
Enviralab Sterility Module Unidirectional Flow Clean Bench ...
Upload
independentCategory
view
6download
0
Aquaculture, 119 (1994) 409-423 Elsevier Science B.V., Amsterdam
409
AQUA 60069
Induction of sterility in coho salmon (Oncorhynchus kisutch) by androgen immersion
before first feeding
Francesc Piferrefi’ , Manuel Carrillob, Silvia Zanuyb, Igor I. Solar” and Edward M. Donaldson”
a West Vancouver Laboratory, West Vancouver, BC, Canada bInstituto de Acuicultura de Torre de la Sal, Ribera de Cabanes, Castell6n, Spain
(Accepted 1 October 1993)
ABSTRACT
Based on the observation that in coho salmon (Oncorhynchus kisutch) the sensitivity to the effects of exogenous steroids is highest around hatching, and that it decreases as ontogenetic development progresses, immersion in a dilute solution of androgen prior to first feeding was examined as a means of inducting sterility in this species. Two approaches were tested: 17a-methyltestosterone (MT) was administered either by continuous immersion for 30 days, from just before hatching to just before first feeding, at doses ranging from 3.12 to 100 ,ug/l, or as discrete 2-h immersions at doses of 1 or 10 mg/l, once or twice per week, for a total of 5 or 10 immersions, respectively. Sampling of fish at about 7 months of age showed that these treatments had no significant effect on survival or length, whereas weight was slightly reduced only in the groups exposed to the highest amount of androgen. At the doses tested, discrete immersions resulted in a higher yield of sterile fish (maximum 92.3%) than continuous immersions (maximum 5 1.2%). In a subsequent experiment, 17ol-methyldihydrotestos- terone was compared with MT in discrete immersions, but the percentage of sterile fish produced was not different from that obtained with MT. These results show that androgen immersion of coho salmon before fust feeding can result in a high proportion of sterile fish without compromising their viability. The benefits for aquaculture derived from sterilizing salmon before first feeding are discussed.
INTRODUCTION
Options for controlled sex differentiation in salmonid aquaculture include the production of monosex female stocks and the production of sterile stocks (Donaldson and Benfey, 1987). Recently, there has been an increased em- phasis on the development of sterilization technologies to improve the eff- ciency of commercial aquaculture operations, as well as a means of prevent- ing reproductive interaction between genetically-modified fish and wild fish (Devlin and Donaldson, 1992; Donaldson et al., 1993 ).
‘Present address and correspondence to: Dr. F. Piferrer, University of California at San Diego, 9500 Gilman Dr., La Jolla, CA 92093-0947, USA.
0044-8486/94/$07.00 0 1994 Elsevier Science B.V. All rights reserved. SSDI0044-8486(93)E0239-6

410 F. PIFERRER ET AL.
Hormonal sterilization is one of several potential methods for the large- scale sterilization of salmonids, and it can be achieved by administration of an androgen, usually 17a-methyltestosterone (MT), at higher doses and for longer periods of time than those required for masculinization (Donaldson and Benfey, 1987). Treatment regimes for hormonal sterilization of salmo- nids and other economically-important species, have included, in the major- ity of cases, dietary androgen supplementation for periods extending over several weeks. In coho salmon (Oncorhynchus kisutch), for instance, the pre- viously tested androgen treatment regimes extended for 2-3 months from first feeding or until the fish reached 3 g in body weight (Goetz et al., 1979; Don- aldson and Hunter, 1982; Shelbourn et al., 1992a,b).
In the case of direct masculinization and feminization, Piferrer and Don- aldson ( 1989, 1992) have demonstrated that the sensitivity to exogenous steroids changes considerably during ontogenesis. In coho salmon, this sen- sitivity was highest around the time of hatching and decreased as ontogenetic development progressed. This implies the existence of a labile period in which the gonads are highly sensitive to the action of exogenously administered sex steroids. Treatment after the labile period requires higher dosages to counter- act the normal course of sex differentiation and may not be effective at all (Piferrer and Donaldson, 1993 ) . Therefore, based on these observations, and with the objective of reducing treatment regimes, we tested the feasibility of sterilizing coho salmon by hormonal immersions during the labile period in order to eliminate the need for subsequent dietary androgen treatment. Ad- ditionally, since androgen treatment in pre-smolts can interfere with seawater adaptation, as shown in various salmonid species (Miwa and Inui, 1986; Ikuta et al., 1987; Lundqvist et al., 1989; Shelbourn et al., 1992a,b), treatment ex- clusively before yolk absorption and first feeding would not be expected to influence seawater adaptation, particularly in smolts transferred at zero age.
To test whether sterilization by immersion treatment alone was feasible, two different approaches were examined. Fish were treated with MT from the late eyed-egg stage through the end of the alevin period, i.e. until first feeding, either by continuous immersion at relatively low doses or by discrete admin- istration of MT at substantially higher doses than those used for direct mas- culinization. In a subsequent study, we determined whether the results ob- tained with discrete immersions could be improved by the administration of the non-aromatizable anclrogen 17cu-methyldihydrotestosterone (MDHT) , since we had obtained evidence of its higher masculinizing potency with re- spect to MT (Piferrer et al., 1993). In this paper, the effects of these treat- ments on survival, growth and gonadal development are reported, illustrating their increasing effects in both testes and ovaries until fully sterilized fish were obtained.

