
Mechanical ventilation interacts with endotoxemia to induce extrapulmonary organ dysfunction

doi:10.1182/blood-2003-03-0713Prepublished online February 26, 2004;
Escolar, Bernd Jilma and Nigel S KeyOmer Aras, Arun Shet, Ronald R Bach, Jessica L Hysjulien, Arne Slungaard, Robert P Hebbel, Gines in human endotoxemiaInduction of microparticle- and cell-associated intravascular tissue factor
(973 articles)Phagocytes � (2497 articles)Hemostasis, Thrombosis, and Vascular Biology �
(1086 articles)Gene Expression �Articles on similar topics can be found in the following Blood collections
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
digital object identifier (DOIs) and date of initial publication. theindexed by PubMed from initial publication. Citations to Advance online articles must include
final publication). Advance online articles are citable and establish publication priority; they areappeared in the paper journal (edited, typeset versions may be posted when available prior to Advance online articles have been peer reviewed and accepted for publication but have not yet
Copyright 2011 by The American Society of Hematology; all rights reserved.20036.the American Society of Hematology, 2021 L St, NW, Suite 900, Washington DC Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

1
INDUCTION OF MICROPARTICLE- AND CELL-ASSOCIATED
INTRAVASCULAR TISSUE FACTOR IN HUMAN ENDOTOXEMIA
Omer Aras*,†, Arun Shet*,†, Ronald R Bach*,¶ , Jessica L. Hysjulien*,†, Arne
Slungaard*,† , Robert P. Hebbel*,†, Gines Escolar¢, Bernd Jilma‡, Nigel S. Key*,†
Departments of *Medicine (Hematology, Oncology and Transplantation) and †Vascular
Biology Center, University of Minnesota, Minneapolis, Minnesota; ¶Research Service,
Minneapolis VA Medical Center, Minneapolis, Minnesota; ¢Servicio de Hemoterapia-
Hemostasia, Hospital Clinic, University of Barcelona, Barcelona, Spain; and ‡Department
of Clinical Pharmacology- TARGET, Vienna University, Austria
This work was supported by grant PO-1 HL65578 (N.S.K.) from the National Heart,
Lung, and Blood Institute, National Institutes of Health.
Running Title: Intravascular tissue factor in endotoxemia
Scientific heading: Hemostasis, Thrombosis, and Vascular Biology
Word Count: Abstract: 195
Text: 5,294
Address correspondence to: Dr. Nigel S. KeyUniversity of Minnesota Medical SchoolMayo Mail Code 480420 Delaware St SE Minneapolis, MN 55455, USATel: 612-624-8903 Fax: 612-625-6919 E-mail: [email protected]
Blood First Edition Paper, prepublished online February 26, 2004; DOI 10.1182/blood-2003-03-0713
Copyright (c) 2004 American Society of Hematology
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

2
ABSTRACT
The precise role of intravascular tissue factor (TF) remains poorly defined, due to the
limited availability of assays capable of measuring circulating TF procoagulant activity
(PCA). As a model of inflammation-associated intravascular thrombin generation, we
studied 18 volunteers receiving an infusion of endotoxin. A novel assay which measures
microparticle (MP)-associated TF PCA from a number of cellular sources (but not
platelets) demonstrated an eight-fold increase in activity at 3-4 hours after endotoxin
administration (p < 0.001), with a return to baseline by 8 hours. TF antigen positive MPs
isolated from plasma were visualized by electron microscopy. Inter-individual MP-
associated TF response to LPS was highly variable. In contrast, a previously described
assay that measures total (cell and MP-borne) whole blood TF PCA demonstrated a more
modest increase, with a peak in activity (1.3 fold over baseline; p < 0.00001) at 3-4
hours, and persistence for > 24 hours. This surprisingly modest increase in whole blood
TF activity is likely explained by a profound although transient LPS-induced
monocytopenia. MP-associated TF PCA was highly correlated with whole blood TF PCA
and total number of circulating MPs, and whole blood TF PCA was highly correlated
with TF mRNA levels.
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

3
Glossary of Abbreviations:
TF: Tissue factor; PCA: Procoagulant activity; MP: Microparticle; LPS:
Lipopolysaccharide; PRP: platelet-rich plasma; PFP: platelet-free plasma; 1B10: IgM
monoclonal mouse anti-human fibroblast surface protein; HUVEC: Human umbilical
vein endothelial cell; MVEC: Human dermal microvascular endothelial cell; BOEC:
human blood outgrowth endothelial cell; CACD: citric acid-citrate-dextrose; EM:
electron microscopy
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

4
INTRODUCTION
Tissue factor (TF), a 47kDa transmembrane glycoprotein, is the principal activator of
coagulation in vivo. The procoagulant activity (PCA) of factor VIIa (FVIIa) is markedly
enhanced after binding to TF, with the subsequent activation of factors IX and X. Current
evidence suggests that basal activation of the coagulation system in vivo is mediated by
the TF/FVIIa complex. Specifically, plasma levels of factors IX and X activation peptides
are markedly suppressed following administration of a potent monoclonal antibody to TF
in normal chimpanzees.1,2 Furthermore, endotoxin-induced activation of coagulation in
these animals can be blocked by the same monoclonal antibody, indicating that this
response is also mediated via the TF pathway.3
These observations do not address whether contact between TF and FVIIa occurs
in the intravascular or extravascular compartment, either at baseline or during
endotoxemia. TF antigen is not normally detectable in endothelial cells or monocytes in
vivo. 4,5 Despite the demonstrated ability of these cells to synthesize TF following
exposure to endotoxin in vitro, 6,7 TF antigen expression could only be demonstrated in
splenic endothelial cells in baboons infused with lipopolysaccharide (LPS).8 However,
recent evidence suggests that intravascular TF may be important in both the initiation and
propagation phases of coagulation.9-12 In these studies, whole blood was shown to contain
circulating TF-bearing microparticles (MPs) derived from leucocytes which may be
transferred to a growing platelet-rich thrombus. Numerous studies have documented the
presence of intravascular TF-bearing MPs in selected disease states.13-16 TF may also be
present in the alpha-granules of circulating platelets.17
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

5
MPs are derived from cells undergoing activation or apoptosis. The most rigorous
criteria for definition of MPs are by size (< 1.0-1.5µm), and by expression of negatively
charged phosphatidylserine in the outer leaflet of the membrane bilayer.18 The
physiologic and pathophysiologic functions of these MPs have yet to be defined,
although evidence suggests that they may be able to facilitate cell-cell cross-talk.19,20
The purpose of this study was to characterize the time course of intravascular TF
expression in both the cellular and MP fractions of blood during human endotoxemia, an
accepted model of the early coagulation/inflammatory changes during sepsis.21 Whole
blood TF PCA was measured using a previously described method.22,23 This assay is very
sensitive and specific for TF PCA and avoids the potential for artifactual increase in TF
activity which may occur during the isolation of monocytes.24 However, since all blood
cells are converted to sub-cellular fragments and MPs by freeze-thawing, the assay does
not distinguish cell-borne TF from MP- associated TF. We therefore developed a new
assay that specifically quantifies MP-associated TF PCA in platelet-free plasma (PFP).
Our data suggest that after endotoxin exposure, there is an early and robust increase in
MP-associated TF PCA, accompanied by a more modest but sustained elevation in whole
blood TF PCA.
MATERIALS AND METHODS
Materials
LPS from Escherichia coli 055:B5 and A23187 were from Sigma Chemical Co. (St.
Louis, MO). Human FVIIa and FX were from Enzyme Research Lab. Inc. (South Bend,
IN, USA). S2756 was purchased from Chromogenix (Molndal, Sweden). IgM
monoclonal mouse anti-human fibroblast surface protein (1B10), mouse IgM isotype
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

