
BAHA Usage in Auditory Rehabilitation of Patients with External Auditory Canal Stenosis
Upload
independentCategory
view
2download
0
IFO
XAa
tFb
2
AmmpsitLecpbirtrqrtwmsMvindftI
Kr
Elrer
*EAros
Neuroscience 154 (2008) 390–396
0d
NFLUENCES OF UN-MODULATED ACOUSTIC INPUTS ONUNCTIONAL MATURATION AND CRITICAL-PERIOD PLASTICITY
F THE PRIMARY AUDITORY CORTEXpoS2adtraaOmdsMwaitCdtc
Atnn
T
RsutoVfIodwtadmlitp
. ZHOU,a,b N. NAGARAJAN,a B. J. MOSSOPa
ND M. M. MERZENICHa*
W. M. Keck Center for Integrative Neuroscience, Coleman Labora-ory, and Department of Otolaryngology, University of California, Sanrancisco, CA 94143, USA
College of Life Sciences, East China Normal University, Shanghai00062, China
bstract—Sensory experiences contribute to the develop-ent and specialization of signal processing capacities in theammalian auditory system during a “critical period” ofostnatal development. Earlier studies have shown that pas-ive exposure to tonal stimuli during this postnatal epochnduces a large-scale expansion of the representations ofhose stimuli within the primary auditory cortex (A1) [ZhangI, Bao S, Merzenich MM (2001) Persistent and specific influ-nces of early acoustic environments on primary auditoryortex. Nat Neurosci 4:1123–1130]. Here, we show that ex-osing rat pups through the normal critical period epoch andeyond to continuous, un-modulated, moderate-level tones
nduces no such representational distortion, and in fact dis-upts the normal development of frequency response selec-ivity and tonotopicity all across area A1. The area of cortexesponding selectively to continuously exposed sound fre-uencies was actually reduced, when compared with ratseared in normal environments. Strong exposure-driven plas-icity characteristic of the critical period could be inducedell beyond the normal end of the critical period, by simplyodulating the tonal stimulus. Thus, continuous tone expo-
ure, like continuous noise exposure [Chang EF, MerzenichM (2003) Environmental noise retards auditory cortical de-elopment. Science 300:498 –502], ineffectively induces crit-cal period plasticity, and indefinitely blocks the closure of aormally-brief critical period window. These findings againemonstrate the crucial role of temporally structured inputsor inducing the progressive cortical maturational changeshat result in the closure of the critical period window. © 2008BRO. Published by Elsevier Ltd. All rights reserved.
ey words: cortical map, critical period, development, envi-onment, frequency receptive field, plasticity.
arlier studies have documented an early, several-day-ong postnatal epoch—a “critical period”— during which theat primary auditory cortex (A1) can be plastically remod-led on a large scale by merely exposing the pup toepeated sounds. Beyond this period, passive sound ex-
Corresponding author.-mail address: [email protected] (M. M. Merzenich).bbreviations: A1, primary auditory cortex; BDNF, brain-derived neu-
otrophic factor; BW20, bandwidth measured at 20 dB above thresh-
7ld; CF, characteristic frequency; P, postnatal day; pps, pulses perecond.
306-4522/08$32.00�0.00 © 2008 IBRO. Published by Elsevier Ltd. All rights reseroi:10.1016/j.neuroscience.2008.01.026
390
osure has less influence on the progressive developmentf frequency tuning and tonotopicity in A1 (de Villers-idani et al., 2007; Nakahara et al., 2004; Zhang et al.,001, 2002; Zhou and Merzenich, 2007). Studies havelso indicated that exposure-based cortical remodelinguring the critical period strongly depends on the modula-ion of sounds in the environment. For example, exposingat pups to pulsed tones of a specific frequency results inn over-representation of that frequency in A1 (Zhang etl., 2001; de Villers-Sidani et al., 2007; Han et al., 2007).n the other hand, if rat pups are exposed to temporallyodulated white noise, the tonotopic organization of A1 isegraded while, paradoxically, there is a premature clo-ure of the critical period (Zhang et al., 2002; Zhou anderzenich, 2007). If rat pups are exposed to continuoushite noise, the spectral and temporal response selectivitynd cortical inhibition are sustained in a degraded and
mmature state, and the critical period window is main-ained open, indefinitely (Chang and Merzenich, 2003;hang et al., 2005). Here, we extend those studies, byocumenting the consequences of rearing pups throughhis critical period epoch and beyond in the presence ofontinuous, moderate-level tonal stimuli.
