
Associations between Canine Male Reproductive Parameters ...
210
Associations between Canine Male Reproductive Parameters and Serum Vitamin D and Prolactin Concentrations by Adria Julianne Kukk A Thesis presented to The University of Guelph In partial fulfillment of requirements for the degree of Doctor of Veterinary Science in Population Medicine Guelph, Ontario, Canada ¤ Adria Julianne Kukk, December, 2011
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Associations between Canine Male Reproductive Parameters ...

Associations between Canine Male Reproductive Parameters and
Serum Vitamin D and Prolactin Concentrations
by
Adria Julianne Kukk
A Thesis presented to
The University of Guelph
In partial fulfillment of requirements for the degree of
Doctor of Veterinary Science in
Population Medicine
Guelph, Ontario, Canada
Adria Julianne Kukk, December, 2011

ABSTRACT
ASSOCIATIONS BETWEEN CANINE MALE REPRODUCTIVE PARAMETERS
AND SERUM VITAMIN D AND PROLACTIN CONCENTRATIONS
Adria Julianne Kukk Advisor: University of Guelph, 2011 Professor C.J. Gartley Maintaining reproductive health and diagnosing and treating conditions of
infertility in stud dogs is important in canine theriogenology. However, there is
still a great deal to be learned about reproductive physiology and factors that
affect reproductive organs and semen quality in dogs. This thesis is an
investigation of two factors in the male dog; serum 25-hydroxy Vitamin D
(25OHVD) and prolactin (PRL) concentrations, and their possible associations
with benign prostatic hyperplasia (BPH), prostate volume and/or sperm
morphology and motility characteristics.
28 (Vitamin D Study) and 29 (28 plus one for the Prolactin study) client
dogs of various breeds from the Ontario Veterinary College and Graham Animal
Hospital in Southwestern Ontario, Canada were enrolled in the study from March
to December 2009. Of these dogs 22 were successfully collected for semen.
BPH was diagnosed using prostate volume measured by ultrasound, as well as
clinical signs including blood in the ejaculate. Semen analysis was performed

using manual microscopic techniques for morphology and computer assisted
sperm analysis equipment for motility.
In the vitamin D study, no associations were found between BPH and
serum 25OHVD concentrations. In contrast, several sperm motility (motility,
progressive motility, beat cross frequency (BCF), distance average path (DAP),
curvilinear distance (DCL), linear distance (DSL), average path velocity (VAP),
curvilinear velocity (VCL) and straight line velocity (VSL), amplitude lateral head
displacement (ALH) and average orientation change (AOC)) and morphology
characteristics (percentage normal sperm, head defects and detached heads)
had desirable outcomes with 25OHVD concentrations between 120-180 nmol/l.
Using bivariable analysis, positive associations were observed with 25OHVD and
some semen quality characteristics from 4 to 8 years of age (motility, progressive
motility, BCF, DCL, VCL, ALH, AOC) and at transformed prostate volumes
smaller than or equal to 4.5 (motility, progressive motility, DCL, VCL, and normal
morphology) while negative associations of these semen parameters were found
at ages greater than 8 years and transformed prostate volumes greater than or
equal to 5.5. Head defects were negatively associated with 25OHVD. Vitamin D
may have an impact on spermatogenesis and normal sperm physiology that
warrants further research.
The prolactin study showed no statistically significant associations
between serum PRL and BPH and serum PRL and sperm motility characteristics.
However, two sperm morphology characteristics (percentage proximal droplets
and percentage midpiece defects) had significant negative associations with PRL

concentrations. Age interaction with PRL was also a factor in the percentage of
midpiece defects with desirable outcomes associated at 4 years of age compared
with older ages. Overall, undesirable outcomes occurred at PRL concentrations
less than 2.5 ng/ml. In conclusion, both 25OHVD and PRL may have important
roles in spermatogenesis and normal sperm physiology in the dog.

v
ACKNOWLEDGEMENTS The past few years have been an exciting and important part of my life. I
would not have been able to complete this great undertaking without the support
and knowledge of a great many people. First I would like to thank my advisor
Dr. Cathy Gartley for her wealth of knowledge in the field of theriogenology, for
her mentorship, her kindness and lastly, for her sense of humour to make me
laugh even on the greyest of days. Also, I would like to thank members of my
advisory committee: Drs. Tracey Chenier, Andria Jones, Stephanie Nykamp and
Heather Chalmers for their patience and expertise in guiding me through my
research and for their critical advice in pushing me to create a body of work I can
be proud of. For his statistical expertise, I am grateful to William Sears whose
help was invaluable in the analysis of my data.
For their friendship, support and encouragement during my D.V.Sc.
programme I would like to thank Drs. Rasa Levstein and Cyril Stephen, our
veterinary technicians Jim Rahn and Karen DiCaro, as well as my fellow
graduate students Drs. Mariana Diel De Amorim and Leslie Gonzalez. I would
also like to thank Dr. Walter Johnson for his encouragement and for sharing his
knowledge of bovine theriogenology. A special thank-you to Sally, Linda, Julie
and Karla in the Population Medicine office, and Mary Elliot for helping me with
the bureaucratic aspects of graduate studies and for making my life that much
easier.
To Dr. Carol Graham, all the staff at Graham Animal Hospital, and to all
our clients who agreed to enroll their dogs in this study, I extend a warm

vi
appreciation for their participation. Without them there wouldn’t have been any
data to analyse. I wish to also thank the Pet Trust for providing the funding to
make this study possible.
Lastly, I would like to thank two very special people in my life, my mother
Eva and my husband Anthony (Toncsi) for the much needed emotional support
during my graduate programme. Their patience and unending belief in me gave
me the strength I needed to achieve my goals.

vii
DECLARATION OF WORK PERFORMED
I declare that with the exception of the items listed below, all work presented in
this thesis was performed by me.
The Animal Health Laboratory, University of Guelph, performed urinalysis and
culture and sensitivities for urine, prostatic fluid and semen.
The Endocrinology Division of the Diagnostic Center of Population and Animal
Health, Michigan State University, performed the radioimmunoassay for Vitamin
D analysis.
The New Animal Science Laboratory, Washington State University, performed
the enzyme-linked immunosorbent Assay for Prolactin analysis.

viii
TABLE OF CONTENTS
CHAPTER ONE ………………………………………………………………… 1
INTRODUCTION, RESEARCH OBJECTIVES AND LITERATURE
REVIEW …………………………………………................................... 1
1. INTRODUCTION …………………………………………………….. 1
2. RESEARCH OBJECTIVES..………………………………………… 3
3. LITERATURE REVIEW.……………………………………………… 4
3.1 PHYSIOLOGY OF CANINE SPERMATOGENESIS AND
METHODS OF SEMEN ANALYSIS………………………… 4
3.1.1 Reproductive Physiology of the Testes and the Role of
Hormones and Local Factors In Male Reproduction….. 4
3.1.2 Spermatogenesis ………………………………………. 13
3.1.3 Factors Affecting Semen Quality.…………………… 16
3.1.4 Methods of Semen Evaluation……………………… 17
3.2 THE CANINE PROSTATE GLAND ……………………….. 31
3.2.1 Anatomy and Physiology of the Prostate Gland ……... 31
3.2.2 Pathophysiology of Benign Prostatic Hyperplasia…….. 33
3.2.3 Conventional Treatment of Benign Prostatic
Hyperplasia ……………………………………………….. 39
3.2.4 Ultrasonography of the Prostate.……………………….. 42
3.3 VITAMIN D AND ITS ROLE IN MALE REPRODUCTION.. 44
3.3.1 Vitamin D Metabolism ………………………………… 45
3.3.2 Vitamin D and the Prostate …………………………… 49
3.3.3 Role of Vitamin D in Sperm Production ……………… 54
3.4 PROLACTIN AND ITS ROLE IN MALE REPRODUCTION.. 56
3.4.1 Prolactin and Ovarian Physiology.……………………. 56
3.4.2 Prolactin and Male Physiology.………………………. 58
3.4.2.1 Prolactin and the Prostate.…………………………. 58
3.4.2.2 Prolactin, Male Fertility and Semen Quality ……… 60

ix
References ………………………………………………………… 67
CHAPTER TWO …………………………………………………………… 81
VITAMIN D, BENIGN PROSTATIC HYPERPLASIA, PROSTATE
VOLUME AND SEMEN PARAMETERS IN THE DOG………… 81
Abstract.……………………………………………………………... 81
1. Introduction……………………………………………………….. 82
2. Materials and Methods.………………………………………….. 84
3. Results.……………………………………………………………. 92
4. Discussion and Conclusions.…………………………………… 98
References ………………………………………………………….. 143
CHAPTER THREE.…………………………………………………………… 147
PROLACTIN, BENIGN PROSTATIC HYPERPLASIA,
PROSTATE VOLUME AND SEMEN PARAMETERS IN THE DOG.. 147
Abstract.………………………………………………………………... 147
1. Introduction …………………………………………………………. 148
2. Materials and Methods ……………………………………………. 151
3. Results ……………………………………………………………… 155
4. Discussion and Conclusions ……………………………………… 157
References.…………………………………………………………….. 178
CHAPTER FOUR …………………………………………………………….. 180
SUMMARY AND CONCLUSIONS ………………………………… 180
References……………………………………………………………. 184
APPENDICES ……………………………………………………………….. 185

x
LIST OF TABLES
Table 1.1 Selected Physical and Chemical Analyses of Prostatic Fluid in Dogs with Healthy Prostates.…………………………………………… 33 Table 1.2 Genes influenced by Vitamin D receptor ligands and their effects 48 Table 1.3 Prolactin values in Dogs (ELISA) ……………………………………65 Table 2.1 Technical Parameters for SpermVision™ CASA …………………111 Table 2.2 Classification scheme for determination of the presence of BPH by ultrasonography in 29 dogs, March-December 2009, Ontario, Canada ………………………………………………………….112 Table 2.3 Characteristics of the study population of 28 dogs, in Ontario, Canada, March-December 2009.……………………………………….113 Table 2.4 Characteristics of the study population of 22 dogs from whom semen was collected in Ontario, Canada March-December 2009…………...114 Table 2.5 Breed, Vitamin D concentration, age and fertility data on 22 dogs from whom semen was collected in Ontario, Canada, 2009…………115 Table 2.6 Mean serum Vitamin D concentration, with respect to type of diet, with associated test-statistic, p-value and confidence intervals………116 Table 2.7 Mean serum Vitamin D concentration, with respect to time of year of sampling, with associated test-statistic, p-value and confidence intervals……………………………………………………………………..117 Table 2.8 Mean serum Vitamin D concentration, with respect to number of hours spent outdoors at time of year of sampling, with associated test-statistic, p-value and confidence intervals…………………………………………118 Table 2.9 Mean serum Vitamin D concentration, with respect to BPH status, and associated test-statistic, p-value and confidence intervals……….119 Table 2.10 Univariable simple linear regression analyses of sperm motility parameters with serum 25-hydroxy vitamin D as explanatory variable, with associated coefficient of determination (R2), test statistic and p-value…………………………………………………………………120

xi
Table 2.11 Non-significant bivariable linear regressions of motility with explanatory variable regression coefficients (where applicable), coefficients of determination for the overall model (R2), test statistics, and the associated p-values..……………………………………………………………………121 Table 2.12 Three bivariable linear regressions of morphology parameters and associated coefficients of determination (R2), test-statistic and p-values………………………………………………………………..123 Table 3.1 Technical Parameters for SpermVision™ CASA …………………..166 Table 3.2 Classification scheme for determination of the presence of Benign Prostatic Hyperplasia (BPH) by ultrasonography in 29 dogs, March-December 2009, Ontario, Canada.………………………………167 Table 3.3 Breed, prolactin concentration, age and fertility data on 22 dogs from whom semen was collected in Ontario, Canada, 2009………….168 Table 3.4 Mean serum prolactin concentrations and confidence intervals with respect to benign prostatic hyperplasia status………………………….169 Table 3.5 Univariable analyses using simple linear regression modeling of semen motility parameters and prolactin with coefficients of determination (R2) of the overall model, test statistic and associated p-value………………………………………………………………….......170 Table 3.6 Three bivariable linear regressions of motility parameters with coefficients of determination of overall model (R2), test statistics, and associated p-values………………………………….............................171 Table 3.7 Univariable analyses of sperm morphology parameters and PRL with coefficients of determination (R2) of the overall model, test-statistic and associated p-values……………….………………………………………173 Table 3.8 Three bivariable linear regressions of morphology parameters, coefficients of determination (R2) of the overall model, test statistics and associated p-values…………………………………………………… 174 Table 3.9 Three non-significant bivariable linear regressions of proximal droplets with variable coefficients where PRL slope coefficients were significant, coefficients of determination for the overall model (R2) and associated p-values………………………………………………………………… 175

xii
LIST OF FIGURES
Figure 2.1 Predicted sperm motility as a response to normal serum 25OHVD range in dogs…………………………………………124 Figure 2.2 Progressive motility as a response to normal serum 25OHVD range in dogs………………………………………………...125 Figure 2.3 Predicted sperm motility in response to normal serum 25OHVD range at multiple ages……………………………………….126 Figure 2.4 Predicted sperm motility in response to normal serum 25OHVD range at various transformed prostate volumes…………..127 Figure 2.5 Predicted progressive motility in response to normal serum 25OHVD range at various ages………………………………..128 Figure 2.6 Predicted progressive motility in response to normal serum 25OHVD range at various transformed prostate volumes…..129 Figure 2.7 Predicted BCF in response to normal serum 25OHVD range at various ages……………………………………………………………130 Figure 2.8 Predicted DCL in response to normal serum 25OHVD range at various ages…………………………………………………………….131 Figure 2.9 Predicted DCL in response to normal serum 25OHVD range at various transformed prostate volumes……………………………….132 Figure 2.10 Predicted VCL in response to normal serum 25OHVD range at various ages……………………………………………………………..133 Figure 2.11 Predicted VCL in response to normal serum 25OHVD range at various transformed prostate volumes………………………………..134 Figure 2.12 Predicted DSL in response to normal serum 25OHVD range at various transformed prostate volumes……………… ……………….135 Figure 2.13 Predicted ALH in response to normal serum 25OHVD range at various ages……………………………………………………………..136 Figure 2.14 Predicted AOC in response to normal serum 25OHVD range at various ages……………………………………………………………..137

xiii
Figure 2.15 Predicted Normal sperm in response to normal serum 25OHVD range…………………………………………………………138 Figure 2.16 Predicted Loose Heads in response to normal serum 25OHVD range…………………………………………………………139 Figure 2.17 Predicted Normal sperm in response to normal serum 25OHVD range at various ages………………………………………140 Figure 2.18 Predicted Normal sperm in response to normal serum 25OHVD range at various transformed prostate volumes…………141 Figure 2.19 Predicted head defects in response to normal serum 25OHVD range at various transformed prostatic volumes…………142 Figure 3.1 Predicted percentage of proximal droplets in response to serum PRL ………………………………………………………………176 Figure 3.2 Predicted percentage of midpiece defects in response to serum PRL……………………………………………………………….177

xiv
LIST OF ABBREVIATIONS:
ABP Androgen binding protein IL Interleukin ALH Amplitude lateral head
diplacement IVF In vitro fertilization
AOC Average orientation change LH Luteinizing hormone AR Androgen receptor LIN Linearity BCF Beat cross frequency MMP Matrix metalloprotease BPH Benign prostatic hyperplasia NF Necrosis factor BTB Blood-testis barrier NRR Non-return rates Ca Calcium P Phosphorus CASA Computer assisted sperm
analysis PR Progesterone receptor
CBC Complete blood count PRL Prolactin CNRR Corrected non-return rates PTH Parathyroid hormone DAP Mean distance PVN Periventricular nucleus DCL Curvilinear distance RXR Retinoid X receptor DHT Dihydrotestosterone SCN Suprachiasmatic nucleus DSL Straight line distance SE Seminiferous epithelium E2 Estradiol STR Straightness coefficient ER Estrogen receptor TGF Transforming growth factor
beta FGF Fibroblast growth factor TNF Tumor necrosis factor alpha FGF-7 Keratinocyte growth factor VAP Mean velocity FITC Fluorescein isothiocyanate VCL Curvilinear velocity FSH Follicle stimulating hormone VDR Vitamin D receptor GH Growth hormone VDRE Vitamin D response element GnRH Gonadotrophin releasing
hormone VSL Straight line velocity
HPTA Hypothalamic-pituitary-testes axis
WOB Wobble coefficient
HPTH Hypothalamus 3 HSDH 3-beta hydroxysteroid dehydrogenase
IGF Insulin-like growth factor 25OHVD 25-hydroxy Vitamin D

1
CHAPTER ONE
INTRODUCTION, RESEARCH OBJECTIVES AND LITERATURE
REVIEW
1. INTRODUCTION
Prostatic diseases impact both general and reproductive health and are
influenced by androgens produced by the testes. Ironically, the very hormone
that is necessary for sperm production and male behaviours, such as libido, can
also have a negative impact on fertility and reproduction. Benign prostatic
hyperplasia (BPH) is the most common age-related condition in intact male dogs.
The condition affects approximately 80% of sexually intact dogs over five years
old and is characterized by an enlarged prostate as a result of both hyperplasia
and hypertrophy of prostatic epithelial cells [1, 2]. BPH may affect fertility,
especially the ability to cryopreserve semen in valuable stud dogs, however, it
also increases the susceptibility of the prostate to infection via ascension of
normal bacterial flora into an abnormal prostate and consequent prostatitis [3].
Surgical castration is the treatment of choice but represents a severe financial
and genetic loss for dog breeders. Unlike human males with the similar condition,
an enlarged prostate is not associated with impairment of urination but can
eventually lead to signs of hematuria, constipation and discomfort [2, 3].
Treatment with estrogens is known to decrease prostate size, however, severe
side effects such as squamous metaplasia of the prostate, thrombocytopenia,

2
leukopenia and fatal aplastic anemia discourage their use. Human approved
treatments used off-label in veterinary medicine are often cost prohibitive. There
is also very little published research on specific effects on physiological factors
related to spermatogenesis and semen quality in the dog. Recent human
research suggests that both Vitamin D and prolactin (PRL) have an influence on
the size of the human prostate and semen quality [4-7]. These findings have
suggested that investigation into prostate size and any possible association with
Vitamin D and/or prolactin may aid development of new treatments for BPH and
poor semen quality in the dog.

3
2. RESEARCH OBJECTIVES
The overall goal of this study was to investigate and identify contributing
etiologies of BPH that could lead to new prevention and treatment strategies.
A. Vitamin D study
I. Determine whether an association exists between serum
concentrations of 25-hydroxyvitamin D and BPH in male dogs.
II. Determine whether an association exists between serum
concentrations of 25-dihydroxyvitamin D and semen quality in
dogs.
B. Prolactin Study
I. Determine whether an association exists between serum
prolactin concentrations and BPH in male dogs.
II. Determine whether an association exists between
serum prolactin concentrations and semen quality in male dogs.

4
3. LITERATURE REVIEW
3.1 PHYSIOLOGY OF CANINE SPERMATOGENESIS AND METHODS OF
SEMEN ANALYSIS
3.1.1 Reproductive Physiology of the Testes and the Role of Hormones and
Local Factors In Male Reproduction
Male reproductive physiology controls the processes by which
spermatozoa are produced, transported and deposited into the female tract for
subsequent fertilization of the oocyte. Any interference with these processes,
whether physical, biological or chemical, can consequently affect fertility.
Spermatogenesis is controlled by the hypothalamic-pituitary-testicular axis, which
will be discussed in detail. The remainder of the reproductive tract, including
epididymides, deferent ducts, urethra, prostate gland, and penis is responsible
for the maturation, storage and/or transport of spermatozoa and seminal fluid to
be deposited in the female during estrus. Although male physiology shares
many similarities across different mammalian species, the dog has its own set of
unique variations.
The testes are found retroperitoneally and contained within the externally
located scrotum. The external location of the testes is crucial in maintaining
optimal temperatures for sperm production, as the process of spermatogenesis is
sensitive to the relatively high internal body temperature [8]. Thermoregulation is

5
further controlled through counter-current heat exchange between the arterial
and venous blood supplying the testes, via the complicated network of vessels
that form the pampiniform plexus [8]. The activity of scrotal sebaceous glands,
the thinness and lack of hair of the scrotal skin, and most importantly the
mechanical repositioning of the testis, with relation to the body wall, via the
cremaster muscle located within the spermatic cord and dartos of the scrotum,
further contributes to thermoregulation [8].
The testes contain three main cell types that are responsible for sperm
production: germ cells or spermatogonia, Sertoli and Leydig cells. The latter two
cell types are under hormonal control via the hypothalamic-pituitary-testicular
axis (HPTA). Gonadotrophin releasing hormone (GnRH) from the hypothalamus,
stimulates the secretion of pituitary luteinizing hormone (LH) and follicle
stimulating hormone (FSH). These hormones act on Leydig and Sertoli cells,
respectively, stimulating production of testosterone from Leydig cells and
synthesis of androgen binding protein (ABP) and aromatization of testosterone to
estrogen in Sertoli cells [9]. Testosterone released into the peripheral circulation
is converted to dihydrotestosterone (DHT) through 5 -reductase activity in a
variety of tissues, such as the prostate and hair follicles, and is responsible for
many of the secondary sex characteristics, especially during male embryonic
development [8, 9]. Negative feedback of the HPTA is mediated via both
testosterone and estrogen that act directly on the hypothalamus. Inhibin, another
hormone product of Sertoli cells, also acts via negative feedback on the pituitary

6
to limit FSH release. In this way a fine-tuned control of spermatogenesis is
achieved [9].
The Sertoli cell is responsible for the support and protection of developing
spermatozoa, by supplying the necessary nutrients and protecting the haploid
and antigenically foreign spermatocytes, spermatids and spermatozoa from
immune attack by the male’s defenses [10]. This barrier, known as the blood-
testis barrier (BTB), is crucial for the survival of the developing spermatozoa that
reside within the intercellular spaces of adjacent Sertoli cells and which are
separated from each other in their different developmental stages and the
extracellular environment via intercellular tight-cell junctions between Sertoli cells
[10, 11]. Testosterone is responsible for the adhesion between the developing
spermatocyte and Sertoli cell, maintaining the integrity of the BTB through
epidermal growth factor and its receptor [12]. The Leydig cells, also known as
interstitial cells, are separated from the Sertoli cells and spermatogonia by a
basement membrane and are supplied and supported by connective tissue,
lymphatics and blood vessels with which they have close contact [8]. The
specialized needs and functions of these differing testicular cell types mirror the
morphological and histological arrangement of the testes.
The Leydig cells, connective tissues and vessels are part of the interstitial
compartment of the testicular parenchyma, and as previously mentioned are
separated from Sertoli and germ cells by a basement membrane. The
seminiferous epithelium (SE) is present on the opposite side of the basement
membrane and is divided into three compartments: basal, deep and peripheral

7
adluminal [8, 13]. The importance of these compartments will be further
explained in the section on spermatogenesis. The seminiferous epithelium forms
a convoluted network of tubules within the microscopic lobules of the testicular
parenchyma that eventually join with rete tubules in the centre of the testis to
form the macroscopic structure called the mediastinum [8]. This is the major
collection region for spermatozoa within the testes, before they are further
transported through the efferent ductules to the head of the epididymis where the
final stages of spermatozoal maturation take place.
In addition to the long feedback loop with respect to the HPTA, certain
local factors have attracted attention in regulation of Sertoli and Leydig cell
function. These include, but are not exclusive to the inflammatory cytokines
tumor necrosis factor alpha (TNF ) and interleukins (IL), and growth factors such
as growth hormone (GH), transforming growth factor beta (TGF ), and insulin-
like growth factor (IGF) [12, 14]. Unfortunately, studies relating to these local
regulating factors of spermatogenesis are lacking in dogs.
In vitro immunohistochemical studies with testicular tissue have isolated
TNF (a 50kDa homotrimer originally isolated from endotoxin-stimulated
macrophages that caused necrosis in tumor tissue) from round and elongated
spermatids, pachytene spermatocytes (see section on spermatogenesis),
interstitial macrophages in mice [15], and in Sertoli cells in rats [16]. Its receptor
(TNF -R) has been shown to be present in both Sertoli cells in pigs [17] and
Leydig cells in the rat [18]. In porcine Sertoli cell culture under FSH stimulation,
an up-regulation of TNF -R in Sertoli cells has been reported [17] and suggests

8
a role of FSH in TNF sensitization in Sertoli cells. Apoptosis of germ cells
through FSH mediated expression of TNF has also been shown [19]. In Leydig
cells, TNF inhibits steroidogenesis via the nuclear factor- B (NF- B) pathway in
mouse cell culture to block the enzymes P450scc, p450c17 and 3 -
hydroxysteroid dehydrogenase (3 HSD) that are necessary in the multi-stage
conversion of cholesterol to testosterone [18]. Up-regulation of the androgen
receptor (AR) in rat Sertoli cells through the same NF- B pathway in DNA-protein
binding studies [20] is yet another effect of TNF in the possible regulation of
spermatogenesis and the interaction between Sertoli, Leydig and germ cells.
Of particular importance, TNF has been shown to down-regulate certain
proteins involved in maintaining the BTB, suggesting a restructuring of tight
junctions to maintain spermatocytes in their separate developmental states as
they move through to the luminal surface of the seminiferous epithelium (SE)
[12]. This is achieved in part by enhancing plasminogen activator/plasminogen
activator inhibitor system present in Sertoli and germ cells to induce phagocytosis
of residual bodies by the Sertoli cells during spermiation [12, 21,{Liu, 2007
#740}]. Induction of matrix metalloproteases (MMPs) necessary in endocytosis
and degradation of tight junction proteins causes reversible increases in the
permeability of tight junctions in the blood-testes barrier (BTB) [16]. Much of the
data on TNF has been compiled from studies examining rodent and swine
testicular tissue in culture, and it is yet unknown whether similar mechanisms of
action exist in the dog and/or how these factors may interact in vivo at
physiological levels in this species.

9
Li et al (2003) studied the function of TNF in in vivo studies in rats [23].
Using a recombinant human TNF at a dose of 0.5 micrograms per testis
(determined previously to be within physiological levels by the criteria that effects
were reversible and non-cytotoxic to Sertoli cells [16]) and an acute high dose of
2 micrograms per testis, immunohistochemical, immunoblotting and electron
microscopic techniques were used to identify a multitude of tight junction proteins
and observe seminiferous SE structure. The contralateral testis was used as a
vehicle-treated or non-treated control. Tissue samples were collected at time 0,
7 hours – 3 days, 3-5 days, 5-8 days and 14-60 days post-treatment. Decreases
in occludin and zona occludins-1 along with disruption of actin filament structure
related to tight junctions were found by day 3 for both treatment doses.
Complete recovery of the blood-testis barrier with germ cell repopulation was
complete by days 14-60 post-treatment. Increases in intercellular spaces
between Sertoli cells and thinning of the SE, damage to the tubules, the
presence of spermatids and spermatocytes within the SE lumen and depletion in
numbers of these cell types by day 5 were consistent with tight junction
disruption. Functional BTB integrity was also determined by testing diffusion of
fluorescent dye (FITC) administered systemically. FITC was found in the lumen
of the SE while in control testes FITC remained confined behind the BTB. By day
8, occludin and zona occludin-1 levels, as well as FITC permeability, returned to
normal showing no difference among treated, vehicle and non-treated controls.
In summary, although greater than normal physiological doses of TNF result in
dramatic disruption of Sertoli cell tight junctions, the presence of small amounts

10
of TNF in physiological normal testis suggests it is needed for normal function
of the BTB. It is possible that TNF helps in physiological reorganizing of the
Sertoli cell tight junctions to allow normal progress of developing spermatocytes
through the SE for eventual release of spermatids into the SE lumen. It is
important to note that only the results of the acute high dose treatment with TNF
were reported. Although physiological doses of TNF were investigated and
found to be significant and similar to the results obtained using
supraphysiological doses, it is unclear why more detailed reporting of these
results was absent.
The transforming growth factor beta (TGF ) family consists of three
isoforms and all three are expressed in a developmental and stage dependent
manner by early spermatids, pachytene spermatocytes and Sertoli cells, as well
as Leydig and myoid cells in the rat and porcine testes [24]. Along with TNF ,
they can disrupt the BTB in a reversible manner by degradation, endocytosis and
reorganization of tight junction transmembrane proteins to allow movement of
developing spermatocytes through the seminferous epithelium [12, 24]. Xia et al
(2009) used similar methodology to the study by Li et al (2003) and produced
similar outcomes with respect to functional permeability of the BTB when
exposed to TGF 3 [25]. However, they went one step further using an in vitro
endocytosis assay to measure amounts of endocytosed tight junction proteins
(occludin, junction adhesion molecule-A and N-cadherin) over time in Sertoli cells
when treated with endogenous amounts of TGF 3, TNF and germ cell culture
medium. The study found increases in the endocytosed proteins by 10 minutes

11
post-treatment when compared with controls. This effect was reversed by 180
minutes post-treatment. It is apparent that cross-talk between the developing
sperm cells and Sertoli cells is necessary for the gradual breakdown of these
tight junctions as spermatocytes make their journey across the SE to the lumen
for eventual release. The current literature supplies strong evidence for this
paracrine communication between developing germ cells, Sertoli and Leydig cells
and their regulatory roles in tight junction physiology.
Interleukin-1, traditionally known for its role in inflammation and produced
from macrophages, has been discovered to play a part in normal physiological
processes, and more specifically, spermatogenesis [26]. The 17kDa IL-1
subtype has been studied in detail and determined to have multifunctional
properties including acting as a growth factor for Sertoli cells [27] and
spermatogonia [28], stimulating production of other cytokines such as IL-6
(associated with apoptosis of germ cells [29]) and activin A (part of the TGF
family) in Sertoli cells, and regulation of steroid production by Leydig cells [26].
Interleukin 1 is produced by Sertoli cells and shown to be dependent on the
presence of germ cells [30]. Colon et al (2005) were able to confirm previous
studies relating IL-1 with increased Leydig cell steroidogenesis [31] with
addition of IL-1 to cultured immature rat Leydig cells where they observed a 4.3
fold increase in testosterone and a 2.86 fold increase in DHT [32] concentrations.
This stimulatory effect on androgen production is only seen in immature rat cell
culture while the opposite effect is seen in mature Sertoli cells and may be the
primary mechanism necessary for testicular maturation in pubescence [32].

