
Cyanogenic glycosides: a case study for evolution and application of cytochromes P450
21
Abstract Cyanogenic glycosides are ancient bio- molecules found in more than 2,650 higher plant species as well as in a few arthropod species. Cyanogenic glycosides are amino acid-derived b-glycosides of a-hydroxynitriles. In analogy to cyanogenic plants, cyanogenic arthropods may use cyanogenic glycosides as defence compounds. Many of these arthropod species have been shown to de novo synthesize cyanogenic glycosides by biochemical pathways that involve identical intermediates to those known from plants, while the ability to sequester cyanogenic glycosides appears to be restricted to Lepidopteran species. In plants, two atypical multifunctional cyto- chromes P450 and a soluble family 1 glycosyl- transferase form a metabolon to facilitate channelling of the otherwise toxic and reactive intermediates to the end product in the pathway, the cyanogenic glycoside. The glucosinolate path- way present in Brassicales and the pathway for cyanoalk(en)yl glucoside synthesis such as rhod- iocyanosides A and D in Lotus japonicus exem- plify how cytochromes P450 in the course of evolution may be recruited for novel pathways. The use of metabolic engineering using cyto- chromes P450 involved in biosynthesis of cyano- genic glycosides allows for the generation of acyanogenic cassava plants or cyanogenic Arabid- opsis thaliana plants as well as L. japonicus and A. thaliana plants with altered cyanogenic, cyanoalkenyl or glucosinolate profiles. Keywords Metabolic engineering Metabolons CYP79 Systems biology Plant–insect interactions Cyanogenic glycosides are ancient biomolecules Cyanogenic glycosides (A1) are amino acid- derived b-glycosides of a-hydroxynitriles. They have been found in more than 2,650 higher plant S. Bak (&) M. Morant A. V. Morant S. Saito N. Bjarnholt M. Zagrobelny K. Jørgensen S. Osmani H. T. Simonsen R. S. Pe ´ rez T. B. van Heeswijck B. L. Møller Plant Biochemistry Laboratory, Department of Plant Biology, and Center for Molecular Plant Physiology (PlaCe), Royal Veterinary and Agricultural University, 40 Thorvaldsensvej, 1871 Frederiksberg Copenhagen, Denmark e-mail: [email protected] S. M. Paquette Department of Biological Structure, University of Washington, HSB G-514/Box 357420, Seattle, WA 98195-7420, USA B. Jørgensen Research Group Cell Wall Biology and Molecular Virology, Department of Genetics and Biotechnology, Danish Institute of Agricultural Sciences, 40 Thorvaldsensvej, 1871 Frederiksberg Copenhagen, Denmark Phytochem Rev (2006) 5:309–329 DOI 10.1007/s11101-006-9033-1 123 Cyanogenic glycosides: a case study for evolution and application of cytochromes P450 Søren Bak Susanne Michelle Paquette Marc Morant Anne Vinther Morant Shigeki Saito Nanna Bjarnholt Mika Zagrobelny Kirsten Jørgensen Sarah Osmani Henrik Toft Simonsen Raquel Sanchez Pe ´rez Torbjørn Bordier van Heeswijck Bodil Jørgensen Birger Lindberg Møller Received: 24 May 2006 / Accepted: 22 September 2006 / Published online: 15 November 2006 Ó Springer Science+Business Media B.V. 2006
Transcript of Cyanogenic glycosides: a case study for evolution and application of cytochromes P450

Abstract Cyanogenic glycosides are ancient bio-
molecules found in more than 2,650 higher plant
species as well as in a few arthropod species.
Cyanogenic glycosides are amino acid-derived
b-glycosides of a-hydroxynitriles. In analogy to
cyanogenic plants, cyanogenic arthropods may use
cyanogenic glycosides as defence compounds.
Many of these arthropod species have been shown
to de novo synthesize cyanogenic glycosides by
biochemical pathways that involve identical
intermediates to those known from plants, while
the ability to sequester cyanogenic glycosides
appears to be restricted to Lepidopteran species.
In plants, two atypical multifunctional cyto-
chromes P450 and a soluble family 1 glycosyl-
transferase form a metabolon to facilitate
channelling of the otherwise toxic and reactive
intermediates to the end product in the pathway,
the cyanogenic glycoside. The glucosinolate path-
way present in Brassicales and the pathway for
cyanoalk(en)yl glucoside synthesis such as rhod-
iocyanosides A and D in Lotus japonicus exem-
plify how cytochromes P450 in the course of
evolution may be recruited for novel pathways.
The use of metabolic engineering using cyto-
chromes P450 involved in biosynthesis of cyano-
genic glycosides allows for the generation of
acyanogenic cassava plants or cyanogenic Arabid-
opsis thaliana plants as well as L. japonicus and
A. thaliana plants with altered cyanogenic,
cyanoalkenyl or glucosinolate profiles.
Keywords Metabolic engineering ÆMetabolons Æ CYP79 Æ Systems biology ÆPlant–insect interactions
Cyanogenic glycosides are ancient biomolecules
Cyanogenic glycosides (A1) are amino acid-
derived b-glycosides of a-hydroxynitriles. They
have been found in more than 2,650 higher plant
S. Bak (&) Æ M. Morant Æ A. V. Morant ÆS. Saito Æ N. Bjarnholt Æ M. Zagrobelny ÆK. Jørgensen Æ S. Osmani Æ H. T. Simonsen ÆR. S. Perez Æ T. B. van Heeswijck Æ B. L. MøllerPlant Biochemistry Laboratory, Department of PlantBiology, and Center for Molecular Plant Physiology(PlaCe), Royal Veterinary and AgriculturalUniversity, 40 Thorvaldsensvej, 1871 FrederiksbergCopenhagen, Denmarke-mail: [email protected]
S. M. PaquetteDepartment of Biological Structure, University ofWashington, HSB G-514/Box 357420, Seattle, WA98195-7420, USA
B. JørgensenResearch Group Cell Wall Biology and MolecularVirology, Department of Genetics andBiotechnology, Danish Institute of AgriculturalSciences, 40 Thorvaldsensvej, 1871 FrederiksbergCopenhagen, Denmark
Phytochem Rev (2006) 5:309–329
DOI 10.1007/s11101-006-9033-1
123
Cyanogenic glycosides: a case study for evolutionand application of cytochromes P450
Søren Bak Æ Susanne Michelle Paquette Æ Marc Morant Æ Anne Vinther Morant ÆShigeki Saito Æ Nanna Bjarnholt Æ Mika Zagrobelny Æ Kirsten Jørgensen ÆSarah Osmani Æ Henrik Toft Simonsen Æ Raquel Sanchez Perez ÆTorbjørn Bordier van Heeswijck Æ Bodil Jørgensen Æ Birger Lindberg Møller
Received: 24 May 2006 / Accepted: 22 September 2006 / Published online: 15 November 2006� Springer Science+Business Media B.V. 2006

species distributed among 130 families in pteri-
dophytes (ferns), gymnosperms and angiosperms
(flowering plants; Conn 1981; Siegler and Brinker
1993) implying that in plants the ability to
synthesize cyanogenic glycosides is at least
300 million years old. The widespread occurrence
of cyanogenic glycosides implies that they are
ancient biomolecules in terrestrial plants. Cyano-
genic glycosides are classified as phytoanticipins.
Upon disruption of plant tissue containing cya-
nogenic glycosides they are degraded by b-glyco-
sidases and a-hydroxynitrilases, resulting in
release of toxic hydrogen cyanide, an aldehyde
or ketone and glucose. This binary system—two
sets of components that separately are chemically
inert—provides plants with an immediate chem-
ical defence response to herbivores and patho-
gens that cause tissue damage (Møller and Seigler
1998; Jones et al. 2000; Morant et al. 2003; Nielsen
and Møller 2004; Poulton and Møller 1989).
Cyanogenic ferns and gymnosperm species con-
tain aromatic cyanogenic glycosides derived from
either tyrosine or phenylalanine, whereas angio-
sperms contain aliphatic as well as aromatic
cyanogenic glycosides derived from isoleucine/
valine/leucine or tyrosine/phenylalanine, respec-
tively (Fig. 1). In addition, a few plant species
such as Passiflora (Malphigiales, eurosids I) con-
tain cyanogenic glycosides derived from the
non-protein amino acid cyclopentenyl glycine
(Jaroszewski et al. 2002a, b). The specific pres-
ence of aromatic cyanogenic glycosides in ferns
and gymnosperms indicate that the cyanogenic
glycosides initially in nature were aromatic, and
that these served as progenitors for aliphatic
cyanogenic glycosides. This evolutionary path is
supported by the fact that ancestral angiosperms
like Magnoliales contain tyrosine-derived cyano-
genic glycosides. Within monocotyledons, Liliales
are known to contain aromatic cyanogenic glyco-
sides, and within Poales both aromatic and
aliphatic cyanogenic glycosides occur. In eudicots,
a wide distribution of aromatic as well as aliphatic
cyanogenic glycosides is observed, but the amino
acid precursor used within a given family is
generally conserved. The presence of aliphatic
cyanogenic glycosides in Poales and eudicots
raises the question of whether aliphatic
cyanogenic glycosides evolved independently at
least twice or whether they evolved before the
radiation of monocotyledons and eudicots. Typ-
ically, plants contain cyanogenic glycosides
derived from a single amino acid: tyrosine,
phenylalanine, leucine, valine or isoleucine,
except that valine- and isoleucine-derived cyano-
genic glycosides typically co-occur. It is only in
some polyploid species like hexaploid wheat
(Triticum aestivum, Poales) that aromatic and
aliphatic cyanogenic glycosides co-occur, but in
this case the co-occurrence is a result of the
allopolyploidity of the hexaploid wheat genome,
where the presence of either aliphatic or aromatic
cyanogenic glycosides in the diploid and tetra-
ploid ancestors have been retained during domes-
tication of wheat (our unpublished results).
Cyanogenic glycosides are not only found in
plants, but are also present in animals within a
limited number of anthropod clades (reviewed by
Zagrobelny et al. 2004; Fig. 2). A few species of
Diploda (millipedes), Chilopoda (centipedes),
Coleoptera (beetles) and Heteroptera (true bugs)
synthesize aromatic cyanogenic glycosides while
more than 200 species within Lepidoptera (but-
terflies and moths) synthesize aliphatic cyano-
genic glycosides. Accordingly, older animal
lineages and plant lineages contain aromatic
cyanogenic glycosides while relatively more
recent lineages as Lepidopteran species and
angiosperms have acquired the capacity to
contain aliphatic cyanogenic glycosides.
In analogy to plants, cyanogenic animals store
the cyanogenic glycosides in their defensive
sections and use them in their own defence
against predators. Within Lepidoptera, several
species have been identified that are able to
sequester cyanogenic glycosides from their host
plants and in some cases also are able to carry out
de novo synthesis of cyanogenic glycosides when
the amount of cyanogenic glycosides in the host
plant is not sufficient to maintain desired levels in
the insect. This implies a close co-evolution of
Lepidopteran species with their preferred host
plants, and that the ability to synthesize cyano-
genic glycosides has been lost in some Lepidop-
teran species. The radiation of Lepidopteran
species beginning 210 million years ago on a
gymnosperm dominated flora precedes the
radiation of angiosperms beginning 140 million
310 Phytochem Rev (2006) 5:309–329
123
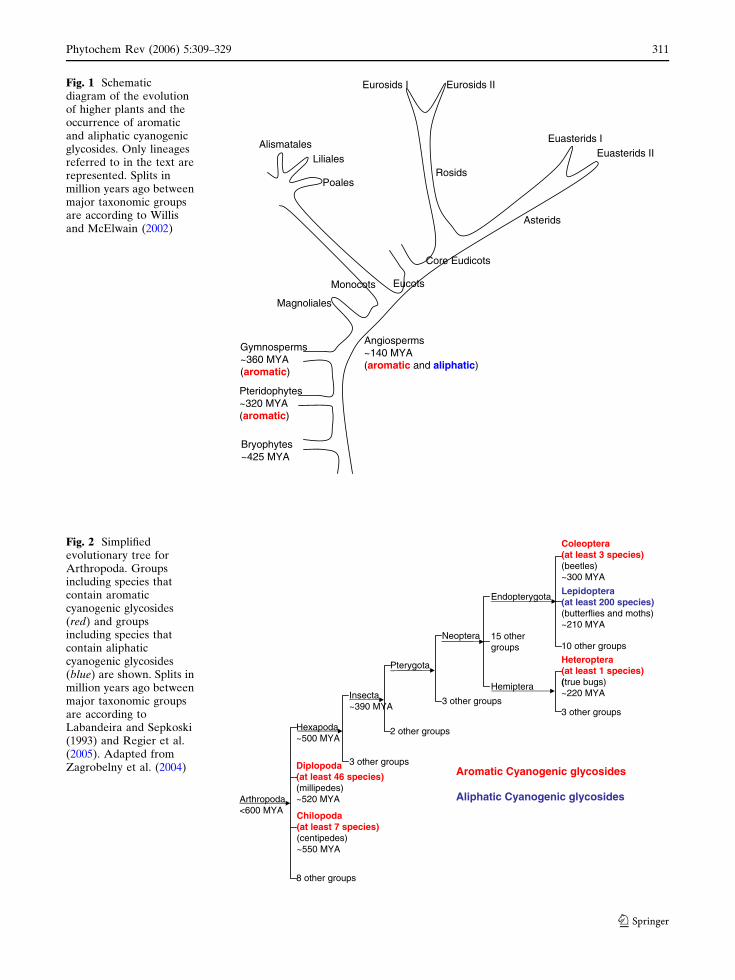
Pteridophytes~320 MYA(aromatic)
Gymnosperms~360 MYA(aromatic)
Angiosperms~140 MYA(aromatic and aliphatic)
Monocots Eucots
Core Eudicots
Rosids
Asterids
Eurosids I Eurosids II
Euasterids IEuasterids II
Poales
Liliales
Magnoliales
Alismatales
Bryophytes~425 MYA
Fig. 1 Schematicdiagram of the evolutionof higher plants and theoccurrence of aromaticand aliphatic cyanogenicglycosides. Only lineagesreferred to in the text arerepresented. Splits inmillion years ago betweenmajor taxonomic groupsare according to Willisand McElwain (2002)
Arthropoda<600 MYA
Hexapoda~500 MYA
Diplopoda(at least 46 species)(millipedes)~520 MYA
Chilopoda(at least 7 species)(centipedes)~550 MYA
Insecta~390 MYA
3 other groups
Pterygota
2 other groups
Neoptera
3 other groups
Endopterygota
15 othergroups
Lepidoptera(at least 200 species)(butterflies and moths)~210 MYA
10 other groups
Coleoptera(at least 3 species)(beetles)~300 MYA
Hemiptera
Heteroptera(at least 1 species)(true bugs)~220 MYA
3 other groups
Aromatic Cyanogenic glycosides
Aliphatic Cyanogenic glycosides
8 other groups
Fig. 2 Simplifiedevolutionary tree forArthropoda. Groupsincluding species thatcontain aromaticcyanogenic glycosides(red) and groupsincluding species thatcontain aliphaticcyanogenic glycosides(blue) are shown. Splits inmillion years ago betweenmajor taxonomic groupsare according toLabandeira and Sepkoski(1993) and Regier et al.(2005). Adapted fromZagrobelny et al. (2004)
Phytochem Rev (2006) 5:309–329 311
123

