
Transcriptome Analysis of Agmatine and Putrescine Catabolism in Pseudomonas aeruginosa PAO1
Upload
independentCategory
view
3download
0
Influence of Pseudomonas aeruginosa as PGPR on oxidative stresstolerance in wheat under Zn stress
Faisal Islam a, Tahira Yasmeen a,n, Qasim Ali b, Shafaqat Ali a, Muhammad Saleem Arif a,Sabir Hussain a, Hina Rizvi a
a Department of Environmental Sciences, Government College University, Faisalabad 38000, Pakistanb Department of Botany, Government College University, Faisalabad 38000, Pakistan
a r t i c l e i n f o
Article history:Received 17 December 2013Received in revised form6 March 2014Accepted 11 March 2014
Keywords:PGPRPseudomonas aeruginosaZincAnti-oxidant enzymesOxidative stressWheat
a b s t r a c t
Plant Growth Promoting Rhizobacteria (PGPR), whose role is still underestimated, plays an important (orperhaps essential) role in improving plant growth. The comprehensive understanding of bacterial plantgrowth promoting mechanism helps to get sustainable agriculture production under biotic and abioticstresses. In the present study, plant growth promoting (PGP) bacterial strain Pseudomonas aeruginosahaving maximum inhibitory concentration of 1500 mg kg�1 against Zn was isolated from arable land,irrigated with industrial effluent and evaluated to determine it bioremediation potential. The study wasmainly focused on plant biomass production, nutrient uptake and oxidative stress tolerance in relation tothe activities of antioxidative enzymes and the content of non-enzymatic antioxidants. The oxidativestress tolerance was measured by estimating the MDA accumulation as well as H2O2 production in wheatplants under Zn (1000 mg kg�1) stress and inoculation of soil with Zn resistant Pseudomonas aeruginosa.Zn in rooting medium reduced the plant growth, leaf photosynthetic pigments as well as uptake of N andP. However, content of MDA and H2O2 increased at higher concentration of Zn. Inoculation ofP. aeruginosa improved the uptake of P and N in wheat plants with an increase in leaf chlorophyll,total soluble protein and plant biomass production. Analysis of plant root and shoot disclosed that Znconcentration was significantly lowered in P. aeruginosa inoculated zinc stressed plants as compare tothe plants grown under Zn stress only. The amelioration of adverse effects of Zn stress on biomassproduction due to P. aeruginosa inoculation was related with enhanced antioxidative enzyme activities(SOD, POD and CAT), and the contents of non-enzymatic components such as ascorbic acid and totalphenolics (TPC) as compare to Zn-treated plants. The up-gradation in antioxidative defense mechanism,resulted a reduction in H2O2 and MDA content due to the scavenging of ROS by antioxidants. It wasconcluded that P. aeruginosa is an ideal candidate for bioremediation and wheat growth promotionagainst Zn-induced oxidative stress by improving the availability of necessary nutrient, elicitingantioxidant defense system as well as by lowering the Zn metal uptake.
& 2014 Elsevier Inc. All rights reserved.
1. Introduction
Release of heavy metals by industrial operations such asmining, smelting, combustion of fossil fuels and the sewage sludgecaused a significant increase of metals due to their persistent andnon-degradable nature, leading to their biomagnification in food
chain (Rajkumar et al., 2009). Of different heavy metals, Zn is amajor pollutant in the environment because of its extensive use inindustry without its recovery (Taniguchi et al., 2000). Zn is anessential trace element for all life forms on earth; however, itselevated concentration is also toxic for biota (He et al., 2010). Itsaccumulation in soil also adversely affects the microbial commu-nity. In excess concentration it is absorbed by the plants whicheventually disturbs metabolism, growth, development and cropproduction (John et al., 2008). Although Zn is not redox activeelement, generation of reactive oxygen species (ROS), whichresults in oxidative stress, is one important aspect of its toxicitythat ultimately evokes the plant defense system. Despite theoxidative stress, production of ethylene, nutritional imbalance,chlorosis and stunted growth are also the important features of Zn
Contents lists available at ScienceDirect
journal homepage: www.elsevier.com/locate/ecoenv
Ecotoxicology and Environmental Safety
http://dx.doi.org/10.1016/j.ecoenv.2014.03.0080147-6513/& 2014 Elsevier Inc. All rights reserved.
Abbreviations: AMF, arbuscular mycorrhiza; AsA, ascorbic acid; CAT, catalase;CFU, colony forming units; G-POD, guaiacol peroxidase; GR, glutathione reductase;HM, heavy metals; MDA, malondialdehyde; MIC, minimum inhibitory concentra-tion; N, nitrogen; P. aeruginosa, Pseudomonas aeruginosa; P, phosphorous; PGP,plant growth promoting; PGPR, plant growth promoting rhizobacteria; SOD,superoxide dismutase; TPC, total phenolic contents; Zn, zinc
n Corresponding author. Fax: þ92 41 9200671.E-mail address: [email protected] (T. Yasmeen).
Ecotoxicology and Environmental Safety 104 (2014) 285–293

toxicity. Under these conditions, plant growth promoting bacteria(PGPR) play a significant role in alleviating the metal toxicity andexert ameliorating effects on growth of plant and mineral uptake.Among mechanisms used by PGPRs to play their role includelowering the level of ethylene in plants due to increased activity of1-aminocyclopropane�1-carboxylate (ACC) deaminase, with analternate increase in indole acetic acid that allows the metalstressed plant to expend its root and establish themselves betterat early growth phase, by the production of siderophores. Thesesiderophores help the plant to get Fe under metal stressedcondition to optimize metabolism. PGPRs also result in theproduction of organic acids that help to solubilize the phosphorsin the rhizosphere. Although many free living soil bacteria havethe potential to promote plant growth, mobilization and immobi-lization of metal in rhizosphere however, only limited attemptshave been made to study the effect of free living soil bacteria asinoculant for plant growth-promotion, in the amelioration of toxiceffects of heavy metals on plants in relation to plant growth,photosynthetic pigments and alterations in the activities of anti-oxidant enzymes and the concentration of non-enzymatic com-pounds. The main objective of the present study was the isolationand characterization (ACC, IAA, Siderophore production andPhosphate solubilization) of highly Zn resistant bacteria, metalsbiosorption potential of isolated bacteria and evaluation of theireffects on the wheat growth, metal accumulation as well asantioxidant enzyme activities and antioxidants. It was hypothe-sized that inoculation of Zn resistant bacteria with PGP traits canconfer the Zn tolerance in wheat plants, in correlation with theinduction of antioxidant enzymes (CAT, SOD, G-POD) and non-enzymatic antioxidative compounds with change in the uptake ofmineral nutrient.
2. Materials and methods
2.1. Isolation and screening of zinc resistant bacteria
For isolation and screening of Zn resistant bacterial isolates 56 soil sampleswere collected from agricultural field irrigated with industrial effluents alongPaharang drain, Faisalabad, Pakistan. The bacterial isolation was carried out sameday by the pore plate method (McLellan et al., 2009). Briefly, 100 μL of 106 timesserially diluted soil solution (1 g soilþ9 mL saline buffer i.e. 85 percent NaCl) wasspread on Luria Bertani (LB) media plates and incubated for 24 h at 30 1C. Coloniesof different morphological appearances were selected and transferred to LB mediaplates for purification through the streak plate method (Harley and Prescott, 2002).
2.2. Determination of minimum inhibitory concentration (MIC)
MIC of all the bacterial isolates was determined by gradual increase of Znconcentration in LB media until the bacterial isolate failed to grow on plates overseven days of incubation following the method described by Aleem et al. (2003).This concentration was considered as MIC of respective isolates.
2.3. Metal biosorption test
Biosorption of Zn (II) was evaluated by allowing the bacterial isolates to interactwith Zn in LB broth. One mL of overnight grown bacterial culture (108 CFU mL�1)was inoculated into 250 mL of round bottom flasks containing 50 mL of sterilizedLB broth amended with 50, 100 and 250 mg kg�1 of metal salt (ZnSO4). Flaskshaving medium were incubated at 30 1C for 24 h. After incubation bacterial culturewas centrifuged at 12,000g for 10 min. Residual amount of metal in supernatantwas measured through atomic absorption spectrophotometer.
