
Influence of furostanol glycosides treatments on strawberry (Fragaria x ananassa Duch.) growth and...
10
Scientia Horticulturae 169 (2014) 179–188 Contents lists available at ScienceDirect Scientia Horticulturae journal h om epage: www.elsevier.com/locate/scihorti Influence of furostanol glycosides treatments on strawberry (Fragaria × ananassa Duch.) growth and photosynthetic characteristics under drought condition Raluca-Petronela Caulet a , Gica Gradinariu a , Dorina Iurea b , Aliona Morariu a,∗ a University of Agricultural Sciences and Veterinary Medicine Ias ¸ i, Romania b Biological Research Institute, Ias ¸ i, Romania a r t i c l e i n f o Article history: Received 24 July 2013 Received in revised form 22 February 2014 Accepted 24 February 2014 Keywords: Furostanol glycosides Strawberry Drought Photosynthesis Chlorophyll fluorescence a b s t r a c t Water availability is one of the major limiting factors for plant growth and productivity. In recent years, efforts to search for specific and practical approaches to improve the adaptability of plants to this envi- ronmental condition have been made. In this study we have examined the influence of two furostanol glycoside application on the capacity for acclimation to drought of two strawberry (Fragaria × ananassa Duch.) cultivars, Real and Magic, grown in greenhouse conditions. Two regimes of irrigation were used: well-watered (80% WHC) and drought stress (50% WHC) for one month. Two furostanol glycoside aque- ous solutions (G1 and G2) were applied by foliar spraying during late afternoon at 3 days intervals on 5 mature plants for each variant. Changes in plant growth and photosynthesis (gas exchange and chlorophyll fluorescence measurements) were studied. In drought conditions, glycoside application induced a decrease in leaf size and increase in roots length, resulting in significant increases of root/shoot ratio. The two glycoside treatments alleviated the effect of drought and improved photosynthetic rate (A) and water use efficiency (A/E) in both cultivars. In addition, G1 treatment reduced the severity of drought effect by increasing the stomata responsiveness thus maintaining a higher relative water content (RWC), while G2 treatment led to a lower decrease of stomatal conductance (g s ) and transpiration rate (E) and a significant decrease in internal CO 2 (C i ) concentration compared to untreated plants subjected to stress. The chlorophyll fluorescence measure- ments indicate that furostanol glycoside treatments mitigated the reduction of photosystem II efficiency (PS II) and electron transport rate (ETR) caused by water stress, compared to untreated plants. More- over, under drought stress, treated plants recorded higher values of non-photochemical processes (NPQ) than untreated ones. One can safely assume that glycoside treatments induced an enhancement of non- photochemical processes as well as an alleviation of the drought-induced alteration in PS II by increasing the efficiency of light utilization and dissipation of excitation energy in the PSII antennae. © 2014 Elsevier B.V. All rights reserved. 1. Introduction Drought is one of the most commonly limiting environmen- tal factors affecting plant growth and productivity. In response Abbreviations: WHC, water holding capacity; G1, glycoside 1; G2, glycoside 2; A, photosynthetic rate; E, transpiration rate; gs , stomatal conductance; WUE = (A/E), water use efficiency; C i , internal CO2; RWC, relative water content; PSI, photosys- tem I; PSII, photosystem II; Fv/Fm, maximal quantum yield of PSII efficiency; PSII, quantum efficiency of PSII; ETR, electron transport rate; NPQ, non-photochemical process; PAR, photosynthetic active radiation; RuBP, ribulose-biphosphate; ROS, reactive oxygen species. ∗ Corresponding author. Tel.: +40 232407370. E-mail address: [email protected] (A. Morariu). to water deficit, plants developed drought avoidance and drought tolerance strategies (Chaves et al., 2003). Morphological adapta- tions in response to water stress involve reduction of leaf area for minimization of water loss (Singer et al., 2003) and increase of investments in the root system (Jackson et al., 2000). The lat- ter occurs as a consequence of altering assimilates partitioning by directing a supplementary amount of phytoassimilants to the roots for increasing water and nutrient uptake (Starck et al., 1995). The strawberry is a plant with a large demand for water, due to a shallow root system, large leaf area and fruits with a high content of water. It is well known that plant roots perform many essential functions, including water and nutrient uptake, storage of reserves, synthesis of specific compounds, etc. (López-Bucio et al., 2003). The size and the architecture of the root system determine http://dx.doi.org/10.1016/j.scienta.2014.02.031 0304-4238/© 2014 Elsevier B.V. All rights reserved.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Influence of furostanol glycosides treatments on strawberry (Fragaria x ananassa Duch.) growth and...

I(u
Ra
b
a
ARRA
KFSDPC
1
t
Awtqpr
h0
Scientia Horticulturae 169 (2014) 179–188
Contents lists available at ScienceDirect
Scientia Horticulturae
journa l h om epage: www.elsev ier .com/ locate /sc ihor t i
nfluence of furostanol glycosides treatments on strawberryFragaria × ananassa Duch.) growth and photosynthetic characteristicsnder drought condition
aluca-Petronela Cauleta, Gica Gradinariua, Dorina Iureab, Aliona Morariua,∗
University of Agricultural Sciences and Veterinary Medicine Ias i, RomaniaBiological Research Institute, Ias i, Romania
r t i c l e i n f o
rticle history:eceived 24 July 2013eceived in revised form 22 February 2014ccepted 24 February 2014
eywords:urostanol glycosidestrawberryroughthotosynthesishlorophyll fluorescence
a b s t r a c t
Water availability is one of the major limiting factors for plant growth and productivity. In recent years,efforts to search for specific and practical approaches to improve the adaptability of plants to this envi-ronmental condition have been made. In this study we have examined the influence of two furostanolglycoside application on the capacity for acclimation to drought of two strawberry (Fragaria × ananassaDuch.) cultivars, Real and Magic, grown in greenhouse conditions. Two regimes of irrigation were used:well-watered (80% WHC) and drought stress (50% WHC) for one month. Two furostanol glycoside aque-ous solutions (G1 and G2) were applied by foliar spraying during late afternoon at 3 days intervalson 5 mature plants for each variant. Changes in plant growth and photosynthesis (gas exchange andchlorophyll fluorescence measurements) were studied.
In drought conditions, glycoside application induced a decrease in leaf size and increase in roots length,resulting in significant increases of root/shoot ratio. The two glycoside treatments alleviated the effectof drought and improved photosynthetic rate (A) and water use efficiency (A/E) in both cultivars. Inaddition, G1 treatment reduced the severity of drought effect by increasing the stomata responsivenessthus maintaining a higher relative water content (RWC), while G2 treatment led to a lower decreaseof stomatal conductance (gs) and transpiration rate (E) and a significant decrease in internal CO2 (Ci)concentration compared to untreated plants subjected to stress. The chlorophyll fluorescence measure-ments indicate that furostanol glycoside treatments mitigated the reduction of photosystem II efficiency
(�PS II) and electron transport rate (ETR) caused by water stress, compared to untreated plants. More-over, under drought stress, treated plants recorded higher values of non-photochemical processes (NPQ)than untreated ones. One can safely assume that glycoside treatments induced an enhancement of non-photochemical processes as well as an alleviation of the drought-induced alteration in PS II by increasingthe efficiency of light utilization and dissipation of excitation energy in the PSII antennae.© 2014 Elsevier B.V. All rights reserved.
. Introduction
Drought is one of the most commonly limiting environmen-al factors affecting plant growth and productivity. In response
Abbreviations: WHC, water holding capacity; G1, glycoside 1; G2, glycoside 2;, photosynthetic rate; E, transpiration rate; gs, stomatal conductance; WUE = (A/E),ater use efficiency; Ci , internal CO2; RWC, relative water content; PSI, photosys-
em I; PSII, photosystem II; Fv/Fm, maximal quantum yield of PSII efficiency; �PSII,uantum efficiency of PSII; ETR, electron transport rate; NPQ, non-photochemicalrocess; PAR, photosynthetic active radiation; RuBP, ribulose-biphosphate; ROS,eactive oxygen species.∗ Corresponding author. Tel.: +40 232407370.