12
Indeed, Svechnikov et al (2001) studied age related effects of IL-1 in 40 and 80
day old rats with suppression of testosterone secretion in the adult Sertoli cell
culture [31], while Colon et al (2005) reported IL-1 action on increasing
testosterone and DHT concentrations is potentiated by GH and IGF-1 in
immature rat cell cultures [32]. As with TNF and TGF , IL-1 also plays a role
in the regulation of the BTB through recycling of tight junction proteins within
Sertoli cells [26]. Sarkar et al (2008) reported that although steady-state levels of
tight junction proteins were not affected by IL-1 treatment of cultured Sertoli
cells, a loss of F-actin filament structure, identified using immunofluorescence
staining, was observed starting at day 8 and persisted until day 45 [33]. In
addition, diffuse localization of tight junction proteins (such as occludin and
junction adhesion molecule-1) away from the BTB and increased permeability as
assessed using the FITC technique described previously were significant findings
and were not seen in the non-treated controls [33]. Although similar results were
obtained to TNF and TGF studies of tight junction physiology, IL-1 actions
were delayed and not readily reversible in the rat [33].
The growth hormones GH and IGF-1 have also been associated with
spermatogenesis and semen quality. Their main effects appear to involve
survival and functioning of Leydig cells and are produced locally by Leydig and
Sertoli cells [14]. GH stimulates testosterone secretion by Leydig cells in rats
[34] and IGF-1 stimulates testosterone secretion in rat Leydig cells [35] and
increases the number of LH-receptors (LH-R) present in porcine Leydig cells in
culture [36]. Their effects are more dramatic in immature compared with adult

13
Leydig cell culture in both species [35, 36]. A recent in vitro study by Yoon et al
(2011) in horses however, found no effect of IGF-1 alone or in combination with
LH in prepubertal horses although a synergistic increase in testosterone was
observed in post-pubertal equine Leydig cell culture with LH than with LH alone
[37]. This indicates a less important role for IGF-1 during sexual development
compared with post-pubertal steroidogenesis in this species and is supported by
previous study [38]. It is important to note that incubation with IGF-1 was much
shorter in the equine study (24 hours) compared with a three day and 48 hour
incubation period in the rat and pig studies, respectively [35, 36] and may not
have been long enough to elicit a significant change. As with the other local
factors studied with respect to spermatogenesis, there is little or no information
as to their presence or role in spermatogenesis in the dog. Further research
within this area is needed if spermatogenesis in the canine is to be understood
and how other factors may impact the finely tuned control of reproductive
physiology in this species.
3.1.2 Spermatogenesis
Spermatogenesis is the process by which the primary germ cells are
transformed into the antigenically and chromosomally distinct spermatozoa that
are capable of fertilizing an oocyte. The process of spermatogenesis is
commonly broken down into several distinct stages: mitosis, meiosis,
spermiogenesis and spermiation [39, 40]. During mitosis primitive type A
spermatogonia in the basal compartment actively replenish themselves [39], to

14
overcome the consequences of normal aging and apoptosis. In this way, there is
a continuous supply of primary germ cells to maintain sperm production,
accounting for the male’s ability to produce sperm throughout his lifetime. Some
of these type A spermatogonia are selected through mechanisms involving
testosterone, retinoic acid and stem cell factors, and Dazl, Sox3 and UTP14
genes to become type B spermatogonia [41, 42]. Type B spermatogonia
continue to undergo mitosis and their last mitotic replication results in the primary
spermatocytes. At this transitional or preleptotene meiotic stage the primary
spermatocyte is present in the basal compartment [39].
Meiosis begins with the leptotene stage and at this time the primary
spermatocyte traverses the BTB or junctional complex (adherins and tight
junctions) between adjacent Sertoli cells and comes to lie in the deep adluminal
compartment [9, 39]. Meiosis I proceeds similarly to mitosis to produce the
diploid secondary spermatocyte (2n) and subsequently meiosis II, which occurs
behind the BTB, results in haploid round spermatids (1n) that are now
antigenically distinct from the other cells in the body. In this way four spermatids
are produced from one primary spermatocyte.
Spermiogenesis is divided into four phases: Golgi, cap, acrosome and
maturation. In the Golgi phase, the Golgi tubules manufacture the enzymes that
will be necessary for the development of the acrosome system and dictate its
position over the sperm nucleus [39]. Capping is the next process by which the
acrosome comes into contact with the nuclear envelope and covers the nucleus
over one third of its length. Following ‘capping’ the acrosomal phase continues

15
with the migration of the acrosomal system over the ventral surface of the
elongating spermatid [39]. The last phase of spermiogenesis is less dramatic as
the acrosome thins and its migration ends as it covers the majority of the
nucleus. The nuclear material continues to condense and excess cytoplasm is
removed from the cell and phagocytosed by the Sertoli cell. Also during this
stage the centriole elongates to become the tail and is oriented towards the
lumen. Spermiation is the final stage by which the Sertoli cell releases the
spermatid and excess cytoplasm and organelles are expelled from the spermatid
[39] resulting in a spermatozoon.
The timing of spermatogenesis has been described by the cell
associations or stages of the spermatogenic cycle (i.e. the time from beginning of
spermatogenesis until spermatid release from the associated Sertoli cell). Since
each Sertoli cell has many stages of developing spermatocytes, patterns of
cellular development are recognized and referred to as cell stages. In the dog,
one cell cycle contains eight stages and is 13.6 days in length [10, 42]. Total
duration of spermatogenesis is 61.2 days and is equivalent to 4.5 spermatogenic
cycles [42, 43].
The final stages of sperm maturation occur in the epididymis and are also
under the control of androgens. In the epididymis sperm acquire motility and
fertilization capacity through epididymal cell production of glycoproteins and
transpeptidases (antioxidant regulation) that promote condensation of DNA. In
the dog this process lasts approximately 10 days [44]. During transport from the
head to the tail of the epididymis the last remnants of cytoplasm travel from the

16
proximal to the distal portion of the sperm flagellum for release prior to final
maturation.
3.1.3 Factors Affecting Semen Quality
There are several factors that can affect semen quality in otherwise
healthy dogs. These include inbreeding, age of dog, breed, collection technique
and frequency of collection. In a study by Santos et al (2006), a knobbed
acrosome defect and poor fertility was observed in four closely related Miniature
Schnauzer dogs where coefficients of inbreeding were 5.3, 10.5, 2.5 and 19.8%
[45]. England et al (2010) did not see any significant correlations between
parents (n=24) and offspring (n=24) with respect to the five semen parameters
studied (percentages of normal motility, percentage morphologically normal
sperm, sperm concentration, total sperm output or total number of live
morphologically normal sperm in a group of normal dogs with no inbreeding
present). Moderate narrow sense heritability (h2) was found for percentage
normal motility (h2= 0.57) and percentage live morphologically normal sperm
(h2=0.21), and total number of live morphologically normal sperm (h2=0.47)
suggesting that breeding stock should be chosen with these parameters in mind.
These two studies suggest that genetics may play a part in semen quality
potential. Age is also thought to impact semen quality with young (< one year of
age) [46] and old dogs having poorer semen quality due to sexual immaturity and
degenerative changes in the testes (testicular neoplasia and decreased
testosterone production) [46, 47], respectively. However, Peters et al (2000)

17
were unable to find differences in spermatogenic changes in the testis with
respect to age in otherwise healthy dogs [48] . Breed effects on semen quality
mainly concern total sperm output and varies greatly with size of the testes,
meaning that larger dogs with larger testes produce larger number of sperm [44].
Semen quality can be negatively affected by the absence of a bitch in heat and
unfamiliar and stressful environments that can influence libido [44]. Frequency of
collection in the dog has been shown to result in optimal semen quality when
done once every two to five days [49]. Collection approximately one hour apart
results in a 70% decrease in total sperm numbers [50]. Although, no literature
exists on the effect of sexual abstinence on the quality of semen collected,
decreased sperm motility and lower numbers of morphologically normal sperm
may be present due to sperm cell senescence. As many factors have a role to
play in semen quality it is important to minimize the effects of those that can
manipulated and taking them into consideration when performing semen
evaluation and investigating possible causes of altered semen quality in the dog,
as well as in other species.
3.1.4 Methods of Semen Evaluation
Several methods of semen analysis are available, ranging from simple
light microscopy to computer assisted semen analysis (CASA) techniques using
various staining techniques to ultrastructural evaluation using electron
microscopy [51-53]. The three main properties of sperm commonly used for
semen evaluation are concentration, motility and morphology. Although

18
evaluation of these qualities does not give a complete picture of the fertilization
potential in every scenario, it can provide an adequate screening technique for
sperm defects. However, to the average veterinarian in reproductive practice,
light microscopy remains the most common technique when evaluating canine
semen [54, 55]. For research purposes, computer assisted techniques have the
advantage of standardization of evaluation, objectivity, repeatability, accuracy
and precision that cannot be achieved otherwise [56].
Determination of Sperm Concentration.
Measurement of sperm concentration in itself is not a measure of sperm
quality as ejaculate volume is dependent on the accessory sex gland secretion,
which in turn is influenced by time of year of collection, duration and intensity of
teasing, and frequency of collection in species such as the horse [57, 58]. In the
dog, semen volume results from prostatic secretions and depends chiefly on the
collector and separation of the second fraction from the rest of the ejaculate with
total volumes ranging from one to 80 millilitres and concentrations ranging from 4
to 400 million per millilitre [51]. England et al (1999) collected semen from a
mixed group of 65 dogs twice, approximately one hour apart and compared the
volumes of the second fraction [50]. First ejaculate second fraction volumes
averaged 1.9ml with a standard deviation of 1.3ml. They reported a significant
decrease in second ejaculate second fraction volume (mean=1.7ml and standard
deviation=1.2ml) as well as concentration and total sperm numbers in the study
population, with German Shepherd Dogs having a significantly higher second

19
collection volume, concentration and total sperm numbers than other breeds.
Wildt et al (1982) also observed a decrease in second fraction volume in inbred
dogs [59] although this finding was not statistically significant. Both these studies
showed that volume of the ejaculate can be highly variable without an impact on
reproductive parameters. However, by measuring volume and concentration,
total sperm numbers can be determined by multiplying the two values together.
Total sperm numbers are dependent on scrotal width and grams of testicular
tissue [44], which explains the discrepancy in sperm output between large and
small dogs. Ultimately, total sperm number is one measure of testicular and
reproductive function in the male.
Traditionally, sperm concentration is determined using a counting chamber
with a grid network i.e. Neubauer or Improved Neubauer chamber, standard
dilution of the semen sample and the use of a dilution media that kills or
immobilizes sperm (Unopette™, Becton-Dickinson) [54, 55]. Other electronic
counters such as densimeters and computerized assisted semen counters (within
CASA systems) have been developed, however, all are standardized using the
traditional hemocytometer method. Schafer-Somi and Aurich (2007) compared
concentrations of dog semen measured by hemocytometer, SpermaCue® and
SpermVision® analyser and found that they are all highly correlated with a p-
value < 0.01 with no significant difference between the methods studied or with
dilution using different extenders [60]. The ease of use of the latter methods
mentioned has allowed for quick and easy determination of sperm numbers in
large semen handling and processing facilities, where speed is essential.

20
However, the gold standard remains the hemocytometer method. A review of the
physics behind sample flow in capillary loaded slides in CASA systems compared
to hemocytometer methods explains differences in concentration measurements
due to the Segre-Silberberg effect [61]. In the shallower and longer chambers
used in CASA (20 µm) compared to the hemocytometer (100 µm) different
velocity gradients due to capillary action are created and result in increased
sperm at a defined distance from the chamber walls. It is then necessary for a
compensation factor to be applied for higher correlation between the two
methods (r2=0.936 compared with r2=0.984) [62]. This should be taken into
consideration when using CASA for sperm concentration measurements.
Evaluation of Sperm Morphology.
Sperm morphology has been used as a measure of reproductive
competency and fertility in multiple species [63, 64] and normal sperm
percentages in an ejaculate should range in excess of 80% [51]. A 2008
Swedish retrospective study by Al-Makhzoomi et al (2008) investigated semen
collected (107 ejaculates) and fertility history of 12 proven bulls used for artificial
insemination over a period of 22 months of progeny testing [65]. Fifty-six day
non-return rates (NRR) were corrected (CNRR) for several factors such as
season, inseminator, estrous control program, breed and parity and used as a
measure of bull fertility. The investigators found percentage of normal
morphology was positively correlated to fertility (CNRR) (r=0.22, p<0.05).
Percentage of head abnormalities was negatively associated with CNRR (r=-

21
0.23, p<0.05), pear-shaped head defects were negatively associated with NRR
and CNRR (r=-0.55, p<0.05), loose sperm heads with NRR and CNRR (r=-0.32,
p<0.01), and sperm heads of variable size and NRR (r=-0.27, p<0.05). In the
same study, spermatozoa with double-folded tails were also negatively
associated with NRR (-0.21, p<0.05). Other defects associated with infertility
include those located in the midpiece as well as the presence of proximal
droplets [66]. Enciso et al (2011) were also able to establish a correlation
between major sperm defects in bulls (double forms, pear-shaped heads, narrow
base of head, small abnormal heads, free heads, tail stump, whip tail, proximal
droplets, and midpiece ‘Dag’ defects) and DNA damage by validating a sperm
chromatic dispersion test and comparing it to sperm morphology assessed by
eosin-aniline blue staining techniques under light microscopy [67]. Assessment
of sperm morphology therefore can be considered a marker for reproductive
potential and DNA integrity.
An early study in a group of 42 bitches bred to 67 stud dogs (34 different
breeds), compared number of pregnant versus non-pregnant bitches inseminated
with differing percentages of normal sperm (50%, 55%, 60%, 65% and 70%)
using the 2 statistic and the greatest strength of association ( coefficient) to
determine the limit beyond which fertility was adversely affected [68]. Those
dogs that had greater than or equal to 60% normal sperm (using both light and
electron microscopic techniques and the sperm morphology criteria as described
by Blom (1973) [69]) had better pregnancy rates than those with less than 60%
normal sperm ( 2=4.54, coefficient = 0.42)) after correcting for those bitches

22
proven to be infertile (n=4) [68]. As well, percentage of head defects was highest
in the lower fertility group (<60% normal sperm) in addition tail defects, proximal
droplets and other midpiece defects were also associated with this group.
However, the statistical significance of the individual sperm defects is invalidated
as the investigators decided (in a non-repeatable way) which abnormalities were
most significant in the case where multiple abnormalities were present or which
was the most predominant defect amongst those of equal of significance. In
further support of morphological assessment of sperm in dogs, Rijsselaere et al
(2007) took breeding histories of dogs and divided them into three groups: fertile
(produced at least one litter in the three months leading up to the study),
subfertile (low pregnancy rates of <50% and/or dogs were unable to fertilize
bitches which became pregnant when mated to other dogs) or not used for
breeding yet [70]. Fresh semen was collected and stained using eosin-nigrosin
staining technique and compared to the breeding histories obtained. Fertile dogs
had significantly (p<0.05) higher percentages of normal sperm (mean SD, 63.3%
28.5) than subfertile (mean SD, 29.4% 29.9) dogs. Difficulty in assessing
fertility in dogs is challenging due to the limited number of breedings for dogs
compared with other species such as cattle and that conception failure is often
due to poor breeding management in the bitch [71]. Therefore the arbitrary
nature of the categorization of fertility from this study may not be an accurate
representation, although the results are consistent with those found in cattle.
Pena et al (2007) showed significantly decreased zona binding, compared with
the control dog, in a Basque Shepherd dog that failed to impregnate three

23
bitches of which two became subsequently pregnant after being mated with
another male [72]. Of the 98% abnormal sperm found on semen analysis, 88%
of these had proximal droplets and transmission electron microscopy revealed no
head, acrosomal or neck defects. This last study specifically pinpoints an
individual defect associated with decreased fertility as measured by a fertility
assay. The information from multiple species including dogs supports the
assessment of sperm morphology as a measure of reproductive potential and
fertility in the male.
Sperm morphology is traditionally evaluated by identifying defects related
to head, midpiece and tail region under light microscopy using special staining
techniques and/or phase contrast under high power (1000X magnification) oil
immersion. Among the stains commonly used are modified Wright’s-Giemsa
stains (DiffQuik or Protocol ) and eosin-nigrosin, the latter also known as a
vital/acrosome differential stain. In eosin-nigrosin stained specimens, live,
normal spermatozoa are unstained and appear white against a dark purple
background with a well defined smooth apical ridge indicating a non-reacted
normal acrosome while those with acrosomal damage appear darker or pink with
the apical ridge absent. Other useful vital stains use combinations, such as;
Trypan blue and Giemsa or Trypan blue, Bismark Brown and Rose Bengal stains
[52]. These staining techniques are more time consuming to use, however, they
are useful in detecting specific acrosomal defects. The use of these stains is
limited to evaluation of fresh semen, as many of the cryopreservatives used in
fresh-chilled and frozen semen have been shown to be incompatible with each

24
other [52]. Morphometric analysis using CASA is most beneficial to evaluation of
the sperm head; mainly the dimensions of length (L) and width (W), and
measures of head perimeter (p). From these measures the area (A), ellipticity
(L/W A/P2), elongation [(L - W)/(L + W LW/4A) of
the sperm head can be determined [73, 74]. However, normal reference ranges
with standardized classification systems have not yet been established based on
these parameters.
Fluorescein staining techniques may be used in conjunction with CASA.
The advantages of these techniques are that 1) cryopreserved semen can be
evaluated without interference from the media used in preservation of samples,
2) proportions of live-dead sperm can be determined, 3) acrosomal integrity,
capacitation status, mitochondrial function, chromosome fragmentation and DNA
content can be determined [52]. These characteristics of sperm are detected
through fluorescein staining used in conjunction with fluorescence microscopy
and/or flow cytometry. The disadvantage of these systems is the cost of
equipment and necessity for standardization and validation, so they are not
routinely used in clinical practice [56].
Morphological defects can be grouped as primary or secondary, referring
to whether it is a defect of spermatogenesis or occurs during epididymal
maturation and transport, respectively, or after ejaculation due to environmental
or processing effects. Primary sperm defects that are associated with poor
fertility include midpiece defects. Proximal droplets associated with poor fertility
can be considered primary or secondary depending on whether maturation of

25
sperm or hidden midpiece defects are the source of the abnormality [52]. This
particular classification, however, does not always determine the severity of the
defect and whether it affects the fertility of the whole ejaculate.
The concept of compensable versus non-compensable sperm defects has
become a more acceptable form of classification. It is based on the notion that
by increasing numbers of normal sperm within an ejaculate, the chances for
fertilization are increased [66]. For example, increasing the total number of
normal sperm in the ejaculate could compensate for those sperm defects that do
not permit binding and/or penetration of the zona pellucida. This would lead to
increased numbers of normal sperm that can fertilize ova, thereby increasing the
chances of fertilization. On the other hand, defects that allow for normal
fertilization of the ovum but have their impact on embryo development would be
considered non-compensable. This is due to the fact that the percent chance of
fertilization of these types of defects does not change with increasing sperm
numbers.
CASA and Sperm Motility.
Computer assisted sperm analysis (CASA) is becoming the gold standard
in semen analysis since the digitization of data allows for increased objectivity,
precision and accuracy of the parameters measured and the potential for
decreasing inter- and intra-laboratory variation through validation and
standardization of analysis [56]. Briefly, CASA consists of a microscope
connected with a video camera with a video frame grabber card (50-60

26
frames/sec) that inputs data as pixels and data is then digitized into a computer
with specialized software to measure concentration, motility and/or morphology of
sperm. Most CASA systems use phase contrast/dark field microscopy that
illuminates sperm as white against a black background making it possible to
identify the centroid (determined by Fourier analysis as the centre of gravity of an
object [75]) of the sperm head (pixel dimensions can be set for different species
with identification of a tail) for tracking purposes, while other systems use
fluorescent dyes (concentration and morphology studies) for identifying sperm.
Software algorithms, through identification of the centroid, identifies successive
fields by predicting the probability of the zone within which the sperm is likely to
move, determines the minimum number of centroids needed for analysis within a
given timeframe, determines the minimum distance between video frames that
indicates the sperm is moving and determines the number of forward video
frames to be looked at to find a missing centroid [76]. In this way the CASA
system is able to track and record data from multiple individual sperm
trajectories.
Although not without issues such as agreement among research
laboratories regarding the validation and standardization of parameters used in
the software programs of different systems, CASA still holds promise in both
human and veterinary clinical practice. The parameters of concentration, motility
and morphology can be assessed; however, there is little information to be found
in the literature on validated computerized morphological assessment of sperm in
veterinary research [56, 77]. Using any kind of CASA system requires that the

27
sample be diluted. Iguer-Ouada (2001) and Verstegen (2002) found that motility
parameters were not affected between the concentrations studied (50-300 X
106/ml), however, accuracy and repeatability of the measures were affected at
concentrations below 50 X 106/ml [56, 77, 78].
Some systems have been validated for dog semen such as the Hamilton-
Thorne sperm analyzer [78] and Sperm Vision® [60]. It has been shown that
type of specimen chamber, temperature, frame rate, number of fields analyzed,
sperm concentration (see above) and semen handling techniques and extenders
can all have an effect on the measurements of concentration and motility [56, 60,
77]. It is recommended that a minimum number of 100 cells should be analyzed
for results to be valid, especially in oligospermic samples, as larger numbers of
cells analysed reduces the coefficient of variation and increases the precision of
the results [56]. High numbers of sperm per examined field can interfere with
estimations due to the affect of collisions, sperm exiting the field, motion in the
wake of moving spermatozoa and the ability of the system to differentiate and
register individual cells. Misidentification of foreign particles or debris can also
occur especially when using semen extenders with egg yolk.
The main motion parameters measured in most CASA systems include
the following: 1) Straight line distance (DSL, µm), 2) Curvilinear distance (DCL,
µm), 3) Mean distance (DAP, µm), 4) Curvilinear velocity (VCL µm/s) which is the
instantaneous velocity of the spermatozoa along its path, 5) Linear velocity (VSL
µm/s) the velocity of the spermatozoa along its straight line path, 6) Mean
velocity (VAP µm/s), 7) Straightness (STR%), a ratio of the VSL/VAP which gives

28
the straightness of the path taken as a percentage of the optical straightness
(100%), 8) Linearity (LIN = VSL/VCL) which corresponds to how close the cell
travels along to the straight line path, 9) Wobble coefficient (WOB=VAP/VCL) or
the oscillation of the curvilinear path along the mean path, 10) Amplitude of
lateral head displacement (ALH µm), 11) Average orientation change of sperm
head (AOC, degrees), and 12) Beat cross frequency (BCF Hz) or the frequency
with which the sperm head crosses its average pathway [60]. All these
parameters give a detailed description of sperm motion and are ultimately used
as tools in measuring fertility potential and reproductive health in the male and
assessing the sperm motion necessary to travel through the female tract to reach
the oocyte and fertilize it.
In a study by Farrell et al (1987) of 11 artificial insemination (AI) bulls
ranging in age between 6 and 11 years, sperm motility parameters from 44 total
ejaculates (211 956 total services) measured by CASA were compared to 59-
day non-return rates in cows and adjusted for whether semen was used in heifers
or cows or whether the females were enrolled in Dairy Herd Information or not
[79]. The results obtained showed significant positive associations between
fertility and the motility parameters of total motility, progressive motility, ALH,
VAP, VCL, VSL, BCF, LIN using multiple linear regression modeling and
achieved r2 values ranging from 0.34 to 0.98. It is important to note that non-
return rates between herds were also accounted for in order to increase the
accuracy of the estimates of fertility. The high r2 estimates from this study were
produced using combinations of motility parameters while only total motility in

29
univariable analysis with fertility had significant correlation alone. It is apparent
that fertility is dependent on the collective nature of these parameters in this
species.
Love et al (2011) looked at several fertility measures in the stallion
(percent pregnant per season, percent pregnant per first cycle and percent
pregnant per cycle) and motility parameters (total motility, progressive motility,
VAP and VSL) and found significant correlations with total motility and
progressive motility with all three fertility parameters [80], however, significance
of other motility parameters varied amongst the three fertility parameters
measured. The most sensitive measure of fertility in the stallion in this study was
determined to be percent pregnant per first cycle and percent pregnant per cycle
when compared to seasonal pregnancy rate. This study in particular shows the
importance of definitions of fertility and the difficulty in assessing outcomes from
CASA motility parameters. Care should be taken when interpreting those
findings and correlating them across studies and species.
CASA has importance in in vitro fertilization (IVF) and embryo transfer as
a tool to overcome subfertility in humans. A study looking at differences in sperm
motility between good (fertilization rate >50%) and poor (fertilization rate 50%)
fertilization rates found that significant associations exist between these two
groups and the motility measurements of ALH, VCL, and VSL [81] but not BCF,
VAP, LIN, and STR. Earlier, Aitken et al (1985) attempted to determine which
characteristics of sperm motion allowed it to penetrate cervical mucus. The study
measured the motion characteristics of sperm and determined bovine cervical

30
mucous sperm concentrations after incubation with a known volume and
concentration of sperm. The authors determined that a large ALH was necessary
for cervical mucus penetration and movement in the bovine [82]. These findings
were further supported by studies showing that in human IVF, larger ALH means
were correlated with higher fertilization success rates and were attributed to an
ability to penetrate the cumulus oophorus and zona pellucida of the oocyte [83,
84]. ALH would appear to be necessary to both in vivo movement through the
barriers encountered in the female reproductive tract and through the barriers
encountered with the oocyte in IVF. In the dog, it has been found that most of
motility parameters measured by CASA had correlation to fertility except for BCF,
LIN, and STR measured by the Hamilton-Thorne semen CASA [70]. Fertile dogs
in this instance were defined as those dogs having produced one litter in the
three months leading up to the study and sub-fertile if pregnancy rates were
below 50%. A significant correlation (r=0.44) between percentage of
morphologically normal sperm and percentage progressive motility has been
shown in the dog [85].
In general the motility parameters that have consistently been associated
with fertility across species include total and progressive motility, ALH, and VSL
and may explain why these parameters are most often studied with relation to
fertility. However, it is important to note that in most studies multiple interactions
with other motility parameters are found, suggesting that fertility is correlated to
combinations of motion characteristics necessary for sperm to travel to the
female oocyte. In this way CASA results should be interpreted according to

31
whether natural breeding/insemination or assisted reproductive techniques are to
be used. Although CASA measures motility in an artificial environment it does
not measure how motility is affected within the female reproductive tract, nor
does it evaluate the genetic competency or fertilizing ability of sperm. Although,
correlated with fertility, motility measured by CASA is merely one piece of a
complex puzzle and cannot account for the multitude of factors affecting fertility
of a male.
3.2 THE CANINE PROSTATE GLAND
3.2.1 Anatomy and Physiology of the Prostate Gland
The dog has only one accessory sex gland, the prostate gland. As a
result it contributes over 97% of the ejaculate fluid [1]. The prostate is positioned
retroperitoneally in the pelvis and surrounds the urethra near the neck of the
bladder. The rectum is located dorsally and the symphysis pubis ventrally to the
gland. Its craniocaudal limits are variable and dependent on the size of the
gland; which increases normally with age, and can therefore be present in a
completely pelvic position or abdominally [86]. It has a symmetrically bilobed
structure with a fibrous tissue capsule. It is supplied by the prostatic artery, a
branch of the internal pudendal artery and drains via the prostatic and urethral
veins. Nervous innervation from the parasympathetic branches of the pelvic
nerve stimulates glandular secretion while sympathetic innervation causes
contraction of smooth muscle and emptying of the gland through multiple

32
ductules into the pelvic urethra, on the ventral aspect of the gland, near the
openings of the vasa deferens in the colliculus seminalis [8, 86].
Histologically the prostate is separated into stromal and glandular parts.
The stroma is made up of a network of delicate fibrous connective tissue strands,
smooth muscle, blood vessels and nerves that are continuous with the capsule,
extend throughout the substance of the gland and coalesce near the urethra.
The glandular part consists of columnar epithelial cells and is organized into
tubuloalveolar lobules that empty through a duct system into the urethra through
several small openings on the colliculus seminalis, as previously described [8,
86].
Prostatic secretions contain components that nourish and maintain sperm
such as fructose, as well as other constituents that provide a mechanical
transport medium through which sperm travel through the female reproductive
tract. In addition, prostatic secretions have anti-bacterial properties [87].
Selected prostatic components and their normal ranges are listed in Table 1.1.
The main protein secreted by the prostate is canine prostate-specific esterase
which accounts for 90% of the total protein secretion by this gland [88]. Although
this enzyme differs in amounts between normal and diseased prostatic tissue it
cannot be used to differentiate among major prostatic diseases [88]. It is still
unknown what role this enzyme plays in male dog reproduction.

33
Table 1.1. Selected Physical and Chemical Analyses of Prostatic Fluid in Dogs with Healthy Prostates
Mean SD (range) Sample Size
pH 6.2 0.3 (5.5-7.1) n = 43
Specific gravity 1.018 0.005 (1.008-1.028) n = 40
Cholesterol mg/dl 27 17.0 (8.0-73.0) n = 29
Zinc ( g/ml) 62.3 35.3 (10.3-120.6) n = 20
Copper ( g/ml) 7.1 4.8 (1.3-19.5) n = 20
Iron ( g/ml) 0.7 0.5 (0-1.6) n = 20
Magnesium ( g/ml) 16.4 9.5 (3.4-40.0) n = 20
Data adapted from Branam et al [89]
The prostate is an androgen-dependent organ as evidenced by complete
atrophy of the gland following castration [86]. It is dependent mainly on the
steroid hormone dihydrotestosterone (DHT) which is converted from testosterone
by the enzyme 5 -reductase produced within the gland. In fact, re-growth of the
atrophied gland can be achieved via supplementation with a combination of
exogenous estradiol (E2) and DHT [90]. DHT is also the main hormone
responsible for the differentiation and development of the gland from the
urogenital sinus during embryogenesis [8].
3.2.2 Pathophysiology of Benign Prostatic Hyperplasia (BPH)
BPH, as the name implies, is a benign enlargement of the prostate gland
that is due to hyperplasia, as well as hypertrophy, of mainly prostatic epithelial
cells in the dog. There appears to be no breed predilection for BPH [91]. Early

34
studies of Beagle dogs have shown maturity of the prostate, according to
histological pattern of developed alveoli and maximum size, occurs by 1.5 years
of age with a remarkable three-fold increase in prostatic size compared with
immature dogs. One study suggested that starting as early as 2-3 years of age
and by 4 years of age >40% of dogs have histological evidence of BPH with 80%
and 100% of Beagles having confirmed BPH by 4 and 7 years of age,
respectively [1]. However, this study was limited to Beagles and may not have
relevance in other breeds. A complex version of BPH has also been described in
which dilation of ducts and the presence of cysts and inflammatory cells occur
with degeneration of epithelial cells [1]. Although the same study also correlated
a decline in secretory volume with age and state of the prostate, many factors
play a role in ejaculate volume that can be difficult to control for. A definite
correlation between prostate size, age and weight has been proven in multiple
studies [47, 92-94].
Androgen:estrogen ratio appears to play a significant role in the
development of BPH. This theory is supported by reported declines in serum
testosterone and DHT concentrations while 17 -estradiol concentrations remain
unchanged [1]. Using histology to identify immature, normal and BPH tissue
(defined as having both hyperplastic and hypertrophic elements) and measuring
individually pooled serum hormone levels using radioimmunoassay in 42 beagles
(8 months to 9 years of age) over three weeks, Brendler et al (1983) were able to
compare mean serum hormone levels amongst the three prostatic groups. They
also compared mean testosterone and E2 with age and found a general trend of

35
decreasing testosterone (with no changes in mean E2) that followed histological
changes and weight increases in the prostate suggesting altered androgen
metabolism in prostatic hyperplasia/hypertrophy. It is important to note that
statistical analysis was not undertaken in this study and significance of these
results is unknown. Experimentally induced BPH requires the administration of
estradiol and DHT, rather than DHT alone [95]. Castrated and control dogs were
divided into groups receiving different hormone treatments given three days a
week for a total of 40 weeks after prostate size regressed at 4 weeks post-
surgery. Testosterone and DHT alone did not produce BPH while E2 alone
caused a squamous metaplasia of the gland. In contrast, the combination of E2
and DHT but not E2 and testosterone caused hyperplastic and hypertrophic
changes in the prostate. This information further suggests androgen:estrogen
ratio to be of significance. It is likely that although DHT and testosterone serum
levels have been shown to be decreased in dogs with BPH, they are still
necessary in the pathophysiology of BPH. This may be due to increased uptake
and utilization of testosterone and DHT in prostatic tissue causing a decrease in
peripheral serum concentrations of these hormones.
A study investigating enzymes of steroid hormone metabolism within the
prostate in normal young (age range 17-40 years) and aged men (age range 60-
80 years) showed that aged men with or without BPH had decreased prostatic
tissue enzyme activity of 3 ( ) hydroxysteroid dehydrogenase (HSDH) compared
with normal young men [96]. The reduction in enzymatic activity suggests that
altered androgen to estrogen ratios found in BPH are due to decreased

36
degradation of stromally-produced dihydrotestosterone in BPH, rather than an
increase in DHT production, which might occur through conversion from
testosterone under the influence of increased 5 -reductase activity. The
mechanisms which lead to alterations in HSDH activity in BPH have not been
elucidated.
It has been suggested that lack of degradation of DHT mediated by
estradiol correlates with an abundance of estrogen receptors (ER) present in
healthy and hyperplastic prostatic stromal and epithelial tissues [97, 98].
Compared to normal prostatic tissue there is a significant decrease in the
percentage of ERs in hyperplastic and neoplastic canine prostates [98]. These
findings suggest a more indirect role of estrogens in the pathology of BPH. It
has been postulated that free radical production by increases in relative estrogen
concentrations cause tissue damage by which DHT induces abnormal prostatic
growth [99]. Interestingly, progesterone receptors (PR) were also identified for
the first time in diseased conditions of the prostate suggesting as yet an unknown
role of progesterone in BPH development [98].
Similarly to the testes, biological effects of testosterone and estrogen in
hyperplastic prostatic tissue may be mediated through several growth factors
[100, 101]. The most important of these growth factors include fibroblast growth
factors (FGF) of which keratinocyte growth factor (FGF-7) appears to be the most
potent mitogenic factor in hyperplastic tissue [102]. IL, IGF and TGF appear to
be important in BPH [100]. These autocrine mediators between stromal and

37
epithelial tissues are considered to be responsible for the increase in prostatic
growth seen in BPH.
Using ELISA and comparing results from normal (peripheral and transition
zones) and hyperplastic human prostatic tissue, Ropiquet et al (1999) were able
to assess whether over-expression of FGF-2 and FGF-7 was present in
hyperplastic tissue [103]. They found a significant increase in both FGFs
compared to normal prostatic tissue. In addition, they investigated the effect of
FGF-7 on epithelial and FGF-2 on stromal growth in prostatic tissue based on
results from other studies showing specificity of these FGFs for distinct cell
populations within the prostate [104, 105]. The results showed significant
increases in total epithelial cell numbers when treated with FGF-7 compared with
controls as well as significant increases in stromal cell numbers when treated
with FGF-2. ILs have been shown to stimulate both these growth factors in
culture. IL-1 was observed to be produced by prostatic epithelial cells and
stimulates FGF-7 production in stromal cells [106], while IL-8, also produced by
prostatic epithelial cells, stimulates FGF-2 in stromal cell culture [107]. These
two growth factors appear to play a significant role in the pathogenesis of
hyperplasia in humans where both stromal and epithelial cells show abnormal
growth. Increases in and irritation of smooth muscle in BPH, as a result of
inflammation, are important in the lower urinary tract symptoms (caused by
increased urethral pressure) that are seen in men [108]. This clinical sign,
however, is not one normally seen in the dog and therefore stromal components
may not have the same importance. In the dog epithelial cells are the main cell

38
type involved in the pathophysiology of BPH, although there is a minimal stromal
component suggesting a greater role of FGF-7 and not FGF-2 in pathogenesis of
BPH in the dog.
IGFs, and perhaps more specifically, IGF binding proteins (IGFBP) seem
to work in conjunction with TGF in regulation, or lack thereof, in BPH. TGF
has been localized mainly in prostatic epithelial cells and expressed in larger
amounts in BPH [109]. Cohen et al (2000) were able to show a 15-fold increase
in IGFBP3 in normal stromal cells compared with a two-fold increase in BPH
when treated with TGF [110]. These finding were correlated to a 68% growth
inhibition in normal prostatic culture compared to 26% in the BPH culture [111].
IGFs have also been localized in prostatic tissue and found to be up to ten-fold
higher in stromal cells from BPH tissue compared with normal tissue [112].
Therefore, growth of tissue in BPH is most likely mediated through multi-factor
complex processes where there are not only increases in growth factors but also
in inhibitory factors that interfere with normal apoptotic processes. The result is
an overall increase in the size of the gland.
Unlike BPH in men, BPH in the dog involves predominantly glandular
rather than stromal components [113]. Growth of the prostate proceeds in a
peripherally diffuse pattern in the dogs and does not interfere with urinary tract
function [1]. Mechanical occlusion of the prostatic urethra occurs in men where
localized nodular stromal proliferation in the so-called transition zone near the
urethral sphincter causes compression of the urethra and impaired urination
[114]. This is exacerbated by the fixed nature of the prostate. Blood in the

39
ejaculate is the most common clinical sign of BPH in dogs and the outwardly
expanding gland rarely causes compression of the rectum and constipation.
More commonly as the non-fixed gland increases in size and weight it assumes a
more abdominal position, especially when the bladder is full, and in some
instances can be difficult to palpate per rectum.
3.2.3 Conventional Treatment of Benign Prostatic Hyperplasia
There are at present several treatment modalities for BPH, however, side
effects and/or relapses post-treatment are common [86]. Due to the benign
nature of the condition, treatment is only sought or recommended where clinical
signs are present. The treatments for BPH in the dog centre on the hormonal
component of the condition. Ultimately, there is down-regulation of testosterone
and/or DHT production as these are the major pathophysiological components of
the condition. Castration, whether by physical or chemical means, is the most
dramatic recourse for complete remission and involution of the gland by
eliminating the main source of androgens. This solution however, is not
considered to be desirable in the valuable breeding animal or where the risks of
surgery/anesthesia far outweigh the benefits. Pharmacological treatments have
focused on three modes of action: 1) 5 reductase inhibitors (decrease DHT in
the prostate), 2) androgen receptor antagonism and 3) GnRH modulators [86,
91].
The azasteroid Finasteride, a 5 reductase inhibitor, has become the most
widely used drug for treatment of BPH in dogs for several reasons. In a double-

40
blinded drug trial, nine dogs with BPH confirmed by having clinical signs of blood
in the ejaculate or constipation, were treated with finasteride (five dogs, 5 mg/day
PO for 16 weeks) or placebo (four dogs, 5 mg powdered sugar in gelatin
capsule/day PO for 16 weeks) [115]. Using both ultrasound (prostate volume)
and radiological (prostate diameter as a percentage of the distance from the
sacral promontory to the pubis) measurements of prostate size at 8 and 16
weeks of treatment, the investigators observed a 24% decrease in prostate
diameter and a 41% decrease in prostate volume by 8 weeks of treatment that
did not differ significantly by the end of the 16 week treatment period. Control
dogs experienced no significant changes in prostate diameter or volume at either
the 8 or 16-week treatment periods. Serum DHT decreased by 50% in the
treatment group by 8 weeks and remained unchanged through the completion of
the study while serum testosterone did not change. No changes in semen quality
were noted except for a decrease in semen volume that was not significant. It
was noted that the five finasteride treated dogs bred successfully during and after
finasteride treatment. None of the dogs showed changes in libido or clinical
signs of blood in the ejaculate or constipation. All clinical signs i.e. blood in the
ejaculate and constipation used to diagnose BPH in individual dogs were
alleviated within four and one week(s) after the beginning of treatment,
respectively. The successful treatment and alleviation of BPH and its clinical
signs in this drug trial, in combination with no evidence of side effects relating to
semen quality, fertility and libido have made finasteride the drug of choice in
treating BPH in dogs.