2.4. Evaluation for PGPR characters
For indole acetic acid (IAA) production, Zn resistant bacterial isolates weregrown in LB broth with the addition of 0.5 mg L�1 tryptohan. One mL of overnightgrown culture (108 CFU mL�1) was added in 20 mL of LB broth and incubated at150g for 96 h. Salkowski's reagent was prepared by dissolving 4.5 g of FeCl3 in10.8 mol of H2SO4 L�1. After incubation, the broth was centrifuged at 6000g for
10 min. The pellet was discarded and supernatant was collected. One mL ofsupernatant and 2 mL of Salkowski's reagent (Gordon and Weber, 1951) weremixed in sterilized glass tube at room temperature (25 1C) and left for 25 min. Theabsorbance of developed pink color was read at 530 nm (Halo DB-20 UV–visDouble beam spectrophotometer) (Sheng et al., 2008). The IAA concentration in theculture was determined using a calibration curve of pure IAA as a standardfollowing linear regression analysis.
The siderophore production of isolated bacteria was determined by the chromeazurol-S (CAS) method (Schwyn and Neilands, 1987). Siderophore production wasmeasured by orange halo zone production around the bacterial isolates on blueagar. The production of α-ketobutyrate (α-KB) by the enzymatic cleavage of ACCwas determined by measuring its absorbance in bacterial culture at 540 nm andcomparing with the absorbance of known concentration of pure α-KB (Belimov etal., 2005). Protein concentration in bacterial culture was estimated by the methodof Bradford (1976) and then the enzymatic activity was expressed as 1 M α-KB mg�1 h�1 (Jalili et al., 2009; He et al., 2012).
For the quantitative measurement of P solubilization, the freshly preparedbacterial culture (108 CFU mL�1) was inoculated in Pikovskaya's broth containing2.5 g of Tricalcium Phosphate (TCP). The culture was incubated at 30 1C with aconstant shaking of 200g for seven days. The culture supernatant was obtained bycentrifugation at 6000g and used to determine P-solubilization through thecalorimetric method of Fiske and Subbarow (1925). Color reagent (750 mL),prepared by mixing 500 mL of 1.5 percent (w/v) ammonium molybdate in a5.5 percent (v/v) sulfuric acid solution and 125 μL of a 2.7 percent (w/v) ferroussulfate solution, was added to the sample solution (750 mL) and the production ofphosphomolybdate was measured spectrophotometrically at 700 nm (Halo DB-20UV–vis Double beam spectrophotometer).
2.5. Bacterial identification
Highly resistant bacterial isolate ZN3 (Pseudomonas aeruginosa) having maxi-mum plant growth promoting abilities was selected from a pool of bacterial isolatesand identified through 16S rRNA gene sequencing. Crude DNA of the isolates wasextracted following the method as described by Cheneby et al. (2004). The PCRreaction was performed in a total volume of 25 μL containing 2.5 μL buffer (10 X),0.5 μL of dNTPs (12.5 mM), 1 μL of each primers 27 F (50-AGA GTT TGA TCC TGG CTCAG-30) and 1492 R (50-GGT TAC CTT GTT ACG ACT T-30) of 10 pmoles μL�1 each,0.1 μL of Taq DNA polymerase, 1 μL MgSO4 (50 mM), 17.23 μL of water and 1 μL oftemplate. Amplification was performed in a PCR machine programmed as initialdenaturation at 94 1C for 4.5 min followed by 30 cycles consisting of denaturationat 94 1C for 30 s, primer annealing at 57 1C for 30 s and primer extension at 68 1Cfor 90 s and final extension at 68 1C for 10 min. Amplified PCR products of 16Sribosomal gene were separated on 1 percent agarose gel in 1� TBE (Trisborate-EDTA) buffer. A 100 bp DNA ladder (Promega) was run as a size marker. The gel wasviewed under UV light and photographed using the Eagle Eye gel documentationsystem (Model 3). Amplified PCR product of 16S rRNA gene of selected isolate ZN3(Pseudomonas aeruginosa) was purified using QIA quick PCR purification kit(QIAGEN) following the standard protocol recommended by the manufacturer, toprepare for sequencing. 16S rRNA gene sequence of bacterial isolate ZN3 wascompared with the known nucleotide sequences in the GenBank database usingBlastN (http://www.ncbi.nlm.nih.gov/BLAST). ZN3 showed 99 percent resemblancewith Pseudomonas aeruginosa. So, on the basis of resemblance with specific groupof bacteria we conclude isolate ZN3 as Pseudomonas aeruginosa. The sequenceswere deposited in the GenBank database under the Accession number KC797117.
2.6. Plant material, growth and treatment conditions
For the pot experiment, soil was collected from the botanical garden ofGovernment College University, Faisalabad, Pakistan (31124/N, 73104/E) in March2012. The soil was sieved (2 mm) and sterilized by autoclaving at 121 1C for 15 minat 15 psi. After sterilization the soil was artificially contaminated with ZnSO4
powder (Zn 1000 mg kg�1) and thoroughly mixed. The contaminated and non-contaminated soil was packed in 2 kg plastic bags separately and left in greenhouse for 160 days for metal stabilization. During this period the average soilmoisture was maintained at 60 percent. Other properties of the botanical gardensoil include the pH 7.4, EC 2.3 dS m�1, organic matter 0.43 percent and available P8.3 mg kg�1 while the Zn content was 20 mg kg�1.
Seeds of winter wheat (Triticum aestivum L.) were obtained from NIAB,Faisalabad, Pakistan. Healthy seeds were surface sterilized with 1 percent mercuricchloride solution for 3 min and then washed with sterilized double distilled water.The seeds were sown in earthen pots lined with polythene layer containing 1.5 kgof autoclaved soil. All the pots (twelve pots) were placed in natural light (sunlight)with average 21 1C and 8 1C temperature for day and night respectively. Averagehumidity during the study period was 60 percent. The experiment was laid out incomplete randomized design (CRD) with four treatments as T0 (Control containsnon-contaminated soil), T1 (Zn amended soil), T2 (Non-contaminated soilþP.aeruginosa), T3 (Zn amended soilþP. aeruginosa). There were three replicationsfor each treatment (four to five plants per pot). For treatment T0 and T1 sterilizedseeds were sown while for treatment T2 and T3, seeds were further dipped for 1 h
F. Islam et al. / Ecotoxicology and Environmental Safety 104 (2014) 285–293286

in bacterial culture (108 CFU mL�1) before sowing. After emergence of seedling,2 mL of bacterial inoculum (108 CFU mL�1) was poured along sides of emergingseedling.
2.7. Colonization of P. aeruginosa after inoculation
P. aeruginosa colonization in rhizosphere soil was determined by the method ofHe et al. (2009). Briefly, 1 g of rhizospheric soil was removed from the roots andshaken in 10 mL sterile water for 30 min. The resulting suspension was evaluatedfor CFU according to the dilution plate method by adding 250 mg kg�1 Zn in LBmedia. The plates were incubated for seven days at 28 1C. The re-isolated strainswere identified for colony characteristics and heavy metal resistance against theparent strain.
2.8. Plant biomass, zinc, nitrogen and phosphorus contents
Plants were harvested 45 days after seed sowing (DAS) and roots were washedwith deionized water and blot dried. Roots and shoots were separated and biomass(dry weight) was recorded after drying in an electric heating oven at 70 1C for fivedays. Oven dried 100 mg plant material was taken into a digestion flask anddigested with nitric acid following Allen et al. (1986) method. The Zn concentrationin plant samples was measured using atomic absorption spectrophotometer inflame mode (air–acetylene) while nitrogen (N) and phosphorus (P) contents inplants were measured following the method ascribed by the Association of OfficialAnalytical Chemists (AOAC, 1990).
2.9. Biochemical studies
Leaf photosynthetic pigments (chlorophyll a, b and total chlorophyll) and totalsoluble protein were estimated according to the method of Arnon (1949) andBradford (1976), respectively.
2.10. Assay of antioxidant enzymes
For the extraction of enzymes fresh leaves (0.5 g) were homogenized in 10 mLof ice cold potassium phosphate buffer (pH 7.0) in an ice bath by grinding with amortar and pestle. The homogenate was centrifuged at 12,000g for 20 min at 4 1C.The supernatant was stored at 4 o C and used for the determination of variousantioxidant enzymes.
