad, making their flight particularly agile. These Rhinolophids are brown and grayish black in colour. Their forearm ranges from 37 – 58 mm, body mass from 4.4 – 22.5 g and head and body length (HB) from 36 – 72 mm.
Figure No. 2: Wing and skull morphology of Rhinolophids of KMTR

Table No. 1c: Cranial measurements of studied Rhinolophids
Expansions of the abbreviations are given below:
SIZE : Sum of the natural logs of Condylo-canine length, Zygomatic breadth and Temporal height; GTL : Greatest length of the skull; CBL : Condylobasal length; CCL : Condylo-canine length; ZB : Zygomatic breadth; BB : Breadth of the braincase; PC : Postorbital constriction; M : Mandible length; MSW : Minimum skull width; PSW : Posterior skull width; TH : Temporal height ; UC : Upper canine
Name of the bat species SIZE
(log)
Cranial measurements (mm)
GTL CBL CCL ZB BB PC M MSW PSW TH UC
Rhinolophus rouxii 7.3 21.71 19.15 18.34 11.17 10.29 3.12 14.73 2.7 8 6.8 3.1
Rhinolophus pusillus 6.5 17.44 15.56 14.24 8.22 8.02 2.45 10.93 2.2 7 5 3.1
Rhinolophus lepidus 6.7 17.17 15.04 14.1 8.28 7.86 2.5 10.63 2.3 7 5.2 1.9
Rhinolophus beddomei 7.8 27.44 23.86 23.45 13.48 11.86 2.72 18.93 3.1 9.6 8.7 3.5

Table No. 1d: Dental measurements of studied Rhinolophids of KMTR
Name of the bat species
Dental measurements (mm) Dental formula (i-incisors, c-canine,
p-premolar, m-molar) Upper jaw Lower jaw
PC PM1 PM3 CMn CMn M DD DT CPH CM Conl CC Con cor
Rhinolophus rouxii
2.83 7 8.4 8.03 8.69 14.7 1.9 0.07 3.01 5.09 4.3 13.3 4 -2- 1 -2-4 123 i ---- c --- pm ------- m -------- =32 12- 1 -234 123
Rhinolophus pusillus
2.45 4.9 5.1 5.8 6.01 10.03 1.2 0.06 2.1 2.9 3.9 9.5 2.1 -2- 1 -2-4 123 i ---- c --- pm ------- m -------- =32 12- 1 -234 123
Rhinolophus lepidus
2.5 5.1 5.9 6.1 6.9 10.9 1.5 0.06 2.9 3.8 2.4 10.2 2.4 -2- 1 -2-4 123 i ---- c --- pm ------ m -------- =32 12- 1 -234 123
Rhinolophus beddomei
2.72 7.3 9.1 8.9 10.01 18.93 2.38 1.14 4.51 5.9 4.2 16.9 5.1 -2- 1 -2-4 123 i ---- c --- pm ------ m -------- =32 12- 1 -234 123
Expansions of the abbreviations are given below:
PC : Postorbital constriction; PM1 : Palatal width from M1 ; PM3 : Posterior palatal width; CMn : Maxillary toothrow; CMn : Mandibular toothrow; CM : Condyle to lower molar toothrow; DD : Dentary depth; DT : Dentary thickness; Concor : Condyle to coronoid process Conl : Condyle length; CC : Condyle to canine; M : Mandible length; CPH : Coronoid process height;