EXPERIMENTAL PROCEDURES
ll experiment procedures used in this study were approved byhe Animal Care and Use Committees at the University of Califor-ia, San Francisco. Every effort was made to minimize both theumber of animals used and their suffering.
one exposure
at pups (Sprague–Dawley) and their mothers were placed inound-shielded test chambers from postnatal day (P) 7 to P35nder continuous pure tone stimuli (either 3.5- or 7-kHz). Note thathis exposure extends for about 22 days beyond the postnatal timef normal closure of the “critical period” in A1 in the rat (deillers-Sidani et al., 2007). The sound signal was generated by a
unction generator (BK Precision Dynascan Corporation, Chicago,L, USA) and amplified to a calibrated level in the free sound fieldf �65-dB SPL. No significant harmonic signals or other soundistortions were recorded via wide sampling in the chamber inhich the tonal stimulus was delivered. Animals were give access
o water and food ad libitum under an 8-h/16-h light/dark cycle. Nobnormalities in the behavior of either the mother or pups could beetected during sound exposure. The weights of all pups andothers were continuously monitored, and there was no weight
oss compared with naïve rats, indicating normal lactation. Activ-ties during waking and the sleep behaviors of these rats indicatedhat the sound stimuli were not stressful. After the end of theeriod of continuous-tone exposure at P35, the tonal stimulus (a
-kHz tone) in a subset of rats was continuously modulatedved.

(liPtetr
E
EdaiTsdcneuawd
shfmcoaADltposfc
C
FtatodFtwl2fsh
arctCoic
tlwrrmwrrtTt
Er
R7qdrmIoqcs(rewseoNCeth
itwttsrtcTigb(Pfap
X. Zhou et al. / Neuroscience 154 (2008) 390–396 391
50-ms duration tone pips with 5-ms ramps at �65-dB SPL, de-ivered at five pulses per second (pps), followed by a 1-sec silentnterval to minimize adaptation effects) for 2 weeks (i.e. from36–P49). In addition, one group of naïve rats was also exposed
o a pulsed 7-kHz tone, from P36–P49. These secondarily-sound-xposed animals were then returned to standard housing condi-ions for a subsequent 2-week period (i.e. P50–P63) before neuralesponse recording in A1.
lectrophysiological recording
lectrophysiological recording of cortical responses was con-ucted at �P35 and �P63 for matched groups of exposed ratsnd naïve control rats. Rats were initially anesthetized with an i.p.
njection of sodium pentobarbital (50 mg/kg body weight).hroughout the surgical procedures and during the recordingession, a state of areflexia was maintained with supplementaloses of 8 mg/ml dilute pentobarbital injected i.p. The trachea wasannulated to ensure adequate ventilation, and the cisterna mag-um was drained of cerebrospinal fluid to minimize cerebraldema. The skull was secured in a head holder leaving the earsnobstructed. After reflecting the right temporalis muscle, theuditory cortex was exposed and the dura resected. The cortexas maintained under a thin layer of viscous silicone oil to preventesiccation.
Cortical responses were recorded with parylene-coated tung-ten microelectrodes (1–2 mohm at 1 kHz; FHC Inc., Bowdoin-am, ME, USA). Recording sites were chosen to evenly samplerom the auditory cortex while avoiding blood vessels, and werearked on a magnified digital image of the cortical surface vas-
ulature. At each recording site the microelectrode was loweredrthogonally into the cortex to a depth of 480–550 �m (layers 4nd 5), where vigorous stimulus-driven responses were recorded.coustic stimuli were generated using TDT System III (Tucker-avis Technologies, Inc., Alachua, FL, USA) and delivered to the
eft ear through a calibrated earphone (STAX 54) with a soundube positioned inside the external auditory meatus. A softwareackage (SigCal, SigGen, and Brainware; Tucker-Davis Technol-gies, Inc.) was used to calibrate the earphone, generate acoustictimuli, monitor cortical response properties online, and store dataor offline analysis. The evoked spikes of a neuron or a smallluster of neurons was collected at each site.
ortical mapping and data analysis
requency tuning curves were reconstructed by presenting pureones of 50 frequencies (1–30 kHz, 25-ms duration, 5-ms ramps)t eight sound intensities (0- to 70-dB SPL in 10-dB increments) tohe contralateral ear in a random, interleaved sequence at a ratef two pps. The characteristic frequency (CF) of a cortical site wasefined as the frequency at the tip of the V-shaped tuning curve.or flat-peaked tuning curves, CF was defined as the midpoint of
he plateau at threshold. For tuning curves with multiple peaks, CFas defined as the frequency at the most sensitive tip (i.e. with
owest threshold). Response bandwidths (bandwidth measured at0 dB above threshold, BW20s) of tuning curves were measuredor all sites. The response latency was defined as the time fromtimulus onset to the earliest response, using peri-stimulus timeistograms of responses to all tone pips.
The area of A1 was defined as previously described (Zhang etl., 2002). To generate A1 maps, Voronoi tessellation (a Matlaboutine; The MathWorks, Inc., Natick, MA, USA) was performed toreate tessellated polygons, with electrode penetration sites atheir centers. Each polygon was assigned the characteristics (i.e.F) of the corresponding penetration site. In this way, every pointn the surface of the auditory cortex was linked to the character-
stics experimentally derived from a sampled cortical site that was
losest to this point. dWe used a previously defined index to quantitatively describehe precision of tonotopicity in A1 (Zhang et al., 2001, 2002). Theine connecting the two most anterior and posterior penetrationsithin A1 was used as a reference for the tonotopic axis. We then
otated each map to orient the tonotopic axis horizontally. Afterotation, new x coordinates of penetrations in each rat were nor-alized to be within a range from 0.0–1.0, and penetration sitesere plotted according to their CFs and x coordinates. The loga-
ithmic frequency range (1–30 kHz) was converted to a linearange (0–1). We defined the index as the average minimal dis-ance from each data point to the line connecting (0,0) and (1,1).he larger the index, the more disordered the tonotopicity of the
onotopic map.