years ago (Labandeira 1993; Labandeira et al.
1994) implying that the ability to de novo
synthesize aliphatic cyanogenic glycosides in
some Lepidopteran species has been replaced by
sequestration as angiosperms containing aliphatic
cyanogenic glycosides proliferated.
The ability of Lepidoptera species to feed on
cyanogenic plants relies on the ability to metab-
olize toxic products formed by catabolism of
cyanogenic glycosides. While most animals rely
on detoxification of HCN by either b-cyanoala-
nine synthase or rhodanese, some Lepidopteran
species, like the neotropical butterfly (Heliconius
sara) have acquired an additional route to metab-
olize the cyanogenic glycosides via thiol
compounds. H. sara is able to sequester the
cyclopentenyl-derived cyanogenic glucoside,
epivolkenin (B4) from its host plant Passiflora
auriculata. In addition, it possesses the ability to
de novo synthesize the aliphatic cyanogenic
glycosides linamarin and lotaustralin. H. sara
metabolizes epivolkenin to the corresponding
thiol compound sarauriculatin by enzymatic
replacement of the nitrile group by a thiol. The
precise nature of this reaction is not known, but it
may enable H. sara to avoid release of free
hydrogen cyanide (Engler et al. 2000). The
replacement and possible ability to metabolize
the nitrile function into, e.g. ammonia and carbon
dioxide may constitute a nitrogen reservoir to
optimize the insect’s primary metabolism. This
adds yet another layer of complexity to the
co-evolution of cyanogenic glycoside metabolism
in insects and plants.
Biosynthesis of cyanogenic glycosides in higherplants and arthropods
In plants, the biosynthetic pathway for cyano-
genic glycosides is highly channelled (Møller and
Conn 1980) and catalysed by two unusual multi-
functional cytochromes P450 (P450) (Halkier and
Møller 1991; Sibbesen et al. 1994; Koch et al.
1995; Halkier et al. 1995; Sibbesen et al. 1995;
Kahn et al. 1997; Bak et al. 1998a) and a UDP-
glucosyl transferase (UGT; Jones et al. 1999;
Hansen et al. 2003; Fig. 3). The enzymes and the
corresponding structural genes were initially
identified from sorghum (Sorghum bicolor,
Poales) which synthesizes the tyrosine-derived
cyanogenic glucoside dhurrin (B3). While plant
cytochromes P450 are generally N-terminally
anchored membrane proteins, plant UGTs are
generally regarded as soluble cytosolic enzymes.
The first committed step in the dhurrin pathway
in sorghum is catalysed by the multifunc-
tional CYP79A1. This P450 catalyses two sequen-
tial N-hydroxylations of tyrosine followed by
a dehydration and a de-carboxylation reac-
tion to produce the corresponding aldoxime,
(Z)-p-hydroxyphenyl acetaldoxime. The second
cytochrome P450 in the pathway, CYP71E1,
catalyses a NADPH-dependent dehydration reac-
tion followed by a C-hydroxylation to produce the
cyanohydrin p-hydroxymandelonitrile, which is
HO
COOH
NH2
CYP79A1
HO
NOH
CYP71E1
HO
OH
CN
UGT85B1
HO
O
CN O
HO
OHOH
HO
Dhurrin
Fig. 3 The biosynthetic pathway of the tyrosine-derivedcyanogenic glucoside dhurrin in sorghum
312 Phytochem Rev (2006) 5:309–329
123
subsequently glycosylated by the family 1 glyco-
syltransferase UGT85B1 to produce dhurrin.
In vitro experiments involving simultaneous
administration of dual radiolabelled intermedi-
ates demonstrated that the dhurrin pathway is
highly channelled (Møller and Conn 1980). Our
working hypothesis is that CYP79A1 and
CYP71E1 and the otherwise soluble UGT85B1
form a metabolon, a multienzyme complex, to
facilitate channelling of the toxic and unstable
intermediates in the pathway (Nielsen and Møller
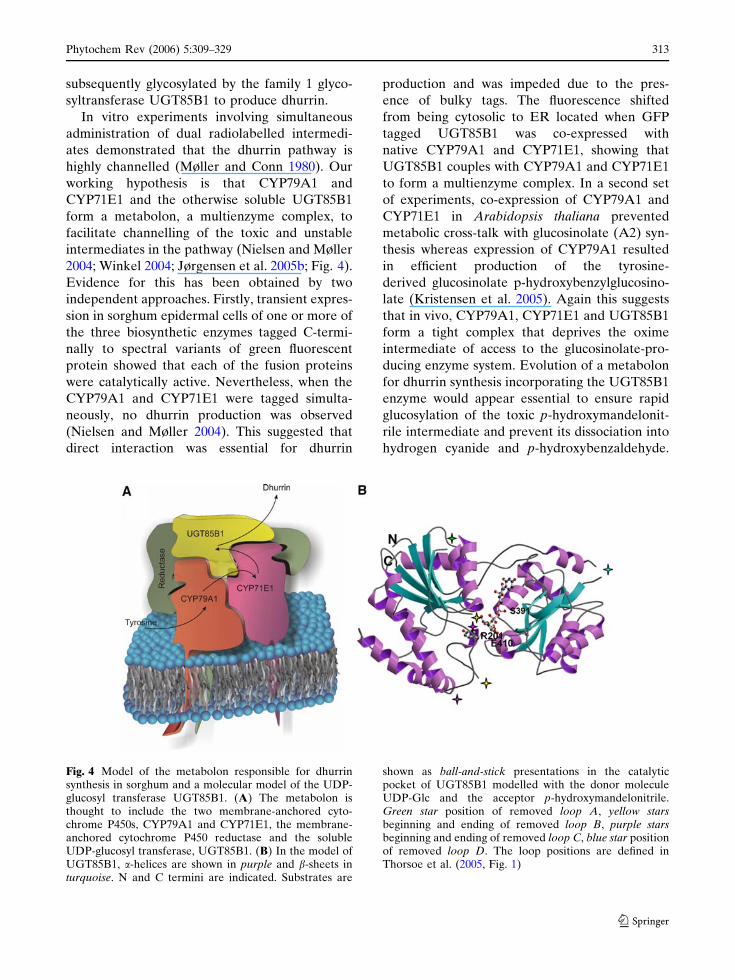
2004; Winkel 2004; Jørgensen et al. 2005b; Fig. 4).
Evidence for this has been obtained by two
independent approaches. Firstly, transient expres-
sion in sorghum epidermal cells of one or more of
the three biosynthetic enzymes tagged C-termi-
nally to spectral variants of green fluorescent
protein showed that each of the fusion proteins
were catalytically active. Nevertheless, when the
CYP79A1 and CYP71E1 were tagged simulta-
neously, no dhurrin production was observed
(Nielsen and Møller 2004). This suggested that
direct interaction was essential for dhurrin
production and was impeded due to the pres-
ence of bulky tags. The fluorescence shifted
from being cytosolic to ER located when GFP
tagged UGT85B1 was co-expressed with
native CYP79A1 and CYP71E1, showing that
UGT85B1 couples with CYP79A1 and CYP71E1
to form a multienzyme complex. In a second set
of experiments, co-expression of CYP79A1 and
CYP71E1 in Arabidopsis thaliana prevented
metabolic cross-talk with glucosinolate (A2) syn-
thesis whereas expression of CYP79A1 resulted
in efficient production of the tyrosine-
derived glucosinolate p-hydroxybenzylglucosino-
late (Kristensen et al. 2005). Again this suggests
that in vivo, CYP79A1, CYP71E1 and UGT85B1
form a tight complex that deprives the oxime
intermediate of access to the glucosinolate-pro-
ducing enzyme system. Evolution of a metabolon
for dhurrin synthesis incorporating the UGT85B1
enzyme would appear essential to ensure rapid
glucosylation of the toxic p-hydroxymandelonit-
rile intermediate and prevent its dissociation into
hydrogen cyanide and p-hydroxybenzaldehyde.
Fig. 4 Model of the metabolon responsible for dhurrinsynthesis in sorghum and a molecular model of the UDP-glucosyl transferase UGT85B1. (A) The metabolon isthought to include the two membrane-anchored cyto-chrome P450s, CYP79A1 and CYP71E1, the membrane-anchored cytochrome P450 reductase and the solubleUDP-glucosyl transferase, UGT85B1. (B) In the model ofUGT85B1, a-helices are shown in purple and b-sheets inturquoise. N and C termini are indicated. Substrates are
shown as ball-and-stick presentations in the catalyticpocket of UGT85B1 modelled with the donor moleculeUDP-Glc and the acceptor p-hydroxymandelonitrile.Green star position of removed loop A, yellow starsbeginning and ending of removed loop B, purple starsbeginning and ending of removed loop C, blue star positionof removed loop D. The loop positions are defined inThorsoe et al. (2005, Fig. 1)
Phytochem Rev (2006) 5:309–329 313
123