6
control for 1B10, and alkaline phosphatase-conjugated goat anti-mouse IgM were from
Sigma. Rhodamine Red-X goat anti-mouse IgG +IgM (H+L), FITC-conjugated goat anti-
rabbit IgG (H+L), FITC-labeled goat anti-mouse IgG +IgM(H+L), 12nm colloidal gold
goat anti-mouse IgG + IgM (H+L), and 6nm colloidal gold donkey anti-rabbit IgG (H+L)
antibodies were from Jackson Immuno-Research Laboratories Inc. (West Grove, PA). CY
5-Annexin V and anti-CD144 (VE cadherin) for endothelial cells (55-7H1) were from
Pharmingen (BD Biosciences, Lexington, KY), PE-labeled anti-CD 14 (IgG1) was from
Biosource (Camarillo, CA), and labeled isotype control antibodies (IgG1-FITC and -PE)
were from R&D systems (Minneapolis, MN). Fluorescein isothiocyanate (FITC)-labeled
MoAb against TF (CJ4068) was obtained from American Diagnostica (Greenwich, CO).
The rabbit polyclonal blocking antibody to TF (and pre-immune IgG control) have been
previously described.25 Innovin™ (recombinant re-lipidated human TF) was from Dade
(Dade International Inc, Miami, FL).
Study design and subjects
The study design was approved by the Institutional Human Subjects Committee. Written
informed consent was obtained from all participants. Criteria for selection of volunteers
and study protocol were as previously described. 26-28 Eighteen healthy male volunteers
ages 18 to 35 (body mass index range 19 to 27.4 kg/m2) were invited to participate in this
study. These subjects were admitted to the study ward after an overnight fast. After
baseline blood sampling, 2 ng/kg LPS (National Reference Endotoxin, Escherichia coli;
USP Convention, Rockville, MD) was administered as an intravenous infusion over 1-2
minutes. Venous blood samples were collected into EDTA-Vacutainer tubes (Becton
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

7
Dickinson, San Jose, CA) at baseline (3 hours before) and at 1, 2, 3, 4, 8, and 24 hours
after LPS administration, applying minimal venostasis. After discarding the first 2 ml,
aliquots were transferred immediately to polypropylene tubes and frozen at -70° C for
later assay of whole blood TF PCA. Platelet-free plasma (PFP) was obtained by double
centrifugation, first at 1,200×g for 15 minutes, and subsequently at12,000×g for 12
minutes at 20°C. PFP was stored at -70°C for future assay of MP-associated TF PCA, as
described below.
Healthy volunteer samples, prepared and stored identically, were used for
comparison. These individuals had similarly not taken any medication for at least ten
days prior to blood collection.
In vitro stimulation of whole blood with LPS
Following informed consent, heparinized whole blood was collected using a 19G needle
(after discarding the first 3 mL) from healthy donors. Whole blood samples were
stimulated with LPS (10ng/ml) at 37°C, with gentle rocking, for up to 24 hours. PFP was
then obtained by sequential centrifugation as described above.
Cell cultures
Human umbilical vein endothelial cells (HUVEC) were isolated and cultured as
previously described.29 Human dermal microvascular endothelial cells (MVEC) were
isolated from newborn human foreskin and cultured as previously described.30 A culture
of human blood outgrowth endothelial cells (BOEC) was established from venous blood
of a healthy donor from which buffy coat mononuclear cells were isolated using
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

8
Histopaque-1077 (Sigma Chemical, St Louis, MO), as previously described.31 Human
aortic smooth muscle cells and human skin fibroblasts were from ATCC (Manassas, VA).
Smooth muscle cells were routinely identified by their appearance on light microscopy,
and by immunostaining with antibody (1:200 dilution) to human smooth muscle actin
(Oncogene Research Products, Boston, MA).
Peripheral blood cell isolation
Platelets were obtained from citric acid-citrate-dextrose (CACD) anticoagulated blood by
centrifugation at 130 ×g for 20 minutes. PRP was then diluted 1:1 with 0.1mol/L CACD,
pH 6.5 and centrifuged at 830×g for a further 20 minutes. The pelleted platelets were
washed in CACD and resuspended in hepes buffer. Erythrocytes, mononuclear, and
polymorphonuclear (PMN) cells were isolated from EDTA-anticoagulated blood using
PMN isolation medium (Robbins Scientific Co. Sunnyvale,CA) by density separation.
Monocytes were isolated by CD 14 immuno-magnetic beads and a magnetic column
(MACS Miltenyi) according to the manufacturer's instructions (Miltenyi Biotec, Auburn,
CA).
Cellular antigen detection by flow cytometry
Binding of 1B10 antibody to cells of interest was assessed by flow cytometry. Briefly,
106 cells were incubated with 1B10 or control murine IgM monoclonal antibody in the
dark at room temperature for 30 minutes, washed once and incubated with FITC-labeled
goat anti-mouse IgG + IgM. After washing twice with PBS and fixing with 2%
paraformaldehyde, samples were acquired on a Becton Dickenson FACS Calibur and
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

9
analyzed using Cellquestpro™ software. Mean fluorescent intensity (MFI) of the positive
cell populations for 1B10 antigen were recorded.
Generation of microparticles following in vitro cellular activation
Cultured fibroblasts and smooth muscle cells, and isolated platelets and red blood cells
were induced to generate MPs by exposure to calcium ionophore (10µM A23187) for 10
minutes.32,33 Control cells were exposed to vehicle alone. Supernatants were then
removed, and subjected to double centrifugation to remove contaminating intact cells.
Similarly, MPs were harvested from the supernatant of HUVECs following exposure to
10ng/mL TNF-α (or vehicle control) for 4 hours,34 and from monocytes following
exposure to 10 ng/mL LPS (or vehicle control) for 4 hours.35 The MP-rich supernatant
was stored at -80°C for assay of MP-associated TF PCA. Baseline MP-associated TF
PCA was defined as the TF activity in the supernatant of cells that had been exposed to
neither activator nor relevant vehicle control.
Isolation of MPs from PFP
PFP was diluted (1:400) with 0.2µ-filtered wash buffer (10mM Hepes, 144mEq NaCl,
5.3mEq KCl, 2.5mM CaCl, 0.5% BSA and 0.01% sodium azide, pH 7.4) and
ultracentrifuged twice at 100,000×g for 60 minutes at 20°C. After careful removal of all
but 1 mL of the supernatant, MPs were then re-suspended by gentle vortexing and
pipetting. MP suspensions were stored at -80°C for flow cytometry.
Flow cytometric analysis of MPs
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

10
Samples of MPs isolated from blood were analyzed by flow cytometry using a Becton
Dickinson FACS Calibur, and analyzed using cellquestpro™ software (San Jose, CA)
using described methods.36 For MP quantification, we used an internal standard
consisting of calibrated polypropylene beads (Bangs Laboratories, Inc., Fisher, IN) added
to the sample before acquisition. A known quantity (250,000) of 7.2 µm beads was added
to each sample and acquisition was terminated after 10,000 beads were counted. Events
falling in the pre-defined MP gate (i.e. ≤ 1.0 µm) (Figure 1a) that were annexin V(+)
(Figure 1b) were defined and enumerated as MPs. Similar enumeration of cell-derived
MPs was performed on blood samples that been stimulated with LPS both in vitro and in
vivo. The number of MPs per mL of plasma was calculated as described elsewhere.36 To
enumerate total MPs and monocyte-derived MPs, aliquots of the isolated washed MPs
were incubated with Cy 5-labeled annexin V + anti-CD 14 PE (Figure 1d, 1f). For each
sample, control labeling was performed in parallel by incubating an isotype control
MoAb or annexin V Cy 5 in EDTA-containing buffer (to prevent Ca2+ dependent annexin
V binding). Endothelial cell-derived MPs were similarly defined as those events that
were specifically labeled with both annexin V Cy5 and anti-CD144-PE (Figure 1h).
Capture assay for MP-associated TF PCA
Fifty microliters of 1B10 antibody (40µg/ml in PBS) were added to each well of a 96-
well microtiter plate (Nunc MaxiSorp) and incubated overnight at 4°C. After 2 washes
with PBS, 365 µL of 3% bovine serum albumin in PBS was added to each well, and
incubated overnight at 4°C.
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