RESULTS
ffects of continuous tone exposure on frequencyepresentations in A1
at pups were reared in continuous pure tones (3.5- or-kHz; n�3 for each group) from P7–P35. A1 was subse-uently electrophysiologically mapped in fine detail imme-iately after sound exposure cessation. Naïve rats (n�4)eared under standard housing conditions were alsoapped at the same ages. As shown in Fig. 1A (left; Group
II), A1 in naïve rats consisted of iso-frequency bandsriented approximately orthogonal to a rostro-caudal fre-uency-representation gradient. Cortical neuron tuningurves recorded in most sites (93%) within A1 were V-haped, with a clear CF defined at the low threshold peakFig. 1Ae, f). In exposed rats, by contrast, tuning curvesecorded from a large number of sites (49% for 3.5-kHzxposed rats and 46% for 7-kHz exposed rats) within A1ere flat-peaked or multipeaked (Fig. 1Aa–d). Compari-on of BW20 showed that tuning curves recorded inxposed rats were more broadly tuned than were thosef naïve rats (Fig. 1Ba; ANOVA with post hoc Student-ewman-Keuls test, all P�0.05–0.001), except for lowestF category centered at 1.75-kHz (ANOVA, P�0.1). How-ver, there was no significant difference in BW20s be-ween rats exposed to 3.5- and 7-kHz (ANOVA with postoc Student-Newman-Keuls test, all P�0.05).
At first glance, the A1 area tuned to higher frequenciesn exposed rats appeared to be larger, while the area tunedo lower frequencies appeared smaller, when comparedith naïve rats (Fig. 1A, left; Group I or II vs. III). To verify
hat observation, we compared the percentage of A1 areahat was tuned to different frequencies in each group. Ashown in Fig. 1Bb, the percentage of A1 area in exposedats was larger than in naïve rats at CF categories cen-ered at 14- and 28-kHz, but was smaller at CF categoriesentered at 1.75-, 3.5-, and 7-kHz (�2�21.7, P�0.006).his result was further confirmed by quantitative compar-
sons of distribution for all CFs obtained from differentroups (Fig. 1Bc). Significant rightward shift of CF distri-ution for exposed rats compared with naïve ratsKruskal-Wallis with post hoc Bonferroni’s test, both�0.001) indicates decreased areas of representation
or lower frequencies, but increased representationalreas of higher frequencies, induced by continuousure-tone exposure. Again, there were no significant
ifferences in the percentages of A1 areas tuned to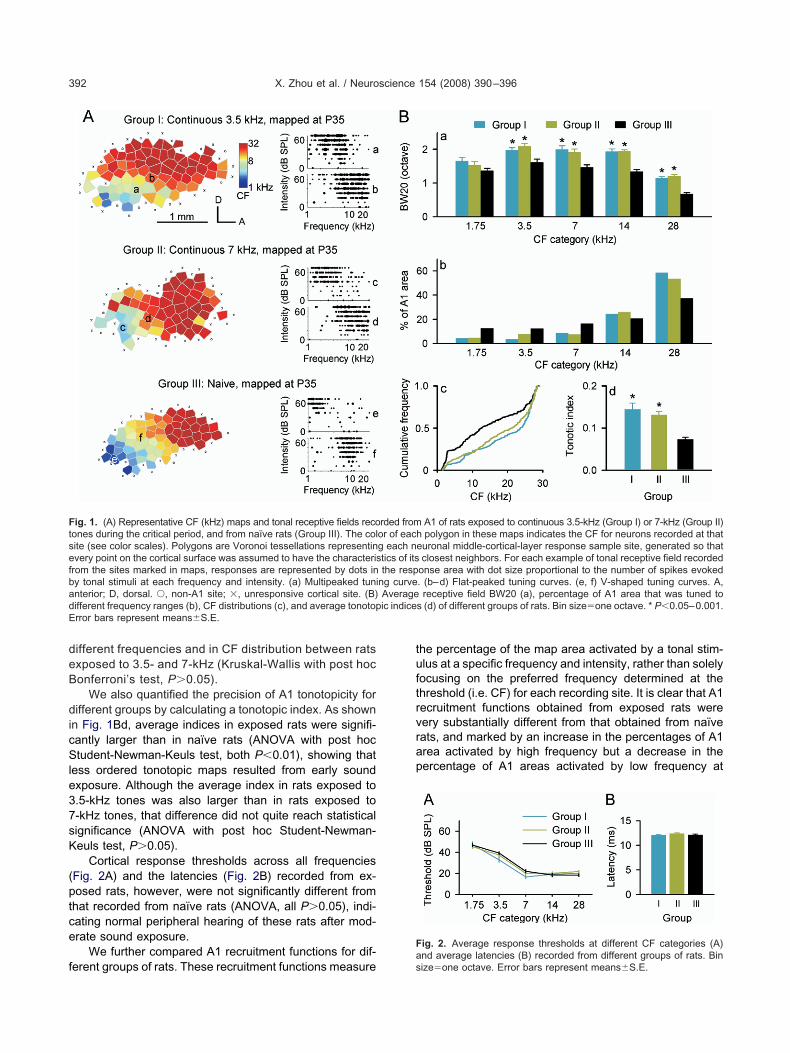
deB
dicSle37sK
(ptce
f
tuftrvrap
Ftsefbad ic indiceE
F
X. Zhou et al. / Neuroscience 154 (2008) 390–396392
ifferent frequencies and in CF distribution between ratsxposed to 3.5- and 7-kHz (Kruskal-Wallis with post hoconferroni’s test, P�0.05).