Currently, crystal structures of plant cytochromes
P450 are not available hampering the elucidation
of how CYP79A1 and CYP71E1 couple. How-
ever, a structural model of sorghum UGT85B1
was built based on hydrophobic cluster analysis
and the crystal structures of two bacterial GTs,
GtfA and GtfB, which each showed approxi-
mately 15% overall amino acid sequence identity
to UGT85B1 (Thorsoe et al. 2005). Despite the
overall low sequence identity, the UGT85B1
model enabled predictions about amino acid
residues important for catalysis and sugar donor
specificity. Furthermore, the UGT85B1 model
identified a hyper variable loop that contained a
hydrophobic patch that may mediating binding of
UGT85B1 to CYP79A1, and CYP71E1 within the
dhurrin metabolon. The mechanism that enables
docking of UGT85B1 into a metabolon is now
being investigated by molecular modelling and
site-directed mutagenesis (Thorsoe et al. 2005)
combined with reconstitution experiments con-
ducted using nanodisc technology (Bayburt and
Sligar 2003) where CYP79A1 and CYP71E1 have
been assembled into nanodiscs together with
sorghum cytochrome P450 reductase.
Many cyanogenic arthropod species have been
shown to de novo synthesize cyanogenic
glycosides by biochemical pathways that involve
similar or identical intermediates to those known
from plants, indicating that a homologous
pathway is present in animals (Duffey 1981;
Davis and Nahrstedt 1987; Zagrobelny et al.
2004; Feyereisen 2005). Older studies have shown
that all of the radiolabelled amino acid, N-
hydroxy amino acid, oxime and nitrile precursors
injected into Heliconius and Zygaena larvae
species or millipedes (Duffey and Towers 1978;
Holzkamp and Nahrstedt 1994) were not metab-
olized into cyanogenic glycosides. Most of these
precursors are now known to be highly unstable
compounds that are readily metabolized and
detoxified in planta (Bak et al. 2000). In addition,
the detection methods used were indirect, and
less sensitive than the LC-MS methods used in
the more recent plant studies. Feeding experi-
ments with Zygaena filipendulae larvae following
application of radiolabelled valine and isoleucine
directly to leaf surfaces of the host plant showed
that in the larvae the radiolabelled valine and
isoleucine ingested with the leaves are not
detected in intermediates but primarily ends up
in the cyanogenic glycosides linamarin and
lotaustralin. This supports a channelled pathway
in insects (Zagrobelny et al. in press) as seen in
plants.
Phylogeny of CYP79s in higher plants
CYP79 orthologs have been identified and iso-
lated from a number of cyanogenic angiosperms
(Fig. 5) using bioinformatics approaches, and
their function confirmed by reconstitution exper-
iments with heterologously expressed enzymes
(Halkier et al. 1996; Bak et al. 1999, 2000;
Andersen et al. 2000; Nielsen and Møller 2000).
No clear orthologs of sorghum CYP71E1 and
UGT85B1 have been reported. Nor have any
genes involved in cyanogenesis been identified in
arthropods. Accordingly, information on evolu-
tion of cyanogenic glycosides at the molecular
level is currently restricted to the CYP79 family in
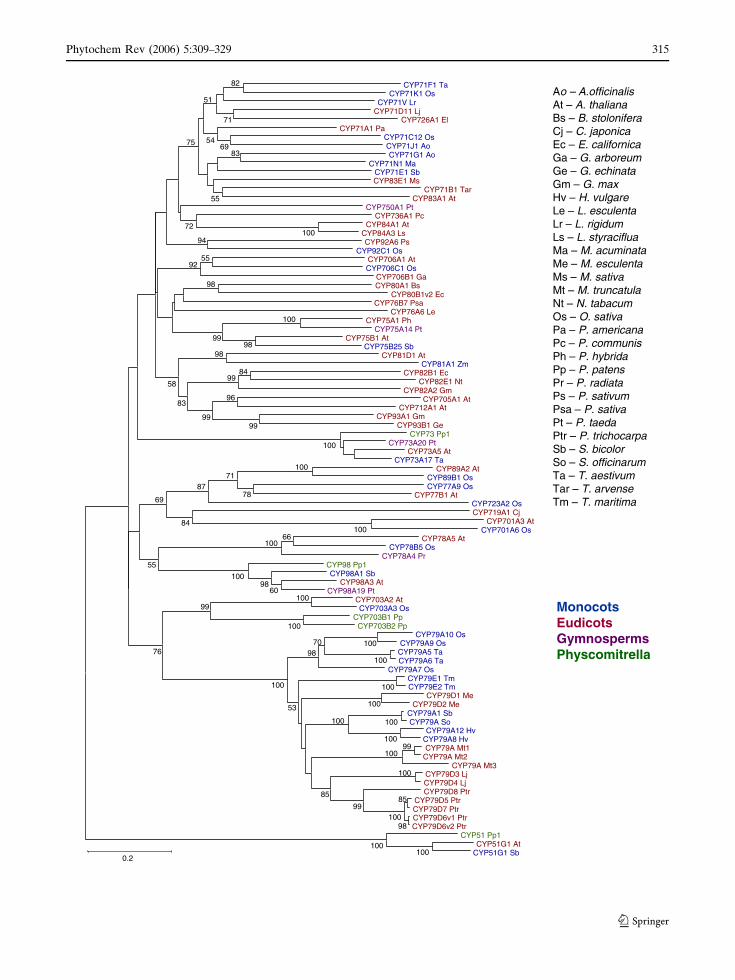
plants. Phylogenetic reconstruction, using CYP79
sequences and selected clan 71 family members
Fig. 5 Neighbour-join phylogenetic tree illustrating thephylogenetic relationships of the P450s involved incyanogenesis against a background of A-type cytochromeP450 sequences. Sequences are from monocots (blue),eudicots (red), gymnosperms (purple) and moss (green).Protein sequences were taken from GenBank and http://drnelson.utmem.edu/biblioD.html and aligned usingClustalX 1.83, and phylogenetic analysis was carried outusing MEGA 3.1 (Kumar et al. 2004). Bootstrap values areindicated in % and based on 1,000 iterations and the treehas been rooted with CYP51s as the outgroup. Bootstrapvalues less than 50% are excluded from the tree. Thealignment for this sequence can be accessed in PDF formatat http://www.p450.kvl.dk/Review2006/Figure5_Align-ment.pdf. Ao Asparagus officinalis, At Arabidopsis thali-ana, Bs Berberis stolonifera, Cj Coptis japonica, EcEschscholzia californica, Ga Gossypium arboreum, GeGlycyrrhiza echinata, Gm Glycine max, Hv Hordeumvulgare, Le Lycopersicon esculentum, Lr Lolium rigidum,Ls Liquidambar styraciflua, Ma Musa acuminata, MeManihot esculenta, Ms Medicago sativa, Mt Medicagotruncatula, Nt Nicotiana tabacum, Os Oryza sativa, PaPersea americana, Pc Pyrus communis, Ph Petunia xhybrida, Pp Physcomitrella patens, Pr Pinus radiata, PsPisum sativum, Psa Pastinaca sativa, Pt Pinus taeda, PtrPopulus trichocarpa, Sb Sorghum bicolor, So Saccharumofficinarum, Ta Triticum aestivum, Tar Thlaspi arvense,Tm Triglochin maritima
c
314 Phytochem Rev (2006) 5:309–329
123
CYP71F1 TaCYP71K1 Os
CYP71V LrCYP71D11 Lj
CYP726A1 ElCYP71A1 Pa
CYP71C12 OsCYP71J1 AoCYP71G1 Ao
CYP71N1 MaCYP71E1 SbCYP83E1 Ms
CYP71B1 TarCYP83A1 At
CYP750A1 PtCYP736A1 Pc
CYP84A1 AtCYP84A3 LsCYP92A6 Ps
CYP92C1 OsCYP706A1 AtCYP706C1 Os
CYP706B1 GaCYP80A1 Bs
CYP80B1v2 EcCYP76B7 Psa
CYP76A6 LeCYP75A1 Ph
CYP75A14 PtCYP75B1 At
CYP75B25 SbCYP81D1 At
CYP81A1 ZmCYP82B1 Ec
CYP82E1 NtCYP82A2 Gm
CYP705A1 AtCYP712A1 At
CYP93A1 GmCYP93B1 Ge
CYP73 Pp1CYP73A20 Pt
CYP73A5 AtCYP73A17 Ta
CYP89A2 AtCYP89B1 OsCYP77A9 Os
CYP77B1 AtCYP723A2 OsCYP719A1 Cj
CYP701A3 AtCYP701A6 Os
CYP78A5 AtCYP78B5 Os
CYP78A4 PrCYP98 Pp1CYP98A1 Sb
CYP98A3 AtCYP98A19 Pt
CYP703A2 AtCYP703A3 Os
CYP703B1 PpCYP703B2 Pp
CYP79A10 OsCYP79A9 Os
CYP79A5 TaCYP79A6 Ta
CYP79A7 OsCYP79E1 TmCYP79E2 Tm
CYP79D1 MeCYP79D2 Me
CYP79A1 SbCYP79A So
CYP79A12 HvCYP79A8 HvCYP79A Mt1
CYP79A Mt2CYP79A Mt3
CYP79D3 LjCYP79D4 LjCYP79D8 Ptr
CYP79D5 PtrCYP79D7 PtrCYP79D6v1 PtrCYP79D6v2 Ptr
CYP51 Pp1CYP51G1 At
CYP51G1 Sb
98
85
100
100
100
100100
100
100
100
99100
100
99
7098
85
53
100
100
66100
100
100
10060
98100
100
100
100100
100
8499
9899
99
99
96
99
98
94
5592
78
7187
84
83
83
82
76
71
69
55
54
51
75
72
69
55
58
98
Ao – A.officinalisAt – A. thalianaBs – B. stoloniferaCj – C. japonicaEc – E. californicaGa – G. arboreumGe – G. echinataGm – G. maxHv – H. vulgareLe – L. esculentaLr – L. rigidumLs – L. styracifluaMa – M. acuminataMe – M. esculentaMs – M. sativaMt – M. truncatulaNt – N. tabacumOs – O. sativaPa – P. americanaPc – P. communisPh – P. hybridaPp – P. patensPr – P. radiataPs – P. sativumPsa – P. sativaPt – P. taedaPtr – P. trichocarpaSb – S. bicolorSo – S. officinarumTa – T. aestivumTar – T. arvenseTm – T. maritima
MonocotsEudicotsGymnospermsPhyscomitrella
0.2
Phytochem Rev (2006) 5:309–329 315
123

from a number of angiosperms, showed that at
least within angiosperms the first committed step
is catalysed by an orthologous enzyme that
predates the segregation of angiosperms and
gymnosperms (Fig. 5). The tree is rooted with
ubiquitous obtusifoliol 14a-demethylase CYP51,
from Physcomitrella, Sorghum as well as Arabid-
opsis, to represent a bryophyte, a monocotyledon
and a eudicotyledon, respectively. The clan 71
constitutes a monophyletic clade predicted to
primarily encompass P450s involved in natural
products biosynthesis (secondary metabolism).
This clade proliferated vastly during the
course of plant land colonization over the last
430 million years through multiple gene, genome
segment and genome duplications and rearrange-
ments (Paquette et al. 2003; Nelson et al. 2004;
Galbraith and Bak 2005) in the continuous
processes of neo- and subfunctionalizations. The
CYP79 family branches off as a discrete clade
relatively deep in the tree which reflects the
ancient nature of cyanogenic glycosides. Deep in
the tree, the CYP73 and CYP98 families involved
in biosynthesis of lignin and other phenylpropa-
noids (Humphreys and Chapple 2002) branch off,
as well as the CYP701 family involved in biosyn-
thesis of gibberellins (Helliwell et al. 1998); the
occurrence of these pathways most likely predate
cyanogenic glucoside biosynthesis in terrestrial
plants. As CYP79 sequences from cyanogenic
gymnosperms and pteridophytes are as yet not
available, the analysis is restricted to angiosperms.
This may influence the position of the individual
CYP79s in the 79 clade as well as the position of
the CYP79 clade in relation to the CYP73, CYP98
and CYP703 clades from which sequences are
indeed available from bryophytes, gymnosperms
as well as angiosperms. From monocotyledons,
CYP79 sequences encoding enzymes with aro-
matic as well as aliphatic amino acids as substrates
are available from both Poales and Alismatales
(triglochin). The only known CYP79 sequence
from eudicotyledons is restricted to eurosids I in
Rosids and have aliphatic amino acids as sub-
strates, though aromatic cyanogenic glycosides are
present in eurosides. In contrast to most other
plant P450s, members of the CYP79 family are
characterized by unique amino acid substitutions
in the otherwise generally conserved ‘‘PERF’’ and
heme-binding domain (Werck-Reichhart et al.
2002; Bak et al. 1998b). These substitutions have
facilitated the isolation of cDNAs from a number
of plants using family CYP79 specific primers
(Andersen et al. 2000; Nielsen and Møller 2000)
and has provided a tool to readily identify CYP79s
among other cytochrome P450 sequences in
databases (Nielsen and Møller 2000; Forslund
et al. 2004). In the PERF region of the CYP79
family, the otherwise conserved apolar and aro-
matic phenylalanine residue has been replaced by
the positively charged amino acid residue histidine
giving the CYP79 family consensus PERH.
Similarly, in the heme-binding consensus se-
quence PFGXGRRXCXG, three substitutions of
hydrophobic amino acids with polar serine or thre-
onine residues are observed, affording a CYP79
consensus of (S/T)F(S/T)TGRRGCXG (see full-
sequence alignment at http://www.p450.kvl.dk/
Review2006/Figure5_Alignment.pdf and see
sequence logos at http://www.p450.kvl.dk//
p450.shtml#logos). The identification of CYP79s
from eudicots, other than rosids, and especially
from gymnosperms and pteridophytes will reveal
how far back these unique substitutions can be
tracked. The PERF and heme-binding regions are
part of the active site and the significance of more
polar amino acid residue in the PERF and heme-
binding region is currently not understood. How-
ever, the substitutions may be related to the
unusual catalytic activities of the CYP79 family
and/or to the fact that amino acid substrates as
well at the generated intermediates are unusually
hydrophilic compared to most other cytochrome
P450 substrates. The availability of crystal struc-
tures of a number of mammalian P450s (Williams
et al. 2003; Scott et al. 2003; Williams et al. 2004)
may facilitate homology modelling and docking of
substrates to further the understanding of the
impact of these changes, analogously to the
information gained by the sorghum UGT85B1
model (Fig. 4; Thorsoe et al. 2005).
Glucosinolates constitute another subclass of
N-containing natural products. They are present
in Brassicales within eurosides II (Bak et al.
1998b; Halkier and Gershenzon 2006). They are
derived from a subset of protein amino acids that
partly overlap with those used for synthesis of
cyanogenic glycosides as well as from chain
316 Phytochem Rev (2006) 5:309–329
123