HORMONAL STERILIZATION OF COHO SALMON 411
MATERIALS AND METHODS
Animals Mixed sex coho salmon fertilized eggs were placed in vertically-stacked in-
cubators (Heath Tecna Corp., Kent, WA), supplied with water at ambient river temperature, until they reached the eyed-egg stage (330”C-days). The eyed-eggs were divided into groups of 150 each and placed in plexiglass boxes fitted with a 1 -mm2 plastic mesh top and bottom as described by Goetz et al. ( 1979). The water temperature was then raised gradually to 10 + 1 “C and maintained at this range throughout the period of hormonal treatments. In this species, hatching takes place around 450’ C-days and, at the water tem- perature described above, it lasts over a period of about 5 days. The day at which 50% of the eggs had hatched is subsequently referred as “hatching”. Timing of hormonal treatments is reported as days before or after hatching.
The alevins remained in the boxes until all hormonal treatments were com- pleted 4 weeks after hatching, and just before first feeding. At this time, they were transported to the Rosewall Creek Experimental Fish Hatchery (Fanny Bay, BC). There, each group of fish was reared in 50-litre fiberglass tanks supplied with well water at approximately 8 a C, under a natural photoperiod and hand-fed Oregon Moist Pellets of the appropriate size 4 times a day.
Androgen treatments The synthetic androgens used were 17a-methyltestosterone (MT) and 1 ICY-
methyldihydrotestosterone (MDHT) (Sigma Chemical Co., St. Louis, MO). Stock solutions of the steroids were prepared by dissolving the appropriate amount of each hormone in 95% ethanol. Aliquots of each stock solution were added to a plastic bucket containing 5 litres of water. Final steroid concentra- tions for each treatment are specified below. Hormonal treatments were car- ried out by immersing the plexiglass chambers containing the eyed-eggs or alevins inside the buckets. Compressed air was gently forced through an air- stone to provide both adequate oxygenation and mixing. The temperature during the immersion treatments was kept constant at 1.0 2 lo C by circulating well water around the plastic buckets.
Experiment 1. This experiment was designed to test whether sterilization could be achieved by continuous immersion in water containing androgen at low doses. Four groups of late eyed-eggs were immersed in MT solutions at the following final concentrations (in hg/l): 3.12 (Group A), 6.25 (Group B), 25 (Group C) and 100 (Group D). Every 5 days, the water in the buckets containing the steroid solutions was replaced and new steroid solutions were made. In this experiment, immersion treatment was continuous for 30 days, covering the period from 4 days before hatching (411 “C-days) to 25 days after hatching (705 o C-days), just before swim-up and first feeding.

412 F. PIFERRER ET AL.
Experiment 2. This experiment was designed to test whether sterilization could be achieved by multiple discrete immersions in water containing androgen at high doses. Four groups, initially at the eyed-egg stage, were subjected to 2-h immersion treatments with MT at 1 or 10 mg/l final concentration, once or twice per week (total of 5 or 10 immersions, respectively): Group E was treated with 1 mg/l once per week; Group F, with 1 mg/l twice per week; Group G, with 10 mg/l once per week; and Group H, with 10 mg/l twice per week. Groups treated once per week received immersions in MT solution 7 days before hatching and 1,6, 13 and 20 days after hatching. Groups treated twice per week, besides the treatments specified above, also received MT 2 days before hatching and 3, 11, 17 and 24 days after hatching.
Experiment 3. This experiment was similar in design to Experiment 2, i.e. using discrete androgen administration. In this case, the synthetic androgens MT and MDHT were compared. Group I was given a 2-hour immersion treatment in MT at 1 mg/l twice per week (like group F); Group J, MT at 10 mg/l twice per week (like Group H) ; Group K, MDHT at 1 mg/l once per week, Group L, MDHT at 1 mg/l twice per week; Group M, MDHT at 10 mg/l once per week, and Group N, MDHT at 10 mg/l twice per week. Groups treated once per week received the androgen 7 days before hatching and 1,7, 14 and 2 1 days after hatching. Groups treated twice per week received addi- tional treatments 2 days before hatching and 3, 10, 17 and 24 days after hatching.
Experiments 1 and 2 were carried out simultaneously, while experiment 3 was conducted in a subsequent year. Untreated groups containing 150 fish served as controls in each case.
Sampling Sampling took place at 18 weeks (Experiments 1 and 2 ) or 23 weeks (Ex-
periment 3) after first feeding. This was designed to allow sufficient time to elapse after the end of the treatments and therefore to clearly establish the gonadal phenotype, since previous hormonal manipulations in this species have shown that the gonadal proportions do not usually change in subsequent samplings, as evidenced by the sterile condition of the gonads and absence of gonadal regeneration in treated fish (Hunter et al., 1982; Shelboum et al., 1992b). Survival at sampling was calculated as the percentage of fish remain- ing with respect to the 150 eyed-eggs allocated to each treatment. Between 30 and 45 fish were randomly taken from each tank and killed with an overdose of the anaesthetic 2-phenoxyethanol (Syndel Laboratories, Vancouver, BC), individually weighed (precision + 0.0 1 g ) and measured (fork length to + 0.1 cm). Differences in average weight and length of fish from each group were analyzed by one-way analysis of variance. Unplanned comparisons among means were carried out by Tukey’s multiple range test (Sokal and Rohlf,

HORMONAL STERILIZATION OF COHO SALMON 413
198 1) . Linear regression was used to determine the possible relationship be- tween the dose administered and the final mean weight and length among the treated groups in Experiments 1 and 2.