We also quantified the precision of A1 tonotopicity forifferent groups by calculating a tonotopic index. As shown
n Fig. 1Bd, average indices in exposed rats were signifi-antly larger than in naïve rats (ANOVA with post hoctudent-Newman-Keuls test, both P�0.01), showing that
ess ordered tonotopic maps resulted from early soundxposure. Although the average index in rats exposed to.5-kHz tones was also larger than in rats exposed to-kHz tones, that difference did not quite reach statisticalignificance (ANOVA with post hoc Student-Newman-euls test, P�0.05).
Cortical response thresholds across all frequenciesFig. 2A) and the latencies (Fig. 2B) recorded from ex-osed rats, however, were not significantly different fromhat recorded from naïve rats (ANOVA, all P�0.05), indi-ating normal peripheral hearing of these rats after mod-rate sound exposure.
We further compared A1 recruitment functions for dif-
ig. 1. (A) Representative CF (kHz) maps and tonal receptive fields recoones during the critical period, and from naïve rats (Group III). The colite (see color scales). Polygons are Voronoi tessellations representinvery point on the cortical surface was assumed to have the characterisrom the sites marked in maps, responses are represented by dots iny tonal stimuli at each frequency and intensity. (a) Multipeaked tuninterior; D, dorsal. Œ, non-A1 site; �, unresponsive cortical site. (B)ifferent frequency ranges (b), CF distributions (c), and average tonotoprror bars represent means�S.E.
erent groups of rats. These recruitment functions measureas
he percentage of the map area activated by a tonal stim-lus at a specific frequency and intensity, rather than solelyocusing on the preferred frequency determined at thehreshold (i.e. CF) for each recording site. It is clear that A1ecruitment functions obtained from exposed rats wereery substantially different from that obtained from naïveats, and marked by an increase in the percentages of A1rea activated by high frequency but a decrease in theercentage of A1 areas activated by low frequency at
A1 of rats exposed to continuous 3.5-kHz (Group I) or 7-kHz (Group II)h polygon in these maps indicates the CF for neurons recorded at thateuronal middle-cortical-layer response sample site, generated so thatclosest neighbors. For each example of tonal receptive field recorded
onse area with dot size proportional to the number of spikes evoked. (b–d) Flat-peaked tuning curves. (e, f) V-shaped tuning curves. A,receptive field BW20 (a), percentage of A1 area that was tuned to
s (d) of different groups of rats. Bin size�one octave. * P�0.05–0.001.
ig. 2. Average response thresholds at different CF categories (A)
rded fromor of eacg each ntics of itsthe resp
ng curveAverage
nd average latencies (B) recorded from different groups of rats. Binize�one octave. Error bars represent means�S.E.
iImtqAblp
Df
Aoc2eopamg3erntopV
twvm
rwatssbeaVG4
egfmepGpAfarAanu
rBn
Ft r a [1/4]-( .
