
Factors influencing protein tyrosine nitration—structure-based predictive models
Upload
independentCategory
view
1download
0
Inhibition of Endothelial Nitric Oxide Synthase Activity byProline-Rich Tyrosine Kinase 2 in Response to
Fluid Shear Stress and InsulinBeate Fisslthaler, Annemarieke E. Loot, Annisuddin Mohamed, Rudi Busse,† Ingrid Fleming
Abstract—In native and primary cultures of endothelial cells, fluid shear stress elicits the tyrosine phosphorylation of theendothelial NO synthase (eNOS), however, the consequences of this modification on enzyme activity are unclear. Wefound that fluid shear stress induces the association of eNOS with the proline-rich tyrosine kinase 2 (PYK2) inendothelial cells and that the eNOS immunoprecipitated from eNOS- and PYK2-overexpressing HEK293 cells wastyrosine-phosphorylated on Tyr657. In mouse carotid arteries, the overexpression of wild-type PYK2, but not adominant-negative PYK2, decreased eNOS activity (�50%), whereas in murine lung endothelial cells, the downregu-lation of PYK2 (small interfering RNA) increased ionomycin-induced NO production. Mutation of Tyr657 to thephosphomimetic residues aspartate (D) or glutamate (E) abolished enzyme activity, whereas a nonphosphorylatablemutant (phenylalanine [F]) showed activity comparable to the wild-type enzyme. Moreover, normal flow-inducedvasodilatation was apparent in carotid arteries from eNOS�/� mice overexpressing either the wild-type eNOS or theY657F mutant, whereas no flow-induced vasodilatation was apparent in arteries expressing the Y657E eNOS mutant.Insulin also activated PYK2 and stimulated eNOS in endothelial cells expressing the Y657F mutant but not wild-typeeNOS. These data indicate that PYK2 mediates the tyrosine phosphorylation of eNOS on Tyr657 in response to fluidshear stress and insulin stimulation and that this modification attenuates the activity of the enzyme. The PYK2-dependent inhibition of NO production may serve to keep eNOS activity low and limit the detrimental consequences ofmaintained high NO output, ie, the generation of peroxynitrite. (Circ Res. 2008;102:1520-1528.)
Key Words: blood flow � insulin � mechanotransduction � nitric oxide � nitric oxide synthases � phosphorylation
Over the last 10 years, it has become clear that theendothelial nitric oxide (NO) synthase (eNOS) is regu-
lated both by changes in the intracellular concentration of freeCa2�, as well as by the phosphorylation of the enzyme.1 Mostis known about the role played by Ser1177 (human se-quence), which is situated in the reductase domain of theenzyme and Thr495, located in the calmodulin (CaM)-binding domain, in the regulation of NO production, thephosphorylation of which appear to play a reciprocal role inthe regulation of eNOS activity.2,3
There are several potentially phosphorylatable tyrosineresidues in eNOS, and there have been numerous reportsshowing that tyrosine kinase inhibitors attenuate endothelialNO production and flow-induced vasodilatation.4–6 It is clearthat the enzyme can be tyrosine-phosphorylated in endothe-lial cells treated with tyrosine phosphatase inhibitors,6,7
H2O2,7,8 or exposed to fluid shear stress,9 as well as in cellsoverexpressing v-Src.8,10 Indeed, Src was reported to phos-phorylate a tyrosine residue (Tyr83, bovine sequence; Tyr81,human sequence) in the oxygenase domain of eNOS in
bovine aortic endothelial cells. This modification was re-ported to be associated with an increase in NO production butbecause no differences in maximal eNOS activity weredetected between the wild-type and the phenylalanine(Tyr81Phe) eNOS mutant, the authors proposed that tyrosinephosphorylation does not modify eNOS activity directly butmay modulate the sensitivity of the enzyme to Ca2�, alterprotein–protein interactions, or change its subcellularlocalization.8
Given that the above-mentioned study was performedusing multipassaged cells overexpressing a specific tyrosinekinase and that we have previously only been able to detectendogenous eNOS tyrosine phosphorylation in primary cul-tures of endothelial cells,9 the aim of this study was toidentify the amino acid(s) in eNOS that is tyrosine-phosphor-ylated following endothelial cell stimulation by physiologicalstimuli, as well as the consequences of this modification onenzyme activity. Because fluid shear stress is known tostimulate the activation of c-Src11 and the proline-rich ty-rosine kinase (PYK2),12 we concentrated initially on the
Original received May 30, 2007; resubmission received January 14, 2008; revised resubmission received April 10, 2008; accepted May 6, 2008.From the Vascular Signalling Group, Institut fur Kardiovaskulare Physiologie, Goethe-Universitat, Frankfurt am Main, Germany.This manuscript was sent to Mark Taubman, Consulting Editor, for review by expert referees, editorial decision, and final disposition.†Deceased.Correspondence to Ingrid Fleming PhD, Vascular Signalling Group, Institut fur Kardiovaskulare Physiologie, Goethe-Universitat, Theodor-Stern-Kai
7, D-60590 Frankfurt am Main, Germany. E-mail [email protected]© 2008 American Heart Association, Inc.
Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/CIRCRESAHA.108.172072
1520
by guest on September 11, 2016
http://circres.ahajournals.org/D
ownloaded from
by guest on Septem
ber 11, 2016http://circres.ahajournals.org/
Dow
nloaded from
by guest on September 11, 2016
http://circres.ahajournals.org/D
ownloaded from
by guest on Septem
ber 11, 2016http://circres.ahajournals.org/
Dow
nloaded from

role of these kinases in the regulation of endothelial NOproduction.
Materials and MethodsMaterialsThe specific eNOS antibody used for Western blotting or immuno-precipitation was from Santa Cruz Biotechnology (Heidelberg,Germany), the specific phospho-Tyr657 eNOS antibody was gener-ated by Eurogentec (Searing, Belgium), and the anti-PYK2 was fromBD Biosciences (Heidelberg, Germany). The immunoprecipitationof tyrosine phosphorylated proteins was performed using a mixtureof antibodies (from Cell Signaling, Santa Cruz Biotechnology,Transduction, and Millipore). N�-Nitro-L-arginine (L-NA), N�-nitro-L-arginine methyl ester (L-NAME), insulin, ionomycin, and all othersubstances were from Sigma (Deisenhofen, Germany).
Cell Culture and TransfectionHuman umbilical vein cells, porcine aortic endothelial cells, andmurine lung endothelial cells were isolated and cultured as de-scribed.13 Confluent cultures were transferred to culture mediumcontaining 2% FCS and either maintained under static conditions orexposed to shear stress (12 dyne cm�2) in a cone-plate viscosimeter.6RFL6 fibroblasts were obtained from Dr Ellen Closs (Mainz,Germany) and cultured in F-12 Nutrient Mixture (Ham, Invitrogen)containing 10% FCS. Plasmids containing the PYK2 WT, or theK457M mutation of PYK2 were kindly provided by Dr Ivan Dikic(Frankfurt, Germany) and the overexpression plasmid for Src kinasewas from Dr Andre Blaukat (Heidelberg, Germany).
Immunoprecipitation and ImmunoblottingeNOS, tyrosine-phosphorylated proteins, or PYK2 were immunopre-cipitated with the appropriate antibodies. Detergent (Triton X-100)-soluble proteins or immunoprecipitates were heated with SDS-PAGEsample buffer and separated by SDS-PAGE, and specific proteinswere detected by immunoblotting as described.13
Metabolic LabelingPorcine aortic endothelial cells were incubated in phosphate-freeTyrode’s solution containing H3
32PO4 (Hartmann Analytic, Braun-schweig, Germany; 30 �mol/L, 0.125 mCi/mL, 12 hours) before theapplication of fluid shear stress (12 dyne cm�2, 10 minutes) or insulin(10 nmol/L, 15 minutes). The immunoprecipitation of eNOS,SDS-PAGE, and phospho–amino acid detection was performed asdescribed.6
Identification of the Phosphorylated TyrosineResidue in eNOSHEK293 cells were cotransfected with eNOS and PYK2 or Srcplasmids. After 48 hours, the cells were lysed with a hypotonic lysisbuffer (5 mmol/L Tris-HCl, pH 7.4, 2 mmol/L EDTA with proteaseinhibitors, 2 mmol/L orthovanadate and 10 nmol/L okadaic acid).The membrane fraction was isolated by centrifugation, and NADP-binding proteins were purified by application of the supernatant to aADP–sepharose column.14 The eluted proteins were concentrated,suspended in radioimmunoprecipitation assay buffer, and tyrosinephosphorylated proteins were immunoprecipitated with a mixture ofphospho-tyrosine–specific antibodies. Immunoprecipitates were sep-arated by SDS-PAGE, and the silver-stained protein band at 140 kDawas excised for further analysis (tryptic digest, immobilized metal-affinity chromatography and matrix-assisted laser desorption ioniza-tion–mass spectrometry [MALDI-MS]) by TopLab (Martinsried,Germany).