E-mail address: [email protected] (A. Morariu).
ttp://dx.doi.org/10.1016/j.scienta.2014.02.031304-4238/© 2014 Elsevier B.V. All rights reserved.
to water deficit, plants developed drought avoidance and droughttolerance strategies (Chaves et al., 2003). Morphological adapta-tions in response to water stress involve reduction of leaf areafor minimization of water loss (Singer et al., 2003) and increaseof investments in the root system (Jackson et al., 2000). The lat-ter occurs as a consequence of altering assimilates partitioning bydirecting a supplementary amount of phytoassimilants to the rootsfor increasing water and nutrient uptake (Starck et al., 1995).
The strawberry is a plant with a large demand for water, dueto a shallow root system, large leaf area and fruits with a high
content of water. It is well known that plant roots perform manyessential functions, including water and nutrient uptake, storage ofreserves, synthesis of specific compounds, etc. (López-Bucio et al.,2003). The size and the architecture of the root system determine
1 orticu
tg1dhfp
hhgPa2aacmsaet
ht(s((ea
afosNmMdacAfwebnune
oarteopeutasDp
80 R.-P. Caulet et al. / Scientia H
he plant’s ability to access water and nutrients, factors that limitrowth and, thus, yield in many agricultural ecosystems (Lynch,995; Price et al., 1997). Therefore, an enhancement of processeseveloped in roots is one of the most important agronomic traits,aving as final goal the decrease of costs related to irrigation and
ertilization, increasing in this way economic efficiency and cropsrofitability.
During the last decade, treatments with biostimulants based onumic acids (Neri et al., 2002) or chitosan (Górnik et al., 2008),ave been found to be efficient in strawberry and grapevine roots’rowth stimulation, both in well watered and drought conditions.ositive results in alleviation of drought effects have been obtainedt pot-grown strawberry plants treated with ABA (Jensen et al.,009) and paclobutrazol (Navarro et al., 2007), both of them actings antitranspirants, promoting stomatal closure and inducing thectivation of drought avoidance mechanisms. Moreover, anotherlass of compounds has been proved to enhance drought toleranceechanisms by improving osmoregulation. For example, Ambiol (a
ynthetic antioxidant compound) had a significant positive effect inlleviating drought symptoms by manitol accumulation (Roussost al., 2010). Similar results have been previously obtained in plantsreated with seaweed extracts (Blunden et al., 1997).
Plants physiological responses to water deficit are related toolding a positive turgor pressure of the mesophyll cells by con-rol of stomatal aperture in order to decrease transpiration rateBosabalidis and Kofidis, 2002), or by osmotic adjustment thatustained turgor maintenance through accumulation of osmolyteglycinebetaine, sugars, sugar alcohols, proline, manitol, etc.)Serraj and Sinclair, 2002), as well as through reduction of tissuelasticity (Savé et al., 1993) that allows a higher degree of stomatalperture.
In drought conditions, the decrease of the stomatal conductancects as a limitative factor for photosynthesis by limiting CO2 dif-usion in the mesophyll (Cornic and Massacci, 1996). A low levelf internal CO2 (Ci) leads to inhibition of Rubisco or ATP synthe-is (Lawlor and Cornic, 2002) and slows down the oxidation ofADPH, disturbing electron transport between PS II and PS I, whichay cause photoinhibition at PS II reaction centres (Flexas andedrano, 2002), therefore light energy is in excess and can produce
amages to the structures of the photosynthesis apparatus. Plantsdapt to this situation by developing a system through which theyan dissipate this energy through the xanthophyll cycle (Demmig-dams and Adams, 1996). Therefore, several studies have been
ocused on maintaining the efficiency of the specific light use inater stress conditions. Good results in reducing of the drought
ffect, by increasing excitation energy dissipation as heat, haveeen reported by Hu et al., 2013 in an experiment with brassi-osteroids treatments in pepper. The encouraging effects obtainedntil now promote further research in the domain of discoveringew classes of compounds with adaptive effect to stress conditionsspecially in the field of ecological agriculture.
Furostanol glycosides are a large group of steroid compounds,f plant origin, with exhibited anti-fungal (Liu et al., 2003)nd nematicidal activities (Vasil’eva et al., 2005). It has beeneported that furostanol glycosides extracted from Dioscorea del-oidea induce activation of nonspecific defensive responses asnhancement of xanthophyll pigments biosynthesis and increasesf antioxidant enzyme activity, which improved the resistance oflants to biotic stress (Vasil’eva et al., 2005). Furostanol glycosidesxtracted from seeds of Lycopersicon sp. and Capsicum sp., had stim-lative effect on vegetative growth and moreover, have been showno increase the photosynthetic pigment content of grapevine and
pple trees (Munteanu et al., 2008). Our preliminary experimentshow that furostanol glycosides extracted from Lycopersicon sp. andigitalis sp. stimulated root system development and influencedhotosynthetic processes and increased water use efficiency, factlturae 169 (2014) 179–188
that suggests the activation of some adaptive mechanisms towardswater deficit conditions (Caulet et al., 2013).
Therefore, the present study aims at evaluating the influence oftreatments with furostanol glycosides on strawberries’ growth anddevelopment in water stress conditions. Changes in photosynthesiswere studied both in terms of CO2 assimilation rate as well as func-tionality of the photosynthetic apparatus, as assessed by chlorophylla fluorescence measurements.
2. Materials and methods
2.1. Plant material and experimental design
The experiment was conducted at the Faculty of Horticul-ture of Iasi, Romania, during November 2012–March 2013. Twostrawberry cultivars (Fragaria × ananassa Duch.), Real and Magicwere used in this experiment. In order to have the most uni-form biological material we decided to use young daughter plants(runners) with 2 leaves and 5–7 roots (initial plant biomass was2.10–2.54 g). Plants were grown in greenhouse conditions, in 6 Lplastic pots filled with a mixture of soil:sand:litter in (1:1:1, v/v/v).Photosynthetic active radiation (PAR) was 400–500 �mol m2 s−1,relative humidity 52%, temperature 22–23 ◦C and CO2 concentra-tion 380 �mol mol−1.
After two weeks of adaptation to greenhouse conditions, plantswere divided into two groups, well-watered and drought-stressed.Well-watered plants were maintained at 80% WHC (Water holdingcapacity) while drought stress was imposed by withholding waterfrom plants, until soil humidity reached 50% WHC; these conditionshave been maintained for 4 weeks. Each pot was weighed dailyto maintain the desired water levels in soil by adding appropriatevolumes of water.
2.2. Preparation of furostanol glycoside and treatment
The two furostanol glycosides, provided by the BiologicalResearch Institute from Iasi, were extracted and purified from seedsof Lycopersicon sp. – G1 (M = 1082) and leaves of Digitalis sp. – G2(M = 1230). The dry plant material was crushed and extracted with60% EtOH (70–80 ◦C). After concentration under reduced pressure,the ethanol extract was partitioned between water and n-butanol.Chromatographic separations of the organic phase on Sephadex G-50 and silica gel L 540 M gave G1 and G2 compounds respectivelythat were detected with Ehrlich reagent.
The G1 and G2 compounds were eluted with 70% EtOH and werefurther subjected to resin“Dowex-8” and “Amberlite IR-120B toneutralize the acid solution and then evaporated to dryness to givecrude glycoside. Determination of structures of the isolated con-stituents was performed using spectral analysis of IR, 1H and 13CNMR spectra including 2D NMR spectroscopic techniques (COSY,HETCOR and COLOC).
Foliar treatments with furostanol glycosides were applied withaqueous solutions in concentration of 0.03 mM, which have alreadybeen proved to be the optimum for these plants in our preliminaryexperiments. Treatments were applied on mature plants in lateafternoon, with 3 d intervals, using a hand-held sprayer. In orderto avoid interferences with different moisture levels, “untreated”plants were sprayed with distilled water. 10 plants have been usedfor each variant.
2.3. Growth parameters
At the end of the experimental period, newly developed leaves,leaf length, total leaf area, root number/plant and their length, weredetermined and measured.