11
Fifty microliters of sample (PFP or conditioned cell supernatant) were added to
each well and incubated for 24 h at 4°C. Plates were then washed 4 times with 360
µL/well of PBS. Tissue factor activity associated with the captured MPs was evaluated
by the activation of FX (150nM) by FVIIa (5nM) in the presence of 5 mM CaCl2. After a
6 h incubation at 37°C, the reaction was terminated by the addition of 25mM EDTA
(25µL). The prewarmed S-2765 chromogenic substrate for FXa (S2765) was then added
at a final concentration of 1.47 mM and incubated for 1 h at 37°C. The absorbance was
measured in an ELISA plate reader at 405nm. A standard curve was constructed using
known concentrations of FXa incubated for 1 hour with the chromogenic substrate.
Confirmation that the PCA was indeed due to TF was established by including an
inhibitory polyclonal antibody to TF in separate duplicate wells prior to addition of
Factors VIIa and X. For all samples, residual “non specific” procoagulant activity
(usually <5%) was subtracted from “total” procoagulant activity, and the remaining
activity was taken as a measure of TF-dependent PCA.
Electron microscopy (EM) and immuno-gold labeling of MPs
The MP pellet was pre-fixed with 2% glutaraldehyde, post-fixed with 2% OsO4, and then
washed and dehydrated in graded alcohols. The pellet was then embedded in Epon resin.
Sections were cut and photographed on a Philips 400 T electron microscope (Phillips
Electronic Instruments, Inc., Mahwah, NJ).
For immuno-gold labeling, the MP pellet was re-suspended in wash buffer and
incubated with a rabbit polyclonal anti-TF antibody (1:500 dilution), followed by 6nm
anti-rabbit IgG-Gold (1:250 dilution). After immuno-labeling, the samples were layered
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

12
for 10 minutes on 0.1% polylysine treated formbar coated grids. Grids with adherent
vesicles were rinsed in PBS and fixed with 0.1% glutaraldehyde in PBS. Grids were then
examined in the electron microscope. For CD14 labeling of MPs, the pellet was
incubated with a mouse anti-CD14 IgG antibody (1:100 dilution), followed by 12 nm
anti-mouse IgG (1:100). For 1B10 labeling of MPs, the pellet was incubated with 1B10
antibody (1:250), followed by 12 nm gold-labeled anti-mouse IgM (1:100). For double
labeling to TF and 1B10 or CD14, anti-TF antibody followed by 1B10 or anti-CD14
antibody were sequentially added as described above. Prior to addition of the secondary
antibodies, grids were washed x3 with PBS. The specificity of immuno-labeling was
verified using control rabbit IgG, mouse IgG, mouse IgM, or secondary gold-tagged
antibody alone.
Whole blood TF PCA
Whole blood TF PCA was assayed on frozen whole blood samples exactly as previously
described,22,23 except for the use of a commercially available thromboplastin, Innovin™,
in place of relipidated human brain TF as the standard in the two stage clotting assay. The
concentration of TF apoprotein in the Innovin™ reagent was first measured using a
previously described ELISA assay.25 Repeated estimates of the reagent lot utilized in
these experiments gave a value of 93.5 ng/mL.
Whole blood TF mRNA expression
TF mRNA expression was performed according to Franco et al 37 with modifications.
After preparing total RNA with the QuiAmp RNA easy kit (Quiagene, Valencia, CA),
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

13
mRNA was directly transcribed into cDNA using the RT- Reagent kit (Applied
Biosystems, Foster City, CA) and stored at - 80°C until analysis. For TF mRNA
quantification the Abi Prism 7700 (Applied Biosystems) was used. Primers were
designed by Primer Express Software (Applied Biosystems) and synthesised based on the
human TF cDNA sequence(Applied Biosystems): 680 F 5-CCCGAACAGTTAACC
GGAAGA-3 and 773R 5-GCTCCAATGATGTAGAATATTTCTCTGA-3, TaqMan
probe 711T FAM CTCCTGGCCCATACACTCTACCGGG TAMRA. 18s was
used as a housekeeping gene for multiplexing (Applied Biosystems) because of its stable
expression under endotoxemia (unpublished data). TF-mRNA was normalised against the
reference gene (18s) and data expressed as fold-increase over baseline values. Dilution
curves of TF-mRNA obtained from LPS-incubated blood samples revealed linearity
(r=0.999) of the assay up to 37.5 cycles, which was set as the limit of sensitivity (and also
as a baseline for conservative statistical calculations).
Statistics
Data are expressed as mean ± SEM, unless otherwise indicated. Non-normally distributed
data was analyzed with the Mann-Whitney U test. The Bonferroni method was used to
correct for the multiple comparisons. Bivariate correlations between parameters were
calculated with the use of Spearman’s rank correlation test. P <0.05 was considered to be
statistically significant. Statistical analysis was performed with SPSS (version 10.0,
Chicago, IL).
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

