
Effects of Fabric Counts and Weave Designs on the Properties ...
Upload
independentCategory
view
0download
0
www.elsevier.com/locate/yviro
Virology 330 (20
Influence of CD4+ T cell counts on viral evolution in HIV-infected
individuals undergoing suppressive HAART
Eric Lorenzoa,*, Maria C. Colona, Sharilyn Almodovara, Irvin M. Maldonadoa, Sandra Gonzaleza,
Sonia E. Costaa, Martin D. Hilla, Rafael Mendozab, Gladys Sepulvedab, Richard Yanagiharac,
Vivek Nerurkarc, Rakesh Kumara, Yasuhiro Yamamuraa, Walter A. Scottd, Anil Kumara
aAIDS Research Program, Ponce School of Medicine, Ponce 00732, Puerto RicobDepartment of Health Immunology Clinic, Ponce 00731, Puerto Rico
cRetrovirology Research Laboratory, University of Hawaii at Manoa, Hawaii 96816, USAdDepartment of Biochemistry and Molecular Biology, University of Miami 33124, USA
Received 19 June 2004; returned to author for revision 6 August 2004; accepted 3 September 2004
Available online 14 October 2004
Abstract
We analyzed the viral C2–V4 envelope diversity, glycosylation patterns, and dS/dN ratios of plasma HIV-1 in an attempt to better
understand the complex interaction between viral quasispecies and the host-selective pressures pre- and post-HAART. Phylogenetic analysis
of the envelope gene of five patients revealed monophyletic clustering in patients with higher CD4+ T cell counts and sequence intermingling
in those with lower CD4+ T cells in relation to the stage of HAART. Our analyses also showed clear shifts in N-linked glycosylation patterns
in patients with higher CD4+ T cells, suggesting possible distinct immunological pressures pre- and post-HAART. The relative preponderance
of synonymous/nonsynonymous changes in the envelope region suggested a positive selection in patients with higher CD4+ T cells, whereas
lack of evidence for positive selection was found in the patients with lower CD4+ T cells. An exception to the last analysis occurred in the
only patient who reached complete viral suppression, maybe due to drug pressure exerted over the pol gene that may obscure the immune
pressure/selection at the envelope in this analysis. All these indications may suggest that even when HAART generates viral suppression,
quasispecies evolve in the envelope gene probably resulting from host-selective pressure.
D 2004 Elsevier Inc. All rights reserved.
Keywords: HIV; Envelope; HAART; CD4+ T cell counts; Phylogeny; N-linked glycosylation; Synonymous and nonsynonymous nucleotide substitution
Introduction
Human immunodeficiency virus type 1 (HIV-1) repli-
cation is characterized by a high degree of viral sequence
variation, primarily due to mutations introduced by the
error-prone reverse transcriptase that leads into development
of distinct viral quasispecies. Therefore, even in a single
individual, the virus exists as highly related, although
genetically distinct quasispecies (Kuiken et al., 1993;
0042-6822/$ - see front matter D 2004 Elsevier Inc. All rights reserved.
doi:10.1016/j.virol.2004.09.015
* Corresponding author. AIDS Research Program, Molecular Virology
Laboratory, Department of Biochemistry, Ponce School of Medicine, PO
Box 7004, Ponce, 00732, Puerto Rico. Fax: +1 787 841 1040.
E-mail address: [email protected] (E. Lorenzo).
Lorenzo et al., 2004; Malim and Emerman, 2001; Nowak
et al., 1991). Phylogenetic methodologies have been
extensively used to reconstruct an evolutionary history of
these diverse viral quasispecies (DeBry et al., 1993; Hillis et
al., 1994; Lorenzo et al., 2001, 1996; Ou et al., 1992). A
critical consequence of HIV-1 variability has been deter-
mined to be high degree of viral sequence diversity in the
envelope gene that results in the co-receptor usage shift.
HIV-1 co-receptor usage has been shown to be an important
determinant of viral tropism and pathogenesis. During acute
infection, the virus uses CCR5 co-receptor (R5) in vivo,
whereas later in the infection, CXCR4-utilizing virus (X4)
appears mixed with R5 quasispecies (Connor et al., 1997;
Stevenson, 2003). The emergence of X4 utilizing strains
04) 116–126

E. Lorenzo et al. / Virology 330 (2004) 116–126 117
presages cell depletion and clinical deterioration (Philpott et
al., 2001; Stevenson, 2003). Genetic analysis of patient viral
populations such as the study presented here has been
extensively used to gain information on the pathogenesis of
the infection as well as to clarify in vivo evolution of HIV-1
(Delwart et al., 1997; Shankarappa et al., 1999). Never-
theless, the exact nature and significance of the mechanisms
that govern intra-host evolution and pathogenesis remain
largely unknown.
The HIV evasive ability to counter the immune system
has also been associated in part with both the rapid
variability of the HIV envelope protein sequence and the
masking of epitopes by glycosylation (Nara et al., 1991;
Quinones-Kochs et al., 2002; Wei et al., 2003). The N-
glycosylation sites are significant for the binding of
carbohydrates to the viral envelope to mask viral protein
epitopes from the immune response, and thus enabling the
HIV-1 to escape neutralization. Indeed, one of the most
important functions of N-linked glycosylation is protection
of otherwise accessible neutralization epitopes of the viral
envelope from neutralizing antibodies (Nab) (Bolmstedt et
al., 1996). Recently, it has been shown that mutations in N-
linked glycosylation sites rendered a neutralization antibody
sensitive virus to be completely resistant to pre-existing
neutralizing antibodies (Stipp et al., 2000; Wei et al., 2003).
This pattern of escape led these researchers to postulate an
bevolving glycan shieldQ mechanism of neutralization
escape in which selected changes in glycosylation patterns
prevent Nab but not receptor binding (Wei et al., 2003). This
evolving glycosylation pattern of the envelope gene
represents a new mechanism contributing to HIV-1 persis-
tence in the face of a dynamic antibody repertoire. Although
the number of N-linked glycosylation sites varies with the
strain of HIV-1, there are usually 24 such potential sites in
the envelope gene (Quinones-Kochs et al., 2002), and their
somewhat conserved locations suggest generally strong and
persistent selective pressures. Clear changes in these
glycosylation sites will suggest shifts in immune pressure
(Kemal et al., 2003; Wei et al., 2003).