41
Treatments involving androgen receptor antagonism have also shown
efficacy in decreasing prostate size in BPH. Osaterone and other androgen
receptor antagonists have been shown to decrease prostate size in dogs [116].
A clinical trial examined 142 dogs that were treated with either osaterone or
delmadinone for 180 days. Osaterone had a significantly greater decrease in
prostate volume (38%) compared with delmadinone (28%) with maximum
reduction in volume by 14 days of treatment for osaterone. Complete remission
of clinical symptoms of BPH occurred in 50% of the dogs by day 14 with 83%
remissions by the end of the study period. Major side effects included increased
appetite and changes in behaviour in a small percentage of the dogs. In another
study by Tsutsui et al (2001) examined semen parameters of beagle dogs during
treatment with osaterone and found no significant differences in sperm count,
morphology, motility, semen volume or pH [117]. Significant decreases in serum
testosterone and LH were observed during the treatment period, however these
levels were maintained during treatment and no adverse side effects were
observed. Although osaterone has shown good clinical efficacy with little or no
side effects it is not currently available as a treatment option in North America.
Progestins, with antiandrogen action, such as delmadinone have also shown
similar efficacy in reduction in prostate volume [116]. Side effects of
delmadinone treatment include inhibition of secretion of adrenocorticotropic
hormone at the hypothalamic-pituitary level, changes in maturation of epididymal
spermatozoa, increases in appetite [116], as well as detrimental effects on sperm
morphology, motility and sperm numbers at higher doses [91]. These effects do

42
not make it desirable when other pharmacologically safe alternatives are
available.
GnRH agonists, such as deslorelin, act by initially causing an increase in
GnRH secretion with subsequent increases in LH/FSH and testosterone. After
the initial increase in hormone up-regulation, a negative feedback inhibition of
GnRH ensues causing decreases in LH/FSH and testosterone. Although,
deslorelin causes a decrease in prostate volume [118], major side effects on
fertility through down-regulation of hormones regulating reproduction in the male
does not make it a desirable option in the breeding dog. At the present time no
GnRH antagonists for BPH treatment are commercially available in Canada or
the USA.
In summary, only those drugs that provide a decrease in prostate volume
and clinical signs of BPH without affecting fertility are desired in the stud dog.
The effects and side effects of the drug treatments mentioned here are reversible
with discontinuation of treatment. This holds true for the decrease in prostate
volume and therefore long-term administration is required to maintain a desirable
prostate size as well as fertility in the stud dog. This has led to the use of
azasteroids with inhibitory action on 5 -reductase activity to be the drug of
choice in breeding animals.
3.2.4 Ultrasonography of the Prostate
Traditional methods of determining size, symmetry and quality of the
prostate gland are subjective, such as digital palpation per rectum, as well as

43
semi-quantitative means using radiography [91, 119]. However, with the
introduction of, and advancements in, ultrasonography, an efficient and non-
invasive tool for determining prostate size, symmetry and lesions has become
available [93, 120, 121]. Due to the pelvic position of the prostate in human
males, ultrasonography is performed using a specialized transrectal probe to
evaluate prostate volume. Ultrasonographic measurements are highly
correlated to volumes measured in cadaver specimens in humans [120].
Although 95% accuracy can be achieved using planimetric measurements with
sequential cross-sectional images at 4 mm apart, this is considered far too time-
consuming and tedious in a clinical veterinary setting. Acceptable accuracy can
be attained by using length, width and height of the gland and using a simple
elliptoid equation to calculate volume [120, 122].
In the veterinary setting transrectal ultrasound is not commonly used in
small animal evaluation thus specific ultrasound probes for this purpose do not
exist. Therefore, ultrasonography of pelvic structures is performed mainly using
a transabdominal approach, and is common practice for prostatic evaluation in
dogs. As previously described, the canine prostate can have a pelvic and/or
abdominal position due to lack of pelvic fixation as seen in men. The greatest
difficulty in evaluation of the completely pelvic prostate in dogs is the positioning
of the ultrasound probe, which must be angled obliquely on the abdomen
towards the pelvic inlet [94, 121]. This makes achieving a true transverse cross
section of the gland difficult. However, relatively accurate estimations of prostate
size can be achieved as the canine prostate can be manipulated via digital

44
manipulation per rectum without causing distress in the patient [93]. There is little
information in the literature on ultrasonography of the canine prostate with
respect to correlation of ultrasound measurements versus actual prostate
volume. However, both Atalan et al (1999) and Kamolpatana et al (2000) found
a high correlation between estimated prostate volume and actual volume
measured through caliper measurement of length, width and height using a
modified formula for the volume of an ellipse (R = 0.76) or through the use of
water displacement technique (R2=0.94) of the dissected prostate glands [94,
121]. As a result, it has been shown that ultrasonography is a fairly accurate
method for evaluation of prostate volume and for monitoring size changes during
treatment for prostate disease [121] and more specifically BPH.
3.3 VITAMIN D AND ITS ROLE IN MALE REPRODUCTION
Currently, the non-traditional roles of Vitamin D have become important in
cancer research due to its ability to affect the normal life cycle of cell populations
within multiple tissues ensuring that natural cell death, or apoptosis, occurs within
these populations [123]. Although BPH is not categorized as a cancerous
process, it shares similarities in that cell proliferation exceeds that of cell death
leading to an increase in both the number and size of prostatic cells. This
unchecked rate of growth, albeit a non-fatal one, has an effect on susceptibility to
infection [91], as well as a secondary impact on reproduction. Treatment can be
expensive, consequent prostatitis a risk, and the inability to use assisted

45
reproductive techniques, such as semen freezing [124], a concern in valuable
breeding animals.
3.3.1 Vitamin D Metabolism
The traditional role of Vitamin D in calcium and phosphate homeostasis
(Ca:P) and bone metabolism was first elucidated by Edward Mellanby in 1918
while researching rickets in children [125]. Unlike humans, dogs do not
synthesize Vitamin D through the action of ultraviolet radiation on the skin [126,
127]. Therefore, dogs rely heavily upon dietary intake of Vitamin D. The majority
of commercial dog foods meet, and more frequently exceed, the minimum daily
requirements for this vitamin (500-5000 IU Vitamin D/kg food/day) set out by the
National Research Council and Association of American Feed Control Officials
[126]. In some cases commercial dog foods come close to the maximum
allowable amounts beyond which toxicity occurs. Toxicity may manifest as
cardiac arrhythmias, stiff joints and renal disease [128].
Vitamin D is a fat-soluble vitamin, and is ingested from the diet as Vitamin
D3 from animal sources such as liver and fish oils. Hydroxylation to the more
active form, 25-hydroxycholecalciferol (25OHD3), occurs via 25-hydroxylation in
the liver, both directly in response to decreased blood calcium and phosphorous
concentrations and indirectly through an increase in Parathyroid Hormone (PTH)
when decreased blood calcium concentrations occur [126]. Further conversion of
25OHD3, occurs via renal 1 -hydroxylase activity to 1,25(OH)2D3, also known as
calcitriol. Calcitriol works directly to increase intestinal absorption and renal

46
reabsorption of calcium and phosphorous and finally to dissolve both
components from bone reserves [126]. Perhaps the most important and
significant role of Vitamin D through this pathway is the prevention of rickets, a
condition affecting growth in the young that leads to softening and deformation of
developing bone tissue [125]. Alternatively, excess serum Ca and P triggers
hydroxylation by the kidney to 24,25(OH)2D3, resulting in deposition of Ca and P
in bone [125].
In recent years, emphasis on Vitamin D research has grown owing to its
association with decreased cancer mortality risk [125, 129, 130]. Garland was
the first to show an association between decreased risk of colorectal cancer and
geographic latitude [130, 131]. This stimulated further study into the anti-cancer
effects of Vitamin D. As a result, the non-traditional role of Vitamin D has come
to light; mainly its anti-proliferative effects and its role as a mediator of cell
differentiation and apoptosis in multiple tissues [132, 133]. Identification of
1,25(OH)2D3 and the enzyme responsible for its formation, 1 -hydroxylase,
ubiquitously throughout human tissues, including the prostate gland [134]
encouraged further research into the role of Vitamin D. These tissues include
human and rat kidney, stomach, large intestinal epithelium, mammary gland,
ovary, prostate gland, and pancreatic duct cells [135]. However, studies are
lacking with respect to these extra tissue effects of Vitamin D in the dog.
Tissue concentrations of 1,25(OH)2D3 are dependent on the circulating
serum 25OHVD3. Deficiency, or more importantly insufficiency, in circulating
25OHD3 results in a lack of Vitamin D effects in individual tissues through

47
decreased binding to the Vitamin D receptor (VDR) [135]. Consequently, there is
a break in the chain of inhibition or induction of transcription of specific genes
related to regulation of the cell cycle (proliferation, invasiveness, metastatic
potential, differentiation and apoptosis) and angiogenesis, all important factors in
cancer physiology [123].
The VDR is a cytosolic receptor that binds with calcitriol to form a complex
that further binds with the retinoid X receptor (RXR) to form a heterodimer. The
formation of the heterodimer is obligatory for translocation into the nucleus and
binding to the VDR response elements (VDRE) of certain genes [133]. The
genes in question can respond in three different ways: 1) bind to promoter
regions of certain genes, 2) bind to negative VDREs (nVDRE) or 3) antagonize
other transcription factors. As a result, multiple actions involving genes with
protein actions involved in bone remodeling, calcium binding, metabolism,
adhesion, anti-proliferation and differentiation may be achieved, with emphasis
on those that are anti-inflammatory and anti-proliferative in nature. The VDR
regulates numerous gene sequences. For instance, up-regulating gene
transcription of osteocalcin or down-regulating gene transcription of PTH [132]
stimulates osteoblasts to increase bone formation or allows for increased renal
elimination of Ca and P, respectively. The VDR-RXR heterodimer can also act
independently by directly inhibiting transcription factors such as the two nuclear
factors (NF): NF-AP and NF- B. The regulatory role of Vitamin D on certain
genes and their effects are summarized in Table 1.2.

48
Table 1.2 Genes influenced by Vitamin D receptor ligands and their effects.
Gene Protein Function
Anti-Inflammation
Anti-Proliferation
negVDRE
Osteocalcin Bone matrix protein
Osteopontin Bone matrix protein
RANKL Bone remodeling
IL-2 EGF-R PTH
CA II Bone remodeling
IL-12 c-myc PTHrP
Calbindin-9k Calcium binding
TNF- K16 Rel B
24-hydroxylase
Metabolism IFN-
mCYP3A1 Metabolism GM-CSF mCYP3A11 Metabolism hCYP3A4 Metabolism 3 integrin Adhesion
P21 Anti-Proliferation
Involucrin Differentiation PLC 1 Differentiation IGFBP-3 Anti-
Differentiation
Adapted from Nagpal et al 2005[133]
LEGEND IL = Interleukin TNF = Tumor Necrosis Factor IFN = Interferon GM-CSF = Granulocyte Macrophage Colony Stimulating Factor IGFBP = Insulin-like Growth Factor Binding Protein PLC = Phospholipase C EGF-R = Epidermal Growth Factor Receptor

49
The VDR has also been identified in multiple human tissues such as
epithelial cells of the epidermis, hair follicles, the female reproductive tract,
mammary gland, colon and lung; endocrine cells of the thyroid, pancreas and
ovary; in cardiac muscle cells; adipocytes and in cancer cell lines [132]. More
importantly, with regards to male reproduction, it has been found in rat and
human testicular tissue, seminiferous tubules and individual spermatozoa, along
with its associated activation enzymes [136-141]. VDR has also been isolated
and identified in rat and human prostatic tissue [139, 142, 143]. With regards to
female reproductive health, female calcitriol deficient mice experience hypoplasia
of the uterus and ovaries [144].
3.3.2 Vitamin D and the Prostate
Prostate cancer in men has been given high priority in the research
literature [4]. Recent cancer statistics reveal that prostate cancer is the second
leading diagnosed cancer in men and its mortality rate is second only to lung
cancer [145]. An American report in 2000 showed three out of four men had
chronic symptoms of BPH in their seventh decade while 6.5 million out of 27
million between the ages of 50 to 79 years sought treatment of symptoms [146].
The first strong epidemiological evidence linking Vitamin D to cancer risk,
through a negative association with sunlight exposure, was found by Garland et
al while looking at colorectal cancer in people [147]. In 1990, Schwartz and
Hulka (1990) hypothesized a similar relationship between prostate cancer and
geographical location showing a negative correlation to amount of ultraviolet

50
radiation exposure [148]. Hanchette and Schwartz (1992) later found evidence
to support this hypothesis while examining prostate cancer risk and geographical
distribution patterns in the ethnic white male population of the United States of
America [130]. This discovery resulted in extensive study of Vitamin D and its
role in human prostatic disease mainly prostatic carcinoma and later BPH [4].
In humans normal, cancerous and hyperplastic prostatic tissue have been
shown to have the VDR, 1 hydroxylase activity and 1,25(OH)2D3 intracellularly
[142]. Indeed, human prostate tissue itself, independent of kidney 1-hydroxylase
activity, can synthesize its own intracellularly active 1,25(OH)2D3 from 25OHD3
[134]. However, in one human prostatic cancer cell line it was revealed that 1
hydroxylase activity was absent and no anti-proliferative effects of Vitamin D
were noted. Furthermore, this study showed concentrations of 1,25(OH)2D3 in
normal and BPH tissue to be comparable to kidney tissue [134], suggesting an
adaptation might exist in some prostatic carcinomas to avoid the anti-proliferative
effects of Vitamin D by removal of this enzyme.
Prostatic carcinoma is relatively rare in both intact and neutered male
dogs with approximately 4% of the male dog population having subclinical
evidence of carcinoma according to the South African study by Mukaratira et al
(2007) [149]. BPH in this species has been accepted as having a similar
prevalence rate to that of men; greater than 80% in intact male dogs over the age
of four years in a review by Memon (2007) [150]. In an early study by Brendler,
BPH was confirmed histologically in 88% of beagles over the age of six years [1].
Atalan et al (1999) found 36/60 (60%) of the dogs in an ultrasound study of

51
prostate volume to have histological BPH while the rest of the dogs were
diagnosed by histology with normal prostates (17/60 or 28%), prostatic neoplasia
(4/60 or 7%) and prostatitis (3/60 or 5%) [94]. However, there is a lack of
prevalence studies using histology to diagnose BPH in the general intact dog
population and prevalence is based heavily on the findings in early beagle
studies and anecdotal evidence [151]. The limitation of prevalence studies is due
to the lack of invasive techniques used to definitively confirm non-clinical BPH.
The benign nature of the condition does not justify the costs or risks involved.
There is also a lack of defined prostate dimension parameters for different
bodyweights and ages when using ultrasonography to measure prostate size as
is present in BPH in men.
Vitamin D studies involving the prostate have concentrated on the effect of
the vitamin in cancerous tissue. Due to the risk of hypercalcemic effects that
occur with high doses of naturally occurring Vitamin D, several experimental
Vitamin D agonists have been developed which demonstrate high affinity for the
VDR in prostatic tissue without this potential side effect. These studies have
shown some promise in reducing PSA concentrations in early clinical trials [134,
152]. However, the effects of Vitamin D on BPH have only more recently
attracted attention [4, 153-155].
As stated earlier, BPH is an androgen dependent condition and Vitamin D
is thought to work downstream from the androgen receptor (AR) to inhibit
production of certain growth factors and pro-inflammatory cytokines [4, 133].
Crescioli et al (2003) studied cultured rat BPH cells using the experimental

52
Vitamin D analogue BXL-353 [153]. They found that, in the presence of 10
nmol/l testosterone, increasing doses of BXL-353 caused a dose-dependent
reduction in the number of BPH cells, as a percentage of pre-treatment numbers,
after 48 hours incubation. In the same study, BXL-353 AR receptor binding and
its ability to convert testosterone to DHT were also studied to determine whether
the mechanism of action was through the AR or 5 -reductase activity,
respectively. Finasteride-treated cell cultures were used as controls. BXL-353
demonstrated an inability to bind to the AR in the presence R1881 (a synthetic
androgen), through a competitive binding assay. No changes in testosterone
and DHT concentrations from prostatic cell cultures were observed for the
Vitamin D agonist while finasteride-treated controls maintained previously
recorded 50% inhibitory concentration for both isoforms of 5 -reductase. These
results indicate an androgen-independent mechanism for reduced cellular growth
and cellular apoptosis in BPH cells.
FGF-7 and IGF are implicated in the mechanism by which Vitamin D
exerts its action in BPH cells. Crescioli et al (2000) looked at FGF-7 action on
human BPH cells in culture [156]. First, they tested FGF-7 treatment alone on
BPH cell culture and found increased cell proliferation that proceeded in a dose-
dependent manner. Next they treated these cell cultures with both BXL-353 and
native Vitamin D and found an equal and significant decrease in the number of
BPH cells in the FGF-7 treated cultures for both the Vitamin D analogue and
Vitamin D. With this evidence, BXL-353 was further studied to determine
whether an effect on FGF-7-receptor signaling was present. They showed a

53
significant reduction in tyrosine phosphorylation of the FGF-7-receptor induced
by FGF-7 in culture, thereby inhibiting primary pathways in the stimulation of
gene expression relating to cell proliferation. This same group also investigated
the effect of the analogue on IGF-1 mediated cell proliferation and found that
BXL-353 was able to achieve this in a dose dependent manner [157]. Therefore,
action of Vitamin D and its analogues appear to be mediated through the
pathways down-stream of androgen action and target the paracrine factors
related to cell growth in prostatic tissue. Crescioli et al (2004) found similar
results with the experimental Vitamin D analogue BXL-628 (elocalcitol), yet had
less hypercalcemic effect than other Vitamin D analogues [154]. In pre-clinical
trials with elocalcitol, Adorini et al (2007) used a small group of male beagle dogs
(n=4) to study the effects of 5 /kg/day of elocalcitol for 9 months [158]. Prostate
weights, expressed as a percentage of prostate weight to bodyweight, were
reduced in this group, however these values did not reach statistical significance.
This may have been due to the low number of dogs used in the study. Of note,
serum calcium levels did not differ between pre-treatment and treatment values.
To date, no other canine studies related to Vitamin D analogues or their effects
on BPH are available.
Inflammation has been shown to be a factor in human BPH and may
explain many of the lower urinary tract symptoms experienced in men. In canine
studies to date, BPH is considered to be without the inflammatory component
present in the human form of the condition [1, 151, 159]. The similarities of BPH
between men and dogs in the spontaneous and cellular proliferation observed in

54
both has been the reason for the dog prostate being used as a primary model to
study in human pre-clinical pharmaceutical research. With the roles of Vitamin D
being elucidated in human research and the potential for treatment of the
condition of BPH being one of these roles, research into Vitamin D and BPH in
dogs is reasonable and warranted.
3.3.3 Role of Vitamin D in Sperm Production
The effects of Vitamin D on semen quantity and quality characteristics and
thus its possible role in male fertility have been studied. Early research in rats
showed that Vitamin D deficiency in male rats greatly affected their ability to
impregnate females [136]. The researchers found an absence of sperm in
vaginal swabs after mating in Vitamin D deficient rats, and a lower fertility rate
even in those where vaginal sperm were found after mating (45% pregnancy
rate), compared with their Vitamin D supplemented controls (73% pregnancy
rate). Vitamin D deficient male rats also had reduced testicular and epididymal
sperm count along with lower concentrations of Sertoli cell testicular glutamyl
transpeptidase activity from whole testes [160]. Histologically, decreased Leydig
cell numbers and degenerative changes in the germinal epithelium were noted in
Vitamin D rats compared to supplemented controls [160].
Further confirmation of a role for Vitamin D within testicular tissue
occurred with the identification of a nuclear VDR in the Sertoli cells [140]. Of
note, however, was that sperm count and sperm motility were dramatically
reduced in VDR null mice when compared with controls. Histologically, thinning

55
of the seminiferous epithelium, dilated seminiferous tubules and decreased or
infrequent spermatogenesis was noted in the testes of VDR null mice.
Interestingly, those VDR null mice that had calcium supplementation and normal
serum calcium concentrations did not show signs of decreased fertility.
In human males the VDR has been identified in the head of sperm cells
but is lacking in the neck and tail region [138]. Cholesterol efflux, a priming event
in the phosphorylation of proteins leading to human sperm capacitation, was
increased in the presence of calcitriol [161]. Also, an increase in phosphorylation
of tyrosine and threonine suggests that the VDR has a role in capacitation and
survival of sperm [143, 161]. Other researchers were able to identify the VDR
and the enzymes of Vitamin D metabolism in the human testis, epididymis,
prostate, and seminal vesicles in varying degrees [139].
The VDR gene is highly conserved among humans, mice and rats [162].
In particular, exons 1e, 1a and 1d of the human VDR gene are conserved in
multiple species, including the dog [163]. Although differences amongst species
exist it is reasonable to assume that the VDR function coded for in these areas
also conserved. Therefore presence of the VDR in similar tissues across species
may be associated with similar functions. Taken together, these studies suggest
an important role for Vitamin D in reproductive development and fertility in mice,
rats and humans. Although Vitamin D has not been investigated in dogs, there is
sufficient evidence to warrant research in this area.

56
3.4 PROLACTIN AND ITS ROLE IN MALE REPRODUCTION
Literature exists on the role of prolactin (PRL) in the female of multiple
species including women, rats and the bitch [5, 164] however PRL has only
recently attracted some interest in the literature regarding the male dog [165-
168]. The physiological role of PRL and its specific target organs have not been
fully identified in any species. Hyperprolactinemia, caused by a prolactinoma or
the use of some medications such as specific serotonin reuptake inhibitors
(SSRIs), is associated with symptoms of decreased libido and infertility in human
males [5, 139, 169].
3.4.1 Prolactin and Ovarian Physiology
Prolactin is a 23 kDa pituitary peptide hormone related to GH and
placental lactogen and produced by the lactotroph cells of the anterior pituitary
[5]. Just as with the other pituitary hormones like LH and FSH, study of PRL in
the male may prove that similar characteristics related to its regulation and action
are conserved not just among species but also between the two sexes. Prolactin
secretion is pulsatile in women and pregnant rats, following a diurnal pattern, and
thought to be regulated by the influence of the light/dark cycle on the regulatory
suprachiasmatic nucleus (SCN) of the brain [5]. Although pulsatile, a 24-hour
circadian rhythm is absent in the bitch [168]. The pattern of secretion in the dog
follows an ultradian and circannual rhythm of secretion with higher concentrations
occurring during periods of increased light [168]. The SCN influences the

57
hypothalamus (HTH) by stimulating the release of dopamine (DA) – the major
inhibitory factor of PRL release. Alternatively, oxytocin acting on the
periventricular nucleus (PVN) has been shown to be a major stimulus for PRL
release. Nerve fibres from the SCN have connections with both the HTH and
PVN oxytocin-containing neurons in rats [170]. Interestingly, in pregnant women
PRL is also produced in extra-pituitary sites, mainly by the decidua, while in
bitches there is no evidence of a source other than the pituitary [171, 172].
With respect to comparative physiology between the ovary and testes it is
important to note that PRL action in the ovary has been linked to influencing
enzymes involved in steroid hormone conversion causing decreases in E2 and
increasing P4 production in the granulosa cells through down-regulation of 20 -
hydroxysteroid dehydrogenase and up-regulation of LH-R in these cells to
maintain the corpus luteum. In two studies of fertility in female PRL and PRL
receptor (PRL-R) knock-out mice, researchers found that the PRL knock-out
group experienced normal cycles and ovulatory patterns but could not maintain
pregnancy [173] while the PRL-R knock-out mice had reduced ovulation,
fertilization and arrest of pre-implantation development [174]. It is the
mechanisms of PRL action at the cellular level where parallels may exist between
female and male physiology showing similar patterns in Leydig cells and/or
Sertoli cells.

58
3.4.2 Prolactin and Male Physiology
3.4.2.1 Prolactin and the Prostate
Considerable evidence exists for a role of PRL in prostate physiology.
Human and rat prostatic epithelial tissue expresses both PRL and its receptor
PRLR [175, 176]. Early studies on the canine prostate detected intracellular
endogenous PRL and binding sites for exogenous PRL within prostatic epithelial
cells [177]. PRL has been shown to be a necessary component for prostatic
epithelial growth and survival in human and rat tissue culture [5, 178]. Original
studies in mice found that hyperprolactinemia increased in the weight of the
prostate gland, by nine to twenty times, as well as histological evidence of
prostatic hyperplasia [179]. Both in vivo and in vitro studies showed PRL
affected growth and differentiation of the prostate [180] and sensitized the
prostatic epithelial cells to androgen effects. This was thought to occur due to
synergism between testosterone and PRL producing an increase in 5 -reductase
activity [181]. These same changes were not noted in canine studies. Increased
cell proliferation was seen only in those cultures supplemented with bovine and
dog serum alone and not with any steroid hormones or PRL [182]. However,
clinical studies of 5 -reductase inhibitors have shown a dramatic effect in size
reduction of the canine prostate through decreased DHT concentrations [115].
Helmerich et al (1976) showed that pretreatment with PRL significantly
decreased prostatic tissue concentrations of DHT and subsequently increased
tissue testosterone [183]. To determine a PRL effect on prostatic androgen

59
metabolism investigators used recently (24 hours) castrated dogs and divided
them into three groups: 1) control, 2) pre-treatment for 3 days with 500 IU PRL
intramuscularly and 3) pre-treatment with 5 mg bromocriptine per os for 3 days.
All dogs were treated at 96 hours post-castration with tritiated testosterone (H3-
T), sacrificed one-hour post H3-T treatment and prostate tissue was harvested
and measured for testosterone and DHT concentrations. PRL pretreatment
significantly increased testosterone concentration over DHT when compared with
controls. Pretreatment with bromocriptine (a dopamine agonist), significantly
increased prostatic DHT and decreased testosterone compared with controls.
Dopamine agonists are compounds that bind to dopamine receptors and cause
increased or enhanced dopamine effects and therefore PRL secretion is
decreased due to the inhibitory action of dopaminergic stimulation. As a result,
the study showed that PRL, through some mechanism, decreases conversion of
testosterone to DHT; the hormone known to be primarily responsible for
hyperplastic changes in the prostate.
Robertson et al (2003) determined that the castration-induced regression
of the prostate in PRLR knockout mice was greater than in the normal controls,
suggesting that PRL and testosterone act together in development of the ventral
lobe of the prostate. The authors suggested PRL has a regulating effect of
prostate development under normal physiological conditions of testosterone
[184]. In PRLR null mice the ventral half of the prostate was 20% heavier
compared to controls. However, the ratio of epithelial cells to stroma within the
dorsal lobes was decreased in PRLR null mice. This difference in size between

60
PRL null and control mice disappeared at one year of age suggesting there is a
transient affect of PRL during prostate development.
Hyperprolactinemia due to prolactinomas in men has not been shown to
increase prostate size [185]. Although there was no correlation between
testosterone and prostate size in patients with prolactinoma, they had
significantly smaller prostates and lower testosterone concentrations than their
normal age-matched controls, suggesting lowered androgen levels in these
patients contributes to a decrease in prostate size. This effect is most likely PRL-
mediated inhibition of androgen production [186], however further study is
warranted.
Conflicting findings on the role of PRL in male prostate physiology are
apparent in the literature. This may be attributed to dose-dependent effects,
chronicity of diseases, as seen with other pituitary hormones such as GnRH
agonists or species-specific differences in prostate physiology. The PRL
feedback loop appears to have increased delays when compared with other
hormones [170, 187]. Nevertheless, PRL does appear to have a role in prostatic
physiology. Studies of prostatic conditions such as BPH and cancer in dogs
should therefore include PRL as a hormone of interest.
3.4.2.2 Prolactin, Male Fertility and Semen Quality
Hyperprolactinemia is associated with decreased semen quality and libido
in both rodents and men [185, 186, 188-191] and, depending on the severity of
the hyperprolactinemia and chronicity of the condition, hypogonadism can be

61
present. The main effects on semen quality include oligozoospermia,
asthenozoospermia and teratozoospermia in these species [185, 186, 188-191].
Many of these conditions can be corrected using dopamine agonists or PRL
antagonists (see review by De Rosa et al (2003) [192]). This impaired
reproductive function in pathological conditions of PRL does not explain normal
reproductive function, yet it has aided in its understanding. A study by Shafik et
al (1994) used supra-physiological doses of ovine PRL (600 g/kg/week IM for 6
months) to induce sterility in dogs [193]. By the end of the six-month study all
dogs were azoospermic. At three months declines in total motility (<70%) and
normal morphology (<60%) were observed, however, no other semen
parameters were studied. It is important to note that by three months after
cessation of treatments semen numbers, motility and normal morphology
returned to pre-treatment values. This study looked at contraceptive effects of
PRL in the dog, so it is unknown whether naturally occurring hyperprolactinemia
in the dog causes infertility and whether native canine PRL has a similar effect,
however, these results are similar to those in men experiencing
hyperprolactinemia.
In contrast to the female, PRL deficiency appears to have no effect on
male fertility. In a study by Steger et al (1998), mature (2-3 months old) male
PRL null and control mice were individually housed for three weeks with two
mature virgin females [194]. Fertility was measured by percentage of live litters
produced from each group. From the control group 17/18 (94%) females
delivered live litters and 32/33 (97%) delivered litters from the PRL null group

62
while litter size did not differ between the two groups (9.68 ± 0.39 and 10.06 ±
0.63 for PRL null and control males, respectively). It is unknown whether any
compensable defects in semen quality were present that might have been
overcome through increased normal sperm numbers via increased number of
matings. Long-term effects of PRL deficiency from this study were also not
accounted for. PRL deficiency may also impact fertility rates where issues of
subfertility in the female are in question, however it is possible that PRL effects
enhance but are not necessary to normal reproductive physiology in the male.
A recent review by Gill-Sharma (2009) details the feedback mechanisms
and action of PRL in the male in both the rat and human [186]. PRL has both a
short and long feedback loop involving the HPTA and the hypophyseal-pituitary-
gonadal axis and appears to be complex.
Although there appear to be multiple regulators of PRL in the female the
only regulator in the male of any importance to date appears to be dopamine.
Dopamine, a neuropeptide hormone, is a potent inhibitor of PRL secretion and is
hypothalamic in origin. PRL down-regulates its own secretion by stimulating
dopaminergic neurons to inhibit its own release. PRL primarily acts directly on
the testis, chiefly the interstitial cells of Leydig [195]. It up-regulates and
increases affinity of LH receptors in Leydig cells [195], causing an increase in
testosterone secretion, although a direct role in increasing serum testosterone is
still controversial [196, 197]. Testosterone produced by Leydig cells then acts on
Sertoli cells to produce E2 and increases production of androgen binding proteins

63
(ABP) [198]. Testosterone then acts via hypothalamic (dopamine) and pituitary
routes to inhibit PRL secretion.
Stimulators of PRL secretion include the following components: Serotonin,
hin, Met enkephalin, Leu enkephalin, thyrotropin-releasing hormone
(TRH), GnRH, substance P, E2, epidermal growth factor, fibroblast growth factor,
cholecystokinin, angiotensin II, and prolactin-releasing peptides (PrRPs) by direct
action at the pituitary level [199-208]. The main hormone of PRL up-regulation,
however, appears to be E2 through aromatization of testosterone. Estradiol acts
directly on lactotrophs but also has a negative feedback on GnRH causing
reduced LH and FSH secretion. In addition, PRL also has an inhibitory effect on
GnRH[209]. The presence of high concentrations of PRL and E2 seen in
hyperprolactinemia is thought to be the mechanism by which serum testosterone,
LH, and FSH are lowered in these conditions. At physiological concentrations E2
feedback does not affect testosterone concentrations mainly due to PRL action
on Leydig cells.
Prolactin reference ranges and ultradian and circannual patterns of PRL
secretion have been published for the male dog [165, 167, 168, 210], although a
difference in the type of assay used in these studies should be noted. The
ranges published by Corrada et al (2006), using homologous enzyme
immunometric assay [167], are summarized in Table 1.3. Of note, Beagle dogs
had, on average, much higher PRL concentrations than both the crossbred dogs
and German Shepherd Dogs sampled in this study. This same group was able to
show a circannual variation in PRL concentrations with an association of higher

64
concentrations with increased daylight hours (November, December and January
compared to May, June, July in the southern hemisphere). Kreeger et al (1992)
recorded nadir during the fall months of October and November [210], but taking
latitude into account, both studies saw similar patterns with respect to hours of
daylight.
Although mean PRL concentration measured using radioimmunoassay
(RIA) did not differ widely among breeds in individuals with normospermia,
Urhausen et al (2009) saw a significant difference between these values in Fox
Terriers compared with Great Danes, with the latter being lower [165]. It was
difficult to draw breed specific differences, however, due to the inbreeding of the
Fox Terriers enrolled in the study. A sharp increase in PRL concentrations was
observed after thyroid stimulating hormone (TSH) stimulation in this same study.
Mean values of PRL remained within the range specified by Corrada et al (2006),
yet it is important to note that sampling was only done once prior to TSH
stimulation [165]. Therefore normal PRL fluctuation was not accounted for.
Although semen quality parameters are known to be adversely affected in human
males with hyperprolactinemia, the study by Koivisto et al (2009) determined that
semen parameters and libido remained unchanged after induced short-term
hyperprolactinemia [211]. In this study, six male beagle dogs between the ages
of 12 months to two years were divided into five three-week treatment periods: 1)
pre-treatment, 2) metoclopramide 2mg/kg/ q8h po, 3) cabergoline 5 g/kg q24h
po, 4) post-treatment period 1 and 5) post-treatment period 2. Fasting blood
samples were collected in the morning twice weekly and hormone analysis was

65
done using RIA. PRL levels significantly increased with metoclopramide
treatment (mean ± SD: 6.5 ± 1.6 ng/ml (p < 0.05)), a known stimulator of PRL
secretion through dopamine antagonism, compared with pre-treatment (mean ±
SD: 4.5 ± 1.1 ng/ml) values. Cabergoline treatment resulted in a decrease in
PRL concentration (mean ± SD: 3.0 ± 0.6 ng/ml) compared with pre-treatment
values (p < 0.05). Post-treatment values did not differ from pre-treatment ones
(mean ± SD 4.4 ± 0.8 ng/ml). It is important to note that although the
investigators described the mean elevation in PRL as mild hyperprolactinemia it
does not appear to be significantly different from the high range of normal
determined previously (6.0 ng/ml). Indeed, mild hyperprolactinemia in men is
defined as PRL levels in excess of 20 ng/ml [188], although it is difficult to
extrapolate these values among species. It is also unknown whether a three-
week period of induced hyperprolactinemia may accurately describe possible
long-term effects of PRL on semen quality.
Table 1.3 Prolactin values in Dogs (ELISA)
Range (Mean SE) (ng/ml) 0 - 6.0 (2.7 0.2)
Mean Baseline SE (ng/ml) 1.4 0.6
Pulse frequency (peaks/6hr) 1-2
Pulse duration (mins) (Mean SE) 15-75 (45 11)
Pulse amplitude (ng/ml) (Mean SE) 1.7-2.4 (1.7 0.4)
Adapted from Corrada et al 2006[167]

66
It appears that the few studies of PRL and fertility conducted in male dogs
seem to correlate well with the findings of most human and rat studies. The
presence of PRL and its receptors in prostate tissue in all species, influences of
PRL on prostatic androgen metabolism and semen parameters in states of
excessive hyperprolactinemia appear to be conserved. However, the in vivo and
cellular mechanisms of PRL physiology in the prostate and in spermatozoal
development, especially in the dog, require further investigation.

