
Cystathionine -synthase is essential for female reproductive function
32
1 Cystathionine β-synthase is essential for female reproductive function Mario A. Guzmán, María A. Navarro, Ricardo Carnicer, Alfonso J. Sarría, Sergio Acín, Carmen Arnal, Pedro Muniesa, Joaquín C. Surra, José M. Arbonés-Mainar, Nobuyo Maeda and Jesús Osada. From the Departamento de Bioquímica y Biología Molecular y Celular (M.A.G., R.C., M.A.N., A.S., S.A., J.C.S., J.M.A.M., J.O.), Departamento de Anatomía y Embriología (P.M.), Departamento de Patología Animal (C.A.) Facultad de Veterinaria, Universidad de Zaragoza; Department of Pathology (N.M.), University of North Carolina at Chapel Hill, USA. Correspondence to Dr. Jesús Osada, Department of Biochemistry and Molecular Biology, Veterinary School, University of Zaragoza, Miguel Servet, 177, E-50013 Zaragoza, Spain. Fax number: 34-976-761612 Telephone number: 34-976-761644 E-mail: [email protected] © The Author 2006. Published by Oxford University Press. All rights reserved HMG Advance Access published September 19, 2006 by guest on March 17, 2014 http://hmg.oxfordjournals.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Cystathionine -synthase is essential for female reproductive function

1
Cystathionine β-synthase is essential for
female reproductive function
Mario A. Guzmán, María A. Navarro, Ricardo Carnicer, Alfonso J. Sarría, Sergio Acín, Carmen Arnal, Pedro Muniesa, Joaquín C. Surra, José M. Arbonés-Mainar, Nobuyo
Maeda and Jesús Osada.
From the Departamento de Bioquímica y Biología Molecular y Celular (M.A.G., R.C., M.A.N., A.S., S.A., J.C.S., J.M.A.M., J.O.), Departamento de Anatomía y Embriología
(P.M.), Departamento de Patología Animal (C.A.) Facultad de Veterinaria, Universidad de Zaragoza; Department of Pathology (N.M.), University of North Carolina at Chapel Hill,
USA.
Correspondence to Dr. Jesús Osada, Department of Biochemistry and Molecular Biology, Veterinary School, University of Zaragoza, Miguel Servet, 177, E-50013 Zaragoza, Spain.
Fax number: 34-976-761612 Telephone number: 34-976-761644
E-mail: [email protected]
© The Author 2006. Published by Oxford University Press. All rights reserved
HMG Advance Access published September 19, 2006 by guest on M
arch 17, 2014http://hm
g.oxfordjournals.org/D
ownloaded from

2
In human reproduction, hyperhomocysteinaemia has been reported as a risk factor for early
pregnancy loss and congenital birth defects. Hyperhomocysteinaemia is also recognized as
a cause of maternal obstetric complications such as preeclampsia. The role of plasma
hyperhomocysteinemia in female fertility was examined using cystathionine beta synthase
knockout (cbs KO) mice. Cbs KO females were infertile, showed alterations in the estrus
cycle, and an increased progesterone response during pseudopregnancy induction. Both cbs
KO ovaries and ovulated oocytes showed no major morphological alterations. However,
placental and uterine masses were decreased at day 18 of pregnancy and showed
morphological abnormalities. In cbs-KO pregnant females the number of uterine
implantation sites was not decreased despite the low number of surviving embryos. Fertility
was restored when cbs deficient ovaries were transplanted to normal ovarectomized
recipients. We detected an increased uterine expression of Grp78, a marker of endoplasmic
reticulum stress, that was accompanied by decreased levels of uterine cbs mRNA in both
hyperhomocysteinemic heterozygous (fertile) and homozygous (non fertile) females. Our
results indicate that cbs -/- female infertility is a consequence of uterine failure and
demonstrate that uterine endoplasmic reticulum stress and cbs expression are not
determinant of infertility suggesting that uterine dysfunction is a consequence of either
hyperhomocysteinemia, or other factor(s) in the uterine environment of cbs -/- animals. In
summary, these studies demonstrate the potential importance of homocysteine levels for
uterine handling of embryos.
by guest on March 17, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

3
In epidemiological and meta-analysis studies, hyperhomocysteinemia has been found to be
a risk factor for placental mediated diseases, such as preeclampsia, spontaneous abortion
and placental abruption (1, 2). In case- control studies, it has been reported that
homocysteine levels are frequently elevated in pregnancies complicated by preeclampsia,
or umbilical placental vascular disease over those present in the normal pregnancy group
(3-5).
Homocysteine is a non-protein sulfur amino acid whose utilization through the
trans-sulfuration pathway is its main metabolic fate, requiring the activity of cystathionine
β-synthase. Alternatively Hcy can be converted to methionine through the remethylation
pathway. Two metabolic reactions using different methyl donors are known: one uses
betaine and the reaction is catalyzed by betaine: homocysteine methyltransferase, and the
other catalyzed by 5-methyltetrahydrofolate: homocysteine methyltransferase uses as donor
5-methyltetrahydrofolate (6). These metabolic pathways are expressed in different tissues
during development. Thus, cystathionine β-synthase is expressed in liver and decidual
tissue while methionine synthase is expressed in all embryonic tissues, being the liver the
primary site of activity for these enzymes (7, 8). Mice deficient in cbs enzyme develop
hyperhomocysteinemia and provide an extraordinary tool to evaluate its effects during
pregnancy. Embryonic development of homozygous animals lacking the enzyme appears to
be normal (9) but there is an increased mortality during postnatal development that results
in the death of 90% of the mice by day 21. Animals surviving this critical period usually
become adults, but while males are fertile females are not. The availability of these females
allows to evaluate the role of hyperhomocysteinemia in female infertility and to define the
molecular and cellular events that may be involved. The endeavour to characterize those
events is the basis of the present report.
by guest on March 17, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