The majority of positive selection in the HIV-1 has been
found to occur in the envelope region of the genome, as
opposed to the gag or pol region (Choisy et al., 2004).
Positive selection in the HIV-1 env gene has been
demonstrated by comparisons of the abundance of non-
synonymous (dN) and synonymous (dS) pairwise variability
in the DNA sequence (Bonhoeffer et al., 1995; Nielsen,
1999). The relative ratio of nonsynonymous and synon-
ymous changes in the HIV env gene seems to be generated
by a balance between selective pressure placed on the virus
quasispecies mainly by the immune system, which favors
amino acid diversification, and the negative selection
against missense mutation that breaks the structural con-
straints of the functional Env protein (Anastassopoulou et
al., 2003; Shankarappa et al., 1999).
Patients under HAART have been extensively followed
for the emergence of drug-resistance by sequencing pol and
identifying the associated mutations (Demeter et al., 1998;
Gallant et al., 2003; Little et al., 2002; Tirado et al., 2004;
Unal et al., 1996). However, there is only limited
information available regarding the evolution of other viral
genes in patients on HAART. Interestingly, the envelope
gene, one of the most immunogenic regions of the HIV-1, is
linked to numerous viral characteristics including trans-
mission, viral entry, co-receptor usage, tropism, and immune
escape (Rangel et al., 2003). However, few research studies
have been focused in the envelope gene of the HIV-1 after
the advent of HAART. In an effort to improve our
understanding of the complex interaction between viral
quasispecies and the host-selective immune pressure, we
analyzed the env gene viral diversity, glycosylation patterns,
and selective pressures in plasma pre- and post-HAART.
Results
We sequenced and analyzed HIV-1 C2–V4 env of 148
clones from plasma virus in five patients who were sampled
before receiving any antiretroviral drug therapy (time 1) and
after achieving viral suppression due to the therapeutic
regimen (time 2) (Fig. 1). Patients A and B were clinically
classified as categories A and B (CDC, 1993), respectively,
with N400 CD4+ T cell counts; and patients C, D, and E
were category C with CD4+ T cell counts b200 cells/ml
blood, at the time of enrollment. All participants exhibited
viral suppression 6–10 months after commencement of
HAART. Patients A and E showed emergence of rebound-
ing virus after initial suppression (time 3), whereas patients
B, C, and D maintained undetectable plasma viral loads
throughout study (Table 1). Viral sequences at second time
point for patient A are unavailable for this study due to the
limited volume of plasma accessible for the amplification of
HIV-1 RNA. We observed T-tropic variants during