elongated derivatives of these amino acids. As in
cyanogenic glucoside synthesis, the conversion of
these parent amino acids into the corresponding
oximes is catalysed by CYP79’s (Bak et al.
1998b). CYP79s in glucosinolate containing spe-
cies are clearly phylogenetically related to
CYP79s from cyanogenic species, and this most
likely represents recruitment of genes from the
cyanogenic glucoside pathway to the more re-
cently evolved glucosinolate pathway (Bak et al.
1998b). Interestingly, glucosinolates are also
found in Drypetes belonging to Malphigiales,
eurosides I. This has given rise to the hypothesis
of a parallel evolution of glucosinolate biosyn-
thesis in rosids from a cyanogenic ancestor
(Rodman et al. 1998). The isolation of genes
encoding enzymes in glucosinolate biosynthesis
from Drypetes should resolve this issue.
In the glucosinolate pathway in Arabidopsis,
the oximes generated by CYP79s are further
metabolized by CYP83A1 or CYP83B1 (Bak and
Feyereisen 2001; Bak et al. 2001; Hansen et al.
2001; Naur et al. 2003). In sorghum, the oxime is
metabolized by CYP71E1 to the cyanohydrin.
CYP83A1 and CYP83B1 share more than 55%
sequence identity at the amino acid level. Accord-
ing to the generally approved nomenclature
system (Nelson et al. 1996), they should then
have been placed in the same subfamily. How-
ever, it has been decided to retain the two
subfamilies to prevent confusion in the literature.
In addition, based on their amino acid sequence
identity and by their clustering within the CYP71
family in phylogenetic trees, the CYP83 family
clearly belongs to the CYP71 family (Fig. 5).
Mainly due to historical reasons as mentioned for
CYP83B1, the CYP83 family has been kept.
Though CYP83s in Arabidopsis and CYP71E1
in sorghum both have oximes as substrates, the
catalytic mechanism does not appear to be
conserved. In cyanogenic plants, oximes are
dehydrated to a nitrile and subsequently hydrox-
ylated to produce an a-hydroxynitrile. It has not
been resolved whether both of these partial
reactions require NADPH or whether the overall
requirement for NADPH is restricted to the
second partial reaction which is a classical cyto-
chrome P450 C-hydroxylation reaction. In the
glucosinolate pathway, oximes are by a yet
unknown mechanism converted to a thiohydroxy-
mate via a proposed reactive aci-nitro compound
(Ettlinger and Kjær 1968; Bak et al. 2001; Bak
and Feyereisen 2001). If the mechanistics of the
CYP71E1 catalysed dehydration reaction in cya-
nogenic glucoside synthesis was resolved, this
may serve to shed new light on the identity of the
intermediates in glucosinolate synthesis. How-
ever, it remains an open question whether the
CYP83s in glucosinolate biosynthesis have
evolved from a CYP71E1 ortholog, or whether
glucosinolate-producing plants have recruited
other CYP71 homologs to catalyse this step.
Indole-3-acetaldoxime, derived from CYP79B2
and/or CYP79B3 metabolism of tryptophan, is
apart from being an intermediate in indole
glucosinolate biosynthesis an intermediate in
biosynthesis of the main phytoalexin camalexin
in Arabidopsis. CYP71B15, which is a relatively
close homolog to both CYP71E1 and CYP83A1/
B1 (Fig. 5) catalyses the conversion of indole-
3-acetaldoxime-derived dihydrocamalexic acid to
camalexin (A3; Schuhegger et al. 2006), implying
that the CYP71 family is a major player in the
recruitment of enzyme catalysts for evolution of
novel natural products biosynthetic pathways in
plants.
The CYP71 family is by far the most
expanded cytochrome P450 family in plants,
and this combined with the apparent lack of
unique sequence substitutions as seen for the
CYP79 family has despite the increasing num-
ber of plant genome and EST sequencing
projects prevented the identification of clear
CYP71E1 orthologs in other species apart from
the closely related sugar cane and rice. Simi-
larly, no clear UGT85B1 orthologs have as yet
been identified.
Cyanoalk(en)yl glycosides: a subclass of natural
products related to cyanogenic glycosides
Lotus japonicus contains high levels of rhodiocy-
anosides A (C1) and D as well as of the
cyanogenic glycosides linamarin (B1) and
lotaustralin (B2; Forslund et al. 2004). Rhodiocy-
anosides A and D belong to a group of
cyanoalkenyl glycosides structurally related to
Phytochem Rev (2006) 5:309–329 317
123

cyanogenic glycosides. In contrast to the
cyanogenic glycosides, cyanoalk(en)yl glycosides
are not cyanogenic, i.e. they do not release HCN
by treatment with b-glucosidases. This reflects
that these glycosides are not derived from
cyanohydrins and that they are hydrolysed into
stable nitriles by b-glucosidases. Rhodiocyano-
sides A and D were first isolated from Rhodiola
species. Rhodiola extracts are used in Chinese
traditional medicine, and the rhodiocyanosides
have been proposed to function as inhibitors of
histamine release in allergic responses (Yoshika-
wa et al. 1995, 1996a, b). As cyanogenic glycosides
are ancient, several animals and fungi have
evolved to be able to use the otherwise toxic
HCN as a source of nitrogen. Accordingly, for the
plant the presence of cyanogenic glycosides may
in fact be disadvantageous. The transition to
cyanoalk(en)yl glycosides may counteract the
adaptation of herbivores and microbes to HCN.
Rhodiocyanosides are derived from the amino
acid isoleucine (Forslund et al. 2004). This implies
that the biosynthesis of rhodiocyanosides is
catalysed by the very same multienzyme complex
involved in lotaustralin synthesis. However, when
rhodiocyanosides are synthesized, the putative
nitrile intermediate undergoes two hydroxyla-
tions and a dehydration reaction (Forslund et al.
2004). The precise mechanism is not understood,
but we suggest that the initial hydroxylation
produces a product that in contrast to the cyano-
hydrin is not easily released from the active site.
This affords the possibility of a dehydration
reaction to proceed while the intermediate
remains bound in the catalytic site. The rhodio-
cyanosides may then be formed by a subsequent
hydroxylation reaction. Metabolite profiling of
Lotus, Rhodiola and also Ribes species by LC-MS
demonstrated that the rhodiocyanosides co-occur
with lotaustralin (Bjarnholt et al. our unpublished
results). Hydroxylated cyanoalkenyl glycosides
such as sarmentosin (C2) have been identified in
Sedum sarmentosum (Fang et al. 1982; Lechten-
berg and Nahrstedt 1999). A similar complex
profile is known from barley, which in addition
to the leucine-derived cyanogenic glucoside
epi-heterodendrin contains the cyanoalkenyl
glycosides osmaronin, dihydroosmaronin (C4),
epidermin and sutherlandin (Nielsen et al.
2002). Our current working hypothesis is that in
species containing cyanoalk(en)yl glycosides,
CYP71E1 orthologs have been duplicated and
subsequently evolved to catalyse two sequential
hydroxylations accompanied by a dehydration
reaction. The availability of model plants like L.
japonicus containing rhodiocyanoside A and D
will allow the elucidation of how this new
pathway has evolved from a cyanogenic glucoside
predisposition on the molecular level. Again,
elucidation of these reaction events may serve
to cast new light on the intermediates involved in
the conversion of oximes to thiohydroxymates in
glucosinolate synthesis.
The proposed evolution of glucosinolates,
camalexin and cyanoalk(en)yl glycosides from a
cyanogenic glucoside predisposition illustrates
how the ancient pathway for cyanogenic gluco-
side synthesis has given rise to new classes and
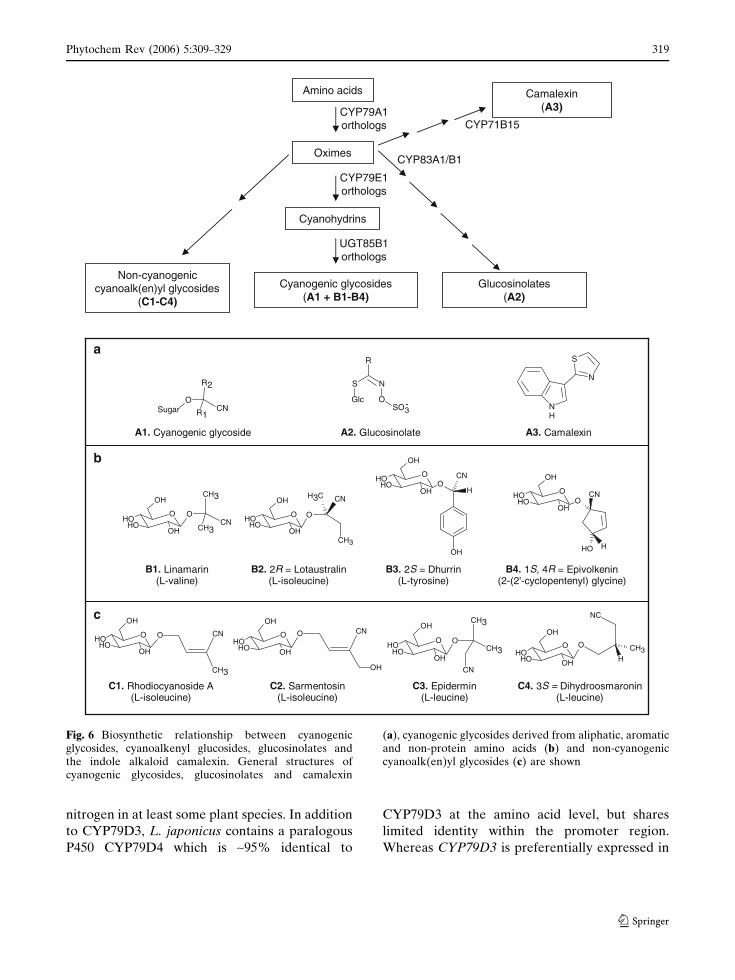
subclasses of natural products (Fig. 6).
Localization of CYP79s and cyanogenic
glycosides
In L. japonicus, the cyanogenic glycosides accu-
mulate in aerial tissues (Forslund et al. 2004).
CYP79D3 transcripts are primarily found in
young and developing tissues where the concen-
tration of cyanogenic glycosides per mg fresh
weight is also the highest. In a similar manner,
sorghum CYP79A1 is predominantly expressed in
the young developing seedling (Busk and Møller
2002). The level of cyanogenic glycosides per mg
fresh weight decreases in sorghum and L. japo-
nicus as a function of developmental age. This
indicates that synthesis primarily occurs in young
and developing tissues and that the levels found
in older plant parts decreases because de novo
synthesis proceeds at a lower rate than catabolic
turnover. The presence of high amounts of
cyanogenic glycosides in young and developing
tissues supports the proposed function as defence
compounds. In cassava, sorghum and eucalyptus,
the level of cyanogenic glycosides increase under
high nitrogen levels (Gleadow and Woodrow
2000; Busk and Møller 2002; Gleadow and
Woodrow 2002; Jørgensen et al. 2005a) indicating
that cyanogenic glycosides may also serve a
function as storage compounds for reduced
318 Phytochem Rev (2006) 5:309–329
123
nitrogen in at least some plant species. In addition
to CYP79D3, L. japonicus contains a paralogous
P450 CYP79D4 which is ~95% identical to
CYP79D3 at the amino acid level, but shares
limited identity within the promoter region.
Whereas CYP79D3 is preferentially expressed in
Amino acids
CYP79A1orthologs CYP71B15
CYP83A1/B1
Camalexin (A3)
Oximes
Cyanogenic glycosides(A1 + B1-B4)
Non-cyanogeniccyanoalk(en)yl glycosides
(C1-C4)
Glucosinolates(A2)
CYP79E1orthologs
Cyanohydrins
UGT85B1orthologs
CH3CN
CH3
O
OHOH
OH
OH
O
B1. Linamarin(L-valine)
B2. 2R = Lotaustralin(L-isoleucine)
B3. 2S = Dhurrin(L-tyrosine)
B4. 1S, 4R = Epivolkenin(2-(2'-cyclopentenyl) glycine)
H
OH
OCN
OHOH
OH
OH
O
HOH
OCN
OHOH
OH
OH
O
CH3
CNCH3
O
OHOH
OH
OH
O
a
SO3OGlc
S N
R
-Sugar R1
CN
R2
O
A1. Cyanogenic glycoside A2. Glucosinolate
N
S
NH
A3. Camalexin
b
c
CN
CH3
CH3
O
OHOH
OH
OH
O
C3. Epidermin(L-leucine)
C4. 3S = Dihydroosmaronin(L-leucine)
C1. Rhodiocyanoside A(L-isoleucine)
C2. Sarmentosin(L-isoleucine)
CNO
CH3
OHOH
OH
OH
O CN
OH
O
OHOHOH
OH
O
H
NC
CH3O
OHOHOH
OH
O
Fig. 6 Biosynthetic relationship between cyanogenicglycosides, cyanoalkenyl glucosides, glucosinolates andthe indole alkaloid camalexin. General structures ofcyanogenic glycosides, glucosinolates and camalexin
(a), cyanogenic glycosides derived from aliphatic, aromaticand non-protein amino acids (b) and non-cyanogeniccyanoalk(en)yl glycosides (c) are shown
Phytochem Rev (2006) 5:309–329 319
123