Small Interfering RNAPYK2 gene expression silencing in mouse endothelial cells, wasachieved by transfection of small interfering (si)RNA duplexes(Stealth Select RNAi, Invitrogen, Paisley, UK) with TransGene IIreagent (MoBiTec, Gottingen, Germany) as described.15 As a nega-
tive control, cells were treated with negative universal controloligonucleotides with medium GC content (Invitrogen).
Measurement of NO Formation and eNOS ActivityHEK293 cells were cotransfected with eNOS and either LacZ, c-Src,or PYK2 expression plasmids. The formation of NO in the trans-fected cells was determined by electron spin resonance spectroscopyusing DETC as spin trap.16 The activity of the eNOS in themicrosomal protein fraction of eNOS-overexpressing HEK293 cellswas determined by the conversion of 3H-L-arginine to L-citrulline asdescribed.3
Determination of cGMP ConcentrationCultured lung endothelial cells from wild-type or eNOS�/� micewere infected with eNOS-overexpressing adenoviruses or treatedwith siRNA, suspended by treatment with Accutase, and washed inHEPES-buffered Tyrode solution containing the phosphodiesteraseinhibitor 3-isobutyl-1-methylxanthine (IBMX) (1 mmol/L) and su-peroxide dismutase (100 U/mL). Approximately 5�104 cells wereadded to RFL6 fibroblasts cultured in 24-well plates and incubated(37°C) in the absence or presence of L-NA, insulin (1 �mol/L, 30minutes), or ionomycin (100 nmol/L, 2 minutes). Thereafter, theincubation was stopped by the addition of trichloroacetic acid (6%),and the concentration of cGMP was determined by a specificradioimmunoassay (GE Healthcare Buchler GmbH, Braunschweig,Germany).
Plasmids and Generation ofRecombinant AdenovirusesMyc-tagged human eNOS cDNA (GenBank accession no.NM_000603) in pcDNA3.1myc/His was used for in vitro mutagen-esis using the QuickChange XL site-directed mutagenesis kit(Stratagene, Amsterdam, Netherlands) with the oligonucleotides(mutated tyrosine codon indicated in bold letters) 5�-GGCTCCCGGGCATTCCCCCACTTCTG-3� (sense Y-F); 5�-GGCTCCCGGGCAGACCCCCACTTCTG-3� (sense Y-D); 5�-GGCTCCCGGGCAGAGCCCCACTTCTG-3� (sense Y-E) incombination with the corresponding antisense primers. pAdshuttle-CMV plasmids containing the wild-type human PYK2 (GenBankaccession no. BC036651) or K457M PYK2, as well as the wild-typeand Y657D, Y657E, and Y657F eNOS mutants, were generatedusing the pAdEasy system (kindly provided by Dr Bert Vogelstein,Baltimore, Md), and the viruses were generated as described.17
Adenoviral Transduction of Endothelial CellsSubconfluent endothelial cell cultures were infected with adenovi-ruses overexpressing PYK2 wild-type or the point mutants describedabove or eNOS wild-type or eNOS Y657 point mutants asdescribed.18
Adenoviral Infection of Mouse Carotid ArteriesMale C57BL/6 or eNOS�/� mice, 6 to 9 weeks of age (Charles River,Sulzfeld, Germany) were anesthetized with isoflurane. The carotidarteries were perfused with saline solution, partially freed of con-nective tissue, and incubated with virus solution (3�106 pfu in 20�L). Flow-induced vasodilatation was assessed 40 to 44 hours afterinfection as described.19
In Vitro PhosphorylationPYK2 was overexpressed in HEK293 cells, then immunoprecipitatedfrom Triton X-100 soluble proteins, and resuspended in kinasereaction buffer (40 mmol/L HEPES, pH 7.0; 80 mmol/L NaCl; 8%glycerine; 0.8 mmol/L EDTA; 0.8 mmol/L dithiothreitol; 5 mmol/LMgCl2). The kinase reaction contained 5 �g of either 1 of thepeptides (corresponding to human eNOS amino acids 647 to 666)biotinNH-CVFGLGSRAYPHFCAFARAV-COOH (Y657) orbiotinNH-CVFGLGSRAFPHFCAFARAV-COOH (Y657F) as sub-strate and 2 �Ci �-32P-ATP. After 30 minutes at 30°C, the reactionwas separated by 16.5% Tris/Tricine PAGE (cathode buffer: 0.1
Fisslthaler et al Tyrosine Phosphorylation of eNOS by PYK2 1521
by guest on September 11, 2016
http://circres.ahajournals.org/D
ownloaded from
mol/L Tris; 0.1 mol/L Tricine; 0.1% SDS; anode buffer: 0.2 mol/LTris/HCl, pH 8.9). The gel was extensively washed, and the peptidesand proteins were visualized by silver staining. After drying, the gelwas exposed to a x-ray film. The bands were excised, and theradioactive incorporation was quantified by scintillation counting.
StatisticsData are expressed as means�SEM. Statistical evaluation wasperformed with Student’s t test for unpaired data and 1-way ANOVAfollowed by a Bonferroni t test or ANOVA for repeated measureswhere appropriate. Values of P�0.05 were consideredstatistically significant.
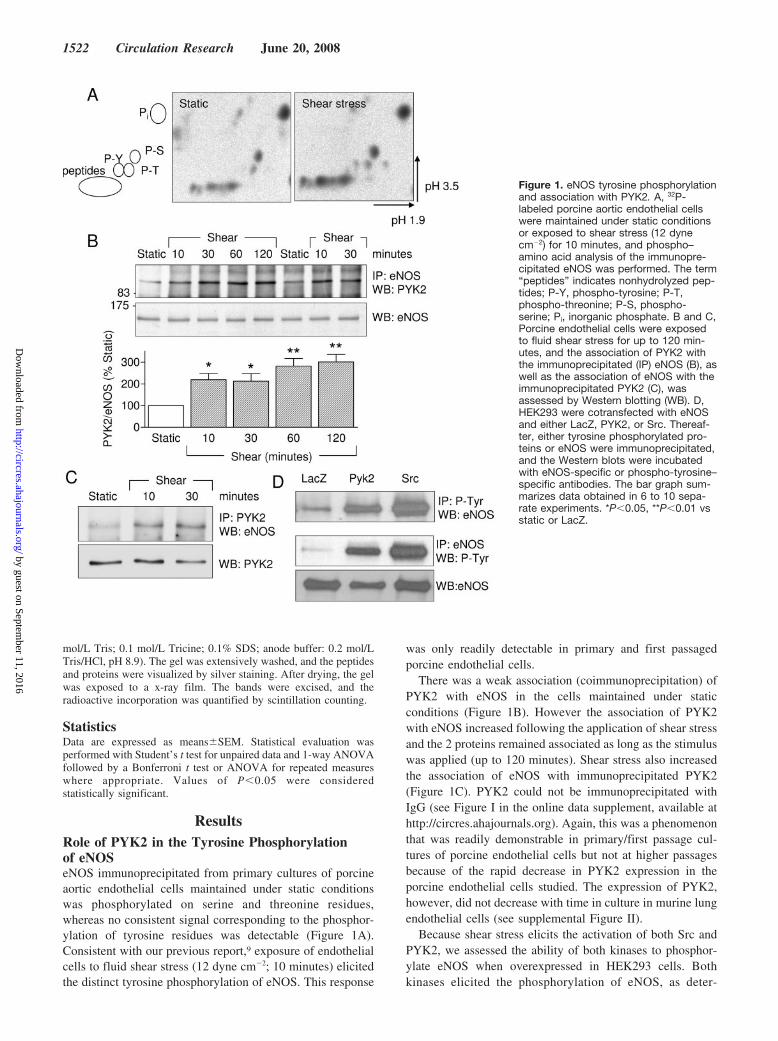
ResultsRole of PYK2 in the Tyrosine Phosphorylationof eNOSeNOS immunoprecipitated from primary cultures of porcineaortic endothelial cells maintained under static conditionswas phosphorylated on serine and threonine residues,whereas no consistent signal corresponding to the phosphor-ylation of tyrosine residues was detectable (Figure 1A).Consistent with our previous report,9 exposure of endothelialcells to fluid shear stress (12 dyne cm�2; 10 minutes) elicitedthe distinct tyrosine phosphorylation of eNOS. This response
was only readily detectable in primary and first passagedporcine endothelial cells.