orticu
u
wrm
2
pdmBcces
2
meMmwadFtsacrr
2
wwwwfw
2
caaTpr1
2
of(
c
R.-P. Caulet et al. / Scientia H
For leaf area measurements, Image J software for Windows wassed.
For the root/shoot ratio determination, the soil was gentlyashed from roots, and the plants were divided into leaves and
oots which were oven dried at 70 ◦C until they reached a constantass.
.4. Gas exchange measurements
Gas exchange and chlorophyll fluorescence measurements wereerformed between 9:00 and 11:00 a.m. on the upper most fullyeveloped leaf of each plant. Gas exchange parameters wereeasured using a portable photosynthetic system (LCi 600, ADC
ioScientific Ltd., England) and net photosynthetic rate (A), intra-ellular CO2 concentration (Ci), transpiration rate (E), and stomatalonductance (gs) were recorded. Water use efficiency (WUE) wasstimated as the quotient of the photosynthetic rate over the tran-piration rate.
.5. Chlorophyll fluorescence measurements
Chlorophyll fluorescence was measured using a portable pulseodulated fluorometer (PAM 2500 from Walz, Germany). Before
ach measurement, leaves were dark-adapted for at least 30 min.inimal fluorescence (F0) was measured under a weak pulse ofodulating light over a 0.8 s period, and maximal fluorescence (Fm)as induced by a saturating pulse of light (8000 �mol m−2 s−1)
pplied over 0.4 s. The maximal quantum efficiency of PSII wasetermined as Fv/Fm, where Fv is the difference between F0 andm. An actinic light source (800 �mol m−2 s−1) was then appliedo achieve steady-state photosynthesis and to obtain Fs (steady-tate fluorescence yield), after which a second saturation pulse waspplied for 0.4 s to obtain F ′
m (light-adapted maximum fluores-ence) and quantum efficiency of PSII (�PSII), electron transportate ETR and non-photochemical quenching coefficient (NPQ) wereecorded.
.6. Determination of RWC
For determination of RWC, 10 leaves/variant were removed,eighed quickly and immediately immersed into double distilledater to saturate them with water for the next 24 h. Then the leavesere oven dried to a constant weight at 70 ◦C and their dry weightas determined. RWC was calculated according to the following
ormula: RWC = (fresh weight − dry weight)/(turgor weight − dryeight).
.7. Photosynthetic pigments’ content determination
Fresh leaves (0.5 g) were homogenized in 80% acetone andentrifuged at 6000 g for 10 min. The supernatant was collectednd the absorbances of the acetone extracts were measuredt 663, 646 and 470 nm using a UV–vis spectrophotometer.he chlorophyll a (Chl a), chlorophyll b (Chl b), total chloro-hylls, total carotenoids (Car) content, Chl a/Chl b and Chl/Caratios were calculated following the equations of Lichtenthaler,987.
.8. Statistical analysis
Results are reported as the mean ± standard error (SE) valuesf three independent experiments. Statistical analyses were per-
ormed with the software Statistical Package for the Social SciencesSPSS) version 20.A one-way analysis of variance (ANOVA) was conducted toompare the effect of glycoside treatments (untreated, G1 and G2
lturae 169 (2014) 179–188 181
variants) on growth parameters and chlorophyll content in wellwatered and draught conditions. Homogeneity of the variances waschecked using Levene tests, before ANOVA.
Where we found a significant effect of glycoside treatment atp < 0.05 significant differences among the treatment means werecalculated by the LSD (least significant difference) test, at p < 0.05.Tests assume equal variances. The overall effects of drought andglycoside application and their interactions on morphological andphysiological parameters were determined by means of a two-wayANOVA at p < 0.05, p < 0.01 and p < 0.001.
3. Results
3.1. Effect of drought stress and furostanol glycosides on plantgrowth parameters
The influence of water deficit on morphological characteris-tics of both strawberry cultivars has been observed rather onthe dimension than the number of roots and leaves. Irrespectiveof cultivar in untreated plants, drought did not affect the roots’number, but a significant increase (p > 0.05) of roots’ length hasbeen determined (Table 1). Moreover, no significant changes inleaves number have been determined in Magic and only a smallreduction (p > 0.05) has been recorded in Real, while leaf size andtotal leaf area were significantly reduced (p > 0.001). As expect-ing, these changes led to a significant increase of root/shoot ratio(Table 1).
Our previous results showed that glycoside treatments havea growth stimulating effect on strawberry plants (Caulet et al.,2013). Present results revealed that even in drought conditionsglycoside treated plants exhibited longer roots in greater numberthan untreated ones, while the number and leaf area were similar(Table 1). Therefore, it can be stated that the drought effect on mor-phologic parameters of treated plants was not significant, exceptthe root/shoot ratio, that increased significantly (p > 0.001) in bothcultivars.
Both glycoside treatments influenced plants growth processesin a similar manner, but the effect was more pronounced in G2treated variants than in G1 treated ones. Significant differenceshave been observed in Magic cultivar, in water stress conditions,when G2 treated plants, exhibited a higher number of roots, longerthan those of G1 treated plants. Likewise, leaf area of G2 treatedvariants has been more reduced in drought conditions, which ledto a significant higher increase of root shoot/ratio compared to G1treated plants.
3.2. Effect of drought stress and furostanol glycosides on gasexchange parameters
The relative water content (RWC) decreased in the leaves ofplants subjected to water deficit. This decrease was similar in bothcultivars and reached to 65% in Real and 63% in Magic after 4 weeksof water stress. Untreated plants cultivated under drought condi-tion had a significant lower photosynthetic activity, compared tothe well watered controls. At minimal RWC values, a pronounceddecrease of net photosynthesis rate up to 55% of control has beenobserved (Fig. 1C). Stomatal conductance and transpiration rateresponses to water deficit follows the same trend, but was less sen-sitive to RWC decrease and as consequence, water use efficiencywas reduced (Fig. 1E).
Both G1 and G2 alleviated drought effect in both cultivars,
but their influence was different in relation to glycoside type.Our results showed that in G1 treated plants RWC was signifi-cantly higher than in G2 treated ones. Although treatment withboth furostanol glycosides had no significant effect on stomatal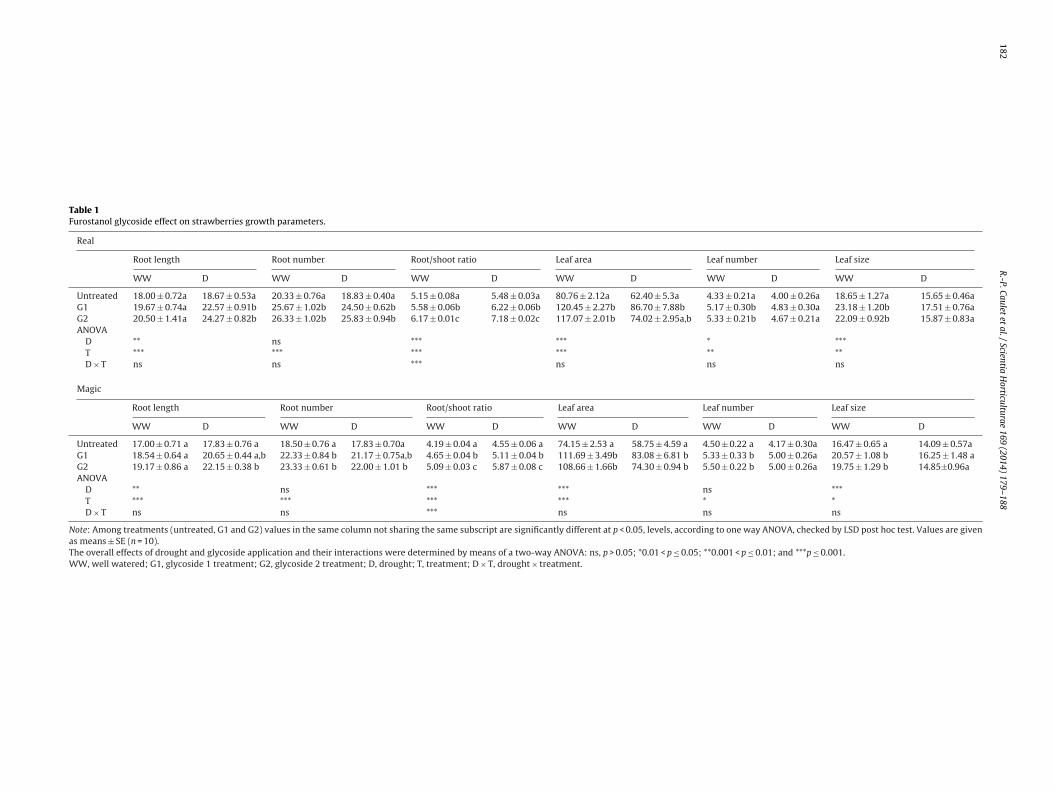
182
R.-P.