advanced disease of patients C and E, in agreement with
Kreisberg et al. (2001) and Schuitemaker et al. (1992).
Utilizing most stringent genotyping method (Jensen et al.,
2003), we observed disappearance of these T-tropic variants
after HAART, in agreement with Philpott et al. (2001) and
Skrabal et al. (2003).
Phylogenetic reconstructions pre- and post-HAART
Phylogenetic analysis by the NJ method of the plasma
quasispecies in the five HIV-1-infected participants showed
monophyletic groups that were separated from nucleotide
sequences of the other patients with 100% bootstrap values,
excluding the possibility of laboratory contamination (Fig.
2). There was a correlation between the clustering tendency
of sequences and the CD4+ T cell count profile in four of five
patients. NJ phylogenetic reconstruction provided evidence
of clear temporal clustering of the C2–V4 env sequences pre-
and post-HAART in patients A, B, and C. On the other hand,
sequence intermingling was observed in patients D and E in
E. Lorenzo et al. / Virology 330 (2004) 116–126118
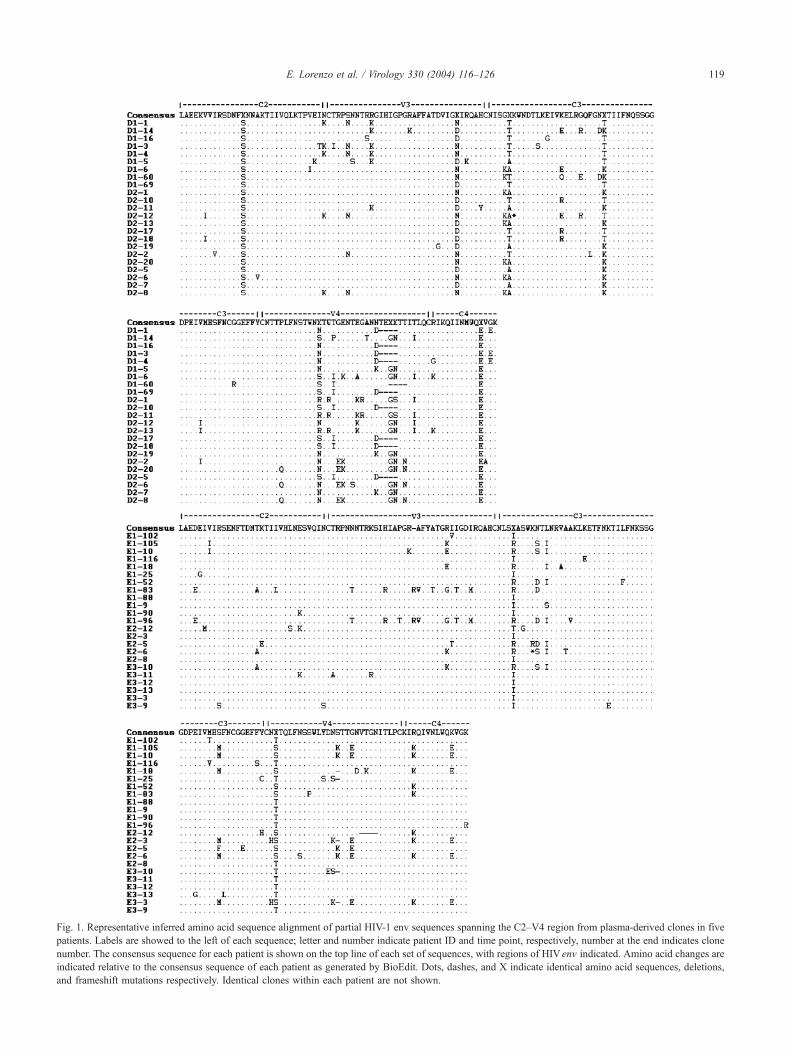
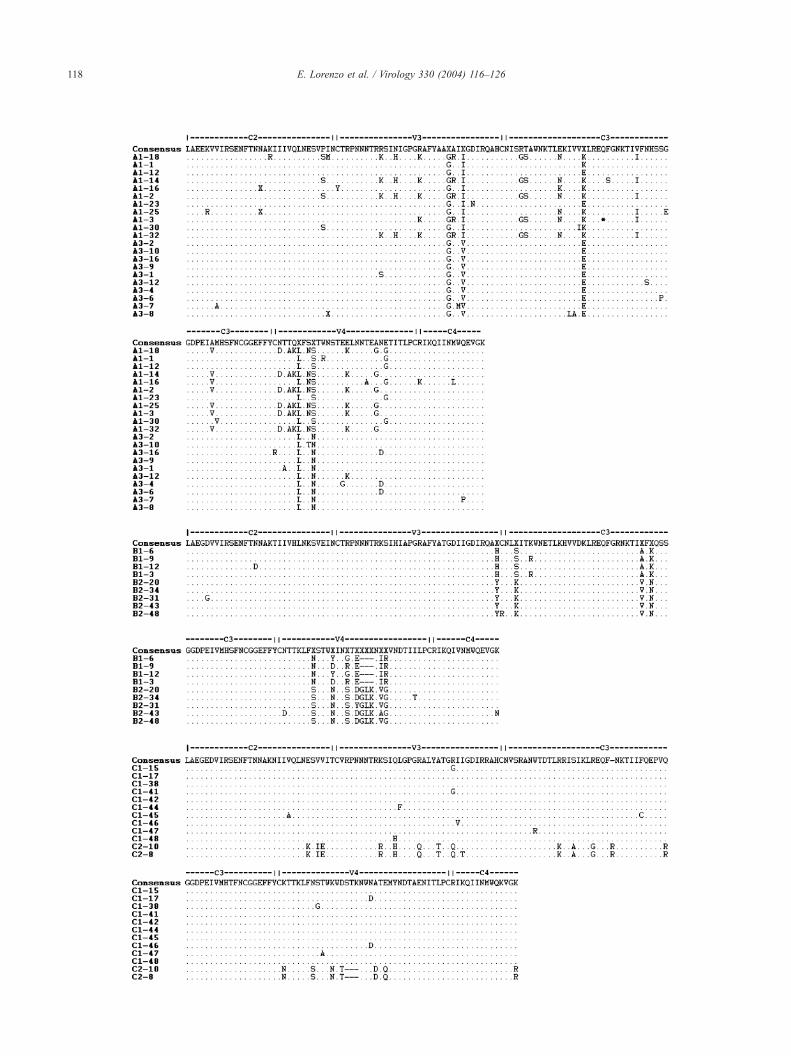
Fig. 1. Representative inferred amino acid sequence alignment of partial HIV-1 env sequences spanning the C2–V4 region from plasma-derived clones in five
patients. Labels are showed to the left of each sequence; letter and number indicate patient ID and time point, respectively, number at the end indicates clone
number. The consensus sequence for each patient is shown on the top line of each set of sequences, with regions of HIV env indicated. Amino acid changes are
indicated relative to the consensus sequence of each patient as generated by BioEdit. Dots, dashes, and X indicate identical amino acid sequences, deletions,
and frameshift mutations respectively. Identical clones within each patient are not shown.
E. Lorenzo et al. / Virology 330 (2004) 116–126 119
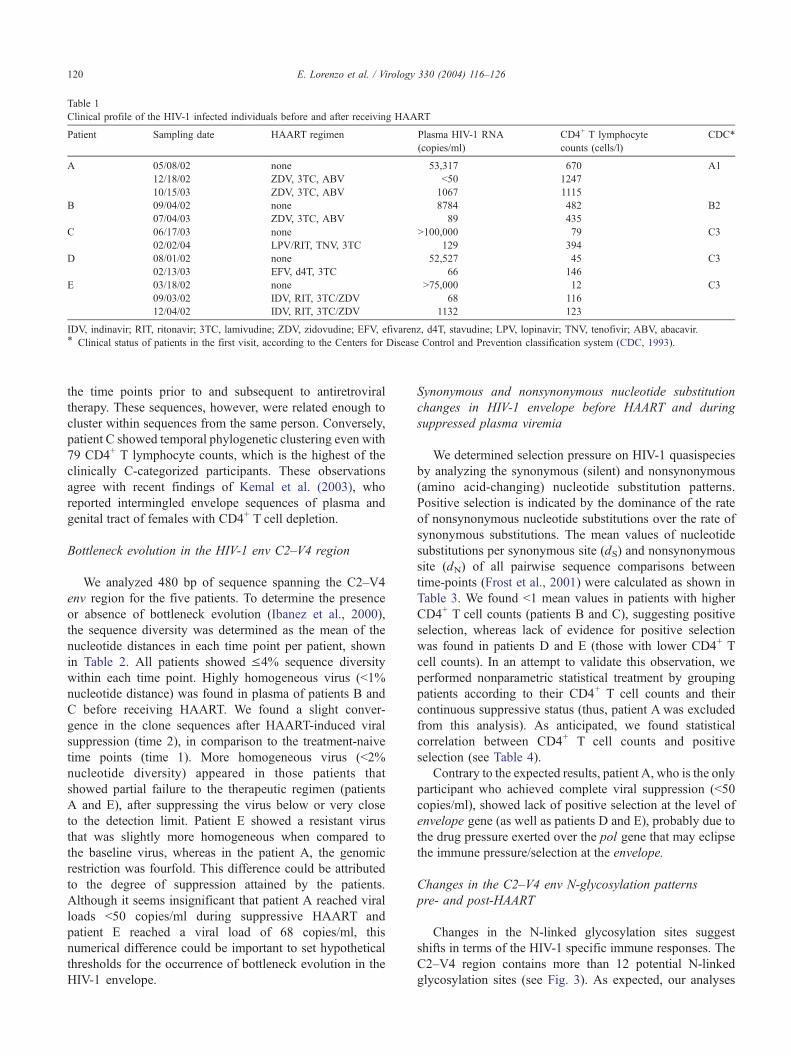
Table 1
Clinical profile of the HIV-1 infected individuals before and after receiving HAART
Patient Sampling date HAART regimen Plasma HIV-1 RNA
(copies/ml)
CD4+ T lymphocyte
counts (cells/l)
CDC*
A 05/08/02 none 53,317 670 A1
12/18/02 ZDV, 3TC, ABV b50 1247
10/15/03 ZDV, 3TC, ABV 1067 1115
B 09/04/02 none 8784 482 B2
07/04/03 ZDV, 3TC, ABV 89 435
C 06/17/03 none N100,000 79 C3
02/02/04 LPV/RIT, TNV, 3TC 129 394
D 08/01/02 none 52,527 45 C3
02/13/03 EFV, d4T, 3TC 66 146
E 03/18/02 none N75,000 12 C3
09/03/02 IDV, RIT, 3TC/ZDV 68 116
12/04/02 IDV, RIT, 3TC/ZDV 1132 123
IDV, indinavir; RIT, ritonavir; 3TC, lamivudine; ZDV, zidovudine; EFV, efivarenz, d4T, stavudine; LPV, lopinavir; TNV, tenofivir; ABV, abacavir.* Clinical status of patients in the first visit, according to the Centers for Disease Control and Prevention classification system (CDC, 1993).