X. Zhou et al. / Neuroscience 154 (2008) 390–396 393
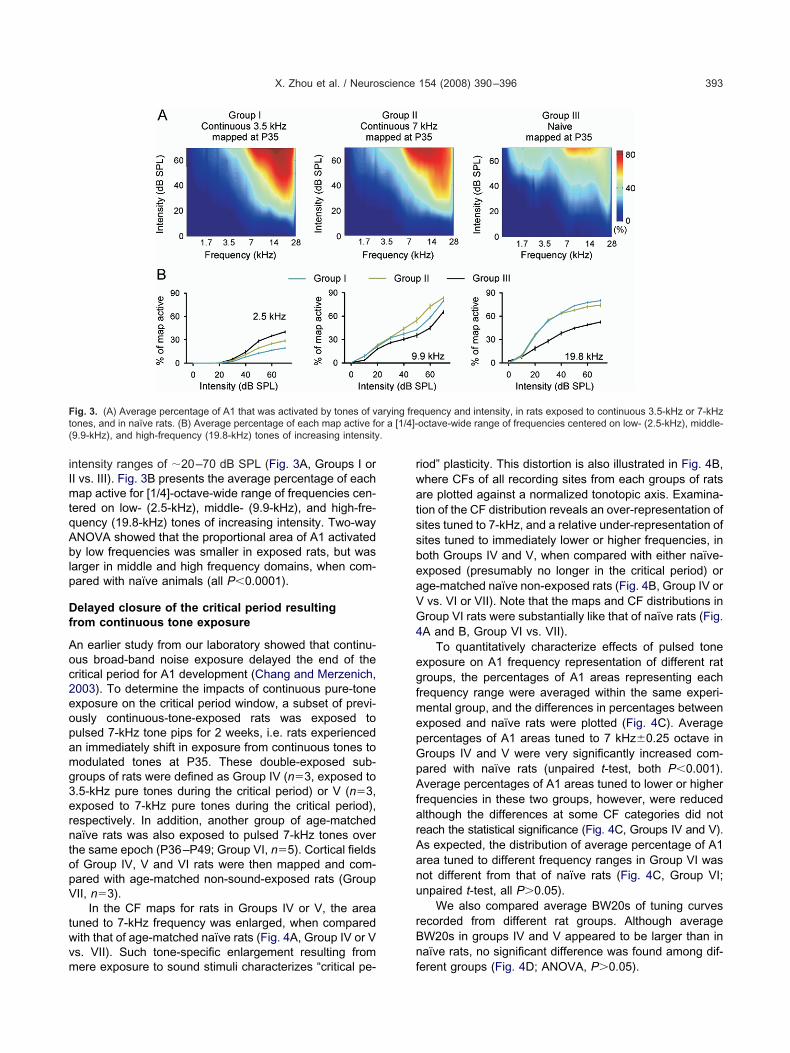
ntensity ranges of �20–70 dB SPL (Fig. 3A, Groups I orI vs. III). Fig. 3B presents the average percentage of eachap active for [1/4]-octave-wide range of frequencies cen-
ered on low- (2.5-kHz), middle- (9.9-kHz), and high-fre-uency (19.8-kHz) tones of increasing intensity. Two-wayNOVA showed that the proportional area of A1 activatedy low frequencies was smaller in exposed rats, but was
arger in middle and high frequency domains, when com-ared with naïve animals (all P�0.0001).
elayed closure of the critical period resultingrom continuous tone exposure
n earlier study from our laboratory showed that continu-us broad-band noise exposure delayed the end of theritical period for A1 development (Chang and Merzenich,003). To determine the impacts of continuous pure-tonexposure on the critical period window, a subset of previ-usly continuous-tone-exposed rats was exposed toulsed 7-kHz tone pips for 2 weeks, i.e. rats experiencedn immediately shift in exposure from continuous tones toodulated tones at P35. These double-exposed sub-roups of rats were defined as Group IV (n�3, exposed to.5-kHz pure tones during the critical period) or V (n�3,xposed to 7-kHz pure tones during the critical period),espectively. In addition, another group of age-matchedaïve rats was also exposed to pulsed 7-kHz tones overhe same epoch (P36–P49; Group VI, n�5). Cortical fieldsf Group IV, V and VI rats were then mapped and com-ared with age-matched non-sound-exposed rats (GroupII, n�3).
In the CF maps for rats in Groups IV or V, the areauned to 7-kHz frequency was enlarged, when comparedith that of age-matched naïve rats (Fig. 4A, Group IV or Vs. VII). Such tone-specific enlargement resulting from
ig. 3. (A) Average percentage of A1 that was activated by tones of vones, and in naïve rats. (B) Average percentage of each map active fo9.9-kHz), and high-frequency (19.8-kHz) tones of increasing intensity
ere exposure to sound stimuli characterizes “critical pe- f
iod” plasticity. This distortion is also illustrated in Fig. 4B,here CFs of all recording sites from each groups of ratsre plotted against a normalized tonotopic axis. Examina-ion of the CF distribution reveals an over-representation ofites tuned to 7-kHz, and a relative under-representation ofites tuned to immediately lower or higher frequencies, inoth Groups IV and V, when compared with either naïve-xposed (presumably no longer in the critical period) orge-matched naïve non-exposed rats (Fig. 4B, Group IV orvs. VI or VII). Note that the maps and CF distributions inroup VI rats were substantially like that of naïve rats (Fig.A and B, Group VI vs. VII).
To quantitatively characterize effects of pulsed tonexposure on A1 frequency representation of different ratroups, the percentages of A1 areas representing eachrequency range were averaged within the same experi-ental group, and the differences in percentages betweenxposed and naïve rats were plotted (Fig. 4C). Averageercentages of A1 areas tuned to 7 kHz�0.25 octave inroups IV and V were very significantly increased com-ared with naïve rats (unpaired t-test, both P�0.001).verage percentages of A1 areas tuned to lower or higher
requencies in these two groups, however, were reducedlthough the differences at some CF categories did noteach the statistical significance (Fig. 4C, Groups IV and V).s expected, the distribution of average percentage of A1rea tuned to different frequency ranges in Group VI wasot different from that of naïve rats (Fig. 4C, Group VI;npaired t-test, all P�0.05).