14
RESULTS
Characterization of the MP-associated TF PCA capture assay
Since the solid phase capture assay was dependent on broad cellular recognition of MPs
by the 1B10 antibody, we first determined whether the antigen recognized by 1B10 is
expressed by cell types that could be the source of circulating intravascular MP-borne TF.
1B10 is a complement fixing murine IgM monoclonal antibody that was originally
produced by immunization of mice with cultured human thymic fibroblasts.38 The antigen
recognized by this antibody was known to be expressed on fibroblasts, as well as tissue
macrophages and peripheral blood monocytes, but not epithelial cells or lymphocytes.38,39
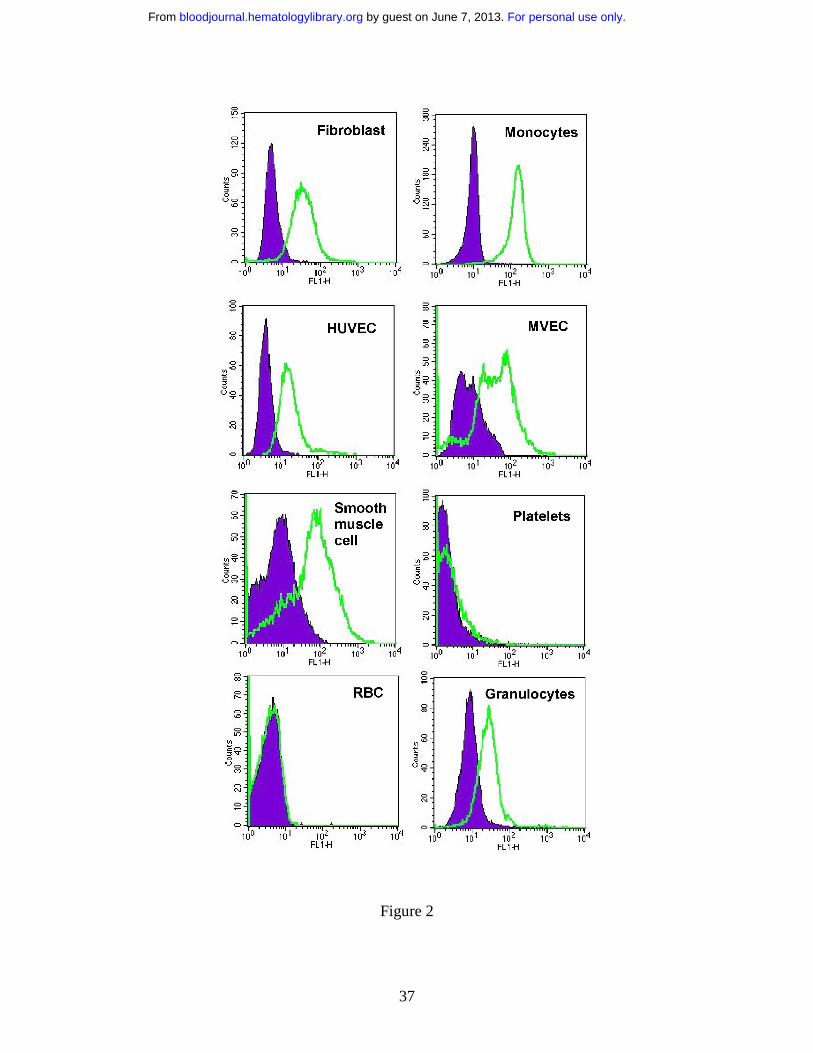
By flow cytometry, we found that 1B10 bound not only to fibroblasts and
monocytes, but also to smooth muscle cells, granulocytes and endothelial cells, regardless
of whether the latter were of large vessel (umbilical vein) or small vessel (foreskin) origin
(Figure 2). On the other hand, red blood cells and platelets were not recognized. Direct
demonstration of 1B10 binding to MPs by flow cytometry could not be satisfactorily
demonstrated due to high background fluorescence in filtered buffer samples containing
unlabeled 1B10 (or control murine IgM) in the presence of the secondary anti-mouse
antibody. No such fluorescence was detected with buffer ± secondary antibody. This
artifact was presumed to be due to aggregation or multimerization of IgM antibody.
In order to determine whether the MP capture assay would detect MP-associated
TF PCA produced by the individual cell types of interest, we prepared MPs from each
cell type after in vitro activation. As previously described, TF-bearing MPs were easily
detectable in the supernatant of smooth muscle cells and fibroblasts, even without cellular
activation 32,33 (Table 1). Constitutive generation of TF-bearing MPs was barely
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

15
detectable in supernatants from monocytes and endothelial cells, but increased after
activation by LPS and TNF-α, respectively, as previously described.34,35 Red cells and
platelets which produce MPs in response to calcium ionophore stimulation, but were not
recognized by 1B10, were included as a negative control.
MP-associated TF PCA is detectable in normal subjects
PFP was obtained from 40 normal volunteers (25 male, 15 female, age range 19 to 54)
and assayed in duplicate for MP-associated TF PCA. All samples had detectable TF
PCA, with a mean Xa generation of 3.96±2.58 pmol/L/hr (mean ± SD). No significant
gender difference in TF activity was detected (3.70±2.28 vs 4.40±3.04 pmol/L/hr, males
vs females, respectively).
In Vitro stimulation of whole blood by LPS leads to increased MP-associated TF
PCA
Heparin-anticoagulated whole blood from 5 normal volunteers was exposed to 10 ng/mL
LPS in vitro for up to 24 hrs at 37°C. PFP was prepared from these samples, and assayed
for MP-associated TF PCA. As shown in figure 3, there was a time-dependent increase in
TF activity in the PFP fraction, although the magnitude of the response varied markedly
between individuals. Since monocytes are presumably the sole source of inducible
activity, the variability in MP-borne TF response to LPS is seemingly analogous to the
previously described variability in monocyte-borne TF, which has been termed the
‘high/low LPS responder phenomenon’.40-42 On repeated testing, we found the MP tissue
factor response to be highly consistent for any given individual (data not shown).
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

16
Circulating in vivo MP-associated TF PCA increases after endotoxin administration
MP-associated TF PCA was detectable in all volunteers at baseline, and increased in most
subjects after endotoxin administration. Mean TF activity increased approximately 8-fold
with a peak at 3 to 4 hours, and a return to baseline levels by 8 hours (Figure 4a). Similar
to the in vitro experiment, considerable inter-individual variability of TF expression in
MPs was observed (Figure 4b), probably reflecting the 'high' and 'low' LPS responder
phenomenon. It should be noted that a marked (> 90%) but transient decrease in
peripheral blood monocyte counts occurs between 1.5 and 6 hours after LPS
administration in this model.27,28 The rapid disappearance of MP-borne TF in vivo
compared to the in vitro experiment probably reflects clearance of MPs. Visual
inspection of Figure 4a produces a crude estimate of the in vivo half life of MPs of < 2
hours.
Increase in total numbers of circulating MPs occurs after endotoxin administration
We determined whether endotoxemia produced an increase in the total number of
circulating MPs in a time course that mirrored that of MP-associated TF PCA. Flow
cytometry was used to enumerate MPs in blood obtained from 13 randomly selected
study participants at baseline and at various time intervals up to 8 hours. Additionally, we
specifically enumerated monocyte-derived MPs, defined as those events that were also
CD14 (+). As shown in Table 2, 3-4 hours after LPS administration, the total number of
MPs had increased approximately 2-fold (from 727 ±131 x 103 MPs/ml at baseline to
1469 ± 267 x 103 MPs/ml at 3 hours; P< 0.05). A parallel increase in the number of CD14
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

17
(+) monocyte-derived MPs (from 104 ±42 x 103 MPs/ml at baseline to 227 ±57 x 103
MPs/ml at 3 hours; P < 0.05) was also apparent. Both the total number of MPs, as well as
the number of CD14 (+) MPs had returned to baseline values by 8 hours. There was a
significant linear correlation between the total number of MPs and the number of CD14
(+) monocyte-derived MPs (Table 3). In addition, MP-associated TF PCA was highly
correlated with the total number of MPs, though not with the number of CD14 (+) MPs.
Because monocyte-derived MPs always accounted for < 20% of total MPs (Table
2), we wondered whether LPS-activated endothelial cells (EC) might contribute to the
expanded pool of circulating MPs at 3-4 hours. In fact, while an increase in EC-derived
(annexin V (+)/VE cadherin (+)) MPs was detected after LPS administration in 3
individuals, the absolute numbers were relatively small (3±3 x 103 MPs/ml at baseline,
48±35 x 103 MPs/ml at 3 hours, and 84±84 x 103 MPs/ml at 8 hours) (Table 2). In a
previous study, 36 we found that platelets account for the majority of circulating MPs in
plasma from normal individuals, although we did not systematically enumerate platelet-
derived MPs in this study.
The number of TF (+) (annexin V (+)/anti TF (+)) MPs was enumerated in 3
individuals (Table 2). Unlike our experience in patients with sickle cell disease, 36 our
attempts to define the cellular origin of TF (+) MPs by triple labeling experiments
produced numbers that were too small to accurately quantify.
Identification of TF on circulating MPs from endotoxin-treated volunteers
Additional experiments were performed to confirm that the TF PCA detected by the
capture assay was in fact associated with circulating MPs. Firstly, PFP from two of the
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