Foraging and feeding strategy
A suite of morphological factors influences foraging behaviour in insectivorous bats, including body mass and the size and shape of the skull, jaws and wings (Freeman, 1979). Large bats with large jaws are capable of eating a wider range of prey sizes than small bats, resulting in a broader feeding niche. When food resources are plentiful, such differences are less apparent (Anthony and Kunz, 1977).
Differences in skull and dental morphology that correlate with dietary hardness are found among insectivores (Freeman, 1979; Dumont, 1999). The differentiation of the teeth and mandibular joint provide a guide to the type of diet that differ species of bats consume as well as the general dietary preferences within genus (Neuweiler, 2001). The Rhinolophus beddomei has wider palate than length. To consume large size prey they are adapted with broader palate rather than longer. This structure presumably made them to prefer hard food item like beetles, moth, caddish fly etc. The other study species R.rouxii, R.pusillus and R.lepidus seem to have narrow palate as they feed on butterflies, bugs, wasps and small moths, the soft prey items. This type of feeding behaviour correlated to the dental and skull structures were confirmed by the earlier studies (Philips, 1980; Advani, 1981; Whitaker et al., 1996; Bates and Harrison, 1997). The present study also confirms that the major dietary consumption of these two species is Lepidoptera and Coleoptera. The other study species R.lepidus and R.pusillus are soft eaters and their major feed items are dipterans and hemipterans.
The large difference in physical properties (size, hardness, toughness) of food item demands adaptations in cranial morphology and the performance of the feeding apparatus (Freeman, 1979). The assessment of physical properties of food items thus offer tremendous insights into the range of variation in cranial and mandibular adaptations, to suit food utilization patterns among insectivores (Strait, 1993). Studies of mastication (Storch, 1968) and analytical predictions of bite forces (Reduker, 1983) have also provided important bases for the ecomorphological adaptations to prey selection. The exhibited inter specific variation in bite force of the rhinolophids cause the species specific variation in preferred prey selection. Extensive studies of Dumont, (1999); Aguirre et al. (2002) and Dumont, (2003) on ecomorphological adaptations on food selection and trophic niche partitioning also substantiate the results.
R.beddomei has higher coronoid process allowing for the attachment of well developed jaw muscle which is essentially robust to counter the reaction forces while biting hard prey like beetles. These types of feeding behaviour in R.beddomei were strongly confirmed by Phillips (1980).
Bogdanowicz et al. (1998) said that the well developed coronoid process allows increased leverage and accommodated large jaw muscle. Increase in the size of temporal muscle can allow an animal to resist the increased stress of struggling, crush hard shelled items and facilitate more extensive use of canines (Bates and Harrison, 1997; Whitaker et al., 1999; Advani, 1981; Philips, 1980). But the other studied rhinolophids exhibit lesser coronoid process with thinner jaws. This characteristic assumes that these rhinolophids prefer soft food items like butterflies, grasshoppers, etc. Findings of Brosset (Brosset, 1962) Phillips (Phillips, 1980) and Jones (Jones et al., 1994) also confirmed the rhinolophid feed analysis results.

www.scrutinyjournals.com Ponmalar and Vanitharani / SIRJ-BES 1:5 (2014)
22
Figure No. 3a: Percentage frequency of insect consumption and the sample identified parts from the dietary items of Rhinolophus ruxii

www.scrutinyjournals.com Ponmalar and Vanitharani / SIRJ-BES 1:5 (2014)
23
Figure No. 3b: Percentage frequency of insect consumption and the sample identified parts from the dietary items of Rhinolophus pusillus

www.scrutinyjournals.com Ponmalar and Vanitharani / SIRJ-BES 1:5 (2014)
24
Figure No. 3c: Percentage frequency of insect consumption and the sample identified parts from the dietary items of Rhinolophus Lepidus

www.scrutinyjournals.com Ponmalar and Vanitharani / SIRJ-BES 1:5 (2014)
25
Figure No. 3d: Percentage frequency of insect consumption and the sample identified parts from the dietary items of Rhinolophus beddomei