leaves, CYP79D4 is preferentially expressed in
root tissue although at an apparent relatively
lower level. The two enzymes possess the same
kinetic parameters when heterologously ex-
pressed in yeast cells (Forslund et al. 2004). The
role of expression of a fully functional paralog
in an otherwise apparent acyanogenic tissue is
currently not understood. However, as a legume
L. japonicus interacts with Rhizobium bacteria to
form nitrogen-fixing nodules under low nitrogen
levels. Our analyses of CYP79D3 and CYP79D4
expression as well as the distribution of cyano-
genic and cyanoalkenyl glycosides in L. japonicus
has not included Rhizobium infected roots but
only tissue from hydroponically grown plants
supplemented with high nitrogen levels. The
possibility that cyanogenic or cyanoalkenyl gly-
cosides may serve as signalling compounds in
symbiotic nitrogen fixation or as transporters of
reduced nitrogen regulated by the nitrogen supply
and Rhizobium interaction remains an intriguing
question. Generation of transgenic L. japonicus
lines with CYP79D3 and CYP79D4 promoters
fused to reporter genes is in progress and will aid
to elucidate the functions of cyanogenic and
cyanoalkenyl glycosides in interactions with
micro-organisms and herbivores (Saito et al.,
our unpublished results).
In cassava, cyanogenic glycosides are prefer-
entially synthesized in the aerial parts and subse-
quently transported to the roots for storage
(Jørgensen et al. 2005a). The mechanism of
transport and the possible existence of a specific
transport form remain elusive. Studies in Hevea
cotyledons suggest that transport of the cyano-
genic glucoside linamarin requires an initial
conversion into the diglucoside linustatin (Selmar
et al. 1988). In contrast to the monoglucoside, the
diglucoside is not hydrolysed by the apoplastic
b-glucosidases and thus fulfil the prerequisite of a
non-degradable transport form. In L. japonicus
and sorghum, the localization of CYP79s at the
tissue level has been determined by RT-PCR and
northern-blotting analyses. In cassava, the local-
ization of CYP79D1 and CYP79D2 transcripts at
the cellular level has been visualized by FITC
labelling by in-tube in situ PCR in 3-month-old
plants (Jørgensen et al. 2005a). The first unfolded
leaf and its petiole have the highest cyanogenic
glucoside biosynthetic activity. In these two
tissues, the transcripts were localized within the
epidermis and the adjacent two cortex cell layers,
and in the endodermis and pericycle cell layers
(Fig. 7). CYP79D1 and CYP79D2 are also highly
expressed in tissues surrounding laticifers and
phloem cells, in regions between vascular bun-
dles, in parenchymatic cells in the vascular tissue
and especially in between the protoxylem and
metaxylem cells. The expression of CYP79D1
Fig. 7 Cellular localization of CYP79D1 in the petiolefrom the first fully unfolded cassava leaf as monitored byin-tube in situ RT-PCR analysis using light microscopy of100 lm transverse sections. (A) Petiole control to visualizethe different cell types. (B) Petiole labelling backgroundcontrol with FITC as observed without primers in the RTreaction. (C) Expression of CYP79D2 in petiole asmonitored by FITC labelling. (D) Expression of CYP79D1in petiole as monitored by FITC labelling. (E) Expressionof CYP79D1 in petiole as monitored by alkaline phospha-tase labelling. e Epidermis, p phloem, l laticifers, enendodermis. Bars = 200 lm. Adapted from Jorgensenet al. (2005)
320 Phytochem Rev (2006) 5:309–329
123

and CYP79D2 in epidermal and cortex cells and
around laticifers represents the function of cya-
nogenic glycosides as biochemical protectants as
they are expressed in conjunction with cassava
linamarin b-glucosidases apoplastically localized
in laticifers. Expression around the phloem and
vascular bundles could represent sites of biosyn-
thesis targeted for transport to the tubers.
Both L. japonicus and cassava possess two
CYP79Ds. In L. japonicus, the two paralogs
represent a clear case of subfunctionalization
where the promoters have diverged while the
coding regions are highly conserved as are their
catalytic parameters. No ESTs for CYP79D4 has
been identified, but RT-PCR experiments reveal
expression in roots (Forslund et al. 2004). In
contrast, the two cassava paralogs are only ~85%
identical at the amino acid level, have the same
catalytic parameters and are co-expressed
(Andersen et al. 2000; Jørgensen et al. 2005a).
The presence of two cassava paralogs is most
likely a consequence of the allotetraploid genetic
background of cassava with one allele originating
from each of the parental lines.
Metabolic engineering of cyanogenic glycosides
The pathway for conversion of a parent amino
acid into the corresponding cyanogenic glucoside
involves six intermediates of which several are
labile. From a genetic point of view, the cyano-
genic glucoside pathway is quite simple as two of
the enzymes responsible for these conversions are
multifunctional. Accordingly, the entire pathway
is encoded by just three structural genes. These
three genes are available from sorghum (Tatter-
sall et al. 2001; Kristensen et al. 2005). This gives
cyanogenic glycosides a pioneering position in
metabolic engineering of natural products in
plants because most other pathways for complex
natural product, e.g. alkaloids have not yet been
fully elucidated and because a substantially larger
number of genes are involved (Kutchan 2005).
The ability to engineer up to 4% dry weight of the
tyrosine-derived cyanogenic glucoside dhurrin
into transgenic A. thaliana plants by ectopic
expression of sorghum CYP79A1, CYP71E1 and
UGT85B1 (Tattersall et al. 2001; Kristensen et al.
2005) has been recently reviewed (Jørgensen
et al. 2005b; Kutchan 2005; Morant et al. 2003;
Memelink 2005). The transfer of the entire high
flux dhurrin pathway from sorghum to Arabidop-
sis by genetic engineering proceeded essentially
without inadvertent effects on the metabolome
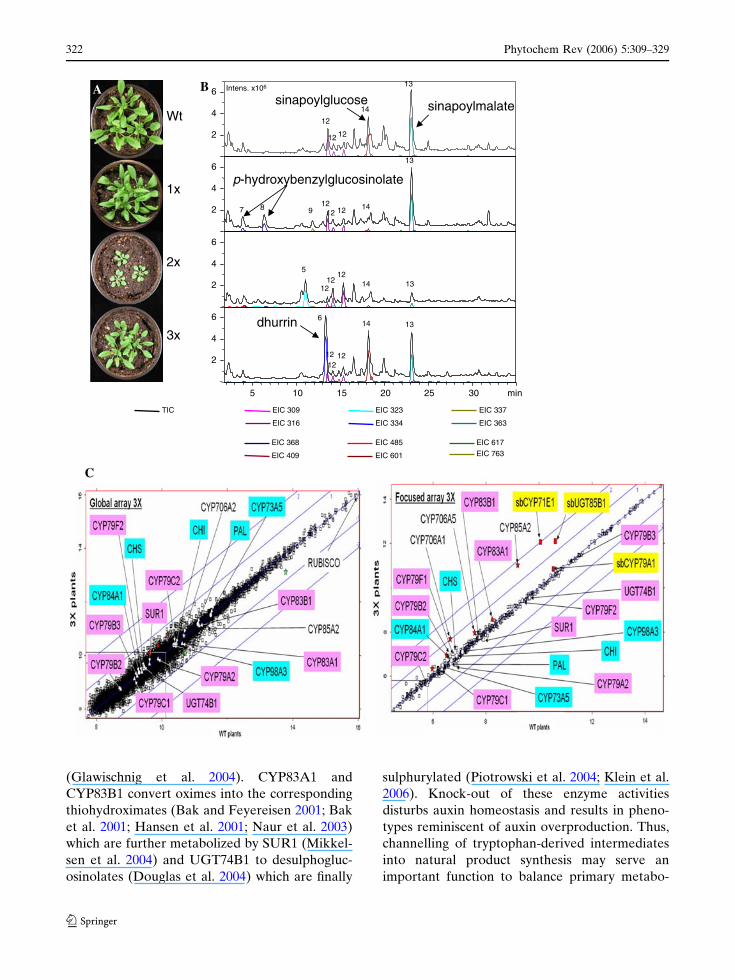
and transcriptome (Kristensen et al. 2005; Fig. 8).
This demonstrates that effective encapsulation of
toxic intermediates by metabolon formation is
also achievable after heterologous expression in a
plant species that would not in nature produce the
same class of natural product (Tattersall et al.
2001; Kristensen et al. 2005). Insertion of an
incomplete pathway (sorghum CYP79A1 and
CYP71E1) resulted in stunted plants, transcrip-
tome alterations, accumulation of numerous new
glycosides derived from detoxification of inter-
mediates in the dhurrin pathway, and in loss of
the Brassicaceae specific UV protectants sinapoyl
glucose and sinapoyl malate (Bak et al. 2000;
Tattersall et al. 2001; Kristensen et al. 2005).
When separately introduced into Arabidopsis,
sorghum CYP79A1 established a highly efficient
interaction with the down-stream glucosinolate-
producing enzymes and resulted in accumulation
of large amounts of p-hydroxybenzylglucosinolate
(up to 3% dry weight; Bak et al. 1999). This gluc-
osinolate is not normally present in Arabidopsis
and thereby changes the overall glucosinolate
profile of Arabidopsis (Petersen et al. 2001). The
feasibility to redirect tyrosine utilization in
Arabidopsis into glucosinolate or cyanogenic
glucoside production without loss of plant fitness
(Bak et al. 1999; Tattersall et al. 2001; Mikkelsen
et al. 2002; Mikkelsen and Halkier 2003; Kristen-
sen et al. 2005) demonstrates the existence of
inherent routes for transport and storage of new
natural product classes introduced into plants by
genetic engineering and an inherent ability to
redirect and optimize the flux of intermediates to
counteract imbalances in primary and secondary
metabolism in plants. The ability to accommodate
altered levels of intermediates, however, depends
on the type of metabolic cross-talk introduced. In
Arabidopsis, tryptophan-derived oximes are key
intermediates in formation of the phytohormone
indole acetic acid (Woodward and Bartel 2005) as
well as in biosynthesis of glucosinolates (Bak
et al. 2001) and the indole alkaloid camalexin
Phytochem Rev (2006) 5:309–329 321
123
(Glawischnig et al. 2004). CYP83A1 and
CYP83B1 convert oximes into the corresponding
thiohydroximates (Bak and Feyereisen 2001; Bak
et al. 2001; Hansen et al. 2001; Naur et al. 2003)
which are further metabolized by SUR1 (Mikkel-
sen et al. 2004) and UGT74B1 to desulphogluc-
osinolates (Douglas et al. 2004) which are finally
sulphurylated (Piotrowski et al. 2004; Klein et al.
2006). Knock-out of these enzyme activities
disturbs auxin homeostasis and results in pheno-
types reminiscent of auxin overproduction. Thus,
channelling of tryptophan-derived intermediates
into natural product synthesis may serve an
important function to balance primary metabo-
dhurrin
sinapoylglucose sinapoylmalate
B Intens. x106
5 10 15 20 25 30 min
7 8 9 1212
12 14
13
12
12
12
14
13
1212
1214 13
5
1212 12
14 136
p-hydroxybenzylglucosinolate
Wt
1x
2x
3x
A
2
4
6
6
4
2
6
4
2
6
4
2
EIC 316
EIC 309
EIC 363
EIC 337
EIC 368
EIC 409
EIC 485
EIC 601
EIC 617
EIC 763
EIC 334
EIC 323 TIC
C
322 Phytochem Rev (2006) 5:309–329
123