67
References: [1] C. B. Brendler, et al., "Spontaneous benign prostatic hyperplasia in the
beagle. Age-associated changes in serum hormone levels, and the morphology and secretory function of the canine prostate," J Clin Invest, vol. 71, pp. 1114-23, May 1983.
[2] R. A. Read and S. Bryden, "Urethral bleeding as a presenting sign of benign prostatic hyperplasia in the dog: a retrospective study (1979-1993)," J Am Anim Hosp Assoc, vol. 31, pp. 261-7, May-Jun 1995.
[3] D. R. Krawiec and D. Heflin, "Study of prostatic disease in dogs: 177 cases (1981-1986)," J Am Vet Med Assoc, vol. 200, pp. 1119-22, Apr 15 1992.
[4] L. Adorini, et al., "Vitamin D receptor agonists target static, dynamic, and inflammatory components of benign prostatic hyperplasia," Ann N Y Acad Sci, vol. 1193, pp. 146-52, Apr 2010.
[5] A. Bachelot and N. Binart, "Reproductive role of prolactin," Reproduction, vol. 133, pp. 361-9, Feb 2007.
[6] M. Blomberg Jensen, et al., "Vitamin D is positively associated with sperm motility and increases intracellular calcium in human spermatozoa," Hum Reprod, vol. 26, pp. 1307-17, Jun 2011.
[7] M. De Rosa, et al., "Six months of treatment with cabergoline restores sexual potency in hyperprolactinemic males: an open longitudinal study monitoring nocturnal penile tumescence," J Clin Endocrinol Metab, vol. 89, pp. 621-5, Feb 2004.
[8] S. D. Johnston, et al., Sexual differentiation and normal anatomy of the dog 1st. ed. Philadelphia: WB Saunders, 2001.
[9] D. E. Noakes, et al., Reproduction in Male Animals 8th ed. London: WB Saunders, 2001.
[10] L. D. Russell and J. P. Malone, "A study of Sertoli-spermatid tubulobulbar complexes in selected mammals," Tissue Cell, vol. 12, pp. 263-85, 1980.
[11] M. Cambrosio Mann, et al., "Blood-tissue barriers in the male reproductive tract of the dog: a morphological study using lanthanum nitrate as an electron-opaque tracer," Cells Tissues Organs, vol. 174, pp. 162-9, 2003.
[12] C. Y. Cheng, et al., "Regulation of spermatogenesis in the microenvironment of the seminiferous epithelium: new insights and advances," Mol Cell Endocrinol, vol. 315, pp. 49-56, Feb 5 2010.
[13] H. E. Duarte, et al., "Ultrastructural characteristics of the testicular capillaries in the dog (Canis familiaris, L.)," Anat Histol Embryol, vol. 24, pp. 73-6, Jun 1995.
[14] K. L. Hull and S. Harvey, "Growth hormone: roles in male reproduction," Endocrine, vol. 13, pp. 243-50, Dec 2000.
[15] S. K. De, et al., "Expression of tumor necrosis factor-alpha in mouse spermatogenic cells," Endocrinology, vol. 133, pp. 389-96, Jul 1993.

68
[16] M. K. Siu, et al., "The interplay of collagen IV, tumor necrosis factor-alpha, gelatinase B (matrix metalloprotease-9), and tissue inhibitor of metalloproteases-1 in the basal lamina regulates Sertoli cell-tight junction dynamics in the rat testis," Endocrinology, vol. 144, pp. 371-87, Jan 2003.
[17] C. Mauduit, et al., "Tumor necrosis factor alpha receptor p55 is under hormonal (follicle-stimulating hormone) control in testicular Sertoli cells," Biochem Biophys Res Commun, vol. 224, pp. 631-7, Jul 25 1996.
[18] C. Y. Hong, et al., "Molecular mechanism of suppression of testicular steroidogenesis by proinflammatory cytokine tumor necrosis factor alpha," Mol Cell Biol, vol. 24, pp. 2593-604, Apr 2004.
[19] M. S. Theas, et al., "Tumour necrosis factor-alpha released by testicular macrophages induces apoptosis of germ cells in autoimmune orchitis," Hum Reprod, vol. 23, pp. 1865-72, Aug 2008.
[20] F. J. Delfino, et al., "NF-kappaB and TNF-alpha stimulate androgen receptor expression in Sertoli cells," Mol Cell Endocrinol, vol. 201, pp. 1-12, Mar 28 2003.
[21] B. Le Magueresse-Battistoni, et al., "Tumor necrosis factor-alpha regulates plasminogen activator inhibitor-1 in rat testicular peritubular cells," Endocrinology, vol. 138, pp. 1097-105, Mar 1997.
[22] Y. X. Liu, "Involvement of plasminogen activator and plasminogen activator inhibitor type 1 in spermatogenesis, sperm capacitation, and fertilization," Semin Thromb Hemost, vol. 33, pp. 29-40, Feb 2007.
[23] M. W. Li, et al., "Tumor necrosis factor {alpha} reversibly disrupts the blood-testis barrier and impairs Sertoli-germ cell adhesion in the seminiferous epithelium of adult rat testes," J Endocrinol, vol. 190, pp. 313-29, Aug 2006.
[24] W. Y. Lui, et al., "TGF-betas: their role in testicular function and Sertoli cell tight junction dynamics," Int J Androl, vol. 26, pp. 147-60, Jun 2003.
[25] W. Xia, et al., "TGF-beta3 and TNFalpha perturb blood-testis barrier (BTB) dynamics by accelerating the clathrin-mediated endocytosis of integral membrane proteins: a new concept of BTB regulation during spermatogenesis," Dev Biol, vol. 327, pp. 48-61, Mar 1 2009.
[26] P. P. Lie, et al., "The biology of interleukin-1: emerging concepts in the regulation of the actin cytoskeleton and cell junction dynamics," Cell Mol Life Sci, Jul 9 2011.
[27] C. Petersen, et al., "Interleukin-1 is a potent growth factor for immature rat sertoli cells," Mol Cell Endocrinol, vol. 186, pp. 37-47, Jan 15 2002.
[28] P. Pollanen, et al., "Interleukin-1 alpha stimulation of spermatogonial proliferation in vivo," Reprod Fertil Dev, vol. 1, pp. 85-7, 1989.
[29] M. Bialas, et al., "The role of IL-6, IL-10, TNF-alpha and its receptors TNFR1 and TNFR2 in the local regulatory system of normal and impaired human spermatogenesis," Am J Reprod Immunol, vol. 62, pp. 51-9, Jul 2009.
[30] C. K. Jonsson, et al., "Constitutive expression of interleukin-1alpha messenger ribonucleic acid in rat Sertoli cells is dependent upon

69
interaction with germ cells," Endocrinology, vol. 140, pp. 3755-61, Aug 1999.
[31] K. V. Svechnikov, et al., "Age-dependent stimulation of Leydig cell steroidogenesis by interleukin-1 isoforms," Mol Cell Endocrinol, vol. 182, pp. 193-201, Sep 2001.
[32] E. Colon, et al., "Stimulation of steroidogenesis in immature rat Leydig cells evoked by interleukin-1alpha is potentiated by growth hormone and insulin-like growth factors," Endocrinology, vol. 146, pp. 221-30, Jan 2005.
[33] O. Sarkar, et al., "Interleukin 1 alpha (IL1A) is a novel regulator of the blood-testis barrier in the rat," Biol Reprod, vol. 78, pp. 445-54, Mar 2008.
[34] M. Kanzaki and P. L. Morris, "Growth hormone regulates steroidogenic acute regulatory protein expression and steroidogenesis in Leydig cell progenitors," Endocrinology, vol. 140, pp. 1681-6, Apr 1999.
[35] S. J. Gelber, et al., "Effects of insulin-like growth factor-I on androgen production by highly purified pubertal and adult rat Leydig cells," J Androl, vol. 13, pp. 125-30, Mar-Apr 1992.
[36] F. Chuzel, et al., "Transcriptional regulation of the lutropin/human choriogonadotropin receptor and three enzymes of steroidogenesis by growth factors in cultured pig Leydig cells," Eur J Biochem, vol. 239, pp. 8-16, Jul 1 1996.
[37] M. J. Yoon and J. F. Roser, "A synergistic effect of insulin-like growth factor (IGF-I) on equine luteinizing hormone (eLH)-induced testosterone production from cultured Leydig cells of horses," Anim Reprod Sci, vol. 126, pp. 195-9, Jul 2011.
[38] M. F. Hess and J. F. Roser, "A comparison of the effects of equine luteinizing hormone (eLH), equine growth hormone (eGH) and human recombinant insulin-like growth factor (hrIGF-I) on steroid production in cultured equine Leydig cells during sexual maturation," Anim Reprod Sci, vol. 89, pp. 7-19, Oct 2005.
[39] R. A. Hess and L. Renato de Franca, "Spermatogenesis and cycle of the seminiferous epithelium," Adv Exp Med Biol, vol. 636, pp. 1-15, 2008.
[40] E. W. Wong and C. Y. Cheng, "Polarity proteins and cell-cell interactions in the testis," Int Rev Cell Mol Biol, vol. 278, pp. 309-53, 2009.
[41] Q. Zhou and M. D. Griswold, "Regulation of spermatogonia," 2008. [42] R. H. Foote, et al., "Spermatogenesis in the dog," Anat Rec, vol. 173, pp.
341-51, Jul 1972. [43] L. D. Russell, Ed., Histological and histopathological evaluation of the
testis. Clearwater, FL: Cache River Press, 1990, p.^pp. Pages. [44] T. T. Olar, et al., "Relationships among testicular size, daily production
and output of spermatozoa, and extragonadal spermatozoal reserves of the dog," Biol Reprod, vol. 29, pp. 1114-20, Dec 1983.
[45] N. R. Santos, et al., "The knobbed acrosome defect in four closely related dogs," Theriogenology, vol. 66, pp. 1626-8, Oct 2006.
[46] R. W. James and R. Heywood, "Age-related variations in the testes and prostate of beagle dogs," Toxicology, vol. 12, pp. 273-9, Mar-Apr 1979.

70
[47] L. A. Lowseth, et al., "Age-related changes in the prostate and testes of the beagle dog," Vet Pathol, vol. 27, pp. 347-53, Sep 1990.
[48] M. A. Peters, et al., "Spermatogenesis and testicular tumours in ageing dogs," J Reprod Fertil, vol. 120, pp. 443-52, Nov 2000.
[49] J. H. Boucher, et al., "The evaluation of semen quality in the dog and the effects of frequency of ejaculation upon semen quality, libido, and depletion of sperm reserves," Cornell Vet, vol. 48, pp. 67-86, Jan 1958.
[50] G. C. England, "Semen quality in dogs and the influence of a short-interval second ejaculation," Theriogenology, vol. 52, pp. 981-6, Oct 15 1999.
[51] S. D. Johnston, et al., Semen Collection, Evaluation and Preservation, 1st ed. Philadelphia: WB Saunders, 2001.
[52] A. I. Martinez, "Canine fresh and cryopreserved semen evaluation," Anim Reprod Sci, vol. 82-83, pp. 209-24, Jul 2004.
[53] T. Rijsselaere, et al., "New techniques for the assessment of canine semen quality: a review," Theriogenology, vol. 64, pp. 706-19, Aug 2005.
[54] J. L. Freshman, "Semen collection and evaluation," Clin Tech Small Anim Pract, vol. 17, pp. 104-7, Aug 2002.
[55] M. V. Root Kustritz, "The value of canine semen evaluation for practitioners," Theriogenology, vol. 68, pp. 329-37, Aug 2007.
[56] J. Verstegen, et al., "Computer assisted semen analyzers in andrology research and veterinary practice," Theriogenology, vol. 57, pp. 149-79, Jan 1 2002.
[57] B. W. Pickett, et al., "Reproductive physiology of the stallion. VI. Seminal and behavioral characteristics," J Anim Sci, vol. 43, pp. 617-25, Sep 1976.
[58] L. Multigner, et al., "Secular sperm trends in stallions between 1981 and 1996," J Androl, vol. 20, pp. 763-8, Nov-Dec 1999.
[59] D. E. Wildt, et al., "Influence of inbreeding on reproductive performance, ejaculate quality and testicular volume in the dog," Theriogenology, vol. 17, pp. 445-52, Apr 1982.
[60] S. Schafer-Somi and C. Aurich, "Use of a new computer-assisted sperm analyzer for the assessment of motility and viability of dog spermatozoa and evaluation of four different semen extenders for predilution," Anim Reprod Sci, vol. 102, pp. 1-13, Nov 2007.
[61] C. Kuster, "Sperm concentration determination between hemacytometric and CASA systems: why they can be different," Theriogenology, vol. 64, pp. 614-7, Aug 2005.
[62] D. H. Douglas-Hamilton, et al., "Capillary-loaded particle fluid dynamics: effect on estimation of sperm concentration," J Androl, vol. 26, pp. 115-22, Jan-Feb 2005.
[63] W. Ombelet, et al., "Sperm morphology assessment: historical review in relation to fertility," Hum Reprod Update, vol. 1, pp. 543-57, Nov 1995.
[64] R. G. Saacke, et al., "Relationship of seminal traits and insemination time to fertilization rate and embryo quality," Anim Reprod Sci, vol. 60-61, pp. 663-77, Jul 2 2000.

71
[65] A. Al-Makhzoomi, et al., "Sperm morphology and fertility of progeny-tested AI dairy bulls in Sweden," Theriogenology, vol. 70, pp. 682-91, Sep 1 2008.
[66] P. J. Chenoweth, "Genetic sperm defects," Theriogenology, vol. 64, pp. 457-68, Aug 2005.
[67] M. Enciso, et al., "Major morphological sperm abnormalities in the bull are related to sperm DNA damage," Theriogenology, vol. 76, pp. 23-32, Jul 1 2011.
[68] E. E. Oettle, "Sperm morphology and fertility in the dog," J Reprod Fertil Suppl, vol. 47, pp. 257-60, 1993.
[69] E. Blom, "[The ultrastructure of some characteristic sperm defects and a proposal for a new classification of the bull spermiogram (author's transl)]," Nord Vet Med, vol. 25, pp. 383-91, Jul-Aug 1973.
[70] T. Rijsselaere, et al., "Effect of body weight, age and breeding history on canine sperm quality parameters measured by the Hamilton-Thorne analyser," Reprod Domest Anim, vol. 42, pp. 143-8, Apr 2007.
[71] S. D. Johnston, Root Kustritz M.V., Olsen, P.N.S., Clinical approach to infertility in the bitch., 1st ed. Philadelphia: WB Saunders, 2001.
[72] A. I. Pena, et al., "Infertility in a dog due to proximal cytoplasmic droplets in the ejaculate: investigation of the significance for sperm functionality in vitro," Reprod Domest Anim, vol. 42, pp. 471-8, Oct 2007.
[73] M. Dahlbom, et al., "Morphometry of normal and teratozoospermic canine sperm heads using an image analyzer: work in progress," Theriogenology, vol. 48, pp. 687-98, Sep 1997.
[74] I. Nunez-Martinez, et al., "Identification of sperm morphometric subpopulations in the canine ejaculate: do they reflect different subpopulations in sperm chromatin integrity?," Zygote, vol. 15, pp. 257-66, Aug 2007.
[75] G. C. Ostermeier, et al., "Measurement of bovine sperm nuclear shape using Fourier harmonic amplitudes," J Androl, vol. 22, pp. 584-94, Jul-Aug 2001.
[76] P. Kathiravan, et al., "Objective sperm motion analysis to assess dairy bull fertility using computer-aided system--a review," Reprod Domest Anim, vol. 46, pp. 165-72, Feb 2011.
[77] M. Iguer-Ouada and J. P. Verstegen, "Validation of the sperm quality analyzer (SQA) for dog sperm analysis," Theriogenology, vol. 55, pp. 1143-58, Mar 15 2001.
[78] M. Iguer-ouada and J. P. Verstegen, "Evaluation of the "Hamilton Thorn computer-based automated system" for dog semen analysis," Theriogenology, vol. 55, pp. 733-49, Feb 1 2001.
[79] P. B. Farrell, et al., "Quantification of bull sperm characteristics measured by computer-assisted sperm analysis (CASA) and the relationship to fertility," Theriogenology, vol. 49, pp. 871-9, Mar 1998.
[80] C. C. Love, "Relationship between sperm motility, morphology and the fertility of stallions," Theriogenology, vol. 76, pp. 547-57, Aug 2011.

72
[81] Y. Hirano, et al., "Relationships between sperm motility characteristics assessed by the computer-aided sperm analysis (CASA) and fertilization rates in vitro," J Assist Reprod Genet, vol. 18, pp. 213-8, Apr 2001.
[82] R. J. Aitken, "Diagnostic value of the zona-free hamster oocyte penetration test and sperm movement characteristics in oligozoospermia," Int J Androl, vol. 8, pp. 348-56, Oct 1985.
[83] C. Jeulin, et al., "Sperm factors related to failure of human in-vitro fertilization," J Reprod Fertil, vol. 76, pp. 735-44, Mar 1986.
[84] T. A. Bongso, et al., "Effect of sperm motility on human in vitro fertilization," Arch Androl, vol. 22, pp. 185-90, 1989.
[85] J. Ellington, et al., "Computer-assisted sperm analysis of canine spermatozoa motility measurements," Theriogenology, vol. 40, pp. 725-33, Oct 1993.
[86] J. Smith, "Canine prostatic disease: a review of anatomy, pathology, diagnosis, and treatment," Theriogenology, vol. 70, pp. 375-83, Aug 2008.
[87] R. Schoonees, et al., "The effect of androgen on canine prostatic secretion," J Surg Oncol, vol. 4, pp. 169-79, 1972.
[88] C. Gobello, et al., "Serum and seminal markers in the diagnosis of disorders of the genital tract of the dog: a mini-review," Theriogenology, vol. 57, pp. 1285-91, Mar 1 2002.
[89] J. E. Branam, et al., "Selected physical and chemical characteristics of prostatic fluid collected by ejaculation from healthy dogs and from dogs with bacterial prostatitis," Am J Vet Res, vol. 45, pp. 825-9, Apr 1984.
[90] A. Bruengger, et al., "Smooth muscle cell of the canine prostate in spontaneous benign hyperplasia, steroid induced hyperplasia and estrogen or tamoxifen treated dogs," J Urol, vol. 130, pp. 1208-10, Dec 1983.
[91] S. D. Johnston, et al., "Prostatic disorders in the dog," Anim Reprod Sci, vol. 60-61, pp. 405-15, Jul 2 2000.
[92] S. J. Berry, et al., "Effects of aging on prostate growth in beagles," Am J Physiol, vol. 250, pp. R1039-46, Jun 1986.
[93] Y. Ruel, et al., "Ultrasonographic evaluation of the prostate in healthy intact dogs," Vet Radiol Ultrasound, vol. 39, pp. 212-6, May-Jun 1998.
[94] G. Atalan, et al., "Ultrasonographic estimation of prostatic size in canine cadavers," Res Vet Sci, vol. 67, pp. 7-15, Aug 1999.
[95] D. P. DeKlerk, et al., "Comparison of spontaneous and experimentally induced canine prostatic hyperplasia," J Clin Invest, vol. 64, pp. 842-9, Sep 1979.
[96] W. Bartsch, et al., "Enzymes of androgen formation and degradation in the human prostate," Ann N Y Acad Sci, vol. 595, pp. 53-66, 1990.
[97] V. Grieco, et al., "The distribution of oestrogen receptors in normal, hyperplastic and neoplastic canine prostate, as demonstrated immunohistochemically," J Comp Pathol, vol. 135, pp. 11-6, Jul 2006.
[98] F. Gallardo-Arrieta, et al., "Ultrastructural changes in prostate cells during hormone-induced canine prostatic hyperplasia," Ultrastruct Pathol, vol. 30, pp. 435-42, Nov-Dec 2006.

73
[99] M. L. Winter and J. G. Liehr, "Possible mechanism of induction of benign prostatic hyperplasia by estradiol and dihydrotestosterone in dogs," Toxicol Appl Pharmacol, vol. 136, pp. 211-9, Feb 1996.
[100] K. L. Lee and D. M. Peehl, "Molecular and cellular pathogenesis of benign prostatic hyperplasia," J Urol, vol. 172, pp. 1784-91, Nov 2004.
[101] M. C. Pollan, et al., "Growth factor expression in early stages of benign prostatic hyperplasia upon exposure to sustained delivery of androgens," Biomed Sci Instrum, vol. 39, pp. 329-34, 2003.
[102] G. Yan, et al., "Heparin-binding keratinocyte growth factor is a candidate stromal-to-epithelial-cell andromedin," Mol Endocrinol, vol. 6, pp. 2123-8, Dec 1992.
[103] F. Ropiquet, et al., "FGF7 and FGF2 are increased in benign prostatic hyperplasia and are associated with increased proliferation," J Urol, vol. 162, pp. 595-9, Aug 1999.
[104] M. T. Story, et al., "Characteristics of FGF-receptors expressed by stromal and epithelial cells cultured from normal and hyperplastic prostates," Growth Factors, vol. 10, pp. 269-80, 1994.
[105] M. Ittman and A. Mansukhani, "Expression of fibroblast growth factors (FGFs) and FGF receptors in human prostate," J Urol, vol. 157, pp. 351-6, Jan 1997.
[106] D. Giri and M. Ittmann, "Interleukin-1alpha is a paracrine inducer of FGF7, a key epithelial growth factor in benign prostatic hyperplasia," Am J Pathol, vol. 157, pp. 249-55, Jul 2000.
[107] D. Giri and M. Ittmann, "Interleukin-8 is a paracrine inducer of fibroblast growth factor 2, a stromal and epithelial growth factor in benign prostatic hyperplasia," Am J Pathol, vol. 159, pp. 139-47, Jul 2001.
[108] M. Caine, "Alpha-adrenergic mechanisms in dynamics of benign prostatic hypertrophy," Urology, vol. 32, pp. 16-20, Dec 1988.
[109] N. Kyprianou, et al., "Apoptotic versus proliferative activities in human benign prostatic hyperplasia," Hum Pathol, vol. 27, pp. 668-75, Jul 1996.
[110] D. W. Cohen, et al., "Expression of transforming growth factor-alpha and the epidermal growth factor receptor in human prostate tissues," J Urol, vol. 152, pp. 2120-4, Dec 1994.
[111] P. Cohen, et al., "Transforming growth factor-beta induces growth inhibition and IGF-binding protein-3 production in prostatic stromal cells: abnormalities in cells cultured from benign prostatic hyperplasia tissues," J Endocrinol, vol. 164, pp. 215-23, Feb 2000.
[112] G. Dong, et al., "Decreased expression of Wilms' tumor gene WT-1 and elevated expression of insulin growth factor-II (IGF-II) and type 1 IGF receptor genes in prostatic stromal cells from patients with benign prostatic hyperplasia," J Clin Endocrinol Metab, vol. 82, pp. 2198-203, Jul 1997.
[113] G. Bartsch and H. P. Rohr, "Comparative light and electron microscopic study of the human, dog and rat prostate. An approach to an experimental model for human benign prostatic hyperplasia (light and electron microscopic analysis)--a review," Urol Int, vol. 35, pp. 91-104, 1980.

74
[114] I. Leav, et al., "Role of canine basal cells in postnatal prostatic development, induction of hyperplasia, and sex hormone-stimulated growth; and the ductal origin of carcinoma," Prostate, vol. 48, pp. 210-24, Aug 1 2001.
[115] K. Sirinarumitr, et al., "Effects of finasteride on size of the prostate gland and semen quality in dogs with benign prostatic hypertrophy," J Am Vet Med Assoc, vol. 218, pp. 1275-80, Apr 15 2001.
[116] M. Albouy, et al., "Efficacies of osaterone and delmadinone in the treatment of benign prostatic hyperplasia in dogs," Vet Rec, vol. 163, pp. 179-83, Aug 9 2008.
[117] T. Tsutsui, et al., "Effect of osaterone acetate administration on prostatic regression rate, peripheral blood hormone levels and semen quality in dogs with benign prostatic hypertrophy," J Vet Med Sci, vol. 63, pp. 453-6, Apr 2001.
[118] C. Gobello, "Dopamine agonists, anti-progestins, anti-androgens, long-term-release GnRH agonists and anti-estrogens in canine reproduction: a review," Theriogenology, vol. 66, pp. 1560-7, Oct 2006.
[119] C. A. Bradbury, et al., "Relationship between prostatomegaly, prostatic mineralization, and cytologic diagnosis," Vet Radiol Ultrasound, vol. 50, pp. 167-71, Mar-Apr 2009.
[120] A. J. Hendrikx, et al., "Ultrasonic determination of prostatic volume: a cadaver study," Urology, vol. 34, pp. 123-5, Sep 1989.
[121] K. Kamolpatana, et al., "Determination of canine prostatic volume using transabdominal ultrasonography," Vet Radiol Ultrasound, vol. 41, pp. 73-7, Jan-Feb 2000.
[122] H. P. Schlemmer and S. Corvin, "Methods for volume assessment of prostate cancer," Eur Radiol, vol. 14, pp. 597-606, Apr 2004.
[123] E. Giovannucci, "Strengths and limitations of current epidemiologic studies: vitamin D as a modifier of colon and prostate cancer risk," Nutr Rev, vol. 65, pp. S77-9, Aug 2007.
[124] T. Rijsselaere, et al., "Effect of blood admixture on in vitro survival of chilled and frozen-thawed canine spermatozoa," Theriogenology, vol. 61, pp. 1589-602, May 2004.
[125] S. B. Mohr, "A brief history of vitamin d and cancer prevention," Ann Epidemiol, vol. 19, pp. 79-83, Feb 2009.
[126] H. A. Hazewinkel and M. A. Tryfonidou, "Vitamin D3 metabolism in dogs," Mol Cell Endocrinol, vol. 197, pp. 23-33, Nov 29 2002.
[127] K. L. How, et al., "Dietary vitamin D dependence of cat and dog due to inadequate cutaneous synthesis of vitamin D," Gen Comp Endocrinol, vol. 96, pp. 12-8, Oct 1994.
[128] R. J. Mellanby, et al., "Hypercalcaemia in two dogs caused by excessive dietary supplementation of vitamin D," J Small Anim Pract, vol. 46, pp. 334-8, Jul 2005.
[129] C. F. Garland, et al., "Vitamin D for cancer prevention: global perspective," Ann Epidemiol, vol. 19, pp. 468-83, Jul 2009.

75
[130] C. L. Hanchette and G. G. Schwartz, "Geographic patterns of prostate cancer mortality. Evidence for a protective effect of ultraviolet radiation," Cancer, vol. 70, pp. 2861-9, Dec 15 1992.
[131] C. F. Garland and F. C. Garland, "Do sunlight and vitamin D reduce the likelihood of colon cancer?," Int J Epidemiol, vol. 9, pp. 227-31, Sep 1980.
[132] M. R. Haussler, et al., "The nuclear vitamin D receptor: biological and molecular regulatory properties revealed," J Bone Miner Res, vol. 13, pp. 325-49, Mar 1998.
[133] S. Nagpal, et al., "Noncalcemic actions of vitamin D receptor ligands," Endocr Rev, vol. 26, pp. 662-87, Aug 2005.
[134] G. G. Schwartz, et al., "Human prostate cells synthesize 1,25-dihydroxyvitamin D3 from 25-hydroxyvitamin D3," Cancer Epidemiol Biomarkers Prev, vol. 7, pp. 391-5, May 1998.
[135] M. Peterlik, et al., "Calcium, vitamin D and cancer," Anticancer Res, vol. 29, pp. 3687-98, Sep 2009.
[136] G. G. Kwiecinski, et al., "Vitamin D is necessary for reproductive functions of the male rat," J Nutr, vol. 119, pp. 741-4, May 1989.
[137] S. T. Corbett, et al., "Vitamin D receptor found in human sperm," Urology, vol. 68, pp. 1345-9, Dec 2006.
[138] S. Aquila, et al., "Human sperm anatomy: ultrastructural localization of 1alpha,25-dihydroxyvitamin D receptor and its possible role in the human male gamete," J Anat, vol. 213, pp. 555-64, Nov 2008.
[139] M. Blomberg Jensen, et al., "Vitamin D receptor and vitamin D metabolizing enzymes are expressed in the human male reproductive tract," Hum Reprod, vol. 25, pp. 1303-11, May 2010.
[140] J. Merke, et al., "Nuclear testicular 1,25-dihydroxyvitamin D3 receptors in Sertoli cells and seminiferous tubules of adult rodents," Biochem Biophys Res Commun, vol. 127, pp. 303-9, Feb 28 1985.
[141] K. Kinuta, et al., "Vitamin D is an important factor in estrogen biosynthesis of both female and male gonads," Endocrinology, vol. 141, pp. 1317-24, Apr 2000.
[142] M. F. Holick, "The vitamin D deficiency pandemic and consequences for nonskeletal health: mechanisms of action," Mol Aspects Med, vol. 29, pp. 361-8, Dec 2008.
[143] J. A. Johnson, et al., "Immunohistochemical detection and distribution of the 1,25-dihydroxyvitamin D3 receptor in rat reproductive tissues," Histochem Cell Biol, vol. 105, pp. 7-15, Jan 1996.
[144] W. Sun, et al., "Defective female reproductive function in 1,25(OH)2D-deficient mice results from indirect effect mediated by extracellular calcium and/or phosphorus," Am J Physiol Endocrinol Metab, vol. 299, pp. E928-35, Dec 2010.
[145] A. Jemal, et al., "Cancer statistics, 2010," CA Cancer J Clin, vol. 60, pp. 277-300, Sep-Oct 2010.
[146] J. T. Wei, et al., "Urologic diseases in America project: benign prostatic hyperplasia," J Urol, vol. 173, pp. 1256-61, Apr 2005.

76
[147] C. F. Garland, et al., "Serum 25-hydroxyvitamin D and colon cancer: eight-year prospective study," Lancet, vol. 2, pp. 1176-8, Nov 18 1989.
[148] G. G. Schwartz and B. S. Hulka, "Is vitamin D deficiency a risk factor for prostate cancer? (Hypothesis)," Anticancer Res, vol. 10, pp. 1307-11, Sep-Oct 1990.
[149] S. Mukaratirwa and T. Chitura, "Canine subclinical prostatic disease: histological prevalence and validity of digital rectal examination as a screening test," J S Afr Vet Assoc, vol. 78, pp. 66-8, Jun 2007.
[150] M. A. Memon, "Common causes of male dog infertility," Theriogenology, vol. 68, pp. 322-8, Aug 2007.
[151] G. Bartsch, et al., "Light-microscopic stereologic analysis of spontaneous and steroid-induced canine prostatic hyperplasia," J Urol, vol. 137, pp. 552-8, Mar 1987.
[152] T. M. Beer, et al., "High-dose weekly oral calcitriol in patients with a rising PSA after prostatectomy or radiation for prostate carcinoma," Cancer, vol. 97, pp. 1217-24, Mar 1 2003.
[153] C. Crescioli, et al., "Inhibition of spontaneous and androgen-induced prostate growth by a nonhypercalcemic calcitriol analog," Endocrinology, vol. 144, pp. 3046-57, Jul 2003.
[154] C. Crescioli, et al., "Inhibition of prostate cell growth by BXL-628, a calcitriol analogue selected for a phase II clinical trial in patients with benign prostate hyperplasia," Eur J Endocrinol, vol. 150, pp. 591-603, Apr 2004.
[155] M. Maggi, et al., "Pre-clinical evidence and clinical translation of benign prostatic hyperplasia treatment by the vitamin D receptor agonist BXL-628 (Elocalcitol)," J Endocrinol Invest, vol. 29, pp. 665-74, Jul-Aug 2006.
[156] C. Crescioli, et al., "Effect of a vitamin D3 analogue on keratinocyte growth factor-induced cell proliferation in benign prostate hyperplasia," J Clin Endocrinol Metab, vol. 85, pp. 2576-83, Jul 2000.
[157] C. Crescioli, et al., "Des (1-3) IGF-I-stimulated growth of human stromal BPH cells is inhibited by a vitamin D3 analogue," Mol Cell Endocrinol, vol. 198, pp. 69-75, Dec 30 2002.
[158] L. Adorini, et al., "Inhibition of prostate growth and inflammation by the vitamin D receptor agonist BXL-628 (elocalcitol)," J Steroid Biochem Mol Biol, vol. 103, pp. 689-93, Mar 2007.
[159] B. Chughtai, et al., "Inflammation and benign prostatic hyperplasia: clinical implications," Curr Urol Rep, vol. 12, pp. 274-7, Aug 2011.
[160] S. Sood, et al., "Effect of vitamin D deficiency on testicular function in the rat," Ann Nutr Metab, vol. 36, pp. 203-8, 1992.
[161] S. Aquila, et al., "Human male gamete endocrinology: 1alpha, 25-dihydroxyvitamin D3 (1,25(OH)2D3) regulates different aspects of human sperm biology and metabolism," Reprod Biol Endocrinol, vol. 7, p. 140, 2009.
[162] J. A. Halsall, et al., "In silico analysis of the 5' region of the Vitamin D receptor gene: functional implications of evolutionary conservation," J Steroid Biochem Mol Biol, vol. 103, pp. 352-6, Mar 2007.

77
[163] F. Jehan, et al., "Exons and functional regions of the human vitamin D receptor gene around and within the main 1a promoter are well conserved among mammals," J Steroid Biochem Mol Biol, vol. 103, pp. 361-7, Mar 2007.
[164] K. Verstegen-Onclin and J. Verstegen, "Endocrinology of pregnancy in the dog: a review," Theriogenology, vol. 70, pp. 291-9, Aug 2008.
[165] C. Urhausen, et al., "Concentrations of prolactin, LH, testosterone, TSH and thyroxine in normospermic dogs of different breeds," Reprod Domest Anim, vol. 44 Suppl 2, pp. 279-82, Jul 2009.
[166] A. R. Gunzel-Apel, et al., "Effects of gonadectomy on prolactin and LH secretion and the pituitary-thyroid axis in male dogs," Theriogenology, vol. 71, pp. 746-53, Mar 15 2009.
[167] Y. Corrada, et al., "Prolactin reference range and pulsatility in male dogs," Theriogenology, vol. 66, pp. 1599-602, Oct 2006.
[168] Y. Corrada, et al., "Secretory patterns of prolactin in dogs: circannual and ultradian rhythms," Reprod Domest Anim, vol. 38, pp. 219-23, Jun 2003.
[169] A. Ciccarelli, et al., "PRL secreting adenomas in male patients," Pituitary, vol. 8, pp. 39-42, 2005.
[170] M. Egli, et al., "Rhythmic secretion of prolactin in rats: action of oxytocin coordinated by vasoactive intestinal polypeptide of suprachiasmatic nucleus origin," Endocrinology, vol. 145, pp. 3386-94, Jul 2004.
[171] R. L. Jones, et al., "Localization and temporal expression of prolactin receptor in human endometrium," J Clin Endocrinol Metab, vol. 83, pp. 258-62, Jan 1998.
[172] A. C. Okkens, et al., "Evidence for the non-involvement of the uterus in the lifespan of the corpus luteum in the cyclic dog," Vet Q, vol. 7, pp. 169-73, Jul 1985.
[173] N. D. Horseman, et al., "Defective mammopoiesis, but normal hematopoiesis, in mice with a targeted disruption of the prolactin gene," EMBO J, vol. 16, pp. 6926-35, Dec 1 1997.
[174] C. J. Ormandy, et al., "Null mutation of the prolactin receptor gene produces multiple reproductive defects in the mouse," Genes Dev, vol. 11, pp. 167-78, Jan 15 1997.
[175] M. T. Nevalainen, et al., "Androgen-dependent expression of prolactin in rat prostate epithelium in vivo and in organ culture," FASEB J, vol. 11, pp. 1297-307, Dec 1997.
[176] M. T. Nevalainen, et al., "Prolactin and prolactin receptors are expressed and functioning in human prostate," J Clin Invest, vol. 99, pp. 618-27, Feb 15 1997.
[177] M. F. El Etreby, et al., "Role of the pituitary gland in experimental hormonal induction and prevention of benign prostatic hyperplasia in the dog," Cell Tissue Res, vol. 204, pp. 367-78, 1979.
[178] T. J. Ahonen, et al., "Prolactin is a survival factor for androgen-deprived rat dorsal and lateral prostate epithelium in organ culture," Endocrinology, vol. 140, pp. 5412-21, Nov 1999.