The effects of androgen administration on the resulting phenotype were evaluated by making histological preparations of the gonads, as described by Piferrer and Donaldson ( 1989 ) . Haematoxylin-eosin-stained 5-p sections of the gonads of each fish were examined in duplicate at two different loca- tions separated about 1 cm apart, to cover most of the gonadal span at the age sampled. This was designed to account for possible variations in gonadal morphology along the cranial-caudal axis. Histological examination of the microscopic slides was carried out by two independent observers. Since the treatments applied resulted in the appearance of several gonadal morpholo- gies, the gonads of treated fish were classified as follows (see detailed descrip- tion of characteristics concerning each gonadal type in the “Results” sec- tion): Type I, normal testis; Type II, affected testis; Type III, normal ovary; Type IV, affected ovary; Type V, intersex; and Type VI, sterile. A conserva- tive approach was always taken in the classification of fish showing gonads belonging to two different types. Thus a fish in which one gonad was an af- fected testis or affected ovary and the other was a sterile gonad, was classified as Type II or IV, respectively, but not as Type VI, even when the effects of androgen treatment were severe in the non-sterilized gonad.
Differences in the sex ratio between control and treated groups were ana- lyzed with the chi-square test (Sokal and Rohlf, 198 1) . In addition, in every experiment, differences in treatment efficiency were analyzed by a modifica- tion of the method of Goodman ( 1964) for contrasts among multinomial populations. The modification served the purpose of comparing separately whether each gonadal type mentioned above appeared with the same fre- quency as a consequence of the different hormonal treatments. Thus, in each experiment, the hormonal treatments were assigned to the I class (i= 1,2, . . . . 4 for Experiments 1 and 2; i= 1, 2, . . . . 6 for Experiment 3 ), and the six ob- served gonadal types mentioned above the J class (j= 1,2, . . . . 6). Then, the null hypothesis (HO), which assumes that the I multinomial populations (hormonal treatments) have the same effectiveness, states that:
II~j=II~j=... =II,,=rLj (forj= 1,2, . . . . J),
where II, is the population relative frequency in the ijth combination, and II, is the mean relative frequency in each j class (gonadal type). The statistic employed was:
X2=-&= 1,Z,...,I(Q -niPj 12/niPj,
where ylij was the number of fish with gonads in each ifih combination, nj the number of fish in each treatment (i.e., the sample size) and pi was calculated as the weighted arithmetic mean of frequencies:

414 F. PIFERRER ET AL.
Pj =-L w,...,A Pijln,
where n was the total number of fish examined in each experiment and pij was the observed relative frequency in the ijth combination, i.e., for simplicity:
Pj =&i= ,,2 ,..., &n.
Therefore, the statistic x2 had a chi-square distribution with I- 2 degreesof freedom (number of hormonal treatments in each experiment minus 1, and minus 1 for the estimated pi).
RESULTS
In Experiments 1 and 2, survival from the late eyed-egg stage to sampling was 80.3% in the control group, while the treated groups had a survival rate l-6% lower than that of the control group, but not related to the amount of androgen administered. Survival in Experiment 3 ranged from 70% to 74%, including both the control group and all but one of the treated groups, and was similarly not related to the amount of androgen administered, with the exception of Group N (MDHT at 10 mg/l twice per week) which had a sur- vival of 64%,
The mean weight and length of the control and treated groups of each ex- periment are shown in Table 1. In Experiment 1, continuous treatment with MT did not affect weight’ or length in a statistically significant manner. Nevertheless, the weight and length of fish treated with MT at 3.12 lug/l (Group A) were slightly higher than those of the controls. Further, the weight and length of fish treated with MT from 6.25 to 100 pg/l (Groups B, C and D), decreased as the dose increased (r= -0.845 for weight; Y= -0.857 for length), although such regressions were not statistically significant (P= 0.154 and P= 0.142, respectively). In Experiment 2, fish treated with discrete im- mersions once per week with MT at 1 mg/l (Group E) showed a slight in- crease in weight and length with respect to the controls. Conversely, groups treated twice per week (Groups F and H) showed a significantly lower mean weight than the controls (P< 0.05). In Experiment 3, mean weight and length of treated groups decreased as the dose of either MT or MDHT increased, but differences with the control group were statistically significant only for the groups treated with the highest amount of each androgen (MT: Group J, P-c 0.05; MDHT: group N, PC 0.0 1).