4
RESULTS
Reproductive Phenotype of cbs Female Mutants. In a continuous six-month mating
study, males lacking cbs were normally fertile (Table 1), as judged by test matings with
known fertile females during which they produced litters in due time, with an average litter
size of 6 pups. On the other hand, females lacking cbs showed a significantly decreased
number of pups per litter (P< 0.001), since five homozygous females had only two born
pups, that died soon after birth, compared to 39 for heterozygous cbs females (Table 1).
Thus, the homozygous cbs females exhibit infertility despite the detection of vaginal plugs
on multiple occasions, demonstrating that the infertility was not caused by any impairment
of sexual behaviour. When exploration of the hypothalamic/pituitary-ovarian axis was
assessed, after mating females with vasectomized mice, it revealed significantly higher
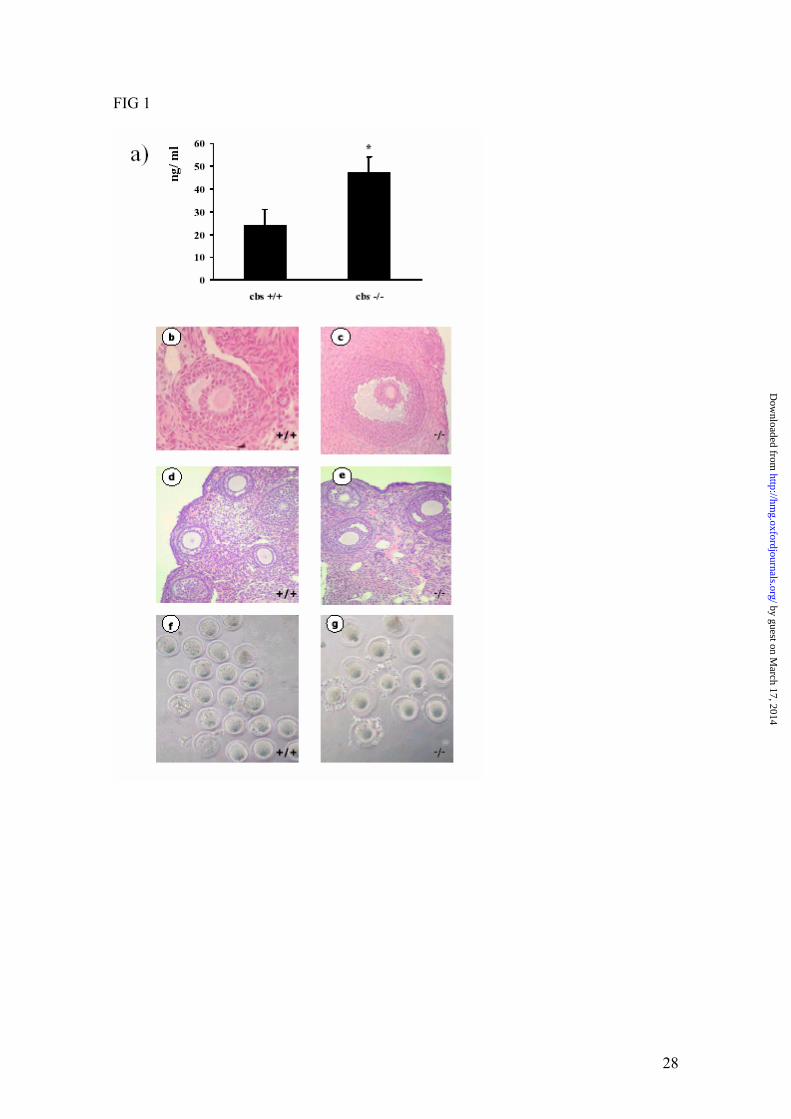
plasma progesterone levels 6 days after mating in pseudopregnant KO females than in their
normal counterparts (Fig. 1a), suggesting that the hormonal milieu in the -/- animals is not
normal. Consequently, we next analyzed the ovaries to determine if any defects in ovarian
structure and function could be indicative of ovarian failure. We found no obvious
histological abnormalities in cbs KO ovaries after normal estrus (Fig. 1 b and c), or after
superovulation experiments (Fig. 1 d and e). However, although morphologically normal
corpora lutea were present in the ovaries of stimulated cbs -/- females, suggesting that
follicles responded to the ovulatory surge of hCG, there was a decreased number of
developed follicles with respect to wild type females indicating a diminished response to
pregnant mare serum gonadotropin. Histochemical analysis of ovaries from superovulated
females also showed an increased oil red O staining of lipids in the ovarian corpora lutea of
cbs KO relative to those of wild-type animals (data not shown). While the cause for this
difference in lipid accumulation in response to superovulation remains unknown it appears
by guest on March 17, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

5
to suggest that no defect on cholesteryl ester storage, necessary to sustain adequate steroid
hormone production for pregnancy, underlies female cbs KO infertility. When the estrus
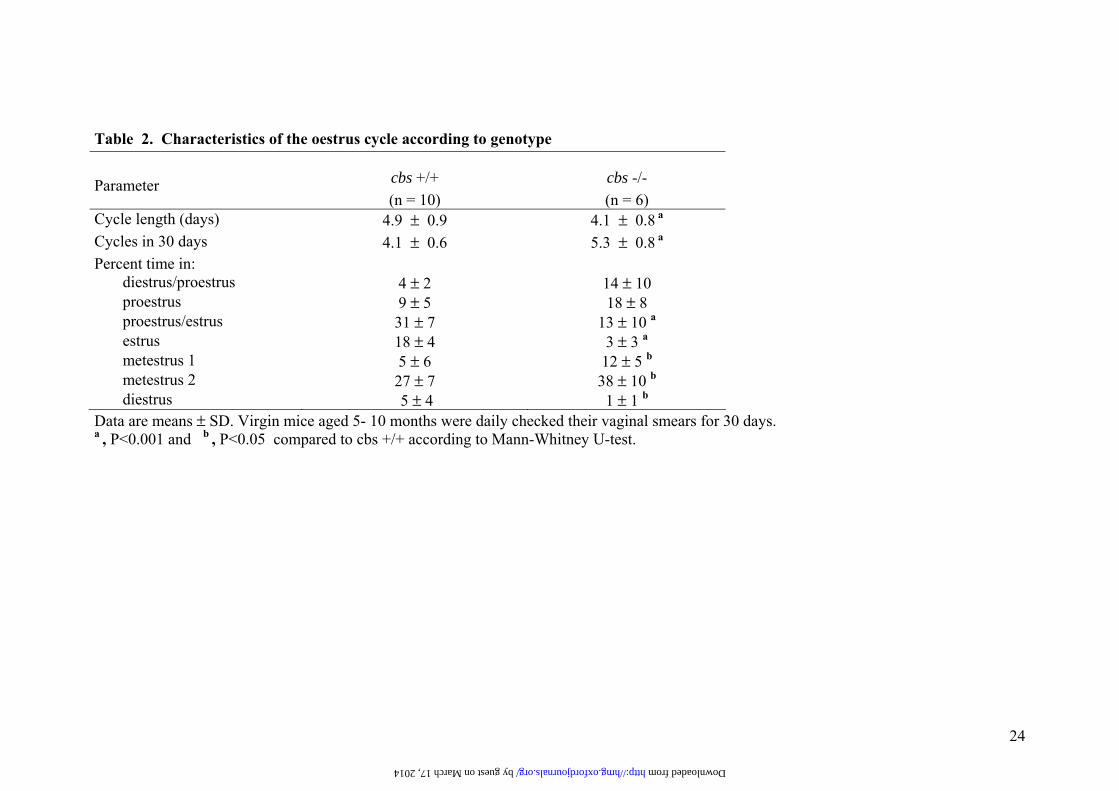
cycle was studied we detected a shortened and irregular estrus cycle in infertile cbs KO
females (Table 2), with a decreased time of estrus and diestrus periods, and a prolonged
metestrus. However, these differences in estrus cycle had no effect over the number of
oocytes ovulated during normal estruses, although the yield was significantly lower when
cbs deficient females were superovulated (Table 3). Oocytes presenting morphological
alterations were absent in females of both genotypes in pseudopregnancy experiments (data
not shown) and represented an equal proportion (8% of total oocyte count) in
superovulation experiments for both genotypes. No differences of collected oocytes were
observed between the two genotypes attending to the presence of polar extrusion body
either during normal estrus (58 ± 7 % for control vs 60 ± 7 for cbs deficient mice) or
superovulation (36 ± 6 % for control vs 40 ± 7 for cbs deficient mice) experiments. No
differences regarding internal diameter of oocytes showing polar body were observed
between the two genotypes (85 ± 3 and 84 ± 4 μm for control [n= 62] and cbs deficient [n=
32] mice, respectively) in pseudopregnancy and supervolulation experiments. However, we
observed that the perivitelline space was higher in cbs deficient than in control oocytes in
both types of experiments (Fig. 1 f and g).
Exploration of Pregnancy. Since there were no apparent differences between normal and
cbs KO females with respect to the extent of normal ovulation and number of oocytes
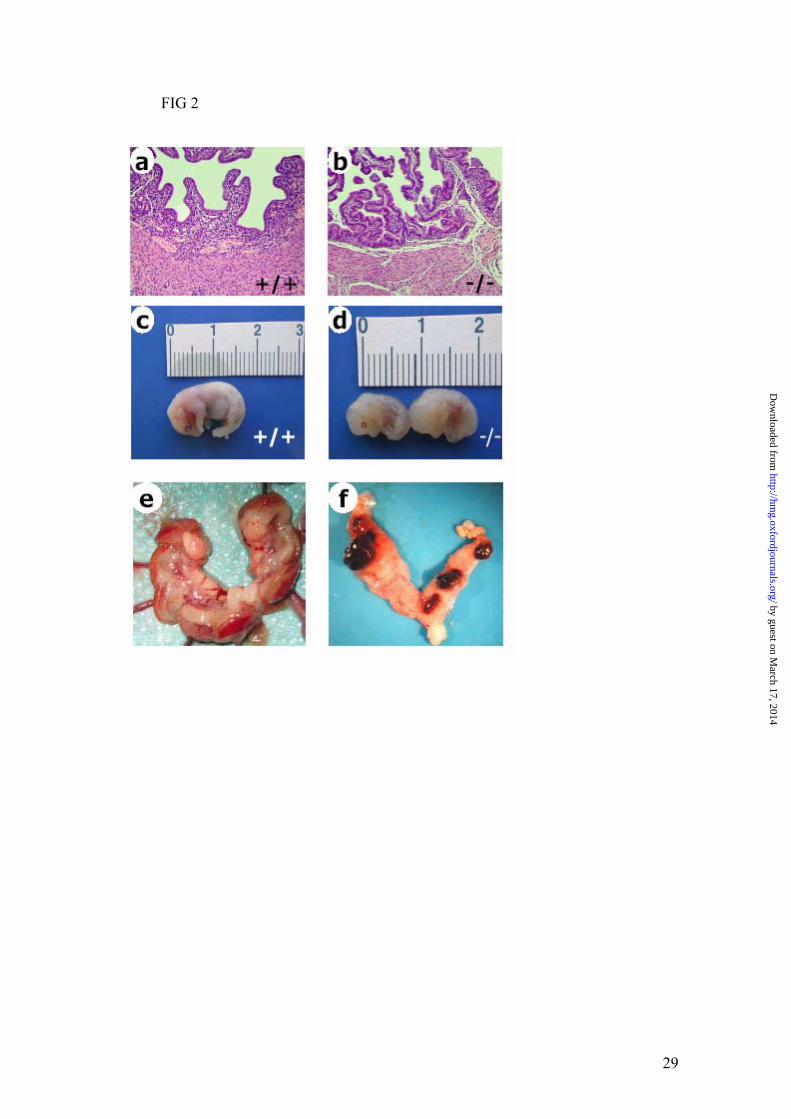
harvested after natural mating, we analysed uteri of gravid females at the 18th day
postmating (Table 4). A striking significant decrease in mass of gravid uterus was found in
cbs -/- compared to wild type females. In addition, there was a dramatic decrease in the
by guest on March 17, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