We also compared average BW20s of tuning curvesecorded from different rat groups. Although averageW20s in groups IV and V appeared to be larger than inaïve rats, no significant difference was found among dif-
quency and intensity, in rats exposed to continuous 3.5-kHz or 7-kHzoctave-wide range of frequencies centered on low- (2.5-kHz), middle-
arying fre
erent groups (Fig. 4D; ANOVA, P�0.05).

Ifisirp2rctmfiaaeArqfsttw
atcte
sm1sqcspd2aMoaoadn
FPP(ti rea of repV ent group
X. Zhou et al. / Neuroscience 154 (2008) 390–396394
DISCUSSION
n this study, we determined the consequences, for theunctional maturation of cortical field A1, of rearing rat pupsn the presence of continuous, moderate-level pure-tonetimuli. Continuous tone rearing had markedly differentmpacts on the response selectivity and the frequencyepresentations of A1 than did rearing in the presence ofulsed tones (de Villers-Sidani et al., 2007; Han et al.,007; Zhang et al., 2001). i) Frequency receptive fieldsecorded in rats exposed to continuous tones were typi-ally flat-peaked or multipeaked, and were far less selec-ive for sound frequency when compared with naïve ani-als. In pulsed tone-exposed rats, however, receptive
elds were substantially like that of naïve rats. A CF waslmost always easily determinable, and they were almostlways continuous and single-peaked. ii) Continuous tonexposure disrupted the normal progressive refinement of1 tonotopic maps, and it distorted map topographies,
esulting in larger-than-normal areas tuned to higher-fre-uencies and smaller-than-normal areas tuned to lower-requencies. There was no clear relationship between ob-erved changes and the frequency of the continuous toneshat these rat pups were exposed to. By contrast, A1onotopic reorganization in pulsed tone exposure cases
ig. 4. (A) Representative CF maps obtained from Groups IV, V, VI7–P35, then pulsed 7-kHz tones for a subsequent 2-week-long period35, to pulsed 7-kHz tones. Group VI naïve rats were exposed to pulse
B) Distributions of CFs plotted against a normalized tonotopic axis ino 7-kHz in both Groups IV and V (arrows), but not in Groups VI and Vn all four rat groups. Note the large-scale growth in the proportional aII rats. * P�0.001. (D) Average BW20 for all recording sites in differ
as highly frequency-dependent, and always resulted in t
n over-representation of exposed frequency and a reduc-ion in the representation of immediately adjacent frequen-ies. iii) Continuous tone exposure delayed the closure ofhe critical period window for A1 development. Pulsed tonexposure accelerated A1 maturation.
It has long been argued that the temporal patterns ofensory inputs are critically important for cortical develop-ent (Buonomano and Merzenich, 1998; Katz and Shatz,996; Rauschecker, 1999; Weliky and Katz, 1997). Earliertudies have shown that the postnatal sharpening of fre-uency receptive fields and the formation of an orderly,ontinuous frequency representation in A1 require expo-ure to normally variable auditory inputs during the criticaleriod (Chang and Merzenich, 2003; Chang et al., 2005;e Villers-Sidani et al., 2007; Keuroghlian and Knudsen,007; Nakahara et al., 2004; Reale et al., 1987; Stantonnd Harrison, 1996; Zhang et al., 2001, 2002; Zhou anderzenich, 2007). Selective strengthening and eliminationf neuronal connections through activity-dependent syn-ptic plasticity contribute significantly to the establishmentf feature-selective cortical response properties (Zhang etl., 2002). We speculate that inappropriate activity-depen-ent modifications on cortical excitatory and inhibitory con-ections under continuous tone exposure might contribute
rats. Group IV rats were exposed to continuous 3.5-kHz tones from49). Group V, rats were exposed to continuous 7-kHz tones, then afterones from P36 to P49. Group VII were non-sound-exposed naïve rats.groups of rats. Note that there was increased A1 area that was tunedferences in percentages of the A1 area tuned to different frequenciesresentation of 7-kHz-tuned neurons in Group IV or V vs. Group VI ors of rats. Error bars represent means�S.E.
and VII(P36–P
d 7-kHz tdifferentII. (C) Dif
o the degraded response selectivity and disrupted tono-

ttehi2Cmda1ctpaes
lirjN(tcasaeinohl
mpgfsais1repr1eczeAstdi
pAmmca
dt(wcTsttaicpup
AtC
B
C
C
C
C
C
d
E
F
H
H
I
X. Zhou et al. / Neuroscience 154 (2008) 390–396 395
opic organization observed in this study, as well as tohose reported in early studies (Kenet et al., 2007; Zhangt al., 2002; Zhou and Merzenich, 2007). On the otherand, experience-dependent changes can also be induced
n the sub-cortical central auditory system (Chiu et al.,003; Lorke et al., 2003; Poon and Chen, 1992; Sanes andonstantine-Paton, 1983, 1985; Yu et al., 2007). In theouse inferior colliculus, for example, click stimuli duringevelopment was shown to block the normal sharpening ofuditory receptive fields (Sanes and Constantine-Paton,983, 1985). Early sound exposure also resulted in aluster of neurons tuned to the exposed tone frequency inhe rat inferior colliculus (Poon and Chen, 1992). It isrobable that observed post-exposure changes in A1 arelso partly the result of feed-forward responses reflectingxperience-dependent changes in various sub-corticaltructures.