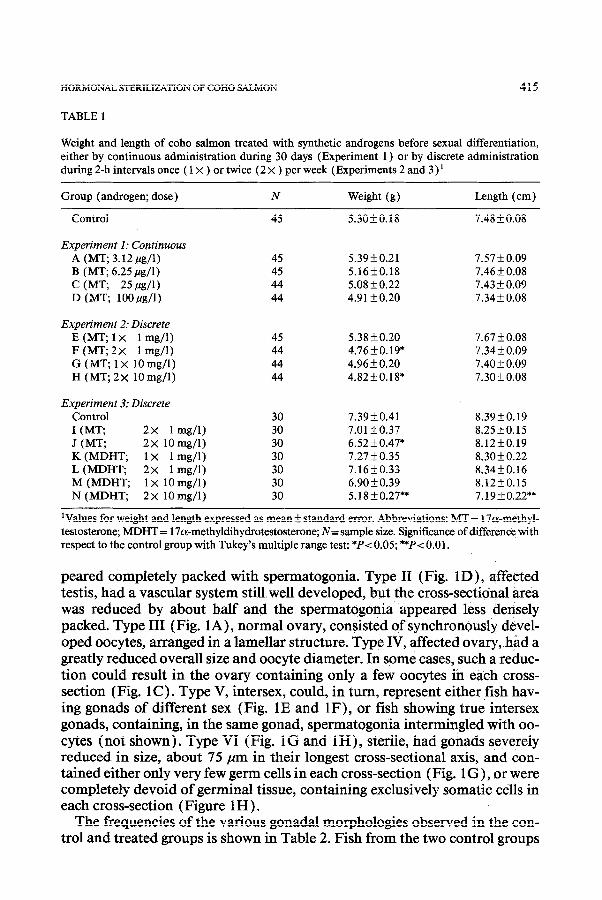
Histological preparations of gonadal cross-sections from fish from the con- trol and treated groups were classified into six types. Fig. 1 illustrates the in- creasing effects of androgen treatment on both testes and ovaries until sterile fish were produced. Type I (Fig. 1B ), normal testis, had a typically developed vascular system, consisting of the gonadal vein and artery, and the sperm duct. These testes were about 280 pm in their longest cross-sectional axis and ap-
HORMONAL STERILIZATION OF COHO SALMON 415
TABLE 1
Weight and length of coho salmon treated with synthetic androgens before sexual differentiation, either by continuous administration during 30 days (Experiment 1) or by discrete administration during 2-h intervals once ( 1 x ) or twice (2 x ) per week (Experiments 2 and 3 ) 1
Group (androgen; dose) N Weight (9) Length (cm)
Control 45 5.30f0.18 7.48 f 0.08
Experiment I: Continuous A (MT; 3.12 pg/l) B (MT; 6.25 fig/l) C (MT; 25 /*g/l) D (MT; 100&l)
45 45 44 44
Experiment 2: Discrete E (MT; 1 x 1 mg/l) 45 F (MT;Zx 1 mg/l) 44 G (MT; 1 x 10 mg/l) 44 H (MT; 2x 10 mg/l) 44
Experiment 3: Discrete Control I (MT; 2x 1 m&l) J (MT; 2x lOmg/l) K (MDHT; lx 1 mg/l) L (MDHT; 2x 1 mg/l) M (MDHT, lx lOmg/l) N (MDHT; 2x lOmg/l)
30 30 30 30 30 30 30
5.39kO.21 7.57kO.09 5.16&0.18 7.46 f 0.08 5.08 +0.22 7.43 2 0.09 4.9lkO.20 7.34& 0.08
5.38 kO.20 7.67kO.08 4.76f0.19* 7.34f 0.09 4.96 k 0.20 7.40 + 0.09 4.82f0.18* 7.30 f 0.08
7.39kO.41 8.39kO.19 7.01 f0.37 8.25kO.15 6.52&0.47* 8.12f0.19 7.27kO.35 8.30f 0.22 7.16kO.33 8.34f0.16 6.90 f 0.39 8.12+0.15 5.18f0.27” 7.19+0.22**
‘Values for weight and length expressed as mean f. standard error. Abbreviations: MT= 17a-methyl- testosterone; MDHT= 17a-methyldihydrotestosterone; N=sample size. Significance of difference with respect to the control group with Tukey’s multiple range test: *PC 0.05; ““P< 0.0 1.
peared completely packed with spermatogonia. Type II (Fig. 1 D), affected testis, had a vascular system still well developed, but the cross-sectional area was reduced by about h.alf and the spermatogonia appeared less densely packed. Type III (Fig. 1 A), normal ovary, consisted of synchronously devel- oped oocytes, arranged in a lamellar structure. Type IV, affected ovary, had a greatly reduced overall size and oocyte diameter. In some cases, such a reduc- tion could result in the ovary containing only a few oocytes in each cross- section (Fig. 1 C ) . Type V, intersex, could, in turn, represent either fish hav- ing gonads of different sex (Fig. 1E and IF), or fish showing true intersex gonads, containing, in the same gonad, spermatogonia intermingled with oo- cytes (not shown). Type VI (Fig. 1G and lH), sterile, had gonads severely reduced in size, about 75 pm in their longest cross-sectional axis, and con- tained either only very few germ cells in each cross-section (Fig. 1 G), or were completely devoid of germinal tissue, containing exclusively somatic cells in each cross-section (Figure 1 H ) .
The frequencies of the various gonadal morphologies observed in the con- trol and treated groups is shown in Table 2. Fish from the two control groups

F. PIFEXRER ET AL.
Fig. 1. Cross-sections of gonads of coho salmon treated with synthetic androgens during early devel- opment, which illustrate the sequence of increasing effects of androgens on ovaries (photomicro- graphs A and C) and on testes (photomicrographs B and D) until sterile gonads were produced (pho- tomicrographs G and H). (A) Normal ovary (Type III; from control group). Observe synchronous oocyte development and the arrangement of ovarian iamellae. (B) Normal testis (Type I; from con- trol group). Note the arrangement of the vascular system. The gonad is fully packed with spermato- gonia. (C) Affected ovary (Type IV), as a consequence of treatment with androgen, resulting in a marked reduction of its size, with fewer oocytes per cross-section.