6
percentage of surviving foetuses in homozygous pregnant females despite a similar number
of implantation sites. The similar number of implantation sites (Table 4) observed in wild-
type and cbs deficient mice supports the notion that the minimal morphological alterations
of cbs-KO oocytes are not responsible for cbs-deficient female infertility. However,
histological evaluation of uterus (Fig. 2 a and b) disclosed a normal endometrium, muscular
layer, and perimetrium. The decrease in gravid uterine weight was accompanied by
significant decreases in placental and foetal weights (Table 4 and Fig 2 c and d) during cbs
KO pregnancies when compared to control animals. Cbs KO embryos showed a Theiler
Stage of 24-25 (Fig 2 c and d) compared to the 26 of wild-type embryos. Histological
assessment of placentas disclosed some abnormalities 18 th day placentas from cbs -/-
when compared to wild-type females (Fig. 3 a and b). The thickness of the decidual layer
was reduced in placentas from cbs -/- females, indicating reduced trophoblast invasion in
these females. The labyrinthine zone was enlarged while the junctional zone was smaller in
placentas from cbs deficient females. That these changes are an unlikely result from lack of
cbs expression in placental tissue of cbs -/- embryos would appear to be indicated by the
very low levels of cbs transcripts detected in placentas from normal animals by
semiquantitative RT-PCR (data not shown). Placental cbs expression was found to be even
lower than uterine expression (see below), suggesting that cbs may be not a major
determinant of placental function at the moment studied.
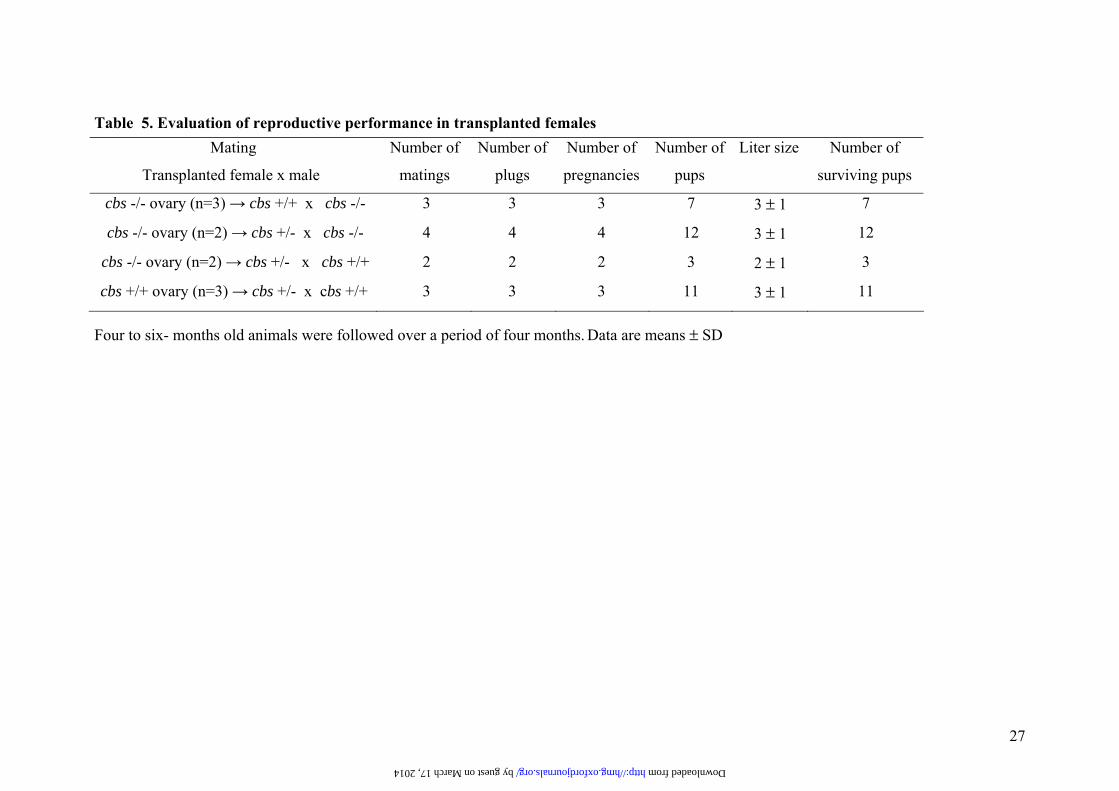
To further discriminate between uterine or placental contributions to the pregnancy failure
observed in cbs deficiency, ovary transplantation experiments were performed. The data
obtained from these experiments are shown in Table 5. Once the cbs -/- ovary was
transferred to the environment of a wild type or heterozygous animal, both the size of the
offspring and the percentage of surviving animals were indistinguishable from those of
by guest on March 17, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