Caulet et
al. /
Scientia H
orticulturae 169
(2014) 179–188
Table 1Furostanol glycoside effect on strawberries growth parameters.
Real
Root length Root number Root/shoot ratio Leaf area Leaf number Leaf size
WW D WW D WW D WW D WW D WW D
Untreated 18.00 ± 0.72a 18.67 ± 0.53a 20.33 ± 0.76a 18.83 ± 0.40a 5.15 ± 0.08a 5.48 ± 0.03a 80.76 ± 2.12a 62.40 ± 5.3a 4.33 ± 0.21a 4.00 ± 0.26a 18.65 ± 1.27a 15.65 ± 0.46aG1 19.67 ± 0.74a 22.57 ± 0.91b 25.67 ± 1.02b 24.50 ± 0.62b 5.58 ± 0.06b 6.22 ± 0.06b 120.45 ± 2.27b 86.70 ± 7.88b 5.17 ± 0.30b 4.83 ± 0.30a 23.18 ± 1.20b 17.51 ± 0.76aG2 20.50 ± 1.41a 24.27 ± 0.82b 26.33 ± 1.02b 25.83 ± 0.94b 6.17 ± 0.01c 7.18 ± 0.02c 117.07 ± 2.01b 74.02 ± 2.95a,b 5.33 ± 0.21b 4.67 ± 0.21a 22.09 ± 0.92b 15.87 ± 0.83aANOVA
D ** ns *** *** * ***T *** *** *** *** ** **D × T ns ns *** ns ns ns
Magic
Root length Root number Root/shoot ratio Leaf area Leaf number Leaf size
WW D WW D WW D WW D WW D WW D
Untreated 17.00 ± 0.71 a 17.83 ± 0.76 a 18.50 ± 0.76 a 17.83 ± 0.70a 4.19 ± 0.04 a 4.55 ± 0.06 a 74.15 ± 2.53 a 58.75 ± 4.59 a 4.50 ± 0.22 a 4.17 ± 0.30a 16.47 ± 0.65 a 14.09 ± 0.57aG1 18.54 ± 0.64 a 20.65 ± 0.44 a,b 22.33 ± 0.84 b 21.17 ± 0.75a,b 4.65 ± 0.04 b 5.11 ± 0.04 b 111.69 ± 3.49b 83.08 ± 6.81 b 5.33 ± 0.33 b 5.00 ± 0.26a 20.57 ± 1.08 b 16.25 ± 1.48 aG2 19.17 ± 0.86 a 22.15 ± 0.38 b 23.33 ± 0.61 b 22.00 ± 1.01 b 5.09 ± 0.03 c 5.87 ± 0.08 c 108.66 ± 1.66b 74.30 ± 0.94 b 5.50 ± 0.22 b 5.00 ± 0.26a 19.75 ± 1.29 b 14.85±0.96aANOVA
D ** ns *** *** ns ***T *** *** *** *** * *D × T ns ns *** ns ns ns
Note: Among treatments (untreated, G1 and G2) values in the same column not sharing the same subscript are significantly different at p < 0.05, levels, according to one way ANOVA, checked by LSD post hoc test. Values are givenas means ± SE (n = 10).The overall effects of drought and glycoside application and their interactions were determined by means of a two-way ANOVA: ns, p > 0.05; *0.01 < p ≤ 0.05; **0.001 < p ≤ 0.01; and ***p ≤ 0.001.WW, well watered; G1, glycoside 1 treatment; G2, glycoside 2 treatment; D, drought; T, treatment; D × T, drought × treatment.

R.-P. Caulet et al. / Scientia Horticulturae 169 (2014) 179–188 183
Fig. 1. Influence of furostanol glycosides treatments on gas exchange parameters of strawberry plants under water stress conditions. Different letters above the bars denotestatistically significant differences between treatments (untreated, G1 and G2) at the p < 0.05 level according to one-way ANOVAs, checked by LSD test. Values are given asmeans ± SE (n = 10). WW, well watered; D, drought; Real Untr., real untreated; Real G1, real treated with G1; Real G2, real treated with G2; Magic Untr., magic untreated;M
c0a(
agic G1, magic treated with G1; Magic G2, magic treated with G2.
onductance and transpiration rate, a significant increase (p, > 0.05) of photosynthetic rate in treated plants had been detectednd as consequence, values of WUE were significantly higherp < 0.05) (Fig. 1A–E).
Intercellular CO2 partial pressure (Ci) varied with treatmentsand water regimes. Irrespective of soil humidity, G2 treated plantshad lower Ci values than G1 and untreated variants (Fig. 1F).Drought significantly increased Ci values in untreated and G1

1 orticu
tv
3fl
aw((N
vchcw(iuohqa
3
tarcw
ncsni
tdosi(
ccpi
4
aipRua(
p
84 R.-P. Caulet et al. / Scientia H
reated plants, while a decrease of this parameter in G2 treatedariants has been recorded.
.3. Effect of drought stress and furostanol glycosides onuorescence parameters
The photosynthetic responses to drought of the 3 treatments as measure of quantum yield of PS II are shown in Fig. 2. In well-atered conditions furostanol glycoside treatments significantly
p < 0.05) increased �PS II (Fig. 2C) and electron transfer rate (ETR)Fig. 2B), without any significant changes in Fv/Fm (Fig. 2A), andPQ (Fig. 2D).
In both studied genotypes, drought increased the minimumalues of basal fluorescence F0 and reduced maximum fluores-ence values Fm (data not shown); as a result lower Fv/Fm ratioas been recorded (Fig. 2A). Moreover, quantum yield of PS II effi-iency (�PS II) and ETR decreased in both cultivars simultaneouslyith increment of non-photochemical quenching processes (NPQ)
Fig. 2A–D). Although drought determined �PS II and ETR reductionn both Real and Magic cultivars, significant (p > 0.05) higher val-es of these parameters in treated plants compared with untreatednes were recorded in both cultivars. Likewise, treated plantsad significantly higher values (up to 25%) of non-photochemicaluenching (NPQ), that reflect the higher capacity to dissipate excessbsorbed energy as heat, than untreated ones (Fig. 2D).
.4. Chlorophyll and carotenoid contents
Drought determined a significant reduction of Chl a, Chl b andotal Chl (p < 0.001) in both cultivars (Table 2). However, no alter-tion of Chl a/Chl b ratio was recorded, indicating that water deficiteduced Chl a and Chl b to similar extent. Also, total carotenoid con-entration was significantly lower in both studied genotypes underater stress conditions.
Although, in well watered conditions glycoside treatments hado significant effect on pigment content, a significant increases ofhlorophyll content (p > 0.001) (both Chl a and Chl b) under watertress having been determined in both cultivars. However, a sig-ificant decrease of Chl a/Chl b ratio was observed due to greater
ncrease of Chl b compared to Chl a.On the other way, in glycoside treated variants, carotenoid con-
ent was higher irrespective of water regime, leading to significantecreases in Chl/Car ratio. Moreover, drought induced reductionf carotenoid content (up to 20% in untreated plants) had beentrongly alleviated in glycoside treated plants, reaching up to 10%n G2 treated ones therefore Chl/Car ratio was strongly reducedp < 0.001).