lism. A disturbance of oxime metabolism affects
phenylpropanoid metabolism and the monomer
composition of lignin, but the nature of this
interaction is not yet understood (Hemm et al.
2003). The ability to redirect aromatic amino
acids into new metabolic pathways also reflects
the plasticity of the shikimate pathway. In plants,
this pathway accommodates up to 20% of the flux
of fixed carbon and is primarily regulated by feed-
back inhibition to facilitate metabolic alterations
of aromatic compound in plants (Herrmann
1995).
Ectopic expression of cassava CYP79D2 in
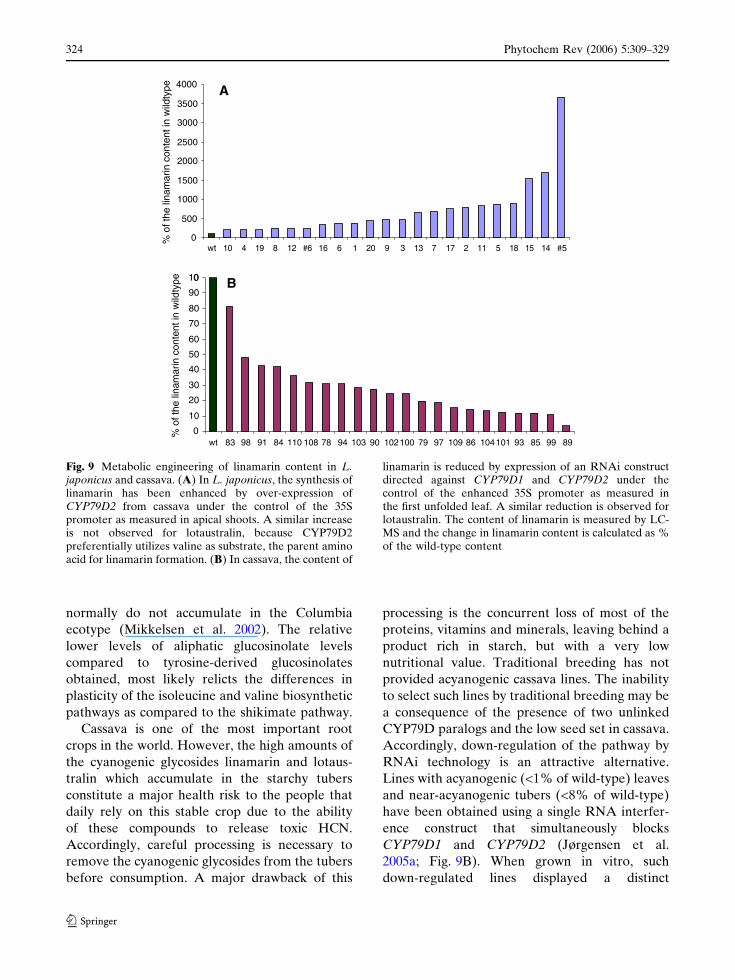
transgenic L. japonicus plants resulted in a ~20-
fold up-regulation of the valine-derived cyano-
genic glucoside linamarin (Forslund et al. 2004;
Fig. 9A). A similar increase of isoleucine-derived
lotaustralin and the cyanoalkenyl glycosides rhod-
iocyanoside A and D was not observed. Accord-
ingly, the relative differences in catalytic
efficiency (Kcat/Km) of cassava CYP79D2 as
compared to L. japonicus CYP79D3 towards the
substrates valine and isoleucine is a major deter-
minator of the ratio of linamarin and lotuastralin
obtained. The cassava CYP79D2 enzyme has
about the same catalytic efficiency towards valine
and isoleucine, and accordingly generates an
approximately 1:1 ratio of linamarin to lotaustr-
alin in this species (Andersen et al. 2000). In
contrast, L. japonicus CYP79D3 has a ~6-fold
higher catalytic efficiency towards isoleucine as
compared to valine (Forslund et al. 2004). This
results in a correspondingly lower relative level of
linamarin to lotaustralin. Accordingly, the trans-
genic L. japonicus lines accumulated much more
linamarin and approached the linamarin to
lotaustralin ratio in cassava. A surprising result
of the ectopic expression of cassava CYP79D2 in
L. japonicus was the accumulation of linamarin
and lotaustralin in roots, demonstrating that roots
indeed do have the capacity to synthesize cyano-
genic glycosides and that presence of the CYP79
step exerts quantitative and qualitative control of
the flux through the pathway. The cyanoalkenyl
glycosides rhodiocyanoside A and D did not
co-accumulate in roots, which is a strong indica-
tion of the presence of two separate biosynthetic
pathways for cyanogenic and alkenyl glucoside
synthesis in roots and aerial tissues in L. japoni-
cus. The possibility that the root content of
linamarin and lotaustralin was derived from
translocation from the aerial parts to the roots
cannot be ruled out. However, the presence in
L. japonicus of two CYP79D paralogs with
different expression patterns supports the pres-
ence of two independent pathways. Unfortu-
nately, the CYP71E1 orthologs in L. japonicus
have not yet been identified and this hampers the
understanding of how L. japonicus organizes the
synthesis of cyanogenic and cyanoalkenyl glyco-
sides in aerial tissues; either as parallel metabo-
lons or as a single promiscuous metabolon
that synthesizes both sets of glycosides simulta-
neously.
Ectopic expression of cassava CYP79D2
has also been used to alter the glucosinolate
profile in Arabidopsis (Mikkelsen and Halkier
2003). In analogy to metabolic cross-talk
observed by expression of sorghum CYP79A1,
the CYP79D2 expressing transgenic lines accu-
mulate up to 0.3% dry weight of the valine- and
isoleucine-derived isopropyl- and 1-methylpro-
pyl-glucosinolates, two glucosinolates that
Fig. 8 Metabolic engineering of dhurrin synthesis intransgenic Arabidopsis plants with marginal inadvertenteffects on the metabolome and transcriptome. Impact onvisual phenotype (A), metabolome (B) and transcriptomeas a result of expression of sorghum CYP79A1 either alone(1·) or combined with sorghum CYP71E1 (2·) or withsorghum CYP71E1 and UGT85B1 (3·) as compared towild-type (WT). (B) LC-MS analysis of the metabolitecontent of the plants shown in (A) as monitored by thetotal ion chromatograms (TIC, black lines) and extractedion chromatograms (EIC, coloured lines). Metabolitesreferred to in the text are high lighted: Sinapoyl glucose(14), sinalpoylmalate (13), dhurrin (6) and p-hydroxy-benzylglucosinolate (7 and 8), additional labelling as inKristensen et al. (2005). c Scatter plot of trancriptomeanalyses of wild-type (WT) versus dhurrin-producing A.thaliana plants (3· plants). The global arrays contains~29,000 probes while the focussed array contains probesfor 453 selected genes in secondary metabolism, aromaticamino acid biosynthesis, all 246 full-length A. thalianacytochromes P450, except CYP84A4, and all 112 family 1UGT except for members of the UGT80 and UCT81families. Phenylpropanoid marker genes are shown in blue,glucosinolate marker genes in pink and sorghum transg-enes in yellow (not present on global array). Genes scoredas up-regulated are shown in red and genes down-regulated in green. Signals below two time’s averagebackground are boxed. Blue lines indicate twofold orfourfold up- or down-regulated expression levels. Adaptedfrom Kristensen et al. (2005)
b
Phytochem Rev (2006) 5:309–329 323
123
normally do not accumulate in the Columbia
ecotype (Mikkelsen et al. 2002). The relative
lower levels of aliphatic glucosinolate levels
compared to tyrosine-derived glucosinolates
obtained, most likely relicts the differences in
plasticity of the isoleucine and valine biosynthetic
pathways as compared to the shikimate pathway.
Cassava is one of the most important root
crops in the world. However, the high amounts of
the cyanogenic glycosides linamarin and lotaus-
tralin which accumulate in the starchy tubers
constitute a major health risk to the people that
daily rely on this stable crop due to the ability
of these compounds to release toxic HCN.
Accordingly, careful processing is necessary to
remove the cyanogenic glycosides from the tubers
before consumption. A major drawback of this
processing is the concurrent loss of most of the
proteins, vitamins and minerals, leaving behind a
product rich in starch, but with a very low
nutritional value. Traditional breeding has not
provided acyanogenic cassava lines. The inability
to select such lines by traditional breeding may be
a consequence of the presence of two unlinked
CYP79D paralogs and the low seed set in cassava.
Accordingly, down-regulation of the pathway by
RNAi technology is an attractive alternative.
Lines with acyanogenic (<1% of wild-type) leaves
and near-acyanogenic tubers (<8% of wild-type)
have been obtained using a single RNA interfer-
ence construct that simultaneously blocks
CYP79D1 and CYP79D2 (Jørgensen et al.
2005a; Fig. 9B). When grown in vitro, such
down-regulated lines displayed a distinct
0
500
1000
1500
2000
2500
3000
3500
4000
wt 10 4 19 8 12 #6 16 6 1 20 9 3 13 7 17 2 11 5 18 15 14 #5
% o
fthe
linam
arin
con
tent
inw
ildty
pe A
0
10
20
30
40
50
60
70
80
90
100
wt 83 98 91 84 110 108 78 94 103 90 102100 79 97 109 86 104101 93 85 99 89
% o
fthe
linam
arin
con
tent
inw
ildty
pe B
Fig. 9 Metabolic engineering of linamarin content in L.japonicus and cassava. (A) In L. japonicus, the synthesis oflinamarin has been enhanced by over-expression ofCYP79D2 from cassava under the control of the 35Spromoter as measured in apical shoots. A similar increaseis not observed for lotaustralin, because CYP79D2preferentially utilizes valine as substrate, the parent aminoacid for linamarin formation. (B) In cassava, the content of
linamarin is reduced by expression of an RNAi constructdirected against CYP79D1 and CYP79D2 under thecontrol of the enhanced 35S promoter as measured inthe first unfolded leaf. A similar reduction is observed forlotaustralin. The content of linamarin is measured by LC-MS and the change in linamarin content is calculated as %of the wild-type content
324 Phytochem Rev (2006) 5:309–329
123