18
highest LPS-responding individuals was ultracentrifuged (400,000×g for 1 hr), after
which the supernatant plasma was carefully decanted, and assayed for MP-associated TF
PCA. As seen in Figure 5, this maneuver removed essentially all of the detectable TF
PCA from solution. Electron microscopic examination of the pellet obtained by
ultracentrifugation demonstrated the presence of TF antigen on MPs as shown in Figure
6b. These in vivo generated MPs are seen as vesicular structures with diameters ranging
from 0.1 to 0.5 µm, which do not contain mitochondria or other recognizable cellular
organelles (Figure 6a). Immuno-gold labeling of MPs demonstrated positive labeling
with anti-TF (Figure 6b), anti-CD14 (Figure 6e), and 1B10 (Figure 6h) antibodies as well
as double labeling with 1B10 and anti-TF (Figure 6k). No labeling was observed when
MPs were incubated with control isotype or secondary gold particle labeled antibodies
alone. The presence of MPs that were positive for both CD14 and TF (Figure 6n)
establishes that at least some circulating TF (+) MPs originated from monocytes.
Cell surface PCA may be regulated by several mechanisms, including encryption
of membrane-bound TF.43 Encryption is defined as the ability of TF to bind its ligand,
FVIIa, while failing to express full PCA. This activity may be 'decrypted' by maneuvers
such as trypsinization, freeze-thawing, or exposure to calcium ionophore.43-45 The
phenomenon may at least in part be explained by loss of phospholipid asymmetry in the
membrane bilayer 43 and is linked to the process of MP generation.45 We thus sought to
determine whether expression of MP-associated TF PCA could be further enhanced by
'decryption' maneuvers. Unlike cell-bound TF in cultured cells or freshly isolated
monocytes, where a 10-100 fold increase in apparent TF PCA is typically seen after
freeze-thawing or ionophore stimulation,44 no incremental rise in TF PCA could be
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

19
demonstrated in MPs derived from either cultured cell lines or isolated from PFP of
endotoxin recipients (data not shown). Therefore by these criteria, MP-borne TF
circulates in a form that is already maximally decrypted, a finding consistent with the fact
that membrane phospholipid asymmetry is frequently lost on circulating MPs.18
However, it cannot be assumed that decryption necessarily implies greater functional
importance of the TF in vivo, since inhibition by TFPI may provide a counter-balancing
anticoagulant effect.
Whole blood TF PCA and TF mRNA increase after endotoxin administration
Whole blood TF PCA also increased in a time-dependent manner, with a more prolonged
duration than MP-associated TF PCA (to at least 24 hours) after endotoxin exposure.
Although peak activity was also observed at 4 hours, the absolute increase was relatively
modest (≈ 30% over baseline) compared to MP-associated TF (Figure 7). An
approximate 25-fold increase [95%CI: 12-39 fold] in TF mRNA was noted. This activity
peaked at about 3 hours (p < 0.00001), and was still elevated at 5 hours (p <0.0001) but
had returned to baseline by 24 hours. The increase in whole blood TF PCA was highly
correlated with TF mRNA and MP-associated TF activity, but not with the total number
of MPs, nor the number of monocyte-derived CD14 (+) MPs (Table 3).
DISCUSSION
In these studies, we used two different functional assays to measure circulating
intravascular TF PCA in human volunteers exposed to endotoxin. One was designed to
measure MP-associated TF PCA in PFP, following the capture of these MPs by the
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

20
monoclonal antibody 1B10. This antibody recognizes an unidentified antigen on the
surface of various cell types including monocytes, granulocytes, endothelial cells, smooth
muscle cells, and fibroblasts, but not platelets. Any or all of these cells could be a
potential source of circulating TF-bearing MPs in vivo. Although technical problems
precluded flow cytometry demonstration of 1B10 binding to MPs derived from some or
all of these cells, direct binding was shown by electron microscopy (Figure 6).
Furthermore, the ability of 1B10 to capture MPs is strongly supported by the fact that
measurable TF activity can be pelleted by ultracentrifugation of plasma (Figure 5), or
detected in the cell-free supernatants of cultured cell lines (Table 1).
Circulating MP-associated TF PCA was present in normal controls at baseline,
consistent with the findings of one group,46 but in contrast to another study in which
platelet, red cell, granulocyte, and endothelial cell-derived MPs were found to support
coagulation in a TF-independent manner.47 We did not routinely examine MPs isolated
by ultracentrifugation for non TF-dependent PCA as others have done. However, in
preliminary experiments, we did note that the PCA on MPs isolated from plasma by
ultracentrifugation is often significantly less TF-dependent than the PCA on MPs
captured by the 1B10 antibody (data not shown). While the reason(s) for this discrepancy
remain(s) unclear, we postulate that MP-associated TF PCA (the focus of these studies) is
likely to be augmented by non TF-dependent PCA in vivo.
Endotoxin stimulation of monocytes in vitro leads to the generation of TF-bearing
MPs with a similar time course seen using our MP-associated TF PCA assay (Figure 3).
However, the lack of correlation between MP-associated TF PCA and number of
monocyte-derived MPs in vivo requires explanation. One possibility is that monocyte-
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

21
derived MPs are selectively enriched in TF, 48 or that the increase in MP-associated TF
activity is due to release of MPs bearing TF by cells other than monocytes. Endothelial
cells could certainly contribute to the TF (+) MP pool, and indeed we have demonstrated
that they do so in other disease states. 36 Others have shown that platelet-derived MPs
may contain TF, 13 although we are unable to confirm this observation using our methods.
36 In this study, we identified only a modest number of EC-derived MPs, although
because of low event numbers, we were unable to accurately quantify the proportion of
EC (or monocyte)-derived MPs that were TF (+) by triple labeling flow cytometry
experiments. We did note a significantly larger pool of flow cytometry events that were <
1 µm in size, TF (+), and either VE-cadherin or CD 14 (+) (data not shown). However,
because these events were annexin V (-) -- and therefore failed to meet our criteria for
MPs -- we did not enumerate them. Thus, the possibility also exists that the lack of
correlation between the number of MPs on flow cytometry and MP-associated TF activity
may be explained by the precise criteria we chose to define MPs.
The increase in MP-associated TF PCA coincides with the profound
monocytopenia (> 90% reduction compared to baseline) that occurs from about 1.5 to 6
hours following endotoxin exposure in this model.27,28 However, individual monocytes
remaining in the circulation at 6 hours following endotoxin exposure demonstrate
elevated TF antigen expression, 27 and about a 15-fold increase in monocyte-associated
TF PCA. 49 The monocytopenia appears to be of a sufficiently severe degree to account
for the modest rise in whole blood TF PCA that we observed. The persistent increase in
whole blood TF PCA at a time when MP-associated TF PCA and total number of MPs
had returned to baseline (8-24 hours) suggests that the majority of this activity resides in
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

22
the cell-bound, rather than the non-cell-bound (i.e. microparticle) fraction of blood. This
impression is supported by the tight correlation between TF mRNA and whole blood TF
PCA levels, but not MP-associated TF PCA.
Although it is now accepted that coagulation is activated via the TF pathway in
endotoxemia and sepsis, much of the supporting evidence is indirect, originating from
therapeutic intervention studies. Administration of TF inhibitors -- such as tissue factor
pathway inhibitor (TFPI), 50 active site-inhibited factor VIIa, 51 or recombinant nematode
anticoagulant protein C2 52 -- to human endotoxin volunteers dose-dependently inhibits
thrombin generation in vivo. Direct evidence for the up-regulation of TF expression
includes the demonstration that whole blood TF mRNA levels increase after endotoxin
administration.37 Osterud and colleagues have demonstrated that monocyte TF activity is
increased in certain forms of sepsis,41 but the more widespread use of their assay has been
limited by the fact that it is technically cumbersome and prone to artifactual ex vivo
activation of monocytes during isolation.24 Functional assays of TF based on modified
whole blood clotting times have also been proposed.53,28 Using one such assay, Marsik et
al 28 recently demonstrated that a proportion of the total PCA in whole blood following
endotoxin administration is TF-dependent, although the presence of ‘downstream’
activated clotting factors also contribute to the shortening of the clotting time.
Plasma TF antigen by ELISA has also been used as a measure of TF expression in
vivo, although this assay does not provide any information on the origin or functional
competence of circulating TF. It has been reported that plasma TF antigen levels remain
persistently elevated between 8 and 48 hours after endotoxin administration, 54 although
we were unable to confirm these findings.27 Since only one third to one half of plasma TF
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