HORMONAL STERILIZATION OF COHO SALMON 417
TABLE 2
Gonadal morphology of who salmon treated with synthetic androgens before sexual differentiation, either by continuous administration during 30 days (Experiment 1) or by discrete administration during 2-h intervals once ( 1 x ) or twice (2 x ) per week (Experiments 2 and 3)’
Group (androgen; dose) N Gonadal type (% )
I II III IV V VI
Control Experiment 1: Continuous
A (MT; 3.12&l) B (MT, 6.25 &l) C (MT; 25 pgugll) D (m lOOfig/l)
45 51.1 0.0 48.9 0.0 0.0 0.0
45 44 44 43
Experiment 2: Discrete E (MT, 1 x 1 mg/l) F (MT; 2x 1 mg/l) G (MT; 1 x 10 mg/l) H (MT, 2x 10 mg/l)
41 31 41 42
Experiment 3: Discrete Control 25 52.0 0.0 48.0 0.0 0.0 0.0
I (MT; 2x 1 mg/l) 28 .I (MT, 2x lOmg/l) 26 K (MDHT; 1 X 1 mg/l) 30 L(MDHT; 2x 1 mg/l) 24 M (MDHT; 1 x 10 mg/l) 29 N (MDHT; 2X lOmg/l) 30
6.7 2.2 0.0 68.9 6.7 15.5 9.1 6.8 0.0 47.7 2.3 34.1 9.1 2.2 11.4 38.6 4.6 34.1 9.3 7.0 16.3 11.6 4.6 51.2
ns ns b C ns a
12.2 19.5 0.0 14.6 0.0 53.1 2.8 13.5 0.0 16.2 0.0 67.5 4.9 9.1 0.0 19.5 0.0 65.9 2.4 4.7 0.0 12.0 0.0 80.9
ns ns ns ns ns ns
0.0 14.3 0.0 10.7 0.0 75.0 0.0 3.8 0.0 3.9 0.0 92.3 0.0 16.7 0.0 10.0 0.0 73.3 0.0 4.2 0.0 16.7 0.0 79.1 0.0 6.9 0.0 13.8 0.0 79.3 0.0 10.0 0.0 6.7 0.0 83.3
ns ns ns ns ns ns
‘Abbreviations: MT = 17ct+methyltestosterone; MDHT = 17a-methyldihydrotestosterone; N= sample size. Gonadal types: I, testis; II, affected testis; III, ovary; IV, affected ovary; V, intersex; VI, sterile. The proportion of gonadal Types I and III of all the treated groups was significantly different (PC 0.00 1) with respect to those of their corresponding controls. Within each experiment, significance of differ- ence among the various treatments is indicated for each gonadal type: a=P<0.025; b=P<0.005; c=P<O.OOl; ns=not significant.
Fig. 1. (D) . Affected testis (Type II), of reduced size and with fewer spermatogonia, but with well- developed vascular system. (E and F) Intersex (Type V) . Both the ovary and testis are of reduced size and belong to the same individual. Observe the appearance in the ovary of the vascular system characteristic of testes. (G and H) Sterile gonads (Type VI). The vascular system is greatly reduced, as is the organ size. Germ cells (indicated by arrowheads) are either very scarce and appear degener- ate (G), or completely absent (H). A and C, bar=200 pm; B, D, G and H, bar=50 pm; E and F, bar = 100 pm. Abbreviations: a = gonadal artery, mo = mesovarium, ms = mesorchium, sb = swim bladder, sd = sperm duct, v = gonadal vein, vs = vascular system.
418 F. PIFERRER ET AL.
exhibited gonads exclusively of either Type I or III (normal testis and normal ovary) at a proportion’of 51.1% and 48.9%, respectively, in Experiments 1 and 2, and at a proportion of 52% and 48% in Experiment 3. In sharp con- trast, fish from treated groups had Type I gonads (normal testes) averaging 8.5% in Experiment 1, 5.5% in Experiment 2, and were absent in all groups of Experiment 3. Type II gonads (affected testes) ranged from 2.2% to 7% in Experiment 1 or decreased as the amount administered of MT but not MDHT increased in both Experiments 2 and 3. Interestingly, Type III gonads (nor- mal ovaries) were absent in Groups A and B but then reappeared as the dose of androgen increased in Groups C and D. Further, normal ovaries were ab- sent in fish belonging to all the groups treated with discrete immersions. In Group A, 68.9% of gonads sampled were Type IV (affected ovaries), but the figure decreased as the dose of androgens increased in the remaining groups from the same experiment. In Experiment 2, affected ovaries were present in 12-19.5% of the fish, in a consistent manner, whereas in Experiment 3 they accounted for 3.9-l 6.7% of the lish examined. A low proportion of Type V gonads (intersex) was present in fish treated by continuous immersion, while this condition was not observed in those receiving discrete immersions. In Experiment 1, the proportion of fish exhibiting Type VI gonads (sterile) in- creased with the dose of androgen from 15.5% to 5 1.2%, of which those with gonads with total absence of germ cells (Fig. 1 H) also increased with the dose from 11.1% (Group A) to 4 1.9% (Group D). Fish from groups treated with
100 ,
DISCRETE MDHT IMMERSIONS
.E .D
CONTINUOUS MT IMMERSION