morphological phenotype with long, slender
stems and long internodes as compared to in
vitro grown wild-type plants. The phenotype was
partially restored by increasing the nitrogen
concentration in the media, and completely
restored by transplanting the lines to soil. The
apparent interactions between the levels of
endogenous nitrogen and cyanogenic glycosides
are not understood. The observed conditional
phenotype indicates that cyanogenic glycosides
may possess other roles than simply being phyto-
anticipins involved in plant defence. These may
include roles as, e.g. signalling compounds or an
involvement in balancing nitrogen levels. In a
parallel approach using anti-sense technology and
the leaf-specific CAB1 promoter, transgenic
plants with reduced cyanogenic glucoside content
in leaves was also observed (Siritunga and Sayre
2003, 2004).
In conclusion, CYP79 activity regulates the flux
of intermediates through the cyanogenic gluco-
side pathway and the substrate specificity of the
CYP79s determines the cyanogenic glucoside
profile. Furthermore, CYP79s from cyanogenic
plants may be used to up- or down-regulate the
level of cyanogenic glycosides as well as to
introduce new glucosinolate profiles. Likewise,
insertion of CYP79s from the glucosinolate path-
way into cyanogenic plants may enable synthesis
of cyanogenic glycosides derived from new parent
amino acids. However, the level achieved of such
new cyanogenic glycosides would be expected to
be low because of the predictable strong interac-
tion between the endogenous cyanogenic CYP79
with the down-stream enzymes in the cyanogenic
glucoside pathway.
Concluding remarks and perspectives
The availability of P450 sequences involved in
biosynthesis of cyanogenic glycosides from an
increasing number of cyanogenic plants has
allowed a first glimpse of the molecular phylog-
eny of this ancient pathway. The addition of more
genomic, EST and cDNA sequences will help to
further our understanding at the molecular level
of how this pathway has evolved and how novel
biosynthetic pathways like the cyanoalk(en)yl,
camalexin and glucosinolate pathways have
evolved from a cyanogenic predisposition. The
poplar genome has revealed the presence of five
closely related CYP79D paralogs (Fig. 5) and the
availability of an EST corresponding to either
CYP79D5 or CYP79D7 documents that at least
some of these genes are transcribed. As cassava,
poplar belongs to Malpighiales (eurosids I) and it
is therefore tempting to suggest that poplar
contain linamarin and lotaustralin. The availabil-
ity of the genome of a putatively cyanogenic tree
model provides the necessary tools to study the
impact of cyanogenesis in a tree. Homology
modelling of cytochromes P450 will facilitate an
understanding of the importance of the unique
substitutions discovered in the PERF and heme-
binding regions of the CYP79s and contribute to
the elucidation of the unusual chemistries of the
multifunctional P450s in this pathway. Compara-
tive modelling of CYP71E1 homologs in cya-
noalkenyl and cyanogenic glucoside pathways will
provide novel information as to how new natural
products evolved at the molecular level. The
availability of genomic sequences from rice,
L. japonicus and poplar constitute important
resources for studying regulation of cyanogenic
glycosides under abiotic and biotic stress, modu-
lated by transcription factors and non-coding
RNAs. Technology platforms as untargeted trans-
criptomics and metabolomics will serve to unravel
the regulatory and metabolic networks and hubs
in interactions between primary and secondary
metabolism. This is important as compelling data
suggest that cyanogenic and cyanoalkenyl glyco-
sides may have evolved to serve other functions
than merely as phytoanticipins. The use of met-
abolic engineering of natural product profiles
using P450s provides a powerful tool to elucidate
the delicate plant–insect/microbe interactions as
well as to exploit the use of transgenic plants as
green factories for production of natural products
for the biotechnic and pharmaceutical industries.
Acknowledgements Steen Malmmose, Susanne Jensenand Charlotte Sørensen are thanked for excellent technicalassistance in the greenhouses and laboratories. All formermembers of the Cyanogenic Glycosides group are thankedfor their contributions.
Phytochem Rev (2006) 5:309–329 325
123

References
Andersen MD, Busk PK, Svendsen I, Møller BL (2000)Cytochromes P-450 from cassava (Manihot esculentaCrantz) catalyzing the first steps in the biosynthesis ofthe cyanogenic glucosides linamarin and lotaustra-lin—cloning, functional expression in Pichia pastoris,and substrate specificity of the isolated recombinantenzymes. J Biol Chem 275:1966–1975
Bak S, Feyereisen R (2001) The involvement of two P450enzymes, CYP83B1 and CYP83A1, in auxin homeo-stasis and glucosinolate biosynthesis. Plant Physiol127:108–118
Bak S, Kahn RA, Nielsen HL, Møller BL, Halkier BA(1998a) Cloning of three A-type cytochromes P450,CYP71E1, CYP98, and CYP99 from Sorghum bicolor(L.) Moench by a PCR approach and identification byexpression in Escherichia coli of CYP71E1 as amultifunctional cytochrome P450 in the biosynthesisof the cyanogenic glucoside dhurrin. Plant Mol Biol36:393–405
Bak S, Nielsen HL, Halkier BA (1998b) The presence ofCYP79 homologues in glucosinolate-producing plantsshows evolutionary conservation of the enzymes inthe conversion of amino acid to aldoxime in thebiosynthesis of cyanogenic glucosides and glucosino-lates. Plant Mol Biol 38:725–734
Bak S, Olsen CE, Petersen BL, Møller BL, Halkier BA(1999) Metabolic engineering of p-hydroxybenzylg-lucosinolate in Arabidopsis by expression of thecyanogenic CYP79A1 from Sorghum bicolor. Plant J20:663–671
Bak S, Olsen CE, Halkier BA, Møller BL (2000) Trans-genic tobacco and Arabidopsis plants expressing thetwo multifunctional sorghum cytochrome P450 en-zymes, CYP79A1 and CYP71E1, are cyanogenic andaccumulate metabolites derived from intermediates indhurrin biosynthesis. Plant Physiol 123:1437–1448
Bak S, Tax FE, Feldmann KA, Galbraith DW, Feyerei-sen R (2001) CYP83B1, a cytochrome P450 at themetabolic branch paint in auxin and indole gluco-sinolate biosynthesis in Arabidopsis. Plant Cell13:101–111
Bayburt TH, Sligar SG (2003) Self-assembly of singleintegral membrane proteins into soluble nanoscalephospholipid bilayers. Protein Sci 12:2476–2481
Busk PK, Møller BL (2002) Dhurrin synthesis in sorghumis regulated at the transcriptional level and induced bynitrogen fertilization in older plants. Plant Physiol129:1222–1231
Conn EE (1981) Cyanogenic glycosides. The biochemistryof plants, vol 7. Academic, New York, pp 479–500
Davis RH, Nahrstedt A (1987) Biosynthesis of cyanogenicglucosides in butterflies and moths—effective incor-poration of 2-methylpropanenitrile and 2-methylbu-tanenitrile into linamarin and lotaustralin by Zygaenaand Heliconius species (Lepidoptera). Insect Biochem17:689–693
Douglas GC, Zipp BJ, Ludwig-Muller J, Masuno MN,Molinski TF, Abel S (2004) Arabidopsis glucosyl-
transferase UGT74B1 functions in glucosinolatebiosynthesis and auxin homeostasis. Plant J40:893–908
Duffey SS (1981) Cyanide and arthopods. In: VenneslandB, Conn EE, Knowles CJ, Westley J, Wissing F (eds)Cyanide in biology. Academic, London, pp 385–414
Duffey SS, Towers GHN (1978) Biochemical basis of HCNproduction in millipede Harpaphe haydeniana (Xys-todesmidae: Polydesmida). Can J Zool 56:7–16
Engler HS, Spencer KC, Gilbert LE (2000) Insect metab-olism—preventing cyanide release from leaves. Nat-ure 406:144–145
Ettlinger MG, Kjær A (1968) Sulfur compounds in plants.Recent Adv Phytochem 1:49–144
Fang SD, Yan XQ, Li JF, Fan ZY, Xu XY, Xu Rs (1982)Studies on the chemical-constituents of Sedum sar-mentosum bunge. IV. The structure of sarmentosinand iso-sarmentosin. Acta Chimi Sin 40:273–280
Feyereisen R (2005) Insect cytochrome P450. In: GilbertLI, Latrou K, Gill SS (eds) Comprehensive molecularinsect science, vol 4. Elsevier, Oxford, pp 1–77
Forslund K, Morant M, Jørgensen B, Olsen CE, AsamizuE, Sato S, Tabata S, Bak S (2004) Biosynthesis of thenitrile glucosides rhodiocyanoside A and D and thecyanogenic glucosides lotaustralin and linamarin inLotus japonicus. Plant Physiol 135:71–84
Galbraith DW, Bak S (2005) Functional genomics of thecytochrome P450 gene superfamily in Arabidopsisthaliana. In: Leister D (ed) Plant functional genomics.Haworth, Binghamton, pp 595–620
Glawischnig E, Hansen BG, Olsen CE, Halkier BA (2004)Camalexin is synthesized from indole-3-acetaldoxime,a key branching point between primary and secondarymetabolism in Arabidopsis. Proc Natl Acad Sci USA101:8245–8250
Gleadow RM, Woodrow IE (2000) Temporal and spatialvariation in cyanogenic glycosides in Eucalyptuscladocalyx. Tree Physiol 20:591–598
Gleadow RM, Woodrow IE (2002) Defense chemistry ofcyanogenic Eucalyptus cladocalyx seedlings is af-fected by water supply. Tree Physiol 22:939–945
Halkier BA, Gershenzon J (2006) Biology and biochemistryof glucosinolates. Annu Rev Plant Biol 57:303–333
Halkier BA, Møller BL (1991) Involvement of cytochromeP-450 in the biosynthesis of dhurrin in Sorghumbicolor (L.) Moench. Plant Physiol 96:10–17
Halkier BA, Nielsen HL, Koch B, Møller BL (1995)Purification and characterization of recombinantcytochrome P450TYR expressed at high levels inEscherichia coli. Arch Biochem Biophys 322:369–377
Halkier BA, Sibbesen O, Møller BL (1996) Isolation ofplant and recombinant CYP79. Methods Enzymol272:268–274
Hansen CH, Du LC, Naur P, Olsen CE, Axelsen KB, HickAJ, Pickett JA, Halkier BA (2001) CYP83B1 is theoxime-metabolizing enzyme in the glucosinolate path-way in Arabidopsis. J Biol Chem 276:24790–24796
Hansen KS, Kristensen C, Tattersall DB, Jones PR, OlsenCE, Bak S, Møller BL (2003) The in vitro substrateregiospecificity of recombinant UGT85B1, the cyano-
326 Phytochem Rev (2006) 5:309–329
123