E. Lorenzo et al. / Virology 330 (2004) 116–126120
the time points prior to and subsequent to antiretroviral
therapy. These sequences, however, were related enough to
cluster within sequences from the same person. Conversely,
patient C showed temporal phylogenetic clustering even with
79 CD4+ T lymphocyte counts, which is the highest of the
clinically C-categorized participants. These observations
agree with recent findings of Kemal et al. (2003), who
reported intermingled envelope sequences of plasma and
genital tract of females with CD4+ T cell depletion.
Bottleneck evolution in the HIV-1 env C2–V4 region
We analyzed 480 bp of sequence spanning the C2–V4
env region for the five patients. To determine the presence
or absence of bottleneck evolution (Ibanez et al., 2000),
the sequence diversity was determined as the mean of the
nucleotide distances in each time point per patient, shown
in Table 2. All patients showed V4% sequence diversity
within each time point. Highly homogeneous virus (b1%
nucleotide distance) was found in plasma of patients B and
C before receiving HAART. We found a slight conver-
gence in the clone sequences after HAART-induced viral
suppression (time 2), in comparison to the treatment-naive
time points (time 1). More homogeneous virus (b2%
nucleotide diversity) appeared in those patients that
showed partial failure to the therapeutic regimen (patients
A and E), after suppressing the virus below or very close
to the detection limit. Patient E showed a resistant virus
that was slightly more homogeneous when compared to
the baseline virus, whereas in the patient A, the genomic
restriction was fourfold. This difference could be attributed
to the degree of suppression attained by the patients.
Although it seems insignificant that patient A reached viral
loads b50 copies/ml during suppressive HAART and
patient E reached a viral load of 68 copies/ml, this
numerical difference could be important to set hypothetical
thresholds for the occurrence of bottleneck evolution in the
HIV-1 envelope.
Synonymous and nonsynonymous nucleotide substitution
changes in HIV-1 envelope before HAART and during
suppressed plasma viremia
We determined selection pressure on HIV-1 quasispecies
by analyzing the synonymous (silent) and nonsynonymous
(amino acid-changing) nucleotide substitution patterns.
Positive selection is indicated by the dominance of the rate
of nonsynonymous nucleotide substitutions over the rate of
synonymous substitutions. The mean values of nucleotide
substitutions per synonymous site (dS) and nonsynonymous
site (dN) of all pairwise sequence comparisons between
time-points (Frost et al., 2001) were calculated as shown in
Table 3. We found b1 mean values in patients with higher
CD4+ T cell counts (patients B and C), suggesting positive
selection, whereas lack of evidence for positive selection
was found in patients D and E (those with lower CD4+ T
cell counts). In an attempt to validate this observation, we
performed nonparametric statistical treatment by grouping
patients according to their CD4+ T cell counts and their
continuous suppressive status (thus, patient A was excluded
from this analysis). As anticipated, we found statistical
correlation between CD4+ T cell counts and positive
selection (see Table 4).
Contrary to the expected results, patient A, who is the only
participant who achieved complete viral suppression (b50
copies/ml), showed lack of positive selection at the level of
envelope gene (as well as patients D and E), probably due to
the drug pressure exerted over the pol gene that may eclipse
the immune pressure/selection at the envelope.
Changes in the C2–V4 env N-glycosylation patterns
pre- and post-HAART
Changes in the N-linked glycosylation sites suggest
shifts in terms of the HIV-1 specific immune responses. The
C2–V4 region contains more than 12 potential N-linked
glycosylation sites (see Fig. 3). As expected, our analyses

Fig. 2. Phylogenetic reconstruction of HIV-1 plasma-derived gp120 env C2–V4 nucleotide sequences generated by the neighbor-joining (NJ) method. Numbers
at branch nodes refer to bootstrap values (1000 repetitions). Letters and number correspond to the patient ID and time point, respectively. The number at the end
refers to the clone number. Identical clones were eliminated from this analysis.
E. Lorenzo et al. / Virology 330 (2004) 116–126 121

Table 2
Nucleotide distances of plasma-derived clone sequences of five patients
before and after receiving HAART
Patient Sampling
time
Mean diversity F SD Wilcoxon’s signed
rank test*
A 1 0.037 F 0.024 b0.001
3 0.010 F 0.010
B 1 0.006 F 0.003 0.005
2 0.004 F 0.004
C 1 0.007 F 0.007 0.031
2 0.003 F 0.002
D 1 0.004 F 0.014 0.003
2 0.033 F 0.012
E 1 0.033 F 0.020 0.012
2 0.035 F 0.013
3 0.022 F 0.011
Diversity was determined by using MEGA with Kimura 2-parameter. SD,
standard deviation.* P value.
E. Lorenzo et al. / Virology 330 (2004) 116–126122
showed that in patients with higher CD4+ T cell counts
(patients A–C), there was a clear shift in terms of
glycosylation sites before and during HAART, suggesting
selective immune pressures. Conversely, in patients E and
D, there was indication of poor selective immune pressure
patients, consistent with their lower CD4+ T cell counts.