Superoxide dismutase (SOD) activity was measured through the photoreduc-tion of nitro blue tetrazolium chloride (NBT) (Dhindsa and Matowe, 1981). Thereaction mixture contained 50 μL of 33 mM NBT, 100 μL of 10 mM L-methionine,50 μL of 0.0033 mM riboflavin in 250 μL of 50 mM sodium phosphate buffer. Thereaction mixture was placed under lamp below 15 W for 25 min then stopped thereaction by switching off the light. Non-illuminated and illuminated reactionswithout supernatant served as control. The absorbance was read at 560 nm. Oneunit of SOD activity was defined as the quantity of SOD required to produce a 50percent inhibition of NBT, and the specific enzyme activity was expressed as unitsmg�1 protein.
For the measurement of catalase (CAT), modified method of Aebi (1974) wasfollowed. In 3 mL of reaction mixture of 1.9 mL of 50 mM phosphate buffer (pH 7),100 μL of enzyme extract was added and incubated for 25 min. The absorbance ofmixture was measured at 240 nm by the addition of 1 mL of 6 mM H2O2 in reactionmixture for 2 min. A 0.1 unit min�1 change in absorbance was defined as 1 unit ofCAT activity, and CAT activity was expressed as units mg�1 protein.
G-POD activity was measured following the method of Rao et al. (1996) withslight modification. Three mL of reaction mixture was formed by the addition of2.2 mL of 50 mM phosphate buffer (pH 7), 300 μL of 20 mM of guaiacol with200 μL of enzyme extract. The mixture was incubated for 5 min at 25 1C and then6 mM of H2O2 (300 μL) was added in reaction mixture to start the reaction. Changein the absorbance for 2 min was recorded for the calculation of POD activity. Oneunit of POD activity was defined as an absorbance change of 0.01 unit min�1 andPOD activity was expressed as units mg�1 protein.
2.11. MDA and H2O2 contents
To find out the extent of Zn-induced oxidative damage on membranes, weevaluated changes in lipid peroxidation by measuring the malondialdehyde (MDA)formation in the leaves of wheat plant. Briefly, 1 g of fresh leaves was homogenizedin 20 mL of 0.1 percent trichloroacetic acid (TCA) solution and centrifuged for10 min at 12,000g. One mL of the supernatant was mixed with 4 mL of trichlor-oacetic acid (TCA) containing five percent thiobarbituric acid (TBA) and heated for30 min at 95 1C and cooled on ice. The mixture was centrifuged at 12,000g for10 min and absorbance of the supernatant was taken at 532 and 660 nm (Demiraland Türkan, 2005). Concentration of H2O2 was determined following Velikova et al.(2000). Fresh leaf tissue (0.5 g) was homogenized with 5 mL of 0.1 percent (w/v)TCA in a pre-chilled pestle and mortar and the homogenate was then centrifuged at
12,000g for 15 min. To 0.5 mL of the supernatant, added 0.5 mL of 10 mMpotassium phosphate buffer (pH 7.0) and 1 mL of 1 M KI (potassium iodide) wereadded. The mixture was vortexed and the absorbance was read at 390 nm (Velikovaet al., 2000). The concentration of H2O2 was calculated using a standard curveprepared with known concentrations of H2O2.
2.12. Assay of non-enzymatic antioxidants
The total phenolic contents (TPCs) were measured by adding 0.5 mL of leafextract (extracted in 80 percent acetone) in 2.5 mL of Folin-Ciocalteu reagent (10percent v/v) and 2 mL of Na2CO3 (7.5 percent). The mixture was heated at 45 1C for40 min and the absorbance was measured at 765 nm. The gallic acid was used asstandard and expressed as mg of gallic acid equivalent g�1 extract. The ascorbicacid (AsA) contents in wheat leaves were estimated following the methoddescribed by Mukherjee and Choudhuri (1983).
2.13. Statistical analysis
All the data were presented as mean values of three replicates. The data setsgenerated were analyzed using a statistical package, SPSS (Version 19.0). One-wayANOVA (analysis of variance) was employed followed by Duncan's multiple rangetests to determine the significant difference among means of the treatment at5 percent significance level.
3. Results
3.1. Screening of Zn resistant bacteria and MIC
We isolated more than 200 bacterial colonies on LB media. Thecolonies of different morphologies were streaked on separate LBmedia plates for purification. All the isolated strains showedtolerance to zinc but the variation was observed among theirresistance levels. The number of bacterial isolates decreased downwith the gradual increase in the concentration of Zn in LB media.The ZN3 (P. aeruginosa), ZK3, ZA1 and ZA2 were found highlyresistant having minimum inhibitory concentration (MIC) rangingfrom 1000 to 1500 mg kg�1 against Zn. We also determined theMIC of these bacterial isolates against lead and copper metal andfound that they were also considerably tolerant to these metals(data in supplemental file). The results of Zn (II) bioaccumulationstudy showed that bacterial isolates remove more the 90 percentof metal when were inoculated in 50 mg kg�1 metal solution butthe bioaccumulation efficiency of bacterial isolates were remark-ably decreased when inoculated in 250 mg kg�1 of Zn solution(data in Supplementary materials).
3.2. Indole acetic acid (IAA) production and phosphate solubilizationcapacity
The zinc resistant bacterial isolates were screened for IAA-production and phosphate solubilization ability under Zn stress.The results showed that all the strains were able to utilizeL-tryptophan as precursor to produce IAA (Fig. 1B). Strain ZN3(similar to P. aeruginosa) produced the highest (40.3 mg L�1) IAA,whereas ZK3 produced the lowest (20.7 mg L�1) IAA among thefour isolates tested under control conditions. However, under Zn(1000 mg kg�1) stress conditions this ability to produce IAAreduced significantly in all bacterial strains except ZA1 whichshowed an increase in IAA under Zn stress. In addition to IAAproduction, all isolates demonstrated the potential for phosphatesolubilization (Fig. 1D). The maximum phosphate solubilizationwas achieved by the strain ZN3 (P. aeruginosa) (98 mg L�1)followed by ZA1 as compared with other bacterial strains. Resultsindicated that the presence of 1000 mg kg�1 of Zn also signifi-cantly suppressed the phosphate solubilization by these bacterialisolates, except ZA2 that remained unaffected in regard withthis attribute. Under Zn stress conditions, the minimum decrease
F. Islam et al. / Ecotoxicology and Environmental Safety 104 (2014) 285–293 287
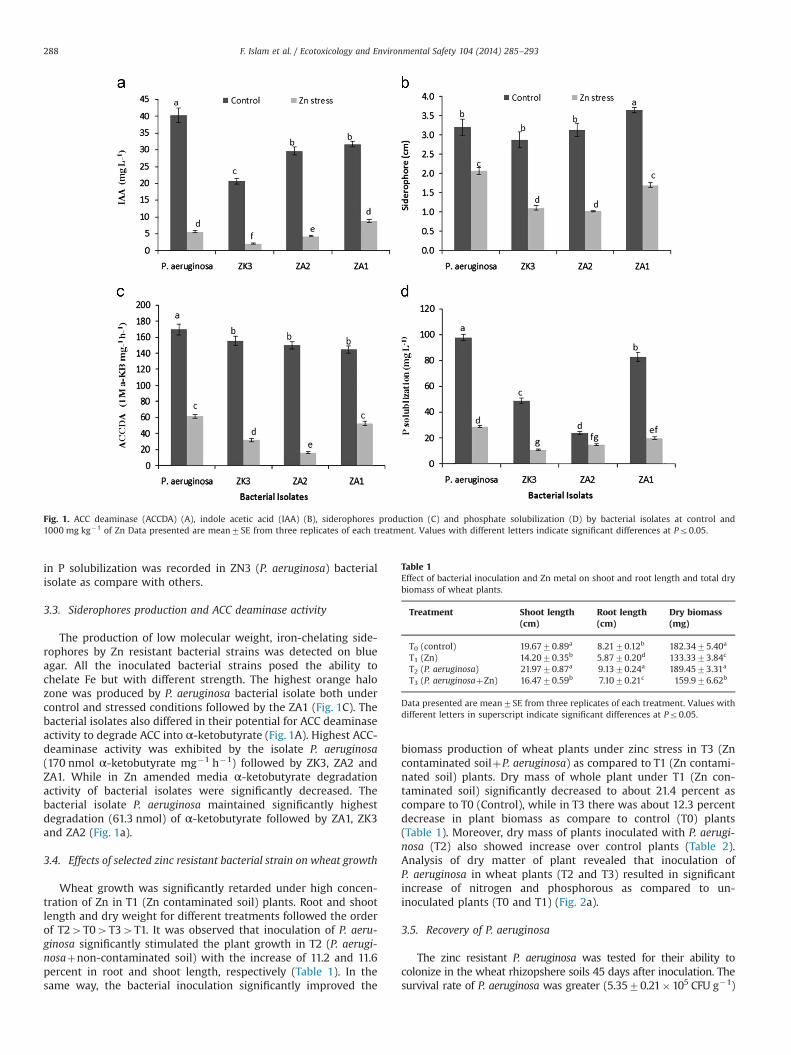
in P solubilization was recorded in ZN3 (P. aeruginosa) bacterialisolate as compare with others.