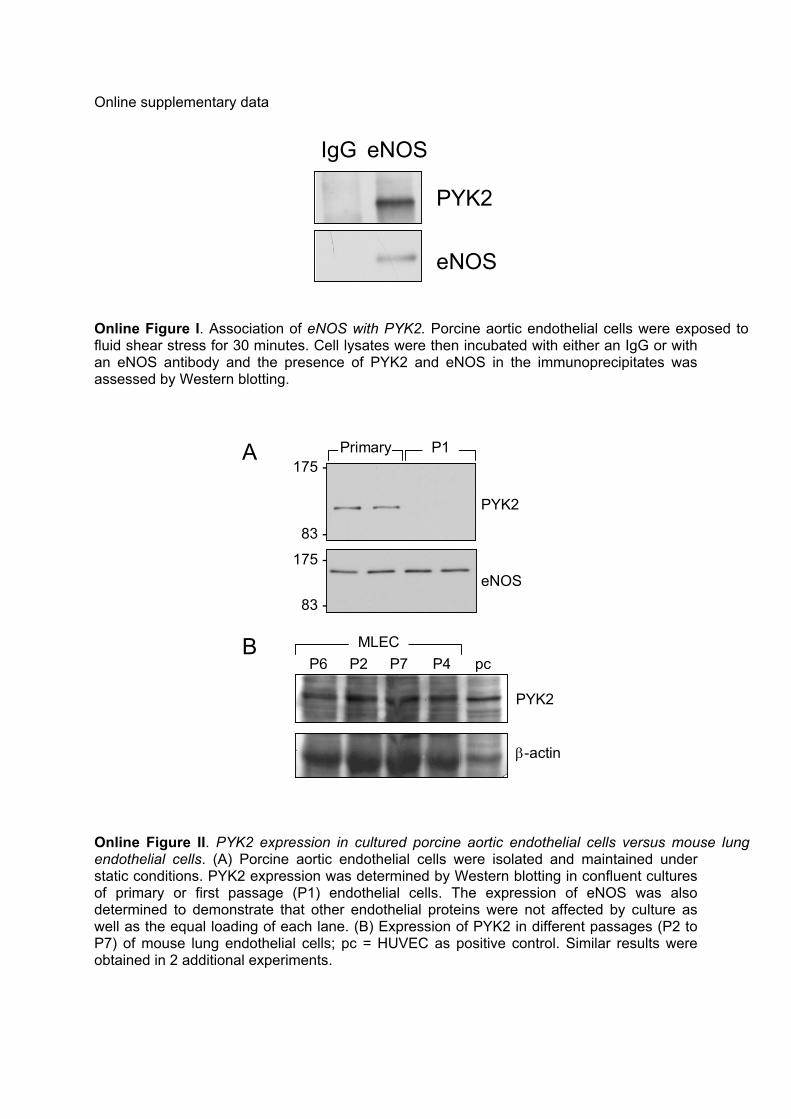
There was a weak association (coimmunoprecipitation) ofPYK2 with eNOS in the cells maintained under staticconditions (Figure 1B). However the association of PYK2with eNOS increased following the application of shear stressand the 2 proteins remained associated as long as the stimuluswas applied (up to 120 minutes). Shear stress also increasedthe association of eNOS with immunoprecipitated PYK2(Figure 1C). PYK2 could not be immunoprecipitated withIgG (see Figure I in the online data supplement, available athttp://circres.ahajournals.org). Again, this was a phenomenonthat was readily demonstrable in primary/first passage cul-tures of porcine endothelial cells but not at higher passagesbecause of the rapid decrease in PYK2 expression in theporcine endothelial cells studied. The expression of PYK2,however, did not decrease with time in culture in murine lungendothelial cells (see supplemental Figure II).
Because shear stress elicits the activation of both Src andPYK2, we assessed the ability of both kinases to phosphor-ylate eNOS when overexpressed in HEK293 cells. Bothkinases elicited the phosphorylation of eNOS, as deter-
Figure 1. eNOS tyrosine phosphorylationand association with PYK2. A, 32P-labeled porcine aortic endothelial cellswere maintained under static conditionsor exposed to shear stress (12 dynecm�2) for 10 minutes, and phospho–amino acid analysis of the immunopre-cipitated eNOS was performed. The term“peptides” indicates nonhydrolyzed pep-tides; P-Y, phospho-tyrosine; P-T,phospho-threonine; P-S, phospho-serine; Pi, inorganic phosphate. B and C,Porcine endothelial cells were exposedto fluid shear stress for up to 120 min-utes, and the association of PYK2 withthe immunoprecipitated (IP) eNOS (B), aswell as the association of eNOS with theimmunoprecipitated PYK2 (C), wasassessed by Western blotting (WB). D,HEK293 were cotransfected with eNOSand either LacZ, PYK2, or Src. Thereaf-ter, either tyrosine phosphorylated pro-teins or eNOS were immunoprecipitated,and the Western blots were incubatedwith eNOS-specific or phospho-tyrosine–specific antibodies. The bar graph sum-marizes data obtained in 6 to 10 sepa-rate experiments. *P�0.05, **P�0.01 vsstatic or LacZ.
1522 Circulation Research June 20, 2008
by guest on September 11, 2016
http://circres.ahajournals.org/D
ownloaded from
mined by the immunoprecipitation of tyrosine phosphory-lated proteins and Western blotting for eNOS or vice versa(Figure 1D).
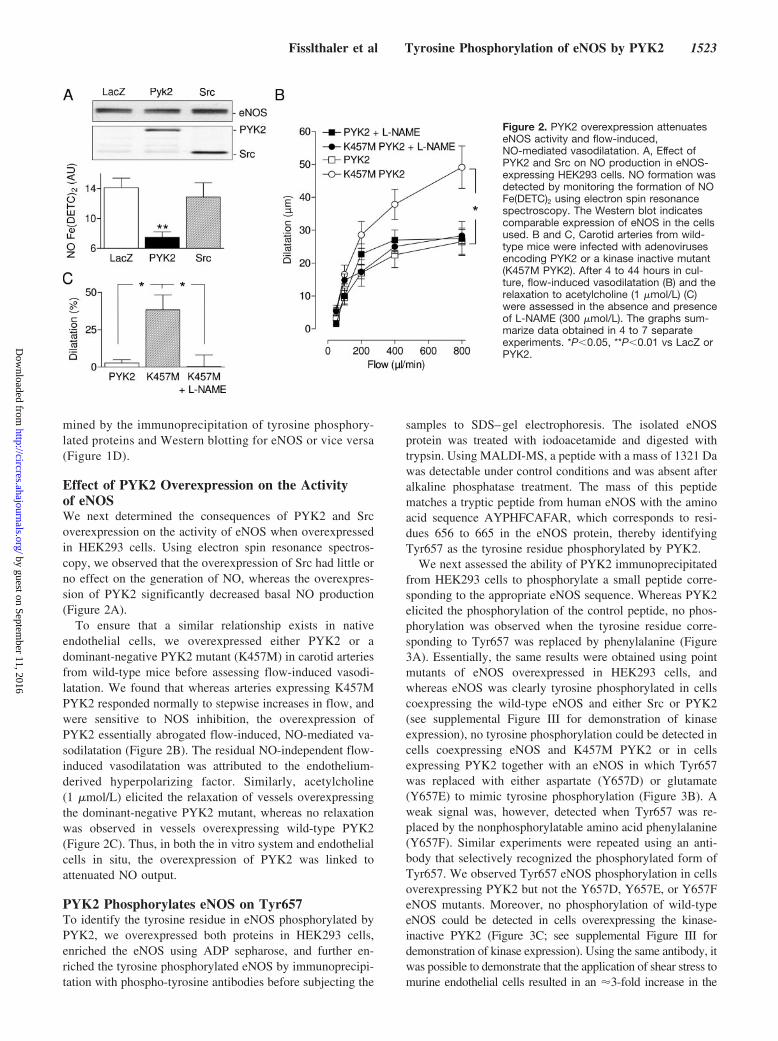
Effect of PYK2 Overexpression on the Activityof eNOSWe next determined the consequences of PYK2 and Srcoverexpression on the activity of eNOS when overexpressedin HEK293 cells. Using electron spin resonance spectros-copy, we observed that the overexpression of Src had little orno effect on the generation of NO, whereas the overexpres-sion of PYK2 significantly decreased basal NO production(Figure 2A).