www.scrutinyjournals.com Ponmalar and Vanitharani / SIRJ-BES 1:5 (2014)
26
Next to feeding apparatus the flight mechanism in bats are associated with partitioning of food resources among bats (Norberg and Rayner, 1987). Bats with different wing designs, varied flight style and performance show considerable diversity in diet preference (Neuweiler, 1984; Fenton, 1990). Body mass, wingspan and wing area are the primary measures of wing design. From these parameters, wing loading and aspect ratio are derived, which describe the size and shape of the wings respectively (Norberg and Rayner, 1987). Interaction between aspect ratio, wing loading and wing tip shape index affect flight (Norberg and Rayner, 1987; Neuweiler, 1989; Habersetzer and Storch, 1989; Rayner et al., 1989; Fenton, 1990). Manoeuverability and agility are the two factors that are strongly influenced by flight adaptation. These two factors explain the ability of bats to change the flight direction without loss of speed and with small turning radius.
Among the studied bat species, the R. beddomei has higher wing loading, lesser manoeuverability and forage in open uncluttered habitat. R. rouxii has wing loading lesser than R.beddomei and they forage in less cluttered habitat whereas the other two study species R. lepidus and R. pusillus have very lesser wing loading, greater maneurability and they exploit cluttered vegetation. They fly among foliage canopy catching insects among branches. Although Rhinolophidae has short wing tips, slow flight and low manoeuvrablity they can fly with same hovering often within clutter (Barrett-Hamilton, 1910; Wallin, 1969). The low wing loading and the short, cambered wings permit highly manoeuvrable flight and the slow light turns, which are essential for insect hawking near vegetation. These bats can also glean on resting insects.
Feeding impact of Rhinolophids in the forests of KMTR
The documented insect parts from the faecal pellet analysis shows the undigested parts of wing fragments, body fragments, leg parts etc. of various pest insects of the forest. The variation in the wing morphology is an indication to prove, bat species forage and cover all the niches. Their diversity in ecomorphology and species specificity avoids competition. A resource partition is possible through their diverse adaptations on wing shape, body size, and echocall sensory cues (Kasso and Balakrishnan, 2013). Studies of Jennings (Jennings et al., 2004) also say that bats choose species-specific types of food items and can detect the same according to their flight ability.
Altogether 9 insect orders were identified from all the faecal samples examined. Scales and body parts of moths (Lepidoptera F = 23 %) were the most dominated item in all the faecal samples of all the rhinolophids. That is followed by Coleoptera (F = 22%) and Diptera (F = 19%). Previous studies of Robinson and Stebbings (1993), on Eptesicus serotinus forest bat of U K also shows that the scales of moth wings were very conspicuous in faeces. In addition coleopteran and lepidopteran insects appear to be the second most important insect orders in the diet of Indiana bats (Kurta and Whitaker, 1996). The present study also shows that there is high frequency of lepidopteran and coleopteran fragments in the faeces of all the four studied species. The major nocturnal pest for the flora of any ecosystem is the caterpillers of moths (Whitaker, 1993). A single moth can lay upto 400-600 eggs. Consumption of a single moth itself causes a cascade effect on forest pest management.
The second higher percentage of identified parts Coleoptera, Diptera, Hymenoptera and Hemiptera. The larval forms of these representatives are the major pests in the forest ecosystem. Almost all the coleopteran beetles are hardwood pests and they infest the trees which cannot be even saved with insecticide treatments. Thereby, to limit beetle population,
www.scrutinyjournals.com Ponmalar and Vanitharani / SIRJ-BES 1:5 (2014)
27
the predation by bats on these insects in the forest ecosystem is very beneficial. In addition, the caterpillars of these pests are defoliators or miners of succulent plant tissues. The larval wood moths tunnel the heartwood of living trees resulting in timber degrade. The white grubs, root-feeding larvae are among the most damaging pests of tall fescue pasture, and turf grass in parts of the United States Australia, and New Zealand (Tashiro, 1987; Potter and Braman, 1991)
The presently studied rhinolophids also consume a major group of dipteran which includes the trill flies, mosquitoes, midges and variety of flies, leg dance flies, snipe flies, hover flies, shore flies, blow flies, dung flies, bat flies etc. The larvae of these dipterans produces various types of galls on stems, twigs, leaves, petioles, flowers, flower-stalks, fruits, and pods, or living under the bark, in decaying wood and bark (Skuhrava et al., 1984).
Hymenopterans like sawflies, leaf cutting wasps, bees and ants, beetles and walking sticks are some of the major defoliators that attack forest trees. Defoliation reduces photosynthesis, interferes with transpiration and translocation within the tree which in turn causes serious loss of tree growth and sometimes tree mortality (Douce et al., 2002)
Earlier studies have proved and indicated that these bat species can consume more than 25% of their body mass of insects each night (Kunz 1988; Fenton 1990; Kasso and Balakrishnan, 2013). They frequently selects nutrient-rich abdomen of insects while discarding the wings, head, and appendages, which greatly increases feeding efficiency and the quantity of insects consumed. In this way, pests are limited by natural ecosystems of which bats play a potential role as pest manager.
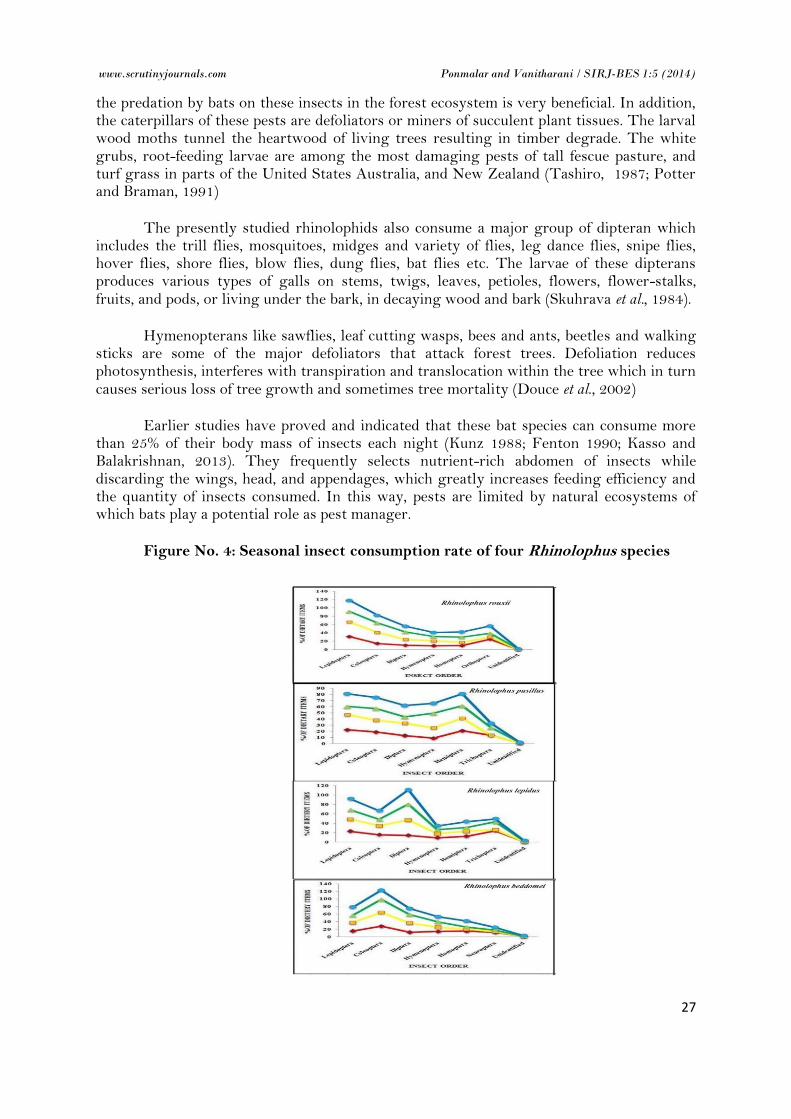
Figure No. 4: Seasonal insect consumption rate of four Rhinolophus species