3.3. Siderophores production and ACC deaminase activity
The production of low molecular weight, iron-chelating side-rophores by Zn resistant bacterial strains was detected on blueagar. All the inoculated bacterial strains posed the ability tochelate Fe but with different strength. The highest orange halozone was produced by P. aeruginosa bacterial isolate both undercontrol and stressed conditions followed by the ZA1 (Fig. 1C). Thebacterial isolates also differed in their potential for ACC deaminaseactivity to degrade ACC into α-ketobutyrate (Fig. 1A). Highest ACC-deaminase activity was exhibited by the isolate P. aeruginosa(170 nmol α-ketobutyrate mg�1 h�1) followed by ZK3, ZA2 andZA1. While in Zn amended media α-ketobutyrate degradationactivity of bacterial isolates were significantly decreased. Thebacterial isolate P. aeruginosa maintained significantly highestdegradation (61.3 nmol) of α-ketobutyrate followed by ZA1, ZK3and ZA2 (Fig. 1a).
3.4. Effects of selected zinc resistant bacterial strain on wheat growth
Wheat growth was significantly retarded under high concen-tration of Zn in T1 (Zn contaminated soil) plants. Root and shootlength and dry weight for different treatments followed the orderof T24T04T34T1. It was observed that inoculation of P. aeru-ginosa significantly stimulated the plant growth in T2 (P. aerugi-nosaþnon-contaminated soil) with the increase of 11.2 and 11.6percent in root and shoot length, respectively (Table 1). In thesame way, the bacterial inoculation significantly improved the
biomass production of wheat plants under zinc stress in T3 (Zncontaminated soilþP. aeruginosa) as compared to T1 (Zn contami-nated soil) plants. Dry mass of whole plant under T1 (Zn con-taminated soil) significantly decreased to about 21.4 percent ascompare to T0 (Control), while in T3 there was about 12.3 percentdecrease in plant biomass as compare to control (T0) plants(Table 1). Moreover, dry mass of plants inoculated with P. aerugi-nosa (T2) also showed increase over control plants (Table 2).Analysis of dry matter of plant revealed that inoculation ofP. aeruginosa in wheat plants (T2 and T3) resulted in significantincrease of nitrogen and phosphorous as compared to un-inoculated plants (T0 and T1) (Fig. 2a).
3.5. Recovery of P. aeruginosa
The zinc resistant P. aeruginosa was tested for their ability tocolonize in the wheat rhizopshere soils 45 days after inoculation. Thesurvival rate of P. aeruginosa was greater (5.3570.21�105 CFU g�1)
Fig. 1. ACC deaminase (ACCDA) (A), indole acetic acid (IAA) (B), siderophores production (C) and phosphate solubilization (D) by bacterial isolates at control and1000 mg kg�1 of Zn Data presented are mean7SE from three replicates of each treatment. Values with different letters indicate significant differences at Pr0.05.
Table 1Effect of bacterial inoculation and Zn metal on shoot and root length and total drybiomass of wheat plants.
Treatment Shoot length(cm)
Root length(cm)
Dry biomass(mg)
T0 (control) 19.6770.89a 8.2170.12b 182.3475.40a
T1 (Zn) 14.2070.35b 5.8770.20d 133.3373.84c
T2 (P. aeruginosa) 21.9770.87a 9.1370.24a 189.4573.31a
T3 (P. aeruginosaþZn) 16.4770.59b 7.1070.21c 159.976.62b
Data presented are mean7SE from three replicates of each treatment. Values withdifferent letters in superscript indicate significant differences at Pr0.05.
F. Islam et al. / Ecotoxicology and Environmental Safety 104 (2014) 285–293288

in only P. aeruginosa inoculation soil (T2) than in Zn stressedP. aeruginosa inoculated soil (T3) (3.170.43�103 CFU g�1). TheP. aeruginosa was not detectable in the uninoculated soils (T0 and T1).
3.6. N, P and Zn uptake in wheat plants with P. aeruginosainoculation
Data presented in Fig. 2a for N and P uptake show thatimposition of Zn stress significantly reduced the uptake of bothnutrients. However, inoculation with P. aeruginosa found to beeffective in reducing the adverse effects of Zn stress on uptake of Nand P. Uptake of Zn in shoot and root increased significantly in Zntreated plants and more increase was recorded in root as com-pared with shoot. A significant reduction of Zn uptake wasrecorded due to inoculation with P. aeruginosa (Fig. 2b).
3.7. Chlorophyll content and total soluble proteins
To examine the biochemical response of bacterial inoculationon wheat plants, chlorophyll a, chlorophyll b, total chlorophyll andtotal soluble proteins were determined. Significant changes inchlorophyll a, b and total chlorophyll were observed in response tobacterial inoculation. The decrease in chlorophyll a, b and totalchlorophyll in T1 (Zn contaminated soil) was 38.8, 46.7, and 40percent, respectively. Whereas, in T3 (Zn contaminated soilþP. aeruginosa) the reduction in chlorophyll a, b and total chlor-ophyll was 10.4, 15.6 and 11.4 percent, respectively. On the otherhand, the bacterial isolates in non-contaminated soil (T2)enhanced the production of Chl a, b and total chlorophyll up to4.6, 10 and 5.5 percent, respectively (Table 2).
The total soluble protein significantly decreased (56.5 percent)in T1 (Zn contaminated soil) whereas, in T3 (Zn contaminatedsoilþP. aeruginosa) the increase in total soluble protein was 80percent as compare to the T1 plants but 21.7 percent decrease was
found compared to the control (T0), though, P. aeruginosa inocula-tions in non-contaminated soil (T2) increased total soluble proteinup to 8.7 percent (Table 2).
3.8. Oxidative stress and antioxidant enzyme's activities
The damages due to oxidative stress in wheat plants wasmeasured in terms of increase or decrease in endogenous levelsof H2O2 and malondialdehyde (MDA) contents. MDA contents tellthe extent of lipid peroxidation of membranes when they areexposed to ROS. A significant change in MDA contents was foundin the leaves of T1 (Zn contaminated soil) and T3 (Zn contaminatedsoilþP. aeruginosa) plants as compare to the control plants. Inleaves of T1 plants MDA contents increased by 145 percent whileP. aeruginosa inoculated Zn stressed plants (T3) showed lessdamage to membranes with 65 percent increase in MDA contentsover control (Table 2). Bacterial and Zn induced changes inhydrogen peroxide contents are also represented in Table 2. Werecorded that P. aeruginosa inoculation significantly reduced theH2O2 contents in T3 (Zn contaminated soilþP. aeruginosa) plantsas compared to the plants grown in Zn contaminated soil (T1).About 55 percent increase in H2O2 contents was measured in T1plants while in T3 plants only 27 percent increase in H2O2 contentswas recorded (Table 2).