2 20 200
CUMULATIVE DOSE (mg/l x h)
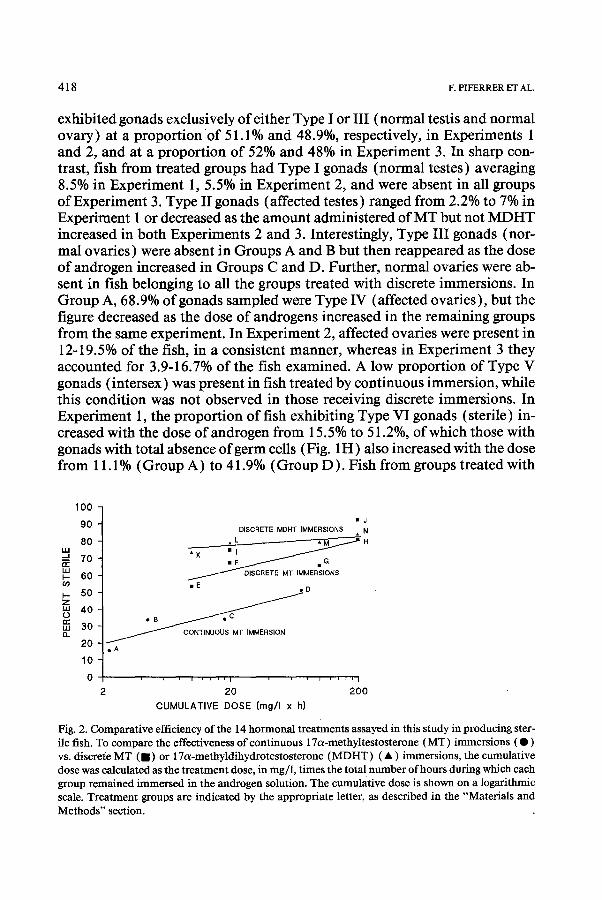
Fig. 2. Comparative efficiency of the 14 hormonal treatments assayed in this study in producing ster- ile fish. To compare the effectiveness of continuous 17a-methyltestosterone (MT) immersions ( 0 ) vs. discrete MT ( n ) or 17a-methyldihydrotestosterone (MDHT) ( A ) immersions, the cumulative dose was calculated as the treatment dose, in mg/l, times the total number of hours during which each group remained immersed in the androgen solution. The cumulative dose is shown on a logarithmic scale. Treatment groups are indicated by the appropriate letter, as described in the “Materials and Methods” section.

HORMONAL STERILIZATION OF COHO SALMON 419
discrete immersions had gonads of Type VI from 53.7% to 80.9% and from 73.3% to 92.3% in Experiments 2 and 3, respectively. Similarly, fish having sterile gonads like those depicted in Fig. 1H ranged from 2 1.6% to 6 1.3% and from 22.6% to 52.5%, respectively. The highest proportion (92.3%) of sterile fish was observed in Group J (MT at 10 mg/l twice per week). The number of sterile fish produced as a consequence of the 14 hormonal treatments as- sayed is also presented in Fig. 2. This figure shows that, when the cummula- tive dose is taken into consideration to allow the comparison of continuous versus discrete immersions, over the dosage ranges tested discrete immer- sions consistently produced about 25% more sterile fish than continuous immersions.
Chi-square tests showed that in all the 14 androgen-treated groups the pro- portion of fish having normal testis or normal ovaries was statistically differ- ent (P< 0.00 1) with respect to their corresponding controls. Chi-square tests of homogeneity to compare whether there were statistically significant differ- ences in the frequencies of each gonadal type as a consequence of the several hormonal treatments assayed, showed no differences in the proportion of go- nads of Types I, II and V in Experiment 1. However, the appearance of go- nads of Type III (normal ovaries), IV (affected ovaries) and VI (sterile) were statistically significant (P< 0.005, P-c 0.00 1 and P< 0.025, respec- tively) among the four experimental groups. In contrast, no significant differ- ences were detected in the appearance of any gonadal type in fish from the groups of Experiments 2 and 3.
DISCUSSION
Previous attempts to sterilize coho salmon with androgens required im- mersion treatments of eyed-eggs and alevins, plus dietary treatment of fry with MT at 1 O-20 mg/kg of food for lo- 12 weeks in order to obtain more than 90% sterile fish (Goetz et al., 1979; Donaldson and Hunter, 1982; Hunter et al., 1982; Shelbourn et al., 1992a,b). Deletion of hormonal immersion treat- ment of either eyed-eggs or alevins reduced the proportion of sterile fish to the range of 78-83% (Donaldson and Hunter, 1982). Dietary treatment alone resulted in 47-52% sterile fish (McBride and Fagerlund, 1973; Goetz et al., 1979), while treatment of eyed-eggs and/or alevins produced only O-26% sterile fish (Donaldson and Hunter, 1982 ) . Thus, earlier studies showed that both immersion treatments during early development and, more importantly, dietary treatment of fry appeared to be necessary to achieve a high percentage of sterile fish. In the present study, the novel use of increased immersion fre- quencies and doses resulted in the production of up to 92.3% sterile fish with- out the need for dietary treatment of fry, hence showing that it is possible to improve treatment regimes of eyed-eggs and alevins. Nevertheless, not all the treatments assayed in our study proved to be equally satisfactory.
Although it has been shown that anabolic steroids at low doses can promote

420 F. PIFERRER ET AL.
growth in fish (Higgs et al., 1982), it is not possible to conclude from this study whether the observed increase in growth of the groups treated with low doses of MT was due to such an effect. On the other hand, the repeatedly- encountered statistically significant decrease in growth in fish treated with MT at 10 mg/l twice per week (Groups H and J) can be viewed as deleterious pharmacological effects of androgen treatment. Survival, however, was not compromised in all but one of the experiment groups.