www.scrutinyjournals.com Ponmalar and Vanitharani / SIRJ-BES 1:5 (2014)
28
The comparative study on seasonal insect consumption rate shows that there is variation in the dietary consumption rate. Seasonal dietary prey selection shows that there is a seasonal trend among the major dietary components. The food availability is higher during the summer time followed by post monsoon, premonsoon and winter. The most dominent food component Lepidoptera, Coleoptera and Diptera shows that there is high availability in summer than in winter. The other food components like hymenopterans are more in premonsoon, homopterans are more in late winter, orthopterans, hemipterans, trichopterans and neuropterans are more in postmonsoon period. Experiments and field observations show that bats can discriminate among various habitats and items of food and adjust their behaviour to utilize, more–or-less optimally to these said varying resources (Barclay, 1985; Norberg, 1981). The wide range of food selection confirmed these bats adaptations and ability to consume various foods in different seasons (Howes, 1974; Zinn and Humphrey, 1981; Jones, 1990; Beck, 1995; Gloor et al., 1995, Taake, 1996, Rydell and Peterson, 1998).
Conclusion
Forest insect outbreaks cause economic damage to forests which in turn affect the wealth of the country. The present impact study shows that there is availability of certain harmful insect pests throughout the year. The role of the studied rhinolophids is of great importance in the forest ecosystem as pest manager. Among the insectivorous bats, the species of Rhinolophus are richly and widely distributed in the forest ecosystem and are likely to control insect pest populations throughout its geographic range. These species eats a number of serious Polyphagus insect pest species of forest trees and thus has a promising role in biological control. Their foraging and roosting areas like caves, tunnels, rock crevices and canopy must be protected. Threats like roost disturbances, firing, deforestation greatly affect their population. Roost protection is the major conservation measure to protect these valuable species. The present study so far has evaluated the importance of these species and their value of pest suppression. This research may thereby lead to improve control measures. The bat conservation will not only control the pest population but also replenish the forest ecosystem.
Acknowledgement
Special thanks and acknowledgement to Tamil Nadu Forest Department, Kalakad Mundanthurai Tiger Reserve, Tirunelveli, for granting permission to conduct the survey and logistical advice. Thanks to Dr. Paul Bates, Dr. David Harrison and Dr. Nikky Thomas, Harrison Institute, UK for their help in the identification of specimens. JV would like to extend her grateful thanks to Mohamed bin Zayed Species Conservation Fund, Abu Dhabi, United Arab Emirates for the financial support to the field work.
Reference
Alankar, S., Jain, R., Agarwal, R. K. and Ahirwar, D., 2008. Clinical trial of herbal drugs and products in India: Past and current status and critical issues. Research Jounal of Pharmacy and Technology, 1(2): 69-74.
Advani, R., 1981. Seasonal fluctuations in the feeding ecology of the Indian false vampire Megaderma lyra lyra (Chiroptera: Megadermatidae) in Rajasthan. Zeits. Saug., 46 (2):90-93