7
female recipients of cbs +/+ ovaries, used as controls of the surgical procedure. No
significant differences in placental function could be drawn to placental genotype as the
average offspring size was similar for recipients of +/+ ovaries mated with +/+ males (+/+
placentas), and recipients of cbs -/- ovaries mated with either cbs -/- (-/- placentas) or wild-
type males (+/- placentas). In addition, the histological abnormalities observed in cbs -/-
placentas at day 18 were recovered when these were obtained from mice obtained by cbs -/-
ovary transferred to wild-type recipients and mating to homozygous males (Fig 3c). Taken
together, the low placental cbs transcript levels detected and more importantly the ovary
transplantation experiments demonstrate that cbs placental expression are dispensable in
order to get pregnancy success, and corroborates that the oocytes produced by the cbs
deficient ovary are quite functional since the liter size was identical between both
genotypes (Table 5). Altogether our results indicate that uterine failure in cbs -/- females is
likely the main cause of infertility.
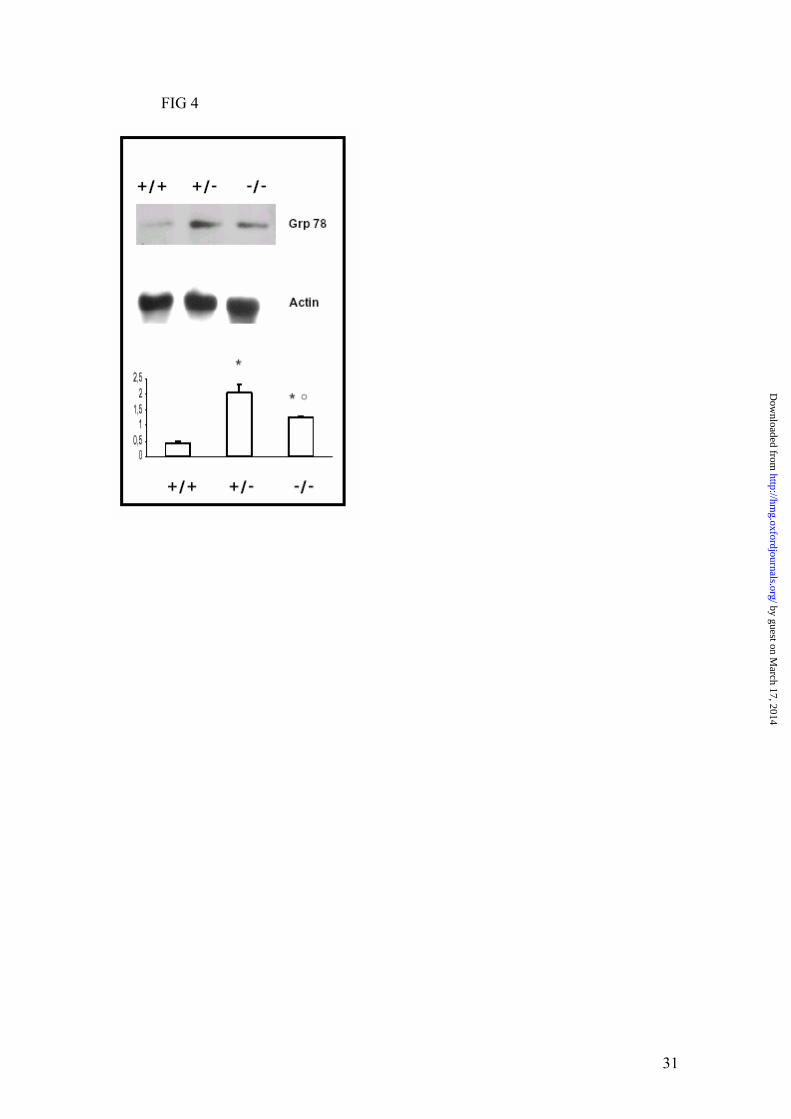
Uterine Endoplasmic Reticulum Stress. To evaluate if uterine dysfunction could be
caused by ERS, Western blotting analysis of Grp78 in uterine homogenates was carried out.
As shown in Fig 4 a, different expression levels of GRP78 were observed depending on the
underlying uterine genotype. Homozygous and heterozygous animals showed elevated
levels of the GRP78 protein considered being a marker of ERS. To explore whether ERS
could be related directly to decreased cbs expression, and since cbs mRNA levels were
below detection using Northern blotting analysis (data not shown), we performed
semiquantitative RT-PCR. As shown in figure 5a, expression levels of cbs mRNA in uteri
from normal pregnant females were significantly lower than those present in liver while, as
expected, no cbs transcripts could be detected in cbs -/- uteri. We also detected a marked
decrease in cbs mRNA levels in heterozygous animals with respect to controls (Fig 5c).
by guest on March 17, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

8
These data indicate that uterine ERS is accompanied by decreased cbs expression in both
cbs +/- and -/- animals. However, whether or not there is any causal relationship between
ERS and decreased cbs expression, uterine stress can not by itself be the cause of infertility
since cbs +/- females are fertile despite the presence of ERS to levels similar of those
present in cbs -/- infertile females. Our data also indicate that uterine cbs expression may be
dispensable for the function of this organ during pregnancy given that no major changes in
cbs transcription associated with pregnancy, as similar low levels of cbs transcripts were
detected in uteri from normal pregnant and non pregnant females (Fig 5b). This view
appears also to be supported by the fact that some KO females are able to deliver in the
absence of CBS expression (Table 1). Taken together our data indicate that cbs -/- uterine
dysfunction during pregnancy does not likely result from lack of cbs expression in this
organ. Instead, it appears to be caused by alterations of the internal milieu in these animals
making hyperhomocysteinemia alone, or together with some other cbs -/- induced
alteration, a good candidate for uterine failure during pregnancy in this animal model.
by guest on March 17, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