Discussion
The present study characterized different quasispecies of
the virus in five HIV-infected individuals. The gene we
analyzed codes for part of the envelope, which is not the
target of HAART regimes of our cohort, implying that we
are primarily measuring the result of a selective pressure
other than HAART. Our results demonstrate the possibility
of involvement of immune pressure in the selection of viral
variants that has been shown by several studies. However,
this is the first indication that during HAART viral evolution
is influenced by not only drug pressure but also immune
pressure in patients with higher CD4+ counts. This
Table 3
Mean synonymous and nonsynonymous nucleotide substitution changes in differ
Patient Sampling
time
Mean dS F SD Wilcoxon’s
signed rank test*
Mean dN
A 1 0.060 F 0.029 0.890 0.400 F3 0.072 F 0.043 0.042 F
B 1 0.013 F 0.005 0.007 0.009 F2 0.015 F 0.006 0.031 F
C 1 0.152 F 0.007 b0.001 0.007 F2 0.038 F 0.010 0.051 F
D 1 0.075 F 0.021 0.081 0.035 F2 0.066 F 0.020 0.031 F
E 1 0.054 F 0.026 0.221 0.037 F2 0.0478 F 0.020 0.37 F3 0.573 F 0.994 b0.001 0.028 F
The frequencies of synonymous substitutions per synonymous site (dS) and no
Synonymous/Nonsynonymous Analysis Program (SNAP) from Los Alamos HIV* P value.
conclusion is based on unambiguous difference in the
envelope sequence in patients having higher CD4+ T cell
counts compared to those who had lower CD4+ T
lymphocyte counts. Our conclusion is also supported by
possible correlation between CD4+ T cell counts in
phylogenetic clustering, dS/dN ratio (an indicator of positive
selection), and changes N-linked glycosylation sites.
Phylogenetic analyses of these plasma sequences
revealed monophyletic clustering of pre- and post-HAART
in patients A, B, and C (higher CD4+ T cell counts).
However, patients D and E (lowest CD4+ T cell counts) did
not cluster in relation to time points. These results suggest
that although all patients experienced a noticeable viral
suppression due to HAART’s pressure over the pol gene,
only the ones with the higher CD4+ T lymphocytes were
able to mount an effective/selective immune pressure over
the envelope gene. This new selective pressure was able to
generate a related, although clearly distinct viral population
within few months. This finding is at least, in principle,
consistent with a recent report that showed women with
monophyletic HIV-1 genomic clusters per compartment
(plasma and vaginal) had consistently higher CD4+ T
counts, while those with lower CD4+ T cell counts display
intermingled quasispecies between compartments (Kemal et
al., 2003). In addition, our findings are in general agreement
with recent studies suggesting that specific immune
responses to HIV (and SIV in model system) are inversely
correlated with disease progression, and that the presence
and help of CD4+ T cells is essential to the disease control
(Garber et al., 2004; Kaech and Ahmed, 2003; Moir et al.,
2003; Rosenberg et al., 1997).
Our analyses also revealed clear shifts in glycosylation
sites in patients A, B, and C. The number of glycosylation
sites is conserved among different HIV strains. Sequence
changes altering potential glycosylation sites have been
shown to play an extremely important role in providing the
capacity to escape from immune recognition (Back et al.,
1994; Cheng-Mayer et al., 1999; Wei et al., 2003). Differ-
ences in glycosylation sites suggest the evolution of two
ent time points
F SD Wilcoxon’s
signed rank test
Mean dS/dN F SD Wilcoxon’s
signed rank test
0.020 0.611 1.930 F 1.315 0.360
0.020 1.635 F 0.555
0.004 0.007 1.866 F 1.059 0.011
0.003 0.400 F 0.207
0.005 b0.001 3.560 F 2.346 b0.001
0.004 0.600 F 0.212
0.010 0.205 2.280 F 0.856 0.728
0.009 2.458 F 1.498
0.016 0.211 1.838 F 1.464 0.014
0.015 1.536 F 0.870
0.016 b0.001 1.555 F 0.4848 0.064
nsynonymous per nonsynonymous site (dN) were estimated by using the
sequence database, http://hiv-web.lanl.gov. SD, standard deviation.

Table 4
Mean of synonymous and nonsynonymous nucleotide substitution changes between time points 1 and 2
Group Time
points
Mean
dS F SD
Wilcoxon’s
signed
rank test
Mean
dN F SD
Wilcoxon’s
signed
rank test
Mean
dS/dN F SD
Wilcoxon’s
signed
rank test
Higher CD4
(Patients B and C)
1 0.015 F 0.007 b0.001 0.007 F 0.005 b0.001 3.332 F 2.281 b0.001
2 0.027 F 0.014 0.042 F 0.011 0.536 F 0.247
Lower CD4
(Patients D and E)
1 0.056 F 0.026 0.070 0.037 F 0.016 0.461 1.880 F 1.422 0.018
2 0.059 F 0.022 0.033 F 0.012 2.106 F 1.369
Patient A was excluded from this analysis because of the unavailability of data for time point 2. SD, standard deviation.
E. Lorenzo et al. / Virology 330 (2004) 116–126 123
separate sets of sequences under distinct immune pressures
before and after HAART. Just as in the previous analysis,
patients D and E (lowest CD4+ counts) did not show clear
shifts in glycosylation sites suggesting lack of an effective
immune pressure on the env gene.
In addition, patients B and C in this study showed a dS/dNratio of b1, suggesting continued positive selection in these
patients, whereas dS/dN ratio (N1) in patients D and E
suggested lack of positive selection. This analysis measures
the variations of the synonymous per nonsynonymous ratio
(dS/dN), which provides a measure at the molecular level of
the selection intensity among amino acid sites (Anastasso-
poulou et al., 2003; Nielsen and Yang, 1998). It has been
suggested that changes in the strength of the immune
response may not result in predictable changes in the dS/dNratio if the selection coefficient is on the same order of
magnitude as the effective population size. Nevertheless,
observations of temporal changes in the dS/dN ratio alone
may still provide partial information about the status of the
immune system (Frost et al., 2001; Nielsen, 1999). The
reason why patient A did not show the expected positive
selection pressure is not fully understood. However, patient
A is the only patient in our study that achieved a viral load
b50 and developed fourfold decrease in genomic diversity
from pre- to post-HAART. At the same time patient A,
together with patient E, is one of the two patients that
exhibited emergence of rebounding virus, and the only in
which we were unable to study the lowest viral load after
therapy started. The time point analyzed in this case is one
in which the viral load is increasing during HAART, which
normally indicates the selection of a resistant virus. This
drug selection is indeed applied to the pol gene, and we
believe it may be eclipsing the immune pressure/selection
applied at the env gene.