To ensure that a similar relationship exists in nativeendothelial cells, we overexpressed either PYK2 or adominant-negative PYK2 mutant (K457M) in carotid arteriesfrom wild-type mice before assessing flow-induced vasodi-latation. We found that whereas arteries expressing K457MPYK2 responded normally to stepwise increases in flow, andwere sensitive to NOS inhibition, the overexpression ofPYK2 essentially abrogated flow-induced, NO-mediated va-sodilatation (Figure 2B). The residual NO-independent flow-induced vasodilatation was attributed to the endothelium-derived hyperpolarizing factor. Similarly, acetylcholine(1 �mol/L) elicited the relaxation of vessels overexpressingthe dominant-negative PYK2 mutant, whereas no relaxationwas observed in vessels overexpressing wild-type PYK2(Figure 2C). Thus, in both the in vitro system and endothelialcells in situ, the overexpression of PYK2 was linked toattenuated NO output.
PYK2 Phosphorylates eNOS on Tyr657To identify the tyrosine residue in eNOS phosphorylated byPYK2, we overexpressed both proteins in HEK293 cells,enriched the eNOS using ADP sepharose, and further en-riched the tyrosine phosphorylated eNOS by immunoprecipi-tation with phospho-tyrosine antibodies before subjecting the
samples to SDS–gel electrophoresis. The isolated eNOSprotein was treated with iodoacetamide and digested withtrypsin. Using MALDI-MS, a peptide with a mass of 1321 Dawas detectable under control conditions and was absent afteralkaline phosphatase treatment. The mass of this peptidematches a tryptic peptide from human eNOS with the aminoacid sequence AYPHFCAFAR, which corresponds to resi-dues 656 to 665 in the eNOS protein, thereby identifyingTyr657 as the tyrosine residue phosphorylated by PYK2.
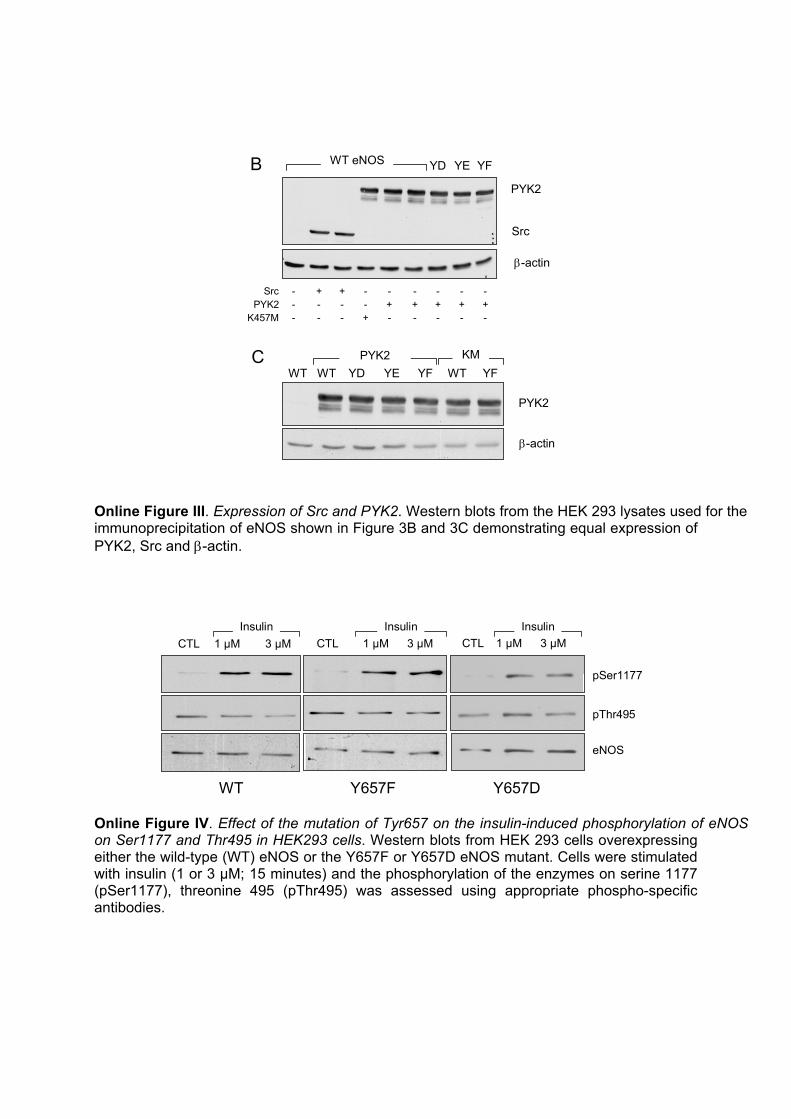
We next assessed the ability of PYK2 immunoprecipitatedfrom HEK293 cells to phosphorylate a small peptide corre-sponding to the appropriate eNOS sequence. Whereas PYK2elicited the phosphorylation of the control peptide, no phos-phorylation was observed when the tyrosine residue corre-sponding to Tyr657 was replaced by phenylalanine (Figure3A). Essentially, the same results were obtained using pointmutants of eNOS overexpressed in HEK293 cells, andwhereas eNOS was clearly tyrosine phosphorylated in cellscoexpressing the wild-type eNOS and either Src or PYK2(see supplemental Figure III for demonstration of kinaseexpression), no tyrosine phosphorylation could be detected incells coexpressing eNOS and K457M PYK2 or in cellsexpressing PYK2 together with an eNOS in which Tyr657was replaced with either aspartate (Y657D) or glutamate(Y657E) to mimic tyrosine phosphorylation (Figure 3B). Aweak signal was, however, detected when Tyr657 was re-placed by the nonphosphorylatable amino acid phenylalanine(Y657F). Similar experiments were repeated using an anti-body that selectively recognized the phosphorylated form ofTyr657. We observed Tyr657 eNOS phosphorylation in cellsoverexpressing PYK2 but not the Y657D, Y657E, or Y657FeNOS mutants. Moreover, no phosphorylation of wild-typeeNOS could be detected in cells overexpressing the kinase-inactive PYK2 (Figure 3C; see supplemental Figure III fordemonstration of kinase expression). Using the same antibody, itwas possible to demonstrate that the application of shear stress tomurine endothelial cells resulted in an �3-fold increase in the
Figure 2. PYK2 overexpression attenuateseNOS activity and flow-induced,NO-mediated vasodilatation. A, Effect ofPYK2 and Src on NO production in eNOS-expressing HEK293 cells. NO formation wasdetected by monitoring the formation of NOFe(DETC)2 using electron spin resonancespectroscopy. The Western blot indicatescomparable expression of eNOS in the cellsused. B and C, Carotid arteries from wild-type mice were infected with adenovirusesencoding PYK2 or a kinase inactive mutant(K457M PYK2). After 4 to 44 hours in cul-ture, flow-induced vasodilatation (B) and therelaxation to acetylcholine (1 �mol/L) (C)were assessed in the absence and presenceof L-NAME (300 �mol/L). The graphs sum-marize data obtained in 4 to 7 separateexperiments. *P�0.05, **P�0.01 vs LacZ orPYK2.