It should be noted that earlier studies have shown thatoud tone (�120-dB SPL) exposure, which typically resultsn a peripheral hearing loss, also induces a large-scaleeorganization of frequency map in the auditory cortex ofuvenile and adult cats (Eggermont and Komiya, 2000;orena et al., 2003). Exposure to a 120-dB SPL pure tone
5- or 6-kHz), for example, induced a shift in cortical CFoward lower frequency and a broadening of the tuningurve, due to changes in the balance between excitationnd inhibition at both cortical and subcortical levels afteround exposure (Norena et al., 2003). In this study, wepplied moderate-level tone stimuli (�65-dB SPL) and nolevation in cortical response threshold has been recorded
n exposed rats compared with naïve animals, indicatingormal peripheral function after sound exposure. We alsobserved larger-than-normal areas which were tuned toigher frequencies in A1 of exposed rats, in addition to
ess-selective tuning curves.The duration of the critical period varies by sensory
odality and in different animal species. While criticaleriod extends for �2 weeks in the rat visual cortex (Fa-iolini et al., 1994), it only lasts a couple of days (P11–P13)or rat A1 when rats are reared with their mother andiblings in a normal cage environment (de Villers-Sidani etl., 2007). The relatively brief normal critical period window
s comparable in length of the critical period for rat somato-ensory cortex (S1 barrel-field) (Rice and Van der Loos,977; Schlaggar and O’Leary, 1993). In addition, the du-ation of the critical period is substantially influenced byarly sensory experiences. Early dark-rearing, for exam-le, results in an indefinite prolongation of the critical pe-iod window for mammalian visual cortex (cats, Cynader,983; Mower, 1991; mice, Iwai et al., 2003; rats, Fagiolinit al., 1994). Continuous noise rearing also retards theritical period development in the rat A1 (Chang and Mer-enich, 2003). We show here that early continuous tonexposure also delays the closure of the critical period for1 development. Thus when these exposed rats wereubsequently exposed to pulsed tone stimuli even beyondhe normal cortical period window, substantial exposure-riven plasticity was still recorded in the A1. One surprise
n these studies was the impacts that tone exposure ap-
eared to have on the progressive functional maturation of1. We had anticipated that continuous tone exposureight block A1 maturation, but only with those regionsost directly engaged by the pure-tone stimulus. To the
ontrary, on these functional grounds, virtually all of A1ppears to have been affected.
It is possible that the observed experience-inducedelay closure of the critical period window may be relatedo the release of the brain-derived neurotrophic factorBDNF) (Chang and Merzenich, 2003; Chang et al., 2005),hich is the primary agent that enables the final corticalhanges that result in critical period closure (Maffei, 2002).he results are also consistent with the finding thattrengthening of locally correlated cortical activity results inhe release of BDNFs that promotes the postnatal matura-ion of the visual cortex (Castren et al., 1992; Hanover etl., 1999). Together, our findings indicate that patterned
nputs play a crucial role for driving changes across theritical period that account for its contributions to earlyostnatal specialization of cortical processing, and for theltimate closure of this powerful, early epoch of corticallasticity.
cknowledgments—This work was supported by National Insti-utes of Health grant NS-10414, the Sandler Fund, and theoleman Fund.
REFERENCES
uonomano DV, Merzenich MM (1998) Cortical plasticity: from syn-apses to maps. Annu Rev Neurosci 21:149–186.
astren E, Zafra F, Thoenen H, Lindholm D (1992) Light regulatesexpression of brain-derived neurotrophic factor mRNA in rat visualcortex. Proc Natl Acad Sci U S A 89:9444–9448.
hang EF, Merzenich MM (2003) Environmental noise retards audi-tory cortical development. Science 300:498–502.
hang EF, Bao S, Imaizumi K, Schreiner CE, Merzenich MM (2005)Development of spectral and temporal response selectivity in theauditory cortex. Proc Natl Acad Sci U S A 102:16460–16465.
hiu TW, Poon PW, Chan WY, Yew DT (2003) Long-term changes ofresponse in the inferior colliculus of senescence accelerated miceafter early sound exposure. J Neurol Sci 216:143–151.
ynader M (1983) Prolonged sensitivity to monocular deprivation indark-reared cats: effects of age and visual exposure. Brain Res284:155–164.
e Villers-Sidani E, Chang EF, Bao S, Merzenich MM (2007) Criticalperiod window for spectral tuning defined in the primary auditorycortex (A1) in the rat. J Neurosci 27:180–189.