23
antigen can be sedimented by high speed centrifugation, it has been proposed that the
assay may in fact measure two forms of TF antigen in plasma –- soluble and MP-
associated.55 This view is supported by a recent study showing that 45-77% of "non cell
bound" TF in pericardial blood of patients undergoing cardiac surgery is MP-associated
and functionally active, whereas the remaining fluid phase TF antigen is non-functional.
56 Because soluble TF lacking the transmembrane domain is not fully functional,57 the
correlation between TF antigen levels and our assay for whole blood TF PCA is likely to
be poor.
The suggestion that MP-associated TF plays an important role in the
coagulopathy of sepsis has been previously addressed using other approaches. Nieuwland
and colleagues demonstrated that patients with meningococcal sepsis have elevated
numbers of circulating MPs that were derived from multiple cell types, especially
granulocytes and platelets. However, 85% or more of TF positive MPs were of
monocytic origin.15 Curiously however, the same group found that the number of TF
positive MPs was decreased in patients with multiple organ dysfunction syndrome and
(non meningococcal) sepsis compared to normals.47 Whether this apparent paradox
reflects the acuity of the various forms of sepsis is unclear.
We also found that the endotoxin-induced increase in TF PCA on circulating MPs
was highly variable between individuals. We believe this finding represents the in vivo
analog of the previously described “high vs low responder” phenotype phenomenon,
which is used to describe the variable monocyte responsiveness to LPS in whole blood in
vitro.40-42 The precise explanation for this phenomenon is unclear, but may reflect
individual differences in plasma,40 leukocyte,42 or platelet 58 factors. The absolute
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

24
increase in TF activity in peripheral blood monocytes was demonstrated to have
prognostic significance in meningococcal sepsis, with higher levels portending a poorer
outcome.42 Whether the same holds true for whole blood and/or MP-associated TF PCA
is an eminently testable hypothesis.
Acknowledgement: The authors wish to thank Dr. Gary Nelsestuen for helpful
discussions. The technical assistance of Julia Nguyen in the isolation and culture of
endothelial cells, Marcie Krumwiede in the preparation of the EM mounts, Nicole
Henderson in sample collection and processing, and Carol Taubert in manuscript
preparation is gratefully acknowledged.
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

25
REFERENCES
1. Bauer KA, Mannucci PM, Gringeri A, et al. Factor IXa-factor VIIIa-cell surface
complex does not contribute to the basal activation of the coagulation mechanism
in vivo. Blood 1992;79:2039-47.
2. ten Cate H, Bauer KA, Levi M, et al. The activation of factor X and prothrombin
by recombinant factor VIIa in vivo is mediated by tissue factor. J Clin Invest
1993;92:1207-12.
3. Levi M, ten Cate H, Bauer KA, et al. Inhibition of endotoxin-induced activation
of coagulation and fibrinolysis by pentoxifylline or by a monoclonal anti-tissue
factor antibody in chimpanzees. J Clin Invest 1994 ;93:114-20.
4. Wilcox JN, Smith KM, Schwartz SM, Gordon D Localization of tissue factor in
the normal vessel wall and in the atherosclerotic plaque. Proc Natl Acad Sci U S
A 1989; 86:2839-43.
5. Drake TA, Morrissey JH, Edgington TS. Selective cellular expression of tissue
factor in human tissues. Implications for disorders of hemostasis and thrombosis.
Am J Pathol 1989;134:1087-97.
6. Rivers RP, Hathaway WE, Weston WL. The endotoxin-induced coagulant activity
of human monocytes. Br J Haematol 1975 ;30:311-6.
7. Colucci M, Balconi G, Lorenzet R, et al. Cultured human endothelial cells
generate tissue factor in response to endotoxin. J Clin Invest 1983;71:1893-6.
8. Drake TA, Cheng J, Chang A, Taylor FB Jr. Expression of tissue factor,
thrombomodulin, and E-selectin in baboons with lethal Escherichia coli sepsis.
Am J Pathol 1993;142:1458-70.
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

26
9. Giesen PL, Nemerson Y. Tissue factor on the loose. Semin Thromb Hemost
2000;26:379-84.
10. Rauch U, Nemerson Y. Tissue factor, the blood, and the arterial wall. Trends
Cardiovasc Med 2000;10:139-43.
11. Balasubramanian V, Grabowski E, Bini A, Nemerson Y. Platelets, circulating
tissue factor, and fibrin colocalize in ex vivo thrombi: real-time fluorescence
images of thrombus formation and propagation under defined flow conditions.
Blood 2002;100:2787-92.
12. Rauch U, Bonderman D, Bohrmann B, et al. Transfer of tissue factor from
leukocytes to platelets is mediated by CD15 and tissue factor. Blood 2000
;96:170-5.
13. Diamant M, Nieuwland R, Pablo RF, Sturk A, Smit JW, Radder JK. Elevated
numbers of tissue-factor exposing microparticles correlate with components of the
metabolic syndrome in uncomplicated type 2 diabetes mellitus. Circulation
2002;106:2442-7.
14. Mallat Z, Benamer H, Hugel B, et al. Elevated levels of shed membrane
microparticles with procoagulant potential in the peripheral circulating blood of
patients with acute coronary syndromes. Circulation 2000;101:841-3.
15. Nieuwland R, Berckmans RJ, McGregor S, et al. Cellular origin and procoagulant
properties of microparticles in meningococcal sepsis. Blood 2000;95:930-5.
16. Berckmans RJ, Nieuwland R, Tak PP, et al. Cell-derived microparticles in
synovial fluid from inflamed arthritic joints support coagulation exclusively via a
factor VII-dependent mechanism. Arthritis Rheum 2002;46:2857-66.
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

27
17. Muller I, Klocke A, Alex M, et al. Intravascular tissue factor initiates coagulation
via circulating microvesicles and platelets. FASEB J 2003;17:476-478
18. Zwaal RF, Schroit AJ. Pathophysiologic implications of membrane phospholipid
asymmetry in blood cells. Blood 1997 ;89:1121-32.
19. Barry OP, FitzGerald GA. Mechanisms of cellular activation by platelet
microparticles. Thromb Haemost 1999;82:794-800.
20. Sabatier F, Roux V, Anfosso F, Camoin L, Sampol J, Dignat-George F.
Interaction of endothelial microparticles with monocytic cells in vitro induces
tissue factor-dependent procoagulant activity. Blood 2002;99:3962-70.
21. DeLa Cadena RA, Majluf-Cruz A, Stadnicki A, Agosti JM, Colman RW,
Suffredini AF. Activation of the contact and fibrinolytic systems after intravenous
administration of endotoxin to normal human volunteers: correlation with the
cytokine profile. Immunopharmacology 1996;33:231-7.
22. Key NS, Slungaard A, Dandelet L, et al. Whole blood tissue factor procoagulant
activity is elevated in patients with sickle cell disease. Blood 1998;91:4216-23.
23. Ozcan M, Morton CT, Solovey A, et al. Whole blood tissue factor procoagulant
activity remains detectable during severe aplasia following bone marrow and
peripheral blood stem cell transplantation. Thromb Haemost 2001;85:250-5.
24. Osterud B. Tissue factor expression in monocytes: in vitro compared to ex vivo.
Thromb Haemost 2000;84:521-2.
25. Bloem LJ, Chen L, Konigsberg WH, Bach R. Serum stimulation of quiescent
human fibroblasts induces the synthesis of tissue factor mRNA followed by the
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