hydrin glucosyltransferase from Sorghum bicolor.Phytochemistry 64:143–151
Helliwell CA, Sheldon CC, Olive MR, Walker AR,Zeevaart JA, Peacock WJ, Dennis ES (1998) Cloningof the Arabidopsis ent-kaurene oxidase gene GA3.Proc Natl Acad Sci USA 95:9019–9024
Hemm MR, Ruegger MO, Chapple C (2003) The Arabid-opsis ref2 mutant is defective in the gene encodingCYP83A1 and shows both phenylpropanoid andglucosinolate phenotypes. Plant Cell 15:179–194
Herrmann KM (1995) The shikimate pathway: early stepsin the biosynthesis of aromatic compounds. Plant Cell7:907–919
Holzkamp G, Nahrstedt A (1994) Biosynthesis of cyano-genic glucosides in the lepidoptera—incorporationof [U-C-14]-2-methylpropanealdoxime, 2S-[U-C-14]-methylbutanealdoxime and D,L-[U-C-14]-N-hydroxy-isoleucine into linamarin and lotaustralin by thelarvae of Zygaena trifolii. Insect Biochem Mol Biol24:161–165
Humphreys JM, Chapple C (2002) Rewriting the ligninroadmap. Curr Opin Plant Biol 5:224–229
Jaroszewski JW, Olafsdottir ES, Wellendorph P, Chris-tensen J, Franzyk H, Somanadhan B, Budnik BA,Jørgensen LB, Clausen V (2002a) Cyanohydrin gly-cosides of Passiflora: distribution pattern, a saturatedcyclopentane derivative from P. guatemalensis, andformation of pseudocyanogenic alpha-hydroxyamidesas isolation artefacts. Phytochemistry 59:501–511
Jaroszewski JW, Olafsdottir ES, Wellendorph P, Christen-sen J, Franzyk H, Somanadhan B, Budnik BA, Jørgen-sen LB, Clausen V (2002b) Natural cyclopentanoidcyanohydrin glycosides, part 23. Cyanohydrin glyco-sides of Passiflora: distribution pattern, a saturatedcyclopentane derivative from P. guatemalensis, andformation of pseudocyanogenic alpha-hydroxyamidesas isolation artefacts. Phytochemistry 59:501–511
Jones PR, Møller BL, Hoj PB (1999) The UDP-glucose:p-hydroxymandelonitrile-O-glucosyltransferase thatcatalyzes the last step in synthesis of the cyanogenicglucoside dhurrin in Sorghum bicolor. Isolation,cloning, heterologous expression, and substrate spec-ificity. J Biol Chem 274:35483–35491
Jones PR, Andersen MD, Nielsen JS, Høj PB, Møller BL(2000) The biosynthesis, degradation, transport andpossible functions of cyanogenic glucosides. In:Romeo JT (eds) Recent advances in phytochemistry:evolution of metabolic pathways. Pergamon, Amster-dam, pp 191–247
Jørgensen K, Bak S, Busk PK, Sorensen C, Olsen CE,Puonti-Kaerlas J, Møller BL (2005a) Cassava plantswith a depleted cyanogenic glucoside content inleaves and tubers. Distribution of cyanogenic gluco-sides, their site of synthesis and transport, andblockage of the biosynthesis by RNA interferencetechnology. Plant Physiol 139:363–374
Jørgensen K, Rasmussen AV, Morant M, Nielsen AH,Bjarnholt N, Zagrobelny M, Bak S, Møller BL(2005b) Metabolon formation and metabolic channel-ing in the biosynthesis of plant natural products. CurrOpin Plant Biol 8:280–291
Kahn RA, Bak S, Svendsen I, Halkier BA, Møller BL(1997) Isolation and reconstitution of cytochromeP450ox and in vitro reconstitution of the entirebiosynthetic pathway of the cyanogenic glucosidedhurrin from sorghum. Plant Physiol 115:1661–1670
Klein M, Reichelt M, Gershenzon J, Papenbrock J (2006)The three desulfoglucosinolate sulfotransferaseproteins in Arabidopsis have different substratespecificities and are differentially expressed. FEBSJ 273:122–136
Koch BM, Sibbesen O, Halkier BA, Svendsen I, MøllerBL (1995) The primary sequence of cytochromeP450tyr, the multifunctional N-hydroxylase catalyzingthe conversion of L-tyrosine to p-hydroxyphenylacet-aldehyde oxime in the biosynthesis of the cyanogenicglucoside dhurrin in Sorghum bicolor (L.) Moench.Arch Biochem Biophys 323:177–186
Kristensen C, Morant M, Olsen CE, Ekstrom CT, Gal-braith DW, Møller BL, Bak S (2005) Metabolicengineering of dhurrin in transgenic Arabidopsisplants with marginal inadvertent effects on themetabolome and transcriptome. Proc Natl Acad SciUSA 102:1779–1784
Kumar S, Tamura K, Nei M (2004) MEGA3: integratedsoftware for molecular evolutionary genetics analysisand sequence alignment. Brief Bioinform 5:150–163
Kutchan TM (2005) Predictive metabolic engineering inplants: still full of surprises. Trends Biotechnol23:381–383
Labandeira CC (1993) The real meaning of insect fossils.Palaios 8:509–511
Labandeira CC, Dilcher DL, Davis DR, Wagner DL(1994) 97-Million years of angiosperm–insect associ-ation—paleobiological insights into the meaning ofcoevolution. Proc Natl Acad Sci USA 91:12278–12282
Labandeira CC, Sepkoshi JJ Jr (1993) Insect diversity inthe fossil record. Science 261:310–315
Lechtenberg M, Nahrstedt A (1999) Cyanogenic glyco-sides. In: Ikan R (ed) Naturally occurring glycosides.Wiley, New York, pp 147–191
Memelink J (2005) Tailoring the plant metabolomewithout a loose stitch. Trends Plant Sci 10:305–307
Mikkelsen MD, Halkier BA (2003) Metabolic engineeringof valine- and isoleucine-derived glucosinolates inArabidopsis expressing CYP79D2 from cassava. PlantPhysiol 131:773–779
Mikkelsen MD, Petersen BL, Olsen CE, Halkier BA(2002) Biosynthesis and metabolic engineering ofglucosinolates. Amino Acids 22:279–295
Mikkelsen MD, Naur P, Halkier BA (2004) Arabidopsismutants in the C–S lyase of glucosinolate biosynthesisestablish a critical role for indole-3-acetaldoxime inauxin homeostasis. Plant J 37:770–777
Møller BL, Conn EE (1980) The biosynthesis of cyano-genic glucosides in higher plants. Channeling ofintermediates in dhurrin biosynthesis by a microsomalsystem from Sorghum bicolor (linn) Moench. J BiolChem 255:3049–3056
Møller BL, Seigler DS (1998) Biosynthesis of cyanogenicglucosides, cyanolipids, and related compounds. In:Singh BK (eds) Plant amino acids, biochemsitry and
Phytochem Rev (2006) 5:309–329 327
123

biotechnology. Marcel Dekker Inc., New York, pp563–609
Morant M, Bak S, Møller BL, Werck-Reichhart D (2003)Plant cytochromes P450: tools for pharmacology,plant protection and phytoremediation. Curr OpinBiotechnol 14:151–162
Naur P, Petersen BL, Mikkelsen MD, Bak S, RasmussenH, Olsen CE, Halkier BA (2003) CYP83A1 andCYP83B1, two nonredundant cytochrome P450 enzy-mes metabolizing oximes in the biosynthesis ofglucosinolates in Arabidopsis. Plant Physiol 133:63–72
Nelson DR, Koymans L, Kamataki T, Stegeman JJ,Feyereisen R, Waxman DJ, Waterman MR, GotohO, Coon MJ, Estabrook RW, Gunsalus IC, NebertDW (1996) P450 superfamily: update on newsequences, gene mapping, accession numbers andnomenclature. Pharmacogenetics 6:1–42
Nelson DR, Schuler MA, Paquette SM, Werck-ReichhartD, Bak S (2004) Comparative genomics of rice andArabidopsis. Analysis of 727 cytochrome P450 genesand pseudogenes from a monocot and a dicot. PlantPhysiol 135:756–772
Nielsen JS, Møller BL (2000) Cloning and expression ofcytochrome P450 enzymes catalyzing the conversionof tyrosine to p-hydroxyphenylacetaldoxime in thebiosynthesis of cyanogenic glucosides in Triglochinmaritima. Plant Physiol 122:1311–1321
Nielsen KA, Møller BL (2004) Cytochromes P450 inPlants. In: Ortiz de Montellano PR (ed) CytochromeP450: structure, mechanism, and biochemistry, 3rdedn. Kluwer Academic/Plenum Publishers, NewYork, pp 553–583
Nielsen KA, Olsen CE, Pontoppidan K, Møller BL (2002)Leucine-derived cyano glucosides in barley. PlantPhysiol 129:1066–1075
Paquette S, Møller BL, Bak S (2003) On the origin of thefamily 1 plant glycosyltransferases. Phytochemisty62:399–413
Petersen BL, Andreasson E, Bak S, Agerbirk N, HalkierBA (2001) Characterization of transgenic Arabidopsisthaliana with metabolically engineered high levels ofp-hydroxybenzylglucosinolate. Planta 212:612–618
Piotrowski M, Schemenewitz A, Lopukhina A, Muller A,Janowitz T, Weiler EW, Oecking C (2004) Desul-foglucosinolate sulfotransferases from Arabidopsisthaliana catalyze the final step in the biosynthesisof the glucosinolate core structure. J Biol Chem279:50717–50725
Poulton J, Møller BL (1989) The biosynthesis of cyano-genic glucosides. In: Dey PM, Harborne JB (eds)Methods in plant biochemistry, vol 9. Academic,London, pp 183–207
Regier JC, Shultz JW, Kambic RE (2005) Pancrustaceanphylogeny: hexapods are terrestrial crustaceans andmaxillopoda are not monophyletic. Proc R Soc B272:395–401
Rodman JE, Soltis PS, Soltis DE, Sytsma KJ, Karol KG(1998) Parallel evolution of glucosinolate biosynthesisinferred from congruent nuclear and plastid genephylogenies. Am J Bot 85:997–1006
Schuhegger R, Nafisi M, Mansourova M, PetersenBL, Olsen CE, Svatos A, Halkier BA, GlawischnigE (2006) CYP71B15 (PAD3) catalyzes the finalstep in camalexin biosynthesis. Plant Physiol141:1248–1254
Scott EE, He YA, Wester MR, White MA, Chin CC,Halpert JR, Johnson EF, Stout CD (2003) An openconformation of mammalian cytochrome P4502B4 at1.6-angstrom resolution. Proc Natl Acad Sci USA100:13196–13201
Selmar D, Lieberei R, Biehl B (1988) Mobilization andutilization of cyanogenic glycosides: the linustatinpathway. Plant Physiol 86:711–716
Sibbesen O, Koch B, Halkier BA, Møller BL (1994)Isolation of the heme-thiolate enzyme cytochrome P-450TYR, which catalyzes the committed step in thebiosynthesis of the cyanogenic glucoside dhurrin inSorghum bicolor (L.) Moench. Proc Natl Acad SciUSA 91:9740–9744
Sibbesen O, Koch B, Halkier BA, Møller BL (1995)Cytochrome P-450TYR is a multifunctional heme-thiolate enzyme catalyzing the conversion of L-tyrosine to p-hydroxyphenylacetaldehyde oxime inthe biosynthesis of the cyanogenic glucoside dhurrinin Sorghum bicolor (L.) Moench. J Biol Chem270:3506–3511
Siegler DS, Brinker AM (1993) Characterisation of cya-nogenic glycosides, cyanolipids, nitroglycosides, or-ganic nitro compounds and nitrile glycosides fromplants. In: Dey PM, Harborne JB (eds) Methods ofplant biochemistry, alkaloids and sulfur compounds.Academic, New York, pp 51–93
Siritunga D, Sayre RT (2003) Generation of cyanogen-freetransgenic cassava. Planta 217:367–373
Siritunga D, Sayre R (2004) Engineering cyanogen syn-thesis and turnover in cassava (Manihot esculenta).Plant Mol Biol 56:661–669
Tattersall DB, Bak S, Jones PR, Olsen CE, Nielsen JK,Hansen ML, Hoj PB, Møller BL (2001) Resistance toan herbivore through engineered cyanogenic gluco-side synthesis. Science 293:1826–1828
Thorsoe KS, Bak S, Olsen CE, Imberty A, Breton C,Møller BL (2005) Determination of catalytic keyamino acids and UDP sugar donor specificity of thecyanohydrin glycosyltransferase UGT85B1 from Sor-ghum bicolor. Molecular modeling substantiated bysite-specific mutagenesis and biochemical analyses.Plant Physiol 139:664–673
Werck-Reichhart D, Bak S, Paquette SM (2002) Cyto-chrome P450. The Arabidopsis book. American Soci-ety of Plant Biologists, Rockville, pp 1–29
Williams PA, Cosme J, Ward A, Angova HC, VinkovicDM, Jhoti H (2003) Crystal structure of humancytochrome P4502C9 with bound warfarin. Nature424:464–468
Williams PA, Cosme J, Vinkovic DM, Ward A, AngoveHC, Day PJ, Vonrhein C, Tickle IJ, Jhoti H (2004)Crystal structures of human cytochrome P450 3A4bound to metyrapone and progesterone. Science305:683–686
328 Phytochem Rev (2006) 5:309–329
123

Willis KJ, McElwain JC (2002) The evolution of plants.Oxford University Press Inc., New York
Winkel BS (2004) Metabolic channeling in plants. AnnuRev Plant Biol 55:85–107
Woodward AW, Bartel B (2005) Auxin: regulation, action,and interaction. Ann Bot (Lond) 95:707–735
Yoshikawa M, Shimada H, Shimoda H, Matsuda H,Yamahara J, Murakami N (1995) Rhodiocyanoside-A and rhodiocyanoside-B, new antiallergic cyanogly-cosides from Chinese natural medicine Si-Li-Hong-Jing-Tian, the underground part of Rhodiolaquadrifida (Pall) Fisch et Mey. Chem Pharm Bull43:1245–1247
Yoshikawa M, Shimada H, Matsuda H, Yamahara J,Murakami N (1996a) Bioactive constituents of Chi-nese natural medicines. 1. New sesquiterpene ketoneswith vasorelaxant effect from Chinese moxa, theprocessed leaves of Artemisia argyi Levl et Vant:moxartenone and moxartenolide. Chem Pharm Bull44:1656–1662
Yoshikawa M, Shimada H, Shimoda H, Murakami N,Yamahara J, Matsuda H (1996b) Bioactive constitu-ents of Chinese natural medicines. 2. Rhodiolae radix.1. Chemical structures and antiallergic activity ofrhodiocyanosides A and B from the underground partof Rhodiola quadrifida (Pall) Fisch et MEY (Crassul-aceae). Chem Pharm Bull 44:2086–2091
Zagrobelny M, Bak S, Rasmussen AV, Jørgensen B,Naumann CM, Møller BL (2004) Cyanogenic gluco-sides and plant–insect interactions. Phytochemistry65:293–306
Zagrobelny M, Bak S, Ekstrøm CT, Oslen CE, MøllerBL: Cyanogenic glycosides in Zygaena filipendulae(Lepidoptera: Zygaenidae) as effected by feeding onwild-type and transgenic Lotus populations withvariable cyanogenic profiles. Insect Biochemistryand Molecular Biology (in press)
Phytochem Rev (2006) 5:309–329 329
123




