www.scrutinyjournals.com Ponmalar and Vanitharani / SIRJ-BES 1:5 (2014)
29
Anthony, E.I.P. and Kunz, T.K., 1977. Feeding strategies of little brown bat, Myotis lucifugus in Southern NewHampshire. Ecology, 58:775-786.
Aguirre, L.F., Herrel, A., Van Damme, R. and Matthyssen, E., 2002. Ecomorphological analysis of trophic niche partitioning in a tropical savanna bat community. Proc. R. Soc. Lond. B Biol. Sci., 269:1271 -1278.
Barclay, R.M.R., 1985. Long- versus short-range foraging strategies of hoary (Lasiurus cinereus) and silver-haired (Lasionycteris noctivagans) bats and the consequences for prey selection. Canadian Journal of Zoology, 63: 2507-2515.
Bates, P.J.J. and Harrison, D.L., 1997. Bats of the Indian subcontinent. Harrison Zool. Mus. Pub., England.
Beck, A., 1995. Fecal analyses of european bat species. Myotis, 32-33: 109–119.
Borror, D., 1992. An Introduction to the study of insects.Harcourt Bace College Publishers, USA.879-896.
Bogdanowicz, W., Kasper, S. and Owen, R.D., 1998. Phylogeny of plecotine bats: reevaluation of morphological and chromosomal data. J. Mammalogy, 79: 78-90.
Brosset, A., 1962. La reproduction des Chiropteres de 1'ouest et du centre de ITnde. Mammalia, 26: 176-213.
Douce, G.K., Moorhead, D.J. and Bargeron, C.T., 2002. Forest Pest Control, The University of Georgia, College of Agricultural and Environmental Sciences, Special Bulletin 16, Revised January.
Dumont, E.R., 1999. The effect of food hardness on feeding behaviour in frugivorous bats (Family Phyllostomidae): an experimental study. Journal of Zoology, 248: 219-229.
Dumont, E.R., 2003. Bats and fruit: an ecomorphological approach. In Bat Ecology (ed. T. H. Kunz and M. B. Fenton), Chicago: University of Chicago Press. pp. 398-429.
Fenton, M.B., 1990. The foraging behaviour and ecology of animal eating bats. Can. J.Zool., 68: 411-422.
Fenton, M.B., 1992. Pellage pattern and crypsis in roosting bats. Taphozous mauritianus and Ephomophorus species. Koe., 35: 49-55.
Fenton, M.B., 2003. Science and the conservation of bats: where to next?. Wildlife Society Bulletin, 31(1): 6–15.
Freeman, P.W., 1979. Specialized insectivory: Beetle-eating and moth-eating molossid bats. Journal of Mammalogy, 60: 467-479.
Gloor, S., Stutz, H.P.B. and Ziswiler, V., 1995. Nutritional habits of the Nocutle bat Nyc-talus noctula (Schreber, 1774) in Switzerland. Myotis., 32-33: 231–242.
Grindal, S.D. and Brigham, R.M., 1999. Impacts of forest harvesting on habitat use by foraging insectivorous bats at different spatial scale. Ecoscience, 6: 25-34.