Both G1 and G2 had the same pattern in chlorophyll contenthanges during the drought period, but in G1 treated variantsarotenoid content was lower compared to that of G2 treatedlants. As a result, significant decrease (p < 0.001) on Chl/Car ratio
n stress condition was observed only in G2 treated plants.
. Discussion
Drought can reduce plant productivity by inhibiting growthnd photosynthesis (Taiz and Zieger, 1998). In our experiments,n water stress conditions, root/shoot ratio increased in untreatedlants as a consequence of leaf area reduction, appearing stronger ateal cultivar than at Magic. The smaller leaf area of plants cultivatednder drought conditions is considered a drought avoidance mech-
nism, which leads to a better control of water loss by transpirationAranda et al., 2005).Furostanol glycoside treatments act like stimulators for growthrocesses, enhancing most of the growth parameters. Similar
lturae 169 (2014) 179–188
effects have been reported in experiments with different biostimu-lants at strawberry (Marfà et al., 2009; Abdel-Mawgoud et al., 2010)lettuce (Kowalczyk et al., 2008; Ertani et al., 2009) and grapevine(Mancuso et al., 2006; Górnik et al., 2008). Besides that glycosidetreatments also determined a significant increase of root/shootratio, due to both the decrease of leaf area as well as stimulationof roots’ growth (Caulet et al., 2013). An increase of investmentin roots results in enhancement of root depth, providing greaterwater uptake, which is crucial for plant survival during drought(Klamkowski and Treder, 2008), fact that led us to believe that thefurostanol glycoside treatments might enhance plants adaptationto water stress.
In our experiment, it was shown that in drought conditionsglycoside treatments provide a higher biomass accumulation inroots, which leads to an increase of root/shoot ratio. This effectwas greater in G2 treated plants than in G1 treated ones. Increas-ing of root/shoot ratio represents a general adaptive strategy ofplants to the water deficit in soil (Buwalda and Lenz, 1992), but ithas been also reported in different experiments with antitranspi-rant substances like paclobutrazol (Chaturvedi et al., 2009). Thissimilarity may suggest an adaptive effect induced by furostanolglycoside treatment on photosynthesis by stomatal closure mod-ulation mechanism. In line with these findings, in well wateredconditions, a slight decrease in photosynthetic rate at glycosidetreated plants compared to untreated ones was observed, fact thatmight be due to a decrease of stomatal conductance and limitationof CO2 diffusion in cells’ mesophyll (Chaves et al., 2003). How-ever, in these variants, higher water use efficiency (A/E) has beendetermined. Greater reduction of transpiration water loss than CO2assimilation rates has been found in experiments with antitranspi-rant compounds at Phaseolus vulgaris L. plants (Iriti et al., 2009).However in G2 treated variants, changes in A, E and gs have beenaccompanied by a slight reduction of internal CO2 concentration.Besides, glycosides treated plants exhibited a greater �PS II andETR, without any significant changes in NPQ, suggesting an increaseof light use efficiency and capacity of CO2 assimilation in Calvincycle (Yu et al., 2004). This can indicate that only stomatal closurelimits the photosynthesis without other changes in the energeticsof photosynthesis process (Flexas and Medrano, 2002).
In our experiments in water stress conditions photosynthesisrate and stomatal conductance decreased significantly (p < 0.01)at untreated plants. Stomatal closure is the first plant response towater limitation that influences gas exchange process. In these con-ditions in both cultivars, a decrease of Fv/Fm and PS II efficiency (�PSII) was recorded. As a result of these decrease, energy productionrate exceeds the rate of its use in photosynthetic electron trans-port chain, thus can damage the reaction centres of PS II (Sandaet al., 2011; Wu and Bao, 2011). In response to these changes,non-photochemical processes (NPQ) were enhanced in order toprevent photodamages of PS II reaction centres through processesof thermal deactivation, downregulating PSII photochemistry, in aso-called dynamic photoinhibition (Osmond, 1994), that brings theelectron transport capacity into balance with carbon metabolism(Epron et al., 1995).
Chlorophyll content was reduced under drought stress in bothcultivars. A reduction in chlorophyll content was reported indrought stressed cotton (Massacci et al., 2008), Catharanthus roseus(Jaleel et al., 2008), sunflower plants (Kiani et al., 2008) and inVaccinium myrtillus (Tahkokorpi et al., 2007).
Both furostanol glycoside treatments induced an increase ofplants adaptability to drought, but this effect has been displayedby two different mechanisms. Our data suggest that G1 treated
plants responded to drought by inducing physiological acclimationresponses, including increased stomata responsiveness (conser-vative manner), thus maintaining a higher RWC than untreatedplants. Increased stomatal responsiveness results in more rapid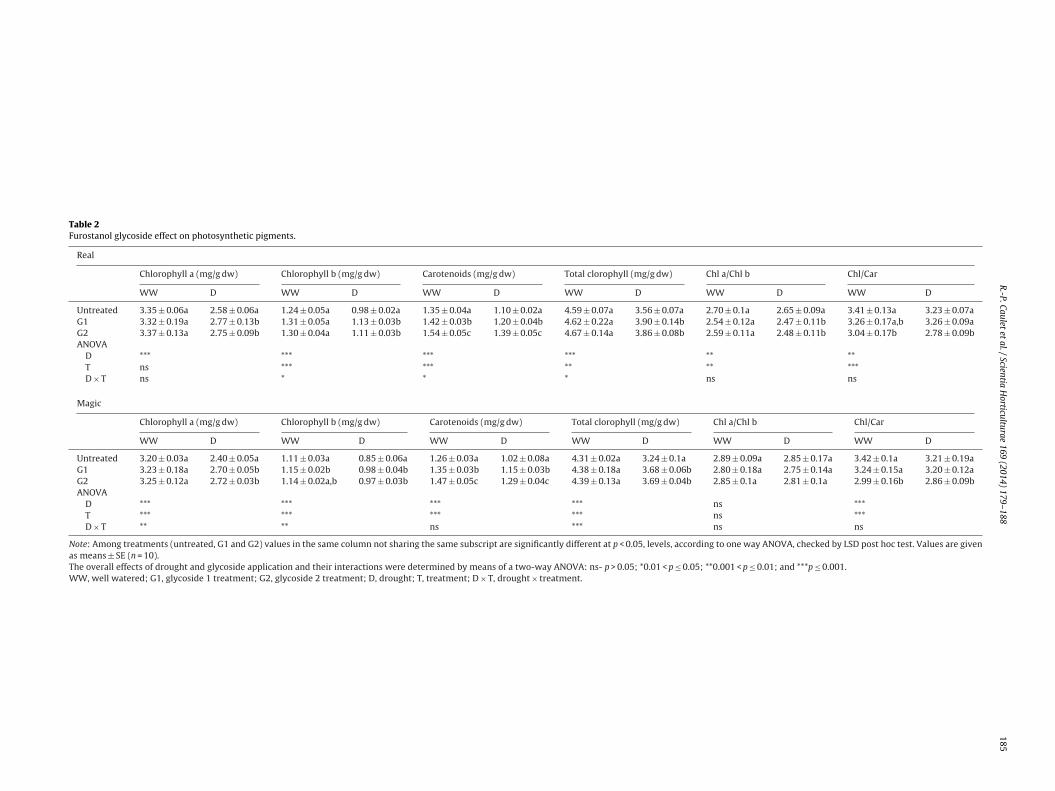
R.-P.
Caulet et
al. /
Scientia H
orticulturae 169
(2014) 179–188
185
Table 2Furostanol glycoside effect on photosynthetic pigments.