9
DISCUSSION
The present manuscript attempts to deepen our understanding of female infertility in
cbs deficient mice. Using diverse observations such as mating monitorization, ovulatory
patterns, histological studies, ovary transplantation, and molecular biology analysis, cbs-
KO female infertility may be assigned to a failure in uterine handling of embryos.
Female infertility is a complex process in which several organs such as the
hypotalamus, hypofisis, ovaries, uteri or embryonic placenta (10, 11) could be involved.
Our exploration of the coordinate action of the hypothalamus/pituitary-ovarian axis through
the induction of pseudo gestation indicates that cbs KO females have an impaired response
at this level since cbs -/- females showed an increased production of progesterone with
respect to control animals (Fig.1) suggesting that corpus luteum is longer maintained and/or
progesterone is poorly removed in these animals. Estrous cyclicity is also a reflection of the
function of the hypothalamic/hypophyseal-ovarian axis. The extension of metestrous 1 and
2, phases characterized by the presence of leukocytosis associated with cornified cells, may
indicate a normal production of estradiol but a resistance to luteolytic rupture (12). These
data are in agreement with the observation of higher progesterone levels in pseudopregnant
cbs KO females (Fig.1), and suggest an unbalanced level of estradiol versus progesterone
that could induce the observed shortening of estrous cycle by eliciting the gonadotropin
surge. Indeed, the observation of normal numbers of maturing follicles in all stages of
development strongly suggests an adequate synthesis of estradiol in cbs KO females.
Several of our observations, such as number of oocytes harvested after natural mating,
normal histology of the ovaries, and restoration of normal pregnancies after ovary
transplantation experiments, are indicating a correct structural organization and function of
this organ. Moreover, the ovaries by themselves, although influencing the female
by guest on March 17, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

10
reproductive performance are not the cause for cbs KO infertility based on data obtained
from ovary transplantation experiments into normal or mild hyperhomocysteinemic
recipients. Despite this conclusion, the insufficient response to the superovulation challenge
and the abnormal estrus cycles are indicative that ovaries are susceptible to environmental
cues present in cbs KO mice such as hyperhomocysteinemia. While it may not be the cause
of the pregnancy failure, it may affect uterine receptivity and pregnancy maintenance. The
latter observations may have potential clinical implications, since women experiencing
metabolic problems similar to those found in cbs -/- females, and undergoing artificial
reproductive technology, may have to suffer the inconveniences of having to collect an
adequate number of oocytes to use for in vitro fertilization and more difficulties to
synchronize hormonal therapy.
A candidate target organ to develop problems upon cbs loss of function is the
placenta, since placental abruption in humans is associated with hyperhomocysteinemia (1,
2). Our experimental data indicate that in cbs KO females there is a decrease of placental
mass that correlates with the presence of weakened layers on histological observation.
However, the occurrence of normal pregnancies in recipients of cbs -/- ovaries mated with
cbs -/- males, where placenta belongs to cbs -/- genotype, indicates that placental
abnormalities detected in cbs KO females are not a consequence of embryo cbs deficiency,
and that cbs heterozygosity of the placental donor cell genome is not critical for foetal
survival. Indeed, the normal development of -/- embryos and the normal liter size in
recipients of cbs -/- ovaries with respect to control animals points to the cbs -/- uterus as the
site of primary failure. The differences in weight of gravid uteri between wild type and
homozygous cbs deficient mice at day 18th of gestation and the striking differences in the
percentages of surviving foetuses are all indicative of an important problem concerning the
by guest on March 17, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

11
uterus. In addition, neither fertilization nor embryo implantation appear to be substantially
compromised in cbs -/- females, as suggested by the existence of similar numbers of
implantation sites between normal and mutant animals (Table 4). However, the existence of
a complicated implantation can be inferred by the appearance of a delayed embryological
stage of cbs -/- embryos (Fig. 2d). Likewise, data from ovary transplantation into normal or
cbs +/- females confirms that mild hyperhomocysteinaemia does not preclude successful
fertilization of oocytes by spermatozoa and suggest it is not the cause for cbs-KO female
infertility.
Deciphering the molecular events involved in reproductive physiology is an
interesting arena where transgenic and knockout mice are very useful models. Thus, it has
been found that defects in preantral and antral follicle growth associate with different
molecules, and molecules implicated in ovulation and corpus luteum have been
characterized (10). Implantation failure has been associated with the absence of
progesterone receptor A and gp130 – a signalling mediator of LIF. Improper
decidualization was found in mice lacking IL-11Rα. Both phenomena were reported in
COX2 and in cannabinoid receptor KO animals (11). Taking into consideration this
scenario, our data provide a new molecular paradigm in which normally implanted, well-
decidualized mice die from a failure in uterine handling due either to: i) a primary uterine
defect caused by total absence of cbs expression in this organ or ii) uterine failure
secondary to internal milieu alterations in these mice. Although we cannot exclude the first
possibility, the fact that uterine cbs mRNA expression levels are very low -at least two
orders of magnitude lower than cbs liver transcript levels-, that there are no major changes
during pregnancy and that some females are able to deliver would suggest that cbs
by guest on March 17, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

12
expression may be dispensable for uterine function. That this assumption could be correct
appears to be sustained also by the fact that lower than normal levels of uterine cbs mRNA,
like those present in cbs +/- females, are compatible with uterine reproductive fitness in
these animals. Therefore we suggest that uterine failure is secondary to alterations in the
internal milieu of cbs -/- females. Severe hyperhomocysteinemia is a good candidate to be
the cause underlying uterine malfunctioning in homozygous cbs deficient mice. A potential
mechanism for uterine dysfunction mediated by hyperhomocysteinemia is likely to be
related with an impairment of the endothelial-dependent vasodilatation response, since it
has been reported that arteries of pregnant mice are more sensitive to the effects of
increased homocysteine than arteries from nonpregnant mice (13). Our data also indicate
the existence of uterine ER stress in both heterozygous and homozygous deficient mice,
since Grp78 (fig 4a) was increased without alteration of Grp94 (data not shown). It has
been claimed that sensitive organs, developing ER stress, firstly increase the former
chaperone (14) what would classify uterus in this category. However, the marked increase
of Grp78, as indicative of ER stress, in heterozygous uterus of fertile females indicates that
this phenomenon is not relevant to the process of uterine handling of embryos, and that
moderate levels of homocysteine in vivo (around 15 μM) are at least as effective as severe
hyperhomocysteinemia (around 200 μM) in arising intracellular signalling. An observation
already found by other authors in other cell types (15).
Mammalian female fertility depends on complex interactions between the reproductive
organs and their environment. Since liver is the primary site of cbs activity (16) its
functional loss could, by inducing alterations of the internal milieu (i.e.
hyperhomocysteinemia), be the underlying cause of cbs KO uterine dysfunction. This
by guest on March 17, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

13
would imply the potential existence of a defective hepatic-uterine axis in cbs KO mice, just
as a defective hepatic-ovarian axis has been suggested to underlie female infertility caused
by abnormal HDLs in the SR-B1 KO mice (17). In addition, considering that the basal
metabolic rate per gram of body weight—the mass-specific rate—is seven times greater or
even higher in mice than in humans (18), the effects observed in mice at the level of 200
μM homocysteine may be observed in humans at the level of around 20 μM. This level can
be easily presented in some common mutations of CBS gene in the human population (19,
20) and whose levels may be aggravated in presence of some environmental habits (21).
Therefore, our data could contribute to understand the human fertility failure associated
with hyperhomocysteinemia and thus may help to overcome it. Although additional
mechanistic studies are necessary, the current data unequivocally establish that
hyperhomocysteinemia is important for normal uterine embryonic development, and
female fertility in mice.
by guest on March 17, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