Activities of different enzymatic antioxidants such as CAT, SODand POD effected significantly due to Zn stress and P. aeruginosainoculation in wheat plants. Zn stress decreased the CAT activitywhile an increase was recorded in SOD and POD compared withcontrol (T0). The increase in the activities of SOD and G-POD was124 and 80 percent respectively while the decrease in CAT was 10percent as compared to the control plants (T0). P. aeruginosainoculation under Zn stress (T3) showed a significant increase inSOD (161 percent), G-POD (180 percent) and CAT (83 percent)activities (Fig. 3). Furthermore, the low damage to cellular
Table 2Chlorophyll a (Chl. a), chlorophyll b (Chl. b), total chlorophyll (T.Chl.) (mg g�1 FW), total soluble proteins (TSP), Malonaldehyde (MDA nmol g�1 f.wt) and Hydrogen peroxide(H2O2 ng g�1 FW) production of wheat plants subjected to Zn stress and Pseudomonas aeruginosa inoculants.
z Chl. a Chl. b T. Chl. TSP (mg g�1 FW) MDA (nmol g�1 FW) H2O2 (ng g�1 FW)
T0 (control) 2.1970.11ab 0.4570.02a 2.6470.10a 23.0071.15a 8.6070.45c 92.3372.91c
T1 (Zn) 1.3470.04c 0.2470.01c 1.5870.07c 10.0071.00c 21.1370.95a 142.0072.65a
T2 (P. aeruginosa) 2.2970.05a 0.5070.02a 2.7870.12a 25.0071.25a 8.3070.45c 100.3375.36c
T3 (P. aeruginosaþZn) 1.9670.11b 0.3870.01b 2.3470.08b 18.0071.32b 13.9670.64b 117.0076.08b
Data presented are mean7SE from three replicates of each treatment. Values with different letters in superscript indicate significant differences at Pr0.05.
Fig. 2. The effect of Pseudomonas aeruginosa inoculation on N and P concentration (percent of dry matter) and Zn uptake (μg g�1 D.W) of wheat plants under the treatments:T0 (non-contaminated soil), T1 (Zn contaminated soil), T2 (non-contaminated soilþPseudomonas aeruginosa) and T3 (Zn contaminated soilþPseudomonas aeruginosa). Datapresented are mean7SE from three replicates of each treatment. Values with different letters indicate significant differences at Pr0.05.
F. Islam et al. / Ecotoxicology and Environmental Safety 104 (2014) 285–293 289

membranes in terms of MDA and H2O2 contents in P. aeruginosainoculated plants is positively associated with their high antiox-idant (enzymatic and non-enzymatic) activities.
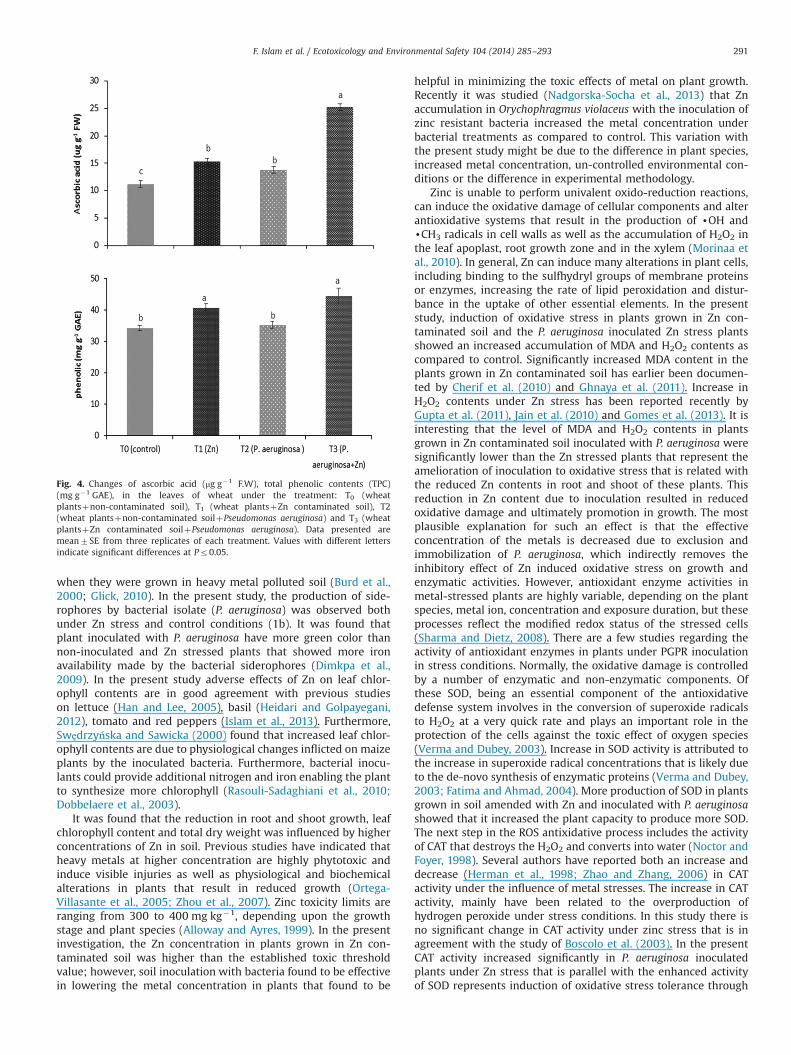
3.9. Non-enzymatic antioxidants
Imposition of Zn stress significantly increased the content ofboth ascorbic acid and total phenolic contents (TPC) in wheatleaves as compared to control (T0) plants. P. aeruginosa inoculationfurther enhanced the contents of ascorbic acid and total phenolicsin Zn stressed plants (T3). However, inoculation dependent sucheffects were not recorded in T2 plants as compared with T0 and T3(Fig. 4).
4. Discussion
The present study reflects the preliminary work done inisolation of heavy metal resistant PGPR from an agricultural landirrigated with industrial effluents. We isolated Zn resistant P.aeruginosa having a MIC of 1500 mg kg�1 and showed multipleplant growth promoting activities. When this bacterium wasinoculated under Zn stress, it significantly increased root andshoot length as well as the total dry weight of plants (Stefan etal., 2013). On the other hand, the reduction of plant growth causedby Zn stress as observed in this study has also been reported insome other plant species such in Trifolium repens (Vivas et al.,2006), and in sugarcane (Jain et al., 2010). Under stress conditions,bacteria in rhizosphere may enhance the plant growth by different
mechanisms such as by optimizing the supply of nutrients,stimulating plant growth by the synthesis of phytohormones IAAand ACC, solubilization of inorganic phosphorus, bioaccumulationor leaching of metals and inhibiting the activity of pathogens(Yang et al., 2009). Typically a bacterium directly affects the plantgrowth and development by using one or more of these mechan-isms (Gamalero et al., 2008). Among these traits, the phytohor-mone auxin (IAA) release by PGPR has various direct effects onplant growth under stress conditions (Brown, 1974; Patten andGlick, 1996; de Garcia Salamone et al., 2005). The main effect ofbacterial IAA production is the improvement of lateral andadventitious roots growth and development that result enhancedmineral and nutrient uptake (Lambrecht et al., 2000; Steenhoudtand Vanderleyden, 2000). Similar to our study, bacterial inocu-lated root elongation has been reported in wheat seedling(Egamberdieva, 2009), canola (Patten and Glick, 2002) as well asin Medicago truncatula under stress conditions. Increased phos-phorous contents (2a) in P. aeruginosa inoculated plants wereobserved that is due to enhanced availability of phosphorus inrooting medium (Jiang et al., 2008) as a P solublization activity ofP. aeruginosa was determined in quantitative analysis (Fig. 1d). Inaddition to the improvement in uptake of necessary micronutri-ents (P, N), PGPR also reduces the heavy metal toxicity bydecreasing the levels of acetylene production because of thepresences of ACC deaminase, that break down the ethyleneprecursor ACC and decreases the plant ethylene level resulting inincreased plant growth (Saleem et al., 2007; Long et al., 2008).Moreover, siderophore producing PGPR help to prevent plantsfrom becoming chlorotic due to the availability of Fe to plants
Fig. 3. Changes of catalase (CAT), superoxide dismutase (SOD), peroxidase (POD) in the leaves of wheat under the treatment: T0 (non-contaminated soil), T1 (Zncontaminated soil), T2 (non-contaminated soilþPseudomonas aeruginosa) and T3 (Zn contaminated soilþPseudomonas aeruginosa). Data presented are mean7SE from threereplicates of each treatment. Values with different letters indicate significant differences at Pr0.05.