Real
Chlorophyll a (mg/g dw) Chlorophyll b (mg/g dw) Carotenoids (mg/g dw) Total clorophyll (mg/g dw) Chl a/Chl b Chl/Car
WW D WW D WW D WW D WW D WW D
Untreated 3.35 ± 0.06a 2.58 ± 0.06a 1.24 ± 0.05a 0.98 ± 0.02a 1.35 ± 0.04a 1.10 ± 0.02a 4.59 ± 0.07a 3.56 ± 0.07a 2.70 ± 0.1a 2.65 ± 0.09a 3.41 ± 0.13a 3.23 ± 0.07aG1 3.32 ± 0.19a 2.77 ± 0.13b 1.31 ± 0.05a 1.13 ± 0.03b 1.42 ± 0.03b 1.20 ± 0.04b 4.62 ± 0.22a 3.90 ± 0.14b 2.54 ± 0.12a 2.47 ± 0.11b 3.26 ± 0.17a,b 3.26 ± 0.09aG2 3.37 ± 0.13a 2.75 ± 0.09b 1.30 ± 0.04a 1.11 ± 0.03b 1.54 ± 0.05c 1.39 ± 0.05c 4.67 ± 0.14a 3.86 ± 0.08b 2.59 ± 0.11a 2.48 ± 0.11b 3.04 ± 0.17b 2.78 ± 0.09bANOVA
D *** *** *** *** ** **T ns *** *** ** ** ***D × T ns * * * ns ns
Magic
Chlorophyll a (mg/g dw) Chlorophyll b (mg/g dw) Carotenoids (mg/g dw) Total clorophyll (mg/g dw) Chl a/Chl b Chl/Car
WW D WW D WW D WW D WW D WW D
Untreated 3.20 ± 0.03a 2.40 ± 0.05a 1.11 ± 0.03a 0.85 ± 0.06a 1.26 ± 0.03a 1.02 ± 0.08a 4.31 ± 0.02a 3.24 ± 0.1a 2.89 ± 0.09a 2.85 ± 0.17a 3.42 ± 0.1a 3.21 ± 0.19aG1 3.23 ± 0.18a 2.70 ± 0.05b 1.15 ± 0.02b 0.98 ± 0.04b 1.35 ± 0.03b 1.15 ± 0.03b 4.38 ± 0.18a 3.68 ± 0.06b 2.80 ± 0.18a 2.75 ± 0.14a 3.24 ± 0.15a 3.20 ± 0.12aG2 3.25 ± 0.12a 2.72 ± 0.03b 1.14 ± 0.02a,b 0.97 ± 0.03b 1.47 ± 0.05c 1.29 ± 0.04c 4.39 ± 0.13a 3.69 ± 0.04b 2.85 ± 0.1a 2.81 ± 0.1a 2.99 ± 0.16b 2.86 ± 0.09bANOVA
D *** *** *** *** ns ***T *** *** *** *** ns ***D × T ** ** ns *** ns ns
Note: Among treatments (untreated, G1 and G2) values in the same column not sharing the same subscript are significantly different at p < 0.05, levels, according to one way ANOVA, checked by LSD post hoc test. Values are givenas means ± SE (n = 10).The overall effects of drought and glycoside application and their interactions were determined by means of a two-way ANOVA: ns- p > 0.05; *0.01 < p ≤ 0.05; **0.001 < p ≤ 0.01; and ***p ≤ 0.001.WW, well watered; G1, glycoside 1 treatment; G2, glycoside 2 treatment; D, drought; T, treatment; D × T, drought × treatment.

186 R.-P. Caulet et al. / Scientia Horticulturae 169 (2014) 179–188
Fig. 2. Influence of furostanol glycosides treatments on chlorophyll a fluorescence parameters of strawberry plants under water stress conditions. Different letters above theb and Ga treateu
slwirltpf
ds1tttrtbst
ars denote statistically significant differences between treatments (untreated, G1re given as means ± SE (n = 10). WW, well watered; D, drought; Real Untr., real unntreated; Magic G1, magic treated with G1; Magic G2, magic treated with G2.
tomatal closure once the leaf water potential falls below certainevels (Jones, 1993). On the other way this treatment improved
ater use efficiency. This can be the effect of the unequal alterationsnduced by water stress in the photosynthetic and transpirationates. A more efficient photosynthetic system is able to function onower concentrations of intercellular CO2 (a consequence of stoma-al closure) and is able to deal with energy excess without essentialroteins or enzymes being damaged when photosynthesis is notunctioning at its maximum (Silim et al., 2001).
Concentration of intercellular CO2 (Ci) is a reliable indicator toiscriminate the changes in the net photosynthetic rate betweentomatal and non-stomatal limitation (Farquhar and Sharkey,982). One of the reasons for the slight increase in Ci at the G1reated plants could be the increased mesophyll resistance for CO2ransport. However, G1 treated plants reach a higher rate of pho-osynthesis at high Ci, thus reflecting the capacity of the leaves toegenerate RuBP, which is associated with improvement of elec-
ron transport activity (Lawlor and Cornic, 2002). Therefore it cane concluded that G1 treatment induces also the activation of non-tomatal mechanisms which accelerates the electron transport andhe efficiency of light absorption.2) at the p < 0.05 level according to one-way ANOVAs, checked by LSD test. Valuesd; Real G1, real treated with G1; Real G2, real treated with G2; Magic Untr., magic
In the glycoside treated plants the reduction of Chla/Chl b ratiowas also observed due to an increase in Chl b contents. Chl a/Chlb ratio is inversely related to the antenna size (Tanaka et al.,2001) that indicate a stimulative effect of this compound on light-harvesting complex, which in turn could maintain a certain levelof photosynthetic efficiency under water deficit (Lawlor and Keys,1993).
In G2 treated plants, a productive strategy of adaptation todrought stress was observed, characterized by maintenance of ahigher transpiration rate and stomatal conductance that allowedCO2 diffusion in mesophyll at lower RWC. This glycoside treat-ment seems to increase plants tolerance to dehydration. Althoughtranspirational water losses were higher than in G1 treated plants,leading to decrease of RWC, water use efficiency increased dueto enhancement of photosynthetic rate. Tissue tolerance to dehy-dration may be achieved by accumulation of compatible solutes,which preserve cellular turgor and maintain metabolic activity dur-
ing water deficits. However, this adaptation is not common in mostof higher plants including crops (Ingram and Bartels, 1996), but itevolved in species with high tolerance to drought like Dygitalis sp.(raw material for G2 glycoside extract) (Galmés et al., 2007). Many
orticu
stwpchta
dniyoO
meawlePettestiIcswp
5
lashnicitwtdtnd
GdcTpt
A
f
R.-P. Caulet et al. / Scientia H
tudies indicated that solute accumulation under water stress con-ributes to inhibition of shoot growth (Serraj and Sinclair, 2002),hile the current experiments demonstrated that in G2 treatedlants the decrease in leaves’ area and number was greater than inontrol or G1 treated variants. The above data are premises for theypothesis that G2 treatments could have the effect of stimulatinghe adaptation of plants to stress by inducing osmotic adjustmentt foliar tissues level.
Despite the drought, in G2 treated variants photosynthesis rateecreased only marginally, being connected with the low inter-al CO2 concentration. The majority of authors agree that the
nitial slope of the CO2 curve is defined by the maximal carbox-lation efficiency of Rubisco and these results suggest an increasef the activation state of photosynthetic enzymes (Ali et al., 2008;gweno et al., 2008).
Particular importance is attached to plant photoprotectionechanisms, which must be activated in drought conditions when
nergy capture by chlorophyll may exceed energy used in CO2ssimilation. The above fact leads to increases in the ROS generationith consequent adverse effects on photosynthetic and other cellu-
ar mechanisms, for example photoinhibition of photosystems (Shit al., 2007) and damage to ATP synthase (Lawlor and Tezara, 2009).rotective and regulatory mechanisms include those that increasenergy dissipation and regulate the energy balance of cells (e.g.he xanthophyll cycle in photosynthetic tissues) and those preven-ing production of – or enhancing breakdown of – ROS (Melchiorret al., 2009). In G2 treated variants, the increase of NPQ in watertressed plants was more pronounced than that recorded at G1reated ones, allowing a higher dissipation of energy, protectingn this way PSII reaction centres and maintaining ETR and �PSI at high levels. Moreover, in these variants a higher carotenoidontent was determined and their major protective role in photo-ynthetic tissue through direct quenching of triplet chlorophyll isell known, which prevents the generation of singlet oxygen androtects from oxidative damage (Farooq et al., 2009).