78
[179] H. Wennbo, et al., "Transgenic mice overexpressing the prolactin gene develop dramatic enlargement of the prostate gland," Endocrinology, vol. 138, pp. 4410-5, Oct 1997.
[180] E. Reiter, et al., "Effects of pituitary hormones on the prostate," Prostate, vol. 38, pp. 159-65, Feb 1 1999.
[181] H. Yamanaka, et al., "Effect of estrogen administration on activities of testosterone 5alpha-reductase, alkaline phosphatase and arginase in the ventral and the dorsolateral prostates of rats," Endocrinol Jpn, vol. 22, pp. 297-302, Aug 1975.
[182] S. Chevalier, et al., "Nonsteroidal serum factors involved in the regulation of the proliferation of canine prostatic epithelial cells in culture," Prostate, vol. 5, pp. 503-12, 1984.
[183] D. Helmerich and J. E. Altwein, "Effect of prolactin and the anit-prolactin bromocriptin on the testosterone uptake and metabolism in androgen-sensitive and insensitive canine organs," Urol Res, vol. 4, pp. 101-5, Nov 10 1976.
[184] F. G. Robertson, et al., "Prostate development and carcinogenesis in prolactin receptor knockout mice," Endocrinology, vol. 144, pp. 3196-205, Jul 2003.
[185] A. Colao, et al., "Outcome of cabergoline treatment in men with prolactinoma: effects of a 24-month treatment on prolactin levels, tumor mass, recovery of pituitary function, and semen analysis," J Clin Endocrinol Metab, vol. 89, pp. 1704-11, Apr 2004.
[186] M. K. Gill-Sharma, "Prolactin and male fertility: the long and short feedback regulation," Int J Endocrinol, vol. 2009, p. 687259, 2009.
[187] M. Egli, et al., "Prolactin secretion patterns: basic mechanisms and clinical implications for reproduction," Reproduction, vol. 140, pp. 643-54, Nov 2010.
[188] G. Corona, et al., "Effect of hyperprolactinemia in male patients consulting for sexual dysfunction," J Sex Med, vol. 4, pp. 1485-93, Sep 2007.
[189] A. Colao and G. Lombardi, "Growth-hormone and prolactin excess," Lancet, vol. 352, pp. 1455-61, Oct 31 1998.
[190] M. De Rosa, et al., "The treatment with cabergoline for 24 month normalizes the quality of seminal fluid in hyperprolactinaemic males," Clin Endocrinol (Oxf), vol. 64, pp. 307-13, Mar 2006.
[191] G. Merino, et al., "Hyperprolactinemia in men with asthenozoospermia, oligozoospermia, or azoospermia," Arch Androl, vol. 38, pp. 201-6, May-Jun 1997.
[192] M. De Rosa, et al., "Hyperprolactinemia in men: clinical and biochemical features and response to treatment," Endocrine, vol. 20, pp. 75-82, Feb-Mar 2003.
[193] A. Shafik, "Prolactin injection, a new contraceptive method: experimental study," Contraception, vol. 50, pp. 191-9, Aug 1994.
[194] R. W. Steger, et al., "Neuroendocrine and reproductive functions in male mice with targeted disruption of the prolactin gene," Endocrinology, vol. 139, pp. 3691-5, Sep 1998.

79
[195] K. Purvis, et al., "Prolactin and Leydig cell responsiveness to LH/hCG in the rat," Arch Androl, vol. 3, pp. 219-30, Nov 1979.
[196] W. J. Huang, et al., "Role of testicular interstitial macrophages in regulating testosterone release in hyperprolactinemia," J Cell Biochem, vol. 88, pp. 766-73, Mar 1 2003.
[197] V. L. Williams, et al., "Common and specific effects of the two major forms of prolactin in the rat testis," Am J Physiol Endocrinol Metab, vol. 293, pp. E1795-803, Dec 2007.
[198] G. Verhoeven and J. Cailleau, "Follicle-stimulating hormone and androgens increase the concentration of the androgen receptor in Sertoli cells," Endocrinology, vol. 122, pp. 1541-50, Apr 1988.
[199] J. P. Advis, et al., "Serotonergic control of prolactin release in male rats," Life Sci, vol. 24, pp. 359-65, Jan 22 1979.
[200] H. C. Christian, et al., "Thyrotrophin-releasing hormone, vasoactive intestinal peptide, prolactin-releasing peptide and dopamine regulation of prolactin secretion by different lactotroph morphological subtypes in the rat," J Neuroendocrinol, vol. 19, pp. 605-13, Aug 2007.
[201] C. M. Leite, et al., "Actions of angiotensin II and dopamine in the medial preoptic area on prolactin secretion," Physiol Res, vol. 57, pp. 109-18, 2008.
[202] Y. J. Jeng, et al., "Combinations of physiologic estrogens with xenoestrogens alter calcium and kinase responses, prolactin release, and membrane estrogen receptor trafficking in rat pituitary cells," Environ Health, vol. 9, p. 61, 2010.
[203] L. Ferland, et al., "Evidence for a role of endorphins in stress- and suckling-induced prolactin release in the rat," Mol Cell Endocrinol, vol. 12, pp. 267-72, Dec 1978.
[204] B. H. Duvilanski, et al., "Interaction between substance P and TRH in the control of prolactin release," J Endocrinol, vol. 166, pp. 373-80, Aug 2000.
[205] M. Dahlhoff, et al., "In Vivo Evidence for Epidermal Growth Factor Receptor (EGFR)-mediated Release of Prolactin from the Pituitary Gland," J Biol Chem, vol. 286, pp. 39297-306, Nov 11 2011.
[206] H. L. Henderson, et al., "Gonadotropin-releasing hormone stimulates prolactin release from lactotrophs in photoperiodic species through a gonadotropin-independent mechanism," Biol Reprod, vol. 78, pp. 370-7, Feb 2008.
[207] C. Spuch, et al., "Fibroblast growth factor-2 and epidermal growth factor modulate prolactin responses to TRH and dopamine in primary cultures," Endocrine, vol. 29, pp. 317-24, Apr 2006.
[208] Y. Zhou, et al., "[Effect of cholecystokinin on prolactin release and its action mechanism in the rats]," Zhongguo Yi Xue Ke Xue Yuan Xue Bao, vol. 17, pp. 412-7, Dec 1995.
[209] H. L. Henderson, et al., "Direct effects of prolactin and dopamine on the gonadotroph response to GnRH," J Endocrinol, vol. 197, pp. 343-50, May 2008.

80
[210] T. J. Kreeger and U. S. Seal, "Circannual prolactin rhythm in intact dogs housed outdoors," Chronobiologia, vol. 19, pp. 1-8, Jan-Mar 1992.
[211] M. B. Koivisto, et al., "Effects of short-term hyper- and hypoprolactinaemia on hormones of the pituitary, gonad and -thyroid axis and on semen quality in male Beagles," Reprod Domest Anim, vol. 44 Suppl 2, pp. 320-5, Jul 2009.

81
CHAPTER TWO
VITAMIN D, BENIGN PROSTATIC HYPERPLASIA, PROSTATE VOLUME AND
SEMEN PARAMETERS IN THE DOG.
Prepared for publication in Theriogenology
Abstract
The role of Vitamin D in normal skeletal metabolism is well-known;
however, other functions of this vitamin in multiple physiological processes,
including reproduction, are still being discovered. The aims of this observational
study were to determine whether Vitamin D is associated with benign prostatic
hyperplasia (BPH), prostate volume and/or semen quality characteristics in the
dog.
Using a convenience sample of healthy dogs, associations between serum
25-hydroxy Vitamin D (25OHVD) and a) BPH, b) prostate volume (n=28) or c)
sperm motility and morphology (n=22) were examined. Using the Student t-test
no difference in 25OHVD between BPH positive dogs and normal dogs was
observed (p=0.59). Bivariable analyses controlling for BPH, revealed no
significant associations between Vitamin D and prostate volume (p=0.51).
Significant associations between Vitamin D and semen quality including motility
(R2=0.55, p=0.0008), progressive motility (R2=0.51, p=0.002), beat cross
frequency (BCF; R2=0.28, p=0.01), distance average path (DAP; R2= 0.38,
p=0.003), curvilinear distance (DCL; R2=0.25, p=0.02), linear distance (DSL;
R2=0.31, p=0.01), average path velocity (VAP; R2=0.35, p=0.005), curvilinear
velocity (VCL; R2=0.25, p=0.02), straight line velocity (VSL; R2=0.27, p=0.02),

82
morphologically normal sperm (R2=0.43,p=0.01) and detached heads (R2=0.31,
p=0.01) were found using univariable analysis. 25OHVD, with an age interaction
term, was significantly associated with the following sperm motility parameters in
bivariable analyses: motility (R2 = 0.57, p=0.0019), progressive motility (R2=0.57,
p=0.002), BCF (R2=0.47, p=0.003), DCL (R2=0.51, p=0.006), VCL (R2=0.51,
p=0.006), amplitude of lateral head displacement (ALH; R2=0.42, p=0.02) and
average orientation change (AOC; R2=0.38, p=0.04), and normal sperm
morphology (R2=0.49, p=0.01). Similarly, 25OHVD and prostatic volume
interaction were associated with motility (R2=0.72, p<0.0001), progressive motility
(R2=0.60, p=0.001), DCL (R2=0.53, p=0.01), VCL (R2=0.52, p=0.01), normal
morphology (R2=0.52, p=0.01), and head defects (R2=0.39, p=0.04).
25OHVD was associated with several sperm motility and morphology
parameters which may suggest a role of 25OHVD on spermatogenesis and
sperm function. Further study of the possible role(s) of Vitamin D in
spermatogenesis, sperm function and sperm physiology is warranted.
Keywords; Vitamin D, semen, prostate, dog, benign prostatic hyperplasia
1. Introduction
Vitamin D has an important role in calcium and phosphorous metabolism
and is integral to normal bone physiology. In severe deficiencies of Vitamin D in
the very young, incomplete mineralization of developing or maturing bone leads
to malformation of the skeleton manifested as rickets. In adults the same
deficiency results in brittle and fragile bones leading to fractures. Research is

83
beginning to suggest a larger role for this vitamin in many different physiological
systems, including reproduction [1-3].
Vitamin D is essential in inducing or inhibiting transcription of numerous
genes that influence cell proliferation, invasiveness, angiogenesis, metastatic
potential, differentiation and apoptosis [4]. The use of experimental Vitamin D
analogues specific to the Vitamin D receptor (VDR) in human prostatic tissue has
resulted in a significant decrease in prostatic volume in men with benign prostatic
hyperplasia [5]. Vitamin D and its receptor have also been identified in human
and rat prostatic tissue, testes and sperm [6-9]. In experimentally induced
Vitamin D deficiency in rats, a decrease in Sertoli cell numbers, degenerative
changes in the seminiferous tubules and decreased sperm motility have been
observed, while Vitamin D deficiency in men has been associated with increased
mortality from prostate and other cancers[10, 11].
In dogs much is known about normal prostate function and physiology as
well as spermatogenesis and sperm function. The effect of fat-soluble vitamins
on reproduction in the stud dogs however, is largely unstudied. The purpose of
this study was to examine whether associations exist between 25-hydroxy
Vitamin D (25OHVD) and BPH, 25OHVD and prostate volume, and 25OHVD and
several sperm motility and morphology characteristics.

84
2. Materials and Methods
2.1 Case Subjects
A convenience sample of 28 healthy intact male dogs of various breeds
(total 23) and fertility histories, with or without indication of prostate disease, were
obtained between the months of March and December of 2009. Dogs were
recruited from cases presented to the Theriogenology Service at the Ontario
Veterinary College and from a private clinic in Southwestern Ontario with a large
breeder clientele. All clients were required to fill out a consent form (Appendix 1)
and brief questionnaire (Appendix 2) outlining their dogs’ fertility history, diet and
amount of time their dogs spent outdoors.
All dogs underwent a complete male breeding soundness examination;
including semen collection and analysis and ultrasonographic examination of the
prostate gland. After completion of the examination, 6 ml of whole blood was
collected via cephalic venipuncture; 2mls was immediately placed into a lavender
top EDTA vial (Vacutainer™ Becton-Dickinson) for complete blood count (CBC)
analysis (Animal Health Laboratory, Ontario Veterinary College). The remainder
was placed in a red top vial (Vacutainer™ 366430 Becton-Dickinson) and allowed
to clot for 20 minutes before centrifugation and collection of serum. Serum was
then frozen and stored at -80°C until analysis. Finally, urine was collected mid-
stream during micturition into a clean vessel for urinalysis. All dogs were
examined and evaluated by the same operator.

85
2.2 Semen collection, dilution and staining.
Twenty-two of the 28 dogs (78.6%) enrolled for the study had semen
successfully collected once with manual stimulation using a latex artificial vagina
attached to a prewarmed sterile glass tube. Semen was divided during ejaculation
into sperm-rich (2nd) and prostatic (3rd) fractions. Semen concentration was
determined using the Unopette® - Neubauer system (Becton-Dickinson,
Rutherford, NJ) as previously described [12]. Sperm-rich semen was diluted 1:1
up to 3:1 with Canipro™ Chill 5 semen extender (Minitube Canada, ON) and
chilled according to product specifications (Appendix 3) and either transported or
stored between 5-8ºC for 1-3 hours prior to computer assisted semen
assessment (CASA) evaluation. Morphological analysis was undertaken using
eosin-nigrosin staining techniques[12] at 1000X magnification under oil (Nikon
Eclipse 50i, Japan) using the classification system of the Society for
Theriogenology (SFT) (Appendix 4). Semen collection and specimen preparation
were performed by the same operator. A small amount of fresh undiluted
unextended sperm-rich semen was submitted to the Animal Health Laboratory at
the Ontario Veterinary College for aerobic bacterial culture and sensitivity testing.
2.3 Motility Analysis
Chilled extended semen samples were slowly warmed to room
temperature over a 10-minute period, and placed on a prewarmed stage of the
microscope (Olympus BX41 (U-Spt, Japan) prior to evaluation. Samples were
given one minute to equilibrate in the slide chamber and then evaluated using the

86
SpermVision™ CASA system (Minitube, America) with the technical settings for
canine semen as described by Schäfer-Somi and Aurich[13]. The present study
used 50-300 cells per field and used a Leja® Standard Count chamber (Leja,
Netherlands) of 2 µl capacity. Table 2.1 details the technical settings used in the
CASA.
The following parameters were assessed using SpermVision™ analysis
(for a complete description of terms see Schäfer-Somi and Aurich, 2007): (1) total
motility expressed as the percentage of spermatozoa with curvilinear velocity
(VCL) >15 µm/s [14], (2) progressive motility (%), (3) amplitude of lateral head
displacement (ALH, µm), (4) average orientation change (AOC, degrees) where
sperm <7 are considered immotile [13], (5) frequency of head displacement or
beat cross frequency (BCF, Hz), (6) mean distance travelled (DAP, µm), (7)
average velocity (VAP, µm/s), (8) curved line distance (DCL, µm), (9) curvilinear
velocity (VCL, µm/s), (10) straight line distance (DSL, µm), (11) linear velocity
(VSL, µm/s), (12) mean straightness coefficient (STR = VSL/VAP X100, %), (13)
linear coefficient (LIN = VSL/VCL X 100, %), and (14) wobble coefficient (WOB =
VAP/VCL X 100, %), and progressive fast spermatozoa (PF), which is defined by
minimal STR for progressive fast sperm = 90% [15]. CASA analysis was
performed by the same individual.
2.4 Prostate Examination
The prostate gland of all dogs was evaluated by the same individual after
semen collection by several methods, including transabdominal ultrasonography,

87
digital palpation per rectum and cytological examination. Culture and sensitivity
of prostatic fluid was performed by the Animal Health Laboratory, Ontario
Veterinary College, Guelph, Ontario.
2.5 Ultrasonography of the Prostate
Dogs were either placed in dorsal recumbency in a V-shaped trough to
stabilize them for examination, or were examined in standing position depending
on patient compliance and size. Using B-mode ultrasonography, measurements
of length, width and height of the prostate gland were taken similarly as described
previously [16, 17] using a MyLab® 5 Portable Ultrasound (Universal Ultrasound,
NY) with a 5-8 MHz curved linear array transducer. The prostate of each dog was
measured three times in transverse and longitudinal sections and the average of
each measurement was used in the analyses of prostate size. Prostate size was
calculated using the volume formula described by Kamolpatana et al (2000) (Vm
= [1/2.6 (length x width x depth)] +1.8] cm3)[17]. Symmetry of the gland, as well
as any lesions present were noted and measured. A BPH case was defined as a
patient with a symmetrical prostate and at least one of the parameters of height,
length and/or width being larger than the set maximum values based on the
clinical experience of the evaluator (see Table 2.2) and the presence of cysts on
ultrasonographic examination; one of the clinical signs of hemospermia, rectal
tenesmus, and dysuria; and/or blood in the prostatic fraction on cytological
examination [18].

88
2.6 Vitamin D Analysis
Vitamin D analysis was performed by the Diagnostic Center for Population
and Animal Health, Michigan State University using a quantitative
radioimmunoassay (RIA) for 25-hydroxyvitamin D (DiaSorin, Stillwater MN)
validated for canines with normal ranges between 60 and 215 nmol/l. All samples
were analysed in a single batch.
The 25OHVD assay consisted of a two-step procedure. Rapid extraction
of 25OHVD and other hydroxylated metabolites from serum of plasma was
accomplished with acetonitrile. Following extraction, the treated samples were
then assayed using an equilibrium RIA procedure. The serum sample, antibody to
25OH-VitD and 125I-labelled 25OHVD tracer were incubated for 90 minutes at 20-
25°C. Phase separation was accomplished after a 20 minute incubation at 20-
25°C with a second antibody precipitating complex. Buffer was added after
incubation and prior to centrifugation to aid in reducing nonspecific binding.
Sensitivity of the assay was 3.7 nmol/l with an inter- and intra-assay coefficient of
variation of 11% and 10%, respectively, with cross-reactivities as follows: 100%
for 25-OH-D2, 25-OH-D3, 24,25-(OH)2-D2, 24,25-(OH)2-D3, 25,26-(OH)2-D2,
and 25,26-(OH)2-D3, 11% cross-reactivity to 1,25-(OH)2-D2, and 1,25-(OH)2-D3,
and 0.8% cross-reactivity to ergo- and cholecalciferol.
2.7 Urinalysis
Urinalysis consisted of a urine test strip (Chemstrip 9 , Roche
Diagnostics, Quebec), microscopic examination of urine sediment, determination

89
of urine specific gravity and bacterial culture and sensitivity (Animal Health
Laboratory, Guelph, ON) in order to rule out cystitis.
2.8 Statistical Analyses
2.8.1 Overall Statistical Analyses
SAS version 9.2 software (SAS Institute Inc. Cary, NC, USA) was used for
all statistical analyses. The GLM procedure was used to analyze the univariable
and multivariable associations between the explanatory variables and outcomes
(see next sections for details). Due to the small sample size and the limited
degrees of freedom available, only two explanatory variables were used in any
model to discourage over-fitting of the model. Models were generated using
backwards elimination in the case of multivariable analyses. The significance
level was set to p<0.05 for all analyses, and 95% confidence intervals were
constructed.
The UNIVARIATE procedure was performed to assess residuals and
ANOVA assumptions in order to determine the fit of the model and whether
transformation of the data was needed to achieve normality. Four tests were
used to test normality: Shapiro-Wilk, Cramér-von Mises, Kolmogorov-Smirnov,
Anderson-Darling tests [19]. Residuals were plotted against the predicted values
and explanatory variables used in the model. Such plots and tests for normality
may reveal outliers, unequal variances or other problems with the assumptions
and may suggest the need for a data transform [19, 20]. Data transformation
included a logit transform (logit of outcome = log((r+k)/(n+k)); r=responding cells,

90
n=number of cell counted, k=small bias correction term = 0.25) in cases of
variables with a percentage value.
Using the Grubbs’ test for outliers [21], any result where the residual value
was greater than 2.58 times the standard deviation of the residuals (one-tailed
test p<0.05) were labeled as outliers and the models were rerun to determine how
both normality of the data and the model changed with removal of these points
(Appendix 5).
2.8.2 Study Population
Descriptive statistics including means, standard deviations (SD), and
standard errors (SE) were used to analyze the characteristics of the study
population. Ages of dogs were simplified using half-year increments as not all
dogs had exact birth dates. Time of year was divided into Summer (March 20-
September 21) and Winter (September 22-March 19) according to the vernal and
autumnal equinoxes in the northern hemisphere, to account for the influence of
natural ultraviolet radiation on vitamin D production via the skin. The Student’s t-
test was used to test whether there was a difference in mean serum 25OHVD
concentrations between dogs of the two seasonal groups. Amount of time spent
outdoors was grouped according to whether the dogs spent greater than or less
than five hours per day during the season of sampling and differences between
groups was analyzed using a one-way ANOVA. Diets were categorized
according to whether a commercial, raw or a combined diet was given to account

91
for other vitamin D sources. Differences in mean serum 25OHVD concentrations
amongst the different diets were analyzed using one-way ANOVA.
2.8.3 BPH and Prostatic Volume
The Student’s t-test was used to test whether there was a difference in
mean serum 25OHVD concentrations between dogs with and without BPH.
Univariable analysis described in section 2.8.1 included the explanatory variable
serum 25OHVD and prostate volume as the outcome variable. Analysis of
prostate volume was analytically controlled for BPH status by including it in the
model. Age and bodyweight were also tested separately as explanatory variables
with serum 25OHVD as the outcome variable. In keeping with previous studies of
prostate volume [16, 17] bivariable regression model of prostate volume with age
and bodyweight as explanatory variables was tested.
2.8.4 Sperm Motility
Univariable analysis of sperm motility described in section 2.8.1 included
total motility, progressive motility, ALH, AOC, BCF, DAP, VAP, DCL, VCL, DSL,
VSL, STR, LIN, and WOB, as outcome variables with serum 25OHVD as the
explanatory variable. All sperm motility parameters were included in bivariable
analyses. Three bivariable regression models were created for each motility
parameter; the first model included age and serum 25OHVD, the second included
bodyweight and serum 25OHVD, and the third included prostate volume and
serum 25OHVD as explanatory variables.

92
2.8.5 Sperm Morphology
Univariable analysis of semen morphology separately modeled normal
sperm, head defects, midpiece defects, tail defects, loose heads, proximal
droplets and distal droplets as outcome variables with serum 25OHVD as the
explanatory variable. All sperm morphology parameters were included in
bivariable analyses. Three bivariable regression models were created for each
sperm motility parameter as the first model included age and serum 25OHVD, the
second included bodyweight and serum 25OHVD, and the third included prostate
volume and serum 25OHVD as explanatory variables.
3. Results
3.1 Vitamin D and the Study Population
Serum vitamin D concentrations of subjects ranged from 70 to 203 nmol/l
with a mean and SD of 140.5 37.1 nmol/l. BPH was found in 82.1% (23/28) of
dogs studied. Ages of dogs ranged from 2 -11 years with a mean and SD of 6 2
years. Bodyweight of dogs ranged from 9.2-78.2Kg with a mean and SD of 35.4
15.8Kg. Tables 2.3 to 2.5 describe the other characteristics of the study
population of dogs. No significant associations between mean serum 25OHVD
concentrations and types of diet (p=0.79), time of year of sampling (p=0.54) and
number of hours spent outdoors in the summer and in the winter (p=0.85) were
observed (Tables 2.6 - 2.8). CBC and urinalyses of all dogs enrolled in the study
were within normal limits. Cultures and sensitivities of urine, prostatic fluid and

93
semen in dogs that gave semen samples, were all negative. All dogs were
healthy having neither illness nor receiving any medications. Fertility histories of
the study population consisted of dogs that had never been bred or had not been
bred within the year prior to evaluation. Body condition of the majority of dogs
was considered ideal except for two dogs (Golden Retriever, Mastiff Cross) that
were categorized as slightly overweight.
3.2 BPH and Prostate Volume
There was no significant association between mean serum 25OHVD
concentration and BPH status (p=0.59) (Table 2.9). Univariable analysis revealed
no significant association between 25OHVD concentration and prostate volume
(R2=0.02, p=0.51). Controlling analytically for BPH in the previous model did not
change the results obtained. There was no significant association with serum
25OHVD as a response to age (R2=0.02, p=0.52) or bodyweight (R2=0.0004,
p=0.92). There was significant association in bivariable analysis of prostate
volume with age and bodyweight (log2Volume=0.13*Age + 0.02*Bodyweight,
R2=0.20, F-value=3.33, p=0.05) without interaction. Overall there was an
increase in prostatic volume with increasing age and bodyweight. Using three
research hounds examined twice one day apart, intra- and inter-assay coefficients
of variation on prostatic measurements and their 95% confidence intervals were
3.6% (2%, 4.6%) and 21.8% (17.3%, 29.7%) for prostatic length, 4.8% (4%, 6%)
and 19.7% (16.5%, 26.4%) for prostatic width, and 4.3% (3.5%, 5.4%) and 21.1%
(16.7%, 28.7%), respectively.

94
3.3 Sperm Motility
Univariable analyses of both total motility (R2=0.55, p=0.0008) and
progressive motility (R2=0.51, p=0.002) had significant association with serum
25OHVD alone (Figures 2.1 and 2.2). For both motility outcomes, the pattern of
association was an increase in the motility parameter at 25OHVD concentrations
between 70 and 100 nmol/l, little or no effect on motility at 25OHVD
concentrations between 100 and 180 nmol/l and a slight decrease in motility at
25OHVD concentrations above 180 nmol/l (all 25OHVD concentrations stated
here are within the normal range for healthy dogs).
Significant univariable associations between 25OHVD and BCF, DAP,
VAP, DCL, VCL, DSL and VSL were also found (Table 2.10); specifically each
had a simple linear positive association with increasing serum 25OHVD. No
significant associations were found among semen motility parameters ALH, AOC,
LIN, STR, and WOB and the explanatory variable serum 25OHVD (Table 2.10).
Bivariable regression analysis revealed significant interaction of age
(R2=0.71, p=0.0001 and R2=0.57, p=0.002) and prostatic volume (R2=0.73,
p=0.0001, and R2=0.60, p=0.0012), but not bodyweight, with serum 25OHVD on
both sperm motility and progressive motility, respectively. Figures 2.3 – 2.6
depict these relationships. At younger ages and smaller prostatic volumes, there
is a noticeable effect of increasing concentrations of serum 25OHVD on
increasing motility and progressive motility at 25OHVD concentrations between
70 and 140 nmol/l. At older ages (i.e.10 years) and larger transformed prostate
volumes (i.e. 5.5) an opposite effect of serum 25OHVD on motility is observed

95
with a decrease in total motility parameters with increasing serum 25OHVD.
Progressive motility follows a similar pattern to total motility with increasing
25OHVD and prostate volume however, there is an increase in progressive
motility with increasing 25OHVD at all ages with less dramatic results between 8
and 10 years of age.
Using bivariable analysis, BCF had a significant association with serum
25OHVD and age of dog as explanatory variables (R2=0.47, p=0.003) but not with
serum 25OHVD and bodyweight nor with serum 25OHVD and prostate volume.
BCF increased slightly with age and increasing serum 25OHVD (Figure 2.7). For
both response variables DCL and VCL, significant interactions were found
between the explanatory variables serum 25OHVD and age, and serum 25OHVD
and prostatic volume but not with serum 25OHVD and bodyweight. Specifically,
at younger ages DCL increased with increasing serum 25OHVD but by 8 years of
age there was little effect of increasing serum 25OHVD on DCL. At 10 years of
age, increasing serum 25OHVD was associated with decreasing DCL (Figure
2.8). Smaller prostate volumes were associated with an increase in DCL with
increasing serum 25OHVD, however, at a larger transformed prostate volume of
5.5 the reverse is true with a decrease in DCL as a response to increasing serum
25OHVD (Figure 2.9). VCL responded to increasing serum 25OHVD in the same
manner as DCL with respect to both age and prostate volume (Figures 2.10-2-
11). A significant association between DSL and prostate volume was found
(R2=0.46, F-value=4.81, p=0.001) (Figure 2.12). Although VSL did not have a

96
significant association with prostate volume it did approach significance (R2=0.28,
F-value=3.42, p=0.06) (Table 2.10).
There was a significant association of each of ALH and AOC with serum
25OHVD and age as explanatory variables but not with serum 25OHVD and
bodyweight nor with serum 25OHVD and prostate volume (Figures 2.13 and
2.14). There was an increase in both ALH and AOC with increasing age and
increasing serum 25OHVD at younger ages and a negative association between
these motility parameters and increasing serum 25OHVD at older ages. There
were no other significant bivariable associations between the remaining motility
parameters DAP, VAP, VSL, STR, LIN, WOB with serum 25OHVD and age,
serum 25OHVD and bodyweight and serum 25OHVD and prostate volume (Table
2.11).
3.4 Outlier Removals
Removal of outliers, as previously defined, within this set of data
influenced two of the significant associations observed (Appendix 5). Sperm total
motility and Vitamin D associations at various prostate volumes changed
significantly with respect to their slope coefficients with removal of one outlier
(Dog 6, a 5-year old Golden Retriever that had never sired a litter even after
multiple breedings). The overall pattern of association was similar to those found
with the outlier maintained although the magnitude of those associations was
decreased. Secondly, removal of the same outlier with respect to the associations
between progressive motility and 25OHVD alone became insignificant (p=0.21).