F. Islam et al. / Ecotoxicology and Environmental Safety 104 (2014) 285–293290
when they were grown in heavy metal polluted soil (Burd et al.,2000; Glick, 2010). In the present study, the production of side-rophores by bacterial isolate (P. aeruginosa) was observed bothunder Zn stress and control conditions (1b). It was found thatplant inoculated with P. aeruginosa have more green color thannon-inoculated and Zn stressed plants that showed more ironavailability made by the bacterial siderophores (Dimkpa et al.,2009). In the present study adverse effects of Zn on leaf chlor-ophyll contents are in good agreement with previous studieson lettuce (Han and Lee, 2005), basil (Heidari and Golpayegani,2012), tomato and red peppers (Islam et al., 2013). Furthermore,Swędrzyńska and Sawicka (2000) found that increased leaf chlor-ophyll contents are due to physiological changes inflicted on maizeplants by the inoculated bacteria. Furthermore, bacterial inocu-lants could provide additional nitrogen and iron enabling the plantto synthesize more chlorophyll (Rasouli-Sadaghiani et al., 2010;Dobbelaere et al., 2003).
It was found that the reduction in root and shoot growth, leafchlorophyll content and total dry weight was influenced by higherconcentrations of Zn in soil. Previous studies have indicated thatheavy metals at higher concentration are highly phytotoxic andinduce visible injuries as well as physiological and biochemicalalterations in plants that result in reduced growth (Ortega-Villasante et al., 2005; Zhou et al., 2007). Zinc toxicity limits areranging from 300 to 400 mg kg�1, depending upon the growthstage and plant species (Alloway and Ayres, 1999). In the presentinvestigation, the Zn concentration in plants grown in Zn con-taminated soil was higher than the established toxic thresholdvalue; however, soil inoculation with bacteria found to be effectivein lowering the metal concentration in plants that found to be
helpful in minimizing the toxic effects of metal on plant growth.Recently it was studied (Nadgorska-Socha et al., 2013) that Znaccumulation in Orychophragmus violaceus with the inoculation ofzinc resistant bacteria increased the metal concentration underbacterial treatments as compared to control. This variation withthe present study might be due to the difference in plant species,increased metal concentration, un-controlled environmental con-ditions or the difference in experimental methodology.
Zinc is unable to perform univalent oxido-reduction reactions,can induce the oxidative damage of cellular components and alterantioxidative systems that result in the production of dOH anddCH3 radicals in cell walls as well as the accumulation of H2O2 inthe leaf apoplast, root growth zone and in the xylem (Morinaa etal., 2010). In general, Zn can induce many alterations in plant cells,including binding to the sulfhydryl groups of membrane proteinsor enzymes, increasing the rate of lipid peroxidation and distur-bance in the uptake of other essential elements. In the presentstudy, induction of oxidative stress in plants grown in Zn con-taminated soil and the P. aeruginosa inoculated Zn stress plantsshowed an increased accumulation of MDA and H2O2 contents ascompared to control. Significantly increased MDA content in theplants grown in Zn contaminated soil has earlier been documen-ted by Cherif et al. (2010) and Ghnaya et al. (2011). Increase inH2O2 contents under Zn stress has been reported recently byGupta et al. (2011), Jain et al. (2010) and Gomes et al. (2013). It isinteresting that the level of MDA and H2O2 contents in plantsgrown in Zn contaminated soil inoculated with P. aeruginosa weresignificantly lower than the Zn stressed plants that represent theamelioration of inoculation to oxidative stress that is related withthe reduced Zn contents in root and shoot of these plants. Thisreduction in Zn content due to inoculation resulted in reducedoxidative damage and ultimately promotion in growth. The mostplausible explanation for such an effect is that the effectiveconcentration of the metals is decreased due to exclusion andimmobilization of P. aeruginosa, which indirectly removes theinhibitory effect of Zn induced oxidative stress on growth andenzymatic activities. However, antioxidant enzyme activities inmetal-stressed plants are highly variable, depending on the plantspecies, metal ion, concentration and exposure duration, but theseprocesses reflect the modified redox status of the stressed cells(Sharma and Dietz, 2008). There are a few studies regarding theactivity of antioxidant enzymes in plants under PGPR inoculationin stress conditions. Normally, the oxidative damage is controlledby a number of enzymatic and non-enzymatic components. Ofthese SOD, being an essential component of the antioxidativedefense system involves in the conversion of superoxide radicalsto H2O2 at a very quick rate and plays an important role in theprotection of the cells against the toxic effect of oxygen species(Verma and Dubey, 2003). Increase in SOD activity is attributed tothe increase in superoxide radical concentrations that is likely dueto the de-novo synthesis of enzymatic proteins (Verma and Dubey,2003; Fatima and Ahmad, 2004). More production of SOD in plantsgrown in soil amended with Zn and inoculated with P. aeruginosashowed that it increased the plant capacity to produce more SOD.The next step in the ROS antixidative process includes the activityof CAT that destroys the H2O2 and converts into water (Noctor andFoyer, 1998). Several authors have reported both an increase anddecrease (Herman et al., 1998; Zhao and Zhang, 2006) in CATactivity under the influence of metal stresses. The increase in CATactivity, mainly have been related to the overproduction ofhydrogen peroxide under stress conditions. In this study there isno significant change in CAT activity under zinc stress that is inagreement with the study of Boscolo et al. (2003). In the presentCAT activity increased significantly in P. aeruginosa inoculatedplants under Zn stress that is parallel with the enhanced activityof SOD represents induction of oxidative stress tolerance through
Fig. 4. Changes of ascorbic acid (μg g�1 F.W), total phenolic contents (TPC)(mg g�1 GAE), in the leaves of wheat under the treatment: T0 (wheatplantsþnon-contaminated soil), T1 (wheat plantsþZn contaminated soil), T2(wheat plantsþnon-contaminated soilþPseudomonas aeruginosa) and T3 (wheatplantsþZn contaminated soilþPseudomonas aeruginosa). Data presented aremean7SE from three replicates of each treatment. Values with different lettersindicate significant differences at Pr0.05.
F. Islam et al. / Ecotoxicology and Environmental Safety 104 (2014) 285–293 291

HalliWell Asada pathway. Similarly G-POD activity was alsoenhanced both in plants grown in Zn contaminated soil inoculatedwith bacteria as well as in plants grown in alone Zn contaminatedsoil. It seems that all antioxidative enzymes are working incombination under P. aeruginosa inoculation that is helpful forbetter growth under such conditions. The enhanced antioxidantenzymes activities under inoculation in the present study mightbe due to the increased gene/mRNA expression of plant antiox-idant enzymes than inoculated plants. As Gururani et al. (2012)found, under multiple stress (salt, heavy metals), that bacterialinoculation enhanced gene/mRNA expression of plant antioxidantenzymes which result an increase in antioxidant enzymes activity,despite decrease in MDA and H2O2 in inoculated plant as compareto un-inoculated stress plants.
In the this study, non-enzymatic antioxidants such as ascorbicacid and total phenolic contents increased in Zn stressed and P.aeruginosa inoculated plants but more increase were in the plantinoculated with bacteria. This increase in ascorbic acid andphenolic in the present study is associated with enhanced growthand negatively with the MDA and H2O2. This shows that enhancedcontent of these compounds in combination with enhanced plantgrowth and reduced H2O2 and MDA contents show their role inplant oxidative stress tolerance that is the function of soilinoculation with bacteria. The findings are similar to Gomeset al. (2013) who noted an increased concentration of ascorbicacid in bacterial inoculation under Zn stress. This increase in totalphenolic could be regarded as a protective reaction for wheatunder Zn stress (Kováčik et al., 2009). So, inoculation of PGPbacterial strains as a biotechnological tool to increase the second-ary metabolite production will be helpful in understanding certainadaptive processes that are poorly understood at present(Cappellari et al., 2013).
Acknowledgment
The authors are thankful to Associate Editor Herbert Allen andanonymous reviews for their valuable suggestions. This work issupported financially by Higher Education Commission (HEC),Pakistan. The results presented in this paper are a part of M. Philstudies of Mr. Faisal Islam.
Appendix A. Supplementary materials
Supplementary materials associated with this article can befound in the online version at http://dx.doi.org/10.1016/j.ecoenv.2014.03.008.