Based on the diversity values of our patients’ viral
sequences before and after HAART, we were not able to
observe any clear viral genomic diversity restriction or
bottleneck evolution in HIV-1 env gene except in patient A.
This might be attributed to the fact that other patients never
achieved undetectable viremia (b50 copies/ml), which might
be important for narrowing the viral diversity in patients on
HAART. Another potentially important implication of the
present study is that establishment of HAART resistance may
be followed by faster virus rebound in patients with low
CD4+ T cell counts than in patients with high CD4+ T cell
counts. The future attempts should be directed towards
characterization of the virus emerging after a prolonged and
complete suppression, which may help in identification of
source of virus replication during HAART, and whether viral
rebound is faster in patients with low CD4+ T cell counts.
Materials and methods
Study subjects
Five HIV-1 infected informed-consenting individuals,
aged from 34 to 56 years were enrolled for the study. Blood
samples were collected from all five patients before
commencement of antiretroviral therapy. They achieved
and maintained almost complete viral suppression after the
initiation of the antiretroviral regimen. In this study, we
analyzed samples pre-HAART (Time 1), during plasma viral
suppression (Time 2) and occasionally during the virolog-
ical failure to the therapy (Time 3). Plasma HIV-1 viral load
measurement was carried out using the standard (detection
limit, 400 copies/ml) and ultrasensitive (detection limit, 50
copies/ml) Amplicor HIV-1 Monitor test (Roche Molecular
Systems, Inc., Branchburg, NJ). CD4+ T cells were
quantified by flow cytometry analyses using a FACSScan
(BD Biosciences, San Jose, CA). The clinical characteristics
of the patients are shown in Table 1.
Genetic characterization of HIV-1 env C2–V4 from plasma
HIV-1 RNA was isolated from plasma by using the
QIAmp Viral RNA Mini Kit (QIAGEN, Inc, Valencia, CA)
following the manufacturer’s indications. For the analysis of
plasma HIV-1 RNAwith viral loads b200 copies/ml, 3 ml of
plasma was applied to the column of the same kit. In order
to amplify the HIV-1 C2–V4 env region, nested PCR was
performed by using the OneStep RT-PCR Kit and HotStar
Taq DNA Polymerase (QIAGEN, Inc) with the external
primer pair ED12 and ED31 (5V-AGTGCTTCCTGCTGCT-CCCAAGAACCCAAG-3V and 5VCCTCAGTCATTAC
ACCAGGCCTGTCCAAAG-3Vrespectively) and the inter-
nal primer pair CV3-R and CV3-F (5V-TGATGGGAGGGG-TATACATT-3V and 5V-CTGTTAAATGGCAGTCTAGC-3Vrespectively). The final amplicon (525 bp) was electro-
phoresed with a low molecular weight marker in 1.2%
agarose, stained with ethidium bromide, and visualized
under UV light for determination of molecular weight.

Fig. 3. Graph of the fraction of N-linked glycosylation sites at each position in the alignment of amino acid sequences from patients A–E. Potential N-
glycosylation sites were determined by using the N-Glycosite tool from Los Alamos HIV sequence database, http://hiv-web.lanl.gov.
E. Lorenzo et al. / Virology 330 (2004) 116–126124
Cloning and sequencing of HIV-1 C2–V4 envelope gene
from plasma
PCR products were ligated and cloned according to the
TOPO TA Cloning Kit protocol (Invitrogen, Carlsbad, CA).
Plasmid DNA was purified by using QIAprep Spin
Miniprep columns (QIAGEN, Inc.). A total of 148 clones
were sequenced with M13 primers using Big-Dye termi-
nator chemistry according to manufacturer’s recommenda-
tions in an Applied Biosystems 3100 Genetic Analyzer
(Applied Biosystems, Foster City, CA).
Sequence analysis
All nucleotide sequences analyses were carried out on
a 480 bp fragment of the env gene spanning the C2–V4
region. Nucleotide sequences were aligned with BioEdit
version 5.0.9 (Hall, 1999) and later hand edited.

E. Lorenzo et al. / Virology 330 (2004) 116–126 125
Phylogenetic reconstructions were generated by the
neighbor-joining (NJ) method (with Kimura two-parame-
ter model and a transition/transversion ratio of 2.0) in
1000 bootstrapped data sets as implemented in the
MEGA version 2.1 (Kumar et al., 2001). The final
graphical output was created with TreeView. Pairwise
nucleotide sequence distance was computed by using
MEGA. The frequencies of synonymous substitutions per
synonymous site (dS) and nonsynonymous per nonsynon-
ymous site (dN) were estimated by using the SNAP tool
from the Los Alamos HIV sequence database, http://hiv-
web.lanl.gov. Potential N-linked glycosylation sites were
determined by using the N-glycosite tool from the Los
Alamos HIV Sequence database. Data were subjected to
nonparametric statistical treatment using Wilcoxon signed
rank test included in the SPSS Version 12.0 software
package (SPSS, Inc., Chicago).
Acknowledgments
We are grateful to the patients for their voluntary
participation in this study. We also thank the Epidemiology
and Biostatistics Core Program at Ponce School of Medicine.
This work was financially supported by National Institutes
of Health grants NIGMS-MBRS (S06-GM08239), NHLBI/
K0-1 (HLO-4371) and NCRR-RCMI (G12RR03050).
References
Anastassopoulou, C.G., Paraskevis, D., Sypsa, V.A., Chryssou, S.E.,
Antoniadou, A., Giamarelou, H., Hatzakis, A., 2003. Genetic evolution
of human immunodeficiency virus type 1 in two spouses responding
successfully to highly active antiretroviral therapy. AIDS Res. Hum.
Retroviruses 19, 65–71.
Back, N.K., Smit, L., De Jong, J.J., Keulen, W., Schutten, M., Goudsmit, J.,
Tersmette, M., 1994. An N-glycan within the human immunodeficiency
virus type 1 gp120 V3 loop affects virus neutralization. Virology 199,
431–438.
Bolmstedt, A., Sjolander, S., Hansen, J.E., Akerblom, L., Hemming, A.,
Hu, S.L., Morein, B., Olofsson, S., 1996. Influence of N-linked glycans
in V4–V5 region of human immunodeficiency virus type 1 glycoprotein
gp160 on induction of a virus-neutralizing humoral response. J.
Acquired Immune Defic. Syndr. Hum. Retrovirol. 12, 213–220.