97
Although, the removal of outliers in large data sets may be advised we maintained
the data due to the small sample size and possibility of removing valuable
information from the analysis.
3.5 Sperm Morphology
The relationship between 25OHVD and morphologically normal sperm
was quadratic, predicting an increase in percent normal morphology with
increasing 25OHVD concentrations at low to mid-normal range of normal serum
25OHVD (see Figure 2.15). However, at high normal values of serum 25OHVD
this effect was lost and a negative effect on normal morphology was observed.
A significant decrease in the percentage of loose heads can be seen with
increasing serum 25OHVD (see Figure 2.16). No other significant associations
with 25OHVD using univariable analyses were found with the remaining
morphology characteristics including midpiece (R2=0.03, F-value=0.52, p=0.48)
and tail (R2=0.002, F-value=0.05, p=0.82) defects, proximal (R2=0.006, F-
value=0.11, p=0.74) and distal (R2=0.02, F-value=0.34, p=0.56) droplets. There
was a tendency towards significance with head defects and 25OHVD (R2=0.17, F-
value=3.61, p=0.07).
Bivariable analyses explained greater variation than univariable analysis
between normal sperm and 25OHVD. Both age and serum 25OHVD and
prostatic volume and serum 25OHVD used in modeling the response of normal
sperm were significant (Table 2.12 and Figures 2.17-2.18). With increasing age
and prostatic volume, response to 25OHVD was greatest at younger ages and

98
smaller prostate volumes, while at the age of 10 years and transformed prostate
volume of 5.5 there was a decrease in percentage of normal sperm with
increasing serum 25OHVD. Prostate volume and 25OHVD interacted and were
significantly associated with percentage of head defects (Table 2.12 and Figure
2.19). The percentage of head defects decreased with increasing 25OHVD at
small prostate volumes, but increased at transformed prostate volume 5.5. The
bivariable regression models for the remaining sperm morphology characteristics
were non-significant (Table 2.12).
4. Discussion and Conclusions
Many of the motility parameters studied have strong correlations to fertility
in many species [22-24] and the associations of Vitamin D with these parameters
have importance in the reproductive health and potential of the stud dog. Among
these parameters total motility, progressive motility, ALH, VAP, VCL, VSL, BCF,
LIN have shown high correlation with AI fertility in multivariable analysis in bulls
[22] while ALH, VCL, and VSL in in vitro human studies have shown correlation
with high fertilization rates [23]. In the dog, it has been found that most of motility
parameters measured by CASA had correlation to fertility except for BCF, LIN,
and STR [24]. Therefore fertility potential, as measured by CASA, and
associations with 25OHVD in the dog can provide new insight into reproductive
health in the stud dog.
Serum 25OHVD had significant positive associations with several motility
parameters in this study in both univariable and bivariable analysis. Total and
progressive motilities in univariable analysis showed almost identical patterns of

99
association with 25OHVD and are in agreement with human studies [25].
25OHVD had positive associations with total and progressive motilities in the low
to mid-range of normal (70-180 nmol/l), and a negative effect at high normal
(>180 nmol/l) concentrations, with the most desirable outcomes in the mid-range
of normal 25OHVD (120-180 nmol/l). Blomberg-Jensen et al (2011) found
similar associations, using a 25OHVD squared term with human sperm, although
their associations were smaller than in our study and a negative effect at higher
concentrations of 25OHVD was not seen [25]. Due to possible interspecies
differences in 25OHVD concentrations, it is difficult to determine whether the
higher 25OHVD concentrations observed in the human study are comparable to
the concentrations observed in our canine population and could a similar effect
have been seen at even higher 25OHVD concentrations (nearer the upper limit)
in men. In addition, the human study analyzed total motility only and found that
in vitro addition of the active Vitamin D tissue form, 1,25-dihydroxyvitamin D
(1,25diOHVD), to mature sperm caused a dose-dependent increase in total
motility, but after a certain concentration (10-7 M) a negative effect was seen.
This finding is supported by studies of sperm survival where a similar biphasic
dose-response curve to 1,25diOHVD was found [6]. This could in part be due to
exceeding Ca2+ stores and exhausting the intracellular Ca2+ influx and ligand
binding and activation of the protein kinase C system responsible for sperm
motility [26]. Blomberg-Jensen et al (2011) also found that addition of
1,25diOHVD caused increased intracellular Ca2+ concentrations in ejaculated
spermatozoa compared with controls. They confirmed that Vitamin D action was

100
through a non-genomic VDR [25]. The results from these studies suggest that
the Vitamin D associated increases in sperm motility and progressive motility we
observed may work independently of VDR mediated gene transcription.
Total and progressive motilities in bivariable analysis with both 25OHVD
and age, and 25OHVD and prostatic volume showed an increase in both motility
parameters at younger ages (4-8 years of age) and smaller transformed prostate
volumes (log2volume = 3.5 - 4.5) and a decrease at older ages (10 years of age)
and larger transformed prostate volumes (log2volume=5.5). Age effects on
motility parameters have not been previously found in the dog [24] although they
have been found in men [27]. Multivariable effects were not investigated in these
studies and the effect of age and 25OHVD with age-25OHVD interaction
observed in our study suggest that age may directly influence Vitamin D
physiology. In a study that investigated age-related changes in Vitamin D
physiology in rat duodenum, an age effect was observed [28]. Measuring both
protein kinase C (PKC) and Ca2+ uptake by cultured duodenal cells from young
(three month-old) and aged (22-24 month-old) rats in the presence and absence
of 1,25diOHVD, aged rats showed higher basal PKC concentrations than
younger rats while basal Ca2+ uptake didn’t differ between the two groups. With
addition of 1,25diOHVD there was a significant increase in both PKC activity and
Ca2+ uptake in the cells from the young rats compared with the aged rats.
Specifically, percentage of PKC activity with 1,25diOHVD treatment was acutely
increased in younger rats while in aged rats there was an acute decrease in
percentage PKC activity returning to normal by 10 and 6 minutes in the young

101
and aged cells, respectively. The results obtained from the rat study suggest
ageing may have a similar effect on sperm total and progressive motilities
through Ca2+/PKC signaling pathways and the patterns of this activity observed
appear to mimic those seen in the motility parameters with 25OHVD of our study.
The results of the rat study also suggest that non-genomic Vitamin D metabolism
is impaired with ageing although it is unknown what effects nuclear Vitamin D
action may have. The association of 25OHVD and prostate volume on motility
parameters may be more difficult to explain. Age and bodyweight together make
up the components that influence prostate volume in the dog in this study as well
as in others [17]. While bodyweight and 25OHVD were not found to have an
association with either total or progressive motility, the opposite was true for age
and 25OHVD and therefore suggests that the age component of prostate volume
may account for the association found. To further support this hypothesis,
clinical BPH with blood in the ejaculate has not been found to have association
with semen quality and fertility [18, 29], although lysis of red blood cells in thawed
frozen semen samples causes detrimental effects on sperm motility and viability
[30]. Studies to determine whether Vitamin D and the VDR are present in the
testes and sperm in the dog and what physiological role Vitamin D plays in sperm
motility are warranted to further detail the associations found in this study.
In univariable analysis of BCF, DAP, DCL VAP, VCL and VSL, each
motility parameter followed a simple positive linear association with serum
25OHVD. It is unknown whether increases in BCF with increasing 25OHVD in
the dog have clinical relevance, as this parameter did not correlate to fertility in

102
this species [24]. DAP and VAP, as well as, DCL and VCL are related
parameters (increases in distance travelled per examination time and the
increase in distance travelled per second) and it is therefore not surprising that
both sets of parameters saw similar increases with increasing 25OHVD
concentrations. It is also important to note that VCL is a component of defining
both total and progressive motilities via CASA and therefore its significance is not
surprising. Increases in VSL were also significant with increasing 25OHVD
concentrations and although DSL association with 25OHVD approached
significance (p=0.06), it was not statistically significant possibly due to a lack of
statistical power due to a small sample size. Fertility has correlated well with
these parameters in dogs [24] and improvements in average, straight and
curvilinear distances and velocities of sperm may be translated as the ability of
sperm to traverse the female reproductive tract efficiently. No other published
studies relating Vitamin D to these motility parameters exist at present. The
mechanism of Vitamin D action on these parameters may occur during crucial
development of the midpiece (mitochondrial/axoneme function) and/or tail
(axoneme) regions of spermatids in the testis. Identification of the VDR in the
midpiece of immature human spermatozoa was found [31] indicating a possible
role of Vitamin D in the development and maturation of sperm in this species and
encourages further study in the dog to determine if the same holds true. A recent
study has shown VDR to be present in the mitochondria of human platelets [32]
suggesting that the VDR localization in the midpiece of human spermatozoa may
be specific to the mitochondria. The presence of the VDR in the tail regions of

103
mature sperm suggests that Vitamin D may influence axoneme structure and this
hypothesis is supported by studies that have shown Vitamin D deficiency during
fetal development in rats alters protein structure of certain cytoskeletal elements,
specifically -tubulins, in rat brain cells [33]. These -tubulin proteins are also
present only in the midpiece and tail regions of human sperm [34]. If Vitamin D
action also holds true for maintaining normal axonemic structure in the sperm tail,
sperm propulsion may therefore be affected. All developmental stages of sperm
have been shown to express the VDR and 25OHVD hydroxylase enzymes in rats
and men, as well as Sertoli and Leydig cells suggesting that Vitamin D has a role,
directly and/or indirectly on sperm maturation. Calcium signaling through
cytosolic VDR may also be the trigger to fuel mechanisms involved in increasing
distance travelled and velocity of sperm, however a negative effect at high
normal 25OHVD concentrations was not observed as with total and progressive
motilities suggesting that this concentration limited mechanism may not be
involved.
There was an increase in BCF with increasing age and 25OHVD in
bivariable analysis. It is uncertain what this association may mean to age-related
male dog fertility and 25OHVD. There were increases in DCL and VCL at
younger ages (4-8 years of age) and smaller prostate volumes (log2volume=3.5-
4.5) while both decreased at older ages (10 years) and larger prostate volumes
(log2volume=5.5) with increasing 25OHVD. Similarly to total and progressive
motility, age-related effects on DCL and VCL may be dependent on Ca2+/PKC
physiology in the mitochondria of the sperm midpiece. It is also possible

104
however, that age-related influences on nuclear Vitamin D effects with respect to
structural protein metabolism may also be involved. As mentioned previously -
tubulins are necessary for axoneme structure in the midpiece and tail regions of
spermatozoa. In a study investigating age-related structural effects in human
fibroblasts, both actin and -tubulin proteins were decreased in senescent
compared with fetal fibroblast culture [35]. F-actin is a structural protein
component of the sperm flagellum and cleavage of F-actin from this structure has
been shown to hamper sperm motility in guinea pigs [36]. In addition, 25OHVD
action on sperm structure in ageing may occur through impaired inhibition of
nuclear factor B (NF B) signaling – known to be a target of nuclear Vitamin D
signaling (see review by Nagpal [37]). NF B has been suggested to be a
mediator through which actin and tubulin decrease and vimentin increases in
senescent human fibroblast cells [35]. Whether a similar mechanism exists in
developing spermatocytes remains unknown. DSL also increased at smaller
transformed prostate volumes and decreased less dramatically at larger
transformed prostate volumes with increasing 25OHVD. Although no affect of
age was seen it is possible that the age component of prostate volume is still a
factor in DSL outcome with 25OHVD. The small sample size and resulting
statistical power of the study may not have been high enough to detect age
effects with 25OHVD. Although, our study found the same positive association of
prostate volume to bodyweight and age as in previous studies [16, 38, 39], it is
possible that bodyweight and age are not the factors of prostate volume
responsible for DSL outcomes with increasing 25OHVD. It is perhaps possible

105
that altered Vitamin D physiology of the enlarged prostate exists. BPH has not
been associated with infertility in the dog, however only a few components of
prostatic fluid have been identified in previous studies [40]. Men with BPH and
chronic pelvic pain syndrome have been shown to have higher concentrations of
IL-8 [41] and the VDR agonist elocalcitol has been shown to decrease these
concentrations in seminal plasma [42]. Whether, seminal plasma components
are altered in BPH in the dog is unknown and the potential influence of Vitamin D
on prostatic secretion and possible semen effects on ejaculated sperm is worth
further study.
Both ALH and AOC increased with increasing 25OHVD at younger ages
(4-6 years) and decreased at older ages (8-10 years). Both parameters are
positively correlated with fertility in dogs [24]. In human sperm, ALH is
associated with fertilizing potential by overcoming the barriers of cervical mucus
and the peri-oocyte envelope before fertilization of the oocyte can take place [43-
45] and increases in this motility parameter suggest a positive role for Vitamin D
in male fertility. Measuring AOC determines whether sperm during analysis are
being passively moved. Sperm with an AOC level of <9.5 have been defined as
being immotile and/or dead [13]. Therefore increases in this parameter indicate
that sperm are able to change direction according to changes and/or obstacles in
their path or are more sensitive and more likely to respond to those obstacles. It
is likely that age-related 25OHVD effects of these parameters are mediated
through the same energy and structural mechanisms as the motility parameters
already mentioned.

106
Normal sperm morphology followed a quadratic pattern similarly to total
and progressive motilities. As with the motility parameters, a positive effect of
25OHVD on normal morphology in the low to mid-range of normal 25OHVD
concentrations (70-160 nmol) and a negative effect in the high end of the range
(>160 nmol/l) were observed. Desirable normal morphology (>80%) was
observed in the 140-180 nmol/l range. As normal spermatogenesis logically
results in production of morphologically normal sperm it is possible that Vitamin D
may exert an effect at the level of spermatocytes, Sertoli and Leydig cells in the
testis of the dog; this is supported by studies in rats that have also shown
reduced fertility and degenerative changes in testis and spermatogonia in
Vitamin D deficiency [2]. Certain local factors such as cytokines and growth
factors are known to have a part in regulation of spermatogenesis [46]. Among
these factors interleukins (ILs), insulin-like growth factors (IGFs), and tumor
necrosis factor- alpha (TNF ) action are influenced by Vitamin D in other tissues
and more specifically, Vitamin D has been shown to negate the effects of these
factors (see review by Nagpal [37]). The inflammatory cytokines IL, IGF and
TNF are thought to be involved in normal tight-junction complex recycling,
however, they can cause disruption of the tight-junction adhesions and
consequently compromise the BTB at greater than physiological normal
concentrations [47-49]. It is possible that maintenance of these tight-junctions is
mediated through Vitamin D and may therefore have a protective function of the
developing sperm from inflammatory insult. Vitamin D has also been shown to
influence enzymes such as phospholipase C (PLC) that are associated with

107
cellular differentiation, while stimulating insulin-like growth factor binding protein 3
(IGFBP-3) secretion to bind and inhibit insulin-like growth factor (IGF) [50].
Perhaps there is a dose-dependent effect of Vitamin D in the testes and germ
cells that causes IGF inhibition at both high and low concentrations yet switches
to stimulate PLC action at physiologically optimal concentrations. The
percentage of detached heads decreased with increasing concentrations of
25OHVD indicating a possible role for Vitamin D in the development of the head
midpiece/tail connection similarly to the prior discussion of normal morphology
and supports our hypothesis for a spermatogenic role of Vitamin D. Although no
other morphological defects of sperm were associated with 25OHVD it is possible
that subtler defects as assessed by scanning or transmission electron
microscopy may have revealed a different outcome with respect to 25OHVD.
Increases in percentage of morphologically normal sperm were associated
with increasing 25OHVD at younger ages (4-8 years) and smaller transformed
prostate volumes (log2volume=3.5-4.5) and decreased with increasing 25OHVD
at older ages (10 years) and larger transformed prostate volumes
(log2volume=5.5). Age and prostate volume influences were most likely due to
the possible age effect on spermatogenesis mentioned previously in the section
on total and progressive motilities. Whether this positive sperm morphological
trait occurs through genomic and/or non-genomic Vitamin D effects remains to be
elucidated. Similarly, percentage of head defects decreased at smaller
transformed prostate volumes (log2volume=3.5-4.5) and increased at larger
transformed prostate volumes (log2volume=5.5). Whether the reason was due to

108
a masked age effect on Vitamin D physiology during spermatogenesis and/or an
influence of Vitamin D associated changes in prostatic fluid composition causing
acrosomal or membrane damage to spermatozoa necessitates further research.
There were a large number of dog breeds compared with the number of
individuals in our study and therefore we were unable to account for breed effect
or breed trends in this study. Nevertheless, the information obtained on Vitamin
D in this population of dogs has the potential to open new avenues of research
and possible treatment for cases of poor or sub-normal semen characteristics in
the dog. It is important to note that the majority of dogs enrolled in the study
were sexually rested for at least one year prior to examination and few had been
used for breeding therefore breeding histories were not analyzed.
Although many forms of Vitamin D exist, 25OHVD most accurately
represents the physiological Vitamin D reserves due to its long half-life of 2-3
weeks and also because the first hydroxylation step in its formation is
unregulated; dependent only on substrate supply [51]. It is this supply of
25OHVD that feeds the conversion to the more active forms of Vitamin D such as
1,25diOHVD in specific tissues such as the prostate and testes. Therefore,
25OHVD was considered to be the rate-limiting step in the enzymatic cascade in
Vitamin D action within these tissues and was thus the chosen substrate to be
assayed [51]. It is also the main circulating form of Vitamin D in the dog.
Day length and ultraviolet (UV) light exposure in the Northern Hemisphere,
as determined by time of year of sampling had no effect on 25OHVD
concentrations in dogs. These data further support other studies determining

109
that dogs are unable to synthesize Vitamin D through the action of UV radiation
on the skin [52, 53]. Vitamin D concentrations were also not influenced by type
of diet, whether raw or commercial indicating appropriate Vitamin D intake by
study subjects was not dependent on type of diet and all diets were able to meet
the daily requirements of the study population.
Experimental trials with Vitamin D analogues in human cases of BPH have
shown a significant shrinkage of enlarged prostates [5, 54, 55]. This study
showed that 25OHVD concentrations were not associated with prostate size or
BPH in the study population of dogs. This lack of significance may be due to the
sample size being too small to detect any differences in the population. BPH was
also not confirmed using histology in this study. Instead, the presence of BPH
was defined according to clinical signs, predetermined cut-off points for prostatic
dimensions on ultrasound examination, and elimination of other pathologies.
Controlling for BPH in prostate volume analysis was done to determine whether
enlargement of the prostate was associated with Vitamin D concentrations
independent of our diagnostic criteria for BPH. Future studies using larger
sample sizes and histological confirmation of BPH are warranted and may reveal
different results.
Adiponectin, a marker for body mass index, has been negatively
associated with Vitamin D concentrations in humans [56]. More specifically,
obese patients with lowered adiponectin concentrations have the strongest
associations with Vitamin D concentrations bordering on deficiency [56].
Adiponectin has also been correlated to body condition scoring in dogs [57].

110
Therefore a possible influence of body condition on Vitamin D concentrations
might exist in the canine. In our study population there were only two dogs
(Golden Retriever and Mastiff cross) that were considered slightly overweight
with the majority of the population falling into the ideal body condition category
and were not considered to be Vitamin D deficient (104 and 115 nmol/l 25OHVD,
respectively). Further study looking at Vitamin D status, body condition score
and adiponectin concentrations in a wide range of dogs would be worthwhile in
understanding Vitamin D physiology in this species. It is important to mention
that, statistically, a great many comparisons were made in our study and the risk
for Type I error (false positives) is increased with the large amount of
associations found to be significant. Unfortunately, there is no reliable method
for correcting for this issue and identifying this type of error in the present study.
Methods that have been suggested i.e. Bonferroni come with their own risk of
increasing Type II error (false negatives).
In conclusion, serum 25OHVD concentrations between 120-180 nmol/l
were associated with desirable semen characteristics that are connected to dog
fertility. A possible new tool in diagnosing and treating abnormalities in the
spermiogram is an exciting concept. Further research aimed at identifying
Vitamin D and/or its receptor in reproductive tissues and the mechanisms by
which they exert their effects would be beneficial to increasing our understanding
of Vitamin D in spermatogenesis and reproductive health in the dog.

111
Table 2.1 Technical Parameters for SpermVision™ CASA
Parameter
Depth of chamber 20um
Light adjustment 96-104
Volume per Chamber 2ul
Temperature 37°C
Sperm concentration Variable
Area of sperm heads 20-60 um2
Number of fields or Cell number 7 or 1000
Number cells/field 50-200
Frame rate 60/sec
Total motility VCL > 15
Progressively Fast VCL >15, STR>0.9
Linear STR >0.9, LIN>5
Immotile AOC <7, DSL <3
Local motility DSL <6
Hyperactive VCL>118, ALH>6.5, LIN<0.5
Non-Linear LIN<=0.5, STR<=0.9
Curvilinear DAP/Radius >=3, LIN<0.5
Incubation time 2 mins
Modified from Schäfer-Somi and Aurich, 2007[13]

112
Table 2.2 Classification scheme for determination of the presence of BPH by ultrasonography in 28 dogs, March-December 2009, Ontario, Canada
Prostatic Measurement
(Maximum of Length, Width or Height)
Bodyweight
> 3.0 cm 20 kg
> 3.5 cm 40 kg
> 4.0 cm > 60 kg
> 4.5 cm 80 kg
Classification based on arbitrary cut-points.

113
Table 2.3 Characteristics of the study population of 28 dogs, in Ontario, Canada March-December 2009.
. *SD=standard deviation SEM=standard error of the mean Min = Minimum value Max = Maximum value
Variable Mean SD SEM Min Max
Age (years) 6.02 2.05 0.38 2.00 11.00
Body Weight (kgs) 35.36 15.84 2.94 9.20 78.20
Vitamin D (nmol/l) 140.50 37.05 5.72 72.00 203.00

114
Table 2.4 Characteristics of the study population of 22 dogs from which semen was collected in Ontario, Canada March-December 2009.
*SD=standard deviation SEM=standard error of the mean Min = Minimum value Max = Maximum value
Variable Mean SD SEM Min Max
Age (years) 6.25 2.09 0.45 4.00 11.00
Body Weight (kgs) 35.47 14.71 3.14 9.20 71.30
Vitamin D (nmol/l) 141.86 35.03 7.47 72.00 189.00
Total sperm (106) 524.26 324.89 69.27 33.00 1000.00
Motility (%) 89.1 15.74 3.36 27.00 98.60
Progressive Motility (%) 80.19 18.57 3.96 13.30 95.70
Normal sperm (%) 61.62 29.58 6.45 0 95.00
Head Defects (%) 10.76 19.97 4.36 0 78.00
Midpiece Defects (%) 8.19 5.90 1.29 0 20.00
Tail Defects (%) 3.38 4.43 0.97 0 16.00
Loose Heads (%) 2.00 3.86 0.84 0 16.00
Proximal Droplets (%) 12.00 18.57 4.05 0 61.00
Distal Droplets (%) 1.67 1.62 0.35 0 5.00
DCL (um) 65.12 15.71 3.43 29.78 94.83
DAP (um) 30.77 5.73 1.25 16.60 40.50
DSL (um) 21.76 4.56 1.00 13.68 33.00
VCL (um/s) 144.30 35.23 7.69 67.64 208.70
VAP (um/s) 68.67 13.44 2.93 38.72 92.77
VSL (um/s) 48.65 10.90 2.38 32.11 75.48
LIN (%) 34.19 7.06 1.54 24 50
STR (%) 70.52 7.69 1.68 57 84
WOB (%) 48.05 5.19 1.13 39 60
BCF (Hz) 21.83 1.95 0.43 18.17 26.53
ALH (um) 5.37 1.16 0.25 2.95 7.36
AOC 17.69 3.75 0.82 11.80 24.99

115
Table 2.5 Breed, Vitamin D concentration, age and fertility data on 22 dogs from which semen was collected in Ontario, Canada, 2009 Dog Breed Vitamin
D (nmol/l)
Age (years)
No. Litters
No. Breedings /year
No. Years Total
1 Shiloh Shepherd 95 5.5 7 1-2 5 2 Labrador Retriever 113 5 0 0 0 3 Great Pyrenees 182 4.5 2 <1 2 4 Australian Shepherd 102 5.5 0 0 0 5 Newfoundland 189 4.5 0 0 0 6 Golden Retriever 72 5 0 Multiple Unknown 7 Golden Retriever 104 9 0 0 0 8 Whippet 179 11 6 <1 7 9
Cavalier King Charles Spaniel
117
9
15 2-3 7
10 Belgian Shepherd 186 8.5 0 0 0 11 Labrador Retriever 97 8 3 0.5 6 12
Chesapeake Bay Retriever
175
6.5
0 0 0
13 Boxer 163 6 0 0 0 14 Labrador Retriever 158 5.5 6 1-2 4 15 Whippet 152 5 0 0 0 16 Mastiff Cross 115 7 0 1 0 17 Golden Retriever 170 4.5 0 0 0 18 Shiloh Shepherd 177 5.5 0 0 0 19 German Shepherd 146 4 0 0 0 20
Welsh Cardigan Blue Corgi
128
10
1 3 Unknown
21 Akita 147 4 2 1 2 22 Samoyed 154 4 0 0 0 No.=number

116
Table 2.6 Mean serum Vitamin D concentration, with respect to type of diet, with associated test-statistic, p-value and confidence intervals. Type of Diet Mean Serum
Vitamin D (nmol/l)
F-value p-value 95% Confidence
Interval
Lower
Limit
Upper
Limit
Raw 145.2 0.24 0.79 95.8 194.6
Commercial 140.5 123.4 157.5
Combined 154.0 100.4 207.6

117
Table 2.7 Mean serum Vitamin D concentration, with respect to time of year of sampling, with associated test-statistic, p-value and confidence intervals. Time of Year Mean Serum
Vitamin D (nmol/l)
t-value p-value 95% Confidence
Interval
Lower
Limit
Upper
Limit
Summer 139.6 -0.63 0.54 119.8 159.5
Winter 148.1 127.5 168.7
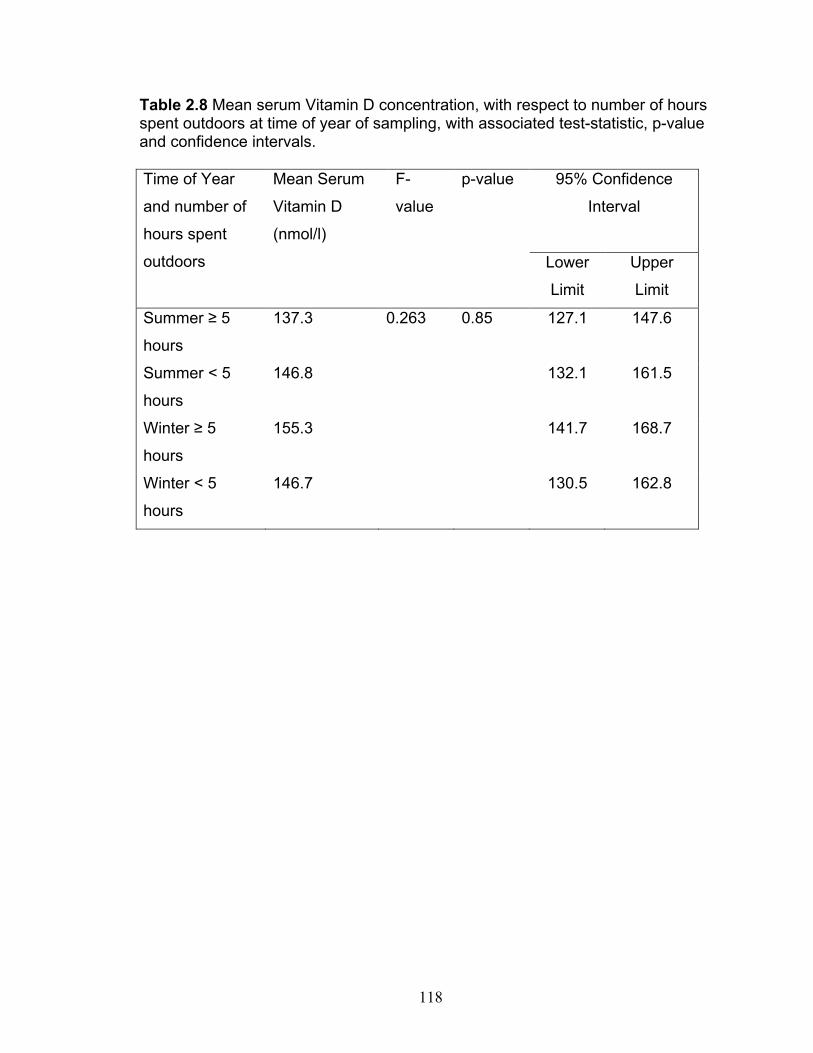
118
Table 2.8 Mean serum Vitamin D concentration, with respect to number of hours spent outdoors at time of year of sampling, with associated test-statistic, p-value and confidence intervals. Time of Year
and number of
hours spent
outdoors
Mean Serum
Vitamin D
(nmol/l)
F-
value
p-value 95% Confidence
Interval
Lower
Limit
Upper
Limit
Summer
hours
137.3 0.263 0.85 127.1 147.6
Summer < 5
hours
146.8 132.1 161.5
Winter
hours
155.3 141.7 168.7
Winter < 5
hours
146.7 130.5 162.8

119
Table 2.9 Mean serum Vitamin D concentration, with respect to BPH status, and associated test-statistic, p-value and confidence intervals. Prostate Status Mean Serum
25OHVD (nmol/l)
t-value p-value 95% Confidence
Interval
Lower
Limit
Upper
Limit
BPH 145.0 0.55 0.59 130.4 159.5
Normal 135.4 81.7 189.1

120
Table 2.10 Univariable simple linear regression analyses of sperm motility parameters with serum 25-hydroxy vitamin D as explanatory variable, with associated coefficient of determination (R2), test statistic and p-value. Motility Parameter R2 value F-value p-value
Total Motility (%) 0.55 10.88 0.0008*
Progressive Motility (%) 0.51 9.32 0.002*
ALH 0.01 2.99 0.10
AOC 0.11 2.45 0.13
BCF (Hz) 0.28 7.23 0.01*
DAP (µm) 0.38 11.61 0.003*
VAP (µm/s) 0.35 10.35 0.005*
DCL (µm) 0.25 6.28 0.02*
VCL (µm/s) 0.25 6.50 0.02*
DSL (µm) 0.31 8.53 0.01*
VSL (µm/s) 0.27 7.16 0.01*
STR 0.00 0.03 0.87
LIN 0.01 0.11 0.75
WOB 0.01 0.18 0.68

121
Table 2.11 Non-significant bivariable linear regressions of motility with explanatory variable regression coefficients (where applicable), coefficients of determination for the overall model (R2), test statistics, and the associated p-values. Explanatory Variable Model Fit Outcome Variable
Explanatory Variable
Regression Coefficient
p-value
R2-value Overall Model
F-value
p-value
Motility 25OHVD 0.02 0.002 Bodyweight -0.01 0.35 0.43 6.65 0.01 Progressive 25OHVD 0.02 0.005 Motility Bodyweight -0.01 0.36 0.38 5.48 0.01 ALH (model 1) 25OHVD - - Bodyweight - - 0.14 1.14 0.26 ALH (model 2) 25OHVD - - Prostate Volume - - 0.14 1.14 0.26 AOC (model 1) 25OHVD - - Bodyweight - - 0.11 1.16 0.33 AOC (model 2) 25OHVD - - Prostate Volume - - 0.13 1.33 0.29 BCF (model 1) 25OHVD 0.03 0.02 Bodyweight -0.01 0.59 0.29 3.63 0.05 BCF (model 2) 25OHVD 0.03 0.01 Prostate Volume 0.75 0.15 0.35 4.95 0.02 DAP (model 1) 25OHVD 0.10 0.004 Age -0.31 0.55 0.39 5.80 0.01 DAP (model 2) 25OHVD 0.10 0.004 Bodyweight 0.04 0.60 0.39 5.72 0.01 DAP (model 3) 25OHVD 0.10 0.004 Prostate Volume 0.56 0.70 0.38 5.62 0.01 DCL 25OHVD - - Bodyweight - - 0.28 3.41 0.06 DSL (model 1) 25OHVD 0.07 0.01 Age 0.34 0.43 0.33 4.51 0.03 DSL (model 2) 25OHVD 0.07 0.01 Bodyweight -0.03 0.64 0.32 4.21 0.03

122
Table 2.11 (continued) Non-significant bivariable linear regressions of motility with explanatory variable regression coefficients (where applicable), coefficients of determination for the overall model (R2), test statistics, and the associated p-values. Explanatory Variable Model Fit Outcome Variable
Explanatory Variable
Regression Coefficient
p-value
R2-value Overall Model
F-value
p-value
VAP (model 1) 25OHVD 0.23 0.006 Age -0.74 0.56 0.37 5.18 0.02 VAP (model 2) 25OHVD 0.23 0.006 Bodyweight 0.06 0.72 0.38 5.00 0.02 VAP (model 3) 25OHVD 0.23 0.006 Prostate Volume 1.17 0.74 0.36 4.99 0.02 VCL (model 1) 25OHVD 0.23 0.006 Bodyweight 0.33 0.49 0.36 5.00 0.02 VSL (model 1) 25OHVD 0.17 002 Age 0.70 0.51 0.29 3.70 0.05 VSL (model 2) 25OHVD 0.17 0.02 Bodyweight -0.07 0.61 0.28 3.57 0.05 VSL (model 3) 25OHVD - - Prostate Volume - - 0.28 3.42 0.06 STR (model 1) 25OHVD - - Age - - 0.25 2.87 0.08 STR (model 2) 25OHVD - - Bodyweight - - 0.12 1.20 0.32 STR (model 3) 25OHVD - - Prostate Volume - - 0.00 0.03 0.97 LIN (model 1) 25OHVD - - Age - - 0.27 3.13 0.07 LIN (model 2) 25OHVD - - Bodyweight - - 0.10 0.94 0.41 LIN (model 3) 25OHVD - - Prostate Volume - - 0.01 0.05 0.95 WOB (model 1) 25OHVD - - Age - - 0.21 2.31 0.13 WOB (model 2) 25OHVD - - Bodyweight - - 0.06 0.51 0.61 WOB (model 3) 25OHVD - - Prostate Volume - - 0.01 0.09 0.92

123
Table 2.12 Three bivariable linear regressions of morphology parameters and associated coefficients of determination (R2), test-statistic and p-values Response (%) Explanatory
Variable with 25OHVD in the model
R2-value F-value p-value of Overall Model
Normal Age 0.49 5.05 0.01* Bodyweight 0.28 3.38 0.06 Prostate Volume
0.52 5.77 0.01*
Head Defects Age 0.17 1.78 0.20 Bodyweight 0.16 1.70 0.21 Prostate Volume
0.39 3.47 0.04*
Midpiece Defects Age 0.07 0.66 0.53 Bodyweight 0.03 0.28 0.76 Prostate Volume
0.03 0.25 0.78
Proximal Droplets Age 0.17 1.74 0.21 Bodyweight 0.10 0.94 0.41 Prostate Volume
0.01 0.06 0.95
Distal Droplets Age 0.24 2.70 0.10 Bodyweight 0.04 0.39 0.69 Prostate Volume
0.20 2.14 0.15
Tail Defects Age 0.02 0.19 0.83 Bodyweight 0.00 0.03 0.96 Prostate Volume 0.03 0.22 0.81

124
Figure 2.1 Predicted sperm motility as a response to normal serum 25OHVD range in dogs. Back-transformation of multiple linear regression model logitMotility=-7.64+0.135VD-0.0004VD2 (R2=0.55, p=0.0008). VD=serum 25-hydroxy vitamin D. CI= 95%Confidence Interval. Predicted Motility (%),
Lower CI, Upper CI

125
Figure 2.2 Progressive motility as a response to normal serum 25OHVD range in dogs. Back-transformation of multiple linear regression model logitPMotility=-7.92+0.128VD-0.0004VD2(R2=0.51, p=0.002). VD=serum 25-hydroxy vitamin D. CI=95% Confidence Interval. Predicted Progressive Motility (%),
Lower CI, Upper CI.

126
Figure 2.3 Predicted sperm motility in response to normal serum 25OHVD range at multiple ages. Back-transformation of multiple linear regression model logitMotility=-46.19 +5.66A+9.72lnVD-1.12A*lnVD (R2 = 0.57, p=0.0019). VD=serum 25-hydroxy vitamin D. CI= 95% Confidence Interval. A=Age Predicted Motility (%), Lower CI, Upper CI.

127
Figure 2.4 Predicted sperm motility in response to normal serum 25OHVD range at various transformed prostate volumes. Back-transformation of multiple regression model logitMotility=-16.7 +3.69Vol +0.14VD-0.03Vol*VD (R2=0.72, p<0.0001). VD=serum 25-hydroxy vitamin D. CI= 95% Confidence Interval. Vol=Prostate Volume.
Predicted Motility (%), Lower CI, Upper CI.

128
Figure 2.5 Predicted progressive motility in response to normal serum 25OHVD range at various ages. Back-transformation of multiple regression model logitPMotility=-8.1+1.11A +0.06VD-0.007A*VD (R2=0.57, p=0.002). VD=serum 25-hydroxy vitamin D. CI=95% Confidence Interval. A=Age. Predicted Progressive Motility (%),
Lower CI, Upper CI.

129
Figure 2.6 Predicted progressive motility in response to normal serum 25OHVD range at various transformed prostate volumes. Back-transformation of multiple regression model logitPMotility=-14.2 +3.0Vol +0.11VD -002Vol*VD (R2=0.60, p=0.001). VD=serum 25-hydroxy vitamin D. CI= 95% Confidence Interval. Vol=Prostate Volume. Predicted Progressive Motility (%), Lower CI, Upper CI.

130
Figure. 2.7 Predicted BCF in response to normal serum 25OHVD range at various ages. Multiple regression model BCF=14.9+0.422A +0.03VD (R2=0.47, p=0.003). VD=serum 25-hydroxy vitamin D. CI=95% Confidence Interval. A=Age
Predicted BCF (%), Lower CI, Upper CI.

131
Figure. 2.8 Predicted DCL in response to normal serum 25OHVD range at various ages. Multiple regression model DCL=-56.5 +0.9VD+14.5A-0.11VD*A (R2=0.51, p=0.006). VD=serum 25-hydroxy vitamin D. CI=95% Confidence Interval. A=Age Predicted DCL (um), Lower CI, Upper CI.

132
Figure 2.9 Predicted DCL in response to normal serum 25OHVD range at various transformed prostate volumes. Back-transformation of multiple regression model DCL=-1790.1+488.7Vol+305.7lnVD -65.9Vol*lnVD-18.8Vol2 (R2=0.53, p=0.01). VD=serum 25-hydroxy vitamin D. CI= 95% Confidence Interval. Vol=Prostate Volume. Predicted DCL (um), Lower CI, Upper CI.

133
Figure 2.10 Predicted VCL in response to normal serum 25OHVD range at various ages. Multiple regression model VCL=-129.7+2.0VD+32.5A -0.11VD*A (R2=0.51, p=0.006). VD=serum 25-hydroxy vitamin D. CI=95% Confidence Interval. A=Age. Predicted VCL (um/s), Lower CI, Upper CI.

134
Figure 2.11 Predicted VCL in response to normal serum 25OHVD range at various transformed prostate volumes. Multiple regression model VCL=-1405.2+525.2Vol+5.6VD -1.2Vol*VD-40.79Vol2 (R2=0.52, p=0.01). VD=serum 25-hydroxy vitamin D. CI=95% Confidence Interval. Vol=Prostate Volume. Predicted VCL (um/s), Lower CI,
Upper CI.

135
Figure 2.12 Predicted DSL in response to normal serum 25OHVD range at various transformed prostate volumes. Multiple regression model DSL=-34.1+10.3Vol+0.403VD-0.075Vol*VD (R2= 0.46, p=0.01). VD=serum 25-hydroxy vitamin D. CI=95% Confidence Interval. Vol=Prostate Volume. Predicted DSL (um), Lower CI, Upper CI.