Fisslthaler et al Tyrosine Phosphorylation of eNOS by PYK2 1523
by guest on September 11, 2016
http://circres.ahajournals.org/D
ownloaded from
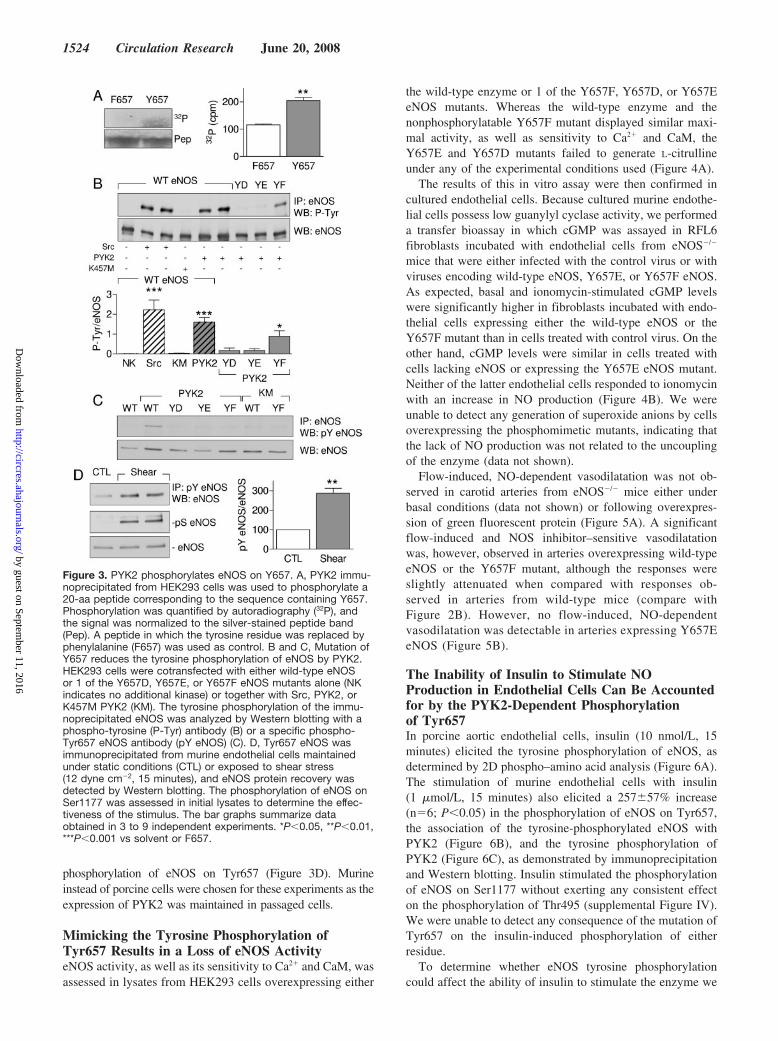
phosphorylation of eNOS on Tyr657 (Figure 3D). Murineinstead of porcine cells were chosen for these experiments as theexpression of PYK2 was maintained in passaged cells.
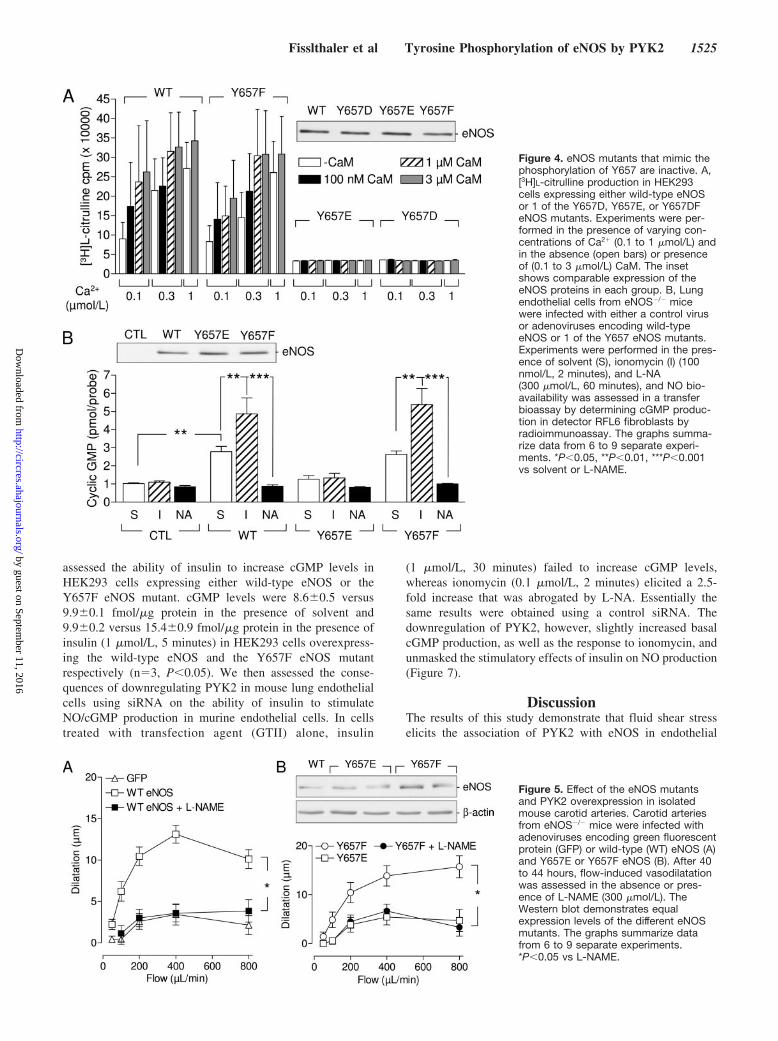
Mimicking the Tyrosine Phosphorylation ofTyr657 Results in a Loss of eNOS ActivityeNOS activity, as well as its sensitivity to Ca2� and CaM, wasassessed in lysates from HEK293 cells overexpressing either
the wild-type enzyme or 1 of the Y657F, Y657D, or Y657EeNOS mutants. Whereas the wild-type enzyme and thenonphosphorylatable Y657F mutant displayed similar maxi-mal activity, as well as sensitivity to Ca2� and CaM, theY657E and Y657D mutants failed to generate L-citrullineunder any of the experimental conditions used (Figure 4A).
The results of this in vitro assay were then confirmed incultured endothelial cells. Because cultured murine endothe-lial cells possess low guanylyl cyclase activity, we performeda transfer bioassay in which cGMP was assayed in RFL6fibroblasts incubated with endothelial cells from eNOS�/�
mice that were either infected with the control virus or withviruses encoding wild-type eNOS, Y657E, or Y657F eNOS.As expected, basal and ionomycin-stimulated cGMP levelswere significantly higher in fibroblasts incubated with endo-thelial cells expressing either the wild-type eNOS or theY657F mutant than in cells treated with control virus. On theother hand, cGMP levels were similar in cells treated withcells lacking eNOS or expressing the Y657E eNOS mutant.Neither of the latter endothelial cells responded to ionomycinwith an increase in NO production (Figure 4B). We wereunable to detect any generation of superoxide anions by cellsoverexpressing the phosphomimetic mutants, indicating thatthe lack of NO production was not related to the uncouplingof the enzyme (data not shown).
Flow-induced, NO-dependent vasodilatation was not ob-served in carotid arteries from eNOS�/� mice either underbasal conditions (data not shown) or following overexpres-sion of green fluorescent protein (Figure 5A). A significantflow-induced and NOS inhibitor–sensitive vasodilatationwas, however, observed in arteries overexpressing wild-typeeNOS or the Y657F mutant, although the responses wereslightly attenuated when compared with responses ob-served in arteries from wild-type mice (compare withFigure 2B). However, no flow-induced, NO-dependentvasodilatation was detectable in arteries expressing Y657EeNOS (Figure 5B).
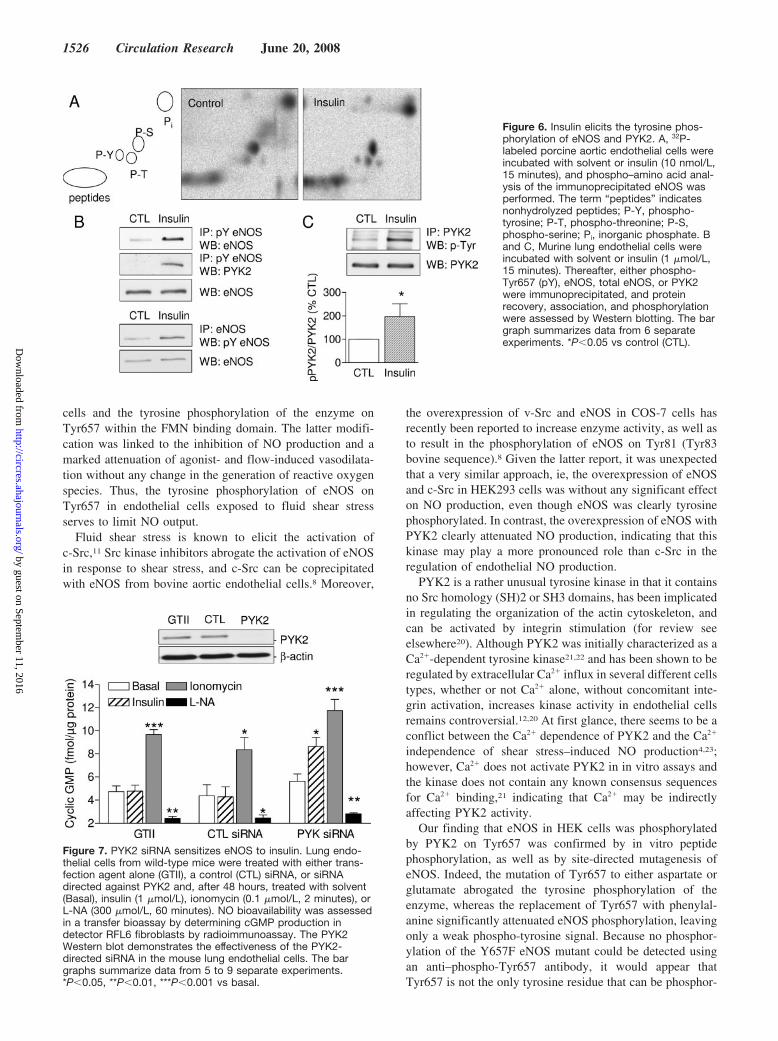
The Inability of Insulin to Stimulate NOProduction in Endothelial Cells Can Be Accountedfor by the PYK2-Dependent Phosphorylationof Tyr657In porcine aortic endothelial cells, insulin (10 nmol/L, 15minutes) elicited the tyrosine phosphorylation of eNOS, asdetermined by 2D phospho–amino acid analysis (Figure 6A).The stimulation of murine endothelial cells with insulin(1 �mol/L, 15 minutes) also elicited a 257�57% increase(n�6; P�0.05) in the phosphorylation of eNOS on Tyr657,the association of the tyrosine-phosphorylated eNOS withPYK2 (Figure 6B), and the tyrosine phosphorylation ofPYK2 (Figure 6C), as demonstrated by immunoprecipitationand Western blotting. Insulin stimulated the phosphorylationof eNOS on Ser1177 without exerting any consistent effecton the phosphorylation of Thr495 (supplemental Figure IV).We were unable to detect any consequence of the mutation ofTyr657 on the insulin-induced phosphorylation of eitherresidue.