Bonhoeffer, S., Holmes, E.C., Nowak, M.A., 1995. Causes of HIV
diversity. Nature 376, 125.
CDC, 1993. Revised classification system for HIV infection and expanded
surveillance case definition for AIDS among adolescents and adults.
Morb. Mort. Wkly. Rep. 41, 961–962.
Cheng-Mayer, C., Brown, A., Harouse, J., Luciw, P.A., Mayer, A.J., 1999.
Selection for neutralization resistance of the simian/human immunode-
ficiency virus SHIVSF33A variant in vivo by virtue of sequence
changes in the extracellular envelope glycoprotein that modify N-linked
glycosylation. J. Virol. 73, 5294–5300.
Choisy, M., Woelk, C.H., Guegan, J.F., Robertson, D.L., 2004. Compara-
tive study of adaptive molecular evolution in different human
immunodeficiency virus groups and subtypes. J. Virol. 78, 1962–1970.
Connor, R.I., Sheridan, K.E., Ceradini, D., Choe, S., Landau, N.R., 1997.
Change in coreceptor use correlates with disease progression in HIV-1-
infected individuals. J. Exp. Med. 185, 621–628.
DeBry, R.W., Abele, L.G., Weiss, S.H., Hill, M.D., Bouzas, M., Lorenzo,
E., Graebnitz, F., Resnick, L., 1993. Dental HIV transmission? Nature
361, 691.
Delwart, E.L., Pan, H., Sheppard, H.W., Wolpert, D., Neumann, A.U.,
Korber, B., Mullins, J.I., 1997. Slower evolution of human immunode-
ficiency virus type 1 quasispecies during progression to AIDS. J. Virol.
71, 7498–7508.
Demeter, L.M., D’Aquila, R., Weislow, O., Lorenzo, E., Erice, A.,
Fitzgibbon, J., Shafer, R., Richman, D., Howard, T.M., Zhao, Y.,
Fisher, E., Huang, D., Mayers, D., Sylvester, S., Arens, M., Sannerud,
K., Rasheed, S., Johnson, V., Kuritzkes, D., Reichelderfer, P., Japour,
A., 1998. Interlaboratory concordance of DNA sequence analysis to
detect reverse transcriptase mutations in HIV-1 proviral DNA. ACTG
Sequencing Working Group. AIDS Clinical Trials Group. J. Virol.
Methods 75, 93–104.
Frost, S., Gunthard, H.F., Wong, J.K., Havlir, D., Richman, D.D., Leigh
Brown, A.J., 2001. Evidence of positive selection driving the evolution
of HIV-1 env under potent antiviral therapy. Virology 284, 250–258.
Gallant, J.E., Gerondelis, P.Z., Wainberg, M.A., Shulman, N.S.,
Haubrich, R.H., St. Clair, M., Lanier, E.R., Hellmann, N.S.,
Richman, D.D., 2003. Nucleoside and nucleotide analogue reverse
transcriptase inhibitors: a clinical review of antiretroviral resistance.
Antivir. Ther. 8, 489–506.
Garber, D.A., Silvestri, G., Barry, A.P., Fedanov, A., Kozyr, N., McClure,
H., Montefiori, D.C., Larsen, C.P., Altman, J.D., Staprans, S.I.,
Feinberg, M.B., 2004. Blockade of T cell costimulation reveals
interrelated actions of CD4+ and CD8+ T cells in control of SIV
replication. J. Clin. Invest. 113, 836–845.
Hall, T.A., 1999. BioEdit: a user-friendly biological sequence alignment
editor and analysis program for Windows 95/98/NT. Nucleic Acids.
Symp. 41, 95–98.
Hillis, D.M., Huelsenbeck, J.P., Cunningham, C.W., 1994. Application and
accuracy of molecular phylogenies. Science 264, 671–677.
Ibanez, A., Clotet, B., Martinez, M.A., 2000. Human immunodeficiency
virus type 1 population bottleneck during indinavir therapy causes a
genetic drift in the env quasispecies. J. Gen. Virol. 81, 85–95.
Jensen, M.A., Li, F.S., van ’t Wout, A.B., Nickle, D.C., Shriner, D., He,
H.X., McLaughlin, S., Shankarappa, R., Margolick, J.B., Mullins, J.I.,
2003. Improved coreceptor usage prediction and genotypic monitoring
of R5-to-X4 transition by motif analysis of human immunodeficiency
virus type 1 env V3 loop sequences. J. Virol. 77, 13376–13388.
Kaech, S.M., Ahmed, R., 2003. Immunology. CD8 T cells remember with a
little help. Science 300, 263–265.
Kemal, K.S., Foley, B., Burger, H., Anastos, K., Minkoff, H., Kitchen, C.,
Philpott, S.M., Gao, W., Robison, E., Holman, S., Dehner, C., Beck, S.,
Meyer III, W.A., Landay, A., Kovacs, A., Bremer, J., Weiser, B., 2003.
HIV-1 in genital tract and plasma of women: compartmentalization of
viral sequences, coreceptor usage, and glycosylation. Proc. Natl. Acad.
Sci. 100, 12972–12977.
Kreisberg, J.F., Kwa, D., Schramm, B., Trautner, V., Connor, R., Schuite-
maker, H., Mullins, J., Ivan’t Wout, A.B., Goldsmith, MA, 2001.
Cytopathicity of human immunodeficiency virus type 1 primary isolates
depends on coreceptor usage and not patient disease status. J. Virol. 75,
8842–8847.
Kuiken, C.L., Zwart, G., Baan, E., Coutinho, R.A., van den Hoek, J.A.,
Goudsmit, J., 1993. Increasing antigenic and genetic diversity of the
V3 variable domain of the human immunodeficiency virus envelope
protein in the course of the AIDS epidemic. Proc. Natl. Acad. Sci. 90,
9061–9065.
Kumar, S., Tamura, K., Jakobsen, I.B., Nei, M., 2001. MEGA2:
molecular evolutionary genetics analysis software. Bioinformatics
17, 1244–1245.
Little, S.J., Holte, S., Routy, J.P., Daar, E.S., Markowitz, M., Collier, A.C.,
Koup, R.A., Mellors, J.W., Connick, E., Conway, B., Kilby, M., Wang,
L., Whitcomb, J.M., Hellmann, N.S., Richman, D.D., 2002. Antire-
troviral-drug resistance among patients recently infected with HIV. N.
Engl. J. Med. 347, 385–394.