. Conclusions
Glycosides treatment is effective in growth stimulation of botheaves and roots system in the strawberry plants. Overall, foliarpplication of furostanol glycosides (especially of G2) leads totimulation of root system development, providing plants with aigher adaptability to environmental conditions. Although exoge-ous application of furostanol glycosides caused slight decrease
n photosynthetic rate, it significantly increases water use effi-iency (A/E) in both cultivars. Both furostanol glycoside treatmentsmproved plants adaptability to drought. Our results demonstratehe ability of G1 compound to increase stomata responsiveness,hich determined a decrease of transpiration rate, and main-
ained higher WUE which can help delay the adverse effects ofrought thus allowing the stressed plants to continue photosyn-hesis under drought conditions. This treatment could activateon-stomatal mechanisms that increase photosynthetic antennaimension, electron transport and efficiency of light absorption.
On the other way, the amelioration of drought effects by2 compound can be ascribed to increasing plants’ tolerance toehydration by maintaining stomata opened and enhancing theirapacity to sustain carbon assimilation under drought condition.his treatment induced higher dissipation of energy and an increaseroduction of carotenoids, protecting in this way PSII reaction cen-res and maintaining ETR and �PS II at high levels.
cknowledgement
The authors would like to thank the Biological Research Instituteor providing the glycosides extracts.
lturae 169 (2014) 179–188 187
References
Abdel-Mawgoud, A.M.R., Tantawy, A.S., El-Nemr, M.A., Sassine, Y.N., 2010. Growthand yield responses of strawberry plants to Chitosan application. Eur. J. Sci. Res.39 (1), 170–177.
Ali, B., Hasan, S.A., Hayat, S.Q., Yadav, S., Fariduddin, Q., Ahmad, A., 2008. A role forbrassinosteroids in the amelioration of aluminium stress through antioxidantsystem in mung bean (Vigna radiata L. Wilczek). Environ. Exp. Bot. 62, 153–159.
Aranda, I., Castro, L., Pardos, M., Gil, L., Pardos, J.A., 2005. Effects of the interactionbetween drought and shade on water relations, gas exchange and morphologicaltraits in cork oak (Quercus suber L.) seedlings. For. Ecol. Manage. 210, 117–129.
Blunden, G., Jenkins, T., Liu, Y., 1997. Enhanced leaf chlorophyll levels in plantstreated with seaweed extract. J. Appl. Phycol. 8, 535–543.
Bosabalidis, A.M., Kofidis, G., 2002. Comparative effects of drought stress on leafanatomy of two olive cultivars. Plant Sci. 163, 375–379.
Buwalda, J.G., Lenz, F., 1992. Effects of cropping, nutrition and water supply on accu-mulation and distribution of biomass and nutrients for apple trees on ‘M9’ rootsystems. Physiol. Plant. 84, 21–28.
Caulet, R.P., Morariu, A., Iurea, D., Gradinariu, G., 2013. Growth and photosyntheticcharacteristics of two strawberry cultivars in response to furostanol glycosidestreatments. Not. Bot. Horti Agrobot. 41 (1), 172–178.
Chaturvedi, A.K., Rajiv, K., Vashistha, A., Prasad, P., Nautiyal, M.C., 2009. Influenceof foliar spray with paclobutrazol and ethepon on growth and photosyntheticbehavior of Saussurea costus (Falc.) Lipsch. – an endangered medicinal and aro-matic herb. Nat. Sci. 7 (8), 53–62.
Chaves, M.M., Maroco, P., João, P., João, S., 2003. Understanding plant responses todrought – from genes to the whole plant. Funct. Plant Biol. 30, 239–264.
Cornic, G., Massacci, A., 1996. Leaf photosynthesis under drought stress. In: Baker, N.(Ed.), Photosynthesis and the Environment. Kluwer Academic Publishers, NewYork, pp. 347–366.
Demmig-Adams, B., Adams, W.W., 1996. The role of xanthophyll cycle carotenoidsin the protection of photosynthesis. Trends Plant Sci. 1, 21–26.
Epron, D., Godard, D., Cornic, G., Genty, B., 1995. Limitation of net CO2 assimilationrate by internal resistances to CO2 transfer in the leaves of two tree species(Fagus sylvatica L. and Castanea sativa Mill.). Plant Cell Environ. 18, 43–51.
Ertani, A., Cavani, L., Pizzeghello, D., Brandellero, E., Altissimo, A., Ciavatta, C., Nardi,S., 2009. Biostimulant activity of two protein hydrolysates in the growth andnitrogen metabolism of maize seedlings. J. Plant Nutr. Soil Sci. 172, 237–244.
Farooq, M., Wahid, A., Kobayashi, N., Fujita, D., Basra, S.M.A., 2009. Plant droughtstress: effects, mechanisms and management. Agron. Sustain. Dev. 29, 153–188.
Farquhar, G.D., Sharkey, T.D., 1982. Stomatal conductance and photosynthesis. J.Annu. Rev. Plant Physiol. 33, 317–345.
Flexas, J., Medrano, H., 2002. Energy dissipation in C3 plants under drought. Funct.Plant Biol. 29, 1209–1215.
Galmés, J., Medrano, H., Flexas, J., 2007. Photosynthesis and photoinhibition inresponse to drought in a pubescent (var. minor) and a glabrous (var. palaui)variety of Digitalis minor. Environ. Exp. Bot. 60, 105–111.
Górnik, K., Mieczysław, G., Romanowska-Duda, B., 2008. The effect of chitosan onrooting of grapevine cuttings and on subsequent plant growth under droughtand temperature stress. J. Fruit Ornam. Plant Res. 16, 333–343.
Hu, W.H., Yan, X.H., Xiao, Y.A., Zeng, J.J., Qi, H.J., Ogweno, J.O., 2013. 24-Epibrassinosteroid alleviate drought-induced inhibition of photosynthesis inCapsicum annuum. Sci. Hort. 150, 232–237.
Ingram, J., Bartels, D., 1996. The molecular basis of dehydration tolerance in plants.Ann. Rev. Plant Physiol. Plant Mol. Biol. 47, 377–403.
Iriti, M., Picchib, V., Rossonia, M., Gomarascac, S., Ludwigd, N., Garganod, M., Faoro,F., 2009. Chitosan antitranspirant activity is due to abscisic acid-dependent sto-matal closure. Environ. Exp. Bot. 66, 493–500.
Jackson, R.B., Sperry, J.S., Dawson, T.E., 2000. Root water uptake and transport: usingphysiological processes in global predictions. Trends Plant Sci. 5, 482–488.
Jaleel, C.A., Manivannan, P., Lakshmanan, G.M.A., Gomathinayagam, M., Panneer-selvam, R., 2008. Alterations in morphological parameters and photosyntheticpigment responses of Catharanthus roseus under soil water deficits. Colloids Surf.B: Biointerfaces 61, 298–303.
Jensen, N.L., Jensen, C.R., Liu, F., Petersen, K.K., 2009. Water relations and abscisicacid in pot-grown strawberry plants under limited irrigation. J. Am. Soc. Hort.Sci. 134 (5), 574–580.
Jones, H.G., 1993. Drought tolerance and water-use efficiency. In: Smith, J.A.C., Grif-fiths, H. (Eds.), Water Deficits: Plant Responses from Cell to Community. BIOSScientific Publishers, Oxford, UK, pp. 193–203.
Kiani, S.P., Maury, P., Sarrafi, A., Grieu, P., 2008. QTL analysis of chlorophyll fluores-cence parameters in sunflower (Helianthus annuus L.) under well-watered andwater-stressed conditions. Plant Sci. 175, 565–573.
Klamkowski, K., Treder, W., 2008. Response to drought stress of three strawberrycultivars grown under greenhouse conditions. J. Fruit Ornam. Plant Res. 16,179–188.
Kowalczyk, K., Zielony, T., Gajewski, M., 2008. Effect of Aminoplant and Asahion yield and quality of lettuce grown on rockwool. In: Dabrowski, Z.T. (Ed.),Biostimulators in Modern Agriculture. WiesJutra, Warszawa, pp. 35–43.