To determine whether eNOS tyrosine phosphorylationcould affect the ability of insulin to stimulate the enzyme we
Figure 3. PYK2 phosphorylates eNOS on Y657. A, PYK2 immu-noprecipitated from HEK293 cells was used to phosphorylate a20-aa peptide corresponding to the sequence containing Y657.Phosphorylation was quantified by autoradiography (32P), andthe signal was normalized to the silver-stained peptide band(Pep). A peptide in which the tyrosine residue was replaced byphenylalanine (F657) was used as control. B and C, Mutation ofY657 reduces the tyrosine phosphorylation of eNOS by PYK2.HEK293 cells were cotransfected with either wild-type eNOSor 1 of the Y657D, Y657E, or Y657F eNOS mutants alone (NKindicates no additional kinase) or together with Src, PYK2, orK457M PYK2 (KM). The tyrosine phosphorylation of the immu-noprecipitated eNOS was analyzed by Western blotting with aphospho-tyrosine (P-Tyr) antibody (B) or a specific phospho-Tyr657 eNOS antibody (pY eNOS) (C). D, Tyr657 eNOS wasimmunoprecipitated from murine endothelial cells maintainedunder static conditions (CTL) or exposed to shear stress(12 dyne cm�2, 15 minutes), and eNOS protein recovery wasdetected by Western blotting. The phosphorylation of eNOS onSer1177 was assessed in initial lysates to determine the effec-tiveness of the stimulus. The bar graphs summarize dataobtained in 3 to 9 independent experiments. *P�0.05, **P�0.01,***P�0.001 vs solvent or F657.
1524 Circulation Research June 20, 2008
by guest on September 11, 2016
http://circres.ahajournals.org/D
ownloaded from
assessed the ability of insulin to increase cGMP levels inHEK293 cells expressing either wild-type eNOS or theY657F eNOS mutant. cGMP levels were 8.6�0.5 versus9.9�0.1 fmol/�g protein in the presence of solvent and9.9�0.2 versus 15.4�0.9 fmol/�g protein in the presence ofinsulin (1 �mol/L, 5 minutes) in HEK293 cells overexpress-ing the wild-type eNOS and the Y657F eNOS mutantrespectively (n�3, P�0.05). We then assessed the conse-quences of downregulating PYK2 in mouse lung endothelialcells using siRNA on the ability of insulin to stimulateNO/cGMP production in murine endothelial cells. In cellstreated with transfection agent (GTII) alone, insulin
(1 �mol/L, 30 minutes) failed to increase cGMP levels,whereas ionomycin (0.1 �mol/L, 2 minutes) elicited a 2.5-fold increase that was abrogated by L-NA. Essentially thesame results were obtained using a control siRNA. Thedownregulation of PYK2, however, slightly increased basalcGMP production, as well as the response to ionomycin, andunmasked the stimulatory effects of insulin on NO production(Figure 7).
DiscussionThe results of this study demonstrate that fluid shear stresselicits the association of PYK2 with eNOS in endothelial
Figure 4. eNOS mutants that mimic thephosphorylation of Y657 are inactive. A,[3H]L-citrulline production in HEK293cells expressing either wild-type eNOSor 1 of the Y657D, Y657E, or Y657DFeNOS mutants. Experiments were per-formed in the presence of varying con-centrations of Ca2� (0.1 to 1 �mol/L) andin the absence (open bars) or presenceof (0.1 to 3 �mol/L) CaM. The insetshows comparable expression of theeNOS proteins in each group. B, Lungendothelial cells from eNOS�/� micewere infected with either a control virusor adenoviruses encoding wild-typeeNOS or 1 of the Y657 eNOS mutants.Experiments were performed in the pres-ence of solvent (S), ionomycin (I) (100nmol/L, 2 minutes), and L-NA(300 �mol/L, 60 minutes), and NO bio-availability was assessed in a transferbioassay by determining cGMP produc-tion in detector RFL6 fibroblasts byradioimmunoassay. The graphs summa-rize data from 6 to 9 separate experi-ments. *P�0.05, **P�0.01, ***P�0.001vs solvent or L-NAME.
Figure 5. Effect of the eNOS mutantsand PYK2 overexpression in isolatedmouse carotid arteries. Carotid arteriesfrom eNOS�/� mice were infected withadenoviruses encoding green fluorescentprotein (GFP) or wild-type (WT) eNOS (A)and Y657E or Y657F eNOS (B). After 40to 44 hours, flow-induced vasodilatationwas assessed in the absence or pres-ence of L-NAME (300 �mol/L). TheWestern blot demonstrates equalexpression levels of the different eNOSmutants. The graphs summarize datafrom 6 to 9 separate experiments.*P�0.05 vs L-NAME.
Fisslthaler et al Tyrosine Phosphorylation of eNOS by PYK2 1525
by guest on September 11, 2016
http://circres.ahajournals.org/D
ownloaded from
cells and the tyrosine phosphorylation of the enzyme onTyr657 within the FMN binding domain. The latter modifi-cation was linked to the inhibition of NO production and amarked attenuation of agonist- and flow-induced vasodilata-tion without any change in the generation of reactive oxygenspecies. Thus, the tyrosine phosphorylation of eNOS onTyr657 in endothelial cells exposed to fluid shear stressserves to limit NO output.
Fluid shear stress is known to elicit the activation ofc-Src,11 Src kinase inhibitors abrogate the activation of eNOSin response to shear stress, and c-Src can be coprecipitatedwith eNOS from bovine aortic endothelial cells.8 Moreover,
the overexpression of v-Src and eNOS in COS-7 cells hasrecently been reported to increase enzyme activity, as well asto result in the phosphorylation of eNOS on Tyr81 (Tyr83bovine sequence).8 Given the latter report, it was unexpectedthat a very similar approach, ie, the overexpression of eNOSand c-Src in HEK293 cells was without any significant effecton NO production, even though eNOS was clearly tyrosinephosphorylated. In contrast, the overexpression of eNOS withPYK2 clearly attenuated NO production, indicating that thiskinase may play a more pronounced role than c-Src in theregulation of endothelial NO production.
PYK2 is a rather unusual tyrosine kinase in that it containsno Src homology (SH)2 or SH3 domains, has been implicatedin regulating the organization of the actin cytoskeleton, andcan be activated by integrin stimulation (for review seeelsewhere20). Although PYK2 was initially characterized as aCa2�-dependent tyrosine kinase21,22 and has been shown to beregulated by extracellular Ca2� influx in several different cellstypes, whether or not Ca2� alone, without concomitant inte-grin activation, increases kinase activity in endothelial cellsremains controversial.12,20 At first glance, there seems to be aconflict between the Ca2� dependence of PYK2 and the Ca2�
independence of shear stress–induced NO production4,23;however, Ca2� does not activate PYK2 in in vitro assays andthe kinase does not contain any known consensus sequencesfor Ca2� binding,21 indicating that Ca2� may be indirectlyaffecting PYK2 activity.