28
appearance of tissue factor antigen and procoagulant activity. J Cell Physiol
1989;139(2):418-23
26. Pernerstorfer T, Hollenstein U, Hansen JB, et al. Lepirudin blunts endotoxin-
induced coagulation activation. Blood 2000;95:1729-34.
27. Pernerstorfer T, Stohlawetz P, Hollenstein U, et al. Endotoxin-induced activation
of the coagulation cascade in humans: effect of acetylsalicylic acid and
acetaminophen. Arterioscler Thromb Vasc Biol 1999;19:2517-23.
28. Marsik C, Quehenberger P, Mackman N, Osterud B, Luther T, Jilma B.
Validation of a novel tissue factor assay in experimental human endotoxemia.
Thromb Res 2003;11194-5:311-5
29. Jaffe EA, Nachman RL, Becker CG, Minick CR. Culture of human endothelial
cells derived from umbilical veins: identification of morphological criteria. J Clin
Invest. 1973;52:2745-2756.
30. Gupta K, Ramakrishnan S, Browne PV, Solovey A, Hebbel RP. A novel
technique for culture of human dermal microvascular endothelial cells under
either serum-free or serum-supplemented conditions: isolation by panning and
stimulation with vascular endothelial growth factor. Exp Cell Res. 1997;230:244-
251.
31. Lin Y, Weisdorf DJ, Solovey A, Hebbel RP. Origins of circulating endothelial
cells and endothelial outgrowth from blood. J Clin Invest. 2000;105:1-77.
32. Carson SD, Perry GA, Pirruccello SJ. Fibroblast tissue factor: calcium and
ionophore induce shape changes, release of membrane vesicles, and redistribution
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

29
of tissue factor antigen in addition to increased procoagulant activity. Blood
1994;84:526-34.
33. Schecter AD, Spirn B, Rossikhina M, et al. Release of active tissue factor by
human arterial smooth muscle cells. Circ Res 2000;87:126-32.
34. Combes V, Simon AC, Grau GE, et al. In vitro generation of endothelial
microparticles and possible prothrombotic activity in patients with lupus
anticoagulant. J Clin Invest 1999;104:93-102.
35. Satta N, Toti F, Feugeas O, et al. Monocyte vesiculation is a possible mechanism
for dissemination of membrane-associated procoagulant activities and adhesion
molecules after stimulation by lipopolysaccharide. J Immunol 1994;153:3245-55.
36. Shet A, Aras O, Gupta K, et al. Sickle blood contains tissue factor positive
microparticles derived from endothelial cells and monocytes. Blood
2003;102(7):2678-83.
37. Franco RF, de Jonge E, Dekkers PE, et al. The in vivo kinetics of tissue factor
messenger RNA expression during human endotoxemia: relationship with
activation of coagulation. Blood 2000;96:554-9.
38. Singer KH, Scearce RM, Tuck DT, Whichard LP, Denning SM, Haynes BF.
Removal of fibroblasts from human epithelial cell cultures with use of a
complement fixing monoclonal antibody reactive with human fibroblasts and
monocytes/macrophages. J Invest Dermatol 1989;92:166-70.
39. Ronnov-Jessen L, Celis JE, Van Deurs B, Petersen OW. A fibroblast-associated
antigen: characterization in fibroblasts and immunoreactivity in smooth muscle
differentiated stromal cells. J Histochem Cytochem 1992;40:475-86..
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

30
40. Nijziel M, van Oerle R, van 't Veer C, van Pampus E, Lindhout T, Hamulyak K.
Tissue factor activity in human monocytes is regulated by plasma: implications
for the high and low responder phenomenon. Br J Haematol 2001;112:98-104.
41. Osterud B, Flaegstad T. Increased tissue thromboplastin activity in monocytes of
patients with meningococcal infection: related to an unfavourable prognosis.
Thromb Haemost 1983;49:5-7.
42. Østerud B. The high responder phenomenon: enhancement of LPS induced tissue
factor activity in monocytes by platelets and granulocytes. Platelets, 1995;6:119
125.
43. Bach RR, Moldow CF. Mechanism of tissue factor activation on HL-60 cells.
Blood 1997;89:3270-6.
44. Bach R, Rifkin DB. Expression of tissue factor procoagulant activity: regulation
by cytosolic calcium. Proc Natl Acad Sci U S A 1990;87:6995-9.
45. Maynard JR, Heckman CA, Pitlick FA, Nemerson Y. Association of tissue factor
activity with the surface of cultured cells. J Clin Invest 1975;55:814-24.
46. Giesen PL, Rauch U, Bohrmann B, et al. Blood-borne tissue factor: another view
of thrombosis. Proc Natl Acad Sci U S A 1999;96:2311-5.
47. Joop K, Berckmans RJ, Nieuwland R, et al. Microparticles from patients with
multiple organ dysfunction syndrome and sepsis support coagulation through
multiple mechanisms. Thromb Haemost 2001;85:810-20.
48. Conde I, Shrimpton CN, Thiagarajan P, Lopez JA. Tissue factor-bearing
microparticles arise from monocyte lipid rafts and can fuse with activated
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

31
platelets, consolidating all of the membrane-bound coagulation reactions on the
platelet surface. J Thromb Haemost 2003 (Suppl):Oct:146
49. Bach R, Jilma B, Mayr F, et al. Time course of tissue factor procoagulant activity
increases on mononuclear cells and platelets following endotoxin-induced
systemic inflammation in humans. Blood 2003 (Suppl):102(11);551a
50. de Jonge E, Dekkers PE, Creasey AA, et al. Tissue factor pathway inhibitor dose-
dependently inhibits coagulation activation without influencing the fibrinolytic
and cytokine response during human endotoxemia. Blood 2000;15;95:1124-9.
51. Jilma B, Marsik C, Mayr F, et al. Pharmacodynamics of active site-inhibited
factor VIIa in endotoxin-induced coagulation in humans. Clin Pharmacol Ther
2002;72:403-10.
52. Moons AH, Peters RJ, Cate H, et al. Recombinant nematode anticoagulant protein
c2, a novel inhibitor of tissue factor-factor VIIa activity, abrogates endotoxin-
induced coagulation in chimpanzees. Thromb Haemost 2002;88:627-31.
53. Santucci RA, Erlich J, Labriola J, et al. Measurement of tissue factor activity in
whole blood. Thromb Haemost 2000;83:445.
54. Taylor FB, Haddad PA, Hack E, et al. Two-stage response to endotoxin infusion
into normal human subjects: Correlation of blood phagocyte luminescence with
clinical and laboratory markers of the inflammatory, hemostatic response. Crit
Care Med 2001;29:326-34.
55. Koyama T, Nishida K, Ohdama S, et al. Determination of plasma tissue factor
antigen and its clinical significance. Br J Haematol 1994;87:343-7
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

32
56. Sturk-Maquelin KN, Nieuwland R, Romijn FP, Eijsman L, Hack CE, Sturk A.
Pro- and non-coagulant forms of non-cell-bound tissue factor in vivo. J Thromb
Haemost 2003;1:1920-1926
57. Fiore MM, Neuenschwander PF, Morrissey JH. The biochemical basis for the
apparent defect of soluble mutant tissue factor in enhancing the proteolytic
activities of factor VIIa. J Biol Chem 1994;269:143-9.
58. Østerud B, Olsen, J.O., Wilsgård, L. The role of arachidonic acid release and
lipoxygenase pathway in lipopolysaccharide-induced thromboplastin activity in
monocytes. Blood Coagulation & Fibrinolysis, 1990;1: 41 46.
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