14
MATERIALS AND METHODS
Animals. Generation of cbs-deficient mice was as described (9). Heterozygous and
homozygous cbs 5- to 6-week-old mice were used in natural matings. CBS-deficient mice
(mixed C57BL/6J x 129Ola background) were housed and fed a normal chow diet and
drinking water supplemented with 0.5% (wt/wt) choline chloride (Sigma Chemical Co.,
Madrid, Spain). All animal studies were approved by the Ethical Committee for Animal
Research of the University of Zaragoza.
PCR Genotyping. Three oligonucleotides were used in PCR amplification of tail DNA to
detect both the endogenous and altered cbs genes simultaneously. The sense primer was 5'-
cbs (GAAGTGGAGCTATCAGAGCA). Downstream primers were neo
(GAGGTCGACGGTATCGATA) and 3'-cbs (CGGATGACCTGCATTCATCT) specific
for the endogenous and altered cbs genes, respectively. PCR amplification results in a 0.5-
kb band from the wild-type gene and a 0.4-kb band from the knockout gene. PCR
amplification was carried out as follows: 94°C 1 minute; 94°C 20 seconds, 59°C 30
seconds, 72°C 30 seconds for 45 cycles followed by a 5-minute elongation period at 72°C.
Estrous Cycles Analysis and Fertility Assessment. Estrous cycles were monitored by
daily vaginal smears from mutant and control mice over a period of 1 month. A small drop
of sterile saline was flushed into the vagina and then dried on a slide. Cells were fixed in
methanol for 40 s, stained with 0.8% eosin for 40 s and 0.2% azur B for another 40 s, and
rinsed to remove excess stains. Smears were taken daily between 9 and 10 a.m. Smears
were evaluated by two experienced observers blinded to the genotype of the mice (12). To
assess fertility of female mice, virgin females were housed continuously with wild-type or
mutant males, and numbers of litters and pups were counted for 6 months.
Superovulation, Pseudopregnancy, Hormonal Assays and Oocyte Analysis.
Superovulation was induced in immature (22–23 days old) mice by i.p. injections of 5 units
of pregnant mare’s serum gonadotropin (Folligon; Intervet, Boxmeer, The Netherlands)
followed by 5 units of human chorionic gonadotropin (Chorulon, Intervet) 48 h later (22).
Pseudopregnancy was induced by mating (confirmed by inspection of vaginal seminal
by guest on March 17, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

15
plug) with vasectomized males and estimation of serum progesterone six days after.
Progesterone concentrations were assayed with an analyzer using a commercial kit
(IMMULITE 2000, Diagnostic Products Corporation, Los Angeles, CA) validated for
mouse sera. In both superovulation and pseudopregnancy experiments, oviducts were
collected. Once recovered from the oviduct, the cumulus–oocyte complexes were treated
with 3 mg/ ml hyaluronidase in M2 medium for 5 min at room temperature to liberate the
oocytes. These were washed three times with fresh medium to remove hyaluronidase and
transferred to a drop of M2 medium covered with paraffin. A complete in fresh observation
of specimens utilising a Leica Wild M650 binocular microscope was achieved rotating
them. Images were captured using a Canon digital camera. Morphometric analyses were
performed using NIH Image software conveniently calibrated with micrometric ocular.
Those oocytes presenting morphological alterations were considered as abnormal (22).
Each sample was assayed for number of oocytes collected, presence of oocytes with polar
body extrusion and the internal oocyte diameter.
Ovary Transplantation. Donor cbs KO or control wild-type mice of similar age and from
the same breeding (5–6 weeks of age) were sacrificed by cervical dislocation, and the
ovaries were removed. Ovaries were maintained in PBS until required for transfer. Hosts
were anesthetized with 2.5% Avertin (intraperitoneally), and ovary transfer surgery was
performed as described previously (22). Basically, ovaries of the host mice were carefully
dissected from the surrounding ovarian bursa. Half ovary from the donor mice was then
inserted into the ovarian bursa. Transplanted hosts were mated with cbs KO or wild-type
males 2 weeks after surgery and tested for fertility. The offspring was genotyped as
described above.
Presence of Sperm in the Itsmus and Embryo Implantation Analysis. Twelve hours
after observation of vaginal seminal plug, sperm from itsmus was flushed into phosphate
buffered medium and semiquantitative assessed. At 18 days postcoitum pregnant mothers
were anesthetized with 2.5% avertin and sacrificed. Ovaries and uterus were harvested.
Once the amniotic sacs and placenta were removed, the gravid uteri were weighted and
prepared for sectioning as described below. Placentas and foetuses were also weighted.
by guest on March 17, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

16
Live and dead foetuses were scored and the number of implant sites analyzed after
immersion of uteri in 0.5% ammonium sulphide solution for 10 min (23).
Histology. Paraffin sections (4 μm) of cbs and WT tissues were stained with hematoxylin
and eosin.
Assesment of cbs Transcript Levels by Semiquantitative RT-PCR. At the moment of
sacrifice, organs were obtained and quickly frozen in liquid nitrogen. Total RNA from liver
or uteri from 18 days of gestation animals was extracted using Trigent reagent MRC
(Sigma, Madrid, Spain) (24). Semiquantitative RT-PCR was performed using the enhanced
avian HS RT-PCR KIT (Sigma), according to manufacturer’s instructions. Reverse
transcription was performed by the two-step protocol using for cDNA synthesis 2.5 μg of
template RNA in a final reaction volume of 20 μl. One μl of the cDNA obtained was then
PCR amplified with cbs or β-actin specific primers. For cbs we designed primers CBS-1
GAA CAC CCC TAT GGT CAG AA and CBS-2 GGA TTT TCG TTC TTC AGT CG
which do not amplify genomic sequences and produce a 388 bp amplification product. The
470 bp PCR product from actin was obtained using primers Actin-1 GAT CAT GTT TGA
GAC CTT CAA CAC C and Actin-2 AGT TTC ATG GAT GCC ACA GGA TTC C. Actin
and cbs amplifications were performed separately to avoid amplicon interference during
amplification. PCR was carried out at 94º for 45sc, annealing at 62º for 45sc, and extension
at 72º for 45 sc. After 29 cycles of amplification the material was subjected to a final
extension step at 72º for 5 min. PCR products were resolved by electrophoresis in 1%
agarose gels and confirmed by DNA sequencing.
Western-Blot. Uterine homogenates from cbs KO or control wild-type mice were analyzed
to test the unfolded protein response. 50 μg of protein were loaded onto a gradient (4-24%)
SDS-polyacrylamide gel. Gel was transferred to PVDF membranes (Millipore, Madrid).
Protein bands were detected using rabbit polyclonal antibodies against mouse GADD 153
(Santa Cruz Biotechnology, Santa Cruz, CA, USA) and Grp78 (Stressgen, Victoria,
Canada) followed by detection with a secondary antibody anti-rabbit IgG peroxidase
conjugate (GE Healthcare) and chemiluminiscence (GE Healthcare, Madrid). Equal loading
was confirmed by using an anti- actin antibody obtained from Sigma. Membranes were
by guest on March 17, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