Our finding that eNOS in HEK cells was phosphorylatedby PYK2 on Tyr657 was confirmed by in vitro peptidephosphorylation, as well as by site-directed mutagenesis ofeNOS. Indeed, the mutation of Tyr657 to either aspartate orglutamate abrogated the tyrosine phosphorylation of theenzyme, whereas the replacement of Tyr657 with phenylal-anine significantly attenuated eNOS phosphorylation, leavingonly a weak phospho-tyrosine signal. Because no phosphor-ylation of the Y657F eNOS mutant could be detected usingan anti–phospho-Tyr657 antibody, it would appear thatTyr657 is not the only tyrosine residue that can be phosphor-
Figure 6. Insulin elicits the tyrosine phos-phorylation of eNOS and PYK2. A, 32P-labeled porcine aortic endothelial cells wereincubated with solvent or insulin (10 nmol/L,15 minutes), and phospho–amino acid anal-ysis of the immunoprecipitated eNOS wasperformed. The term “peptides” indicatesnonhydrolyzed peptides; P-Y, phospho-tyrosine; P-T, phospho-threonine; P-S,phospho-serine; Pi, inorganic phosphate. Band C, Murine lung endothelial cells wereincubated with solvent or insulin (1 �mol/L,15 minutes). Thereafter, either phospho-Tyr657 (pY), eNOS, total eNOS, or PYK2were immunoprecipitated, and proteinrecovery, association, and phosphorylationwere assessed by Western blotting. The bargraph summarizes data from 6 separateexperiments. *P�0.05 vs control (CTL).
Figure 7. PYK2 siRNA sensitizes eNOS to insulin. Lung endo-thelial cells from wild-type mice were treated with either trans-fection agent alone (GTII), a control (CTL) siRNA, or siRNAdirected against PYK2 and, after 48 hours, treated with solvent(Basal), insulin (1 �mol/L), ionomycin (0.1 �mol/L, 2 minutes), orL-NA (300 �mol/L, 60 minutes). NO bioavailability was assessedin a transfer bioassay by determining cGMP production indetector RFL6 fibroblasts by radioimmunoassay. The PYK2Western blot demonstrates the effectiveness of the PYK2-directed siRNA in the mouse lung endothelial cells. The bargraphs summarize data from 5 to 9 separate experiments.*P�0.05, **P�0.01, ***P�0.001 vs basal.
1526 Circulation Research June 20, 2008
by guest on September 11, 2016
http://circres.ahajournals.org/D
ownloaded from

ylated by PYK2 and that the mutation of 1 phosphorylationsite may influence the phosphorylation of other residues inmuch the same way that the mutation of some of the serinephosphorylation sites can be compensated for by the phos-phorylation of others.24
The consequence of eNOS Tyr657 phosphorylation wasnot, as expected, a subtle modification of NO production orthe Ca2� sensitivity of the enzyme but a complete loss ofenzyme activity. However, a clue to why the mutation of thistyrosine residue could have such dramatic effects can befound by considering the mechanisms known to regulate theactivity of the neuronal NOS, which was recently reported tobe determined by a large-scale swinging motion of the FMNdomain to deliver electrons to the catalytic module in theholoenzyme.25 From the crystal structure of neuronal NOS,the phosphorylation of a tyrosine residue (Tyr889, rat neuro-nal NOS sequence), which is in the vicinity of the FMNdomain, could prevent its movement, essentially locking theFMN domain into its electron-accepting position, thus inhib-iting enzyme activity.25 Because Tyr657 is the equivalenttyrosine residue in the human eNOS sequence, it is highlylikely that its phosphorylation would be associated with a lossof NO production.
Perhaps the most physiologically intriguing observationmade during the course of this study relates to the sensitivityof eNOS to insulin. We have repeatedly found that althoughthe stimulation of native or primary cultures of endothelialcells with insulin results in the rapid phosphorylation of Akt,as well as eNOS on Ser1177, it fails to activate eNOS andincrease endothelial cGMP levels26 or elicit relaxation.27
Given that the phosphorylation of Tyr657 inhibited eNOSactivity, we hypothesized that the insulin-induced activationof PYK2 and tyrosine phosphorylation of Tyr657 may func-tionally antagonize the effects of the Ser1177 phosphoryla-tion. Indeed, we found that insulin stimulated the tyrosinephosphorylation of both PYK2 and eNOS in murine endo-thelial cells and that the downregulation of PYK2 (usingsiRNA) rendered eNOS sensitive to insulin. At this stage, itshould be noted, however, that it is impossible to rule out thatthe manipulation of PYK2 may also alter other regulatoryphosphorylation sites on eNOS or the interaction with otherproteins, even though we were unable to detect any conse-quence of Tyr657 mutation in the insulin-induced phosphor-ylation of eNOS on Ser1177.
Although shear stress elicits the activation of eNOS inendothelial cells, the level of NO output is generally low (2-to 4-fold above basal levels) compared to that generatedfollowing agonist stimulation (�20 fold). The kinetics of theresponses are also markedly different because the shearstress–induced production of NO can be demonstrated as longas the stimulus remains constant, whereas that induced byagonists takes the form of a transient burst of NO productionrarely lasting more than a few minutes. Given that shear stresselicits the phosphorylation of a tyrosine residue that nega-tively regulates eNOS activity, it is tempting to speculate thatthis event plays a key role in negatively modulating enzymeactivity, thus keeping NO output low and reducing the risk ofcofactor, ie, tetrahydrobiopterin depletion and the uncouplingof the enzyme. It will be interesting to determine whether
eNOS dysregulation in pathophysiological states is associatedwith changes in the activity of PYK2 and the tyrosinephosphorylation of eNOS.
AcknowledgmentsWe dedicate this article to the memory of R.B. We are indebted toIsabel Winter, Mechtild Piepenbrock, and Katarina Bruch for experttechnical assistance.
Sources of FundingThis study was supported by Deutsche Forschungsgemeinschaftgrant SFB 553, B1, and by EICOSANOX, an integrated projectNetwork of Excellence supported by the European Community’sSixth Framework Programme (contract no. LSHM-CT-2004-005033). The authors are members of the European VascularGenomic Network, a Network of Excellence supported by theEuropean Community’s Sixth Framework Programme (contract no.LSHM-CT-2003-503254).
DisclosuresNone.
References1. Fleming I, Busse R. Molecular mechanisms involved in the regulation of
the endothelial nitric oxide synthase. Am J Physiol Regul Integr CompPhysiol. 2003;284:R1–R12.
2. Harris MB, Ju H, Venema VJ, Liang H, Zou R, Michell BJ, Chen Z-P,Kemp BE, Venema RC. Reciprocal phosphorylation and regulation of theendothelial nitric oxide synthase in response to bradykinin stimulation.J Biol Chem. 2001;19:16587–16591.
3. Fleming I, Fisslthaler B, Dimmeler S, Kemp BE, Busse R. Phosphory-lation of Thr495 regulates Ca2�/calmodulin-dependent endothelial nitricoxide synthase activity. Circ Res. 2001;88:e68–e75.
4. Ayajiki K, Kindermann M, Hecker M, Fleming I, Busse R. IntracellularpH and tyrosine phosphorylation but not calcium determine shear stress-induced nitric oxide production in native endothelial cells. Circ Res.1996;78:750–758.
5. Corson MA, James NL, Latta SE, Nerem RM, Berk BC, Harrison DG.Phosphorylation of endothelial nitric oxide synthase in response to fluidshear stress. Circ Res. 1996;79:984–991.
6. Fleming I, Bauersachs J, Fisslthaler B, Busse R. Ca2�-independent acti-vation of the endothelial nitric oxide synthase in response to tyrosinephosphatase inhibitors and fluid shear stress. Circ Res. 1998;82:686–695.
7. Garcıa-Cardena G, Fan G, Stern DF, Liu J, Sessa WC. Endothelial nitricoxide synthase is regulated by tyrosine phosphorylation and interacts withcaveolin-1. J Biol Chem. 1996;271:27237–27240.
8. Fulton D, Church JE, Ruan L, Li C, Sood SG, Kemp BE, Jennings IG,Venema RC. Src kinase activates endothelial nitric-oxide synthase byphosphorylating Tyr-83. J Biol Chem. 2005;280:35943–35952.