www.scrutinyjournals.com Ponmalar and Vanitharani / SIRJ-BES 1:5 (2014)
30
Habersetzer, J. and Storch, G., 1989. Ecology and echolocation of the Eocene Messel bats. In: Hana´ k V,Hora´cˇek I and Gaisler J, eds. European bat research 1987. Prague: Charles University Press, 213–133.
Howes, C.A., 1974. Notes on the prey and feeding behaviour of the noctule bat. Naturalist., 930: 107–110.
Jennings, N.V., Parsons, S., Arlow, K.E.B. and Annon, M.R.C., 2004. Echolocation calls and wing morphology of bats from the West Indies. Act.Chirop., 6(1): 75-90.
Jones, G., 1990. Prey selection by the greater horseshoe bat (Rhinolophus ferrumequinum): optimal foraging by echolocation? Journal of Animal Ecology, 59(2): 587-602.
Jones, G.C., John, H.L.H. and Moshe, S., 1994. Organisms as ecosystem engineers. Oikos, 69: 373-386.
Kalko, E.K.V., Estrada, V.S., Schmidt, M., Wegmann, M. and Meyer, C.F.J., 2008. Flying high – assessing the use of the aerosphere by bats. Integrative and Comparative Biology, 48, 60–73.
Kasso, M. and Balakrishnan, M., 2013. Ecological and economic importance of bats (ORDER CHIROPTERA), Department of Zoological Sciences, Addis Ababa University, P.O. Box 1176, Addis Ababa, Ethiopia.
Kelm, D.H., Wiesner, K.R. and Von, H.O., 2008. Effects of artificial roosts for frugivorous bats on seed dispersal in a Neotropical forest pasture mosaic. Conservation Biology, 22, 733–741.
Kunz, T.H., 1988. Food habit analysis of insectivorous bats. In: Ecological and behavioural methods for the study of bats (Kunz T. H. ed.), Simithsonian Institution Press, Washington, London. 171-209.
Kurta, A. and Whitaker, J.O., 1996. Diet of endangered Indiana bat, Myotis sodalis in Michigan. Bat Research News, 37:139.
Lobova, T.A., Geiselman, C.K. and Mori, S.A., 2009. Seed Dispersal by Bats in the Neotropics. New York Botanical Garden Press, New York.
Nair, P.K.R., 1989. Agroforestry systems in the tropics. Kluwer Academic publishers. Dordrecht. Netherlands.
Neuweiler, G., 1984. Evolutionary Aspects of Bat Echolocation, J. Comp. Physiol. A., 189: 245–256.
Neuweiler, G., 1989. Foraging ecology and audition in echolocating bats. Trends Ecol. Evol., 4: 160–166.
Neuweiler, G., 2001. The biology of bats, Published by Oxford University Press.
Norberg, U.M., 1981. Allometry of bat wings and legs and comparison with bird wings. Phil. Trans. R. Soc., 292: 359-398.
Norberg, U.M. and Rayner, J.M., 1987. Ecological Morphology and Flight in Bats (Mammalia; Chiroptera): Wing Adaptation, Flight Performance, Foraging Strategy and Echolocation, Phil. Trans. R. Soc. Lond. B, 1987, pp 335–427.