17
exposed to ECL film (GE Healthcare) and films analyzed using a laser LKB 2202
densitometer (GE Healthcare).
Statistical Analysis. Data were analyzed by using either a two-tailed, unpaired Student’s t
test or an unpaired, nonparametric Mann-Whitney U-test test using Instat 3.02 for
Windows software (GraphPad, S. Diego, CA, USA). Values are presented as means ± SD.
by guest on March 17, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

18
ACKNOWLEDGEMENTS
This research was supported by grants CICYT (SAF2004-08173-C03-02), FISS
01/0202; Redes DGA (A-26) and FISS de investigación cooperativa C03/01, Fundación
Española del Corazón and R.C., S.A., M.A.N. and M.A.G.G. were recipient of DGA,
FEGA-FEOGA and PROMEP fellowships. We thank Jesús Navarro, Angel Beltrán, Jesús
Cazo, Carmen Navarro and Clara Tapia from Unidad Mixta de Investigación for their
invaluable help in maintaining animals and Rosario Puyó for her technical assistance. We
thank Dr García de Jalón for helpful suggestions.
The authors declare no conflict of interest.
by guest on March 17, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

19
REFERENCES
1. Ueland, P.M., Nygard, O., Vollset, S.E. and Refsum, H. (2001) The Hordaland
Homocysteine Studies. Lipids, 36, S33-39.
2. Ray, J.G. and Laskin, C.A. (1999) Folic acid and homocyst(e)ine metabolic defects
and the risk of placental abruption, pre-eclampsia and spontaneous pregnancy loss:
a systematic review. Placenta, 20, 519-529.
3. Wang, J., Trudinger, B.J., Duarte, N., Wilcken, D.E. and Wang, X.L. (2000)
Elevated circulating homocyst(e)ine levels in placental vascular disease and
associated pre-eclampsia. Br. J. Obst. Gyn., 107, 935-938.
4. Nelen, W.L. (2001) Hyperhomocysteinaemia and human reproduction. Clin. Chem.
Lab. Med., 39, 758-763.
5. Daly, S., Cotter, A., Molloy, A.E. and Scott, J. (2005) Homocysteine and folic acid:
implications for pregnancy. Semin. Vasc. Med., 5, 190-200.
6. Selhub, J. (1999) Homocysteine metabolism. Annu. Rev. Nutr., 19, 217-246.
7. VanAerts, L.A., Poirot, C.M., Herberts, C.A., Blom, H.J., De Abreu, R.A., Trijbels,
J.M., Eskes, T.K., Peereboom-Stegeman, J.H. and Noordhoek, J. (1995)
Development of methionine synthase, cystathionine-beta-synthase and S- adenosyl-
homocysteine hydrolase during gestation in rats. J. Reprod. Fertil., 103, 227-232.
8. Thompson, H.R., Jones, G.M. and Narkewicz, M.R. (2001) Ontogeny of hepatic
enzymes involved in serine- and folate-dependent one-carbon metabolism in rabbits.
Am. J. Physiol. Gastrointest. Liver Physiol., 280, G873-878.
9. Watanabe, M., Osada, J., Aratani, Y., Kluckman, K., Reddick, R., Malinow, M.R.
and Maeda, N. (1995) Mice deficient in cystathionine beta-synthase: animal models
for mild and severe homocyst(e)inemia. Proc. Natl. Acad. Sci. U. S. A., 92, 1585-
1589.
10. Elvin, J.A. and Matzuk, M.M. (1998) Mouse models of ovarian failure. Rev.
Reprod., 3, 183-195.
by guest on March 17, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

20
11. Paria, B.C., Reese, J., Das, S.K. and Dey, S.K. (2002) Deciphering the cross-talk of
implantation: advances and challenges. Science, 296, 2185-2188.
12. Nelson, J.F., Felicio, L.S., Randall, P.K., Sims, C. and Finch, C.E. (1982) A
longitudinal study of estrous cyclicity in aging C57BL/6J mice: I. Cycle frequency,
length and vaginal cytology. Biol. Reprod., 27, 327-339.
13. Powers, R.W., Gandley, R.E., Lykins, D.L. and Roberts, J.M. (2004) Moderate
hyperhomocysteinemia decreases endothelial-dependent vasorelaxation in pregnant
but not nonpregnant mice. Hypertension, 44, 327-333.
14. Outinen, P.A., Sood, S.K., Pfeifer, S.I., Pamidi, S., Podor, T.J., Li, J., Weitz, J.I. and
Austin, R.C. (1999) Homocysteine-induced endoplasmic reticulum stress and
growth arrest leads to specific changes in gene expression in human vascular
endothelial cells. Blood, 94, 959-967.
15. Werstuck, G.H., Lentz, S.R., Dayal, S., Hossain, G.S., Sood, S.K., Shi, Y.Y., Zhou,
J., Maeda, N., Krisans, S.K., Malinow, M.R. et al. (2001) Homocysteine-induced
endoplasmic reticulum stress causes dysregulation of the cholesterol and
triglyceride biosynthetic pathways. J. Clin. Invest., 107, 1263-1273.
16. Ratnam, S., Maclean, K.N., Jacobs, R.L., Brosnan, M.E., Kraus, J.P. and Brosnan,
J.T. (2002) Hormonal Regulation of Cystathionine beta -Synthase Expression in
Liver. J. Biol. Chem., 277, 42912-42918.
17. Miettinen, H.E., Rayburn, H. and Krieger, M. (2001) Abnormal lipoprotein
metabolism and reversible female infertility in HDL receptor (SR-BI)-deficient
mice. J. Clin. Invest., 108, 1717-1722.
18. Demetrius, L. (2005) Of mice and men.When it comes to studying ageing and the
means to slow it down, mice are not just small humans. EMBO reports, 6, S39-S44.
19. Lee, S.J., Lee, D.H., Yoo, H.W., Koo, S.K., Park, E.S., Park, J.W., Lim, H.G. and
Jung, S.C. (2005) Identification and functional analysis of cystathionine beta-
synthase gene mutations in patients with homocystinuria. J. Hum. Genet., 50, 648-
654.
by guest on March 17, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

21
20. Urreizti, R., Asteggiano, C., Bermudez, M., Cordoba, A., Szlago, M., Grosso, C., de
Kremer, R.D., Vilarinho, L., D'Almeida, V., Martinez-Pardo, M. et al. (2006) The
p.T191M mutation of the CBS gene is highly prevalent among homocystinuric
patients from Spain, Portugal and South America. J. Hum. Genet., 51, 305-313.
21. Kluijtmans, L.A., Young, I.S., Boreham, C.A., Murray, L., McMaster, D., McNulty,
H., Strain, J.J., McPartlin, J., Scott, J.M. and Whitehead, A.S. (2003) Genetic and
nutritional factors contributing to hyperhomocysteinemia in young adults. Blood,
101, 2483-2488.
22. Hogan, B., Beddington, R., Costantini, F. and Lacy, E. (1994) Manipulating the
mouse embryo. In. Cold Spring Harbor Press, Plainview, New York, USA, pp. 130-
188.
23. Manson, M. and Kang, Y.J. (1994) Principles and Methods of Toxicology. In.
Raven Press, New York, USA, pp. 989-1037.
24. Calleja, L., Paris, M.A., Paul, A., Vilella, E., Joven, J., Jimenez, A., Beltran, G.,
Uceda, M., Maeda, N. and Osada, J. (1999) Low-cholesterol and high-fat diets
reduce atherosclerotic lesion development in ApoE-knockout mice. Arterioscler.
Thromb. Vasc. Biol., 19, 2368-2375. by guest on March 17, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