E. Lorenzo et al. / Virology 330 (2004) 116–126126
Lorenzo, E., Herrera, R.J., Lai, S., Fischl, M.A., Hill, M.D., 1996. The Tat
and C2–V3 envelope genes in the molecular epidemiology of human
immunodeficiency virus-1. Virology 221, 310–317.
Lorenzo, E., Collins, T., Fisher, E., Herrera, R.J., 2001. The use of tat and
env sequences from human immunodeficiency virus 1 in phylogenetic
epidemiological studies. Electrophoresis 22, 438–444.
Lorenzo, E., Kumar, R., Hill, M.D., Costa, S., Chaudhary, S., Tirado, G.,
Yamamura, Y., Kumar, A., 2004. Genetic characterization of human
immunodeficiency virus type 1 Tat before and after highly active
antiretroviral therapy. AIDS Res. Hum. Retroviruses 20.
Malim, M.H., Emerman, M., 2001. HIV-1 sequence variation: drift, shift,
and attenuation. Cell 104, 469–472.
Moir, S., Ogwaro, K.M., Malaspina, A., Vasquez, J., Donoghue, E.T.,
Hallahan, C.W., Liu, S., Ehler, L.A., Planta, M.A., Kottilil, S., Chun,
T.W., Fauci, A.S., 2003. Perturbations in B cell responsiveness to CD4+
T cell help in HIV-infected individuals. Proc. Natl. Acad. Sci. 100,
6057–6062.
Nara, P.L., Garrity, R.R., Goudsmit, J., 1991. Neutralization of HIV-1: a
paradox of humoral proportions. FASEB J. 5, 2437–2455.
Nielsen, R., 1999. Changes in dS/dN in the HIV-1 env gene. Mol. Biol.
Evol. 16, 711–714.
Nielsen, R., Yang, Z., 1998. Likelihood models for detecting positively
selected amino acid sites and applications to the HIV-1 envelope gene.
Genetics 148, 929–936.
Nowak, M.A., Anderson, R.M., McLean, A.R., Wolfs, T.F., Goudsmit, J.,
May, R.M., 1991. Antigenic diversity thresholds and the development
of AIDS. Science 254, 963–969.
Ou, C.Y., Ciesielski, C.A., Myers, G., Bandea, C.I., Luo, C.C., Korber,
B.T., Mullins, J.I., Schochetman, G., Berkelman, R.L., Economou,
A.N., et al., 1992. Molecular epidemiology of HIV transmission in a
dental practice. Science 256, 1165–1171.
Philpott, S., Weiser, B., Anastos, K., Kitchen, C.M., Robison, E., Meyer III,
W.A., Sacks, H.S., Mathur-Wagh, U., Brunner, C., Burger, H., 2001.
Preferential suppression of CXCR4-specific strains of HIV-1 by
antiviral therapy. Clin. Invest. 107, 431–438.
Quinones-Kochs, M.I., Buonocore, L., Rose, J.K., 2002. Role of N-
linked glycans in a human immunodeficiency virus envelope
glycoprotein: effects on protein function and the neutralizing antibody
response. J. Virol. 76, 4199–4211.
Rangel, H.R., Weber, J., Chakraborty, B., Gutierrez, A., Marotta, M.L.,
Mirza, M., Kiser, P., Martinez, M.A., Este, J.A., Quinones-Mateu, M.E.,
2003. Role of the human immunodeficiency virus type 1 envelope gene
in viral fitness. J. Virol. 77, 9069–9073.
Rosenberg, E.S., Billingsley, J.M., Caliendo, A.M., Boswell, S.L., Sax,
P.E., Kalams, S.A., Walker, B.D., 1997. Vigorous HIV-1-specific
CD4+ T cell responses associated with control of viremia. Science 278,
1447–1450.
Schuitemaker, H., Koot, M., Kootstra, N.A., Dercksen, M.W., de Goede,
R.E., van Steenwijk, R.P., Lange, J.M., Schattenkerk, J.K., Miedema,
F., Tersmette, M., 1992. Biological phenotype of human immunodefi-
ciency virus type 1 clone at different stages of infection: progression of
disease is associated with a shift from monocytotropic to T-cell-tropic
virus population. J. Virol. 66, 1354–1360.
Shankarappa, R., Margolick, J.B., Gange, S.J., Rodrigo, A.G., Upchurch,
D., Farzadegan, H., Gupta, P., Rinaldo, C.R., Learn, G.H., He, X.,
Huang, X.L., Mullins, J.I., 1999. Consistent viral evolutionary changes
associated with the progression of human immunodeficiency virus type
1 infection. J. Virol. 73, 10489–10502.
Skrabal, K., Trouplin, V., Labrosse, B., Obry, V., Damond, F., Hance,
A.J., Clavel, F., Mammano, F., 2003. Impact of antiretroviral
treatment on the tropism of HIV-1 plasma virus populations. AIDS
17, 809–814.
Stevenson, M., 2003. HIV-1 pathogenesis. Nat. Med. 9, 853–860.
Stipp, H.L., Kumar, A., Narayan, O., 2000. Characterization of immune
escape viruses from a macaque immunized with live-virus vaccine and
challenged with pathogenic SHIVKU-1. AIDS Res. Hum. Retroviruses
16, 1573–1580.
Tirado, G., Jove, G., Kumar, R., Noel, R.J., Reyes, E., Sepulveda, G.,
Yamamura, Y., Kumar, A., 2004. Differential virus evolution in blood
and genital tract of HIV-Infected females: evidence for the involvement
of drug ad non-drug resistance-associated mutations. Virology 324,
577–586.
Unal, A., Lorenzo, E., Brown, M., Smith, L., Matsuura, S., Scott, G., Scott,
W., 1996. Reverse transcriptase mutations in HIV-1-infected children
treated with zidovudine. J. Acquired Immune Defic. Syndr. Hum.
Retrovirol. 13, 140–145.
Wei, X., Decker, J.M., Wang, S., Hui, H., Kappes, J.C., Wu, X., Salazar-
Gonzalez, J.F., Salazar, M.G., Kilby, J.M., Saag, M.S., Komarova, N.L.,
Nowak, M.A., Hahn, B.H., Kwong, P.D., Shaw, G.M., 2003. Antibody
neutralization and escape by HIV-1. Nature 422, 307–312.
Copyright © 2022 FDOKUMEN




