References
Aebi, H., 1974. Catalases. Methods Enzym. Anal. 2, 673–684.Aleem, A., Isar., J., Malik, A., 2003. Impact of long-term application of industrial
wastewater on the emergence of resistance traits in Azotobacter chroococcumisolated from rhizospheric soil. Bioresour. Technol. 86, 7–13.
Allen, E., Grimshaw, H.M., Parkinson, J.A., Grimshaw, C., Roberts, J., 1986. Chemicalanalysis, Methods in Plant Ecology. Blackwell Scientific, London, pp. 285–344.
Alloway, B., Ayres, D., 1999. Heavy metals, Chemical Basis of Environment AirPollution. PWN, Warszawa, pp. 218–246.
AOAC, 1990. Official Methods of Analysis. Association of Official Analytical Chemists,Arlington.
Arnon, D.I., 1949. Copper enzymes in isolated chloroplasts. Polyphenoloxidase inBeta vulgaris. Plant Physiol. 24, 1–15.
Belimov, A., Safronova, V., Demchinskaya, S., Piluzza, G., Bullitta, S., 2005. Cadmium-tolerant plant growth-promoting rhizobacteria associated with the roots of Indianmustard (Brassica juncea L. Czern.). Soil Biol. Biochem. 37, 241–250.
Boscolo, P.R., Menossi, M., Jorge, R.A., 2003. Aluminum-induced oxidative stress inmaize. Phytochemistry 62 (2), 181–189.
Bradford, M.M., 1976. A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein-dye binding.Anal. Biochem. 72, 248–254.
Brown, M., 1974. Seed and root bacterization. Annu. Rev. Phytopathol. 12, 181–197.Burd, G.I., Dixon, G., Glick, B.R., 2000. Plant growth-promoting bacteria that
decrease heavy metal toxicity in plants. Can. J. Microbiol. 46, 237–245.Cappellari, L.R., Santoro, M.V., Nievas, F., Giordano, W., Banchio, E., 2013. Increase of
secondary metabolite content in marigold by inoculation with plant growth-promoting rhizobacteria. Appl. Soil Ecol. 70, 16–22.
Cheneby, D., Perrez, S., Devroe, C., Hallet, S., Couton, Y., Bizouard, F., Iuretig, G.,Germon, J., Philippot, L., 2004. Denitrifying bacteria in bulk and maize-rhizospheric soil: diversity and N2O-reducing abilities. Can. J. Microbiol. 50,469–474.
Cherif, J., Derbel, N., Nakkach, M., Bergmann, Hv., Jemal, F., Lakhdar, Z.B., 2010.Analysis of in vivo chlorophyll fluorescence spectra to monitor physiologicalstate of tomato plants growing under zinc stress. J. Photochem. Photobiol. 101,332–339.
Demiral, T., Türkan, İ., 2005. Comparative lipid peroxidation, antioxidant defensesystems and proline content in roots of two rice cultivars differing in salttolerance. Environ. Exp. Bot. 53, 247–257.
Dhindsa, S.R., Matowe, W., 1981. Drought tolerance in two mosses: correlated withenzymatic defence against lipid peroxidation. J. Exp. Bot. 32, 79–91.
Dimkpa, C.O., Merten, D., Svatos, A., Büchel, G., Kothe, E., 2009. Siderophoresmediate reduced and increased uptake of cadmium by Streptomyces tendae F4and sunflower (Helianthus annuus), respectively. J. Appl. Microbiol. 107,1687–1696.
Dobbelaere, S., Vanderleyden, J., Okon, Y., 2003. Plant growth-promoting effects ofdiazotrophs in the rhizosphere. Crit. Rev. Plant Sci. 22, 07–149.
Egamberdieva, D., 2009. Alleviation of salt stress by plant growth regulators andIAA producing bacteria in wheat. Acta Physiol. Plant 31, 861–864.
Fatima, A., Ahmad, M., 2004. Certain antioxidant enzymes of Allium cepa asbiomarkers for the detection of toxic heavy metals in wastewater. Sci. TotalEnviron. 346, 256–273.
Fiske, C.H., Subbarow, Y., 1925. The colorimetric determination of phosphorus. J.Biol. Chem. 66, 375–400.
Gamalero, E., Berta, G., Massa, N., Glick, B.R., Lingua, G., 2008. Synergistic interac-tions between the ACC deaminase‐producing bacterium Pseudomonas putidaUW4 and the AM fungus Gigaspora rosea positively affect cucumber plantgrowth. FEMS Microbiol. Ecol. 64, 459–467.
Ghnaya, A.B., Hourmant, A., Couderchet, M., Branchard, M., Charles, G., 2011.Modulation of Zn-induced oxidative stress, polyamine content and metalaccumulation in rapeseed (Brassica napus cv. Jumbo) regenerated from trans-versal thin cell layers in the presence of zinc. Int. Res. J. Biotechnol. 2, 062–071.
Glick, B.R., 2010. Using soil bacteria to facilitate phytoremediation. Biotech. Adv. 28,367–374.
Gomes, M, Duarte, D., Carneiro, M.M., Barreto, L.C., Carvalho, M., Soares, A.M.,Guilherme, L.R.G, Garcia, Q.S., 2013. Zinc tolerance modulation in Myracrodruonurundeuva plants. Plant Physiol. Biochem. 67, 1–6.
Gordon, S.A., Weber, R.P., 1951. Colorimetric estimation of indoleacetic acid. PlantPhysiol. 26, 192–195.
Gupta, B., Pathak, G., Pandey, N., 2011. Induction of oxidative stress and antioxidantresponses in Vigna mungo by zinc stress. Russ. J. Plant Physiol. 58, 85–91.
Gururani, M., Upadhyaya, C., Baskar, V., Venkatesh, J., Nookaraju, A., Park, S., 2012.Plant growth-promoting rhizobacteria enhance abiotic stress tolerance insolanum tuberosum through inducing changes in the expression of ros-scavenging enzymes and improved photosynthetic performance. J. PlantGrowth Regul. 32, 245–258.
Han, H., Lee, K., 2005. Plant growth promoting rhizobacteria effect on antioxidantstatus, photosynthesis, mineral uptake and growth of lettuce under soil salinity.Res. J. Agric. Biol. Sci. 1, 210–215.
Herman, B., Biczak, R., Gurgul, E., 1998. Effect of 1, 10-phenanthro- line onperoxidase and catalase activity and chlorophyll, sugar, and ascorbic acidcontents. Biol. Plant 41, 607–613.
Harley, J., Prescott, L., 2002. Laboratory Exercises in Microbiology.He, C.Q., Tan, G.E., Liang, X., Du, W., Chen, Y.L., Zhi, G.Y., Zhu, Y., 2010. Effect of Zn-
tolerant bacterial strains on growth and Zn accumlation in Orychophragmusviolaceus. Appl. Soil Ecol. 44, 1–5.
He, H., Ye, Z., Yang, D., Yan, J., Xiao, L., Zhong, T., Yuan, M., Cai, X., Fang, Z., Jing, Y.,2012. Characterization of endophytic Rahnella sp. JN6 from Polygonum pub-escens and its potential in promoting growth and Cd, Pb, Zn uptake by Brassicanapus. Chemosphere 90, 1960–1965.
He, L.Y., Chen, Z.J., Ren, G.D., Zhang, Y.F., Qian, M., Sheng, X.F., 2009. Increasedcadmium and lead uptake of a cadmium hyperaccumulator tomato bycadmium-resistant bacteria. Ecotox. Environ. Safe 72, 1343–1348.
Heidari, M., Golpayegani, A., 2012. Effects of water stress and inoculation with plantgrowth promoting rhizobacteria (PGPR) on antioxidant status and photosyn-thetic pigments in basil (Ocimum basilicum L.). J. Saudi Soc. Agric. Sci. 11, 57–61.
Islam, M., Sultana, T., Joe, M.M., Yim, W., Cho, J.C., Sa, T., 2013. Nitrogen‐fixingbacteria with multiple plant growth-promoting activities enhance growth oftomato and red pepper. J. basic microbiol. 53, 1004–1015.