The reappearance of fish with normal ovaries in Groups C and D after their absence in Groups A and B is most likely due to the effects of the aromatiza- ble MT, a paradoxical phenomenon previously observed in several species, including coho salmon (Piferrer and Donaldson, 1991). Discrete immer- sions produced a higher proportion of sterile fish than continuous treatment. However, the effectiveness of discrete immersions was markedly less depen- dent on the cumulative dose of androgen employed, as shown by the &i-square tests. At a cumulative dose between 10 and 80 (mg/l x h), Fig. 2 shows that discrete immersions produced, in a consistent manner, about 25% more ster- ile fish than continuous immersion. Yet higher doses by discrete immersion did not significantly increase the proportion of sterile fish. Similarly, MDHT did not result in a significantly higher proportion of sterile fish, although it has been shown that at lower doses MDHT is twice as potent as MT in mas- culinizing Pacific salmon (Piferrer et al., 1993). The present results, there- fore, may reflect the way in which androgens affect gonadal development causing sterilization.
In the context of fish farming, sterile fish could be defined as those with gonads of reduced size where gametogenesis and steroidogenesis has been blocked, at least beyond the time of marketing. Thus, the prevalence of the sterile condition, i.e. the absence of gonadal regeneration, is of relevance. In this experiment, it is assumed that the level of sterility recorded at 7 months would be maintained. In a previous experiment in coho salmon, where both hormonal and dietary treatments were employed, Shelbourn et al. ( 1992a) reported that the proportion of sterile fish was 100% at the end of the treat- ment period. However, at 7 months of age 4.8% of the fish showed signs of gonadal regeneration, exhibiting severely affected testes but not ovaries, and thus were classilied as partially sterile. Nonetheless, gonadal regeneration was not observed after a similar androgen treatment, which maintained 100% ste- rility at 7.5 months of age (Shelboum et al., 1992b). Furthermore, Solar et al. ( 1986) reported that sterilized coho salmon maintained this condition up to 5 years of age. Thus, in the present study, without an actual sample of adult fish, it is not possible to guarantee the absence of gonadal regeneration after the treatments assayed, although, based on previous studies, it is fairly safe to assume that the proportion of sterile fish observed at 7 months of age would be maintained later. It is also interesting to note that in our experiment the cross-sectional area of sterile gonads was approximately 7 times smaller than

HORMONAL STERILIZATION OF COHO SALMON 421
that of comparable control males, whereas in the study by Shelbourn et al. ( 1992b), where much higher amounts of androgens were administered to the fish, the cross-sectional areas of control males and steriles were similar. The marked reduction in cross-sectional area, observed in the gonads of sterile fish, may be due to the fact that in this study androgen treatments were ap- plied during the labile period and before sexual differentiation, thus further emphasizing the effectiveness of administering steroids for the purposes of sex control in fish specifically at this time.
Regarding practical applications, it may be possible, by carefully selecting the androgen, dose and, perhaps, additional treatments, to obtain 100% ster- ile fish exclusively by immersion. Reducing or eliminating dietary treatment would be of benefit because of the inconvenience that the dietary technique would present if applied on a large production scale. The possibility of steri- lizing fish before first feeding would permit salmonid hatcheries to produce their own stock of sterile fish during the incubation period. This would pre- vent the problem of dietary androgen affecting the performance of zero-age smolts. Additionally, limiting the intervention to the hatchery would facili- tate the monitoring of the process, thus improving the consistency of proto- cols and safety for both operators and the environment. Because of its poten- tial advantages, it is suggested that it would be worthwhile to test this method in other teleosts, especially in those species where sexual differentiation does not take.place until several weeks after hatching.
In conclusion, our study may facilitate further development and regulatory approval of androgen sterilization regimes. We have shown that it is possible to obtain a reasonable and consistently high proportion (in the range of 80- 90%) of sterile coho salmon by discrete immersions of late eyed-eggs and alevins in synthetic androgen solutions. The sterilization rates achieved to date would be insufficient to prevent reproductive interaction between genet- ically modified fish and wild fish. However, the treatments described here represent a significant reduction in the current regimes for hormonal sterili- zation, and the results obtained suggest a possible direction for future research.
ACKNOWLEDGEMENTS
We thank Ian Baker and the staff at the Rosewall Creek Experimental Fish Hatchery for the scrupulous maintenance of the fish, and two anonymous reviewers for their constructive criticism of the manuscript. Part of these studies (Experiment 3) were made possible by NATO Collaborative Re- search Grant CRG-890958 to M.C. and E.M.D.
REFERENCES
Devlin, R.H. and Donaldson, E.M., 1992. Containment of genetically altered fish with empha- sis on salmonids. In: C.L. Hew and G.L. Fletcher (Editors), Transgenic Fish. World Scien- tific, Singapore, pp. 229-265.

422 F. PIFERRER ET AL.
Donaldson, E.M. and Benfey, T.J., 1987. Current status of induced sex manipulation. In: D.R. Idler, L.W. Crim and J.M. Walsh (Editors), Proc. Third Int. Symp. Repr. Phys. Fish., St. John’s, Nfld. Aug. 2-7. Memm. Univ. Nfld., St. John’s, Nfld., pp. 108-l 19.
Donaldson, E.M. and Hunter, G.A., 1982. Sex control in fish with particular reference to sal- monids. Can. J. Fish. Aquat. Sci., 39: 99-l 10.