9. Fisslthaler B, Dimmeler S, Hermann C, Busse R, Fleming I. Phosphor-ylation and activation of the endothelial nitric oxide synthase by fluidshear stress. Acta Physiol Scand. 2000;168:81–88.
10. Takenouchi Y, Oo ML, Senga T, Watanabe Y, Machida K, Miyazaki K,Nimura Y, Hamaguchi M. Tyrosine phosphorylation of NOS3 in a breastcancer cell line and Src-transformed cells. Oncol Rep. 2004;11:1059–1062.
11. Okuda M, Takahashi M, Suero J, Murry CE, Traub O, Kawakatsu H,Berk BC. Shear stress stimulation of p130(cas) tyrosine phosphorylationrequires calcium-dependent c-Src activation. J Biol Chem. 1999;274:26803–26809.
12. Tai LK, Okuda M, Abe J, Yan C, Berk BC. Fluid shear stress activatesproline-rich tyrosine kinase via reactive oxygen species-dependentpathway. Arterioscler Thromb Vasc Biol. 2002;22:1790–1796.
13. Fleming I, Fisslthaler B, Dixit M, Busse R. Role of PECAM-1 in theshear-stress-induced activation of Akt and the endothelial nitric oxidesynthase (eNOS) in endothelial cells. J Cell Sci. 2005;118:4103–4111.
14. Martasek P, Liu Q, Liu J, Roman LJ, Gross SS, Sessa WC, Masters BS.Characterization of bovine endothelial nitric oxide synthase expressed inE. coli. Biochem Biophys Res Commun. 1996;219:359–365.
15. Potente M, Fisslthaler B, Busse R, Fleming I. 11,12-Epoxyeicosatrienoicacid-induced inhibition of FOXO factors promotes endothelial prolif-
Fisslthaler et al Tyrosine Phosphorylation of eNOS by PYK2 1527
by guest on September 11, 2016
http://circres.ahajournals.org/D
ownloaded from

eration by down-regulating p27Kip1. J Biol Chem. 2003;278:29619–29625.
16. Lenasi H, Kohlstedt K, Fichtlscherer B, Mulsch A, Busse R, Fleming I.Amlodipine activates the endothelial nitric oxide synthase by alteringphosphorylation on Ser1177 and Thr495. Cardiovasc Res. 2003;59:844–853.
17. He TC, Zhou S, da Costa LT, Yu J, Kinzler KW, Vogelstein B. Asimplified system for generating recombinant adenoviruses. Proc NatlAcad Sci U S A. 1998;95:2509–2514.
18. Michaelis UR, Fisslthaler B, Medhora M, Harder D, Fleming I, Busse R.Cytochrome P450 2C9-derived epoxyeicosatrienoic acids induce angio-genesis via cross-talk with the epidermal growth factor receptor (EGFR).FASEB J. 2003;17:770–772.
19. Dixit M, Loot AE, Mohamed A, Fisslthaler B, Boulanger CM,Ceacareanu B, Hassid A, Busse R, Fleming I. Gab1, SHP2, and proteinkinase A are crucial for the activation of the endothelial NO synthase byfluid shear stress. Circ Res. 2005;97:1236–1244.
20. Orr AW, Murphy-Ullrich JE. Regulation of endothelial cell function BYFAK and PYK2. Front Biosci. 2004;9:1254–1266.
21. Lev S, Moreno H, Martinez R, Canoll P, Peles E, Musacchio J, PlowmanG, Rudy B, Schlessinger J. Protein tyrosine kinase PYK2 involved inCa2�-induced regulation of ion channel and MAP kinase functions.Nature. 1995;376:737–745.
22. Yu H, Li X, Marchetto GS, Dy R, Hunter D, Calvo B, Dawson TL, WilmM, Anderegg RJ, Graves LM, Earp HS. Activation of a novel calcium-dependent protein-tyrosine kinase. Correlation with c-Jun N-terminalkinase but not mitogen-activated protein kinase activation. J Biol Chem.1996;271:29993–29998.
23. Dimmeler S, Fleming I, Fisslthaler B, Hermann C, Busse R, Zeiher AM.Activation of nitric oxide synthase in endothelial cells by Akt-dependentphosphorylation. Nature. 1999;399:601–605.
24. Bauer PM, Fulton D, Boo YC, Sorescu GP, Kemp BE, Jo H, Sessa WC.Compensatory phosphorylation and protein-protein interactions revealedby loss of function and gain of function mutants of multiple serinephosphorylation sites in endothelial nitric oxide synthase. J Biol Chem.2003;278:14841–14849.
25. Garcin ED, Bruns CM, Lloyd SJ, Hosfield DJ, Tiso M, Gachhui R, StuehrDJ, Tainer JA, Getzoff ED. Structural basis for isozyme-specific regu-lation of electron transfer in nitric-oxide synthase. J Biol Chem. 2004;279:37918–37927.
26. Fleming I, Schulz C, Fichtlscherer B, Kemp BE, Fisslthaler B, Busse R.AMP-activated protein kinase (AMPK) regulates the insulin-induced acti-vation of the nitric oxide synthase in human platelets. Thromb Haemost.2003;90:863–867.
27. Randriamboavonjy V, Schrader J, Busse R, Fleming I. Insulin induces therelease of vasodilator compounds from platelets by a nitric oxide-Gkinase-VAMP-3-dependent pathway. J Exp Med. 2004;199:347–356.
1528 Circulation Research June 20, 2008
by guest on September 11, 2016
http://circres.ahajournals.org/D
ownloaded from

Beate Fisslthaler, Annemarieke E. Loot, Annisuddin Mohamed, Rudi Busse and Ingrid Fleming2 in Response to Fluid Shear Stress and Insulin
Inhibition of Endothelial Nitric Oxide Synthase Activity by Proline-Rich Tyrosine Kinase
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 2008 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/CIRCRESAHA.108.1720722008;102:1520-1528; originally published online May 15, 2008;Circ Res.
http://circres.ahajournals.org/content/102/12/1520World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org/content/suppl/2008/05/15/CIRCRESAHA.108.172072.DC1.htmlData Supplement (unedited) at:
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Researchin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on September 11, 2016
http://circres.ahajournals.org/D
ownloaded from
Online supplementary data
PYK2
eNOS
IgG eNOS
Online Figure I. Association of eNOS with PYK2. Porcine aortic endothelial cells were exposed to fluid shear stress for 30 minutes. Cell lysates were then incubated with either an IgG or with an eNOS antibody and the presence of PYK2 and eNOS in the immunoprecipitates was assessed by Western blotting.
175 -
83 -
175 -
83 -
PYK2
eNOS
P1
P6 P2 P7 P4 pcMLEC
PYK2
β-actin
A
B
Primary
Online Figure II. PYK2 expression in cultured porcine aortic endothelial cells versus mouse lung endothelial cells. (A) Porcine aortic endothelial cells were isolated and maintained under static conditions. PYK2 expression was determined by Western blotting in confluent cultures of primary or first passage (P1) endothelial cells. The expression of eNOS was also determined to demonstrate that other endothelial proteins were not affected by culture as well as the equal loading of each lane. (B) Expression of PYK2 in different passages (P2 to P7) of mouse lung endothelial cells; pc = HUVEC as positive control. Similar results were obtained in 2 additional experiments.
PYK2
Src
β-actin
PYK2
β-actin
YD YE YFWT eNOSB
WT WT YD YE YF WT YFPYK2 KMC
Src + +- ------
K457MPYK2 + ++++- -- -
- -- + - -- --
Online Figure III. Expression of Src and PYK2. Western blots from the HEK 293 lysates used for the immunoprecipitation of eNOS shown in Figure 3B and 3C demonstrating equal expression of PYK2, Src and β-actin.
pThr495
WT
pSer1177
CTL
Y657F
3 µM1 µM CTL 3 µM1 µM
Y657D
CTL 3 µM1 µMInsulin Insulin Insulin
eNOS
Online Figure IV. Effect of the mutation of Tyr657 on the insulin-induced phosphorylation of eNOS on Ser1177 and Thr495 in HEK293 cells. Western blots from HEK 293 cells overexpressing either the wild-type (WT) eNOS or the Y657F or Y657D eNOS mutant. Cells were stimulated with insulin (1 or 3 µM; 15 minutes) and the phosphorylation of the enzymes on serine 1177 (pSer1177), threonine 495 (pThr495) was assessed using appropriate phospho-specific antibodies.
Copyright © 2022 FDOKUMEN




