Jain, R., Srivastava, S., Solomon, S., Shrivastava, A.K., Chandra, A., 2010. Impact ofexcess zinc on growth parameters, cell division, nutrient accumulation, photo-synthetic pigments and oxidative stress of sugarcane (Saccharum spp.). ActaPhysiol. Plant 32, 979–986.
Jalili, F., Khavazi, K., Pazira, E., Nejati, A., Rahmani, H.A., Sadaghiani, H.R., Miransari,M., 2009. Isolation and characterization of ACC deaminase-producing
F. Islam et al. / Ecotoxicology and Environmental Safety 104 (2014) 285–293292

fluorescent pseudomonas, to alleviate salinity stress on canola (Brassica napus L.)growth. J. Plant Physiol. 166, 667–674.
Jiang, C., Sheng, X., Qian, M., Wang, Q., 2008. Isolation and characterization of aheavy metal-resistant Burkholderia sp. from heavy metal-contaminated paddyfield soil and its potential in promoting plant growth and heavy metalaccumulation in metal-polluted soil. Chemosphere 72, 157–164.
John, R., Ahmad, P., Gadgil, K., Sharma, S., 2008. Effect of cadmium and lead ongrowth, biochemical parameters and uptake in Lemna polyrrhiza L. Plant SoilEnviron. 54, 262.
Kováčik, J., Klejdus, B., Hedbavny, J., Štork, F., Bačkor, M., 2009. Comparison ofcadmium and copper effect on phenolic metabolism, mineral nutrients andstress-related parameters in Matricaria chamomilla plants. Plant Soil 320,231–242.
Lambrecht, M., Okon, Y., Vande, B.A., Vanderleyden, J., 2000. Indole-3-acetic acid: areciprocal signalling molecule in bacteria–plant interactions. Trends Microbiol.8, 298–300.
Long, H.H., Schmidt, D.D., Baldwin, I.T., 2008. Native bacterial endophytes promotehost growth in a species-specific manner; phytohormone manipulations do notresult in common growth responses. PloS One 3, e2702.
McLellan, T., Marr, E.S., Wondrack, L.M., Subashi, T.A., Aeed, P.A., Han, S., Xu, Z.,Wang, I.K., Maguire, B.A., 2009. A systematic study of 50S ribosomal subunitpurification enabling robust crystallization. Acta Crystallogr. D65, 1270–1282.
Morinaa, F., Jovanovicb, L., Mojovicc, M., Vidovica, M., D, P., SV, J, 2010. Zinc-inducedoxidative stress in Verbascum thapsus is caused by an accumulation of reactiveoxygen species and quinhydrone in the cell wall. Physiol. Plant 140, 209–224.
Mukherjee, S., Choudhuri, M., 1983. Implications of water stress‐induced changes inthe levels of endogenous ascorbic acid and hydrogen peroxide in Vignaseedlings. Physiol. Plant 58, 166–170.
Nadgorska-Socha, A., Kafel, A., Kandziora-Ciupa, M., Gospodarek, J., Zawisza-Raszka,A., 2013. Accumulation of heavy metals and antioxidant responses in Vicia fabaplants grown on monometallic contaminated soil. Environ. Sci. Pollut. Res. Int.20, 1124–1134.
Noctor, G., Foyer, C.H., 1998. Ascorbate and glutathione: keeping active oxygenunder control. Annu. Rev. Plant Biol. 49, 249–279.
Ortega-Villasante, C., Rellán-Alvarez, R., Del-Campo, F.F., Carpena-Ruiz, R.O., Her-nández, LE., 2005. Cellular damage induced by cadmium and mercury inMedicago sativa. J. Exp. Bot. 56, 2239–2251.
Patten, C., Glick, B., 1996. Bacterial biosynthesis of indole-3-acetic acid. Can. J.Microbiol. 42, 207–220.
Patten, C., Glick, B., 2002. The role of bacterial indoleacetic acid in the developmentof the host plant root system. App. Environ. Microbiol. 68, 3795–3801.
Rajkumar, M., Prasad, M., Freitas, H., Ae, N., 2009. Biotechnological applications ofserpentine soil bacteria for phyto-remediation of trace metals. Crit. Rev.Biotechnol. 29, 120–130.
Rao, L., Perez, D., White, E., 1996. Lamin proteolysis facilitates nuclear events duringapoptosis. J. Cell Biol. 135, 1441–1455.
Rasouli-Sadaghiani, M., Hassani, A., Barin, M., Danesh, Y.R., Sefidkon, F., 2010. Effectsof arbuscular mycorrhizal (AM) fungi on growth, essential oil production andnutrients uptake in basil. J. Med. Plants Res. 4, 2222–2228.
de Garcia Salamone, I.E., Hynes, R.K., Nelson, L.M., 2005. Role of cytokinins in plantgrowth promotion by rhizosphere bacteria. In: ZA, S. (Ed.), PGPR: Biocontroland Biofertilization.. Springer, Dordrecht, The Netherlands, pp. 173–195.
Saleem, M., Arshad, M., Hussain, S., Bhatti, A.S., 2007. Perspective of plant growthpromoting rhizobacteria (PGPR) containing ACC deaminase in stress agricul-ture. J. Ind. Microbiol. Biotechnol. 34, 635–648.
Schwyn, B., Neilands, J., 1987. Universal chemical assay for the detection anddetermination of siderophores. Anal. Biochem. 160, 47–56.
Sharma, S., Dietz, K., 2008. The relationship between metal toxicity and cellularredox imbalance. Trends Plant Sci. 14, 43–50.
Sheng, X.F., Xia, J.J., Jiang, C.Y., He, L.Y., Qian, M., 2008. Characterization of heavymetal-resistant endophytic bacteria from rape (Brassica napus) roots and theirpotential in promoting the growth and lead accumulation of rape. Environ. Poll.156, 1164–1170.
Steenhoudt, O., Vanderleyden, J., 2000. Azospirillum, a free‐living nitrogen‐fixingbacterium closely associated with grasses: genetic, biochemical and ecologicalaspects. FEMS Microbiol. Rev. 24, 487–506.
Stefan, M., Munteanu, N., Stoleru, V., Mihasan, M., Hritcu, L., 2013. Seed inoculationwith plant growth promoting rhizobacteria enhances photosynthesis and yieldof runner bean (Phaseolus coccineus L.). Sci. Hortic. 151, 22–29.
Swędrzyńska, D., Sawicka, A., 2000. Effect of inoculation with Azospirillumbrasilense on development and yielding of maize (Zea mays ssp. SaccharataL.) under different cultivation conditions. Pol. J. Environ. Stud. 9, 505–550.
Taniguchi, J., Hemmi, H., Tanahashi, K., Amano, N., Nakayama, T., Nishino, T., 2000.Zinc biosorption by a zinc-resistant bacterium, Brevibacterium sp. strain HZM-1.J. Micro. Biotechnol. 54, 581–588.
Velikova, V., Yordanov, I., Edreva, A., 2000. Oxidative stress and some antioxidantsystems in acid rain-treated bean plants: protective role of exogenous poly-amines. Plant Sci. 151, 59–66.
Verma, S., Dubey, R., 2003. Lead toxicity induces lipid peroxidation and alters theactivities of antioxidant enzymes in growing rice plants. Plant Sci. 164,645–655.
Vivas, A., Biro, B., Ruiz-Lozano, J., Barea, J., Azcon, R., 2006. Two bacterial strainsisolated from a Zn-polluted soil enhance plant growth and mycorrhizalefficiency under Zn-toxicity. Chemosphere 62, 1523–1533.
Yang, J., Kloepper, J., Ryu, C., 2009. Rhizosphere bacteria help plants tolerate abioticstress. Trends Plant Sci. 14, 1–4.
Zhao, F., Zhang, H., 2006. Salt and paraquat stress tolerance results from co-expression of the Suaeda salsa glutathione S-transferase and catalase intransgenic rice. Plant Cell Tissue Organ Cult. 86, 349–358.
Zhou, Z., Huang, S., Guo, K., Mehta, S., Zhang, P., Yang, Z., 2007. Metabolicadaptations to mercury-induced oxidative stress in roots of Medicago sativa L.Inorg. Biochem. 101, 1–9.
F. Islam et al. / Ecotoxicology and Environmental Safety 104 (2014) 285–293 293
Copyright © 2022 FDOKUMEN




