136
Figure 2.13 Predicted ALH in response to normal serum 25OHVD range at various ages. Multiple regression model ALH=-3.1+1.1*A+0.06VD-0.01A*VD (R2=0.42, p=0.02). VD=serum 25-hydroxy vitamin D. CI=95% Confidence Interval. A=Age. Predicted ALH (um), Lower CI, Upper CI.

137
Figure 2.14 Predicted AOC in response to normal serum 25OHVD range at various ages. Multiple regression model AOC=-6.6+3.1*A+0.2VD-0.02A*VD (R2=0.38, p=0.04). VD=serum 25-hydroxy vitamin D. CI=95% Confidence Interval. A=Age. Predicted AOC (degrees), Lower CI, Upper CI

138
Figure 2.15 Predicted Normal sperm in response to normal serum 25OHVD range Back-transformation model logitNormal=-17.79+0.246VD-0.0008VD2 (R2=0.43, p=0.01). Predicted Normal Sperm (%), Lower CI,
Upper CI. VD=serum 25-hydroxy vitamin D. CI= 95% Confidence Interval.

139
Figure 2.16 Predicted Loose Heads in response to normal serum 25OHVD range. Back-transformation of logitLooseHeads=-1.62+0.22VD (R2=0.31, p=0.01). Predicted Loose Heads (%), Lower CI, Upper CI. VD=serum 25-hydroxy vitamin D. CI=95% Confidence Interval.

140
Figure 2.17 Predicted Normal sperm in response to normal serum 25OHVD range at various ages. Back-transformation of multiple regression model logitNormal=-17.59+2.09A +0.12VD-0.01A*VD (R2=0.49, p=0.01). Predicted Normal Sperm (%), Lower CI, Upper CI. VD=serum 25-hydroxy vitamin D. CI=95% Confidence Interval. A=Age.

141
Figure 2.18 Predicted Normal sperm in response to normal serum 25OHVD range at various transformed prostate volumes. Back-transformation of multiple regression logitNormal=-29.2+5.77Vol +0.21VD-0.04Vol*VD (R2=0.52, p=0.01
Predicted Normal Sperm (%), Lower CI, Upper CI. VD=serum 25-hydroxy vitamin D. CI= 95% Confidence Interval. Vol=Prostate Volume.

142
Figure 2.19 Predicted head defects in response to normal serum 25OHVD range at various transformed prostatic volumes. Back-transformation of multiple logitHead=24.5-5.55Vol-0.19VD+0.04Vol*VD (R2=0.39, p=0.04). Predicted Head Defects (%), Lower CI, Upper CI. VD=serum 25-hydroxy vitamin D. CI=95% Confidence Interval. Vol=Prostate Volume.

143
References
[1] G. G. Kwiecinksi, et al., "1,25-Dihydroxyvitamin D3 restores fertility of vitamin D-deficient female rats," Am J Physiol, vol. 256, pp. E483-7, Apr 1989.
[2] G. G. Kwiecinski, et al., "Vitamin D is necessary for reproductive functions of the male rat," J Nutr, vol. 119, pp. 741-4, May 1989.
[3] M. F. Holick, "The vitamin D deficiency pandemic and consequences for nonskeletal health: mechanisms of action," Mol Aspects Med, vol. 29, pp. 361-8, Dec 2008.
[4] E. Giovannucci, "Strengths and limitations of current epidemiologic studies: vitamin D as a modifier of colon and prostate cancer risk," Nutr Rev, vol. 65, pp. S77-9, Aug 2007.
[5] L. Adorini, et al., "Vitamin D receptor agonists target static, dynamic, and inflammatory components of benign prostatic hyperplasia," Ann N Y Acad Sci, vol. 1193, pp. 146-52, Apr 2010.
[6] S. Aquila, et al., "Human sperm anatomy: ultrastructural localization of 1alpha,25-dihydroxyvitamin D receptor and its possible role in the human male gamete," J Anat, vol. 213, pp. 555-64, Nov 2008.
[7] M. Blomberg Jensen, et al., "Vitamin D receptor and vitamin D metabolizing enzymes are expressed in the human male reproductive tract," Hum Reprod, vol. 25, pp. 1303-11, May 2010.
[8] J. A. Johnson, et al., "Immunohistochemical detection and distribution of the 1,25-dihydroxyvitamin D3 receptor in rat reproductive tissues," Histochem Cell Biol, vol. 105, pp. 7-15, Jan 1996.
[9] M. Kivineva, et al., "Localization of 1,25-dihydroxyvitamin D3 receptor (VDR) expression in human prostate," J Steroid Biochem Mol Biol, vol. 66, pp. 121-7, Aug 1998.
[10] S. Sood, et al., "Effect of vitamin D repletion on testicular function in vitamin D-deficient rats," Ann Nutr Metab, vol. 39, pp. 95-8, 1995.
[11] C. F. Garland, et al., "Vitamin D for cancer prevention: global perspective," Ann Epidemiol, vol. 19, pp. 468-83, Jul 2009.
[12] S. D. Johnston, et al., Semen Collection, Evaluation and Preservation, 1st ed. Philadelphia: WB Saunders, 2001.
[13] S. Schafer-Somi and C. Aurich, "Use of a new computer-assisted sperm analyzer for the assessment of motility and viability of dog spermatozoa and evaluation of four different semen extenders for predilution," Anim Reprod Sci, vol. 102, pp. 1-13, Nov 2007.
[14] T. Rigau, et al., "Effects of glucose and fructose on motility patterns of dog spermatozoa from fresh ejaculates," Theriogenology, vol. 56, pp. 801-15, Sep 15 2001.
[15] A. I. Pena, et al., "Studies on the intracellular Ca2+ concentration of thawed dog spermatozoa: influence of Equex from different sources, two thawing diluents and post-thaw incubation in capacitating conditions," Reprod Domest Anim, vol. 38, pp. 27-35, Feb 2003.

144
[16] G. Atalan, et al., "Ultrasonographic estimation of prostatic size in canine cadavers," Res Vet Sci, vol. 67, pp. 7-15, Aug 1999.
[17] K. Kamolpatana, et al., "Determination of canine prostatic volume using transabdominal ultrasonography," Vet Radiol Ultrasound, vol. 41, pp. 73-7, Jan-Feb 2000.
[18] S. D. Johnston, et al., "Prostatic disorders in the dog," Anim Reprod Sci, vol. 60-61, pp. 405-15, Jul 2 2000.
[19] SASInstitute. (2011, Goodness of Fit Tests (Third Edition ed.). Available: http://support.sas.com/documentation/cdl/en/procstat/63104/HTML/default/viewer.htm#procstat_univariate_sect037.htm
[20] R. Sokal and F. Rohlf, Biometry: The principles and practice of statistics in biological research, 3rd ed. New York: W.H. Freeman, 1995.
[21] F. Grubbs, "Procedures for Detecting Observations in Samples," Technometrics, vol. 11, pp. 1-21, 1969.
[22] P. B. Farrell, et al., "Quantification of bull sperm characteristics measured by computer-assisted sperm analysis (CASA) and the relationship to fertility," Theriogenology, vol. 49, pp. 871-9, Mar 1998.
[23] Y. Hirano, et al., "Relationships between sperm motility characteristics assessed by the computer-aided sperm analysis (CASA) and fertilization rates in vitro," J Assist Reprod Genet, vol. 18, pp. 213-8, Apr 2001.
[24] T. Rijsselaere, et al., "Effect of body weight, age and breeding history on canine sperm quality parameters measured by the Hamilton-Thorne analyser," Reprod Domest Anim, vol. 42, pp. 143-8, Apr 2007.
[25] M. Blomberg Jensen, et al., "Vitamin D is positively associated with sperm motility and increases intracellular calcium in human spermatozoa," Hum Reprod, vol. 26, pp. 1307-17, Jun 2011.
[26] K. Schuh, et al., "Plasma membrane Ca2+ ATPase 4 is required for sperm motility and male fertility," J Biol Chem, vol. 279, pp. 28220-6, Jul 2 2004.
[27] Q. X. Zhu, et al., "Turning point of age for semen quality: a population-based study in Chinese men," Fertil Steril, vol. 96, pp. 572-6, Sep 2011.
[28] G. Balogh, et al., "Influence of age on 1,25(OH)2-vitamin D3 activation of protein kinase C in rat duodenum," Mol Cell Endocrinol, vol. 129, pp. 127-33, May 16 1997.
[29] K. Kamolpatana, et al., "Effect of finasteride on serum concentrations of dihydrotestosterone and testosterone in three clinically normal sexually intact adult male dogs," Am J Vet Res, vol. 59, pp. 762-4, Jun 1998.
[30] T. Rijsselaere, et al., "Effect of blood admixture on in vitro survival of chilled and frozen-thawed canine spermatozoa," Theriogenology, vol. 61, pp. 1589-602, May 2004.
[31] M. Blomberg Jensen, et al., "Expression of the vitamin D receptor, 25-hydroxylases, 1alpha-hydroxylase and 24-hydroxylase in the human kidney and renal clear cell cancer," J Steroid Biochem Mol Biol, vol. 121, pp. 376-82, Jul 2010.
[32] F. Silvagno, et al., "Mitochondrial localization of vitamin D receptor in human platelets and differentiated megakaryocytes," PLoS One, vol. 5, p. e8670, 2010.

145
[33] L. Almeras, et al., "Developmental vitamin D deficiency alters brain protein expression in the adult rat: implications for neuropsychiatric disorders," Proteomics, vol. 7, pp. 769-80, Mar 2007.
[34] K. Kadam, et al., "Spatial distribution of actin and tubulin in human sperm nuclear matrix-intermediate filament whole mounts-a new paradigm," Microsc Res Tech, vol. 70, pp. 589-98, Jul 2007.
[35] K. Nishio and A. Inoue, "Senescence-associated alterations of cytoskeleton: extraordinary production of vimentin that anchors cytoplasmic p53 in senescent human fibroblasts," Histochem Cell Biol, vol. 123, pp. 263-73, Mar 2005.
[36] Y. Azamar, et al., "F-actin involvement in guinea pig sperm motility," Mol Reprod Dev, vol. 74, pp. 312-20, Mar 2007.
[37] S. Nagpal, et al., "Noncalcemic actions of vitamin D receptor ligands," Endocr Rev, vol. 26, pp. 662-87, Aug 2005.
[38] S. J. Berry, et al., "Effects of aging on prostate growth in beagles," Am J Physiol, vol. 250, pp. R1039-46, Jun 1986.
[39] C. B. Brendler, et al., "Spontaneous benign prostatic hyperplasia in the beagle. Age-associated changes in serum hormone levels, and the morphology and secretory function of the canine prostate," J Clin Invest, vol. 71, pp. 1114-23, May 1983.
[40] J. E. Branam, et al., "Selected physical and chemical characteristics of prostatic fluid collected by ejaculation from healthy dogs and from dogs with bacterial prostatitis," Am J Vet Res, vol. 45, pp. 825-9, Apr 1984.
[41] G. Penna, et al., "Seminal plasma cytokines and chemokines in prostate inflammation: interleukin 8 as a predictive biomarker in chronic prostatitis/chronic pelvic pain syndrome and benign prostatic hyperplasia," Eur Urol, vol. 51, pp. 524-33; discussion 533, Feb 2007.
[42] G. Penna, et al., "The vitamin D receptor agonist elocalcitol inhibits IL-8-dependent benign prostatic hyperplasia stromal cell proliferation and inflammatory response by targeting the RhoA/Rho kinase and NF-kappaB pathways," Prostate, vol. 69, pp. 480-93, Apr 1 2009.
[43] R. J. Aitken, "Diagnostic value of the zona-free hamster oocyte penetration test and sperm movement characteristics in oligozoospermia," Int J Androl, vol. 8, pp. 348-56, Oct 1985.
[44] C. Jeulin, et al., "Sperm factors related to failure of human in-vitro fertilization," J Reprod Fertil, vol. 76, pp. 735-44, Mar 1986.
[45] T. A. Bongso, et al., "Effect of sperm motility on human in vitro fertilization," Arch Androl, vol. 22, pp. 185-90, 1989.
[46] C. Y. Cheng, et al., "Regulation of spermatogenesis in the microenvironment of the seminiferous epithelium: new insights and advances," Mol Cell Endocrinol, vol. 315, pp. 49-56, Feb 5 2010.
[47] P. P. Lie, et al., "The biology of interleukin-1: emerging concepts in the regulation of the actin cytoskeleton and cell junction dynamics," Cell Mol Life Sci, Jul 9 2011.

146
[48] D. Pretzer, et al., "Growth factors (EGF, IGF-I) modulate the morphological differentiation of adult marmoset (Callithrix jacchus) Sertoli cells in vitro," J Androl, vol. 15, pp. 398-409, Sep-Oct 1994.
[49] W. Y. Lui, et al., "TGF-betas: their role in testicular function and Sertoli cell tight junction dynamics," Int J Androl, vol. 26, pp. 147-60, Jun 2003.
[50] J. C. Fleet, "Rapid, membrane-initiated actions of 1,25 dihydroxyvitamin D: what are they and what do they mean?," J Nutr, vol. 134, pp. 3215-8, Dec 2004.
[51] A. M. Wootton, "Improving the measurement of 25-hydroxyvitamin D," Clin Biochem Rev, vol. 26, pp. 33-6, Feb 2005.
[52] K. L. How, et al., "Dietary vitamin D dependence of cat and dog due to inadequate cutaneous synthesis of vitamin D," Gen Comp Endocrinol, vol. 96, pp. 12-8, Oct 1994.
[53] H. A. Hazewinkel and M. A. Tryfonidou, "Vitamin D3 metabolism in dogs," Mol Cell Endocrinol, vol. 197, pp. 23-33, Nov 29 2002.
[54] E. Colli, et al., "BXL628, a novel vitamin D3 analog arrests prostate growth in patients with benign prostatic hyperplasia: a randomized clinical trial," Eur Urol, vol. 49, pp. 82-6, Jan 2006.
[55] C. Crescioli, et al., "Inhibition of prostate cell growth by BXL-628, a calcitriol analogue selected for a phase II clinical trial in patients with benign prostate hyperplasia," Eur J Endocrinol, vol. 150, pp. 591-603, Apr 2004.
[56] A. Vaidya, et al., "The Independent Association Between 25-Hydroxyvitamin D and Adiponectin and Its Relation With BMI in Two Large Cohorts: The NHS and the HPFS," Obesity (Silver Spring), Jul 14 2011.
[57] R. Ricci and F. Bevilacqua, "The potential role of leptin and adiponectin in obesity: A comparative review," Vet J, May 17 2011.

147
CHAPTER THREE
PROLACTIN, BENIGN PROSTATIC HYPERPLASIA,
PROSTATE VOLUME AND SEMEN PARAMETERS IN THE DOG.
Abstract
Prolactin (PRL) is well known for its role in pregnancy and lactation in the
bitch, however; the role of PRL in male dogs has not been fully characterized.
The aim of this observational study was to determine whether PRL is associated
with benign prostatic hyperplasia (BPH), prostate volume and semen quality
characteristics in stud dogs.
A convenience sample of 29 healthy dogs consisting of 24 different breeds
ranging in age from two to 11 years was studied to determine whether an
association exists between serum PRL concentrations and BPH. Of the dogs
that successfully gave semen samples (n=22; 17 breeds; age range: 4 to 11
years), associations between PRL and multiple characteristics of semen motility
and morphology were also investigated. Student’s t-test revealed no difference
in serum PRL concentrations between dogs with and without BPH (t-value=0.87,
p=0.39). Bivariable linear regression analysis revealed no associations between
prostate volume and PRL when controlling for BPH (R2=0.04, p=0.31). No
significant associations were found between PRL and any of the 14 sperm
motility parameters, as determined by computer assisted motility analysis
(CASA). Two sperm morphology parameters had significant associations with
PRL: percentage proximal droplets (R2=0.23, p=0.03) and percentage midpiece

148
defects with an age interaction (R2=0.42, p=0.03). Serum PRL concentrations
less than 2.5ng/ml were associated with higher percentages of midpiece defects
and proximal cytoplasmic droplets in canine sperm.
Serum PRL concentration ranges in this study of normal healthy dogs
(range: undetectable to 28.24 ng/ml) were higher than previously reported values
(range: undetectable to 6.0 ng/ml) using the same validated ELISA kit. In
conclusion, PRL concentrations had no effect on BPH or sperm motility
parameters, but had a limited negative effect on sperm morphological defects,
namely percentage proximal droplets and midpiece defects.
Keywords: prolactin, prostate, semen, dog, benign prostatic hyperplasia
Abbreviations: Amplitude of lateral head displacement (ALH), Average
orientation change (AOC), Beat cross frequency (BCF), Mean distance (DAP),
Curvilinear distance (DCL), Straight line distance (DSL), Linearity (LIN =
VCL/VSL), Straightness (STR), Mean velocity (VAP), Curvilinear velocity (VCL),
Linear velocity (VSL µm/s), Wobble (WOB=VAP/VCL).
1. Introduction
Prolactin (PRL) is a 199 amino acid (23kDa) protein hormone related to
both growth hormone and placental lactogen [1]. It is well known for its role in
pregnancy/pseudopregnancy and lactation in bitches [2]. Other non-traditional
physiological roles in vertebrate species are still being elucidated including water
and electrolyte balance, growth and development, endocrinology and
metabolism, brain function and behaviour, and immunoregulation [3]. Although

149
its role in reproduction and fertility in men has been studied in relation to
pathological conditions such as prostatic atrophy and teratozoospermia [4, 5],
little is known about the role of canine PRL in the reproductive physiology of the
stud dog.
The PRL feedback loop in male rodents and humans has been shown to
work in the following manner. Prolactin is produced stored and released in the
lactotroph cells of the anterior pituitary similarly to luteinizing hormone (LH) and
follicle stimulating hormone (FSH). PRL upregulates LH receptors in Leydig
cells, thereby stimulating increased secretion of testosterone [1]. Increased
testosterone stimulates estradiol-17 (E2) production by Sertoli cells which in turn
acts on the pituitary lactotroph cells in a positive feedback loop to secrete more
PRL. A direct negative feedback mechanism is accomplished through E2 and
testosterone inhibition of GnRH secretion in the hypothalamus and LH/FSH
secretion in the pituitary. Indirectly, GnRH inhibition of LH/FSH secretion also
down-regulates testosterone secretion [6].
In the rat, prolactin has been suggested to work synergistically with
testosterone to increase 5 -reductase activity in both in vitro and in vivo studies,
and induces growth, differentiation, and hyperplastic changes of the prostate [7-
9]. PRL is considered to have stimulatory effects on prostate growth and
development as its receptors are present in fibromuscular and epithelial cells, as
well as in focal glandular hyperplastic cells, in experimentally induced BPH [10-
12]. PRL and its binding sites have been identified in the canine prostate.
However, PRL differs in its action by down-regulation of 5 -reductase activity,

150
resulting in a decrease in prostatic DHT concentrations in canine prostatic cells
studied in vitro [10]. This suggests a possible role of PRL in atrophy or
decreased size of the prostate by acting on intraprostatic DHT concentrations in
the dog.
The effect of experimentally induced hyperprolactinemia in dogs has been
studied. Shafik et al (1994) used high doses (600ug/kg/week) of ovine PRL on a
population of male beagles to determine whether any contraceptive effects could
be shown [13]. Their treatment resulted in a negative impact on both semen
morphology and motility after two weeks of treatment with degenerative changes
of the seminiferous tubules and complete azoospermia by six months of
treatment [13]. Although treatment of dogs with high doses of a non-canine PRL
can be useful as a tool for contraception, it does not provide information on the
normal reproductive physiology of PRL in the dog nor on possible naturally
occurring PRL related reproductive pathologies.
Semen quality studies in hyperprolactinemic rats and men reported
oligozoospermia, asthenozoospermia and teratozoospermia [4, 5, 14]. In
conditions causing hyperprolactinemia, increased E2 results in low testosterone
concentrations and clinical manifestations of hypogonadism and decreased
semen quality [6]. In the dog, published long-term studies of the effects of
spontaneous or induced hyperprolactinemia with canine PRL on semen quality
do not exist. Short-term hyperprolactinemia using dopamine antagonists did not
alter semen characteristics, while short-term hypoprolactinemia using dopamine
agonists resulted in slight alterations on one sperm motion characteristic (VSL)

151
as assessed by CASA [15]. Clearly, minimal data exists on the role of PRL in
sperm and prostate physiology in dogs.
The purpose of this study was to investigate the associations of PRL with
BPH, prostate volume and semen quality characteristics in stud dogs. We
hypothesized that high serum PRL concentrations would be negatively
associated with BPH, prostate volume and semen quality [16].
2. Materials and Methods
2.1 Case Subjects
A convenience sample of 29 healthy intact male dogs belonging to 24
breeds with varying fertility histories, with or without indication of prostate disease
was obtained between March and December of 2009. Dogs were recruited from
cases presented to the Theriogenology Service at the Ontario Veterinary College
(Guelph, Ontario) and from a small private clinic in Southwestern Ontario with a
large breeder clientele. All clients were required to fill out a consent form
(Appendix 1) and brief questionnaire (Appendix 2) outlining their dogs’ fertility
history, diet and amount of time their dogs spent outdoors. All dogs underwent a
complete male breeding soundness examination, blood and urine collection as
described in the Materials and Methods section in Chapter Two.
2.2 Semen collection, dilution and staining.
Twenty-two of the 29 dogs (75.9%) enrolled in the study had semen
collected. Semen processing was completed according to the Materials and

152
Methods section of Chapter Two.
2.3 Motility Analysis
All semen motility analysis was undertaken according to the protocols set
out in the Materials and Methods section of Chapter Two. Technical settings for
computer assisted semen analysis (CASA) were programmed according to
Schafer-Somi and Aurich (2009) (Table 3.1) [17].
2.4 Prostate Examination
The prostate gland of all dogs was evaluated after semen collection by
several methods, as described in the Materials and Methods section in Chapter
Two.
2.5 Ultrasonography of the Prostate
Ultrasonography of the prostate was accomplished according the Materials
and Methods section in Chapter Two. Table 3.2 describes the criteria by which
BPH was diagnosed according to ultrasound measurements of the prostate.
2.6 Prolactin Assay
Serum samples were batched and shipped overnight on dry ice to the
Endocrinology Laboratory, Washington State University. Prolactin concentrations
were measured using a commercially available validated ELISA for canine
prolactin [16] according to manufacturer instructions (Milenia Canine prolactin

153
(MKVCP-1), Milenia Biotec distributed by Alpco Immunoassays, Salem NH). As
samples were batched and run in the same assay no inter-assay variation was
present. The intra-assay coefficient of variation (CV) was 5.2% with a sensitivity
of 0.4 ng/ml. The lowest PRL concentration standard used in the assay was 2.5
ng/ml.
2.7 Urinalysis
Urinalysis was completed according to the protocols set out in the
Materials and Methods section in Chapter Two.
2.8 Statistical Analyses
2.8.1 Overall Statistical Analysis
Statistical analyses of the data from individual dogs was performed using
SAS version 9.2 software (SAS Institute Inc. Cary, NC, USA). All statistical
analyses of both univariable and bivariable associations were completed using
the general linear model (GLM) procedure as described in the Material and
Methods of Chapter Two. No more than two explanatory variables were used in
any analysis due to the small sample size to avoid over-fitting the data to the
statistical models. Assumptions of normality were tested using the UNIVARIATE
procedure, as described in Chapter Two, and data transformations were used
when necessary. A logit transformation was used in the case of percentage
values and a small bias correction term added in order to accommodate zero
values (Logit outcome variable = log((r+k)/(n-r+k)); r= number of responding

154
cells, n=number of cells counted, k= bias correction term=0.25). In cases where
transformations were not successful in normalizing the data, the Monte Carlo
exact non-parametric test was used to determine if simple univariable correlations
were significant. P-
analyses, and 95% confidence limits were used. The Grubbs’ test for outliers was
used on the residuals[18] and models were rerun with removal of these points as
described in the Materials and Methods in Chapter Two.
2.8.2 Study Population
Descriptive statistics of the characteristics of the study population of dogs
were completed as per the Materials and Methods of Chapter Two.
2.8.3 BPH and Prostate Volume
The Student’s t-test was used to detect whether there was a difference in
mean prolactin concentrations (PRL) between dogs with and without BPH.
Univariable analysis was also completed with age and bodyweight as explanatory
variables with PRL as the response variable. Bivariable analysis was performed
using the explanatory variable PRL and the outcome variable prostate volume,
controlling for benign prostatic hyperplasia analytically by including it in the model.
2.8.4 Sperm Motility
Univariable and bivariable analyses of semen motility as previously
described included the following outcome variables: total motility, progressive

155
motility, ALH, AOC, BCF, DAP, DCL, DSL, VAP, VCL, VSL, LIN, STR, and WOB.
PRL, age, bodyweight and prostatic volume were each tested as explanatory
variables. Bivariable analyses included the explanatory variable combinations of
PRL and age, PRL and prostate volume, and PRL and bodyweight.
2.8.5 Sperm Morphology
Univariable and bivariable analyses of semen morphology included the
following outcome variables: normal sperm, head defects, midpiece defects, tail
defects, loose heads, proximal droplets and distal droplets. PRL, age and
bodyweight were each tested as explanatory variables. Bivariable analyses
included the explanatory variable combinations of PRL and age, PRL and
prostate volume and PRL and bodyweight.
3. Results
3.1 Study Population
The mean serum prolactin concentration of the study population (n=29)
was 4.19 ng/ml 1.16 ng/ml, and concentrations ranged between undetectable to
28.24 ng/ml. The mean serum prolactin concentration of part of the population
from whom semen was collected (n=22) was 5.23 ng/ml ± 1.46 ng/ml. All other
characteristics of the study population of dogs can be found in Table 3.3 and in
Chapter Two (Table 2.4).

156
3.2 BPH and Prostate Volume
The mean serum PRL concentration did not significantly differ between
dogs with BPH and dogs without (t-value=0.87, p=0.39), the details of which are
shown in Table 3.4. No significant univariable associations were observed
between PRL and age (R2=0.04, p=0.31) or bodyweight (R2=0.01, p=0.54).
There was also no significant association between serum PRL and prostate
volume when controlling for BPH in the model (R2=0.04, p=0.31).
3.3 Sperm Motility
No significant associations between PRL and semen motility
characteristics of total motility, progressive motility, ALH, AOC, BCF, DAP, DCL,
DSL, LIN, STR, VAP, VCL, VSL, and WOB were found using univariable simple
linear regression analysis (Table 3.5) or bivariable analyses (Table 3.6).
3.2 Semen Morphology
PRL was significantly associated with proximal droplets in univariable
analyses (Table 3.7, Figure 3.1). Overall there was a decrease in proximal
droplets with increasing PRL concentrations. No other significant univariable
associations between PRL and sperm morphology parameters were found (Table
3.7).
The final bivariable model of midpiece defects was significant with PRL and age
(Table 3.8, Figure 3.2). Overall, there was a decrease in midpiece defects with
increasing PRL concentrations. Percentages of midpiece defects were lowest at

157
4 years of age and highest at 6 years of age with a decrease in midpiece defects
from 8 to 10 years of age. No other bivariable analyses showed significant
associations with serum PRL (Table 3.8). The overall bivariable models with
proximal droplets as a response to PRL were significant, although the regression
slope of the second variable were not (Table 3.9). This result indicated a strong
influence of PRL on the analysis and in these cases the model was further
simplified to its univariable form.
3.3 Outlier Removals
Outliers were identified and removed from the model of normal sperm and
PRL. The results of this can be viewed in Appendix 5. Removal of three outliers
(dogs 6, 1, and 15) revealed associations between normal sperm morphology
and PRL (R2=0.36, p=0.01), normal sperm morphology and PRL with age
(R2=0.66, p=0.0005), and normal sperm morphology and PRL with prostate
volume (R2= 0.52, p=0.006) (see Appendix 5). Univariable analysis, with these
outliers omitted, between normal sperm morphology and PRL showed an
increasing percentage of normal sperm with higher concentrations of PRL within
the range of the study population. Dogs with PRL values less than 2.5 ng/ml had
much lower percentages of normal sperm. In bivariable analyses, there was a
similar pattern of increasing percentage of normal sperm with increasing
prolactin, however, at older ages and higher prostate volumes there was a lower
percentage of normal sperm compared to dogs at younger ages and smaller
prostate volumes. Although, the removal of outliers in large data sets may be

158
advised we maintained the data due to the small sample size and possibility of
removing valuable information from the analysis.
4. Discussion and Conclusions
The physiology and role of prolactin in the stud dog is poorly understood.
The purpose of this observational study was to determine whether PRL is
associated with BPH and prostate volume and associated with sperm motility and
morphology characteristics in intact male dogs of various breeds.
A significant unconditional effect of PRL on proximal droplets was
observed, with a higher percentage of proximal droplets (20%; 95% CI 3.5%-
62%) at serum PRL concentrations below 2.5ng/ml and a small yet significant
decrease in proximal droplet percentages at PRL concentrations at or above 2.5
ng/ml. Proximal droplets, which represent excess cytoplasm in the final
maturation process, move distally from the proximal to distal midpiece region
before being released from the epididymis. Prolactin concentrations were also
significantly associated with midpiece defects of sperm, but this effect was shown
to be dependent on the age of the dog. An increase in percentage of midpiece
defects was seen with PRL concentrations below 2.5 ng/ml throughout all age
groups with significant decreases in those defects at higher PRL concentrations.
Although the general trend in all age groups shows a small yet significant
decrease in midpiece defects as PRL concentrations increase over 2.5ng/ml, it is
unclear why middle-aged dogs (6 years) have the largest percentages of
midpiece defects at PRL concentrations less than 2.5 ng/ml and older dogs (10

159
years) have the lowest. It was initially thought that one Mastiff cross (Dog 16) in
the study with bilateral interstitial cell tumours and a high percentage of midpiece
defects (20%) was influencing the results. However, removal of this dog from the
data set did not alter the results of the analysis. These results relating to
proximal droplets and midpiece defects suggest that PRL may play a role in
epididymal sperm maturation and/or spermatogenesis and that an age
dependent effect on midpiece defects is apparent, although no other evidence
exists to support this hypothesis in the dog.
PRL, and its receptor (PRLR), have been identified in seminiferous
tubules, developing spermatozoa and epididymal tissue in men and rats [19-21]
and in the testicular tissue of rams and pigs [22, 23], although it has yet to be
studied in the dog. PRL has also been shown to increase rat testicular tissue
testosterone levels in vitro by increasing LH receptors in Leydig cells [24]. It is
reasonable to assume then that a similar PRL function and PRLR localization in
the dog exists and may have relevance with the results of our study. This
previous research in humans and other species also supports the hypothesis that
normal PRL levels may be necessary for normal development of sperm both at
the level of spermatogenesis within the seminiferous tubules and maturation of
sperm within the epididymis itself. Alternatively, PRL may act indirectly on sperm
by potentiating or modifying testosterone action during spermatogenesis and
maturation through action on either Sertoli cells and/or epididymal tissue through
the androgen receptors located in these areas. The latter hypothesis may also
explain why reports of hypoprolactinemia on fertility do not exist in men and why

160
fertility rates in male PRL knock-out mice do not show alterations in pregnancy
rates [25]. Although our study detected increased proximal droplets and
midpiece defects at PRL concentrations less than 2.5 ng/ml, the amount of
normal sperm was still in excess of 70% and therefore a perceptible impact on
fertility would be unlikely.
Outlier removals (Appendix 5) showed similar results with respect to
normal sperm and PRL. Again, dogs with PRL values less than 2.5 ng/ml had
substantially lower percentages of normal sperm once these outliers were
removed. This suggests that these PRL values may result in defects of
spermatogenesis as previously discussed. Ultimately, a lack of evidence for
other causes of decreased morphologically normal sperm to support removal of
apparent statistical outliers, as well as a small sample size in this study, were
factors in maintaining them in the analysis.
Serum PRL concentrations in excess of 13 ng/ml in men were associated
with infertility and poor sperm motility and morphology [4]. Serum concentrations
of PRL in excess of 20 ng/ml in men is considered hyperprolactinemia [26].
However, the results of the current study suggest that concentrations as high as
28.24 ng/ml are within the normal reproductive range for dogs as no adverse
effects on semen quality were observed at this concentration. Concentrations
below 2.5ng/ml could be considered hypoprolactinemic as impacts on sperm
quality were observed. Hypoprolactinemia has been associated with metabolic
syndrome in humans [27]. None of our study subjects had a history of diabetes or
presented as obese, however it is possible that subclinical cases may have been

161
present. Thyrotrophin releasing hormone (TRH) is a stimulator of PRL secretion
by lactotroph cells of the pituitary. The absence of clinical signs of
hypogonadism or lethargy, alopecia and skin disorders due to hypothyroidism
suggest that the highest values for PRL obtained in our study were within normal
limits. Prolactinoma, or naturally- or pharmacologically-induced
hyperprolactinemia are rare occurrences in dogs, therefore research about
hyperprolactinemia and its pathology outside of pregnancy and lactation in dogs
is scarce. Overall, the absence of significance in the remaining sperm motility
and morphology parameters may be due to the small sample size in this study.
In order to increase the power and sensitivity of the analysis, a sample size of 88
or greater dogs would be needed. A long-term study to recruit adequate
numbers may resolve this issue. We did not perform a serum biochemical
analysis as part of our evaluation of the study dogs and therefore were not able
to evaluate other conditions including metabolic disease.
Use of the Milenia Canine Prolactin ELISA was chosen as it has recently
been validated for dogs [16] with a published normal reference range
(undetectable to 6.0ng/ml) and is a commercial assay that would be readily
available to practising clinicians. The choice was therefore made with practical
considerations in mind. Although RIA is the gold standard for research purposes,
any clinically significant findings may not be transferable in dealing with clinical
cases of infertility in patients. The high end of the range for PRL differed from the
high end of the range in our study (6.0 ng/ml compared with 28.24 ng/ml).
Although Corrada et al found one individual had higher PRL concentrations

162
compared to the other dogs in that study [16], five dogs from our population (one
each of Samoyed, Labrador Retriever, German Shepherd Dog, Golden Retriever,
and Welsh Cardigan Blue Corgi) had similar PRL concentrations to the excluded
individual. These differences between studies may be due to the differences in
sample size (n=65 compared to n=20) and over-representation of Beagles and
German Shepherd Dogs in the reference range study [16]. Study of a larger
population including many breeds, sizes and ages would clarify whether these
high values are truly normal in the overall intact canine male population. We
could then define hyper- and hypoprolactinemia and establish breed-specific
normal ranges for serum PRL concentrations to aid in the diagnosis and
treatment of reproductive conditions.
No significant associations between PRL and sperm motility
characteristics were observed in our study. This is in contrast to the work of
Koivisto et al, (2009), who found that short-term experimentally induced
hypoprolactinemia with cabergoline was associated with decreased VSL [15].
The discrepancies between studies may be explained by several factors.
Koivisto’s study population consisted of only beagles and did not examine other
breeds. Beagles were shown to have higher serum concentrations of PRL in one
study [16] while in another study Fox Terriers were shown to have significantly
lower PRL concentrations [28], therefore breed effects are relevant. The dogs in
Koivisto’s study were aged 12 months to 2 years compared to our study group of
2 to 11 years. It is possible that our study was able to find age related effects
due to including dogs of broader ages. Koivisto’s study evaluated the effects of

163
experimentally induced hypoprolactinemia using treatment with cabergoline for 3
weeks, while our study represented a one-time observation of semen quality and
PRL levels. A single measurement may not reflect the overall PRL status of our
dogs. Conversely, the cabergoline treatment may not have been long enough to
elicit chronic morphological effects as seen in our study. Methods used to
measure PRL also differed in the two studies, although ranges and means were
comparable with our validated ELISA to RIA studies. In addition, our small
sample size may not have provided enough power to the study to detect changes
in VSL in our study population. Not all dogs examined were able to provide
semen samples for the study. The absence of an estrous bitch during semen
collection and the sexual inexperience of some of the study subjects lead to
failure of adequate sexual stimulation in these cases.
No significant associations between PRL concentrations and BPH or
prostate volume were found in the present study. It is important to note that PRL
release is pulsatile in nature and the lack of multiple blood sampling during the
study may not have given us an accurate representation of overall PRL
concentrations. Further investigation by increasing the number and frequency of
subject sampling and increasing the number of dogs enrolled in a study of this
nature is recommended.
PRL has been shown to decrease in vitro prostatic DHT production in the dog
[10] and may have a role to in prostatic pathology. Whether the mechanism
involved in decreasing prostatic DHT is by direct action of PRL on prostatic
epithelial cells or due to a chronic negative feedback of PRL on testosterone

164
secretion is unknown and cannot be determined by the results obtained here.
The diagnosis of BPH in this study used the generally accepted clinical signs of
enlarged prostatic dimensions by digital palpation per rectum, a predetermined
classification of enlarged prostate with respect to bodyweight, symmetry of the
gland on ultrasound examination and/or the presence of blood in the ejaculate.
BPH is most accurately determined using histology as the gold standard, and
some cases reported here might not have had histological evidence of BPH.
Further research using prostatic biopsy is recommended and may reveal
associations between PRL and BPH. Biopsy and/or fine needle aspirate of the
prostate in our study would have been an unwarranted risk in otherwise healthy
client dogs. We were also unable to control for bodyweight and age
simultaneously in evaluating prostate volume with PRL concentrations.
In rats, PRL has shown delayed long-term feedback on spermatogenesis
and the prostate [29, 30]. Long-term hyperprolactinemia results in decreased
serum testosterone concentrations and teratozoospermia in men [4], so an effect
of PRL on certain aspects of sperm development related to testosterone
concentrations in dogs may be plausible. Analyzing concentrations of other
hormones such as testosterone, estradiol (as a modifier of PRL secretion),
thyroid stimulating hormone (as an index of TRH concentration), and LH/FSH in
our study population, may add to furthering our understanding of PRL in its role
in reproductive physiology. By further investigating and identifying PRL and/or
its receptor in canine testes and sperm cells, one may better understand the
specifics and localization of PRL effects on the dog.