Lawlor, D.W., Cornic, G., 2002. Photosynthetic carbon assimilation and associatedmetabolism in relation to water deficits in higher plants. Plant Cell Environ. 25,
275–294.Lawlor, D.W., Keys, A.J., 1993. Understanding photosynthetic adaptation to changeclimate. In: Fowden, L., Manseld, T., Stoddart, J. (Eds.), Plant Adaptation to Envi-ronment Stress. Chapman and Hall, London, pp. 85–100.

1 orticu
L
L
L
L
LM
M
M
M
M
N
N
O
O
88 R.-P. Caulet et al. / Scientia H
awlor, D.W., Tezara, W., 2009. Causes of decreased photosynthetic rate andmetabolic capacity in water-deficient leaf cells: a critical evaluation of mecha-nisms and integration of processes. Ann. Bot. 103, 561–579.
ichtenthaler, H.K., 1987. Chlorophylls and carotenoids: pigments of photosyntheticmembranes. Methods Enzymol. 148, 350–382.
iu, H.W., Wang, S.L., Cai, B., Qu, G.X., Yang, X.J., Kobayashi, H., Yao, X.S., 2003. Newfurostanol glycosides from the rhizomes of Dioscorea futschauensis R. Kunth. J.Asian Nat. Prod. Res. 4, 241–247.
ópez-Bucio, J., Cruz-Ramírez, A., Herrera-Estrella, L., 2003. The role of nutrientavailability in regulating root architecture. Curr. Opin. Plant Biol. 6, 280–287.
ynch, J., 1995. Root architecture and plant productivity. Plant Physiol. 109, 7–13.ancuso, S., Azzarello, E., Mugnai, S., Briand, X., 2006. Marine bioactive substances
(IPA extract) improve ion fluxes and water stress tolerance in potted Vitis viniferaplants. Adv. Hort. Sci. 20, 156–161.
arfà, O., Cáceres, R., Polo, J., Ródenas, J., 2009. Animal protein hydrolysate as abiostimulant for transplanted strawberry plants subjected to cold stress. ActaHort. 842, 315–318.
assacci, A., Nabiev, S.M., Pietrosanti, L., Nematov, S.K., Chernikova, T.N., Thor, K.,Leipner, J., 2008. Response of the photosynthetic apparatus of cotton (Gossypiumhirsutum) to the onset of drought stress under field conditions studied by gas-exchange analysis and chlorophyll fluorescence imaging. Plant Physiol. Biochem.46 (2), 189–195.
elchiorre, M., Robert, G., Trippi, V., Racca, R., Lascano, H.R., 2009. Superoxide dis-mutase and glutathione reductase overexpression in wheat protoplast: changesin cellular redox state and photooxidative stress tolerance. Plant Growth Regul.57 (1), 57–68.
unteanu, N., Iurea, D., Mustea, M., 2008. Improving the vine crop technolo-gies by using glycoside substances, under conditions of economic efficiencyand environmental protection. Cercetari Agronomice Moldova 63 (3), 135–141.
avarro, A., Sanchez-Blanco, M.J., Banõn, S., 2007. Influence of paclobutrazol onwater consumption and plant performance of Arbutus unedo seedlings. Sci. Hort.111 (2), 133–139.
eri, D., Lodolini, E.M., Savini, G., Sabbatici, P., Bonanomi, G., Zucconi, F., 2002.Foliar application of humic acids on strawberry (cv. Onda). Acta Hort. 594,297–302.
gweno, J.O., Song, X.S., Shi, K., Hu, W.H., Mao, W.H., Zhou, Y.H., Yu, J.Q., Nogues,S., 2008. Brassinosteroids alleviate heat-induced inhibition of photosynthesisby increasing carboxylation efficiency and enhancing antioxidant systems in
Lycopersicon esculentum. J. Plant Growth Regul. 27 (1), 49–57.smond, C.B., 1994. What is photoinhibition? Some insights from comparisons ofshade and sun plants. In: Baker, N.R., Bowyer, J.R. (Eds.), Photoinhibition of Pho-tosynthesis: From Molecular Mechanisms to the Field. BIOS Scientific Publishers,Oxford, pp. 1–24.
lturae 169 (2014) 179–188
Price, A.H., Tomos, A.D., Virk, D.S., 1997. Genetic dissection of root growth in rice(Oryza sativa L.). I. A hydroponic screen. Theor. Appl. Genet. 95, 132–142.
Roussos, A.P., Denaxa, N.K., Damvakaris, T., Stournaras, V., Argyrokastritis, I., 2010.Effect of alleviating products with different mode of action on physiology andyield of olive under drought. Sci. Hort. 125, 700–711.
Sanda, S., Yoshida, K., Kuwano, M., Kawamura, T., Munekage, Y.N., Akashi, K., Yokota,A., 2011. Responses of the photosynthetic electron transport system to excesslight energy caused by water deficit in wild watermelon. Physiol. Plant. 142,247–264.
Savé, R., Penuelas, J., Marfá, O., Serrano, L., 1993. Changes in leaf osmotic and elasticproperties and canopy structure of strawberries under mild water stress. Hort.Sci. 28, 925–927.
Serraj, R., Sinclair, T.R., 2002. Osmolyte accumulation: can it really help increase cropyield under drought conditions? Plant Cell Environ. 25 (2), 333–341.
Shi, Q., Ding, F., Wang, X., Wei, M., 2007. Exogenous nitric oxide protects cucumberroots against oxidative stress induced by salt stress. Plant Physiol. Biochem. 45,542–550.
Silim, S.N., Guy, R.D., Patterson, T.B., Livingston, N.J., 2001. Plasticity in water-useefficiency of Picea sitchensis, P. glauca and their natural hybrids. Oecologia 128,317–325.
Singer, S.M., Helmy, Y.I., Karas, A.N., Abou-Hadid, A.F., 2003. Influences of differentwater stress treatments on growth, development and production of snap bean(Phaseolus vulgaris L.). Acta Hort. 14, 605–611.
Starck, R.L., Wenerick, R.W., Williams, F.M., Wuest, J.P., 1995. The effects of pH,flow rate, and carbon supplementation on manganese retention in mesocosmwetlands. J. Environ. Qual. 24, 816–826.
Tahkokorpi, M., Taulavuori, K., Laine, K., Taulavuori, E., 2007. After-effects of droughtrelated winter stress in previous and current year stems of Vaccinium myrtillusL. Environ. Exp. Bot. 61, 85–93.
Taiz, L., Zieger, E., 1998. Stress physiology. In: Taiz, L., Zieger, E. (Eds.), Plant Physi-ology. , 2nd ed. Sinauer Associates, Inc., Sunderland, MA, pp. 725–757.
Tanaka, R., Koshino, Y., Sawa, S., Ishiguro, S., Okada, K., Tanaka, A., 2001. Over-expression of chlorophyllide a oxygenase (CAO) enlarges the antenna size ofphotosystem II in Arabidopsis thaliana. Plant J. 26, 365–373.
Vasil’eva, I.S., Vanyushkin, S.A., Zinov’eva, S.V., Udalova Zh, V., Volkova, L.A., Nosov,A.M., Paseshnichenko, V.A., 2005. Adaptogenic effects of furostanol glycosidesof Dioscorea deltoidea wall on oxidative processes in tomato plants during bioticstress. Appl. Biochem. Microbiol. 41 (3), 306–311.
Wu, X.L., Bao, W.K., 2011. Leaf growth, gas exchange and chlorophyll fluorescence
parameters in response to different water deficits in wheat cultivars. Plant Prod.Sci. 14, 254–259.Yu, J.Q., Huang, L.F., Hu, W.H., Zhou, Y.H., Mao, W.H., Ye, S.F., Nogués, S., 2004. A roleof brassinosteroids in the regulation of photosynthesis in Cucumis sativus. J. Exp.Bot. 55, 1135–1143.




