22
FIG 1. Histological and functional analysis of ovaries. a) Plasma progesterone
concentrations from pseudopregnant females at 6 day after mating with vasectomized
males, (n = 10 and 6 for wild type and cbs -/-, respectively; *P < 0.05). Hematoxylin and
eosin-stained sections of adult wild-type (b) and cbs -/- (c) ovaries at x400; showing a
follicle. Histological sections of a representative ovary from an immature wild-type female
(d) and from an immature cbs -/- female (e), both after superovulation. Phase-contrast
microscopy of from wild-type (f) or cbs -/- (g) oocytes devoid of cumulus.
FIG 2. Uterine histology in cbs mice. Hematoxylin and eosin-stained sections of wild-
type (a) and cbs -/- (b) uteri from day 18 of gestation at x100 magnification. Embryos were
collected from normal (c) or cbs -/- (d) at day 18 of gestation. Appearance of uterine horns
carrying embryos at 18th day of gestation from normal (e) or cbs -/- (f) with resorbing sites
stained with ammonium sulphide.
FIG 3. Placental histology in cbs mice. Hematoxylin and eosin-stained placental sections
of (a) wild-type, (b) cbs -/- and (c) cbs-/- placenta obtained from a cbs-/- ovary transferred
to a wild-type female recipient taken at day 18 of gestation and observed at x40. The
different layers of the mouse placenta are shown (d, decidual; j, junctional; l, labyrinthine).
FIG 4. Characterization of ER stress according to genotype in uterus. Western blot of
Grp78 according to genotype. * and º P<0.05 vs wild-type and +/-, respectively; according
to one-way ANOVA.
FIG 5. Expression of cbs gene in function of genotype and physiological conditions. A)
Expression of cbs gene in uterus and liver. B) Expression of uterine cbs gene in pregnant
(Up) and non-pregnant (Unp) mice. C) Expression of uterine cbs gene according to
genotype.
by guest on March 17, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

23
Table 1. Evaluation of reproductive performance in female according to genotype
Four to six- months old animals were followed over a period of six months. a , P<0.001 vs cbs -/- x cbs -/- according to one-way ANOVA.
Mating Female x male
Number of matings
Number of plugs
Number of pregnancies
Number of pups
Litter size
Number of surviving pups
cbs -/- (n=5) x cbs -/- 10 10 1 2 0.4 ± 1 0 cbs +/- (n=5) x cbs -/- 10 9 7 39 6 ± 1a 23 cbs +/- (n=5) x cbs +/- 6 6 6 39 6 ± 2a 24 cbs +/+ (n=5) x cbs +/+ 6 6 6 49 8 ± 2a 46
by guest on March 17, 2014 http://hmg.oxfordjournals.org/ Downloaded from
24
Table 2. Characteristics of the oestrus cycle according to genotype Parameter cbs +/+
(n = 10) cbs -/- (n = 6)
Cycle length (days) 4.9 ± 0.9 4.1 ± 0.8 a Cycles in 30 days 4.1 ± 0.6 5.3 ± 0.8 a Percent time in:
diestrus/proestrus 4 ± 2 14 ± 10 proestrus 9 ± 5 18 ± 8 proestrus/estrus 31 ± 7 13 ± 10 a estrus 18 ± 4 3 ± 3 a metestrus 1 5 ± 6 12 ± 5 b metestrus 2 27 ± 7 38 ± 10 b diestrus 5 ± 4 1 ± 1 b
Data are means ± SD. Virgin mice aged 5- 10 months were daily checked their vaginal smears for 30 days. a , P<0.001 and b , P<0.05 compared to cbs +/+ according to Mann-Whitney U-test.
by guest on March 17, 2014 http://hmg.oxfordjournals.org/ Downloaded from

25
Table 3. Ovulation pattern according to genotype in experimental conditions Ovulation cbs +/+ cbs -/-
Spontaneous*
Eggs per mouse 8 ± 1 8 ± 1
Presence of sperm¶ + +
Hormonally induced
Eggs per mouse 28 ± 5 13 ± 2 a Data are means ± SD of three experiments. *Released oocytes were counted 0.5 days after mating to fertile males. ¶Semiquantitative assessment of sperm. A + indicates 2 -5 spermatozoids per field at 400x. a , P<0.02 according to Mann-Whitney U-test.
by guest on March 17, 2014 http://hmg.oxfordjournals.org/ Downloaded from

26
Table 4. Evaluation of pregnancy characteristics at gestation day 18 Mating
Female x male Gravid uterus weight∗
(g) Number of implantation
sites per mice Percentage of
surviving foetuses #Placenta weight
(g) Foetus
weight (g)
cbs +/+ x cbs +/+ (n= 4)
5.5 ± 1.6 6 ± 2 94 ± 6 0.17 ± 0.03 (n= 21)
0.82 ± 0.3 (n= 21)
cbs -/- x cbs -/- (n= 4)
2.1 ± 0.6 a 6 ± 1 18 ± 18 a 0.11 ± 0.03 a
(n= 8) 0.45 ± 0.2 a
(n= 8)
Data are means ± SD. ∗ The gravid uteri were weighted after removing the amniotic sacs and placentas. #Percentage of surviving foetuses was calculated as the ratio of surviving embryos referred to the number of implantation sites and expressed as %. a , P<0.02 according to Mann-Whitney U-test.
by guest on March 17, 2014 http://hmg.oxfordjournals.org/ Downloaded from
27
Table 5. Evaluation of reproductive performance in transplanted females
Four to six- months old animals were followed over a period of four months. Data are means ± SD
Mating
Transplanted female x male
Number of
matings
Number of
plugs
Number of
pregnancies
Number of
pups
Liter size Number of
surviving pups
cbs -/- ovary (n=3) → cbs +/+ x cbs -/- 3 3 3 7 3 ± 1 7
cbs -/- ovary (n=2) → cbs +/- x cbs -/- 4 4 4 12 3 ± 1 12
cbs -/- ovary (n=2) → cbs +/- x cbs +/+ 2 2 2 3 2 ± 1 3
cbs +/+ ovary (n=3) → cbs +/- x cbs +/+ 3 3 3 11 3 ± 1 11
by guest on March 17, 2014 http://hmg.oxfordjournals.org/ Downloaded from
28
FIG 1
by guest on March 17, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from
29
FIG 2
by guest on March 17, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

30
FIG 3
by guest on March 17, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from
31
FIG 4
by guest on March 17, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

32
FIG 5
by guest on March 17, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from




