165
In conclusion, low serum PRL concentration was associated with higher
percentages of proximal droplets and midpiece defects; the latter also increased
with age at PRL concentrations less than 2.5 ng/ml. No effects of serum PRL
concentration on BPH, prostate volume or other semen parameters related to
motility and morphology were observed. PRL appears to have minimal
reproductive effects in the dog and may not have equal importance in
reproductive pathologies seen in other species, such as man, however; further
study on a larger scale is required. The information obtained indicates that in
some cases of sub-fertility related to defects of the sperm midpiece and proximal
droplets, maintenance of PRL concentrations above 2.5 ng/ml may be desirable
to improve reproductive potential in individual dogs.

166
Table 3.1 Technical Parameters for SpermVision™ Computer Assisted Semen Analysis used on samples from 29 dogs, March-December 2009, Ontario, Canada
Parameter
Depth of chamber 20um
96-104
2ul
37°C
Variable
20-60 um2
7 or 1000
50-200
60/sec
VCL > 15
VCL >15, STR>0.9
STR >0.9, LIN >5
AOC <7, DSL <3
DSL <6
VCL>118, ALH>6.5, LIN<0.5
LIN<=0.5, STR<=0.9
DAP/Radius >=3, LIN<0.5
2 mins
Light adjustment
Volume per Chamber
Temperature
Sperm concentration
Area of sperm heads
Number of fields or Cell number
Number cells/field
Frame rate
Total motility
Progressively Fast
Linear
Immotile
Local motility
Hyperactive
Non-Linear
Curvilinear
Incubation time
Modified from Schäfer-Somi and Aurich, 2007[17]

167
Table 3.2 Classification scheme for determination of the presence of benign prostatic hyperplasia by ultrasonography in 29 dogs, March-December 2009, Ontario, Canada
Prostatic Measurement
Maximum length, width or height
Bodyweight
> 3.0 cm 20 kg
> 3.5 cm >20 to 40 kg
> 4.0 cm 60 kg
> 4.5 cm 80 kg
Classification based on arbitrary cut-points.

168
Table 3.3 Breed, prolactin concentration, age and fertility data on 22 dogs from which semen was collected in Ontario, Canada, 2009 Dog Breed Prolactin
(ng/ml) Age (years)
No. Litters
No. Breedings /year
No. Years Total
1 Shiloh Shepherd 4.14 5.5 7 1-2 5 2 Labrador Retriever 0.98 5 0 0 0 3 Great Pyrenees 0.92 4.5 2 <1 2 4
Australian Shepherd
2.02
5.5
0 0 0
5 Newfoundland 4.57 4.5 0 0 0 6 Golden Retriever 5.68 5 0 Multiple Unknown 7 Golden Retriever 0 9 0 0 0 8 Whippet 0 11 6 <1 7 9
Cavalier King Charles Spaniel
0.11
9
15 2-3 7
10 Belgian Shepherd 0.31 8.5 0 0 0 11 Labrador Retriever 17.47 8 3 0.5 6 12
Chesapeake Bay Retriever
6.56
6.5
0 0 0
13 Boxer 2.07 6 0 0 0 14 Labrador Retriever 3.96 5.5 6 1-2 4 15 Whippet 3.46 5 0 0 0 16 Mastiff Cross 2.07 7 0 1 0 17 Golden Retriever 11.49 4.5 0 0 0 18 Shiloh Shepherd 1.59 5.5 0 0 0 19 German Shepherd 9.89 4 0 0 0 20
Welsh Cardigan Blue Corgi
9.59
10
1 3 Unknown
21 Akita 0 4 2 1 2 22 Samoyed 28.24 4 0 0 0

169
Table 3.4 Mean serum PRL concentrations and confidence intervals with respect to Benign Prostatic Hyperplasia (BPH) status Prostate Status Mean serum
PRL (ng/l) t-Value
p-value 95% Confidence Interval
Lower Limit Upper Limit
BPH 4.4 0.87 0.39 1.6 7.3
Non-hyperplastic 3.0 2.6 5.5

170
Table 3.5 Univariable analyses using simple linear regression modeling of semen motility parameters and PRL with coefficients of determination (R2) of the overall model, test statistic and associated p-values. Semen Parameter R2-value F-statistic p-value
Total Motility (%) 0.01 0.23 0.64
Progressive Motility (%) 0.03 0.64 0.43
ALH (um) 0.02 0.42 0.52
AOC 0.05 0.91 0.35
BCF (Hz) 0.08 1.70 0.21
DAP (um) 0.01 0.10 0.75
DCL (um) 0.04 0.79 0.38
DSL (um) 0.00 0.00 0.99
VAP (um/s) 0.01 0.11 0.75
VCL (um/s) 0.04 0.77 0.39
VSL (um/s) 0.00 0.00 0.97
STR 0.01 0.28 0.60
LIN 0.05 0.95 0.34
WOB 0.11 2.26 0.15

171
Table 3.6 Three bivariable linear regressions of motility parameters with coefficients of determination of overall model (R2), test statistics, and associated p-values. Response Explanatory
Variables with PRL in the model
R2-value
F-Statistic p-value of Linear Model
Total Motility Age 0.01 0.12 0.89 Bodyweight 0.03 0.29 0.75 Prostate Volume 0.01 0.11 0.90 Progressive Motility Age 0.03 0.33 0.73 Bodyweight 0.05 0.48 0.62 Prostate Volume 0.05 0.33 0.73 ALH Age 0.10 1.00 0.39 Bodyweight 0.02 0.22 0.80 Prostate Volume 0.02 0.21 0.81 AOC Age 0.13 1.36 0.28 Bodyweight 0.05 0.43 0.66 Prostate Volume 0.06 0.57 0.58 BCF Age 0.20 2.30 0.13 Bodyweight 0.08 0.82 0.46 Prostate Volume 0.16 1.66 0.22 DAP Age 0.02 0.17 0.84 Bodyweight 0.02 0.18 0.84 Prostate Volume 0.01 0.09 0.91 DCL Age 0.10 1.02 0.38 Bodyweight 0.07 0.64 0.54 Prostate Volume 0.04 0.40 0.68

172
Table 3.6 (continued) Three bivariable linear regressions of motility parameters with coefficients of determination of overall model (R2), test statistics, and associated p-values. Response Explanatory
Variables with PRL in the model
R2-value
F-Statistic p-value of Linear Model
DSL Age 0.02 0.19 0.84 Bodyweight 0.00 0.04 0.96 Prostate Volume 0.00 0.02 0.98 VAP Age 0.02 0.17 0.85 Bodyweight 0.01 0.13 0.88 Prostate Volume 0.01 0.09 0.92 VCL Age 0.10 0.98 0.39 Bodyweight 0.06 0.57 0.58 Prostate Volume 0.04 0.39 0.68 VSL Age 0.02 0.14 0.87
Bodyweight 0.01 0.05 0.95 Prostate Volume 0.00 0.01 0.99 STR Age 0.25 2.88 0.08 Bodyweight 0.12 1.26 0.31 Prostate Volume 0.02 0.15 0.85 LIN Age 0.27 3.15 0.07 Bodyweight 0.13 1.25 0.31 Prostate Volume 0.05 0.46 0.64 WOB Age 0.25 2.81 0.09 Bodyweight 0.14 1.41 0.27 Prostate Volume 0.11 1.08 0.36

173
Table 3.7 Univariable analyses of sperm morphology parameters and PRL with coefficients of determination (R2) of the overall model, test-statistic and associated p-values. Morphology
Parameter (%)
R2-value F-Statistic P-value
Normal Sperm 0.00 0.00 0.99
Head Defects 0.00 0.01 0.93
Midpiece Defects 0.11 2.32 0.14
Tail Defects 0.07 1.36 0.26
Loose Heads 0.09 1.78 0.20
Proximal Droplets 0.23 5.50 0.03*
Distal Droplets 0.005 0.08 0.78
* Indicates significant p-

174
Table 3.8 Three bivariable linear regressions of morphology parameters, coefficients of determination (R2) of the overall model, test statistics and associated p-values Response (%) Explanatory
Variables with PRL in the model
R2-value F-Statistic p-value
Normal Sperm Age 0.00 0.01 0.99 Bodyweight 0.00 0.00 1.00 Prostate Volume
0.03 0.27 0.77
Head Defects Age 0.01 0.11 0.90 Bodyweight 0.00 0.01 0.99 Prostate Volume
0.03 0.31 0.74
Loose Heads Age 0.17 1.77 0.20 Bodyweight 0.10 0.98 0.40 Prostate Volume
0.12 1.14 0.34
Tail Defects Age 0.12 1.12 0.32 Bodyweight 0.07 0.65 0.54 Prostate Volume
0.10 0.96 0.40
Distal Droplets Age 0.24 2.69 0.10 Bodyweight 0.03 0.25 0.78 Prostate Volume
0.19 1.96 0.17
Midpiece Defects Age 0.42 3.89 0.03* Bodyweight 0.13 1.24 0.31 Prostate Volume 0.12 1.11 0.35
* Indicates significant p-value

175
Table 3.9 Three non-significant bivariable linear regressions of proximal droplets with variable coefficients where PRL slope coefficients were significant, coefficients of determination for the overall model (R2) and associated p-values. Explanatory Variable Model Fit Model of Proximal Droplets
Explanatory Variable
Regression Coefficient
p-value
R2-value for Overall Model
F-statistic
p-value
Model 1 PRL -0.49 0.08 Age 0.28 0.23 0.30 3.61 0.05 Model 2 PRL -0.55 0.05 Bodyweight -0.03 0.31 0.28 3.30 0.06 Model 3 PRL -0.61 0.04 Prostate
Volume -0.09 0.88 0.24 2.61 0.10

176
Fig 3.1 Predicted percentage of proximal droplets in response to serum PRL. Back-transformation of the linear regression model logitProxDrop=-2.94- 0.60lnPRL (R2=0.23, p=0.03). Predicted Proximal Droplets (%), Lower CI, Upper CI. CI= 95% Confidence Interval.

177
Fig 3.2 Predicted percentage of midpiece defects in response to serum PRL. Back-transformation of the linear regression model logitMidpiece= -8.24+1.93A-0.41lnPRL- 0.15A2 (R2=0.42, p=0.03). Predicted Midpiece Defects (%), Lower CI, Upper CI. CI= 95% Confidence Interval, A=Age

178
References
[1] A. Bachelot and N. Binart, "Reproductive role of prolactin," Reproduction,
vol. 133, pp. 361-9, Feb 2007. [2] C. Gobello, et al., "A review of canine pseudocyesis," Reprod Domest
Anim, vol. 36, pp. 283-8, Dec 2001. [3] C. Bole-Feysot, et al., "Prolactin (PRL) and its receptor: actions, signal
transduction pathways and phenotypes observed in PRL receptor knockout mice," Endocr Rev, vol. 19, pp. 225-68, Jun 1998.
[4] G. Merino, et al., "Hyperprolactinemia in men with asthenozoospermia, oligozoospermia, or azoospermia," Arch Androl, vol. 38, pp. 201-6, May-Jun 1997.
[5] A. Colao, et al., "Outcome of cabergoline treatment in men with prolactinoma: effects of a 24-month treatment on prolactin levels, tumor mass, recovery of pituitary function, and semen analysis," J Clin Endocrinol Metab, vol. 89, pp. 1704-11, Apr 2004.
[6] M. K. Gill-Sharma, "Prolactin and male fertility: the long and short feedback regulation," Int J Endocrinol, vol. 2009, p. 687259, 2009.
[7] E. Reiter, et al., "Effects of pituitary hormones on the prostate," Prostate, vol. 38, pp. 159-65, Feb 1 1999.
[8] K. E. Lane, et al., "Suppression of testosterone and estradiol-17beta-induced dysplasia in the dorsolateral prostate of Noble rats by bromocriptine," Carcinogenesis, vol. 18, pp. 1505-10, Aug 1997.
[9] L. Tangbanluekal and C. L. Robinette, "Prolactin mediates estradiol-induced inflammation in the lateral prostate of Wistar rats," Endocrinology, vol. 132, pp. 2407-16, Jun 1993.
[10] S. Chevalier, et al., "Nonsteroidal serum factors involved in the regulation of the proliferation of canine prostatic epithelial cells in culture," Prostate, vol. 5, pp. 503-12, 1984.
[11] M. F. El Etreby and A. T. Mahrous, "Immunocytochemical technique for detection of prolactin (PRL) and growth hormone (GH) in hyperplastic and neoplastic lesions of dog prostate and mammary gland," Histochemistry, vol. 64, pp. 279-86, 1979.
[12] D. Helmerich and J. E. Altwein, "Effect of prolactin and the anit-prolactin bromocriptin on the testosterone uptake and metabolism in androgen-sensitive and insensitive canine organs," Urol Res, vol. 4, pp. 101-5, Nov 10 1976.
[13] A. Shafik, "Prolactin injection, a new contraceptive method: experimental study," Contraception, vol. 50, pp. 191-9, Aug 1994.
[14] M. Laszczynska, et al., "Evaluation of spermatozoa of the rat in hyperprolactinaemia induced by metoclopramide," Andrologia, vol. 24, pp. 101-8, Mar-Apr 1992.
[15] M. B. Koivisto, et al., "Effects of short-term hyper- and hypoprolactinaemia on hormones of the pituitary, gonad and -thyroid axis and on semen

179
quality in male Beagles," Reprod Domest Anim, vol. 44 Suppl 2, pp. 320-5, Jul 2009.
[16] Y. Corrada, et al., "Prolactin reference range and pulsatility in male dogs," Theriogenology, vol. 66, pp. 1599-602, Oct 2006.
[17] S. Schafer-Somi and C. Aurich, "Use of a new computer-assisted sperm analyzer for the assessment of motility and viability of dog spermatozoa and evaluation of four different semen extenders for predilution," Anim Reprod Sci, vol. 102, pp. 1-13, Nov 2007.
[18] F. Grubbs, "Procedures for Detecting Observations in Samples," Technometrics, vol. 11, pp. 1-21, 1969.
[19] A. Ouhtit, et al., "Visualization of gene expression of short and long forms of prolactin receptor in rat reproductive tissues," Biol Reprod, vol. 49, pp. 528-36, Sep 1993.
[20] W. M. Hair, et al., "Prolactin receptor expression in human testis and accessory tissues: localization and function," Mol Hum Reprod, vol. 8, pp. 606-11, Jul 2002.
[21] E. Hondo, et al., "Prolactin receptor expression in rat spermatogenic cells," Biol Reprod, vol. 52, pp. 1284-90, Jun 1995.
[22] H. N. Jabbour and G. A. Lincoln, "Prolactin receptor expression in the testis of the ram: localisation, functional activation and the influence of gonadotrophins," Mol Cell Endocrinol, vol. 148, pp. 151-61, Feb 25 1999.
[23] P. Guillaumot, et al., "Sertoli cells as potential targets of prolactin action in the testis," Mol Cell Endocrinol, vol. 122, pp. 199-206, Sep 18 1996.
[24] K. Purvis, et al., "Prolactin and Leydig cell responsiveness to LH/hCG in the rat," Arch Androl, vol. 3, pp. 219-30, Nov 1979.
[25] R. W. Steger, et al., "Neuroendocrine and reproductive functions in male mice with targeted disruption of the prolactin gene," Endocrinology, vol. 139, pp. 3691-5, Sep 1998.
[26] A. R. Hoffman, et al., "Patient guide to hyperprolactinemia diagnosis and treatment," J Clin Endocrinol Metab, vol. 96, pp. 35A-6A, Feb 2011.
[27] G. Corona, et al., "Hypoprolactinemia: a new clinical syndrome in patients with sexual dysfunction," J Sex Med, vol. 6, pp. 1457-66, May 2009.
[28] C. Urhausen, et al., "Concentrations of prolactin, LH, testosterone, TSH and thyroxine in normospermic dogs of different breeds," Reprod Domest Anim, vol. 44 Suppl 2, pp. 279-82, Jul 2009.
[29] M. De Rosa, et al., "The treatment with cabergoline for 24 month normalizes the quality of seminal fluid in hyperprolactinaemic males," Clin Endocrinol (Oxf), vol. 64, pp. 307-13, Mar 2006.
[30] M. De Rosa, et al., "Six months of treatment with cabergoline restores sexual potency in hyperprolactinemic males: an open longitudinal study monitoring nocturnal penile tumescence," J Clin Endocrinol Metab, vol. 89, pp. 621-5, Feb 2004.

180
CHAPTER FOUR
SUMMARY AND CONCLUSIONS
This thesis investigated possible associations between serum
concentrations of Vitamin D and prolactin (PRL) and reproductive characteristics
related to prostate volume, sperm morphology and sperm motility in male dogs.
The first part of this research determined whether an association existed
between serum 25OHVD and BPH, as well as between 25OHVD and prostate
volume controlling analytically for BPH. Possible associations between serum
25OHVD and a) semen motility as evaluated by CASA and b) microscopically
determined sperm morphology were also studied. The second part of the
research investigated the same reproductive traits for associations with serum
PRL concentrations.
In both the Vitamin D and PRL studies, we were not able to show any
associations between Vitamin D or PRL with BPH, nor were there associations
between Vitamin D or PRL and prostate volume controlling for BPH analytically.
The low power of the study due to the small sample size may not have permitted
detection of small differences. The diagnosis of BPH was determined based on
transrectal palpation per digitum, ultrasound dimensions of the prostate, and/or
blood present in the third fraction of the ejaculate. However, a lack of histological
diagnosis of BPH as a gold standard may have led to a misdiagnosis of BPH,
thus introducing a misclassification bias. One must also consider that prostate
physiology may differ from that in men, although experimental Vitamin D

181
analogues decreased prostate size in a small number of beagle dogs in a
preliminary drug trial[1]. Increasing the number of dogs studied would allow for
more accurate findings and possibly result in a different outcome with regards to
BPH and prostate health.
Vitamin D was shown to have associations with several measures of
semen motility and morphology. We determined that serum 25OHVD
concentrations for optimal semen quality characteristics ranged from 120-180
nmol/l using a validated and commercially available ELISA for dogs. A negative
effect on semen quality could be observed below or above these concentrations,
although this effect was lessened at concentrations at the high range of normal
and was associated with smaller decreases in semen quality. In the human and
rat, Vitamin D and its receptor have been identified in sperm cells in all stages of
development and testicular tissue, chiefly in Sertoli cells [2-4]. It is possible that
Vitamin D is necessary for normal and healthy development of sperm in all
species, including dogs, and is translated into improvements in normal
morphology and consequently, normal and improved motility at optimal
concentrations.
Prostate volume is correlated with age and both age and prostate volume
were factors that influenced Vitamin D in its semen quality associations. It is
possible that Vitamin D exerts a toxic effect at older ages or that older animals
are unable to use Vitamin D in the same way as younger animals. One rat study
showed a decrease in intestinal Vitamin D action in older compared with younger
rats and may suggest impaired Vitamin D metabolism in older animals [5].

182
Perhaps this is also true for the ageing dog with respect to Vitamin D and semen
quality. Isolation and identification of Vitamin D and its receptor in the
reproductive tissues and sperm cells of the dog at various ages may shed some
more light on the roles and mechanisms of Vitamin D, and may provide
explanations for the age related effects.
The second half of this thesis looked at serum PRL concentrations and
semen quality characteristics. Using the same validated ELISA our serum PRL
concentrations ranges were greater than previously published (non-detectable to
28.24 ng/ml compared with non-detectable to 6.0 ng/ml [6]). Prolactin
concentrations were associated with sperm midpiece defects as well as proximal
droplets, an observation not previously published. Fewer midpiece defects were
found when serum PRL concentrations were greater than 2.5 ng/ml. This new
information suggests that PRL concentrations less than 2.5 ng/ml may be defined
as hypoprolactinemia due to the increase in midpiece defects and proximal
droplets at these levels. Evidence of hyperprolactinemia was not observed in
this study.
It is important to note that PRL does undergo periodic fluctuation in
physiological concentrations and that sampling in this study was undertaken at
different times of the day and during different times of the year. The lack of
associations with the majority of semen parameters may be a result of small
sample size or due to the restriction of having only one PRL measurement per
subject. Taking multiple measurements and controlling for the time of year of

183
sampling may impact serum PRL concentrations, and consequently, semen
quality.
Prolactin has been associated with semen pathologies in cases of chronic
hyperprolactinemia in men through long-term negative feedback loops that cause
decreases in circulating testosterone concentrations. Commercial validated tests
for PRL in the dog have only recently become available to the veterinary
community. Although, hyperprolactinemia has not been previously diagnosed in
dogs it is still possible this condition may occur and have an impact on fertility in
the male dog. Further research is needed to determine whether situations of
PRL excess occur naturally in the dog and what role that might play in male dog
infertility.
In conclusion, this study has revealed new associations with respect to
semen quality characteristics and Vitamin D and PRL in the stud dog. This is an
exciting avenue of research that promises to have important clinical ramifications
in the treatment of canine fertility issues related to semen morphology and
motility characteristics. We hope that these preliminary studies help to further
supplement the arsenal of tools available to optimize reproductive health in the
dog.

184
References: [1] L. Adorini, et al., "Vitamin D receptor agonists target static, dynamic, and
inflammatory components of benign prostatic hyperplasia," Ann N Y Acad Sci, vol. 1193, pp. 146-52, Apr 2010.
[2] S. Aquila, et al., "Human sperm anatomy: ultrastructural localization of 1alpha,25-dihydroxyvitamin D receptor and its possible role in the human male gamete," J Anat, vol. 213, pp. 555-64, Nov 2008.
[3] J. Merke, et al., "Nuclear testicular 1,25-dihydroxyvitamin D3 receptors in Sertoli cells and seminiferous tubules of adult rodents," Biochem Biophys Res Commun, vol. 127, pp. 303-9, Feb 28 1985.
[4] M. Blomberg Jensen, et al., "Vitamin D receptor and vitamin D metabolizing enzymes are expressed in the human male reproductive tract," Hum Reprod, vol. 25, pp. 1303-11, May 2010.
[5] G. Balogh, et al., "Influence of age on 1,25(OH)2-vitamin D3 activation of protein kinase C in rat duodenum," Mol Cell Endocrinol, vol. 129, pp. 127-33, May 16 1997.
[6] Y. Corrada, et al., "Prolactin reference range and pulsatility in male dogs," Theriogenology, vol. 66, pp. 1599-602, Oct 2006.

185
APPENDICES

186
Appendix 1. Investigation of serum vitamin D and prolactin levels in benign prostatic hyperplasia and semen quality in intact male dogs. Client Consent Form. “Investigation of serum vitamin D and prolactin levels in benign
prostatic hyperplasia and semen quality in intact male dogs”
CLIENT CONSENT FORM
Benign Prostatic Hyperplasia (BPH) is a non-cancerous enlargement of the prostate, associated with poor semen quality, and affects approximately 80% of intact male dogs. Recent research into prostate disease in men has shown that vitamin D is associated with a decrease in the occurrence of this condition and better overall prostate health. Prolactin, a reproductive hormone most commonly associated with females during pregnancy and lactation, also has a role in prostate health. In increase in serum prolactin levels in men has been shown to negatively affect fertility and potentiate hyperplasia of the prostate. Interestingly, the prostate gland is similar in both human and canine species. Our objectives are to determine: 1) whether an association exists between BPH and circulating vitamin D and prolactin levels and 2) to provide information to the canine breeding industry regarding the possible effects of vitamin D and prolactin on canine male fertility and prostate health. We request that you complete a brief questionnaire to obtain background information on breeding history and management. Semen collection will be undertaken, after which, both a physical and ultrasound examination will be performed. A blood and urine sample will also be taken for research purposes. The procedures being used in this study are considered neither painful nor invasive. The costs for the procedures and testing will be assumed by the project. All results will be kept in strictest confidence. We appreciate your time and consideration. Dr. Cathy Gartley, Dr. Adria Kukk I agree to participate in the study. Patient Name__________________ Patient ID________ Signature of Owner/Authorized Agent_____________ Date_____________ Signature of Clinician ____________________ Date_____________

187
Appendix 2. Investigation of serum vitamin D and prolactin levels in benign prostatic hyperplasia and semen quality in intact male dogs. Client Questionnaire. Questionnaire
The following questions address necessary background information for our study on the role of vitamin D and prolactin on benign prostatic hyperplasia and fertility in intact male dogs. Please answer all of the questions to the best of your ability. 1. Client name: _____________________________________________________ 2. Dog’s name, breed, and age: ________________________________________
Part 1: Management
3. What sort of diet is your dog on? Please state the brand name if using a commercial diet and
any regular treats or ‘goodies’ they receive daily. Please include the approximate amounts.
i) _____________________________________________________
ii) _____________________________________________________
iii) _____________________________________________________
4. How is your dog housed? For example, does he live in a kennel situation with outdoor runs,
does he live indoors with the family, or a combination of both?
____________________________________________________________________________
____________________________________________________________________________
5. How many hours a day, approximately, does your dog spend out-‐doors? For example leash
walks, in the backyard, in outdoor runs etc.
In the summer: _________________
In the winter: _________________
Page 1 of 2

188
6. Is your dog on any supplements and, if so, what are they?
i) ___________________________________________
ii) ___________________________________________
iii) ___________________________________________
Part 2: Medical history
7. History of past illness (Type/year/duration)
i) _______________________________________
ii) _______________________________________
iii) _______________________________________
8. Is your dog on any medication(s) and, if so, what are they?
i)_________________________________________
ii)_________________________________________
Part 3: Breeding History
9. How many litters has you dog sired?
Per year: _________________
Per career: _________________
10. Average size of litter? _________________
11. Approximately how many breedings/year? Please include fresh chilled/frozen semen
inseminations if applicable. ___________________________
12. How many years has he been at stud in total? ________________
Page 2 of 2

189
Appendix 3 Minitube Chill 5 extender Protocol (Excerpt). Protocol for CaniPRO Chill 5 Culture Medium for Canine Semen (13574/0105) Extender preparation: Warm CaniPRO Chill 5 to room temperature. Semen extension:
Use only the sperm-rich fraction of the ejaculate (2nd fraction) for insemination. Do not include the first fraction of the ejaculate (clear prostatic fraction before sperm rich fraction) or the third fraction (prostatic fraction post-sperm rich fraction) of the ejaculate in the sample because the quality of the semen may decrease.
After collection evaluate semen quality (Concentration, Motility, Morphology). Dilute one part of semen in three to five parts of extenders. (ie: 1ml in 3-5ml of
CaniPRO Chill 5) base on initial semen concentration. Semen can be preserved for up to 5 days at 4°C (39°F) with preservation of
a minimum of 70% initial motility. Avoid changes in temperature during preservation by placing the tube with the extended semen in small water-bath.
At time of insemination warm semen to room temperature and perform a motility evaluation (See insemination instructions below).
For purpose of shipping chilled semen
Store diluted semen at 4°C (39°F) for at least 2 hours before packaging and shipping.
Perform a motility evaluation before shipping and retain a control aliquot for future evaluation.
If needed to reach ideal insemination volume (See insemination instructions below) add some extender in extra 15ml tube in Canine Transport Box.
Canine Semen Concentration (106) Per ML
Ratio of Semen to Extender
250 – 750
1 to 3
750 – 1.25
1 to 4
Above 1.25
1 to 5

190
Appendix 4 SFT Guidelines (Excerpt) GUIDELINES FOR CANINE BREEDING SOUNDNESS EXAMINATION Original authors: Original reviewers: B.J. Purswell, Virginia Tech P.N.S. Olson, University of Minnesota G.C. Althouse, Iowa State University S.D. Johnston, University of Minnesota M.V. Root, University of Minnesota L.E. Evans, Iowa State University Current reviewer: M.V. Root Kustritz, University of Minnesota Other changes: G.C. Althouse is now at University of Pennsylvania, P.N.S. Olson is with the Morris Animal Foundation, and S.D. Johnston is at Western College of Health Sciences. Morphology Morphology most often is assessed by staining the semen sample and observing the cells under 1000X magnification (oil immersion). Morphology may be assessed without staining using a phase contrast microscope after fixing the sample with formol-buffered saline at a dilution of 1:9. Different sample preparations cause artifacts that will be seen as morphologic abnormalities; percentage morphologically normal spermatozoa should be fairly consistent regardless of method [1]. The two most common stains used are a rapid Wright’s Giemsa stain (DiffQuik, Baxter Healthcare, Miami FL) and eosin/nigrosin stain (SFT stain, Lane Manufacturing, Denver CO). To stain with rapid Wright’s Giemsa stain, place one drop of semen on a slide and smear out as for a blood smear. Allow to air dry. Immerse the slide into each of the three solutions in the same order as used for any type of cytology, allowing the slide to sit in each solution for five minutes. Rinse the slide completely and let it air dry before evaluation under oil immersion. To stain with eosin/nigrosin, place one drop of semen and a similar-sized drop of stain on one end of a glass slide. Gently mix the two together with a pusher slide, and draw out as a thin film similar to a blood smear. Allow to air dry as quickly as possible; placing the slide on a hot plate (37oC) and blowing on the slide will hasten the drying process and help prevent staining artifacts [2]. Spermatozoa stained with rapid Wright’s Giemsa stain appear purple on a clear background. Normal spermatozoa stained with eosin/nigrosin stain appear white against a black or violet background. Eosin/nigrosin stain is taken up by spermatozoa that have abnormal plasma membranes and so appear pink against a dark background. It might be assumed that such spermatozoa are non-viable

191
but no significant correlation between “live-dead” ratio and fertility has been described in dogs. Under oil immersion, examine and count at least 100 spermatozoa. The number of normal spermatozoa in 100 is the percentage morphologically normal spermatozoa. Total normal for the sample is calculated by multiplying total number of spermatozoa in the ejaculate by percentage morphologically normal. Commonly accepted values are greater than 80% morphologically normal spermatozoa and greater than 200 million total normal spermatozoa in an ejaculate. Abnormal spermatozoa may be classified as having primary defects (those that occur during spermatogenesis, including defects in head shape, bent midpiece, persistent proximal cytoplasmic droplet, and doubling of any portion of the spermatozoon) or secondary defects (those that occur during epididymal maturation or slide preparation, including detached heads, persistent distal cytoplasmic droplets, and bent tails). Correlation between specific defects and fertility in dogs is poorly defined. References [1] Root Kustritz MV, Johnston SD, Olson PN and Root TK: The effect of stains and investigators on assessment of morphology of canine spermatozoa. J Amer Anim Hosp Assoc 1998;34:348-352. [2] Shaffer HE, Almquist JO. Vital staining of bovine spermatozoa with a eosin-aniline blue staining mixture. J Dairy Sci 1948;3:677-678.

192
Appendix 5. Outlier Removal Statistics
Predicted sperm motility in response to normal serum 25OHVD range at various transformed prostate volumes with removal of dog 6. Back-transformation of multiple regression model logitMotility=-11.7 +2.73Vol +0.11VD-0.02Vol*VD (R2=0.56, p=0.003). CI= 95% Confidence Interval. Vol=Prostate Volume. Predicted Motility (%), Lower CI,
Upper CI

193
Table. Outlier removal of dog 6. Explanatory variable with vitamin D (VD) in the model and corresponding coefficient of determination (R2) and p-value. Resulting analysis revealed no significant associations between serum 25OHVD and progressive motility.
Response Explanatory Variable with Vitamin D in the model
R2-value p-value of Linear Model
Progressive Motility
VD2
0.18
0.21

194
Outlier removal of dog 6, 1 and 15. Predicted normal sperm and serum PRL. Back transformation of linear regression model logitNormal=0.26 + 0.004lnPRL (R2=0.36, p=0.01). Predicted Normal Sperm (%), Lower CI,
Upper CI. CI= 95% confidence interval.

195
Predicted normal sperm in response to serum PRL range at various transformed prostate volumes with removal of dogs 6, 1 and 15. Back-transformation of multiple regression model logitNormal=3.32 -0.52Vol + 0.32lnPRL (R2=0.52, p=0.006). CI= 95% Confidence Interval. Vol=Transformed Prostate Volume. Predicted Normal Sperm (%), Lower CI,
Upper CI.

196
Predicted normal sperm in response to normal serum PRL range at various ages with removal of dogs 6, 1 and 15. Back-transformation of multiple regression model logitNormal=2.66 + 0.25lnPRL - 0.25Age (R2=0.66, p=0.0005). CI=Confidence Interval. PRL=prolactin. Predicted Normal Sperm (%), Lower CI, Upper CI




