33
FIGURE LEGENDS
Figure 1. Flow cytometric analysis and quantification of MPs in both ex vivo LPS
treated whole blood and in vivo endotoxin volunteers.
Determination of forward (FSC) and side-scatter (SSC) characteristics of light
by1.0 µm latex beads in buffer was used to establish the MP (or R1) gate (a).
The R2 (or bead) gate includes 7.2 µm latex beads used for enumeration of
MPs as described in Methods. Detection of PS-positive MPs by annexin-V-
Cy-5 labeling on the Y-axis in relation to light scatter (SSC) on the X-axis (b).
Determination of the limit for negative fluorescence, performed in the
presence of EDTA, was performed as a negative control for annexin V (c).
MPs isolated from whole blood following ex vivo stimulation by LPS are
shown in panels (d) and (e). In the remaining panels ((f) through (i)), MPs
isolated from whole blood of volunteers exposed to endotoxin are shown. In
all cases, MPs were triple labeled for Annexin V (not shown) with anti-
CD14-PE (Y-axis) and anti TF-FITC (X-axis) (d, f), or anti-CD144-PE (Y-
axis) and anti-TF-FITC (X-axis) (h) or relevant isotype control IgG-PE (Y-
axis) and IgG-FITC (X-axis) (e, g) and (i).
Figure 2. Expression of antigen recognized by 1B10 by a variety of cell types.
Fibroblast, human umbilical vein endothelial cells (HUVEC), microvascular
endothelial cells (MVEC) and smooth muscle cells were cultured and CD14
(+) monocytes, granulocytes, platelets and red cells were isolated as described
in Methods. These cells were then stained with 1B10 MoAb or a control
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

34
murine monoclonal IgM, followed by rabbit anti-mouse IgM-FITC and
analyzed by flow cytometry. Labeling with control IgM antibody is shown in
solid filled curve.
Figure 3. Time dependent increase in MP-associated TF PCA in platelet free
plasma following exposure of whole blood to LPS ex-vivo. Heparinized
whole blood samples from 5 healthy controls were stimulated with 10 ng/mL
LPS at 37oC for 24 hours. At various time points after LPS exposure, platelet
free plasma was prepared (as described in Methods) and assayed for MP-
associated TF activity. Each line represents a different individuals.
Figure 4a. Time dependent increase in MP-associated TF PCA in vivo after LPS
administration. EDTA-anticoagulated blood samples for assay of MP-
associated TF PCA were collected at baseline and various time points after
LPS infusion (2ng/kg) in 18 male volunteers. MP-associated TF PCA was
measured in platelet free plasma as described in Methods. Data are presented
as mean±SEM. * P<0.05 versus baseline.
Figure 4b. The MP-borne TF response following LPS exposure is highly variable
between subjects. The data from figure 4a are re-plotted by individual subject
to illustrate the variability of response.
Figure 5. MP-associated TF PCA is recovered in the pellet of ultracentrifuged
platelet free plasma following LPS exposure in vivo. Platelet free plasma
from two volunteers, obtained 3-4 hours after LPS exposure, was subjected to
high speed ultracentrifugation. The re-suspended pellet and supernatant
fractions were then independently assayed for MP-associated TF PCA, as
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

35
described in Methods. An inhibitory polyclonal antibody to TF (″+ Poly TF
Ab″) was included in some wells.
Figure 6. Ultrastructure and immuno-gold labelling of MPs. Preparation of samples
for electron microscopy is described in Methods. MPs were isolated from PFP
of a volunteer receiving endotoxin. In vivo generated MPs are seen as
vesicular structures with diameters ranging from 0.1 to 0.5 µm (6a). MPs
obtained by ultra-centrifugation were incubated with rabbit polyclonal anti-TF
antibody, and/or anti CD14 antibody or 1B10 anibody with appropriate
secondary gold labeled antibodies. In column I positive gold labeling to TF is
demonstrated (6b, 6 nm gold), CD14 (6e, 12 nm gold), 1B10 (6h, 12 nm
gold). Microparticles double labeled for 1B10 and TF (6k), and CD14 and TF
(7n) are also shown. No labeling was observed with control antibodies
(Column II) or secondary gold particle labeled antibodies alone (Column III).
Bars=100nm.
Figure 7. Whole blood TF PCA increases after LPS exposure. EDTA-anticoagulated
whole blood samples from LPS treated volunteers were frozen at -70°C
immediately after collection. Whole blood TF PCA was measured as
described in Methods. Data are presented as geometric means± 95%CI. * p <
0.05 versus baseline.
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

36
Figure 1
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom
37
Figure 2
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

38
Figure 3
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

39
Figure 4a and 4b
B
A
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

40
Figure 5
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

41
Figure 6
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

42
Figure 7
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

43
Table 1. Expression of Microparticle-Associated Tissue Factor Procoagulant
Activity on In vitro Generated Microparticlesa.
Cellular Origin of MPsMP-Associated TF PCA
in Supernatant[Xa], pmol/L/h/106 cells
FibroblastsIonophore (-)
Ionophore (+)>300>300
Smooth Muscle CellsIonophore (-)
Ionophore (+)>300>300
HUVECTNF (-)
TNF (+)4
26CD14(+) Monocytes
LPS (-) LPS (+)
320
PlateletsIonophore (-)
Ionophore (+)00
Red Blood CellsIonophore (-)
Ionophore (+)01
a Cells were isolated as described in Methods. Fibroblasts, smooth muscle cells, red blood
cells and platelets (106 cells in each case) were activated by exposure to 10µM ionophore
(or vehicle control) for 10 min at 37º C. Monocytes were activated by exposure to
10ng/mL LPS (or vehicle control) for 4 hrs at 37º C, and HUVEC by exposure to
10ng/mL TNF-α (or vehicle control) for 4 hrs at 37º C. In every case, the supernatant
was removed and assayed for MP-associated TF PCA as described in Methods.
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

44
Table 2. Number of Circulating Microparticles (MPs× 103/mL) In Endotoxin Treated Volunteers.
Time ( hour)
Total MPs
(n = 13)
CD14 (+) MPs
(n = 13)
VE Cadherin (+)
MPs
(n = 3)
TF(+) MPs
(n = 3)
-3 727±131† 104±42 3±3 26±8
3 1469±267 * 227±57 * 48 ±35 83 ±27
4 1333±235 * 192±52 * - -
8 890±537 91±33 84±84 100±81
† Data are expressed as mean±SEM
* P value <0.05 versus baseline
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom

45
Table 3. Spearman's Rho Correlations In Endotoxin Treated Volunteers.
MP-AssociatedTF PCA(n = 18)
Whole Blood TF PCA(n = 18)
Total Number of MPs(n = 13)
CD14 (+) MPs
(n =13)
TF mRNA
(n =18)
Whole Blood TF PCA
0.34 (0.0001)
Total number of MPs
0.56 (0.0002) 0.03 (0.88)
CD14(+) MPs 0.24 (0.14) 0.04 (0.80) 0.61 (0.00003)
TF mRNA 0.27 (0.06) 0.54 (0.0001) 0.40 (0.09) 0.54 (0.02)
.Data are shown as r value and (p value)
For personal use only. by guest on June 7, 2013. bloodjournal.hematologylibrary.orgFrom
Copyright © 2022 FDOKUMEN




