ggermont JJ, Komiya H (2000) Moderate noise trauma in juvenilecats results in profound cortical topographic map changes in adult-hood. Hear Res 142:89–101.
agiolini M, Pizzorusso T, Berardi N, Domenici L, Maffei L (1994)Functional postnatal development of the rat primary visual cortexand the role of visual experience: dark rearing and monoculardeprivation. Vision Res 34:709–720.
an YK, Kover H, Insanally MN, Semerdjian JH, Bao S (2007) Earlyexperience impairs perceptual discrimination. Nat Neurosci 10:1191–1197.
anover JL, Huang ZJ, Tonegawa S, Stryker MP (1999) Brain-derivedneurotrophic factor overexpression induces precocious critical pe-riod in mouse visual cortex. J Neurosci 19:RC40.
wai Y, Fagiolini M, Obata K, Hensch TK (2003) Rapid critical periodinduction by tonic inhibition in visual cortex. J Neurosci 23:6695–
6702.
K
K
K
L
M
M
N
N
P
R
R
R
S
S
S
S
W
Y
Z
Z
Z
X. Zhou et al. / Neuroscience 154 (2008) 390–396396
atz LC, Shatz CJ (1996) Synaptic activity and the construction ofcortical circuits. Science 274:1133–1138.
enet T, Froemke RC, Schreiner CE, Pessah IN, Merzenich MM(2007) Perinatal exposure to a noncoplanar polychlorinated biphe-nyl alters tonotopy, receptive fields, and plasticity in rat primaryauditory cortex. Proc Natl Acad Sci U S A 104:7646–7651.
euroghlian AS, Knudsen EI (2007) Adaptive auditory plasticity indeveloping and adult animals. Prog Neurobiol 82:109–121.
orke DE, Wong LY, Lai HW, Poon PW, Zhang A, Chan WY, Yew DT(2003) Early postnatal sound exposure induces lasting neuronalchanges in the inferior colliculus of senescence accelerated mice(SAMP8): a morphometric study on GABAergic neurons andNMDA expression. Cell Mol Neurobiol 23:143–164.
affei L (2002) Plasticity in the visual system: role of neurotrophinsand electrical activity. Arch Ital Biol 140:341–346.
ower GD (1991) The effect of dark rearing on the time course of thecritical period in cat visual cortex. Brain Res Dev Brain Res58:151–158.
akahara H, Zhang LI, Merzenich MM (2004) Specialization of primaryauditory cortex processing by sound exposure in the “critical pe-riod.” Proc Natl Acad Sci U S A 101:7170–7174.
orena AJ, Tomita M, Eggermont JJ (2003) Neural changes in catauditory cortex after a transient pure-tone trauma. J Neurophysiol90:2387–2401.
oon PW, Chen X (1992) Postnatal exposure to tones alters the tuningcharacteristics of inferior collicular neurons in the rat. Brain Res585:391–394.
auschecker JP (1999) Auditory cortical plasticity: a comparison withother sensory systems. Trends Neurosci 22:74–80.
eale RA, Brugge JF, Chan JC (1987) Maps of auditory cortex in catsreared after unilateral cochlear ablation in the neonatal period.
Brain Res 431:281–290.ice FL, Van der Loos H (1977) Development of the barrels and barrelfield in the somatosensory cortex of the mouse. J Comp Neurol171:545–560.
anes DH, Constantine-Paton M (1983) Altered activity patternsduring development reduce neural tuning. Science 221:1183–1185.
anes DH, Constantine-Paton M (1985) The sharpening of frequencytuning curves requires patterned activity during development in themouse, Mus musculus. J Neurosci 5:1152–1166.
chlaggar BL, O’Leary DD (1993) Patterning of the barrel field insomatosensory cortex with implications for the specification ofneocortical areas. Perspect Dev Neurobiol 1:81–91.
tanton SG, Harrison RV (1996) Abnormal cochleotopic organizationin the auditory cortex of cats reared in a frequency augmentedenvironment. Auditory Neurosci 7:97–107.
eliky M, Katz LC (1997) Disruption of orientation tuning in visualcortex by artificially correlated neuronal activity. Nature 386:680–685.
u X, Sanes DH, Aristizabal O, Wadghiri YZ, Turnbull DH (2007)Large-scale reorganization of the tonotopic map in mouse auditorymidbrain revealed by MRI. Proc Natl Acad Sci U S A 104:12193–12198.
hang LI, Bao S, Merzenich MM (2001) Persistent and specific influ-ences of early acoustic environments on primary auditory cortex.Nat Neurosci 4:1123–1130.
hang LI, Bao S, Merzenich MM (2002) Disruption of primary auditorycortex by synchronous auditory inputs during a critical period. ProcNatl Acad Sci U S A 99:2309–2314.
hou X, Merzenich MM (2007) Intensive training in adults refines A1representations degraded in an early postnatal critical period. Proc
Natl Acad Sci U S A 104:15935–15940.(Accepted 17 January 2008)(Available online 25 January 2008)
Copyright © 2022 FDOKUMEN




