www.scrutinyjournals.com Ponmalar and Vanitharani / SIRJ-BES 1:5 (2014)
31
Pavey, C.R., Burwell, C.J., Grunwald, J.E., Marshall, C.J. and Neuweiler, G., 2001. Dietary benefits of twilight foraging by insectivorous bats. Hipposideros speoris. Biotropica, 33: 670-681.
Philips, W.W.A., 1980. Manual of the mammal of Sri Lanka. Part-1, wildlife and nature protection Society of srilanka, 1-116.
Potter, D.A. and Braman, S.K., 1991. Ecology and management of turfgrass insects. Annual. Review of Entomology, 36: 383-406.
Rayner, H.C., Atkins, R.C. and Westerman, R.A., 1989. Relief of local stump pain by capsaicin cream. Lancet, 2: 1276-1277.
Reduker, D.W. 1983. Functional analysis of the masticatory apparatus in two species of Myotis. J. Mammal, 64: 277-286.
Robinson, M.F. and Stebbings, R.E., 1993. Food of the serotine bat, Eptesicus serotinus’s faecal analysis a valid qualitative and quantitative technique? Journal of Zoology, 231: 239-248.
Rydell, J. and Petersons, G., 1998. The diet of the noctule bat Nyctalus noctula in Latvia. Z. Säugetierkd, 63: 79–83.
Simmons, N.B., 2005. Order Chiroptera, In: D.E. Wilson and D.M. Reeder (eds.), Mammal Species of the World, 3rd Edition, Volume 1. The John Hopkins University Press, Baltimore, MD, USA. 2142. pp.312-529.
Strait, S.G., 1993. Differences in occlusal morphology and molar size in frugivores and faunivores. Journal of Human evolution, 25: 471–484.
Storch, G., 1968. Funktionsmorphologische Untersuchungen an der Kaumuskulatur und an korrelierten Schadelstrukturen der Chi- ropteren. Abh. Senckenb. Naturforsch. Ges., 517: 1-92.
Skuhrava, M., Skuhravy, V. and Brewer, J.W., 1984. Biology of gall midges, p. 169-222. In Ananthakrishan, T.N. Biology of Insect Galls. Word & IBH. New Delhi, Bombay, Calcutta. 362 p.
Tashiro, H., 1987. Turfgrass insects of the United States and Canada. Cornell University Press, Ithaca NYand London. 391pp.
Taake, K.H., 1996. Beutetiere westfälischer Abendsegler (Nyctalus noctula). Myotis. Bonn., 34: 121–122.
Von, H.O. and Winter, Y., 2003. Glossophagine bats and their flowers: costs and benefits for plants and pollinators. In: Kunz, T.H., Fenton, M.B. (Eds.), Bat Ecology. University of Chicago Press, Chicago, pp. 346–397.
Whitaker, J.O., 1993. Bats, Beetles and Bugs. More bigbrown bats as agricultural pests. Bats, 11: 23.
Whitaker, J.O., Neefus, C. and Kunz, T.H., 1996. Dietary variation in the Mexican free-tailed bat (Tadarida brasiliensis mexicana). Journal of Mammalogy, 77: 716-724.
Zinn, T.L. and Humphrey, S.R., 1981. Seasonal food resources and prey selection of the southeastern brown bat (Myotis austroriparius) in Florida. Florida Scientist, 44: 81-90.

www.scrutinyjournals.com Ponmalar and Vanitharani / SIRJ-BES 1:5 (2014)
32
Wallin, L., 1969. The Japanese bat fauna. Zool Bidrg, 37: 226–440.
Williams, G.K., Perfecto, I. and Vandermeer, J., 2008. Bats limit insects in a Neotropical agroforestry system. Science, 320: 70.




