Donaldson, E.M., Devlin, R.H., Solar, 1.1. and Piferrer, F., 1993. The reproductive containment of genetically altered salmonids. In: J.G. Cloud and G.H. Thorgaard (Editors), Genetic Conservation of Salmonid Fishes. NATO Advanced Study Institute series. Plenum Press, New York, pp. 113- 129.
Goetz, F. W., Donaldson, E.M., Hunter, G.A. and Dye, H.M., 1979. Effects of estradiol- 17p and 17a-methyltestosterone on gonadal differentiation in the coho salmon Oncorhynchus kis- utch. Aquaculture, 17: 267-278.
Goodman, L.A., 1964. Simultaneous confidence intervals for contrast among multinomial pop- ulations. Ann. Math. Stat., 35: 716-725.
Higgs, D.A., Fagerlund, U.H.M., Eales, J.G. and McBride, J.R., 1982. Application of thyroid and steroid hormones as anabolic agents in fish culture. Comp. Biochem. Phys., 73B: 143- 176.
Hunter, G.A. and Donaldson, E.M., 1983. Hormonal sex control and its application to fish culture. In: W.S. Hoar, D.J. Randall and E.M. Donaldson (Editors), Fish Physiology. Vol. 9, Part B: Behaviour and Fertility Control. Academic Press, New York, pp. 223-303.
Hunter, G.A., Donaldson, E.M., Goetz, F.W. and Edgell, P.R., 1982. Production of all-female and sterile coho salmon, and experimental evidence for male heterogamety. Trans. Am. Fish. Sot., 111: 367-372.
Ikuta, K., Aida, K., Okumoto, N. and Hanyu, I., 1987. Effects of sex steroids on the smoltifica- tion of masu salmon (Oncorhynchus masou). Gen. Comp. Endocrinol., 65: 99-l 10.
Lundqvist, H., Borg, S. and Berglund, I., 1989. Androgens impair seawater adaptability in smolting Baltic salmon (Salmo salar). Can. J. Zool., 67: 1733-1736.
McBride, J.R. and Fagerlund, U.H.M., 1973. The use of 17a-methyltestosterone for promoting weight increases in juvenile Pacific salmon. J. Fish. Res. Bd. Can., 30: 1099-l 104.
Miwa, S. and Inui, Y., 1986. Inhibitory effects of 17cu-methyltestosterone and estradiol-17/I on smoltification of sterilized amago salmon ( Oncorhynchus rhodurus) . Aquaculture, 53: 2 l- 39.
Piferrer, F. and Donaldson, E.M., 1989. Gonadal differentiation in coho salmon, Oncorhynchus kisutch, after a single treatment with androgen or estrogen at differehnt stages during onto- genesis. Aquaculture, 77: 251-262.
Piferrer, F. and Donaldson, E.M., 199 1. Dosage-dependent differences in the effect of aroma- tizable and nonaromatizable androgens on the resulting phenotype of coho salmon (Oncor- hynchus kisutch ) . Fish Physiol. Biochem., 9: 145- 150.
Piferrer, F. and Donaldson, E.M., 1992. The comparative effectiveness of the natural and a synthetic estrogen for the direct feminization of chinook salmon (Oncorhynchus tshawyt- scha). Aquaculture, 106: 183-193.
Piferrer, F. and Donaldson, E.M., 1993. Hormonal sex control in Pacific salmon: the impor- tance of treatment timing. In: J.F. Muir and R.J. Roberts (Editors), Recent Advances in Aquaculture, Vol. IV. Blackwell Scientific Publications, Oxford, pp. 67-77.
Piferrer, F., Baker, I.J. and Donaldson, E.M., 1993. Effects of natural, synthetic, aromatizable, and nonaromatizable androgens in inducing male sex differentiation in genotypic female chinook salmon (Oncorhynchus tshawytscha). Gen. Comp. Endocrinol., 91: 59-65.
Shelbourn, J.E., Clarke, W.C., McBride, J.R., Higgs, D.A. and Donaldson, E.M., 1992a. The effect of hormone dose, water temperature and salinity on growth, smolting and histology of underyearling coho salmon (Oncorhynchus kisutch) sterilized by treatment with 17a-meth- yltestosterone. Can. Tech. Rep. Fish. Aquat. Sci., 1869; 15 pp.

HORMONAL STERILIZATION OF COHO SALMON 423
Shelbourn, J.E., Clarke, W.C., McBride, J.R., Fagerlund, U.H.M. and Donaldson, E.M., 199213. The use of 17a-methyltestosterone and 3,5,3’-triiodo-Gthyronine for sterilizing and accel- erating the growth of zero-age coho salmon (Oncorhynchus kisutch). Aquaculture, 103: 85- 99.
Sokal, R. and Rohlf, J., 198 1. Biometry. The Principles and Practice of Statistics in Biological Research, 2nd ed. W.H. Freeman and Co., New York, 859 pp.
Solar, I.I., Baker, I.J., Donaldson, E.M., Hunter, G.A. and Stone, E.T., 1986. Coded wire tag recoveries from the first release of all-female and sterile groups of coho salmon (Oncorhyn- thus kisutch) into the marine environment. Can. Data Rep. Fish. Aquat. Sci., No. 609,29 p.
Copyright © 2022 FDOKUMEN




















